the correlation among the dynamic change of zn2+, znt-1, and brain-derived neurotrophic factor after...
TRANSCRIPT

The Correlation Among the Dynamic Change of Zn2+,ZnT-1, and Brain-Derived Neurotrophic Factor AfterAcute Spinal Cord Injury in Rats
Yansong Wang & Xifan Mei & Li Zhang & Gang Lv
Received: 10 July 2010 /Accepted: 2 September 2010 /Published online: 14 September 2010# Springer Science+Business Media, LLC 2010
Abstract Zinc plays an important role in regulating the expression of brain-derivedneurotrophic factor (BDNF) and its receptor in nervous system, but the correlation amongZn2+, zinc transporter, and BDNF in spinal cord injuries (SCI) is not fully understood. Thepurpose of this study was to investigate the expression of Zn2+, zinc transporter 1 (ZnT-1),and BDNF, as well as their correlation in spinal cord-injured rats. One hundred Wistar malerats were divided into two groups: sham-operated group (as control group) and model group.Spinal cord injury was induced in model groups by hemisection of T9 on the left side.Compared with the control group, the serum zinc levels in SCI model group weresignificantly decreased after surgery, but zinc concentrations in spinal cord were increasedgradually. The mRNA levels of ZnT-1 and BDNF were significantly increased in SCI modelgroup, and there is a positive correlation between them (Spearman rho=0.381, P=0.0204).The correlation found between BDNF and ZnT-1 allows us to speculate that these two factorsmay be physiologically co-regulated, which may provide an idea for the treatment of SCI.
Keywords Zinc . ZnT-1 . Brain-derived neurotrophic factor . Spinal cord injury
Introduction
Spinal cord injuries (SCI) usually causes life-long disability for the patients, such asparaplegia and tetraplegia. Growth factors and their abilities to promote axonal regenerationand remyelination in injured spinal cord have been extensively studied [1–4]. Brain-derivedneurotrophic factor (BDNF) is an important neurotrophic factor, which can improve thepathological state of neurons and promote regeneration of injured neurons, that reducesneuronal apoptosis [5–7]. But, the expression of endogenous BDNF was not enough toparticipate in spinal cord repair. Exogenous BDNF has been applied to repair SCI, whichhas made some achievements [8–10]. However, how to prevent host rejection and cross theblood–brain barrier effectively are the problems that cannot be avoided.
Biol Trace Elem Res (2011) 143:351–358DOI 10.1007/s12011-010-8845-4
Y. Wang : X. Mei : L. Zhang :G. Lv (*)Liaoning Medical University, Jinzhou 121000, People’s Republic of Chinae-mail: [email protected]

Zinc is an essential trace element that plays a key role in multiple metabolic andsignaling pathways [11, 12]. In one study, the expression of BDNF was down-regulated inthe hippocampus and cortex of zinc deficiency rats [13]. Trk activation by zinc isextracellularly mediated by activation of metalloproteinases, which release pro-BDNF fromcells and convert pro-BDNF to mature BDNF [14]. Zinc cannot travel across biologicalmembranes by passive diffusion, specific membrane transporters, and channels involved inits transfer and metabolism [15]. The first putative zinc extruder, zinc transporter 1 (ZnT-1),has been suggested to play a key role in reducing cellular Zn2+ concentration [16]. Despitethe potential importance of ZnT-1 in the CNS, the expression of this protein and itscorrelation with BDNF in the SCI have not been described. The present study was carriedout to investigate the possible role of ZnT-1 in improving the expression of endogenousBDNF in spinal cord-injured rats by regulating Zn2+.
Materials and Methods
Animal Care
The study was approved by the Ethics Committee of Liaoning Medical University andcomplied with the Guidelines for the Care and Use of Laboratory Animals issued by theMinistry of Science and Technology of China in 2006.
One hundred healthy, clean, Wistar male rats, aged 2–3 months, weighing 250±20.5 g,were provided by the Laboratory Animal Center, Shengjing Hospital, China MedicalUniversity [permission No. SYXK (Liao) 2003–0019]. Animals were housed in individualstainless steel cages in a room with a 12–12 h light–dark schedule. The temperature wascontrolled to 21±2°C for the duration of the experiment with a relative humidity of 55%.Animals received sterilized food and water ad libitum.
Experimental Design
A total of 100 rats were randomly and evenly divided into two groups: in SCI model group,50 rats were anesthetized with a 4% chloral hydrate solution (360 mg/kg i.p.), and theirbacks were shaved and disinfected with iodine at the incision sites. Skin and muscles werecut over the caudal thoracic segments with a fine scalpel. The position of the vertebrae wasfixed by insertion of hooks into the connective tissue and muscles around the incision. Alaminectomy was performed at the T9 vertebrae. The spinal cord was then hemisected onthe left side using surgical scissors. The surrounding tissues were subsequently closed withsurgical sutures. About 5 ml of 0.9% NaCl was injected subcutaneously after the surgery.Penicillin (0.2 ml/kg) was administered subcutaneously at the end of the surgery in order toprevent infection. The animals had no significant health problems for weeks afterspinalization. Fifty sham-operated rats were treated with laminectomy at the T9 vertebrae,but the spinal cord was not hemisected.
Determination of Zinc in Blood Serum
Animals were fasted overnight before sampling days. After anesthesia with chloral hydratesolution, 1 ml blood was taken from the femoral artery in each group before surgery and at30 min, 24 h, 7 days, and 14 days after surgery, and the same volume of 0.9% NaCl wasgiven by tail vein injection. Blood was allowed to clot, and serum was prepared through
352 Wang et al.

centrifugation and aliquots were kept frozen at −80°C until analysis. All laboratory wareswere cleaned thoroughly with detergent and tap water, rinsed with distilled water, soaked indilute nitric acid, and then rinsed thoroughly with deionized distilled water. Levels of zincin serum were measured by Spectra Span V. Three Electrodes Direct Current PlasmaAtomic Emission Spectrometer (Beck Man, USA).
Determination of Zinc in Spinal Cord
Fifty rats were randomly and evenly divided into SCI model group and sham-operated group;after being anesthetized, T9 spinal cord was removed before surgery and at 30 min, 24 h,7 days, and 14 days after surgery, rinsed with ice cold saline, weighed, frozen on dry ice, andstored at −80°C. Then, the samples were digested, centrifuged, and assayed as above.
Determination of ZnT-1 mRNA and BDNF mRNA by Real-Time PCR
Real-time PCR was performed in Applied SYBR Green Biosystems 7500 Real-Time PCRSystem (Foster City, CA, USA) following the manufacturer’s instructions. Total cellularRNA from a portion of T9 spinal cord was placed in RNA STAT 60 reagent, and followingtheir protocol, RNA was purified and isolated. After a cycle of 95°C for 10 min, thereactions were cycled 40 times under the following parameters: 95°C for 30 s, then 56°Cfor 1 min, and 72°C for 1 min. A non-template control was run in order to exclude thepresence of genomic DNA, and all experiments were performed three times, using GAPDH(Chemicon International Temecula, CA, USA) as the internal reference. The BDNF, ZnT-1,and GAPDH primer sequences are shown in Table 1.
We performed an analysis using the ΔΔCt′; this procedure can be used since we havedetermined previously that the replication efficiencies (slopes of the calibration or standardcurves) for the genes of interest and housekeeping gene are very close. The selection of theprimers was performed using a Lasergene (DNA Star Inc. WI, USA) program.
Data Analysis
Data were collected in Excel (Microsoft). All data were expressed as the mean ± SD. Forstatistical analysis, differences between the three tested groups were assessed using the one-way analysis of variance and the post hoc Tukey’s test. Spearman’s rho tests were used to
Table 1 BDNF, ZnT-1, and GAPDH primer sequences
Primer Sequence
BDNF Upstream 5′-CAT TAC CTT CCT GCA TCT GTT GG-3′
Downstream 5′-CGT GGA CGT TAC TTC TTT CAT GG-3′
Length (bp) 231
ZnT-1 Upstream 5′-GAC CAG GAG GAG ACC AAC A -3′
Downstream 5′-GAA AAG TAG AAG ACC AAG GCA TT-3′
Length (bp) 263
GAPDH Upstream 5′-AAC AAG CAA CTG TCC CTG AGC-3′
Downstream 5′-GTA GAC AGA AGG TGG CAC AGA-3′
Length (bp) 453
The Dynamic Change of Zn2+, ZnT-1, and BDNF After SCI in Rats 353

detect relationships between BDNF mRNA and ZnT-1 mRNA. P<0.05 was considered toindicate significant differences.
Results
Content of Zinc in Blood Serum
Serum zinc concentrations in both control rats and SCI model rats are presented in Fig. 1.Compared with the control group, the serum zinc levels in SCI model group weresignificantly decreased at the same time points after surgery (P<0.05).
Content of Zinc in Spinal Cord
Zinc concentrations in spinal cord in SCI model group were increased gradually ascompared with the control group 24 h, 7 days, and 14 days after surgery (P<0.05), whichincreased along with the experimental time (Fig. 2).
Expression of ZnT-1 mRNA and BDNF mRNA by Real-Time PCR
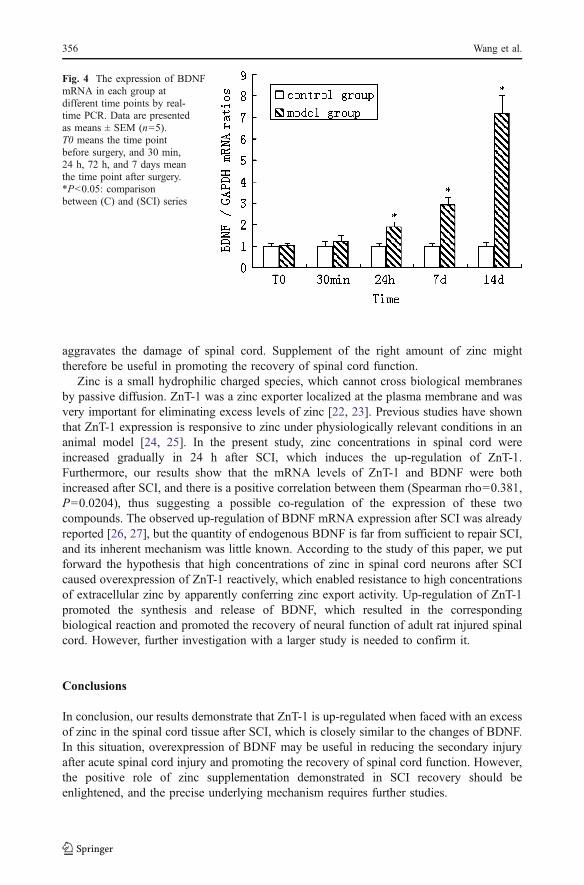
Results from Real-time PCR demonstrated that amplified products were specific (Figs. 3and 4). The mRNA levels of ZnT-1 and BDNF in control group were very low, and bothincreased after the spinal cord was hemisected 24 h later. A statistically significantdifference was observed between the two groups (P<0.05). Concentrations of ZnT-1 andBDNF positively correlated in SCI model group at the same time points after surgery,which was attested by the determination of the Spearman index (Spearman rho=0.381,P=0.0204).
Discussion
Zinc has an extremely diverse array of functions within the mammalian central nervoussystem as a key component of numerous enzymes [17, 18]. Recently, people present that
Fig. 1 Evolution of blood serumzinc levels in both control rats(C) and SCI model rats(SCI). Data are presentedas means ± SEM (n=5).T0 means the time pointbefore surgery, and 30 min,24 h, 72 h, and 7 days meanthe time point after surgery.*P<0.05: comparisonbetween (C) and (SCI) series
354 Wang et al.

zinc may be involved in regulating the expression of BDNF and its receptor in centralnervous system, which plays a key role in the pathophysiology and therapy of SCI [13, 14].However, a study on the correlation among dynamic change of Zn2+ and BDNF after acutespinal cord injury in rats is rare.
In the present study, SCI model group showed more decrease of zinc levels in serumafter surgery as compared with the control group (P<0.05), which was coincident withprevious reports [19]. The possible reasons are as follows: Leucocyte endogenous mediatorreleased from leucocytes during inflammation acts to redistribute the body zinc from theserum to the liver and may produce a drop in serum zinc [20]. Post-traumatic tissue needs alarge number of zinc to participate in, which results in the redistribution and the decrease ofzinc in vivo. At the same time, urinary zinc excretion in injured group increases obviously[21]. The decrease of zinc concentrations in serum was the result of SCI and in turn
Fig. 3 The expression ofZnT-1 mRNA in each groupat different time points by real-time PCR. Data are presentedas means ± SEM (n=5).T0 means the time pointbefore surgery, and 30 min,24 h, 72 h, and 7 days meanthe time point after surgery.*P<0.05: comparisonbetween (C) and (SCI) series
Fig. 2 Zinc concentrations ofspinal cord in both control rats(C) and SCI model rats (SCI).Data are presented as means ±SEM (n=5). T0 means the timepoint before surgery and 30 min,24 h, 72 h, and 7 days mean thetime point after surgery. *P<0.05: comparison between (C)and (SCI) series
The Dynamic Change of Zn2+, ZnT-1, and BDNF After SCI in Rats 355
aggravates the damage of spinal cord. Supplement of the right amount of zinc mighttherefore be useful in promoting the recovery of spinal cord function.
Zinc is a small hydrophilic charged species, which cannot cross biological membranesby passive diffusion. ZnT-1 was a zinc exporter localized at the plasma membrane and wasvery important for eliminating excess levels of zinc [22, 23]. Previous studies have shownthat ZnT-1 expression is responsive to zinc under physiologically relevant conditions in ananimal model [24, 25]. In the present study, zinc concentrations in spinal cord wereincreased gradually in 24 h after SCI, which induces the up-regulation of ZnT-1.Furthermore, our results show that the mRNA levels of ZnT-1 and BDNF were bothincreased after SCI, and there is a positive correlation between them (Spearman rho=0.381,P=0.0204), thus suggesting a possible co-regulation of the expression of these twocompounds. The observed up-regulation of BDNF mRNA expression after SCI was alreadyreported [26, 27], but the quantity of endogenous BDNF is far from sufficient to repair SCI,and its inherent mechanism was little known. According to the study of this paper, we putforward the hypothesis that high concentrations of zinc in spinal cord neurons after SCIcaused overexpression of ZnT-1 reactively, which enabled resistance to high concentrationsof extracellular zinc by apparently conferring zinc export activity. Up-regulation of ZnT-1promoted the synthesis and release of BDNF, which resulted in the correspondingbiological reaction and promoted the recovery of neural function of adult rat injured spinalcord. However, further investigation with a larger study is needed to confirm it.
Conclusions
In conclusion, our results demonstrate that ZnT-1 is up-regulated when faced with an excessof zinc in the spinal cord tissue after SCI, which is closely similar to the changes of BDNF.In this situation, overexpression of BDNF may be useful in reducing the secondary injuryafter acute spinal cord injury and promoting the recovery of spinal cord function. However,the positive role of zinc supplementation demonstrated in SCI recovery should beenlightened, and the precise underlying mechanism requires further studies.
Fig. 4 The expression of BDNFmRNA in each group atdifferent time points by real-time PCR. Data are presentedas means ± SEM (n=5).T0 means the time pointbefore surgery, and 30 min,24 h, 72 h, and 7 days meanthe time point after surgery.*P<0.05: comparisonbetween (C) and (SCI) series
356 Wang et al.

References
1. Fan J, Xiao Z, Zhang H, Chen B, Tang G, Hou X, Ding W, Wang B, Zhang P, Xu R, Jianwu D (2010)Linear ordered collagen scaffolds loaded with collagen-binding neurotrophin-3 promotes axonalregeneration and partial functional recovery after complete spinal cord transaction. J Neurotrauma.doi:10.1089/neu.2010.1281
2. Park HW, Lim MJ, Jung H, Lee SP, Paik KS, Chang MS (2010) Human mesenchymal stem cell-derivedSchwann cell-like cells exhibit neurotrophic effects, via distinct growth factor production, in a model ofspinal cord injury. Glia 58(9):1118–1132
3. Zhang L, Ma Z, Smith GM, Wen X, Pressman Y, Wood PM, Xu XM (2009) GDNF-enhanced axonalregeneration and myelination following spinal cord injury is mediated by primary effects on neurons.Glia 57(11):1178–1191
4. Zhou L, Shine HD (2003) Neurotrophic factors expressed in both cortex and spinal cord induce axonalplasticity after spinal cord injury. J Neurosci Res 74(2):221–226
5. Kamei N, Tanaka N, Oishi Y, Hamasaki T, Nakanishi K, Sakai N, Ochi M (2007) BDNF, NT-3, andNGF released from transplanted neural progenitor cells promote corticospinal axon growth inorganotypic cocultures. Spine 32(12):1272–1278
6. Sasaki M, Radtke C, Tan AM, Zhao P, Hamada H, Houkin K, Honmou O, Kocsis JD (2009) BDNF-hypersecreting human mesenchymal stem cells promote functional recovery, axonal sprouting, andprotection of corticospinal neurons after spinal cord injury. J Neurosci 29(47):14932–14941
7. Koyama R, Yamada MK, Fujisawa S, Katoh-Semba R, Matsuki N, Ikegaya Y (2004) Brain-derivedneurotrophic factor induces hyperexcitable reentrant circuits in the dentate gyrus. J Neurosci 24(33):7215–7224
8. Altar CA, Cai N, Bliven T, Juhasz M, Conner JM, Acheson AL, Lindsay RM, Wiegand SJ (1997)Anterograde transport of brain-derived neurotrophic factor and its role in the brain. Nature 389(6653):856–860
9. Ramer MS, Priestley JV, McMahon SB (2000) Functional regeneration of sensory axons into the adultspinal cord. Nature 403(6767):312–316
10. Vavrek R, Girgis J, Tetzlaff W, Hiebert GW, Fouad K (2006) BDNF promotes connections ofcorticospinal neurons onto spared descending interneurons in spinal cord injured rats. Brain 129(6):1534–1545
11. Kulikowska E, Moniuszko-Jakoniuk J, Miniuk K (1991) Role of zinc in physiologic and pathologicprocesses in the organism. Pol Tyg Lek 46(24–26):470–473
12. Li Y, Hawkins BE, DeWitt DS, Prough DS, Maret W (2010) The relationship between transient zinc ionfluctuations and redox signaling in the pathways of secondary cellular injury: relevance to traumaticbrain injury. Brain Res 1330:131–141
13. Fang H-Y, Jiang Y-G, Liu J, Fang H-T, Pang W (2008) Effects of zinc deficiency on the cAMP/PKA-CREB-BDNF signaling pathway of hippocampus and cortex in rats. Acta Nutrimenta Sinica 30(2):153–156 (In Chinese)
14. Hwang JJ, Park MH, Choi SY, Koh JY (2005) Activation of the Trk signaling pathway by extracellularzinc. Role of metalloproteinases. J Biol Chem 280(12):11995–12001
15. Zhang LY, Wang XL, Sun DX, Liu XX, Hu XY, Kong F (2008) Regulation of zinc transporters bydietary flaxseed lignan in human breast cancer xenografts. Mol Biol Rep 35(4):595–600
16. Sekler I, Moran A, Hershfinkel M, Dori A, Margulis A, Birenzweig N, Nitzan Y, Silverman WF (2002)Distribution of the zinc transporter ZnT-1 in comparison with chelatable zinc in the mouse brain. J CompNeurol 447(3):201–209
17. Failla ML (2003) Trace elements and host defense: recent advances and continuing challenges. J Nutr133(5 Suppl 1):1443S–1447S
18. Chimienti F, Aouffen M, Favier A, Seve M (2003) Zinc homeostasis-regulating proteins: new drugtargets for triggering cell fate. Curr Drug Targets 4(4):323–338
19. Simşek A, Senköylü A, Cila E, Uğurlu M, Bayar A, Oztürk AM, Işikli S, Muşdal Y, Yetkin H (2006) Isthere a correlation between severity of trauma and serum trace element levels? Acta Orthop TraumatolTurc 40(2):140–143 (in Turkish)
20. Pekarek RS, Wannemacher RW Jr, Beisel WR (1972) Effect of leukocytic endogenous mediator (LEM)on the tissue distribution of zinc and iron. Proc Soc Exp Biol Med 140:685–688
21. McClain CJ, Twyman DL, Ott LG, Rapp RP, Tibbs PA, Norton JA, Kasarskis EJ, Dempsey RJ, Young B(1986) Serum and urine zinc response in head-injured patients. J Neurosurg 64(2):224–230
22. McMahon RJ, Cousins RJ (1998) Regulation of the zinc transporter ZnT-1 by dietary zinc. Proc NatlAcad Sci USA 95(9):4841–4846
The Dynamic Change of Zn2+, ZnT-1, and BDNF After SCI in Rats 357

23. Liuzzi JP, Bobo JA, Lichten LA, Samuelson DA, Cousins RJ (2004) Responsive transporter genes withinthe murine intestinal-pancreatic axis form a basis of zinc homeostasis. Proc Natl Acad Sci USA 101(40):14355–14360, Epub
24. Liuzzi JP, Blanchard RK, Cousins RJ (2001) Differential regulation of zinc transporter 1, 2, and 4mRNA expression by dietary zinc in rats. J Nutr 131(1):46–52
25. Pfaffl MW, Windisch W (2003) Influence of zinc deficiency on the mRNA expression of zinctransporters in adult rats. J Trace Elem Med Biol 17(2):97–106
26. Qin DX, Zou XL, Luo W, Zhang W, Zhang HT, Li XL, Zhang H, Wang XY, Wang TH (2006)Expression of some neurotrophins in the spinal motoneurons after cord hemisection in adult rats.Neurosci Lett 410(3):222–227
27. Takahashi M, Palmer TD, Takahashi J, Gage FH (1998) Widespread integration and survival of adult-derived neural progenitor cells in the developing optic retina. Mol Cell Neurosci 12(6):340–348
358 Wang et al.