th17 cells contribute to viral replication in ... cells contribute to viral replication in...
TRANSCRIPT

of June 5, 2018.This information is current as
MyocarditisCoxsackievirus B3-Induced Acute Viral Th17 Cells Contribute to Viral Replication in
He-Ping Guo, Xiang Cheng and Yu-Hua LiaoJi-Hua Dong, Jin-Ping Wang, Jing-Hui Zhang, Min Wang, Jing Yuan, Miao Yu, Qiong-Wen Lin, Ai-Lin Cao, Xian Yu,
http://www.jimmunol.org/content/185/7/4004doi: 10.4049/jimmunol.1001718August 2010;
2010; 185:4004-4010; Prepublished online 27J Immunol
Referenceshttp://www.jimmunol.org/content/185/7/4004.full#ref-list-1
, 11 of which you can access for free at: cites 24 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Errata
/content/193/12/6208.full.pdf /content/187/6/3451.full.pdf
or: next pageAn erratum has been published regarding this article. Please see
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2010 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Th17 Cells Contribute to Viral Replication in CoxsackievirusB3-Induced Acute Viral Myocarditis
Jing Yuan,*,1 Miao Yu,*,1 Qiong-Wen Lin,*,1 Ai-Lin Cao,* Xian Yu,* Ji-Hua Dong,†
Jin-Ping Wang,† Jing-Hui Zhang,‡ Min Wang,* He-Ping Guo,* Xiang Cheng,*,2 and
Yu-Hua Liao*,2
Acute viral myocarditis (AVMC) is characterized by virus-triggeredmyocardial inflammation, andCoxsackievirus B3 (CVB3) is the
primary pathogen. We previously proved that Th17 cells, besides having proinflammatory effects, were involved in AVMC by en-
hancing humoral response. However, the relationship between Th17 cells and CVB3 replication remains unknown. In this exper-
iment, we infected BALB/c mice with CVB3 for establishing AVMC models and then found that, with the increase of viral
replication, the expressions of splenic Th17 cells, serum IL-17, and cardiac IL-17 mRNAwere elevated significantly, accompanied
by the progressive cardiac injuries of AVMC. Furthermore, on day 5, the peak time for viral replication, correlation was positive
between cardiac IL-17 mRNA and CVB3 RNA (correlation index = 0.835; p , 0.01). Although the expressions of Th1 and CD8+
T cells, which could secrete the antiviral cytokine IFN-g and damage the heart, were also elevated, along with Th17 cells, in
AVMC, the neutralization of IL-17 further upregulated the percentages of splenic Th1 and CD8+ T cells and the levels of cardiac
IFN-g mRNA. The cardiac pathological changes were obviously improved after neutralization, with reduced viral replication
followed by decreases in the cardiac inflammatory cytokines IL-17, TNF-a, and IL-1b. These data suggest that Th17 cells contribute
to CVB3 replication in AVMC, and that IL-17 might be an important target for regulating the balance of antiviral immunities. The
Journal of Immunology, 2010, 185: 4004–4010.
Acute viral myocarditis (AVMC), a precursor of dilatedcardiomyopathy leading to heart failure, is a triphasicdisease involving an initial viral infection, followed by
autoimmune response and finally remodeling of cardiac structureand function (1). Coxsackievirus B3 (CVB3), an enterovirus of thePicornaviridae family, is the primary pathogen of viral myocarditis(2). Although an excessively activated immune response triggeredby the virus is the dominant cause of myocyte injury, the virus itselfalso plays an important role in the development of AVMC throughdirect attack on myocardial cells (3).Recently, the unique CD4+ Th cell subset called Th17 cells have
been shown to be involved in several inflammatory diseases, includingexperimental autoimmune myocarditis, experimental autoimmuneencephalomyelitis, and collagen-induced arthritis (4). Besides theproinflammatory effect of Th17 cells, our earlier research suggested
that Th17 cells could facilitate the production of autoantibodies in thedevelopment of AVMC (5, 6). However, the relationship betweenTh17 cells and CVB3 replication remains unknown in AVMC.Therefore, in the current study, we investigated the dynamic
alterations of Th17 cells, viral replication, and related inflam-matory factors in the course of AVMC. At the same time, the IL-17 mAb was given to AVMC mice to further explore whether andhow Th17 cells are associated with viral replication in CVB3-induced AVMC.
Materials and MethodsMice
Male BALB/c mice aged 4 wk were purchased from the experimental animalresearch center (Hubei province, China). All animals were kept in thepathogen-free mouse room in the experimental animal center (Tongji MedicalCollege of Huazhong University of Science and Technology, Wuhan, China),and all experiments were carried out in accordance with guidelines for theCare and Use of Laboratory Animals (Science and Technology Department ofHubei Province, China, 2005).
Virus
The CVB3 (3m strain) was prepared by passage through Hela cells. Thevirus titer determined by PFU assay of HeLa cells was 1 3 107. BALB/cmice were infected by an i.p. injection of 0.2 ml RPMI 1640 (LifeTechnologies, Carlsbad, CA) containing ∼105 PFUs of the virus forestablishing AVMC models.
Interventions and groups
BALB/c mice were randomly divided into four groups: 1) in the AVMCgroup (n = 24), mice were treated with CVB3 and saline (100 ml permouse) i.p. on day 0; 2) in the IL-17 mAb group (n = 24), mice weretreated with CVB3 and anti-mouse IL-17 mAb (100 mg per mouse;eBioscience, San Diego, CA) i.p. on day 0; 3) in the isotype control group(n = 24), mice were treated with CVB3 and isotype control Ig G1Ab (100mg per mouse; eBioscience) i.p. on day 0; and 4) and in the normal group(n = 24), BALB/c mice were not administered anything else. On days 3, 5,7, and 10 of the experiment, six animals of each group were randomly
*Laboratory of Cardiovascular Immunology, Institute of Cardiology, †Laboratory ofVirology, and ‡Laboratory of General Surgery, Union Hospital, Huazhong Universityof Science and Technology, Wuhan, China
1J.Y., M.Y., and Q.-W.L. contributed equally to this work.
2X.C. and Y.-H.L. are joint corresponding authors.
Received for publication May 24, 2010. Accepted for publication July 21, 2010.
This work was supported by Grants 2007CB512000 and 2007CB512005 from theNational Basic Research Program of China (973 Program), Grant 2007A01 from theKey Laboratory of the Chinese Ministry, Grant 2008CDB147 from the Hubei Prov-ince Natural Science Foundation of China, and Grant NCET-09-0380 from the Pro-gram for New Century Excellent Talents, University of China (to X.C.).
Address correspondence and reprint requests to Prof. Yu-Hua Liao and Xiang Cheng,Laboratory of Cardiovascular Immunology, Institute of Cardiology, Union Hospital,Huazhong University of Science and Technology, 1277 Jie-Fang Avenue, Wuhan,430022, China. E-mail addresses: [email protected] and [email protected]
Abbreviations used in this paper: AVMC, acute viral myocarditis; cTnI, cardiactroponin I; CVB3, Coxsackievirus B3; HW/BW, heart weight to body weight; R,correlation index; RORgt, retinoic acid-related orphan receptor gt; T-bet, transcrip-tion factor T-box expressed in T cells.
Copyright� 2010 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1001718
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
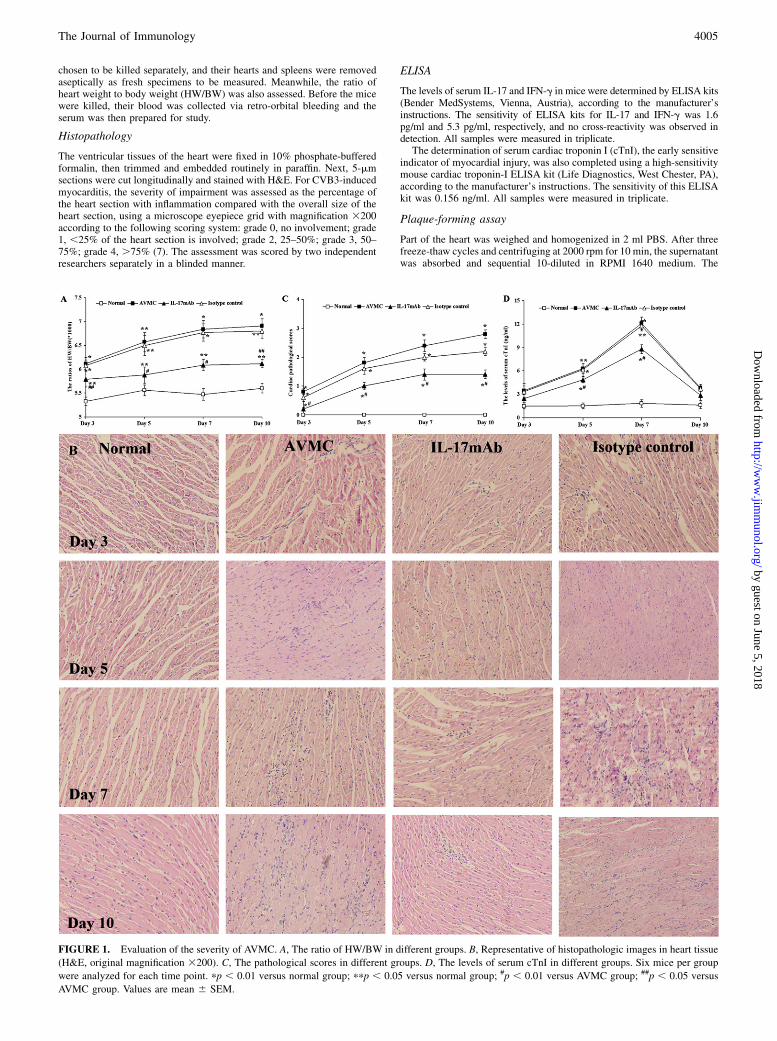
chosen to be killed separately, and their hearts and spleens were removedaseptically as fresh specimens to be measured. Meanwhile, the ratio ofheart weight to body weight (HW/BW) was also assessed. Before the micewere killed, their blood was collected via retro-orbital bleeding and theserum was then prepared for study.
Histopathology
The ventricular tissues of the heart were fixed in 10% phosphate-bufferedformalin, then trimmed and embedded routinely in paraffin. Next, 5-mmsections were cut longitudinally and stained with H&E. For CVB3-inducedmyocarditis, the severity of impairment was assessed as the percentage ofthe heart section with inflammation compared with the overall size of theheart section, using a microscope eyepiece grid with magnification 3200according to the following scoring system: grade 0, no involvement; grade1, ,25% of the heart section is involved; grade 2, 25–50%; grade 3, 50–75%; grade 4, .75% (7). The assessment was scored by two independentresearchers separately in a blinded manner.
ELISA
The levels of serum IL-17 and IFN-g in mice were determined by ELISA kits(Bender MedSystems, Vienna, Austria), according to the manufacturer’sinstructions. The sensitivity of ELISA kits for IL-17 and IFN-g was 1.6pg/ml and 5.3 pg/ml, respectively, and no cross-reactivity was observed indetection. All samples were measured in triplicate.
The determination of serum cardiac troponin I (cTnI), the early sensitiveindicator of myocardial injury, was also completed using a high-sensitivitymouse cardiac troponin-I ELISA kit (Life Diagnostics, West Chester, PA),according to the manufacturer’s instructions. The sensitivity of this ELISAkit was 0.156 ng/ml. All samples were measured in triplicate.
Plaque-forming assay
Part of the heart was weighed and homogenized in 2 ml PBS. After threefreeze-thaw cycles and centrifuging at 2000 rpm for 10 min, the supernatantwas absorbed and sequential 10-diluted in RPMI 1640 medium. The
FIGURE 1. Evaluation of the severity of AVMC. A, The ratio of HW/BW in different groups. B, Representative of histopathologic images in heart tissue
(H&E, original magnification 3200). C, The pathological scores in different groups. D, The levels of serum cTnI in different groups. Six mice per group
were analyzed for each time point. pp , 0.01 versus normal group; ppp , 0.05 versus normal group; #p , 0.01 versus AVMC group; ##p , 0.05 versus
AVMC group. Values are mean 6 SEM.
The Journal of Immunology 4005
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
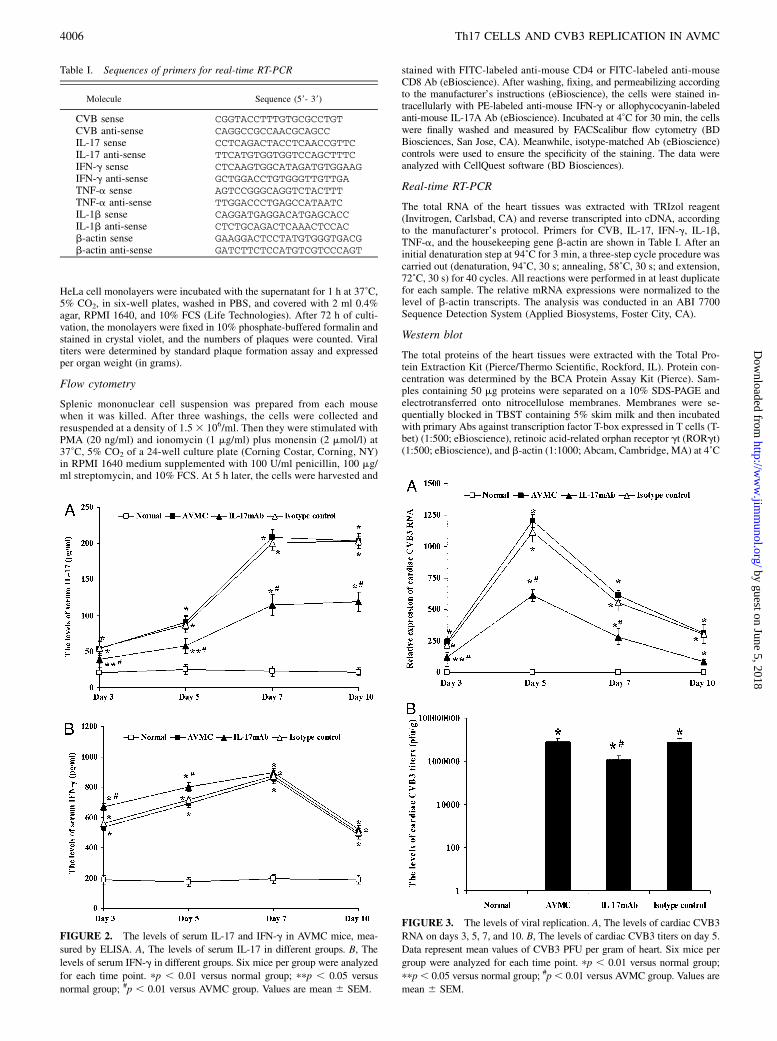
HeLa cell monolayers were incubated with the supernatant for 1 h at 37˚C,5% CO2, in six-well plates, washed in PBS, and covered with 2 ml 0.4%agar, RPMI 1640, and 10% FCS (Life Technologies). After 72 h of culti-vation, the monolayers were fixed in 10% phosphate-buffered formalin andstained in crystal violet, and the numbers of plaques were counted. Viraltiters were determined by standard plaque formation assay and expressedper organ weight (in grams).
Flow cytometry
Splenic mononuclear cell suspension was prepared from each mousewhen it was killed. After three washings, the cells were collected andresuspended at a density of 1.53 106/ml. Then they were stimulated withPMA (20 ng/ml) and ionomycin (1 mg/ml) plus monensin (2 mmol/l) at37˚C, 5% CO2 of a 24-well culture plate (Corning Costar, Corning, NY)in RPMI 1640 medium supplemented with 100 U/ml penicillin, 100 mg/ml streptomycin, and 10% FCS. At 5 h later, the cells were harvested and
stained with FITC-labeled anti-mouse CD4 or FITC-labeled anti-mouseCD8 Ab (eBioscience). After washing, fixing, and permeabilizing accordingto the manufacturer’s instructions (eBioscience), the cells were stained in-tracellularly with PE-labeled anti-mouse IFN-g or allophycocyanin-labeledanti-mouse IL-17A Ab (eBioscience). Incubated at 4˚C for 30 min, the cellswere finally washed and measured by FACScalibur flow cytometry (BDBiosciences, San Jose, CA). Meanwhile, isotype-matched Ab (eBioscience)controls were used to ensure the specificity of the staining. The data wereanalyzed with CellQuest software (BD Biosciences).
Real-time RT-PCR
The total RNA of the heart tissues was extracted with TRIzol reagent(Invitrogen, Carlsbad, CA) and reverse transcripted into cDNA, accordingto the manufacturer’s protocol. Primers for CVB, IL-17, IFN-g, IL-1b,TNF-a, and the housekeeping gene b-actin are shown in Table I. After aninitial denaturation step at 94˚C for 3 min, a three-step cycle procedure wascarried out (denaturation, 94˚C, 30 s; annealing, 58˚C, 30 s; and extension,72˚C, 30 s) for 40 cycles. All reactions were performed in at least duplicatefor each sample. The relative mRNA expressions were normalized to thelevel of b-actin transcripts. The analysis was conducted in an ABI 7700Sequence Detection System (Applied Biosystems, Foster City, CA).
Western blot
The total proteins of the heart tissues were extracted with the Total Pro-tein Extraction Kit (Pierce/Thermo Scientific, Rockford, IL). Protein con-centration was determined by the BCA Protein Assay Kit (Pierce). Sam-ples containing 50 mg proteins were separated on a 10% SDS-PAGE andelectrotransferred onto nitrocellulose membranes. Membranes were se-quentially blocked in TBST containing 5% skim milk and then incubatedwith primary Abs against transcription factor T-box expressed in T cells (T-bet) (1:500; eBioscience), retinoic acid-related orphan receptor gt (RORgt)(1:500; eBioscience), and b-actin (1:1000; Abcam, Cambridge, MA) at 4˚C
Table I. Sequences of primers for real-time RT-PCR
Molecule Sequence (59- 39)
CVB sense CGGTACCTTTGTGCGCCTGTCVB anti-sense CAGGCCGCCAACGCAGCCIL-17 sense CCTCAGACTACCTCAACCGTTCIL-17 anti-sense TTCATGTGGTGGTCCAGCTTTCIFN-g sense CTCAAGTGGCATAGATGTGGAAGIFN-g anti-sense GCTGGACCTGTGGGTTGTTGATNF-a sense AGTCCGGGCAGGTCTACTTTTNF-a anti-sense TTGGACCCTGAGCCATAATCIL-1b sense CAGGATGAGGACATGAGCACCIL-1b anti-sense CTCTGCAGACTCAAACTCCACb-actin sense GAAGGACTCCTATGTGGGTGACGb-actin anti-sense GATCTTCTCCATGTCGTCCCAGT
FIGURE 2. The levels of serum IL-17 and IFN-g in AVMC mice, mea-
sured by ELISA. A, The levels of serum IL-17 in different groups. B, The
levels of serum IFN-g in different groups. Six mice per group were analyzed
for each time point. pp , 0.01 versus normal group; ppp , 0.05 versus
normal group; #p , 0.01 versus AVMC group. Values are mean 6 SEM.
FIGURE 3. The levels of viral replication. A, The levels of cardiac CVB3
RNA on days 3, 5, 7, and 10. B, The levels of cardiac CVB3 titers on day 5.
Data represent mean values of CVB3 PFU per gram of heart. Six mice per
group were analyzed for each time point. pp , 0.01 versus normal group;
ppp, 0.05 versus normal group; #p, 0.01 versus AVMC group. Values are
mean 6 SEM.
4006 Th17 CELLS AND CVB3 REPLICATION IN AVMC
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
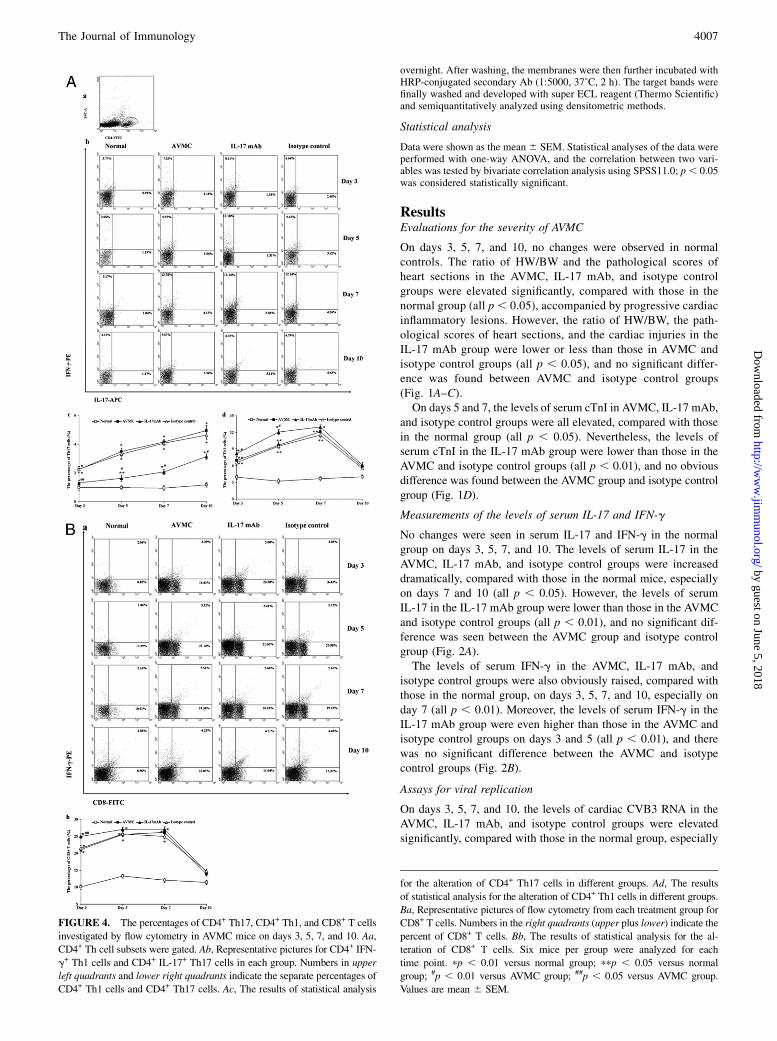
overnight. After washing, the membranes were then further incubated withHRP-conjugated secondary Ab (1:5000, 37˚C, 2 h). The target bands werefinally washed and developed with super ECL reagent (Thermo Scientific)and semiquantitatively analyzed using densitometric methods.
Statistical analysis
Data were shown as the mean6 SEM. Statistical analyses of the data wereperformed with one-way ANOVA, and the correlation between two vari-ables was tested by bivariate correlation analysis using SPSS11.0; p, 0.05was considered statistically significant.
ResultsEvaluations for the severity of AVMC
On days 3, 5, 7, and 10, no changes were observed in normalcontrols. The ratio of HW/BW and the pathological scores ofheart sections in the AVMC, IL-17 mAb, and isotype controlgroups were elevated significantly, compared with those in thenormal group (all p, 0.05), accompanied by progressive cardiacinflammatory lesions. However, the ratio of HW/BW, the path-ological scores of heart sections, and the cardiac injuries in theIL-17 mAb group were lower or less than those in AVMC andisotype control groups (all p , 0.05), and no significant differ-ence was found between AVMC and isotype control groups(Fig. 1A–C).On days 5 and 7, the levels of serum cTnI in AVMC, IL-17 mAb,
and isotype control groups were all elevated, compared with thosein the normal group (all p , 0.05). Nevertheless, the levels ofserum cTnI in the IL-17 mAb group were lower than those in theAVMC and isotype control groups (all p , 0.01), and no obviousdifference was found between the AVMC group and isotype controlgroup (Fig. 1D).
Measurements of the levels of serum IL-17 and IFN-g
No changes were seen in serum IL-17 and IFN-g in the normalgroup on days 3, 5, 7, and 10. The levels of serum IL-17 in theAVMC, IL-17 mAb, and isotype control groups were increaseddramatically, compared with those in the normal mice, especiallyon days 7 and 10 (all p , 0.05). However, the levels of serumIL-17 in the IL-17 mAb group were lower than those in the AVMCand isotype control groups (all p , 0.01), and no significant dif-ference was seen between the AVMC group and isotype controlgroup (Fig. 2A).The levels of serum IFN-g in the AVMC, IL-17 mAb, and
isotype control groups were also obviously raised, compared withthose in the normal group, on days 3, 5, 7, and 10, especially onday 7 (all p , 0.01). Moreover, the levels of serum IFN-g in theIL-17 mAb group were even higher than those in the AVMC andisotype control groups on days 3 and 5 (all p , 0.01), and therewas no significant difference between the AVMC and isotypecontrol groups (Fig. 2B).
Assays for viral replication
On days 3, 5, 7, and 10, the levels of cardiac CVB3 RNA in theAVMC, IL-17 mAb, and isotype control groups were elevatedsignificantly, compared with those in the normal group, especially
FIGURE 4. The percentages of CD4+ Th17, CD4+ Th1, and CD8+ T cells
investigated by flow cytometry in AVMC mice on days 3, 5, 7, and 10. Aa,
CD4+ Th cell subsets were gated. Ab, Representative pictures for CD4+ IFN-
g+ Th1 cells and CD4+ IL-17+ Th17 cells in each group. Numbers in upper
left quadrants and lower right quadrants indicate the separate percentages of
CD4+ Th1 cells and CD4+ Th17 cells. Ac, The results of statistical analysis
for the alteration of CD4+ Th17 cells in different groups. Ad, The results
of statistical analysis for the alteration of CD4+ Th1 cells in different groups.
Ba, Representative pictures of flow cytometry from each treatment group for
CD8+ T cells. Numbers in the right quadrants (upper plus lower) indicate the
percent of CD8+ T cells. Bb, The results of statistical analysis for the al-
teration of CD8+ T cells. Six mice per group were analyzed for each
time point. pp , 0.01 versus normal group; ppp , 0.05 versus normal
group; #p , 0.01 versus AVMC group; ##p , 0.05 versus AVMC group.
Values are mean 6 SEM.
The Journal of Immunology 4007
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
on day 5 (all p, 0.01). However, the levels of cardiac CVB3 RNAin the IL-17 mAb group were lower than those in the AVMC andisotype control groups on days 3 and 5 (all p , 0.05), and nodifference was noted between the AVMC group and isotype controlgroup (Fig. 3A, Table I).On day 5, the levels of cardiac CVB3 titers in the AVMC, IL-17
mAb, and isotype control groups were higher than those in thenormal group (p , 0.01), and that in the IL-17 mAb group waslower than those in the AVMC and isotype control groups (p ,0.05). No difference was found between the AVMC and isotypecontrol groups (Fig. 3B).At the same time points, cardiac CVB3 was not detected in
normal controls (Fig. 3A, 3B).
Investigations for percentages of CD4+ Th17, CD4+ Th1, andCD8+ T cells
On days 3, 5, 7, and 10, the percentages of CD4+ Th17 cells in theAVMC, IL-17 mAb, and isotype control groups were increasedmarkedly, compared with those in the normal group, especially ondays 7 and 10 (all p , 0.05). However, the percentages of CD4+
Th17 cells in the IL-17 mAb group were lower than those in the
AVMC and isotype control groups (all p , 0.05), and there was nosignificant difference between the AVMC group and isotype controlgroup (Fig. 4A).On days 3, 5, and 7, the percentages of CD4+ Th1 cells in the
AVMC, IL-17 mAb, and isotype control groups were also elevated,compared with those in the normal group, especially on day 7 (allp , 0.05). Furthermore, on days 3 and 5, the percentages of CD4+
Th1 cells in the IL-17 mAb group were even higher than those inthe AVMC and isotype control groups (all p , 0.05), and no sig-nificant difference was noted between the AVMC group and isotypecontrol group (Fig. 4A).On days 3, 5, and 7, the percentages of CD8+ T cells in the
AVMC, IL-17 mAb, and isotype control groups rose significantly,compared with those in the normal group (all p , 0.05). Moreover,on day 3, the percentages of CD8+ T cells in the IL-17 mAb groupwere even higher than those in the AVMC and isotype controlgroups (p , 0.05), and no difference was observed between theAVMC and isotype control groups (Fig. 4B).Throughout the course of the experiment, there were no changes
in the percentages of CD4+ Th17 cells, CD4+ Th1 cells, and CD8+
T cells in normal mice (Fig. 4A, 4B).
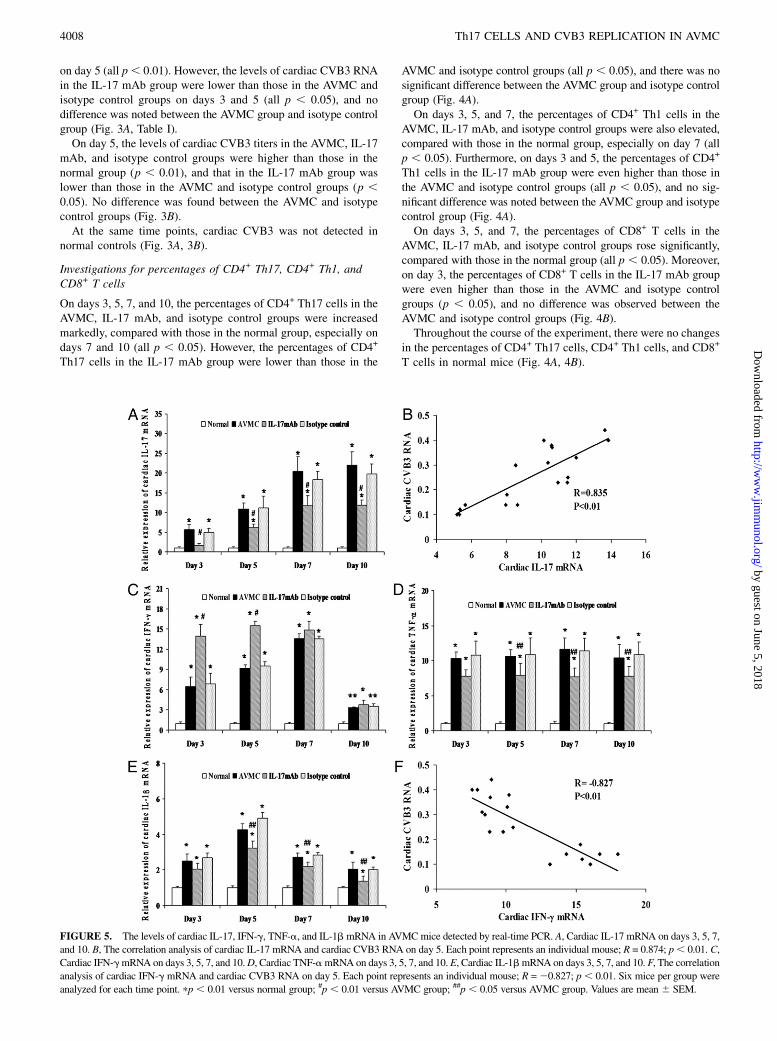
FIGURE 5. The levels of cardiac IL-17, IFN-g, TNF-a, and IL-1b mRNA in AVMCmice detected by real-time PCR. A, Cardiac IL-17 mRNA on days 3, 5, 7,
and 10. B, The correlation analysis of cardiac IL-17 mRNA and cardiac CVB3 RNA on day 5. Each point represents an individual mouse; R = 0.874; p, 0.01. C,
Cardiac IFN-gmRNA on days 3, 5, 7, and 10.D, Cardiac TNF-amRNA on days 3, 5, 7, and 10. E, Cardiac IL-1bmRNA on days 3, 5, 7, and 10. F, The correlation
analysis of cardiac IFN-g mRNA and cardiac CVB3 RNA on day 5. Each point represents an individual mouse; R =20.827; p, 0.01. Six mice per group were
analyzed for each time point. pp , 0.01 versus normal group; #p , 0.01 versus AVMC group; ##p , 0.05 versus AVMC group. Values are mean 6 SEM.
4008 Th17 CELLS AND CVB3 REPLICATION IN AVMC
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
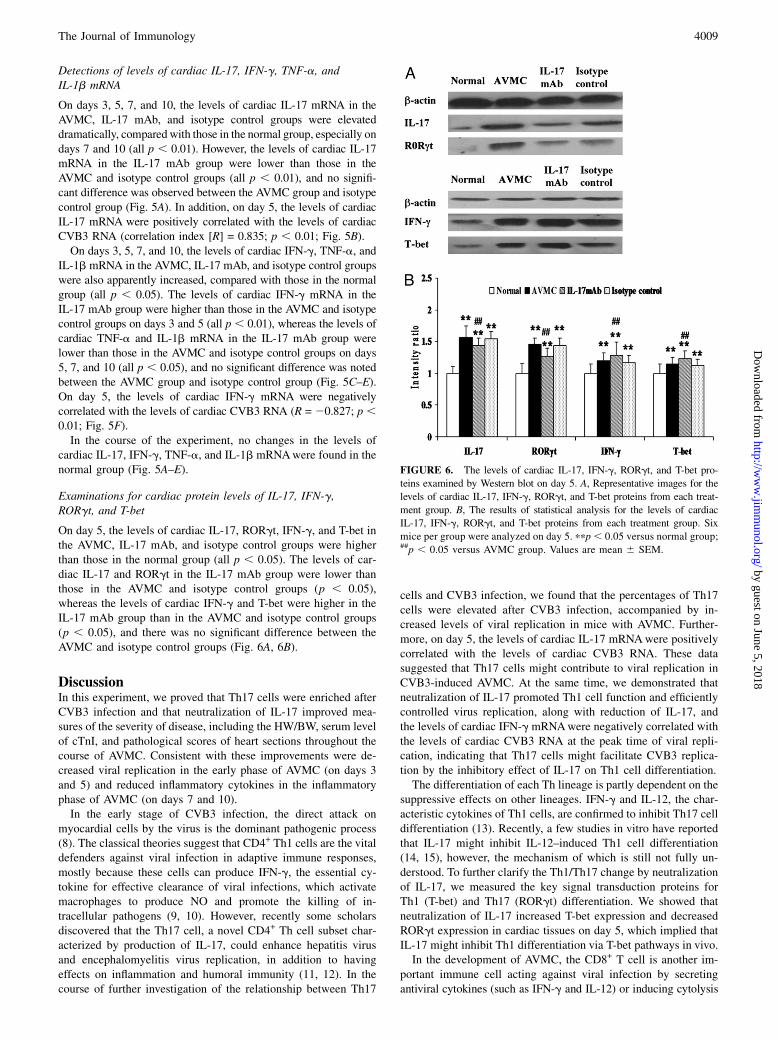
Detections of levels of cardiac IL-17, IFN-g, TNF-a, andIL-1b mRNA
On days 3, 5, 7, and 10, the levels of cardiac IL-17 mRNA in theAVMC, IL-17 mAb, and isotype control groups were elevateddramatically, compared with those in the normal group, especially ondays 7 and 10 (all p , 0.01). However, the levels of cardiac IL-17mRNA in the IL-17 mAb group were lower than those in theAVMC and isotype control groups (all p , 0.01), and no signifi-cant difference was observed between the AVMC group and isotypecontrol group (Fig. 5A). In addition, on day 5, the levels of cardiacIL-17 mRNA were positively correlated with the levels of cardiacCVB3 RNA (correlation index [R] = 0.835; p , 0.01; Fig. 5B).On days 3, 5, 7, and 10, the levels of cardiac IFN-g, TNF-a, and
IL-1b mRNA in the AVMC, IL-17 mAb, and isotype control groupswere also apparently increased, compared with those in the normalgroup (all p , 0.05). The levels of cardiac IFN-g mRNA in theIL-17 mAb group were higher than those in the AVMC and isotypecontrol groups on days 3 and 5 (all p , 0.01), whereas the levels ofcardiac TNF-a and IL-1b mRNA in the IL-17 mAb group werelower than those in the AVMC and isotype control groups on days5, 7, and 10 (all p , 0.05), and no significant difference was notedbetween the AVMC group and isotype control group (Fig. 5C–E).On day 5, the levels of cardiac IFN-g mRNA were negativelycorrelated with the levels of cardiac CVB3 RNA (R = 20.827; p ,0.01; Fig. 5F).In the course of the experiment, no changes in the levels of
cardiac IL-17, IFN-g, TNF-a, and IL-1b mRNAwere found in thenormal group (Fig. 5A–E).
Examinations for cardiac protein levels of IL-17, IFN-g,RORgt, and T-bet
On day 5, the levels of cardiac IL-17, RORgt, IFN-g, and T-bet inthe AVMC, IL-17 mAb, and isotype control groups were higherthan those in the normal group (all p , 0.05). The levels of car-diac IL-17 and RORgt in the IL-17 mAb group were lower thanthose in the AVMC and isotype control groups (p , 0.05),whereas the levels of cardiac IFN-g and T-bet were higher in theIL-17 mAb group than in the AVMC and isotype control groups(p , 0.05), and there was no significant difference between theAVMC and isotype control groups (Fig. 6A, 6B).
DiscussionIn this experiment, we proved that Th17 cells were enriched afterCVB3 infection and that neutralization of IL-17 improved mea-sures of the severity of disease, including the HW/BW, serum levelof cTnI, and pathological scores of heart sections throughout thecourse of AVMC. Consistent with these improvements were de-creased viral replication in the early phase of AVMC (on days 3and 5) and reduced inflammatory cytokines in the inflammatoryphase of AVMC (on days 7 and 10).In the early stage of CVB3 infection, the direct attack on
myocardial cells by the virus is the dominant pathogenic process(8). The classical theories suggest that CD4+ Th1 cells are the vitaldefenders against viral infection in adaptive immune responses,mostly because these cells can produce IFN-g, the essential cy-tokine for effective clearance of viral infections, which activatemacrophages to produce NO and promote the killing of in-tracellular pathogens (9, 10). However, recently some scholarsdiscovered that the Th17 cell, a novel CD4+ Th cell subset char-acterized by production of IL-17, could enhance hepatitis virusand encephalomyelitis virus replication, in addition to havingeffects on inflammation and humoral immunity (11, 12). In thecourse of further investigation of the relationship between Th17
cells and CVB3 infection, we found that the percentages of Th17cells were elevated after CVB3 infection, accompanied by in-creased levels of viral replication in mice with AVMC. Further-more, on day 5, the levels of cardiac IL-17 mRNAwere positivelycorrelated with the levels of cardiac CVB3 RNA. These datasuggested that Th17 cells might contribute to viral replication inCVB3-induced AVMC. At the same time, we demonstrated thatneutralization of IL-17 promoted Th1 cell function and efficientlycontrolled virus replication, along with reduction of IL-17, andthe levels of cardiac IFN-g mRNAwere negatively correlated withthe levels of cardiac CVB3 RNA at the peak time of viral repli-cation, indicating that Th17 cells might facilitate CVB3 replica-tion by the inhibitory effect of IL-17 on Th1 cell differentiation.The differentiation of each Th lineage is partly dependent on the
suppressive effects on other lineages. IFN-g and IL-12, the char-acteristic cytokines of Th1 cells, are confirmed to inhibit Th17 celldifferentiation (13). Recently, a few studies in vitro have reportedthat IL-17 might inhibit IL-12–induced Th1 cell differentiation(14, 15), however, the mechanism of which is still not fully un-derstood. To further clarify the Th1/Th17 change by neutralizationof IL-17, we measured the key signal transduction proteins forTh1 (T-bet) and Th17 (RORgt) differentiation. We showed thatneutralization of IL-17 increased T-bet expression and decreasedRORgt expression in cardiac tissues on day 5, which implied thatIL-17 might inhibit Th1 differentiation via T-bet pathways in vivo.In the development of AVMC, the CD8+ T cell is another im-
portant immune cell acting against viral infection by secretingantiviral cytokines (such as IFN-g and IL-12) or inducing cytolysis
FIGURE 6. The levels of cardiac IL-17, IFN-g, RORgt, and T-bet pro-
teins examined by Western blot on day 5. A, Representative images for the
levels of cardiac IL-17, IFN-g, RORgt, and T-bet proteins from each treat-
ment group. B, The results of statistical analysis for the levels of cardiac
IL-17, IFN-g, RORgt, and T-bet proteins from each treatment group. Six
mice per group were analyzed on day 5. ppp, 0.05 versus normal group;##p , 0.05 versus AVMC group. Values are mean 6 SEM.
The Journal of Immunology 4009
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

of infected cells via Fas and perforin pathways (16, 17). Martin-Orozco et al. (18) considered that Th17 cells could amplify CD8+
T cell activation in lung melanoma. Conversely, other investigationsshowed that Th17 cells could downregulate CD8+ T cell function byrepressing the production of IL-12 and IFN-g (12, 19). In addition,IL-17 could increase the expression of antiapoptotic proteins of theBcl-2 family though the NF-kB signaling pathway, which mightimpair the Fas pathway of CD8+ T cells via protecting theinfected cells from apoptosis (12, 20). In this study, we observedthat neutralization of IL-17 enhanced the production of CD8+
T cells and suppressed the levels of CVB3 replication on day 3,which provided evidence that Th17 cells might contribute to CVB3replication by inhibiting CD8+ T cell activity.Although CD4+ Th1 cells and CD8+ T cells perform a function in
antiviral immunity, they are also believed to exacerbate myocarditisby autoimmunity associated with excessive release of proinflam-matory cytokines, such as TNF-a and IL-1b. Thus, a balance be-tween antiviral immunity and autoimmunity might exist in AVMC(21, 22). To clarify whether the increased proportions of Th1 cellsand CD8+ T cells would lead to aggressive myocarditis in thisexperiment, we detected the expression of TNF-a and IL-1b, whichwere the dominant inflammatory mediators of myocardial injuryin the inflammatory phase of CVB3 infection in AVMC (23, 24).Unlike the elevation in percentages of CD8+ T cells and CD4+
Th1 cells, the levels of cardiac TNF-a and IL-1b mRNA declinedalong with the reduced IL-17 and decreased CVB3 replication,which diminished the severity of AVMC. We then could concludethat IL-17 produced by Th17 cells might play a role in regulatingthe balance between antiviral immunity and autoimmunity inCVB3-induced AVMC and that IL-17 would be a novel therapeutictarget for AVMC.
AcknowledgmentsWe thank all the members of the experimental animal center of the Tongji
Medical College (Huazhong University of Science and Technology) for
excellent animal care.
DisclosuresThe authors have no financial conflicts of interest.
References1. Dennert, R., H. J. Crijns, and S. Heymans. 2008. Acute viral myocarditis. Eur.
Heart J. 29: 2073–2082.2. Deonarain, R., D. Cerullo, K. Fuse, P. P. Liu, and E. N. Fish. 2004. Protective
role for interferon-beta in coxsackievirus B3 infection. Circulation 110: 3540–3543.
3. Liu, P. P., and J. W. Mason. 2001. Advances in the understanding of myocarditis.Circulation 104: 1076–1082.
4. Tesmer, L. A., S. K. Lundy, S. Sarkar, and D. A. Fox. 2008. Th17 cells in humandisease. Immunol. Rev. 223: 87–113.
5. Yuan, J., A. L. Cao, M. Yu, Q. W. Lin, X. Yu, J. H. Zhang, M. Wang, H. P. Guo,and Y. H. Liao. 2010. Th17 cells facilitate the humoral immune response inpatients with acute viral myocarditis. J. Clin. Immunol. 30: 226–234.
6. Yuan, J., M. Yu, Q. W. Lin, A. L. Cao, X. Yu, J. H. Dong, J. P. Wang,J. H. Zhang, M. Wang, H. P. Guo, and Y. H. Liao. 2010. Neutralization of IL-17inhibits the production of anti-ANT autoantibodies in CVB3-induced acuteviral myocarditis. Int. Immunopharmacol. 10: 272–276.
7. Nishio, R., A. Matsumori, T. Shioi, H. Ishida, and S. Sasayama. 1999. Treatmentof experimental viral myocarditis with interleukin-10. Circulation 100: 1102–1108.
8. Wang, Y. X., V. da Cunha, J. Vincelette, K. White, S. Velichko, Y. F. Xu,C. Gross, R. M. Fitch, M. Halks-Miller, B. R. Larsen, et al. 2007. Antiviral andmyocyte protective effects of murine interferon-beta and -alpha2 in coxsack-ievirus B3-induced myocarditis and epicarditis in Balb/c mice. Am. J. Physiol.Heart Circ. Physiol. 293: H69–H76.
9. Jin, B., R. Y.Wang, Q. Qiu, F. Sugauchi, T. Grandinetti, H. J. Alter, and J. W. Shih.2007. Induction of potent cellular immune response in mice by hepatitis C virusNS3 protein with double-stranded RNA. Immunology 122: 15–27.
10. Bogdan, C. 2001. Nitric oxide and the immune response. Nat. Immunol. 2: 907–916.
11. Zhang, J. Y., Z. Zhang, F. Lin, Z. S. Zou, R. N. Xu, L. Jin, J. L. Fu, F. Shi,M. Shi, H. F. Wang, and F. S. Wang. 2010. Interleukin-17-producing CD4(+)T cells increase with severity of liver damage in patients with chronic hepatitisB. Hepatology 51: 81–91.
12. Hou, W., H. S. Kang, and B. S. Kim. 2009. Th17 cells enhance viral persistenceand inhibit T cell cytotoxicity in a model of chronic virus infection. J. Exp. Med.206: 313–328.
13. Harrington, L. E., R. D. Hatton, P. R. Mangan, H. Turner, T. L. Murphy,K. M. Murphy, and C. T. Weaver. 2005. Interleukin 17-producing CD4+ effectorT cells develop via a lineage distinct from the T helper type 1 and 2 lineages.Nat. Immunol. 6: 1123–1132.
14. Toh, M. L., M. Kawashima, S. Zrioual, A. Hot, P. Miossec, and P. Miossec. 2009.IL-17 inhibits human Th1 differentiation through IL-12R beta 2 downregulation.Cytokine 48: 226–230.
15. Nakae, S., Y. Iwakura, H. Suto, and S. J. Galli. 2007. Phenotypic differencesbetween Th1 and Th17 cells and negative regulation of Th1 cell differentiationby IL-17. J. Leukoc. Biol. 81: 1258–1268.
16. Wong, P., and E. G. Pamer. 2003. CD8 T cell responses to infectious pathogens.Annu. Rev. Immunol. 21: 29–70.
17. Kagi, D., F. Vignaux, B. Ledermann, K. Burki, V. Depraetere, S. Nagata,H. Hengartner, and P. Golstein. 1994. Fas and perforin pathways as majormechanisms of T cell-mediated cytotoxicity. Science 265: 528–530.
18. Martin-Orozco, N., P. Muranski, Y. Chung, X. O. Yang, T. Yamazaki, S. Lu,P. Hwu, N. P. Restifo, W. W. Overwijk, and C. Dong. 2009. T helper 17 cellspromote cytotoxic T cell activation in tumor immunity. Immunity 31: 787–798.
19. da Matta Guedes, P. M., F. R. Gutierrez, F. L. Maia, C. M. Milanezi, G. K. Silva,W. R. Pavanelli, and J. S. Silva. 2010. IL-17 produced during Trypanosoma cruziinfection plays a central role in regulating parasite-induced myocarditis. PLoSNegl. Trop. Dis. 4: e604.
20. He, D., H. Li, N. Yusuf, C. A. Elmets, J. Li, J. D. Mountz, and H. Xu. 2010. IL-17 promotes tumor development through the induction of tumor promotingmicroenvironments at tumor sites and myeloid-derived suppressor cells. J.Immunol. 184: 2281–2288.
21. Fairweather, D., S. Frisancho-Kiss, S. A. Yusung, M. A. Barrett, S. E. Davis,R. A. Steele, S. J. Gatewood, and N. R. Rose. 2005. IL-12 protects againstcoxsackievirus B3-induced myocarditis by increasing IFN-gamma and macro-phage and neutrophil populations in the heart. J. Immunol. 174: 261–269.
22. Henke, A., S. Huber, A. Stelzner, and J. L. Whitton. 1995. The role of CD8+T lymphocytes in coxsackievirus B3-induced myocarditis. J. Virol. 69: 6720–6728.
23. Yamada, T., A. Matsumori, and S. Sasayama. 1994. Therapeutic effect of anti-tumor necrosis factor-alpha antibody on the murine model of viral myocarditisinduced by encephalomyocarditis virus. Circulation 89: 846–851.
24. Huber, S. A., J. Polgar, P. Schultheiss, and P. Schwimmbeck. 1994. Augmen-tation of pathogenesis of coxsackievirus B3 infections in mice by exogenousadministration of interleukin-1 and interleukin-2. J. Virol. 68: 195–206.
4010 Th17 CELLS AND CVB3 REPLICATION IN AVMC
by guest on June 5, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
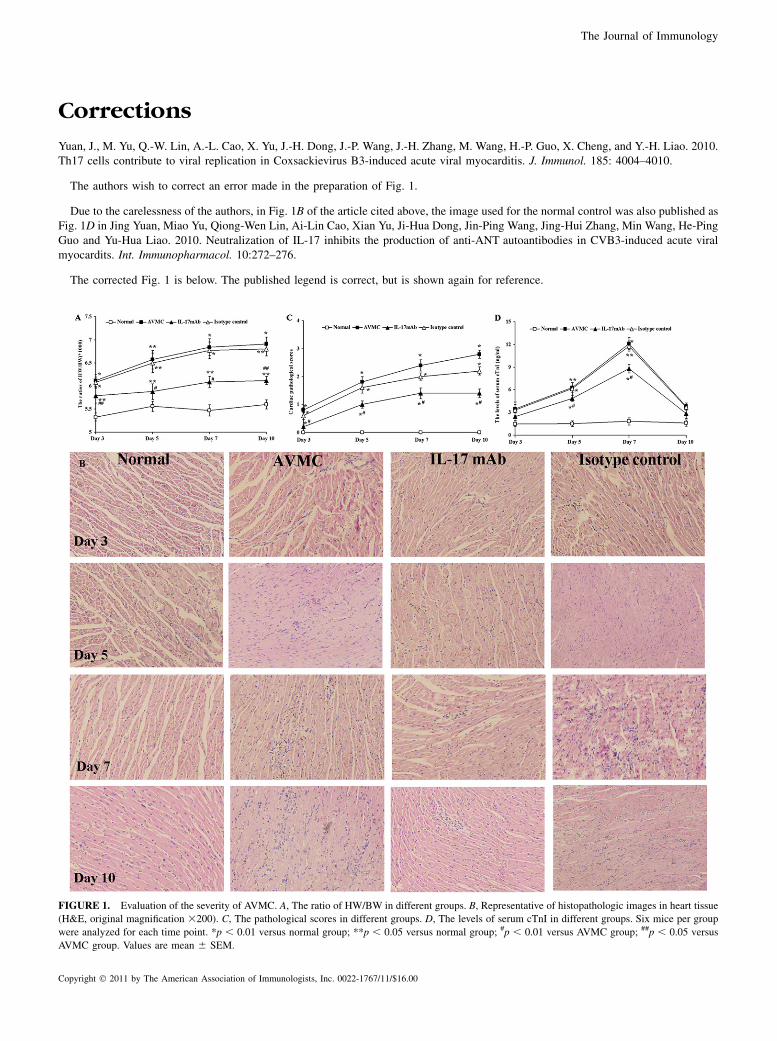
Corrections
Yuan, J., M. Yu, Q.-W. Lin, A.-L. Cao, X. Yu, J.-H. Dong, J.-P. Wang, J.-H. Zhang, M. Wang, H.-P. Guo, X. Cheng, and Y.-H. Liao. 2010.Th17 cells contribute to viral replication in Coxsackievirus B3-induced acute viral myocarditis. J. Immunol. 185: 4004–4010.
The authors wish to correct an error made in the preparation of Fig. 1.
Due to the carelessness of the authors, in Fig. 1B of the article cited above, the image used for the normal control was also published asFig. 1D in Jing Yuan, Miao Yu, Qiong-Wen Lin, Ai-Lin Cao, Xian Yu, Ji-Hua Dong, Jin-Ping Wang, Jing-Hui Zhang, Min Wang, He-PingGuo and Yu-Hua Liao. 2010. Neutralization of IL-17 inhibits the production of anti-ANT autoantibodies in CVB3-induced acute viralmyocardits. Int. Immunopharmacol. 10:272–276.
The corrected Fig. 1 is below. The published legend is correct, but is shown again for reference.
FIGURE 1. Evaluation of the severity of AVMC. A, The ratio of HW/BW in different groups. B, Representative of histopathologic images in heart tissue
(H&E, original magnification 3200). C, The pathological scores in different groups. D, The levels of serum cTnI in different groups. Six mice per group
were analyzed for each time point. *p , 0.01 versus normal group; **p , 0.05 versus normal group; #p , 0.01 versus AVMC group; ##p , 0.05 versus
AVMC group. Values are mean 6 SEM.
Copyright � 2011 by The American Association of Immunologists, Inc. 0022-1767/11/$16.00
The Journal of Immunology

In addition, the strain of virus used was the CVB3m strain (CCTCC GDV115), a mutant of Coxsackie virus B3 (Nancy) strain, asdescribed previously (1, 2).
The authors regret the duplicate publication of this figure and apologize to the scientific community for the need to publish thiscorrection.
References1. Lee, C., E. Maull, N. Chapman, S. Tracy, J. Wood, and C. Gauntt. 1997. Generation of an infectious cDNA of a highly cardiovirulent coxsackievirus B3(CVB3m) and
comparison to other infectious CVB3 cDNAs. Virus Res. 50: 225–235.2. Gauntt, C. J., M. D. Trousdale, D. R. LaBadie, R. E. Paque, and T. Nealon. 1979. Properties of coxsackievirus B3 variants which are amyocarditic or myocarditic for mice.
J. Med. Virol. 3: 207–220.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1190050
3452 CORRECTIONS

Corrections
Yuan, J., M. Yu, Q.-W Lin, A.-L. Cao, X. Yu, J.-H. Dong, J.-P. Wang, J.-H. Zhang, M. Wang, H.-P. Guo, X. Cheng, and Y.-H. Liao. 2010.Th17 cells contribute to viral replication in coxsackievirus B3-induced acute viral myocarditis. J. Immunol. 185: 4004–4010.
The representative FACS plots of Fig. 4Ab (AVMC and Isotype control groups) and Fig. 4Ba (AVMC group) on Day 7 wereincorrect. This correction does not influence the interpretation of the results or the conclusions. The correct figure is published below.The entire figure is shown, but the only change is to the plots referenced above. The legend is correct as published and is also shown belowfor reference.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1490045
Copyright � 2014 by The American Association of Immunologists, Inc. 0022-1767/14/$16.00
The Journal of Immunology
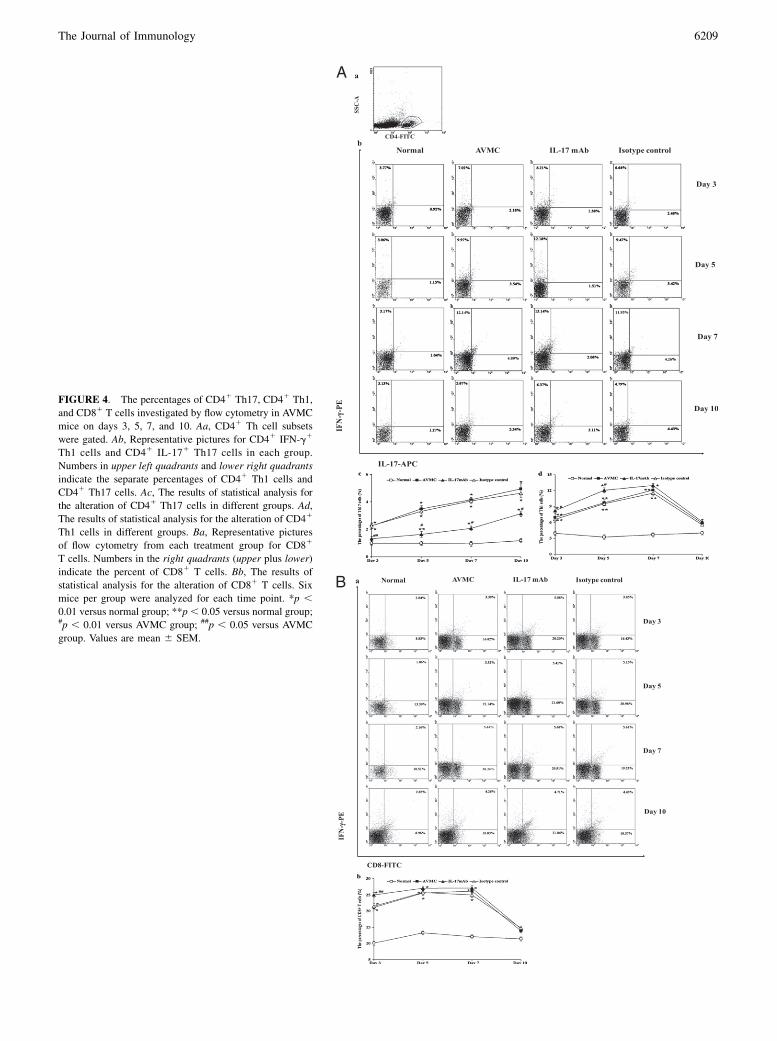
FIGURE 4. The percentages of CD41 Th17, CD41 Th1,
and CD81 T cells investigated by flow cytometry in AVMC
mice on days 3, 5, 7, and 10. Aa, CD41 Th cell subsets
were gated. Ab, Representative pictures for CD41 IFN-g1
Th1 cells and CD41 IL-171 Th17 cells in each group.
Numbers in upper left quadrants and lower right quadrants
indicate the separate percentages of CD41 Th1 cells and
CD41 Th17 cells. Ac, The results of statistical analysis for
the alteration of CD41 Th17 cells in different groups. Ad,
The results of statistical analysis for the alteration of CD41
Th1 cells in different groups. Ba, Representative pictures
of flow cytometry from each treatment group for CD81
T cells. Numbers in the right quadrants (upper plus lower)
indicate the percent of CD81 T cells. Bb, The results of
statistical analysis for the alteration of CD81 T cells. Six
mice per group were analyzed for each time point. *p ,0.01 versus normal group; **p, 0.05 versus normal group;#p , 0.01 versus AVMC group; ##p , 0.05 versus AVMC
group. Values are mean 6 SEM.
The Journal of Immunology 6209