temporal patterns of milk production in antarctic fur seals (arctocephalus gazella)
TRANSCRIPT

J. Zool., Lond. (1 995) 237, 1 - 12
Temporal patterns of milk production in Antarctic fur seals (Arctocephalus gazellu)
J . P. Y. ARNOULD AND I . L. BOYD British Antarctic Survey, Natural Environment Research Council, High Cross, Madingley Road,
Cambridge CB3 OET, UK
(Accepted 21 June 1994)
(With 7 figures in the text)
The timing of milk production in Antarctic fur seals was studied at Bird Island, South Georgia. Like all lactating otariid seals (Pinnipedia: Otariidae), Antarctic fur seal females alternate between short nursing periods ashore and regular foraging trips to sea. Females do not necessarily return to the colony with full mammae, which indicates that mammary volume capacity is unlikely to limit foraging trip duration. Upon arrival at the colony, milk fat (r2 = 0.33, P < 0.04) and protein (2 = 0.60, P < 0.002) content were positively correlated to the time spent at sea. A similar trend was observed in the milk produced on land. The rate of milk energy production was much lower at sea (5.02 f. 0.05 MJ . day-’) than on land (23.66 f 4.4 MJ . day-’). The rate of milk energy production during the foraging trip was negatively correlated to the time spent at sea (2 = 0.29, P < 0.05), whereas the rate of milk energy production on land was positively correlated (rZ = 0.61, P < 0.001) to the duration of the preceding foraging trip. The total amount of milk energy delivered to the pup during each two- day nursing period was positively correlated (rZ = 0.60, P < 0.002) to the duration of the previous foraging trip. The overall rate of milk energy delivery, however, was independent of foraging trip duration. This accords with previous observations that the growth rates of Antarctic fur seal pups are unaffected by maternal foraging trip duration patterns.
Contents Page
Introduction . . . . . . , . . . . . . . . . . . . . . . . . . . . . . . . . 1 Methods . . . . . . . . . . . . . . . . , , . . . . . . . . . . . . . . . . 2 Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 Discussion.. . . . . . . . . . . . a . . . . . . . . . . . . . . . . . . . . 9
Sampling procedures . . . . , . . . . . . . . . . . . . . . . . . . . . . . 9 Milk production . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
References.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
Introduction
To meet the energy and nutrient demands of milk production, maternal food consumption must be increased before and/or during lactation (Sadleir, 1984). During lactation, however, constraints may be placed on maternal foraging behaviour by the offspring. In placental mammals where the dependent offspring remain with the mother while she is foraging (e.g. ‘follower’ ungulates, primates), predator awidance may reduce foraging efficiency as mothers spend more time being vigilant and/or consume food of lower quality in areas of less danger from predators (Sadleir, 1969; Carl & Robbins, 1988). In contrast, in species where mothers forage
1 0 1995 The Zoological Society of London

2 J. P. Y. ARNOULD AND I. L. BOYD
alone and return to the offspring periodically to nurse (e.g. 'hider' ungulates, carnivores, rodents, lagomorphs, chiropterans), mothers may forage unhindered by the dependent offspring and thus have greater access to potential food sources (Broekhuizen & Maaskamp, 1980; Sadleir, 1984; Gittleman, 1988; Gittleman & Thompson, 1988). Transporting milk and/or nutrients for its production, however, incurs an energetic cost as well as a reduction in the time available for foraging (Gentry et al., 1986). Females may be limited in how long they are able to search for food by how much milk and nutrients they are physically capable of carrying and/or by the fasting endurance of their offspring.
Otariid seals (Pinnipedia: Otariidae) have a lactation pattern characterized by short nursing periods alternating with regular foraging trips to sea (Bonner, 1984). Food abundance is patchy in the marine environment, so that seals may have to travel long distances in search of prey, during which time their pups must fast on shore, consuming body reserves for maintenance instead of growth (Gentry & Kooyman, 1986; Gentry et al., 1986). Therefore, to maximize pup growth rate and survivorship, females must balance the amount of time spent in search of food for milk production with the amount of milk energy produced and time spent delivering it.
Recent studies on the Antarctic fur seal (Arctocephalus gazella) at South Georgia have found that foraging trip duration patterns can vary substantially between individuals, with some females consistently undertaking long trips (6-8 days) whereas others regularly make short trips (2-4 days) (Boyd, Lunn & Barton, 1991; Lunn et al., 1993). Despite the differences in foraging trip durations, there are no apparent differences in pup growth rates (Boyd, Lunn & Barton, 1991; Lunn et al., 1993), which suggests that the overall delivery rates of milk energy to the offspring must be similar.
To improve the understanding of the mechanisms that may control relationships between foraging cycle duration and pup growth rates in Antarctic fur seals, we studied the temporal patterns of milk production in this species. Our objectives were to determine: (1) whether foraging trip duration is dependent on or limited by mammary volume capacity (i.e. do females terminate a foraging trip when their mammary glands are full?); (2) the extent to which milk production varies in terms of rate and composition (e.g. fat and protein content); and (3) what maternal characteristics (e.g. body mass, foraging trip duration) influence production.
Methods
The study was conducted at Bird Island (54" 00' S, 38" 02' W), South Georgia, between 21 December 1992 and 20 February 1993. Sixteen females returning from foraging trips of known durations were captured upon arrival at the colony, using standard noose-pole and restraint board techniques (Gentry & Holt, 1982). as they rejoined their pups. They were lightly anaesthetized with a 1 : 1 combination of tiletamine hydrochloride and zolazepam hydrochloride (ZoletilB, Laboratories Vibrac, B.P. 13 0602 1 Nice, Cedex, France) to reduce possible stress effects during the prolonged handling period (Boyd et a/., 1990). Each female was weighed with a spring scale (100 kg f 0.5 kg) and straight-line length was measured to the nearest centimetre with a tape measure. Complete mammary evacuation was then achieved by manual expression of milk from each nipple over a period of 40-60 min with the aid of repeated intramuscular oxytocin injections (0.5-1.0m1, l0 iu . ml-I). During the course of the milking there was a general decrease in the amount of milk which could be expressed and, although oxytocin injections would temporarily reverse this trend, the overall decrease continued. The mammae were, therefore, considered empty if no more milk could be expressed after 2 consecutive injections of oxytocin administered at 10-min intervals. Females and pups were then kept in adjacent enclosures where they could maintain visual, vocal and olfactory contact but could not suckle. Sixteen to 24 h later, females were lightly anaesthetized once more and weighed, and complete

MILK PRODUCTION IN ANTARCTIC FUR SEALS 3
mammary evacuation was achieved by the same procedures as before. Females and pups were released after the female had recovered fully from the anaesthetic (<6 h).
The pups were weighed on a spring balance (20 kg f 0.1 kg) and sexed at the beginning of the study. The volume of milk collected at each sampling was measured and sub-sampled for gross composition analysis before being fed to the pup via a stomach tube. Sub-samples were stored in plastic vials at -20 "C until analysed.
Before all analyses, the frozen milk samples were thawed at room temperature (20 "C) for 12 h and mixed thoroughly with a high-speed stirring rod. All analyses were run in duplicate. Water and ash content were estimated by standard techniques. Water content was determined by drying sub-samples (c. 1 g f 1 mg) in pre-weighed aluminium foil trays for 24 h at 70 "C in an oven and then for a further 24 h at 70 "C in a vacuum oven. The dried samples were then cooled in a desiccator before being weighed again. Ash content was determined by placing sub-samples (c. 1 g f 1 mg) in ceramic crucibles and drying them as for analysis of dry mass. They were then heated in a furnace at: (I) 200 "C for 6 h; (2) 450 "C for 18 h; and (3) 550 "C for 18 h. The samples were then cooled in a desiccator before weighing.
Protein and lipid content were determined by a stoichiometric method (Gnaiger & Bitterlich, 1984; Arnould, Boyd & Clark, In press). Sub-samples (1-2 mg f 0.001 mg) of the dried mass from the water content analysis were packaged into pre-weighed tin boats and stored in a desiccator until analysed on an automatic CHN elemental analyser (Carlo 1106 Elemental analyser) using acetanilide as a standard. The measured C, H and N proportions of dry mass were used to calculate protein and lipid content following the procedures in Gnaiger & Bitterlich (l984), modified to account for the specific characteristics of Antarctic fur seal milk (Arnould et al., In press). Carbohydrates typically represent < 1 % of the total milk volume, contributing to <0.2% of the gross energy (Oftedal, 1984; Oftedal, Boness & Tedman, 1987). Consequently, gross energy content (kJ . g-') of the milk was calculated by multiplying the derived chemical composition by standard values for the energy density of lipid (38.12 kJ . g-') and protein (23.64 kJ . g-') (Perrin, 1958). Chemical composition could not be determined for all the milk volumes collected owing to the loss of 4 samples during storage.
For each individual, the rate of milk volume production while at sea was defined as the volume of milk collected as it came ashore divided by the duration of the preceding foraging trip. The rate of production on land was defined as the volume of milk collected in the second milking divided by the time interval since the first milking.
To estimate how much milk the study females would have produced during a normal attendance period had they not been handled, it was necessary to evaluate the natural timing of milk transfer in this species. Fourteen randomly selected mother-pup pairs (pups: 8 female, 6 male) were captured as they reunited when the female returned to the colony from a foraging trip. The pups were sexed, weighed and individually numbered by bleaching the dorsal fur with peroxide dye (Clairol Born Blonde@, Bristol-Myers Company Ltd, Ickenham, Uxbridge, UK). To facilitate serial weighing of the pups, mothers and pups were kept together in an enclosure where they could suckle normally. The enclosure was surrounded by a raised gantry allowing the pups to be caught easily from above, with minimal disturbance, using a long noose-pole. Pups were weighed as 8-h intervals for 3 days before being released with their mothers.
Results
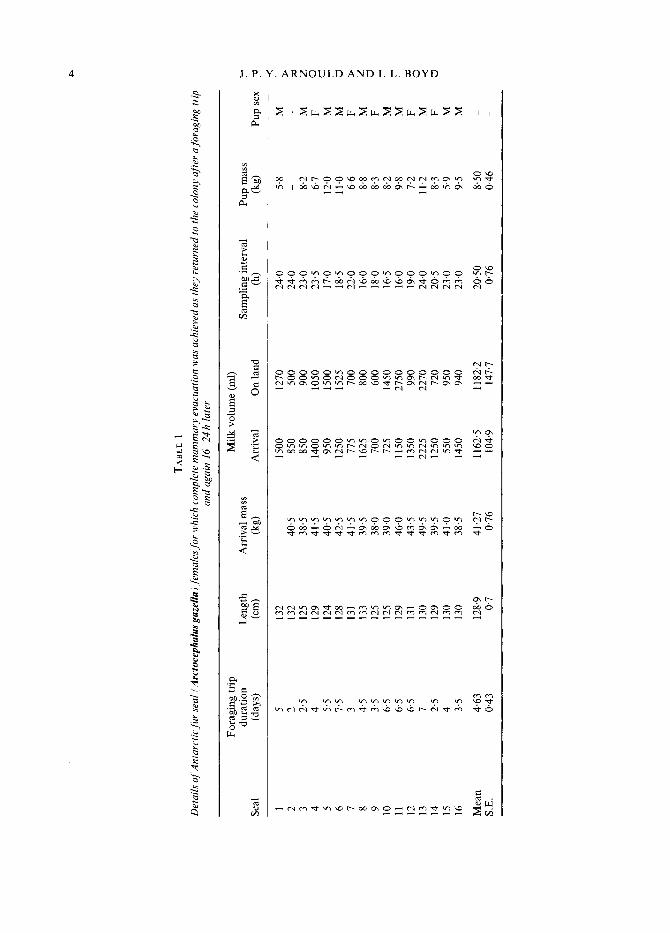
The foraging trip duration, milk volumes obtained and maternal characteristics of the individual females are presented in Table I. The mean foraging trip duration was 4.63 f 0.43 days and females on average returned to the colony with 1.162 f 0.105 litres of milk in their mammae. In seven (44%) of the 16 females sampled, the volume of milk obtained after they had been ashore for 16-24 hours was greater (by up to 200% in one case) than the volume initially carried ashore (Fig. 1). This shows that females do not necessarily return to the colony (i.e. terminate a foraging trip) only when their mammae are full. The volume of milk females carried
P
TA
BL
E
1 D
etai
ls o
f An
tarc
ticju
r se
al (
Arc
toce
phal
us ga
rella
) fem
ales
fiv
whi
ch c
ompl
ete
mam
mar
y ev
acua
tion
was
acl
iiew
d as
th
q re
turn
ed to
the
colo
ny a
fier
a,fo
ragi
ng tr
ip
and
agai
n 16
-24
h la
ter
Fora
ging
trip
M
ilk v
olum
e (m
l) du
ratio
n Le
ngth
A
rriv
al m
ass
Sam
plin
g in
terv
al
Pup
mas
s Se
al
(day
s)
(4
(kg)
A
rriv
al
On
land
(h
) (k
g)
Pup
sex
1 2 3 4 5 6 7 8 9 10
11
12
13
14
15
16
Mea
n S.
E.
5 2 2.5
4 5.5
7.5
3 4.5
3.5
6.5
6.5
6.5
7 2.5
4 3.5
4.63
0.
43
I32
I32
125
129
124
128
131
133
125
125
129
131
130
I29
130
130
128.
9 0.
7
40.5
38
.5
41.5
40
.5
42.5
41
.5
39.5
38
.0
39.0
46
.0
43.5
49
.5
39.5
41
.0
38.5
41.2
7 0.
76
I500
85
0 85
0 14
00
950
1250
77
5 I6
25
700
725
1150
13
50
2225
12
50
550
1450
1 162
5 10
4.9
1270
50
0 90
0 10
50
1500
15
25
700
800
600
1450
27
50
990
2270
72
0 95
0 94
0
11 82
.2
147.
7
244
24.0
23
.0
23.5
17
.0
18.5
22
.0
16.0
18
.0
16.5
16
.0
19.0
24
.0
20.5
23
.0
23.0
20.5
0 0.
76
5.8
8.2
6.7
12.0
11
.0
6.6
8.8
8.3
8.2
9.8
7.2
11.2
8.
3 5.
9 9.
5
8.50
0.
46
-
M
M
F M
M
F M
F M
M
F M
F
M
M
c.
?J
4
9
?J z 0
C
r
U 9 z 0
r
0
4
U
c(
W
MILK PRODUCTION I N ANTARCTIC F U R SEALS
2500-
2ooo-
1500- E v
- 3 B 1000- Y .-
500-
5
o Female pup
0 Pup sex unknown 0 Male pup 0 r2=0.15,P>0.1
0 0 O .
O.oo 0
0 0
0
0 Carried ashore - 16-24 h later 2500
.-. 40- 7
E 30-
F
z H
a,
5 20- ._
10-
a,
- f 1500 B z 1000 2
500
0 i 2 3 4 Li 6 7 /)I 3 9 10 I
2 1 3 14 1 5 16 Seal
FIG. 1 . Volume of milk obtained when completely evacuating the mammae of Antarctic fur seal females as they returned to the colony from foraging trips and again 16-24 h later.
0 1 I I I I I I I I 0 1 2 3 4 5 6 7 8
Foraging trip duration (days)
50 1 I 0
r ‘=0.33, P <0.04
~ 0
0
0 1 I I I I I I I I
0 1 2 3 4 5 6 7 8 Foraging trip duration (days)
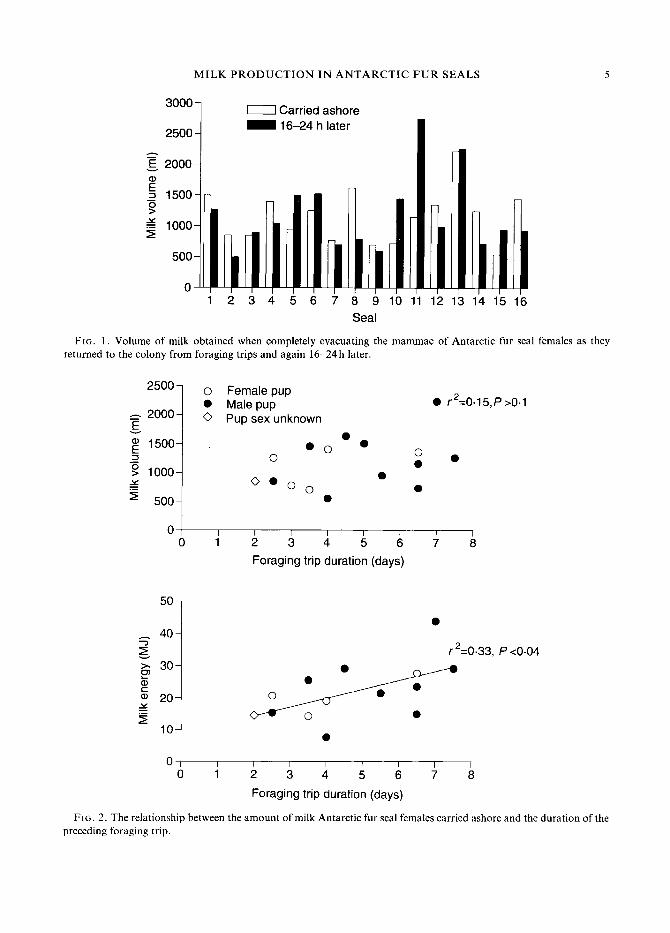
F I G . 2. The relationship between the amount of milk Antarctic fur seal females carried ashore and the duration of the preceding foraging trip.
6 J. P. Y . A R N O U L D A N D I. L. BOYD
20 -
10
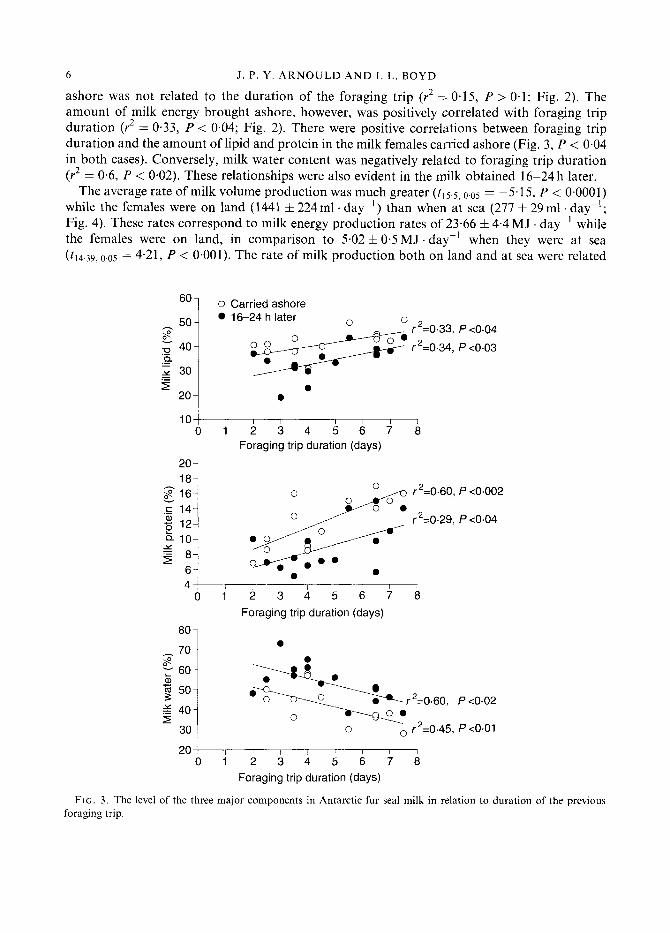
ashore was not related to the duration of the foraging trip (r2 =- 0.15, P > 0.1; Fig. 2). The amount of milk energy brought ashore, however, was positively correlated with foraging trip duration (r2 = 0.33, P < 0.04; Fig. 2). There were positive correlations between foraging trip duration and the amount of lipid and protein in the milk females carried ashore (Fig. 3, P < 0.04 in both cases). Conversely, milk water content was negatively related to foraging trip duration (r2 = 0.6, P < 0.02). These relationships were also evident in the milk obtained 16-24h later.
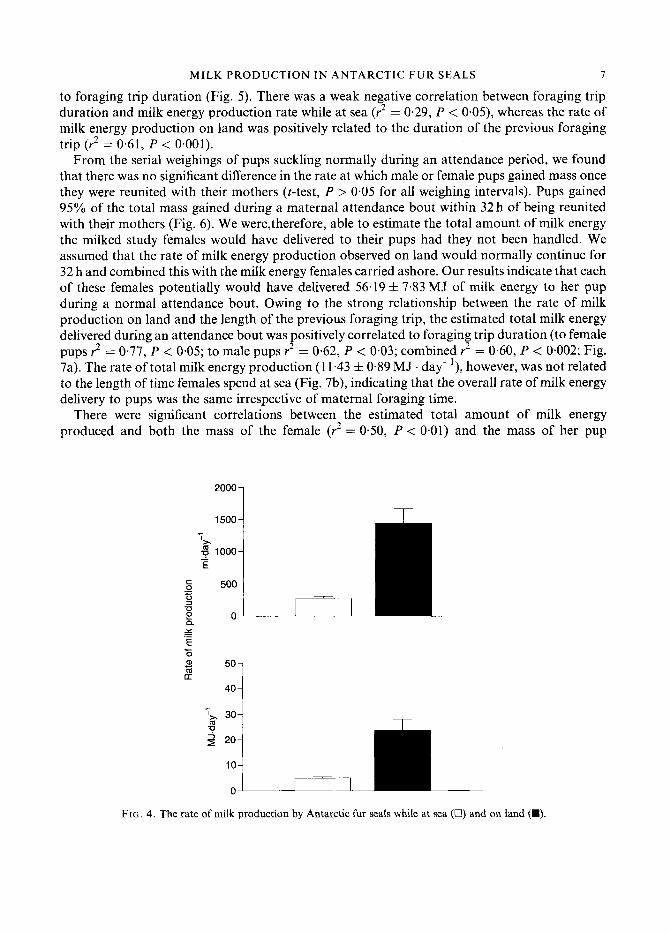
The average rate of milk volume production was much greater ( t I 5 5 , 0.05 = -5.1 5, P < 0,0001) while the females were on land (1441 f 224ml. day-') than when at sea (277 f 29 ml . day-'; Fig. 4). These rates correspond to milk energy production rates of 23.66 f 4.4 MJ . day-' while the females were on land, in comparison to 5.02 f 0.5MJ. day-] when they were at sea (t14.39, 0 0 5 = 4.21, P < 0.001). The rate of milk production both on land and at sea were related
0
1 I I I I I I I
601 o Carried ashore =,, 16-24 h later
70- 8
60-
fii 50- 3 40-
3 30 -
a,
I -- 0 1 2 3 4 5 6 7 8
Foraging trip duration (days)
8o 1
20- I I I I I 0 1 2 3 4 5 6 7 8
Foraging trip duration (days)
FIG. 3 . The level of the three major components in Antarctic fur seal milk in relation to duration of the previous foraging trip.
M I L K PRODUCTION IN ANTARCTIC FUR SEALS
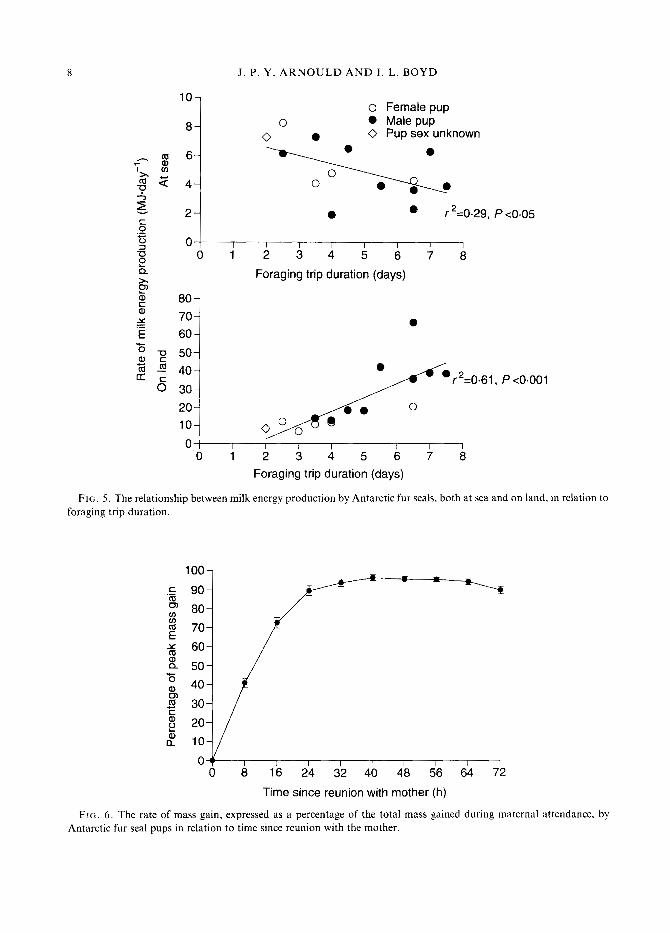
to foraging trip duration (Fig. 5). There was a weak negative correlation between foraging trip duration and milk energy production rate while at sea (r2 = 0.29, P < 0.05), whereas the rate of milk energy production on land was positively related to the duration of the previous foraging trip (r2 = 0.61, P < 0.001).
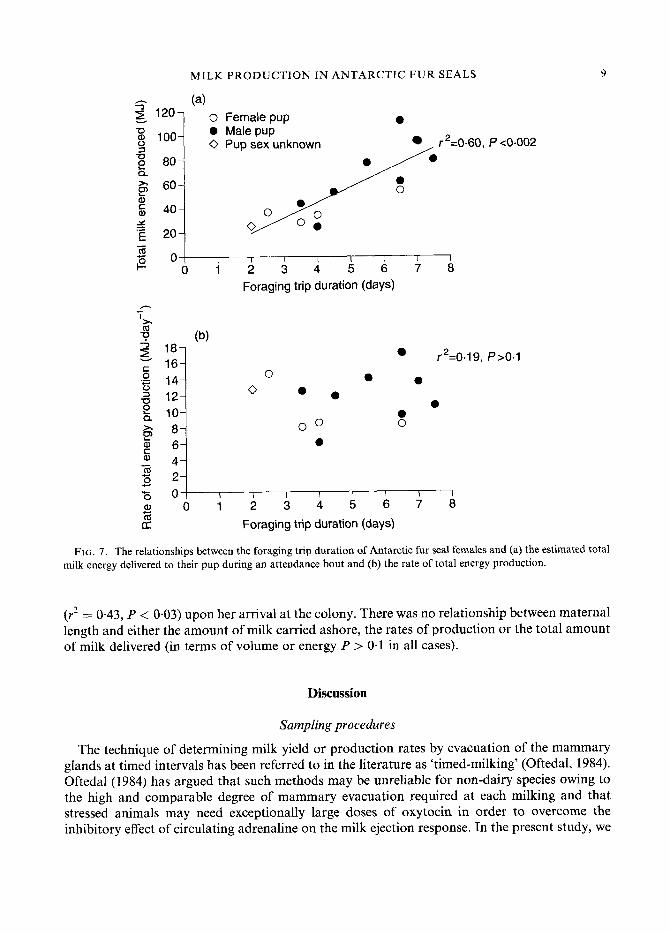
From the serial weighings of pups suckling normally during an attendance period, we found that there was no significant difference in the rate at which male or female pups gained mass once they were reunited with their mothers (t-test, P > 0.05 for all weighing intervals). Pups gained 95% of the total mass gained during a maternal attendance bout within 32 h of being reunited with their mothers (Fig. 6). We were,therefore, able to estimate the total amount of milk energy the milked study females would have delivered to their pups had they not been handled. We assumed that the rate of milk energy production observed on land would normally continue for 32 h and combined this with the milk energy females carried ashore. Our results indicate that each of these females potentially would have delivered 56.19 f. 7.83 MJ of milk energy to her pup during a normal attendance bout. Owing to the strong relationship between the rate of milk production on land and the length of the previous foraging trip, the estimated total milk energy delivered during an attendance bout was positively correlated to foraging trip duration (to female pups r2 = 0.77, P < 0.05; to male pups r2 = 0.62, P < 0.03; combined r2 = 0.60, P < 0.002; Fig. 7a). The rate of total milk energy production (1 1.43 f 0.89 MJ . day-'), however, was not related to the length of time females spend at sea (Fig. 7b), indicating that the overall rate of milk energy delivery to pups was the same irrespective of maternal foraging time.
There were significant correlations between the estimated total amount of milk energy produced and both the mass of the female (r2 = 0.50, P < 0.01) and the mass of her pup
I
*Oo0 1 I T
Y E 0 al m U
._ c
501 c
T
0
FIG. 4. The rate of milk production by Antarctic fur seals while at sea (U) and on land (M).
8
C 0 .- -
J. P. Y . A R N O U L D A N D I . L. B O Y D
a, 80 - a, 70- Y E 60 -
u 50-
C
.- - a , K c
IT _m 6 40- 30-
20 - 10-
0
l01
0 / r2=0.61, P <0.001
0 0 I I I I I I I I
.i o Female pup
0 0 Male pup 0 0 Pup sex unknown 0
0 r2=0.29, P<0.05
O! I I I I I I I I I 0 8 16 24 32 40 48 56 64 72
Time since reunion with mother (h)
FIG. 6 . The rate of mass gain, expressed as a percentage of the total mass gained during maternal attendancc. by Antarctic fur seal pups in relation to time since reunion with the mother.
MILK PRODUCTION IN ANTARCTIC FUR SEALS
(a) 0 Femalepup 0 0 Male pup 0 Pup sex unknown r2=0.60, P ~0.002
v
-0
3 -0
Q
$ 100-
g 80-
3h 60-
5 40- Y 'E 20-
9
o:, e 0 1 2 3 4 5 6 7 8 Foraging trip duration (days)
r*=0.19, Pr0.1
0 0 0
0 0 0 0
0 0 0 0
0
5 0 1 I I I I I I I I .I- a 0 1 2 3 4 5 6 7 8 CT Foraging trip duration (days) a
FIG. 7. The relationships between the foraging trip duration of Antarctic fur seal females and (a) the estimated total milk energy delivered to their pup during an attendance bout and (b) the rate of total energy production.
(r2 = 0.43, P < 0.03) upon her arrival at the colony. There was no relationship between maternal length and either the amount of milk carried ashore, the rates of production or the total amount of milk delivered (in terms of volume or energy P > 0.1 in all cases).
Discussion
Sampling procedures
The technique of determining milk yield or production rates by evacuation of the mammary glands at timed intervals has been referred to in the literature as 'timed-milking' (Oftedal, 1984). Oftedal (1 984) has argued that such methods may be unreliable for non-dairy species owing to the high and comparable degree of mammary evacuation required at each milking and that stressed animals may need exceptionally large doses of oxytocin in order to overcome the inhibitory effect of circulating adrenaline on the milk ejection response. In the present study, we

10 J . P. Y . A R N O U L D A N D 1. L. BOYD
milked the animals while they were lightly anaesthetized so as to reduce any possible stress effects and used a standard criterion to determine complete mammary evacuation. The efficiency of this sampling protocol was verified when, in inspecting a female killed for another study, after 550 ml of milk were obtained by manual expression in the usual manner, we could only find 10 ml in the dissected mammae.
Repeated and high doses of exogenous oxytocin have been shown to affect milk lactose content (Linzell, 1967; Lane et al., 1970) but not lipid or protein content (Ballou et al., 1993). Pinniped milks typically contain negligible amounts of carbohydrate (Oftedal et al., 1987) and the lipid and protein contents observed in this study were within the range previously reported for this species (Costa, 1991; Arnould & Boyd, In press). The observed decrease in lipid and protein content between the milk females carried ashore and that collected a day later is similar to the differences observed between females arriving and departing from the colony in this and other otariid species (Costa & Gentry, 1986; Arnould & Boyd, In press). It appears unlikely, therefore, that our sampling procedures would have biased the observed milk composition.
The validity of our sampling procedure is further supported by the close agreement between our estimate of average milk energy delivered per attendance bout (56.19 MJ), based upon production rates, and that found by Costa (1991) from isotopic measurements of pup milk consumption (63.7MJ, one-sample t-test 12 = 0.959, P > 0.03). Costa (1991) calculated a daily delivery rate of 10.5 MJ . day-’ for a mean foraging trip duration of 4.3 days, whereas we estimated 1 1.43 MJ . day-’ for the 4.63 day mean foraging trip we observed in this study (one- sample t-test t0.05, 12 = 1.057, P > 0.3).
Milk production
Milk lipid and protein content were positively related to foraging trip duration. Trillmich & Lechner (1986) noted that, within the Otariidae, species undertaking short trips have lower milk lipid levels than those undertaking long trips. They suggested that this relationship may be an adaptation to sustain pups during the mothers’ absence. We suggested that this pattern of concentrating milk nutrients may, more importantly, be an adaptation allowing females to maximize the efficiency of nutrient transport to their pups by reducing the load carried back to the colony. The mechanisms by which Antarctic fur seals are able to alter milk composition in this manner are not known.
The results of the present study clearly indicate that females do not necessarily return to the colony with full mammae. Hence, mammary volume capacity is unlikely to be a major factor limiting foraging trip duration. Gentry et al. (1986) postulated that fur seals may terminate foraging trips upon attaining ‘nutritional satiation’ set points. From the present data, however, it is not possible to assess whether foraging trip durations in Antarctic fur seals are determined by such ‘nutritional satiation’ set points.
The rate of milk energy production was much greater while females were ashore than while they were at sea. Consequently, 42-79% of the milk energy delivered to the pup was produced from the female’s body reserves while she was ashore fasting. The longer a female had been at sea. the greater the proportion of the total milk energy delivered that was derived from body stores rather than being carried ashore in the mammae. In addition, the rate at which this milk energy was produced on land increased with foraging trip duration. Hence, females undertaking longer foraging trips maximize the rate of nutrient and energy transport not only by producing concentrated milk but also by carrying most of the ‘milk’ as body lipid and protein stores to

MILK PRODUCTION I N ANTARCTIC F U R SEALS 11
be converted to milk on land. The rate of transfer of this milk is then maximized by increasing the rate of production, in relation to the trip duration, thereby reducing the amount of time females require to remain on shore and, hence, the ‘metabolic overhead’ they incur while fasting (Gentry et al., 1986; Costa, 1991).
When comparing otariid species, Costa (1991) noted that those undertaking short foraging trips return to their pup with less milk energy than those species spending longer periods at sea, and that this is consistent with central place foraging theory. Central place foraging theory states that animals feeding further from a central place should make fewer trips and return with greater amounts of energy per trip than those feeding closer (Orians & Pearson, 1977). In the present study, the total amount of milk energy delivered to the pup during an attendance bout was positively related to the length of the previous foraging trip, indicating that this principle is also valid for the individual variations in foraging behaviour that occur within a single species, the Antarctic fur seal. Consequently, the overall rate of milk energy delivery in Antarctic fur seals was independent of maternal foraging trip duration. This accords with the observation that growth rates of Antarctic fur seal pups, in years of normal food abundance, are unaffected by maternal foraging trip duration patterns (Lunn et al., 1993).
In summary, in comparison with those individuals undertaking short foraging trips, Antarctic fur seal females spending longer at sea deliver more milk energy to their pup during attendance on shore. They store the majority of nutrients acquired for milk production as body reserves and carry the remainder as concentrated milk. They subsequently maximize the rate of nutrient transfer to their offspring by increasing the rate of production of concentrated milk during attendance on land. The mechanisms by which Antarctic fur seal females are able to alter the rate of production, and the composition, of milk in relation to their foraging patterns are not known. While the total amount of energy delivered to the pup during an attendance bout is positively related to foraging trip duration, the overall rate of energy delivery is the same regardless of maternal foraging patterns. Therefore, from the pup’s perspective, there does not appear to be a selective advantage in having a mother that follows a particular pattern of foraging trip duration. It is, however, unknown whether the foraging strategies that females employ give rise to any intrinsic differences in the energetic costs that they incur, and affect their long-term reproductive success.
We thank A. Barnes and J. Cooper for their assistance with the chemical analyses of the milk samples and are grateful to J. P. Croxall, M. A. Fedak and an anonymous reviewer for constructive criticism of this manuscript.
REFERENCES
Arnould, J. P.Y. & Boyd, I. L. (In press). Inter- and intra-annual variation in milk composition in Antarctic fur seals
Arnould, J. P. Y., Boyd, I.L. & Clarke, A. (In press). A simplified method for determining the gross chemical composition
Ballou, L. U., Bleck, J. L., Bleck, G. T. & Bremel, R. D. (1993). The effects of daily oxytocin injections before and after
Bonner, W. N. (1984). Lactation strategies in pinnipeds: problems for a marine mammalian group. Symp. zool. SOC. Lond.
Boyd, I. L., Lunn, N. J. & Barton, T. (1991). Time budgets and foraging characteristics of lactating Antarctic fur seals.
Boyd, I. L., Lunn, N. J., Duck, C. D. & Barton, T. (1990). Response of Antarctic fur seals to immobilization with
(Arctocephalus gazella). Physiol. Zool.
of pinniped milk samples. Can. J . 2001.
milking on milk production, milk plasmin, and milk composition. J. Dairy Sci. 7 6 1544-1549.
NO. 51: 253-272.
J. Anim. Ecol. 60: 577-592.
ketamine, a ketamine-diazepam or ketamine-xylazine mixture, and Zoletil@. Mar. Mammal. Sci. 6: 135- 145.

12 J. P . Y . ARNOULD A N D I. L. BOYD
Broekhuizen, S. & Maaskamp, F. (1980). Behaviour of does and leverets of the European hare (Lepus europaeus) whilst nursing. J . Zool., Lond. 191: 487-501.
Carl, G. R. & Robbins, C. T. (1988). The energetic cost of predator avoidance in neonatal ungulates: hiding versus following. Can. J. Zool. 66: 239-246.
Costa, D. P. (1991). Reproductive and foraging energetics of pinnipeds: implications for life history patterns. In B e h i o u r ofpinnipeds: 300-344. Renouf, D. (Ed.). London: Chapman and Hall.
Costa, D. P. & Gentry, R. L. (1986). Free-ranging energetics of northern fur seals. In Fur seals: maternal strategies on land and at sea: 79-101. Gentry, R. L. & Kooyman, G. L. (Eds). Princeton: Princeton University Press.
Gentry, R. L., Costa, D. P., Croxall, J. P., David, J. H. M., Davis, R. W., Kooyman, G. L., Majluf, P., McCann, T. S. & Trillmich, F. (1986). Synthesis and conclusions. In Fur seals: maternal strategies on land and ut sra: 220-264. Gentry, R.L. & Kooyman, G. L. (Eds). Princeton: Princeton University Press.
Gentry, R. L. & Holt, J. R. (1982). Equipment and techniques for handling northern fur seals. NOAA. Tech. Rep. NMFS spec. scient. Rep. Fish No. 758: 1-16.
Gentry, R. L. & Kooyman, G. L. (1986). Introduction. In Fur seals: maternalstrategies on land and at scv: 3-27. Gentry. R. L. & Kooyman, G. L. (Eds). Princeton: Princeton University Press.
Gittleman, J. L. (1988). Behavioural energetics of lactation in a herbivorous carnivore, the red panda (Ailuru.s,/ulgen.s).
Gittleman, J. L. & Thompson, S. D. (1988). Energy allocation in mammalian reproduction. Am. Zool. 28: 863-875. Gnaiger, E. & Bitterlich, G. (1984). Proximate biochemical composition and caloric content calculated from elemental
CHN analysis: a stoichiometric concept. Oecologia 6 2 289-298. Lane, G. T., Dill, C. W., Armstrong, B. C. & Switzer, L. A. (1970). Influence of repeated oxytocin injections on
composition of dairy cows’ milk. J . Dairy Sci. 53: 427-429. Linzell, J. L. (1967). The effect of very frequent milking and of oxytocin on the yield and composition of milk in fed and
fasted goats. J. Physiol. 190: 333-346. Lunn, N. J.. Boyd, I. L., Barton, T. & Croxall, J. P. (1993). Factors affecting the growth rate and weaning mass of
Antarctic fur seal pups at Bird Island, South Georgia. J . Mammal. 7 4 908-919. Oftedal, 0. T. (1984). Milk consumption, milk yield and energy output at peak lactation: a comparative review. Symp.
zool. Sue. Lond. No. 51: 33-85. Oftedal, 0. T., Boness, D. J. & Tedman, R. A. (1987). The behaviour, physiology, and anatomy of lactation in the
Pinnipedia. Curr. Mammal. 1: 175-245. Orians, G. H. & Pearson, N. E. (1977). On the theory of central place foraging. In Analysis of ecologicalsystems: 153-177.
Horn, D. J., Stairs, G. R. & Mitchell, R. D. (Eds). Columbus: Ohio State University Press. Perrin, D. R. (1958). The calorific value of milk of different species. J. Dairy Res. 2 5 215-220. Sadleir, R. M. F. S. (1969). The ecology of reproduction in wild and domestic mammals. London: Methuen. Sadleir, R. M. F. S. (1984). Ecological consequences of lactation. Acta. zool. fenn. No. 171: 179-182. Trillmich, F. & Lechner, E. (1986). Milk of the Galapagos fur seal and sea lion, with a comparison of the milk of eared
Ethology 7 9 13-24.
seals (Otariidae). J . Zool., Lond. ( A ) 209 281-277.