synaptic relationships involving local axon collaterals of pyramidal neurons in the cat motor cortex
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 336~229-242 (1993)
Synaptic Relationships Involving Local Axon Collaterals of Pyramidal Neurons
in the Cat Motor Cortex
ASAF KELLER AND HIROSHI ASANUMA Department of Anatomy and Cell Biology and Program in Neuroscience,
Uniformed Services University of the Health Sciences, Bethesda, Maryland 20814 (A.K.); The Rockefeller University, New York, New York 10021 (H.A.)
ABSTRACT The intracortical synaptic relationships of pyramidal neurons in the cat motor cortex were
studied by intracellular recording and labeling techniques. Neurons that responded with monosynaptic excitatory postsynaptic potentials (EPSPs) to microstimulation in the somatosen- sory cortex were identified by intracellular recordings. Long-term potentiation (LTP) was evoked in all of these neurons (n = 151, following tetanic stimulation (50 Hz, 5 s) of their afferents from the somatosensory cortex. Three of these cells (cells A-C) were identified as pyramidal neurons, following intracellular injections of Neurobiotin. The intracortical axon collaterals of these labeled cells arborized extensively, forming terminal clusters both in close proximity to the parent soma and along their long, horizontal branches. Terminal clusters in both the proximal and in the distal termination zones of each of the cells were studied by electron microscopy. In their proximal arborization zones, the axon collaterals of the labeled pyramidal neurons synapsed preferentially with dendritic spines belonging to other pyramidal cells. In contrast, in their distal terminal clusters, the axon collaterals of each of the cells formed synapses in different proportions with different postsynaptic targets. The distal axon collaterals of cell A formed 86% of their synapses with pyramidal neurons; those of cell B formed 64% of their synapses with pyramidal cells, the remaining synapses with the dendritic shafts and somata of nonpyramidal neurons, and those of cell C provided most of their output (68%) to nonpyramidal, presumably inhibitory neurons. These findings suggest a high selectivity of intrinsic axon collaterals to form specific patterns of synapses. The patterns of synaptic interactions formed by these intrinsic axon collaterals may be a substrate for shaping and modulating representation maps in the motor cortex.
Key words: intrinsic synaptic circuitry, representation maps, 3-dimensional reconstructions, biocytin
D 1993 Wiley-Liss, Inc.
The aim of the present work is to understand the roles of intrinsic or intracortical synaptic interactions in the func- tions of the motor cortex. Although synapses of local origin may account for as much as 70% of the synapses in certain areas of the cerebral cortex (e.g., Gruner et al., '741, relatively little progress has been made towards elucidating the roles of these intrinsic interactions. However, where data are available, they support a prominent role for intrinsic circuitry in shaping and modulating the response properties of cortical neurons. This has been most clearly demonstrated in recent studies of intrinsic circuitry in the visual cortex (see reviews by Gilbert et al., '90; Martin, '88). These studies demonstrated the existence of extensive projections of axon collaterals, originating from pyramidal neurons, which mediate both intracolumnar and intercolum- nar synaptic interactions (Gilbert et al., '90). Particularly prominent are the horizontal intracortical connections
formed by axon collaterals that extend for several millime- ters distal to their parent soma, commonly forming several clusters of axon terminals (DeFelipe et al., '86; Gilbert and Wiesel, '83; Martin and Whitteridge, '84).
Attempts to decipher the functional organization of the motor cortex have focused primarily on revealing the somatotopic organization of this area (for historical reviews see Asanuma, '89; Humphrey, '86). It is conceivable that motor cortical functions are shaped in large part also by intracortical synaptic circuitry. Our understanding of the functional organization of this cortical area is hindered by the paucity of data on intrinsic connections within the motor cortex. Recent studies have shown that long, horizon-
Accepted May 14,1993. Address reprint requests to Dr. Asaf Keller, Department of Anatomy,
USUHS, 4301 Jones Bridge Road, Bethesda, MD 20814-4799.
o 1993 WILEY-LISS, INC.

230 A. KELLER AND H. ASANUMA
tal axon collaterals provide a substrate for synaptic interac- tions among neurons located in different representation zones in the motor cortex (Huntley and Jones, '91; Keller, '93). Furthermore, there are data indicating that these intracortical interactions mediate the modulation of motor maps that occurs during motor learning or subsequent to nerve damage (Donoghue and Sanes, '91; Jacobs et al., '91; Jacobs and Donoghue, '91).
The present study aims to decipher aspects of intrinsic circuitry in the motor cortex by examining the synaptic relationships of pyramidal neurons in the superficial layers of the cat motor cortex. We have demonstrated recently that these cells are involved in the induction of long-term potentiation (LTP) in the motor cortex (Keller et al., '901, and suggested that this form of plasticity may be involved in processes of motor learning and memory (Asanuma and Keller, '91). In the present study, we identified pyramidal neurons in which LTP was evoked following tetanic stimu- lation of their afferents from the somatosensory cortex. These cells were labeled intracellularly and their intrinsic axonal arbors traced and reconstructed in three dimen- sions. The neuronal elements postsynaptic to these labeled axon collaterals were then identified by serial section electron microscopy. Some of these results were presented in a preliminary form (Keller and Asanuma, '91).
MATERIALS AND METHODS Electrophysiology
Intracellular recordings and labeling from neurons in the cat motor cortex were performed with the aid of procedures detailed previously (Keller et al., '90, '91). In brief, experi- ments were performed on 15 adult cats of both sexes (weight 2.5-3.0 kg), under Nembutal anesthesia (initial dose 35 mgikg i.p.; maintenance dose 2 mglkgih, i.v.1. Glass micropipettes filled with 4% Neurobiotin (Vector) dissolved in 2.5 M KC1 and 0.05 M Tris (pH 7.4, 37°C) were used for intracellular recordings from neurons in the motor cortex (area 4y). An array of 9 platinum-in-glass stimulat- ing electrodes inserted into the peri-ansate region (areas 1-2) was used to deliver intracortical microstimulation (ICMS; 30 LA, 0.2 ms, 1 Hz). When a motor cortical neuron responded to the ICMS with excitatory postsynaptic poten- tials (EPSPs), the responses of EPSPs to double-pulse stimulation (1 to 3 ms intervals) was examined. Facilitation of the second of a pair of EPSPs was measured relative to the latency and amplitude of the first. EPSPs were classi- fied as monosynaptic if the latency of the second response was identical to that of the first. The latencies of all the EPSPs classified as monosynaptic ranged from 0.9 to 2.6 ms (1.9 * 0.41, in agreement with previous studies (Kosar et al., '85). After at least 5 minutes of control recordings, tetanic stimulation (50 Hz, 5 s) was delivered through the responsible stimulating electrode. Immediately following the tetanic stimulation, recordings of EPSPs in response to the test ICMS (1 Hz) were resumed.
Upon completion of the electrophysiological recordings, intracellular injections of Neurobiotin were made by pass- ing depolarizing current pulses (2 nA, 2 Hz, 100 ms) through the recording electrode.
Histology The cats were perfused under Nembutal anesthesia by
transcardial infusion of 350 cc of buffered saline followed by a fixative containing 4% paraformaldehyde, 0.1% glutaralde-
hyde and 0.2% picric acid. The brains were removed and stored in the same fixative solution overnight a t 4°C. Blocks of tissue containing area 4y were cut into 50 pm thick sagittal or tangential sections on a vibratome, and the sections were then processed to reveal Neurobiotinm- labeled cells according to previously described procedures (Keller et al., '90).
Three labeled pyramidal cells were selected for further anatomical analyses. These cells were photographed and then traced, in serial sections, by means of a computer- aided morphometry system (Neurolucida, MicroBright- Field). Sections immediately adjacent to those containing labeled cells were Nissl-stained to identify the boundaries of cytoarchitechtonic areas and of cortical layers.
Electron microscopy For each of the three cells, two regions of axon collaterals
were selected for electron microscopy. One region contained axon collaterals arborizing within the immediate vicinity of the parent soma and proximal dendrites (proximal region), and a second region containing terminal-rich zones located horizontally at least 500 pm distal to the parent soma (distal region). These regions were blocked and prepared for electron microscopic examination (White and Keller, '87). These blocks were cut into long, unbroken series of thin sections (350-620 sections per series) that were mounted on formvar-coated grids.
Axon terminals containing the Neurobiotin reaction prod- uct were identified in these thin sections and photographed on every thin section of each series. These photographs were printed at a final magnification of x 19,500. Analysis of the prints included identification of the neuronal ele- ments postsynaptic to these axon terminals and their classification as either dendritic spines, dendritic shafts or somata (see Results). Some of the postsynaptic processes were subsequently followed and photographed through long series of consecutive sections. The photographs of these profiles were used for a three-dimensional reconstruc- tion of the neuronal elements postsynaptic to labeled terminals. The reconstruction process was done by means of the Neurolucida (MicroBrightField) computer-aided mor- phometry system. The profiles of selected postsynaptic elements were digitized and aligned relative to each other using fiduciary points. Profiles from consecutive sections were displaced from each other in the z-axis by a distance equal to the approximate section thickness (90 nm; Willi- ams and Meek, '66) times the final print magnification. In addition, the locations of all types of synapses on each of the profiles were marked on the digitized image.
To determine the synaptic density of all types of synapses in the neuropil, the dissector technique (Sterio, '84) was applied, as described previously (Keller et al., '92). Using the dissector method, the numerical densities (Nu) of asymmetrical synapses with dendritic spines, dendritic shafts, and somata in the neuropil surrounding each of the labeled terminal clusters were calculated.
RESULTS Electrophysiology
Tetanic stimulation of corticocortical afferents was at- tempted in 15 cells from which stable intracellular record- ings were obtained. All the cells responded with monosynap- tic EPSPs to stimulation in the somatosensory cortex (SCx), as evidenced by the latency of these responses ( < 2.6 msec) and by response to double-shock stimulation (see

INTRINSIC CIRCUITRY OF CAT MOTOR CORTEX 23 1
180
Fig. 1. Time course of long-term potentiation (LTP) induced in a motor cortical neuron (Cell B) that responded with monosynaptic inputs from the somatosensory cortex. The amplitudes of excitatory
postsynaptic potentials (EPSPs) were recorded before and after tetanic stimulation (arrow), and these values are expressed as percentage of mean control EPSP amplitudes.
Materials and Methods). In all 15 cells, tetanic stimulation produced a significant and persistent increase in EPSP amplitudes. An example of the time course of this potentia- tion is depicted in Figure l. In this cell (B), as well as in all other cells, the potentiation was preceded by a transient depression of EPSP amplitudes, lasting 30 seconds to 200 seconds. Immediately thereafter, EPSP amplitudes in- creased and stabilized at a potentiated level (168% to 240% of control amplitudes) for the duration of the recording period (minimum 20 minutes). Throughout the recording period, no changes were observed in the resting membrane potential or in the resistance of the recorded cells.
After documenting the effects of tetanic stimulation in these cells, the cells were filled by intracellular injections of Neurobiotin. Subsequent histochemical processing re- vealed that all the cells were either pyramidal or nonpyrami- dal neurons having somata in layers I1 or I11 of area 47. These findings are in agreement with our previous reports (Keller et al., '90; Sakamoto et al., '871, demonstrating that cells in the superficial layers of the cat motor cortex that receive monosynaptic inputs from the SCx produce LTP in response to tetanic stimulation of these afferents.
Light microscopy Three pyramidal neurons that produced LTP in response
to SCx stimulation were selected for anatomical analyses. Two of the cells (A and B) were sectioned in a parasagittal plane; cell C was sectioned tangentially, in a plane parallel to the pial surface. Portions of the dendritic trees, and all the discernible axon collaterals, were traced and analyzed by means of a three-dimensional computer reconstruction package (see Materials and Methods).
The somata of cells A and C were located in layer 11, and that of cell B in superficial layer 111. The size of these somata was similar, having major diameters ranging from 14 to 18 pm. All the cells had spiny dendrites, including an
apical dendrite that extended up to and ramified in layer I, and a rich basal dendritic tree (Fig. 2). The apical dendrite of cell B, whose parent somata was in layer 111, was elongated, branching mainly in upper layer I1 and in layer I. The apical dendrite of the more superficially located cell A emitted secondary branches from its proximal segments, close to the parent soma in layer 11. The lateral spread of the basal dendrites belonging to cells A and C was 350 and 380 pm, respectively, whereas the basal dendrites of cell B were more restricted, occupying a region of 220 pm around the parent soma.
The axon of each of the cells originated from the soma, and projected towards the white matter, where it could no longer be traced. Along its course towards the white matter, a number of branches left the main axon at a right angle, and projected horizontally, usually within a single layer. In the immediate vicinity of their parent soma, these axon collaterals branched extensively, forming a cluster of axo- nal arborizations. Extensive arborizations were formed also by the long, horizontal branches emitted within the same layer the parent soma was located. Usually, each cell had a single, long, horizontal collateral in layers I1 or 111, and this branch projected in the anteroposterior plane. These hori- zontal collaterals projected for a distance of up to 1 mm, forming several clusters of axon terminals along their route. Usually, a cluster of axon terminals was formed at the distal end of these long, horizontal collaterals. Addi- tional clusters were formed along the path of these collater- als, with no apparent regular periodicity between these clusters. Numerous boutons, presumably axon terminals, were associated with these axon collaterals, primarily within the dense clusters of axonal arborizations. Both boutons terminaux and boutons en passant were identified along these long, horizontal axon collaterals (Fig. 3). The clusters of axon terminals belonging to cells A and C had diameters that were similar to those of the basal dendrites of their

A
B ps w
ps
V
C
V
?
Fig.
2.
Rec
onst
ruct
ions
, fr
om s
eria
l se
ctio
ns,
of t
he p
yram
idal
ne
uron
s (A
-C)
exam
ined
in
this
stu
dy. A
xons
are
pre
sent
ed a
s so
lid
lines
and
the
loca
tions
of
bout
ons
as c
ircl
es. P
artia
lly r
econ
stru
cted
de
ndri
tes a
re re
pres
ente
d as
bro
ken
lines
. Box
ed a
reas
repr
esen
t are
as
sele
cted
for e
lect
ron
mic
rosc
opic
al an
alys
es. I
n A
and
B, c
ortic
al la
yers
ar
e in
dica
ted
by R
oman
num
eral
s, l
amin
ar b
ound
arie
s by
hor
izon
tal
lines
that
repr
esen
t 100
km
. Sca
le b
ar in
C =
100
km
.

INTRINSIC CIRCUITRY OF CAT MOTOR CORTEX 233
Fig. 3. A-C: Micrographs of Neurobiotin-filled axon collaterals belonging to the pyramidal neurons examined. Pia is towards the top. Both boutons terminaux (arrowheads) and boutons en passant (arrows) can be identified. Scale bar = 30 km.
parent somata. The gaps between these clusters also matched the diameter of their corresponding basal den- dritic fields (Fig. 2). This is agreement with a recent report by Lund et al. ('931, demonstrating, in several cortical areas of the macaque monkey, a close correlation between the size of intracortical axon terminal clusters and terminal-free gaps and the lateral spread of basal dendrites belonging to pyramidal cells.
Electron microscopy For each cell, two segments of axon collaterals were
selected for electron microscopy: a proximal segment arbo- rizing in close proximity to the parent soma and apical dendrite, and a distal segment located at least 500 km
lateral to the parent soma. Tissue samples from both the distal and proximal segments were trimmed to include an axon terminal-rich cluster. Electron microscopical analyses of tissue containing proximal and distal clusters included: (1) localization and identification of labeled axon terminals, (2) identification and classification of neural elements postsynaptic to labeled axon terminals, and (3) assessment of the selectivity of labeled axon terminals to form synapses with particular targets.
The local axon collaterals of the labeled cells ranged from 0.4 to 1.5 pm in diameter. The proximal branches of these axon collaterals, arborizing near their parent somata, were unmyelinated, whereas the horizontal branches entered a myelin sheath 20-50 km
Labeled axon terminals.
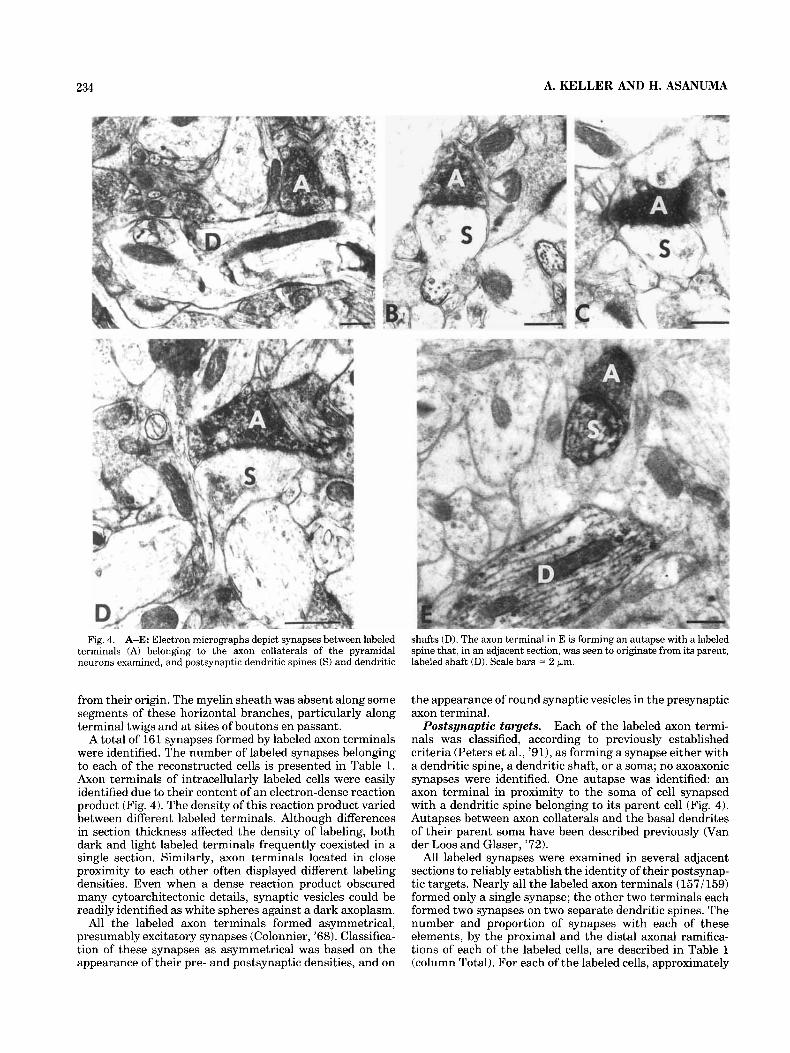
234 A. KELLER AND H. ASANUMA
Fig. 4. A-E: Electron micrographs depict synapses between labeled terminals (A) belonging to the axon collaterals of the pyramidal neurons examined, and postsynaptic dendritic spines ( S ) and dendritic
shafts (D). The axon terminal in E is forming an autapse with a labeled spine that, in an adjacent section, was seen to originate from its parent, labeled shaft (D). Scale bars = 2 km.
from their origin. The myelin sheath was absent along some segments of these horizontal branches, particularly along terminal twigs and at sites of boutons en passant.
A total of 161 synapses formed by labeled axon terminals were identified. The number of labeled synapses belonging to each of the reconstructed cells is presented in Table 1. Axon terminals of intracellularly labeled cells were easily identified due to their content of an electron-dense reaction product (Fig. 4). The density of this reaction product varied between different labeled terminals. Although differences in section thickness affected the density of labeling, both dark and light labeled terminals frequently coexisted in a single section. Similarly, axon terminals located in close proximity to each other often displayed different labeling densities. Even when a dense reaction product obscured many cytoarchitectonic details, synaptic vesicles could be readily identified as white spheres against a dark axoplasm.
All the labeled axon terminals formed asymmetrical, presumably excitatory synapses (Colonnier, '68). Classifica- tion of these synapses as asymmetrical was based on the appearance of their pre- and postsynaptic densities, and on
the appearance of round synaptic vesicles in the presynaptic axon terminal.
Each of the labeled axon termi- nals was classified, according to previously established criteria (Peters et al., '91), as forming a synapse either with a dendritic spine, a dendritic shaft, or a soma; no axoaxonic synapses were identified. One autapse was identified: an axon terminal in proximity to the soma of cell synapsed with a dendritic spine belonging to its parent cell (Fig. 4). Autapses between axon collaterals and the basal dendrites of their parent soma have been described previously (Van der Loos and Glaser, '72).
All labeled synapses were examined in several adjacent sections to reliably establish the identity of their postsynap- tic targets. Nearly all the labeled axon terminals (1571159) formed only a single synapse; the other two terminals each formed two synapses on two separate dendritic spines. The number and proportion of synapses with each of these elements, by the proximal and the distal axonal ramifica- tions of each of the labeled cells, are described in Table 1 (column Total). For each of the labeled cells, approximately
Postsynaptic targets.

INTRINSIC CIRCUITRY OF CAT MOTOR CORTEX 235
TABLE 1. The Number and Proportion of Various Postsynaptic Neuronal Elements Forming Synapses With the Labeled Axon Collaterals Examined in This Study'
Unknown Spiny Nonspiny Total
Cell
A
B
C
All
Location
Proximal Distal Proximal Distal Proximal Distal Proximal Distal Total
Spine Shaft
15 79% 4 21% 17 85% 3 15% 18 82% 4 18% 13 81% 3 19% 12 75% 4 25% 6 33% 12 67%
45 79% 12 21% 36 67% 18 33% 81 30
Spine Shaft
4 100% 0 0% 2 100% 0 0% 2 67% 1 33% 2 100% 0 0% 3 1009, 0 0% 5 83% 1 17% 9 90% 1 10% 9 90% 1 10%
18 2
Spine
1 17% 0 0 0% 1 14% 0 0% 0 0% 1 7% 1 6% 2
Shaft Soma
5 83% 0 0% 0 0 4 80% 1 20% 3 43% 3 43% 3 100% 0 0% 5 56% 4 44%
12 86% 1 7% 8 50% 7 44%
20 8
Spine
20 69% 19 86% 20 67% 16 64% 15 68% 11 33% 55 68% 46 58%
101 63%
Shaft
9 31% 3 14% 9 30% 6 24% 7 32%
18 55% 25 31% 27 34% 52 32%
Soma
0 0% 0 0% 1 3% 3 12% 0 ow 4 12% 1 170 7 9% 8 5%
Grand total
29 22 30 25 22 33 81 80
161
'Location, axon collaterals forming synapses proximal or distal to the location of their parent somata. Unknown, synapses formed with dendritic spines and shafts whose parent cell type was not conclusively defined. Spiny and Nonspiny, synapses formed with the dendritic shafts and spines and with the somata of cells conclusively identified as either spiny neurons (i.e., pyramidal cells) or nonspiny neurons he. , nonpyramidal cells).
equal numbers of axon terminals were studied in the proximal and the distal arborization zones.
Within the proximal arborization zones, ail three cells formed the majority of their synapses with dendritic spines (67-69%), and their remaining synapses with dendritic shafts (30-32%) or with somata (0-3%). There were little differences in the relative proportions of synapses with different neural elements among the proximal axon collater- als of the three cells (Table 1, column Total). The three cells differed, however, in the proportions of synapses they had with different neural elements within their distal arboriza- tions. The distal axon collaterals belonging to cells A and B synapsed preferentially with dendritic spines, but they differed in the proportions of synapses formed with the different cells. Cell A formed 86% of its distal synapses with dendritic spines, and the remaining synapses with dendritic shafts. Cell B formed 64% of its distal synapses with dendritic spines, 24% with dendritic shafts, and 12% with somata. Cell C differed from both the other cells in that the majority of its distal synapses were with dendritic shafts (55%), only 33% of its synapses were with dendritic spines, and the remaining synapses with somata.
A more complete description of the neural elements postsynaptic to labeled axon collaterals was attained by classifying them as belonging to either smooth or spiny postsynaptic dendrites. For this purpose, postsynaptic tar- gets were reconstructed from unbroken series of thin sections. Postsynaptic spines were reconstructed back to their dendrites of origin, and these dendrites were then examined in extensive series of thin sections and recon- structed to assess their shapes and the numbers and distribution of the different types of synapses they formed. A similar analysis was done for postsynaptic dendritic shafts and for somata that formed synapses with labeled axon terminals. The morphologies of the reconstructed elements and the distribution of synapses they received were used for the classification of these postsynaptic tar- gets.
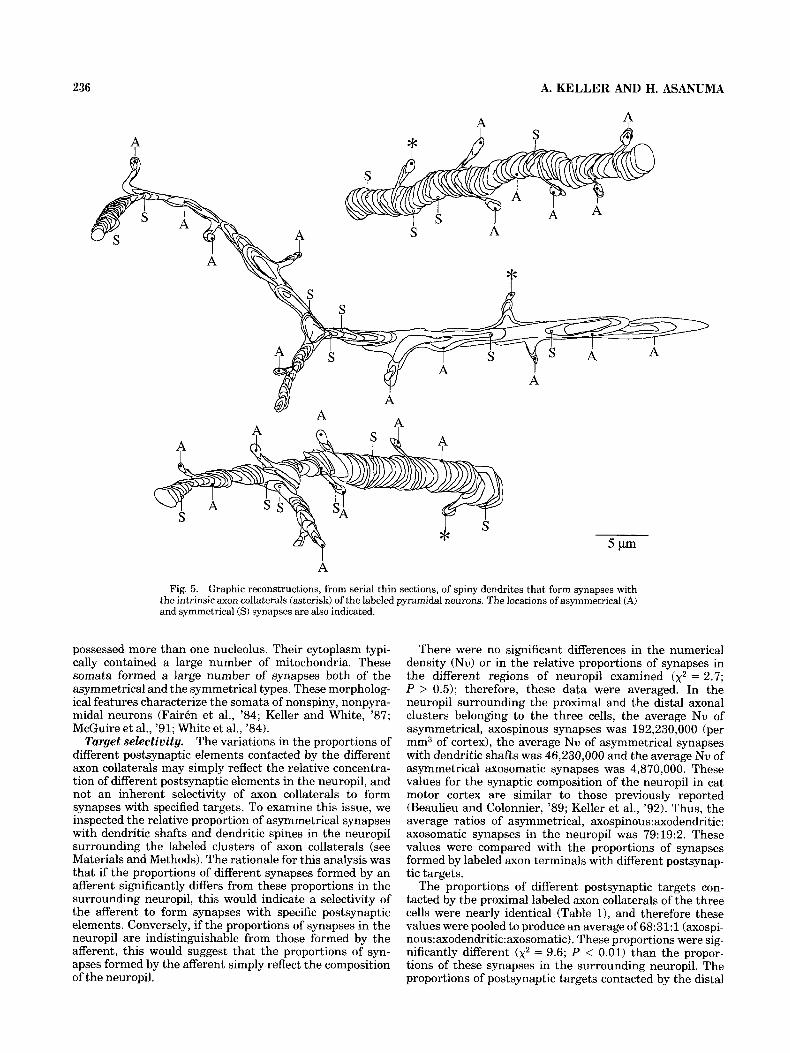
Spinypostsynaptic dendrites. Twenty spines postsynap- tic to labeled axon terminals were reconstructed back to their dendrites of origin (Fig. 5 ) . Most of these dendrites were oriented vertically towards the pial surface, and were usually straight with relatively constant diameters. All of these reconstructed dendrites synapsed only with a single labeled axon terminal. Following serial section analysis, two of the spines postsynaptic to labeled axon terminals were identified as belonging to sparsely spiny dendrites (see below). In the remaining eighteen dendrites, a large num- ber of spine necks were emitted along these dendritic
shafts, each terminating in a spine head receiving a single, asymmetrical synapse. A relatively small number of syn- apses was directly on the shafts of these dendrites; a synapse was formed on the shafts of these spiny dendrites at 3-9 ym intervals (mean = 0.4 synapses/km, n = 18). More than half of these shaft synapses were of the symmet- rical type, and many occurred near dendritic branch points. Similar criteria on the shape of dendritic segments and their complement of synapses have been previously shown to characterize spiny dendrites (Hersch and White, '82; McGuire et al., '84, '91; Saint Marie and Peters, '85; White and Hersch, '82; White et al., '80).
Smooth postsynaptic dendrites. Twenty-two dendritic shafts postsynaptic to labeled axon terminals were recon- structed. Two of these dendritic shafts forming synapses with labeled terminals were classified as belonging to spiny dendrites, based on the criteria outlined above. The remain- ing dendritic shafts were either cylindrical, or they had a beaded appearance, with periodic swellings along the shaft (Fig. 6). Both the smooth and the beaded dendrites were oriented horizontally or obliquely at various angles to the pial surface. The dendritic shafts typically contained a large number of mitochondria, particularly within their beaded segments. Although some of these dendrites exhibited an occasional spine, these spines were sparse ( < 1 spine/5 mm shaft length) and usually lacked a spine neck (sessile spines). These dendritic segments were densely innervated, commonly forming synapses at intervals of 1 km, or less (mean = 1.7 synapses/km, n = 22). The large majority of these synapses were asymmetrical. Some of the recon- structed dendrites formed two (n = 4) or three (n = 1) synapses with labeled axon terminals, and these occurred in close proximity to each other. One smooth dendrite was traced back to its parent soma (Fig. 6). This cell body formed both asymmetrical and symmetrical synapses; since only nonspiny neurons are known to receive asymmetrical synapses on their somata (Fairen et al., '84; Keller and White, '87; Peters and FairBn, '78), this confirms the classification of this cell as a nonspiny neuron.
The morphology of these smooth or sparsely spinous dendrites, and the patterns of synapses they form, are characteristic of nonspiny, nonpyramidal neurons (Keller and White, '87; LeVay, '73; McGuire et al., '84; Peters and Fairen, '78; Somogyi et al., '83).
Postsynaptic somata. Eight labeled axon terminals formed synapses with neuronal somata. Two cell bodies each received three such synapses, and two additional cells each received a single labeled synapse. The nuclei of these somata showed no unique nuclear inclusions and usually
236 A. KELLER AND H. ASANUMA
A
A A W I
A Fig. 5. Graphic reconstructions, from serial thin sections, of spiny dendrites that form synapses with
the intrinsic axon collaterals (asterisk) of the labeled pyramidal neurons. The locations of asymmetrical (A) and symmetrical (S) synapses are also indicated.
possessed more than one nucleolus. Their cytoplasm typi- cally contained a large number of mitochondria. These somata formed a large number of synapses both of the asymmetrical and the symmetrical types. These morpholog- ical features characterize the somata of nonspiny, nonpyra- midal neurons (Fairen et al., '84; Keller and White, '87; McGuire et al., '91; White et al., '84).
The variations in the proportions of different postsynaptic elements contacted by the different axon collaterals may simply reflect the relative concentra- tion of different postsynaptic elements in the neuropil, and not an inherent selectivity of axon collaterals to form synapses with specified targets. To examine this issue, we inspected the relative proportion of asymmetrical synapses with dendritic shafts and dendritic spines in the neuropil surrounding the labeled clusters of axon collaterals (see Materials and Methods). The rationale for this analysis was that if the proportions of different synapses formed by an afferent significantly differs from these proportions in the surrounding neuropil, this would indicate a selectivity of the afferent to form synapses with specific postsynaptic elements. Conversely, if the proportions of synapses in the neuropil are indistinguishable from those formed by the afferent, this would suggest that the proportions of syn- apses formed by the afferent simply reflect the composition of the neuropil.
Target selectivity.
There were no significant differences in the numerical density (Nu) or in the relative proportions of synapses in the different regions of neuropil examined ( x 2 = 2.7; P > 0.5); therefore, these data were averaged. In the neuropil surrounding the proximal and the distal axonal clusters belonging to the three cells, the average Nu of asymmetrical, axospinous synapses was 192,230,000 (per mm3 of cortex), the average Nu of asymmetrical synapses with dendritic shafts was 46,230,000 and the average Nu of asymmetrical axosomatic synapses was 4,870,000. These values for the synaptic composition of the neuropil in cat motor cortex are similar to those previously reported (Beaulieu and Colonnier, '89; Keller et al., '92). Thus, the average ratios of asymmetrical, axospinous:axodendritic: axosomatic synapses in the neuropil was 79:19:2. These values were compared with the proportions of synapses formed by labeled axon terminals with different postsynap- tic targets,
The proportions of different postsynaptic targets con- tacted by the proximal labeled axon collaterals of the three cells were nearly identical (Table 11, and therefore these values were pooled to produce an average of 68:31:1 (axospi- nous:axodendritic:axosomatic). These proportions were sig- nificantly different ( x 2 = 9.6; P < 0.01) than the propor- tions of these synapses in the surrounding neuropil. The proportions of postsynaptic targets contacted by the distal

INTRINSIC CIRCUITRY OF CAT MOTOR CORTEX 237
A
A
Fig. 6. Graphic reconstructions, from serial thin sections, of nonspiny dendrites and a cell body that form synapses with the intrinsic axon collaterals (asterisk) of the labeled pyramidal neurons. The locations of asymmetrical (A) and symmetrical (S) synapses are also indicated.
axon collaterals varied greatly among the cells (Table 1): cell A-86:14:0; cell B-64:24:12; cell C-33:55: 12. The proportions of postsynaptic targets contacted by the distal axon collaterals of cells B and C were significantly different from the proportions of postsynaptic targets in the sur- rounding neuropil (x2 tests; P < 0.0001). The values of these proportions for cell A were not significantly different from the corresponding values in the surrounding neuropil (x2 = 3.9; P > 0.1). These findings indicate that the varia- tions in the proportions of synapses formed by the different axon collaterals reflect an inherent selectivity in the synap- tic targets of these afferents.
DISCUSSION Clustered intrinsic connections
Our results show that pyramidal neurons in the superfi- cial layers of the cat motor cortex possess extensive intrin- sic axon collaterals that arborize to form clusters of axon terminals. These terminal clusters are formed in the imme- diate vicinity of the parent somata, and along the long, horizontal collaterals that project in an anteroposterior plane in the same layer as their parent somata.
This finding is in agreement with other evidence for clustered intrinsic connections in the motor cortex. For example, applications of minute quantities of anterograde or retrograde tracers in a single representation zone of the cat motor cortex reveal the existence of long (up to 3 mm), horizontal axon collaterals that terminate in clusters of axon terminals (Keller, '93). Similarly, intrinsic axon collat- erals that project preferentially in the anteroposterior plane give rise to clusters of axon terminals in the monkey motor cortex (Huntley and Jones, '91; Lund et al., '93). Earlier studies analyzing the distribution of intrinsic axon termi- nals in the motor cortex labeled by lesion-induced degenera- tion (Gatter and Powell, '78), by the application of tracers (DeFelipe et al., '86; Jones et al., '78), or by intracellular injections of dyes (Deschgnes et al., '79; Keller et al., '90; Landry et al., '80) have also provided evidence for a clustered distribution of axon terminals of intrinsic origin in the motor cortex.
Clustered intrinsic connections in other cortical areas. The existence of long, horizontal axon collaterals that terminate in clusters of axon terminals is a common feature of pyramidal neurons in many cortical areas (Feldman, '84). For example, these patterns have been demonstrated in the

238
visual cortex of the monkey (Fitzpatrick et al., '851, cat (Gilbert and Wiesel, '79), rat (Burkhalter, '891, and tree shrew (Rockland et al., '82), in the somatosensory cortex of the cat (Schwark and Jones, '89), mouse (Bernard0 et al., 'go), and monkey (DeFelipe et al., '86), and in the auditory cortex of the cat (Ojima et al., '91; Wallace et al., '91).
These horizontally oriented projections may extend for distances of over 3 mm (e.g., Gilbert and Wiesel, '791, suggesting that they link neurons within separate func- tional columns. Some insight into the function of these horizontal excitatory connections has been provided by studies showing that in most cortical areas examined, these synaptic interactions link functional columns having cells with similar receptive field properties (Gilbert, '92). For example, excitatory interactions in the primary visual cortex occur between cells residing within columns of like orientation selectivity, but not between neurons within columns of different orientation selectivities (Gilbert and Wiesel, '89; Schwarz and Bolz, '91; Ts'o et al., '86). Excitatory connections link regions of the cat auditory cortex that contain neurons having similar frequency pref- erences (Matsubara and Phillips, '88). Excitatory interac- tions between neurons sharing similar receptive field prop- erties have also been identified in the somatosensory cortex of the cat, where they link cells belonging to the same submodality band (Metherate and Dykes, '85).
In addition to the axon collaterals of pyramidal neurons, the axonal arbors of nonpyramidal, GABAergic cells may also contribute to intrinsic, horizontal interactions in the motor cortex. Using retrograde labeling of tritiated GABA, DeFelipe and Jones ('85) demonstrated that GABAergic, intracortical axons are largely restricted to a narrow colum- nar domain in the motor cortex, and do not project exten- sively parallel to the pial surface. However, a contribution to horizontal intrinsic connections can be made by the GABAergic basket cells whose horizontal axon collaterals may extend for more than 1 mm (DeFelipe et al., '85; Jones, '75). Recent studies in the visual cortex have shown that a subset of GABAergic neurons contribute to long range, inhibitory connections (Kritzer et al., '92; McDonald and Burkhalter, '93; but see Matsubara and Boyd, '92 for contrary findings).
Synaptology of axon collaterals in the motor cortex
The goal of the present study was to decipher the patterns of synaptic connections that link neurons in the motor cortex to form functional circuits. To this end, the neural elements postsynaptic to identified axon collaterals of superficial pyramidal neurons were identified. We have examined the output relationships of axon collaterals termi- nating in clusters in close proximity to their parent soma, as well as those in the distal clusters formed along the long, horizontal collateral branches.
Synaptology o f proximal clusters. The proximal branches of the axon collaterals belonging to all three cells examined synapsed preferentially with dendritic spines. These postsynaptic spines are likely to belong to other pyramidal neurons. This conclusion is based on the finding that the majority (92%) of the spines that were traced in serial thin sections were identified as belonging to pyrami- dal neurons.
Relatively few studies have examined the output relation- ships of axon collaterals belonging to pyramidal neurons (see White, '89). Pertinent to the present study are reports
A. KELLER AND H. ASANUMA
that proximal clusters of axon collaterals belonging to pyramidal neurons in the superficial layers of the cat (Kisvarday et al., '86) and monkey (McGuire et al., '91) visual cortex and in layer V of the cat visual cortex (Gabbott et al., '87) synapse preferentially with dendritic spines. Similarly, axon collaterals of corticocortical (Elhanany and White, '90) and callosal (White and Czeiger, '91) neurons in the mouse somatosensory cortex synapse preferentially with spines belonging to pyramidal neurons.
Other types of pyramidal neurons have been reported to form most of their intrinsic synapses with dendritic shafts of non-, or sparsely spinous neurons. Layer VI pyramids in the cat visual cortex form the majority of their intrinsic connections with dendritic shafts, presumably belonging to nonpyramidal neurons (McGuire et al., '84). Similarly, axon collaterals of layer V/VI corticothalamic cells in the mouse barrel cortex (White and Keller, '87), of a layer I11 pyramid in monkey somatosensory cortex (Winfield et al., '811, and of layer I1 pyramids in the opossum piriform cortex (Haber- ly and Presto, '86) all provide input preferentially to nonpyramidal neurons.
In contrast to the unifor- mity in the proportions of postsynaptic targets contacted by the proximal clusters of the axon collaterals belonging to all three cells examined, the distal branches of these collaterals differed significantly in the proportions of their postsynap- tic targets. Thus, Cell A formed the majority (86%) of its synapses with dendritic spines; Cell B provided 64% of its output to spines of pyramidal neurons; the remaining 36% to dendritic shafts and somata of nonpyramids, and Cell C provided 67% of its output to the dendrites and somata of nonpyramidal neurons (Fig. 7). A number of studies have reported that axon collaterals of certain types of pyramidal neurons synapse preferentially with nonpyramidal cells (see above). The relatively high proportion of synapses with the somata of nonpyramidal neurons was surprising, since although axosomatic synapses formed by collaterals of pyramidal neurons have been reported previously (Kisvard- ay et al., '861, these synaptic patterns were thought to be rare in the cerebral cortex (White, '89).
Even less expected was the finding that the proportions of different postsynaptic targets contacted by the proximal versus the distal axonal clusters of the same cell can differ significantly. In the few studies where the synaptic targets of proximal versus distal axon collaterals were compared, similar output patterns were reported in both loci. Axon collaterals belonging to layer I11 pyramids in the monkey visual cortex (McGuire et al., '91), and layer V pyramids in cat visual cortex (Gabbott et al., '87) synapse preferentially with dendritic spines, in both their proximal and their distal termination zones. However, extrinsic pathways affer- ent to the cerebral cortex have been reported to form synapses with different targets in different portions of their arborizations. For example, claustral afferents to the cat visual cortex arborize in layers I and VI, and in layer IV, in layers I and VI they synapse almost exclusively with dendritic spines, whereas in layer IV they synapse in about equal proportions with spines and with shafts (LeVay, '86).
The dendrites of pyra- midal neurons formed only a single synapse with a labeled axon collateral. Although it is not possible to determine with certainty whether the different dendrites that received these synapses belonged to the same cell, this was not considered a likely possibility. The spiny dendrites exam- ined were oriented vertically, towards the pial surface,
Synaptology ofdistal clusters.
Divergence of intrinsic synapses.

INTRINSIC CIRCUITRY OF CAT MOTOR CORTEX 239
7.
/-\ I A
1 Cell A
.-'
64%
y.i$J Cell B v
nonpyramidal X neurons
Fig. 7. Summary diagram depicting the proportions of postsynaptic targets contacted by the axon collaterals of the labeled cells (filled triangles). Postsynaptic targets include spiny dendrites (filled circles), presumably of pyramidal neurons, and the somata and dendrites of nonspiny, presumably inhibitory cells (open circles). A-C correspond to the 3 pyramidal neurons described in this study (Fig. 2).
whereas most of the axon collaterals were oriented horizon- tally (Fig. 3), geometrically constraining the possibility of multiple synapses on a single dendrite. This in agreement with previous studies demonstrating that axon collaterals of pyramidal neurons synapse sparingly with their targets (Gabbott et al., '87; Kisvarday et al., '86; McGuire et al., '84, '91). In contrast, some of the nonpyramidal neurons re- ceived multiple synapses from labeled axon collaterals. These multiple inputs converged to synapse in close proxim- ity to each other, either on dendritic shafts or on somata, strategically placing them to exert potent influences on their postsynaptic targets.
Selectivity of synaptic connections Finding that the proportions of postsynaptic targets of
the distal axon collaterals belonging to the three cells varied to such an extent suggest that each of these axonal branches is highly selective with regard to the neural elements with which it synapses. This specificity is further demonstrated by the fact that the proportions of different synapses
formed by each of the terminal clusters were independent of the synaptic complement in the surrounding neuropil. This indicates that the synaptic targets of each of these terminal clusters are dependent on properties inherent to that afferent, and not on the composition of the neuropil.
A similar conclusion regarding the selectivity of intrinsic axons to synapse with specific types of neural elements has been demonstrated in the monkey visual cortex (McGuire et al., '91), and in the mouse somatosensory cortex (Elhanany and White, '90; White and Czeiger, '91; White and Keller, '87). Similarly, extrinsic afTerents to the cerebral cortex form synapses with specific postsynaptic targets, in propor- tions that are different from and independent of the composition of their surrounding neuropil (White, '89). In the motor cortex, for example, callosal afferents, and affer- ents from the somatosensory and the premotor area each form specific and different proportions of synapses with dendritic spines vs. dendritic shafts, even though the terminal fields of these afferents coexist within the same region (Sloper, '73; Sloper and Powell, '79). Finally, a number of quantitative studies have demonstrated that different types of cortical neurons receive characteristic numbers, types, and proportions of synapses from different pathways (reviewed by White, '89). Taken together, these findings illustrate the high degree of specificity of synaptic interactions in the cerebral cortex, and confound supposi- tions that the synaptology of the cerebral cortex is quasi- random, shaped only by the geometrical constraints of the neuropil (Braitenberg and Schiiz, '91).
It should be noted that the conclusions of the present study are based on a relatively small sample of neurons and synapses. Thus it may be argued that the differences in the proportions of synapses formed by distal axonal clusters reflect a continuum of potential synaptic patterns, and not a specificity. We consider this possibility unlikely for the following reasons: (1) the synaptic patterns formed by the proximal axonal clusters were highly consistent, reflecting a high degree of specificity; (2) different patterns of syn- apses were formed by different cells despite a consistent complement of synapses in the neuropil; and (3) the present data are in agreement with previous demonstrations of specificity of extrinsic and intrinsic cortical pathways (de- scribed above).
The role of intrinsic connection in the motor cortex
Shaping of motor representations. The functional orga- nization of the motor cortex is thought to be composed of discrete efferent zones, each containing neurons associated with the activation of particular muscle groups (Asanuma, '89). A group of muscles may be activated by several efferent zones in the motor cortex, and zones related to a muscle are intermixed with columns related to different muscles (Asanuma and Sakata, '67; Cheney and Fetz, '85; Jankowska et al., '75). The spatial separation of efferent zones related to the same group of muscles suggests that a mechanism exists by which these zones may interact for the execution of movement patterns. The long, horizontal axon collaterals described in the present study may form the anatomical substrate for these interactions. Long, horizon- tal intrinsic connections among different motor representa- tion zones have previously been demonstrated in the cat (Keller, '93) and monkey (Huntley and Jones, '91) motor cortex.

240 A. KELLER AND H. ASANUMA
The proximal axonal clusters of pyramidal neurons syn- apse with other pyramidal neurons, presumably located within the same efferent zone. These synaptic patterns would result in a feed-forward activation of additional excitatory neural components in the same efferent zone. In contrast, the distal axonal clusters of some pyramidal neurons provide input primarily to pyramidal (excitatory) cells, whereas other pyramidal neurons contact nonpyrami- dal (presumably inhibitory) cells. One possibility is that the distal afferents that synapse with pyramidal neurons pro- vide a substrate for feed-forward excitatory interactions between groups of cells related to the same movement pattern. The horizontal axon collaterals of pyramidal cells synapsing with inhibitory neurons may form a basis for lateral inhibition between representation zones related to the activation of different muscles. This arrangement could result in a recruitment of groups of cells related to a particular movement, and a concurrent suppression of cells related to antagonistic movements. Pertinent to this hypoth- esis are findings that intrinsic excitatory interactions in the motor cortex occur preferentially among neurons related to the activation of a single, or neighboring joints (Kwan et al., '87; Murphy et al., '85). Furthermore, blockade of GABAer- gic inhibition in the motor cortex results in disruptions to the spatiotemporal sequences of movement patterns (Mat- sumura et al., '91). Removal of GABAergic inhibition also results in task-related activity in neurons that were previ- ously inactive during the execution of a motor task (Mat- sumura et al., '92).
Several recent stud- ies have demonstrated that the representations of muscles or movements in the motor cortex can be dynamically modulated. For example, these motor maps can be modu- lated in response to repetitive cortical stimulation (Nudo et al., 'go), following motor nerve lesions (Donoghue et al., 'go), intracortical application of pharmacologic agents (Ja- cobs and Donoghue, '911, or training in a motor task (Donoghue and Sanes, '91; Milliken et al., '92; Woody and Black-Cleworth, '73).
One of the mechanisms responsible for this dynamic reorganization may be related to the induction of LTP-like processes, as demonstrated in the present and in our previous studies. Thus, repetitive execution of particular movement patterns may enhance the efficacy of specific somatosensory inputs to the motor cortex, inputs that converge on the pyramidal cells examined in the present study (see Asanuma and Keller, '91). These pyramidal neurons, through their long, horizontal axon collaterals, would subsequently affect synaptic transmission within adjacent efferent zones. Jacobs et al. ('91) have demon- strated that excitatory connections exist between horizon- tally distant sites in the rat motor cortex and that these connections are normally masked by inhibition. Thus, pliability of motor representation maps may involve modu- lation of intracortical inhibition that regulates the efficacy of intrinsic, excitatory connections (Jacobs and Donoghue, '91). The present findings support this hypothesis by demonstrating the existence of horizontal axon collaterals that link neurons in the motor cortex presumably located within different representation zones. The horizontal, feed- forward excitatory connections may be normally masked by the corresponding feed-forward inhibitory inputs. Follow- ing external perturbations, such as training or deafferentia- tion, the excitatory inputs are "unmasked" resulting in an immediate modulation of the motor map.
Plasticity of motor representation.
ACKNOWLEDGMENTS The expert technical assistance of Didi Arissian, Eliza-
beth Ruiz and Daniel Weintraub is gratefully acknowl- edged. Supported by NIH grants NS-31078 (A.K.) and NS-10705 (H.A.). Asaf Keller is a Alfred P. Sloan Research Fellow.
LITERATURE CITED Asanuma, H. (1989) The Motor Cortex. New York: Raven Press. Asanuma, H., and A. Keller (1991) Neurohiological basis of motor learning
and memory. Concepts in Neurosciences 2 - 3 0 , Asanuma, H., and H. Sakata (1967) Functional organization of a cortical
efferent system examined with focal depth stimulation in cats. J. Neurophysiol. 3Or35-54.
Beaulieu, C., and M. Colonnier (1989) Effects of the richness of the environment on six different cortical areas of the cat cerebral cortex. Brain Res. 495:382-386.
Bernardo, K.L., J.S. McCasland, T.A. Woolsey, and R.N. Strominger (1990) Local intra- and interlaminar connections in mouse barrel cortex. J. Comp. Neurol. 291.231-255.
Braitenberg, V., and S. Schiiz (1991) Anatomy of the Cortex: Statistics and Geometry. Berlin: Springer-Verlag.
Burkhalter, A. (1989) Intrinsic connections of rat primary visual cortex: Laminar organization of axonal projections. J. Comp. Neurol. 279t171- 186.
Cheney, P.D., and E.E. Fetz (1985) Comparable patterns of muscle facilita- tion evoked hy individual corticomotoneuronal (CM) cells and by single intracortical microstimuli in primates: Evidence for functional groups of CM cells. J. Neurophysiol. 53:786-804.
Colonnier, M. (1968) Synaptic patterns on different cell types in the different laminae of the cat visual cortex. An electron microscope study. Brain Res.
DeFelipe, J., and E.G. Jones (1985) Vertical organization of y-aminohutyric acid-accumulating intrinsic neuronal systems in monkey cerebral cortex. J. Neurosci. 5:3246-3260.
DeFelipe, J., J. Conley, and E.G. Jones (1986) Long-range focal collateraliza- tion of axons arising from corticocortical cells in monkey sensory-motor cortex. J. Neurosci. 6t3749-3766.
Deschenes, M., A. Lebelle, and P. Landry (1979) Morphological characteriza- tion of slow and fast pyramidal tract cells in the cat. Brain Res. 178.251-274.
Donoghue, J.P., and J. Sanes (1991) Dynamic modulation of primate motor cortex output during movement. Soc. Neurosci. Abst. 21:1022.
Donoghue, J.P., S. Suner, and J.N. Sanes (1990) Dynamic organization of primary motor cortex output to target muscles in adult rats. 11. Rapid reorganization following motor nerve lesions. Exp. Brain Res. 79:492- 503.
Elhanany, E., and E.L. White (1990) Intrinsic circuitry: Synapses involving the local axon collaterals of corticocortical projection neurons in the mouse primary somatosensory cortex. J. Comp. Neurol. 291.43-54.
Fairen, A., J. DeFelipe, and J . Regidor (1984) Nonpyramidal neurons: General account. In A. Peters and E.G. Jones (eds): Cerebral Cortex. Vol 1: Cellular Components of the Cerebral Cortex. New York: Plenum Press, pp. 201-253.
Feldman, M.L. (1984) Morphology of the neocortical pyramidal neuron. In A. Peters and E.G. Jones (eds): Cerebral Cortex Vol. 1: Cellular Compo- nents of the Cerebral Cortex. New York Plenum Press, pp. 123-200.
Fitzpatrick, D., J.S. Lund, and G.G. Blasdel (1985) Intrinsic connections of macaque striate cortex: Afferent and efferent connections of lamina 4C. J. Neurosci. 5:3329-3349.
Gabhott, P.L.A., K.A.C. Martin, and D. Whitteridge (1987) Connections between pyramidal neurons in layer 5 of cat visual cortex (area 17). J. Comp. Neurol. 259:364-381.
Gatter, K.C., and T.P.S. Powell (1978) The intrinsic connections of the cortex of the area 4 of the monkey. Brain 101:513-541.
Gilbert, C.D. (1992) Horizontal integration and cortical dynamics. Neuron 9: 1-13.
Gilbert, C.D., J.A. Hirsch, and T.N. Wiesel (1990) Lateral interactions in visual cortex. Cold Spring Harbor Symp. Quant. 55663-677.
Gilbert, C.D., and T.N. Wiesel (1979) Morphology and intracortical projec- tions of functionally characterized neurons in the cat visual cortex. Nature 28Ot120-125.
9:268-287.

INTRINSIC CIRCUITRY OF CAT MOTOR CORTEX 241
Gilbert, C.D., and T.N. Wiesel (1983) Clustered intrinsic connections in cat visual cortex. J. Neurosci. 3:1116-1133.
Gilbert, C.D., and T.N. Wiesel (1989) Columnar specificity of intrinsic horizontal and corticocortical connections in cat visual cortex. J. Neuro- sci. 9.2432-2442.
Gruner, J.E., J.C. Hisch, and C. Sotelo (1974) Ultrastructural features of the insulated suprasylvan gyrus in the cat. J. Comp. Neurol. 154~1-27.
Haberly, L.B., and S. Presto (1986) Ultrastructural analysis of synaptic relationships of intracellularly stained pyramidal cell axons in piriform cortex. J. Comp. Neurol. 248:464474.
Hersch, S.M., and E.L. White (1982) A quantitative study of the thalamocor- tical and other synapses in layer IV of pyramidal cells projecting from mouse SmI cortex to the caudate-putamen nucleus. J. Comp. Neurol. 211217-255.
Humphrey, D.R. (1986) Representation of movements and muscles within the primate precentral motor cortex: Historical and current prespectives. Fed. Proc. 45.2687-2699.
Huntley, G.W., and E.G. Jones (1991) Relationship of intrinsic connections to forelimb movement representations in monkey motor cortex: A correlative anatomic and physiological study. J. Neurophysiol. 66:390- 413.
Jacobs, K.M., and J.P. Donoghue (1991) Reshaping the cortical motor map by unmasking latent intracortical connections. Science 251t944-947.
Jacobs, K.M., B.W. Connors, and J.P. Donoghue (1991) Layer V contains a substrate for reorganization of motor cortex maps. Soc. Neurosci. Abst. 17:311.
Jankowska, E., Y. Padel, and R. Tanaka (1975) The mode of activation of pyramidal tract cells by intracortical stimuli. J. Physiol. 249:617-636.
Jones, E.G. (1975) Varieties and distribution of non-pyramidal cells in the somatic sensory cortex of the squirrel monkey. J. Comp. Neurol. 160:205-268.
Jones, E.G., J.D. Coulter, and S.H.C. Hendry (1978) Intracortical connectiv- ity of architectonic fields in the somatic sensory, motor and parietal cortex of monkeys. J. Comp. Neurol. 181:291-348.
Keller, A. (1993) Patterns of intrinsic connections between motor represen- tation zones in the cat motor cortex. NeuroReport 4:515-518.
Keller, A., and H. Asanuma (1991) Intrinsic connections of cat motor cortex: Axonal projections of superficial pyramids. Soc. Neurosci. Abst. 17:1112.
Keller, A,, and E.L. White (1987) Synaptic organization of GABAergic neurons in the mouse SmI Cortex. J. Comp. Neurol. 262:1-12.
Keller, A,, K. Arissian, and H. Asanuma (1992) Synaptic proliferation in the motor cortex of adult cats following long-term thalamic stimulation. J. Neurophysiol. 68.295-308.
Keller, A., A. Iriki, and H. Asanuma (1990) Identification of neurons producing LTP in the cat motor cortex: Intracellular recordings and labeling. J. Comp. Neurol. 300:47-60.
Keller, A., E. Miyashita, and H. Asanuma (1991) Minimal stimulus parame- ters and the effects of hyperpolarization on the induction of long-term potentiation in the cat motor cortex. Exp. Brain Res. 87295-302.
Kisvarday, S.F., K.A.C. Martin, T.F. Freund, Z. Magloczky, D. Whitteridge, and P. Somogyi (1986) Synaptic targets of HRP-filled layer 111 pyramidal cells in the cat striate cortex. Exp. Brain Res. 64r541-552.
Kosar, E., R.S. Waters, N. Tsukahara, and H. Asanuma (1985) Anatomical and physiological properties of the projection from the sensory cortex to the motor cortex in normal cats: The difference between corticocortical and thalamocortical projections. Brain Res. 345:68-78.
Kritzer, M.F., A. Cowey, and P. Somogyi (1992) Patterns of inter- and intralaminar GABAergic connections distinguish striate (V1) and ex- trastriate (V2, V4) visual cortices and their functionally specialized subdivisions in the rhesus monkey. J. Neurosci. 1245454564.
Kwan, H.C., J.T. Murphy, and Y.C. Wong (1987) Interaction between neurons in the precentral cortical zones controlling different joint. Brain Res. 400:259-269.
Landry, P., A. Labelle, and M. Deschbes (1980) Intracortical distribution of axonal collaterals of pyramidal tract cells in the cat motor cortex. Brain Res. 191:327-336.
LeVay, S. (1973) Synaptic patterns in the visual cortex of the cat and monkey. Electron microscopy of Golgi preparations. J. Comp. Neurol. 150:53-86.
LeVay, S. (1986) Synaptic organization of claustral and geniculate afferents to the visual cortex of the cat. J. Neurosci. 6r3564-3575.
Lund, J.S., T. Yoshioka, and J.B. Levitt (1993) Comparison of intrinsic connectivity in different areas of macaque monkey cerebral cortex. Cerebral Cortex 32148-162.
Martin, K.A.C. (1988) The Wellcome prize lecture: From single cells to simple circuits in the cerebral cortex. Q. J. Exp. Physiol. 73537-702.
Martin, K.A.C., and D. Whitteridge (1984) Form, function and intracortical projections of spiny neurones in the striate visual cortex of the cat. J. Physiol. 353.463-504.
Matsubara, J.A., and J.D. Boyd (1992) Presence of GABA-immunoreactive neurons within intracortical patches in area 18 of the cat. Brain Res. 583: 16 1-170.
Matsuhara, J.A., and D.P. Phillips (1988) Intracortical connections and their physiological correlates in the primary auditory cortex (AI) of the cat. J. Comp. Neurol. 268t3848.
Matsumura, M., T. Sawaguchi, and K. Kubota (1992) GABAergic inhibition of neuronal activity in the primate motor and premotor cortex during voluntary movement. J. Neurophysiol. 68:692-702.
Matsumura, M., T. Sawaguchi, T. Oishi, K. Ueki, and K. Kubota (1991) Behavioral deficits induced by local injection of bicuculline and muscimol into the primate motor and premotor cortex. J. Neurophysiol. 65:1542- 1553.
McDonald, C.T., and A. Barkhalter (1993) Organization of long-range inhibitory connections within rat visual cortex. J. Neurosci. 13t768-781.
McGuire, B.A., C.D. Gilbert, P.K. Rivlin, and T.N. Wiesel(1991) Targets of horizontal connections in Macaque primary visual cortex. J. Comp. Neurol. 305:370-392.
McGuire, B.A., J.P. Hornung, C.D. Gilbert, and T.N. Wiesel(1984) Patterns of synaptic input to layer 4 of cat striate cortex. J. Neurosci. 4:3021- 3033.
Metherate, R., and R.W. Dykes (1985) Simultaneous recordings from pairs of cat somatosensory cortical neurons with overlapping peripheral recep- tive fields. Brain Res. 341:119-129.
Milliken, G.W., R.J. Nudo, R. Grenda, W.M. Jenkins, and M.M. Merzenich (1992) Expansion of distal forelimb representations in primary motor cortex of adult squirrel monkeys following motor training. SOC. Neurosci. Abst. 18.506.
Murphy, J.T., H.C. Kwan, and Y.C. Wong (1985) Cross correlation studies in primate motor cortex: Synaptic interactions and shared input. Can. J. Sci. Neurol. Sci. 12.11-23.
Nudo, R.J., W.M. Jenkins, and M.M. Merzenich (1990) Repetitive microstim- ulation alters the cortical representation of movements in adult rats. Somatosens. Motor Res. 7:463-483.
Ojima, H., C.N. Honda, and E.G. Jones (1991) Patterns of axon collateraliza- tion of identified supragranular pyramidal neurons in the cat auditory cortex. Cerebral Cortex lt80-94.
Peters, A,, and A. Fairen (1978) Smooth and sparsely-spined stellate cells in the visual cortex of the r a t A study using a combined Golgi-electron microscope technique. J. Comp. Neurol. 181:129-172.
Peters, A,, S.L. Palay, and H.d. Webster (1991) The Fine Structure of the Nervous System: Neurons and Their Supporting Cells. Third Edition. New York: Oxford University Press.
Rockland, K.S., J.S. Lund, and A.L. Humphrey (1982) Anatomical banding of intrinsic connections in striate cortex of tree shrews (tupaia glis). J. Comp. Neurol. 209:41-58.
Saint Marie, R.L., and A. Peters (1985) The morphology and synaptic connections of spiny stellate neurons in monkey visual cortex (area 17): A Golgi-electron microscopic study. J. Comp. Neurol. 233213-235.
Sakamoto, T., L.L. Porter, and H. Asanuma (1987) Long lasting potentiation of synaptic potentials in the motor cortex produced by stimulation of the sensory cortex in the cat: A basis of motor learning. Brain Res. 413:360-364.
Schwark, H.D., and E.G. Jones (1989) The distribution of intrinsic cortical axons in area 3b of cat primary somatosensory cortex. Exp. Brain Res. 78:501-513.
Schwarz, C., and J. Bolz (1991) Functional specificity of a long-range horizontal connection in cat visual cortex: A cross-correlation study. J. Neurosci. 112995-3007.
Sloper, J.J. (1973) An electron microscope study of the termination of afferent connections to the primate motor cortex. J. Neurocytol. 2361- 368.
Sloper, J.J., and T.P.S. Powell (1979) An experimental electron microscopic study of afferent connections to the primate motor and somatic sensory cortices. Phil. Trans. Roy. SOC. Lond. B. 285:199-225.
Somogyi, P., Z.F. Kisvhday, K.A.C. Martin, and D. Whitteridge (1983) Synaptic connections of morphologically identified and physiologically characterized large basket cells in the striate cortex of cat. Neuroscience 10:261-294.

242 A. KELLER AND H. ASANUMA
Sterio, J. (1984) The unbiased estimation of number and sizes of arbitrary particles using the dissector. J. Microsc. 134r127-136.
Ts'o, D.Y., C.D. Gilbert, and T.N. Wiesel (1986) Relationships between horizontal interactions and functional architecture in cat striate cortex as revealed by cross-correlation analysis. J. Neurosci. 6:1160-1170.
Van der Loos, H., and E.M. Glaser (1972) Autapses in neocortex cerebri: Synapses between a pyramidal cell's axon and its own collaterals. Brain Res. 48r355-360.
Wallace, M.N., L.M. Kitzes, and E.G. Jones (1991) Intrinsic interlaminar and intralaminar connections and their relationship to the tonotopic map in cat primary auditory cortex. Exp. Brain Res. 86:527-544.
White, E.L. (1989) Cortical Circuits: Synaptic Organization of the Cerebral Cortex- Structure, Function and Theory. Boston: Birkhauser.
White, E.L., and D. Czeiger (1991) Synapses made by axons of callosal projection neurons in mouse somatosensory cortex: Emphasis on intrin- sic connections. J. Comp. Neurol. 303233-244.
White, E.L., and S.M. Hersch (1982) A quantitative study of thalamocortical and other synapses involving the apical dendrites of corticothalamic projection cells in mouse SmI cortex. J. Neurocflol. lIt137-157.
White, E.L., and A. Keller (1987) Intrinsic circuitly involving the local axonal collaterals of corticothalamic projection cells in mouse SmI cortex. J. Comp. Neurol. 26r213-26.
White, E.L., G. Benshalom, and S.M. Hersch (1984) Thalamocortical and other synapses of non-spiny multipolar cells in mouse SmI cortex. J. Comp. Neurol. 229:311-320.
White, E.L., S.M. Hersch, and M.P. Rock (1980) Synaptic sequences in mouse SmI cortex involving pyramidal cells labeled by retrograde filling with horseradish peroxidase. Neurosci. Lett. 19:149-154.
Williams, M.A., and G.A. Meek (1966) Studies on thickness variation in ultrathin sections. J. Roy. Micr. SOC. 85r337.
Winfield, D.A., R.N.L. Brooke, J.J. Sloper, and T.P.S. Powell (1981) A combined Golgi-electron microscopic study of the synapses made by the proximal axon and recurrent collaterals of a pyramidal cell in the somatic sensory cortex of the monkey. Neuroscience 6:1217-1230.
Woody, C.D., and P. Black-Cleworth (1973) Differences in excitability of cortical neurons as a function of motor projection in conditioned cats. J. Neurophysiol. 36:1104-1116.