sustainable horticulture production in the … · horticultural production in the tropics. 3rd –...
TRANSCRIPT

PROCEEDINGS OF THE HORTICULTURE SEMINAR ON
SUSTAINABLE HORTICULTURAL SUSTAINABLE HORTICULTURAL SUSTAINABLE HORTICULTURAL SUSTAINABLE HORTICULTURAL PRODUCTION IN THE TROPICSPRODUCTION IN THE TROPICSPRODUCTION IN THE TROPICSPRODUCTION IN THE TROPICS
3rd to 6th October 2001, Jomo Kenyatta University of Agriculture and Technology (JKUAT), Juja – KENYA
EDITORS
Wesonga, J.M.; T. Losenge; C.K. Ndung’u, K. Ngamau, F.K. Ombwara and S.G. Agong
(JKUAT)
A. Fricke; B. Hau and H. Stützel (Hannover)
Department of Horticulture Jomo Kenyatta University of Agriculture and Technology, Kenya,
in collaboration with
Institute of Vegetable and Fruit Science
and Institute of Plant Diseases and Plant Protection
University of Hannover, Germany.
DAAD-German Academic Exchange Services

Wesonga, et al., 2002. Procedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics. October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya i
Published by the Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology (JKUAT), P.O. Box 62000, Nairobi, Kenya; Institute of Vegetable and Fruit Science and Institute of Plant Diseases & Plant Protection, University of Hannover, Herrenhaeuser Str. 2, D-30419 Hannover Germany and German Academic Exchange Service, DAAD, P.O. Box 14050, Nairobi, KENYA
©2002 Department of Horticulture, JKUAT; Institute of Vegetable and Fruit Science and Institute of Plant Diseases & Plant Protection, University of Hannover and German Academic Exchange Service, DAAD
ISBN: 9966-923-03-9
Organising Committee
Department of Horticulture, JKUAT, Kenya University of Hannover, Germany
Prof. S.G. Agong Team Leader Dr. Cyrus Kamande Ndungu Chairman Dr. Kamau Ngamau Co-ordinator Mr. Francis Kweya Ombwara Secretary Mr. John Mwibanda Wesonga Member Mr. Turoop Losenge Member
Prof. Hartmut Stutzel Team Leader Prof. Benhard Hau Member Dr. Adreas Fricke Member
Enquiries about this book can be addressed to: Jomo Kenyatta University of Agriculture and Technology Department of Horticulture P.O. Box 62000, Nairobi, Kenya Phone +254 151 52711 Fax +254 151 52164 e-mail: [email protected]
Correct citation: Wesonga, J.M.; T. Losenge; C.K. Ndung’u, K. Ngamau, F.K. Ombwara, S.G. Agong, A. Fricke,
B. Hau and H. Stützel. 2002. Proceedings of the Horticulture Seminar on Sustainable Horticultural Production in the Tropics. 3rd – 6th October 2001. Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology

Wesonga, et al., 2002. Procedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics. October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya ii
TABLE OF CONTENT ITEM PAGE
CORRECT CITATION: .................................................................................................................................... I TABLE OF CONTENT.................................................................................................................................... II PREFACE....................................................................................................................................................... V
OFFICIAL OPENING SPEECH .................................................................................................................... VII MUSHROOM PRODUCTION IN SUSTAINABLE SMALL-SCALE FARMING SYSTEM - OPPORTUNITIES AND CONSTRAINTS: A SURVEY OF UASIN GISHU DISTRICT................................... 1
CHARACTERISTICS OF FRENCH BEAN (PHASEOLUS VULGARIS) PRODUCTION: A CASE STUDY OF KIBIRIGWI IRRIGATION SCHEME IN CENTRAL PROVINCE OF KENYA.................... 5
SNOW PEA AND SUGAR SNAP PRODUCTION IN MT. KENYA REGION: STATUS, CHALLENGES, AND OPPORTUNITIES ....................................................................................................... 5
POST HARVEST HANDLING OF CUT FLOWERS..................................................................................... 10
INTERACTIONS IN PLANT DISEASE EPIDEMICS .................................................................................... 17
DIAMONDBACK MOTH, PLUTELLA XYLOSTELLA (L.), ON PEAS IN KENYA: IMPACT OF THE HOST SHIFT ON THE PEST AND ITS PARASITOID ................................................................... 17
PHAEORAMULARIA LEAF AND FRUIT SPOT OF CITRUS: A MAJOR THREAT TO CITRUS PRODUCTION IN TROPICAL AFRICA, PRESENT STATUS AND FUTURE PERSPECTIVES................. 18
RAPHANUS RAPHANISTRUM L AS A DIVERSIONARY HOST OF BREVICORYNE BRASSICAE L AND BAGRADA CRUCIFERARUM KIRK................................................................................................... 25
TOWARDS SUSTAINABLE CROP PROTECTION THROUGH UTILISING FAVOURABLE CULTURAL PRACTICES: POTENTIAL IN VEGETABLE CROPS............................................................. 30
POTENTIAL FOR ENHANCING THE IMPACT OF AUGMENTATION BIOCONTROL OF TWO KEY CATERPILLAR PESTS ON IMPORTANT VEGETABLE CROPS IN KENYA .................................... 31
HOST PLANT RESISTANCE IN OKRA (ABELMOSCUS ESCULENTUS (L) MOENCH:MALV ACEAE) VARIETIES. .................................................................................................................................................. 32
SOME ASPECTS OF SUSTAINABLE TOMATO PRODUCTION ............................................................... 36
RESPONSE OF TOMATO (LYCOPERSICON ESCULENTUM L.) GENOTYPES TO INDUCED SALT STRESS ....................................................................................................................................................... 37
GENETIC TRANSFORMATION OF TOMATO LINES FOR RESISTANCE OF CUCUMBER MOSAIC VIRUS........................................................................................................................................................... 38
EFFECTS OF DURATION OF SOAKING AND CONCENTRATION LEVEL OF SALICYLIC ACID ON SEED GERMINATION OF CABBAGE (BRASSICA OLERACEA VAR. CAPITATA L.), CUCUMBER (CUCUMIS SATIVUS L.) AND TOMATO (LYCOPERSICON ESCULENTUM MILL.)................................. 39
EFFECT OF IRRIGATION MANAGEMENT ON YIELD, WATER AND NITROGEN USE OF OKRA (ABELMOSCHUS ESCULENTUS L.) .......................................................................................................... 45
GENETIC IMPROVEMENT OF VEGETABLE CROPS FOR SUST AINABLE HORTICULTURAL PRODUCTION.............................................................................................................................................. 53
PROPAGATION OF KEI APPLE (DOVYALIS CAFFRA WARB.); TPRI NURSERY EXPERIENCE.......... 54
SELECTION OF MACADAMIA ROOTSTOCKS LEAST SUSCEPTIBLE TO CHLOROSIS AND NECROSIS ................................................................................................................................................... 62

Wesonga, et al., 2002. Procedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics. October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya iii
SNAP BEAN IMPROVEMENT IN KENYA ................................................................................................... 65
EFFECT OF EXPLANT TYPE ON GUS GENE EXPRESSION AND SHOOT REGENERATION FROM BRUSSELS SPROUTS AFTER EXPOSURE TO AGROBACTERIUM TUMEFACIENS................. 68
THE EFFECT OF PREPARATION METHODS ON THE NUTRITIONAL QUALITY OF SOME VEGETABLES CONSUMED IN KENYA...................................................................................................... 75
PRINCIPLES OF SUSTAINABLE NUTRIENT MANAGEMENT IN VEGETABLE PRODUCTION ............. 80
RATIONALIZATION OF NUTRITIONAL REQUIREMENTS FOR SUSTAINABLE COFFEE PRODUCTION IN KENYA............................................................................................................................ 81
AFRICAN INDIGENOUS VEGETABLES-OPPORTUNITIES AND CONSTRAINTS................................... 81
INFLUENCE OF WATER LEVEL AND GENOTYPE ON TRANSPIRATION OF SPIDER PLANT AND BLACK NIGHTSHADE IN POT EXPERIMENTS ................................................................................ 92
LABORATORY METHODS IN SCREENING CROPS FOR ALUMINIUM TOLERANCE............................ 97
POSSIBLE ROLE OF DAY LENGTH ON GRAPEVINE PERFORMANCE IN THE TROPICS ................... 97
STORAGE AND PROCESSING POTENTIALS FOR HORTICULTURAL PRODUCE IN KENYA.............. 98
COMPARATIVE STUDY OF BANANA (MUSA SPP.) PROPAGATION METHODS AT FARM LEVEL IN KENYA ...................................................................................................................................... 105
FARMING CHARACTERISTICS AND CONSTRAINTS AMONG SMALL SCALE FRENCH BEAN (PHASEOLUS VULGARIS) FARMERS IN MWEA, KENYA...................................................................... 106
PARTICIPATORY PROMOTION OF HORTICULTURE PRODUCTION IN RIFT VALLEY AND WESTERN KENYA .................................................................................................................................... 111
SOCIO-ECONOMIC ANALYSIS OF THE CONSTRAINTS TO ADOPTION OF COMMERCIALIZED SMALLHOLDER HORTICULTURAL PRODUCTION SYSTEM IN THE RIFT VALLEY REGION: A CASE OF NANDI AND UASIN GISHU DISTRICT OF KENYA.............................................................. 112
INCIDENCE OF STRIGA HERMONTHICA INFESTATION DEPTHS ON STRIGA INFECTION INDICES AND MAIZE YIELD COMPONENT............................................................................................. 113
ROLE OF ORGANIC SOIL AMENDMENTS IN THE MANAGEMENT OF ROOT-KNOT NEMATODES ON OKRA ........................................................................................................................... 113
CURRENT FRENCH BEAN PESTS AND DISEASE MANAGEMENT AT MWEA-TEBERE, CENTRAL KENYA ..................................................................................................................................... 118
APPLICATION OF REMOTE SENSING TECHNIQUES IN MANAGEMENT OF BIOTIC CROP STRESS .......................................................................................................................................... 123
USE OF NATURAL PESTICIDES FOR PEST AND DISEASE MANAGEMENT IN VEGATABLE PRODUCTION UNDER SMALL SCALE FARMS...................................................................................... 123
SCREENING OF THREE STRIGA HERMONTHICA STIMULATING FLUORESCENS BACTERIA FOR PHYTOTOXICITY IN COWPEA......................................................................................................... 123
EXPRESSION OF RESISTANCE IN. BRASSICA RAPA SP. INFECTED WITH PLASMODIOPHORA BRASSICAE WOR..................................................................................................................................... 124
VEGETATIVE PROPAGATION OF MACADAMIA NUT (MACADAMIA INTEGRIFOLIA, (M. INTEGRIFOLIA * M. TETRAPHYLLA) HYBRIDS). ............................................................................ 128
THE EFFECT OF DIFFERENT SOILESS SUBSTRATE MIXTURES ON THE ROOTING OF HYPERICUM (HYPERICUM ANDROSAEMAM) ....................................................................................... 131
PROPAGATION OF LILY BULBS FROM SCALES .................................................................................. 132

Wesonga, et al., 2002. Procedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics. October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya iv
GROWTH OF MANGO ROOTSTOCK SEEDLINGS UNDER DIFFERENT WATER REGIMES............... 137
PRELIMINARY STUDIES ON GRAFTING OF TOMA TO (LYCOPERSICON ESCULENTUM MILL.) ON SOLANUM INCANUM L. ..................................................................................................................... 137
TOMATO GRAFTING: OPTIMISATION OF THE TECHNIQUE AND ENVIRONMENTAL CONDITIONS FOR GRAFT SUCCESS ..................................................................................................... 137
EFFECTS OF FORCING METHODS AND ROOT PRUNING ON THE GROWTH OF BUDDED ‘ROUGH LEMON’ ROOTSTOCKS ............................................................................................................ 138
GROWTH RESPONSES OF 'ROUGH LEMON'(CITRUS SINENSIS L.) ROOTSTOCKS TO DIFFERENT CONTAINER SIZES AND IRRIGATION FREQUENCY........................................................ 144
EFFECT OF CONTAINER VOLUME AND NITROGEN APPLICATION ON THE GROWTH OF YOUNG CITRUS SEEDLINGS................................................................................................................... 151
ACHIEVEMENTS IN ROOT AND TUBER CROPS IMPROVEMENT RESEARCH AND FUTURE PROSPECTS IN SOUTHWESTERN ETHIOPIA........................................................................................ 158
AGRONOMIC STUDIES FOR SUSTAINABLE PINEAPPLE (ANANAS COMOSUS) PRODUCTION IN SOUTHWESTERN ETHIOPIA............................................................................................................... 159
EFFECT OF NITROGEN FOR SUSTAINABLE VEGETABLE PRODUCTION: RECENT DEVELOPMENTS ...................................................................................................................................... 159
THE PARTITIONING OF ASSIMILATES IN FABA BEAN: A COMPARISON OF THE DETERMINATE AND INDETERMINATE GROWTH HABIT...................................................................... 160
THE EFFECT OF SILICON AND MANAGANESE ON COWPEA (VIGNA UNGUICULATA) LEAF WATER LOSS.................................................................................................................................. 169
ROLE OF MYCORRHIZAL FUNGI IN THE SUSTAINABLE HORTICULTURAL CROP PRODUCTION IN THE TROPICS .............................................................................................................. 169
ISOLATION AND TESTING OF LOCAL COMMON BEAN RHIZOBIUM FOR IMPROVED N SUPPLY THROUGH NITROGEN FIXATION. ........................................................................................ 170
VARIETY EVALUATION AND IMPROVED AGRONOMIC PRACTICES FOR GINGER (ZINGIBER OFFICINALE ROSC.) IN ETHIOPIA ....................................................................................... 178
CHICKPEA ADAPTATION TO KENYA’S NORTHWESTERN HIGHLANDS............................................ 178
THE EFFECT OF MILD POSTHARVEST CHILLING ON THE EATING QUALITY OF BANANAS.......... 181
POLLINATION CONSIDERATIONS IN SUSTAINABLE INTEGRATED HORTICULTURAL PRODUCTION SYSTEMS OF FRUIT AND SEED VEGETABLES IN THE TROPICS.............................. 186
AGRO-METEOROLOGICAL INFLUENCES IN COFFEE PRODUCTION- IMPLICATIONS FOR IMPROVED COFFEE PRODUCTION IN KENYA ...................................................................................... 190
BIO-ALGEEN: ECONOMIC BENEFITS WITH ENVIRONMENTALLY FRIENDLY PRODUCTS AND PRODUCTION ................................................................................................................................... 191
OFFICIAL CLOSING SPEECH .................................................................................................................. 192
SEMINAR PROGRAM................................................................................................................................ 194
LIST OF PARTICIPANTS........................................................................................................................... 198

Wesonga, et al., 2002. Procedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics. October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya v
PREFACE Horticultural sector remains an important foreign exchange earner to Kenya and contributes significantly in the local diets. Fresh horticultural crops often identified as commodity group for which there is high demand, are of value nutritionally and have potential for local and export markets. Development of this sector will stimulate economic growth as well as provide employment opportunities given the relatively high premium price attached to this crop. Due to growing competition for both domestic and export markets, growers require technologies, which will exert less pressure on the available inputs, environment and guarantee good health. As a consequence, there is need to strongly develop technological options for sustainable production and post-harvest handling. The main objective of the seminar was to bring together the stakeholders in the horticultural industry with a view to strategize and develop a framework for solving the problem of the small-scale farmers. The seminar addressed among other topical issues ways and means of sustaining horticultural production in the tropics under the following themes:
i) Socio-economic issues ii) Plant Propagation iii) Nursery management iv) Plant nutrition v) Plant protection and plant pest systems vi) Crop improvement vii) Crop physiology viii) Postharvest ix) Intergrated horticultural production systems
During the first two days, the scientific exchange was accomplished through oral and poster presentations at Jomo Kenyatta University of Agriculture and Technology, (JKUAT). These discussions continued on field trips on days 3 and 4 to horticultural production sites in Kenya. The Seminar Organising Committee would like to thank all the authors contained in these proceedings. We would also like to express our appreciation to all the individuals who singly or collectively contributed to the organization and the ultimate success of this seminar. Very special thanks go to the German Academic Exchange Service (DAAD) without whose financial support the seminar organization would be a non-starter. We are also indebted to the University of Hannover for the collaboration that made the workshop a great success. We greatly thank the chairpersons of the sessions, rapporteurs, the editorial staff and other Horticulture departmental staff members for their various contributions. Lastly but not least, we wish to sincerely acknowledge the support received from the Vice-Chancellor, Prof. R.W. Michieka, Deputy Vice Chancellors, Prof. S. G. Agong, Prof. H. Thairu and Prof. S.K. Sinei. We also record the support that we received from the Ministry of Agriculture and Rural Development, especially the Deputy Director In charge of Horticulture, Mr. Frederick P. Muema who represented the Minister on the official closing of the seminar.

Wesonga, et al., 2002. Procedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics. October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya vi
Organisers
Jomo Kenyatta University of Agriculture and Technology (JKUAT) Department of Horticulture P.O. Box 62000, Nairobi, Kenya Phone +254 151 52711 Fax +254 151 52164 e-mail: [email protected]
University of Hannover Institute of Vegetable & Fruit Science and Institute of Plant diseases & Plant Protection Herrenhaeuser Str. 2 D-30419 Hannover Phone +49 511 762 2634 Fax +49 511 762 3606 e-mail [email protected]
March, 2002

Wesonga, et al., 2002. Procedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics. October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya vii
OFFICIAL OPENING SPEECH SPEECH BY PROF. S. G. AGONG DEPUTY VICE CHANCELLOR (ADMINISTRATION, PLANNING AND
DEVELOPMENT), JKUAT, ON THE OFFICIAL OPENING OF THE HORTICULTURE SEMINAR ON SUSTAINABLE HORTICULTURAL PRODUCTION IN THE TROPICS OCTOBER 3rd, 2001
The Vice Chancellor, JKUAT, Directors of Institutes and Chairmen of Departments, Distinguished Guests, Ladies and Gentlemen It gives me immense pleasure to be with you today and to preside over the opening occasion of the 3rd National Workshop, which focuses on Sustainable Horticultural Production in the Tropics. According to the Kenya Economic Survey 2001, the agricultural sectors' Gross Domestic Product (GDP) growth note declined from 1.2% in 1999 to negative 2.4% in 2000. The value of horticultural exports declined by 2.1% from Kshs.14, 200 million in 1999 to Kshs.13, 900 million in 2000. The volume of horticultural exports increased marginally 0.2% in 2000. The local value of production was about 3 times as much as the mentioned. The marginal growth is attributed to the decreased output by small-scale producers as a result of inadequate rainfall in the last two years. The Governments financial statement increased budgetary allocation on extension service and from September 2001, all services to the horticultural industry will be exempted from Value Added Tax (VAT). At the local level, this industry attracts substantial encouragement from the Government. However, internationally it has to contend with trade barriers and stiff competition. The government machinery could enhance its functioning further by e.g. improving infrastructure especially in the rural areas and providing incentives to strengthen local self help groups/cooperatives. The Kenya Government is in the process of finalizing proposals for horticultural reforms following recommendations of a horticultural review task force appointed in 1999. When passed, a Bill is expected to provide for the development, promotion and coordination of the industry and establish the Horticultural Crops Development Authority (HCDA). This Authority will define standards of production, handling transportation and storage. Invariably, this will ensure that the horticultural industry gets its rightful recognition as a key player in the country's economy. Ladies and Gentlemen, I appreciate that you are from different disciplines and regions. The wealth of knowledge and experience is vast and it will contribute immensely to ways and means of sustaining horticultural production in the tropics. Your four day workshop will have succeeded if systems that arise from your discussions contribute to enhanced environmental quality and resource base, ensure food security are economically sound enhancing the quality of life for the community. Most of the participants are from universities, which have a primary mission of training, research and extension. It would be prudent to find ways and means of passing the knowledge gained in the next few days to the users - basically the small-scale farmers. Develop partnerships with the farmers and educate them on better and controlled use of chemical inputs, protection of the environment and profitable marketing strategies. In this way, you will have contributed significantly to poverty reduction. The intentions of workshops are continually defeated by ineffective implementation. This country has brilliant

Wesonga, et al., 2002. Procedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics. October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya viii
policies for encouraging this important sector of the economy. The follow up is deterred by insufficient capital. The capital base will be enhanced even as the horticultural sector redefines its business strategy to tap the opportunities presented by business. You have access to technology that is not available to the farmers. Harness knowledge but use it to benefit the overall development of the nations you represent. Finally, I wish to that the sponsors of this workshop, that is, the German Academic Exchange Service (DAAD), Jomo Kenyatta University of Agriculture and Technology and University of Hannover. I wish you fruitful deliberations. With these remarks, Ladies and Gentlemen, it is now my pleasure to declare this workshop officially open. Thank you.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 1
MUSHROOM PRODUCTION IN SUSTAINABLE SMALL-SCALE FARMING SYSTEM - OPPORTUNITIES AND CONSTRAINTS: A SURVEY OF UASIN GISHU DISTRICT Buigut, S. K.
Abstract Kenya’s per capita food production including dietary protein is on the decline. A rapid population growth has put pressure on arable land, resulting in continuous cropping or reduced fallows. Cost constraints the use of inorganic fertilizers by small farmers, leading to declining land productivity. To reverse the trend, technologies that enhance productivity of farm resources and reduce waste are needed. One way of increasing farm resource productivity is to make more efficient use of the abundant agro by products. In the western highlands of Kenya cereal residue, especially maize stover and wheat straw is generally burnt to facilitate land preparation for subsequent crop. Only a little is fed to livestock. Edible mushroom production is an efficient bioconversion technology. It is able to utilize this low quality cereal residue to produce high quality protein. Edible mushrooms are a source of high quality protein (about 19% - 35% protein) with all essential amino acids and are of medicinal value to man. In addition the spent compost is a good soil conditioner and bio-fertilizer. It also fetches good prices - currently 500 Ksh to 600 Ksh/kg in Eldoret. Mushroom production in the western highlands of Kenya has not taken root. A survey of Uasin Gishu district shows that after the exit of EATEC, there are only three small-scale producers in the district. This crop has not been actively promoted in the past by the ministry of agriculture. However, any effort to promote it has to take into consideration several limitations. The first is technical. The ministry of agriculture technical staff in the district is not trained in mushroom production techniques. Only one SMS has had training. At the current prices of 500Ksh/kg – 600Ksh/kg it puts it out of range of most consumers locally. There is need to look at possibilities of accessing external markets. This technology also needs to be vigorously promoted. It is not a technology that can easily diffuse from farmer to farmer. The existing producers (and EATEC previously) pursue an active exclusion principle.
Introduction Kenya’s per capita food production is on the decline (World Bank, 1996; FAO, 1998). A more acute problem than the general decline in food consumption per capita, though more subtle in its manifestation, is the protein deficit. Per caput dietary supply decreased from 64g/day in 1969-71 to 52 g/day in 1990 period (FAO, 1996). From 1970 to 1996, the decline in per caput dietary protein supply is about 19.5%. A number of socio-economic factors viz; a rapid population growth, large rural population, low incomes, illiteracy and a stagnant agricultural technology counter attempts to improve the situation. Rapid population growth estimated at 3.4% for the period 1975 – 1997 (UNDP, 1999) has put enormous pressure on prime arable land. The result has been reduced land holdings, soil fertility decline from continuous cropping or reduced fallows, and untenable utilization of marginal lands. More mouths to feed and decline in land productivity have reduced nutritional level of the farm families. Secondly the amount of surplus agricultural production from small-scale farm system has declined. This in turn has depressed the level of farm income and exacerbated the situation at the farm household level. To reverse the trend, technologies that enhance productivity of farm resources and reduce waste are required. A traditional approach has been the development of higher yielding crop varieties as in cereals. Granted, HYCVs will further increase the productivity of land. But generally, experts believe a tenfold increase of yields is not in sight from genetic engineering and biotechnology as in the first green revolution (Pauli, 1996). In addition, in depleted soil, the full potential of HYCVs is realized only with adequate application of external inputs such as inorganic fertilizers and chemicals. Restricted use of external inputs, and dependence on fallows for maintaining soil fertility and weed control is cited as one of the major constraints to raising productivity in African

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 2
agriculture (FAO, 1986). Most farmers in Africa appreciate the value of fertilizers, but seldom apply them at the recommended rates and at the appropriate times because of high costs, lack of credit, delivery delays and low and variable returns (Sanchez et al, 1997). Three requirements for increasing agricultural production have been identified: x) An enabling environment for small-holder farming sector (infrastructure, education, credit,
inputs, markets and extension services). xi) Reversing soil fertility depletion, and xii) Intensifying and diversifying land use with high value products (Sanchez and Leakey,
1997). At the farm level it makes sense for farmers to reverse the nutrient mining to the point where the marginal costs of nutrient replenishment are covered by marginal benefits. This means the value of increased production brought about by fertilizer should be sufficiently high to cover cost of the fertilizers, compensate for the risk, and provide a reasonable return to the farmer. Fertilizer application by smalll-holder farmers to food crops is often not profitable due to combination of high fertilizer prices, low food crop prices and high risk. Even when fertilizer application is profitable, many farmers cannot afford to purchase fertilizer at the beginning of the season when other basic needs are pressing. Currently research efforts are in the search for alternate organic matter technologies for nutrient management. These include use of plant residues, green manures, and deep-root nutrient recycling systems. The latter two is where agro-forestry is making its contribution in soil fertility management (Buresh and Tian, 1998). However the use of organic materials to increase the carbon reserves of soil under tropical conditions require large amounts of annual additions. Another way of increasing farm resource productivity is to make a more efficient use of the abundant agro by-products and to reduce farm waste to a minimum. Currently in much of Kenya – especially the western highlands, the farming system practiced contributes to the nutrient loss. Crop residues are not returned to the soil. Many farmers use the long stems of crops such as maize stover as building materials or fuel. A sizeable proportion of crop residue gets burned up in the process of land preparation. Only a tenth of the residues and weeds left on the fields after harvest is eaten by livestock (ILRI, 1999). A focus is therefore needed on increasing the productivity of agro by-products. The success of the second green revolution will depend on a scientific research program that focuses on the complete use of all the biomass. One such technology to utilize organic waste is edible mushroom cultivation. Mushroom production provides a number of opportunities for improving the sustainability of small-farming system and rural development. Edible mushroom production is a particularly effective form of bioconversion technology. Edible mushrooms are a source of high quality protein - about 19% to 35% - (Aletor, 1995; Longvah and Deosthale, 1998) with all essential amino acids, and are of medicinal value to man (Chiang and Mshigeni, 1997; FAO, 1983; Oei, 1991). Most of the medicinal extracts from mushrooms are polysaccharides. These extracts are strengtheners of immune system, with little or no side effects. The great attraction of mushroom production is that it can be based on a wide range of agricultural by–products (cereal straw, sisal waste, sawdust, cotton waste, and water hyacinth). These are widely abundant and of little on no value – at least in their original form (Oei, 1991; Murugasen et al, 1995). These materials are mostly resistant to natural biodegradation because they contain mainly cellulose, hemicellulose and lignin. Mushroom mycelia excrete enzyme complexes that can degrade these components. Integrating mushroom production into the farm system will convert these otherwise waste products into high quality protein. This will help alleviate the protein deficit and improve farm incomes.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 3
Currently mushrooms fetch good prices (about 500K.Shs. – 600K.Shs. in Eldoret). This will improve farm incomes and therefore their ability to purchase required inputs. The spent compost left after harvesting of mushroom is a good soil conditioner and bio-fertilizer that can be used in crop production to reduce the required inorganic fertilisers. This will lead to a more sustainable farming system. Steward et al (1998a) found that the application of spent mushroom substrate caused a rapid increase in soil inorganic N concentration, increased both soil pH and CEC, whereas inorganic fertilizers decreased both. It also improves the soil physical properties. Application of spent mushroom substrate improves the environment for plant root development by decreasing soil density, increasing aggregate stability, reducing clod and surface crust formation, and increasing infiltration level (Stewart, 1998b). Spent compost is also a highly nutritious fodder for animals such as poultry (Zhang et al, 1995). Although production of this crop has not developed well in the western parts of the country, a number of communities have harvested wild mushroom for food in the past. Overall little amounts of mushrooms are being cultivated in Africa. Oei (1991) cites several reasons for neglect of mushroom in developing countries: comparatively little scientific study has been done on tropical mushrooms, literature on mushroom growing is expensive and not aimed at developing countries, unavailable technical skills to produce spawn and suitable strains are hard to find. A survey of mushroom production in Uasin Gishu district identified several constraints. After exit of EATEC (East African tanning and extract company), there are only a few (3) small-scale producers in the district. The ministry of agriculture has not actively promoted the crop in the past in the extension packages provided to the farmers or field days. Interested farmers have had to seek the technology on their own. The ministry of agriculture staff is also ill equipped to promote this technology. Only one of the SMS has had training in mushroom production. Another constraint is the market. Currently consumer price in Eldoret town is about 500Ksh - 600Ksh/kg. This puts it out of the range of most local consumers. It cannot compete with other protein sources like beef, beans or eggs for a place in the average family diet list. May be the most serious problem to the promotion of the technology is the exclusion principle adopted by the current producers. Most technologies diffuse from farmer to farmer. The survey noted however that the existing farmers (EATEC too previously) pursue an active exclusion principle. They are not willing to allow other farmers to visit their production sites. And since it is an in-the-house technology it is easy to exclude others. The number of existing producers is also too small to allow much unaided diffusion of the technology.
Helping small farmers make the transition. Mushroom production is generally more input intensive than traditional field crops. Further profitability is more dependent on meeting the consumers’ demand for freshness and quality. The payoff is higher but the risks are much higher, they will be required to work harder, and learn new skills. They will require more than just opportunity to adopt this technology. They will require a more complete support package, which should include: A government support program to promote private firm participation would be an ideal situation. The government through such a program would provide technical help, and linkage between farmers and private companies. Private companies would provide credit, market expertise, and initially an assured price to the farmer. Such an approach has been tried in Thailand and Taiwan (Benzinger, 1996). In Thailand 1986 to promote transition to high value crops (tomatoes, seed production, watermelon) the government supported the private sector through Agro-Production & Marketing Program (APMP). APMP staff acted as catalysts between firms and farmers – they ran

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 4
demonstration plots, taught farmers production techniques, solicited visits by private firms and shared agro-climatic data. The private firms provided a complete package of support measures, which included risk sharing. The development of mushroom and asparagus industries in Taiwan in the late sixties is another example. Neither crop had been grown in Taiwan. The government supported the development of the two industries through TARI (Taiwan agricultural research institute) financed out of the government budget, the local farmers association taught the farmers techniques, inspection and grading of the mushrooms was done at collecting centres jointly run by representatives of farmers association and contracting canner. All canners acted as a cartel in international markets citing a single price. Taiwan dominated the asparagus market for almost twenty years and was the world’s the single largest exporter of canned mushroom. In Kenya HCDA and FPEAK gave a reasonable initial impetus to the horticultural sector - especially cut flowers. The success of these two programs shows that it is possible to help farmers shift to high value crops through a blend of government and private enterprise. The environment of course will dictate what that optimal mix would be.
Acknowledgements. Much thanks to E. Chekwony, P. Orogo, and A. Odhiambo of the MoA, Uasin Gishu district for their support in the survey.
References. Aletor, V. A. (1995). Compositional studies on edible tropical species of mushrooms. Food
chemistry, 54(3), 265-268. Benzinger, V. (1996). Small fields, Big money: two successful programs in helping small farmers
make the transition to high value-added crops. World development, vol. 24 No. 11 pp 1681 – 1693.
Buresh, R. J., and G. Tian (1998). Soil improvement by trees in sub-Saharan Africa. Agroforestry systems (38), 51-76.
Chiang, S. T., and K. E. Mshigeni (1997). Mushroom production in Africa: Prospects. Discovery and innovation, vol. 9, (3/4). 127-129.
FAO (1998). FAO production yearbook, vol. 51, 1998. Statistical series no 142. FAO of the UN. FAO (1996). The sixth world food survey. FAO of UN, Rome. FAO (1986). Raising productivity. African agriculture: the next 25 years. Annex III. FAO of the UN,
Rome. FAO (1983). Growing mushrooms. Oyster mushroom, jews ear mushroom, straw mushroom. FAO
of the UN, regional office for Asia and Pacific, Bangkok. ILRI (1999). Livestock research for development 4. ILRI Newsletter. Longvah, T., Y. G. Deosthale (1998). Compositional and nutritive studies on edible mushroom from
Northeast India. Food chemistry, 63 (3) 331-334. Murugasen, A.G., G. S. Vijayalokshmi, N. Sukumaran, C. Mariappan (1995). Utilization of water
hyacinth for oyster mushroom cultivation. Bioresource technology. 51 (1) 97-98. Oei, P. (1991). Manual on mushroom cultivation. Technologies, species and opportunities in
developing countries. Tool/CTA, Amsterdam. Pauli, G. (1996). The second green revolution. Discovery and innovation. Vol 8 No. 1, 1-2. Sanchez, P. H., and R. R. B. Leakey (1997). Land-use transformation in Africa.Three determinants
for balancing food security with natural resource utilization. Env. J. Agron. 7: 1-9 Sanchez, P. A., K. D. Shepherd, M. J. Soule, F. M. Place, R. J. Buresh, A. N. Izac, A.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 5
U. Mwokunye, F. R. Kwesiga, C. G. Ndiritu, P. L., and Woomer (1997). Soil fertility replenishment in Africa: an investment in natural resource capital. In (eds) Buresh R. J., P. A. Sanchez, and F. Calhoun. Replenishing soil fertility in Africa. SSSA special publication No 51.
Stewart D. P. C., Cameron K. C., and Conforth I. S. (1998a). Effects of spent mushroom substrate on soil conditions and plant growth in an intensive horticultural system: a comparison with inorganic fertilizer. Australian journal of soil research, 36 (2). 185-198.
Stewart D. P. C., Cameron K. C., Conforth I. S., and Sedcole J. R. (1998b). Effects of spent mushroom substrate on soil physical conditions and plant growth in an intensive horticultural system. Australian journal of soil research, 36 (6). 185-198.
UNDP (1999). Human development report 1999. Oxford University press. New York. World Bank (1996). Social indicators of development. The John Hopkins University Press, London. Zhang, C. K., Gong, F., Li, D. S. (1995). A note on the utilization of spent mushroom composts in
animal feeds. Bioresource technology. 52 (1) 89-91.
CHARACTERISTICS OF FRENCH BEAN (PHASEOLUS VULGARIS) PRODUCTION: A CASE STUDY OF KIBIRIGWI IRRIGATION SCHEME IN CENTRAL PROVINCE OF KENYA Wamocho, L. S., Ombwara, F.K., Makokha, A.O; Wesonga, J.M. and Sichangi, P. Jomo Kenyatta University of Agriculture and Technology, P.O. Box 62000, Nairobi, Kenya Email [email protected]
French bean production by farmers on small-scale holdings has recently developed into a major income generating activity in various parts of Central Kenya. French beans are primarily grown as a cash crop destined for export. Most of the agronomic practices of production are similar to those of field beans, which were traditionally planted and cultivated by women. Due to the economic benefits of intensive French bean production, certain production activities have been relinquished to male farmers. A survey was carried out at the Kibirigwi Irrigation Scheme, to determine the different activities and the constraints affecting production of French beans. Most of the respondents had farms of total acreage of 4 acres and below, although the majority were producing French Beans on less than 1 acre. French bean was the major cash crop grown in the scheme for 92.3% of the farmers. Most resident farmers were females constituting 77% of the respondents, although males controlled the income and provision of farm inputs for the commercial enterprise. The implication of this is discussed in this paper.
SNOW PEA AND SUGAR SNAP PRODUCTION IN Mt. KENYA REGION: STATUS, CHALLENGES, AND OPPORTUNITIES Lydia Njuguna, Joseph Kibaki, Margaret Muchui, Caesar M. Kambo, Susan Munene, Simon B. Wepukhulu,,
Abstract Vegetables play a significant role in our economy through export earnings and as a major component of our diets. Snow/snap pea is an important export crop ranking third after French beans and other beans. The contribution of Snow peas in the export market has grown significantly over the years. In 1993, only 284 metric tons of snow peas were exported. The volumes increased to 3,707 metric tons in 1997, earning the country a total of KES 30 million and KES 378.7 million in 1993 and 1997 respectively (HCDA 1993-2,000 , table 1).. Sugar Snaps exports followed a similar upward trend with 556 metric tons valued at KES 56 million in 1995 and 1,237 metric tons valued at KES 172 million in 1999. However, a drop in volumes and value was recorded from 1998-2000 due to adverse weather conditions (El-nino and the drought that followed). Besides foreign exchange earnings, the crop also creates employment especially for the rural population as well as generating income for the farmers. A small volume of the peas is also consumed locally as immature pods and as shelled peas when mature. Mt. Kenya region is one of the most important snow pea production areas in Kenya. Inspite of the crop's importance, little research has been carried out and limited information has been documented on its status and opportunities in Kenya. Two surveys were conducted in 2000-2001 by a multi-disciplinary research team from the National Horticultural Research Center-Thika to establish the status,

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 6
challenges/constraints and research opportunities in production of the crop in the Mt. Kenya region (Naro-moru area in Nyeri district and Timau area in Meru Central district). Farmers were found to grow more than one variety of snow pea or sugar snap per season depending on the exporters’ preference. Generally the medium-tall indeterminate varieties were preferred over the short determinate ones. Among the important constraints was lack of cultivars adapted to different agro-ecological zones and susceptibility of the commonly grown varieties to the major diseases and insect pests. Important diseases and insect pests included powdery mildew, aschochyta blights, stem rots and fusarium wilt, leaf miners, flower thrips and aphids. A wide range of chemicals was used to control various pests. Both men and women were involved in snow pea production and some of the activities were shared while others were sex specific. High post-harvest losses were reported mainly due to diseases and insect pests, poor pod quality, poor post harvest handling and mechanical damage of the pods. A pod disorder commonly referred to as “pittings” was found to cause up to 80% loss. Marketing was a major constraint with farmers being exploited by exporters, middlemen and brokers who control marketing operations. Technology gaps requiring research intervention were identified.
INTRODUCTION Snow/snap peas rank high among the priority vegetables whose research would generate much income. Output of the Horticultural Crops priority setting workshop of 1996, indicated that a total of KES 2.3 billion could be generated from research on varietal development in peas mainly in the high and high - mid altitudes (snow pea/sugar snaps and garden peas) (KARI, 1996). With a view of developing appropriate research proposals, baseline data was gathered from the field. A bi -faceted approach was adopted in the current study: i) An assessment of the status of snow peas on growers fields. ii) On-farm evaluation of new cultivars to identify superior varieties for different zones and for
tolerance / resistance to powdery mildew and aschochyta which are the major diseases affecting snow peas and sugar snaps, as well as acceptance of the introduced varieties by the farmers, exporters and the consumers.
Two baseline studies were carried out in two seasons in October 2000 and in February 2001 in Timau area of Meru district. On-site assessments were done on small (<1 acre) and medium (1-5 acres) scale farms owned by both contracted and non-contracted outgrowers. Agronomic as well as post-harvest and pest and disease assessments were carried out. Socioeconomic data such as major operations, involvement of gender and costs of various operations, labor distributions and usage of inputs was also captured Seven sugar snaps and 3 snow pea varieties were introduced and evaluated in collaboration with the farmers and exporters in Nyeri (Naromoru) and Meru (Timau) districts.
RESULTS Current Status The farmers in both districts were found to grow different varieties across seasons depending on the exporters’ preferences. Generally, the medium-tall indeterminate varieties were preferred over the short determinate dwarf varieties (tables 2a and b). The commonly grown sugar snap variety was Cascadia and Oregon sugar pod 11 was the popular snow pea variety. Amongst the new varieties introduced, sugar snap varieties Sugar Daddy and Super Sugar were preferred in Timau area while in Naro-Moru Sugar Daddy and Sugar star were preferred. Snow pea variety Snowflakes compared well with Oregon Sugar pod 11 in both districts.
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 7
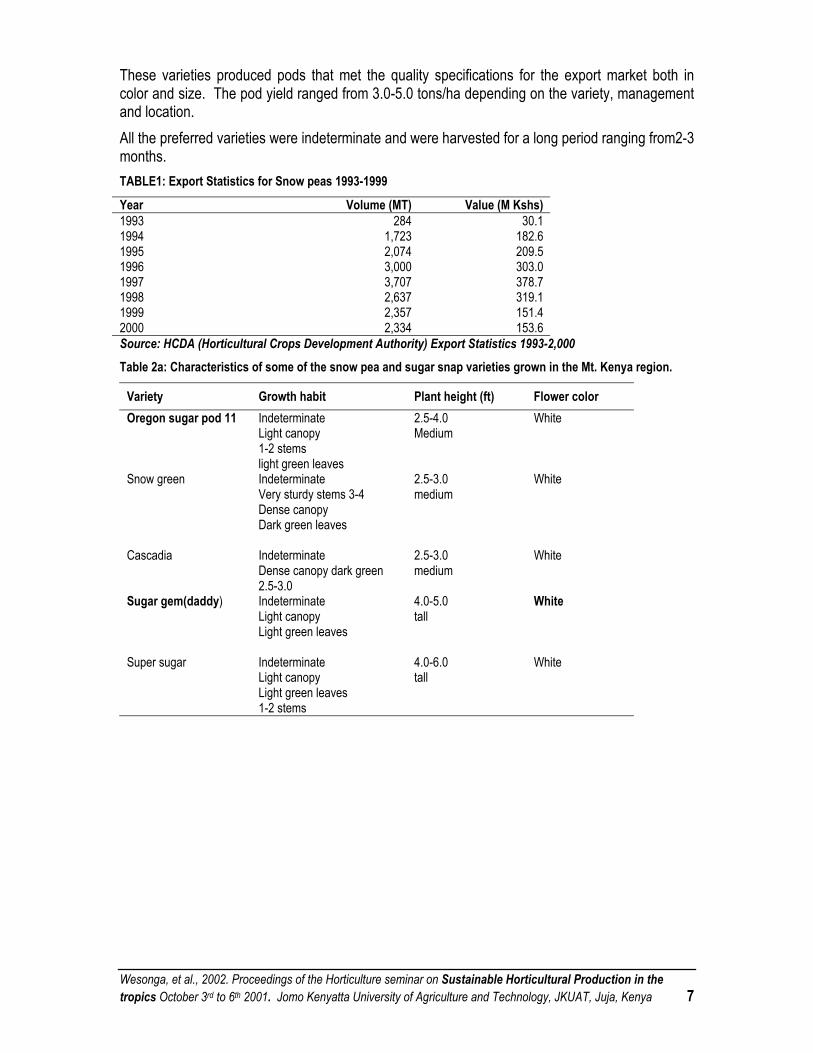
These varieties produced pods that met the quality specifications for the export market both in color and size. The pod yield ranged from 3.0-5.0 tons/ha depending on the variety, management and location. All the preferred varieties were indeterminate and were harvested for a long period ranging from2-3 months. TABLE1: Export Statistics for Snow peas 1993-1999
Year Volume (MT) Value (M Kshs) 1993 284 30.1 1994 1,723 182.6 1995 2,074 209.5 1996 3,000 303.0 1997 3,707 378.7 1998 2,637 319.1 1999 2,357 151.4 2000 2,334 153.6 Source: HCDA (Horticultural Crops Development Authority) Export Statistics 1993-2,000 Table 2a: Characteristics of some of the snow pea and sugar snap varieties grown in the Mt. Kenya region.
Variety Growth habit Plant height (ft) Flower color Oregon sugar pod 11 Indeterminate
Light canopy 1-2 stems light green leaves
2.5-4.0 Medium
White
Snow green Indeterminate Very sturdy stems 3-4 Dense canopy Dark green leaves
2.5-3.0 medium
White
Cascadia Indeterminate Dense canopy dark green 2.5-3.0
2.5-3.0 medium
White
Sugar gem(daddy) Indeterminate Light canopy Light green leaves
4.0-5.0 tall
White
Super sugar Indeterminate Light canopy Light green leaves 1-2 stems
4.0-6.0 tall
White
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 8
Table 2b: Characteristics of some of the snow pea and sugar snap varieties grown in the Mt. Kenya region.
Variety Days to maturity Pod characteristics Yield (Ton/ha)
Oregon sugar pod 11 75-80 Light –green Broad and flat 9.5x2.0cm
3.0-4.0
Snow green 80-85 Dark green Broad and flat 9.0x2.1cm
4.0-5.0
Cascadia 75-80 Round fleshy Dark green 8.0x1.5cm
4.5-5.0
Sugar gem(daddy) 75-80 Round fleshy pods Light green High % of short curved pods 8x1.5cm
3.0-3.5
Super sugar 75-80 Round fleshy pods 8x1.5cm 4.5-5.0
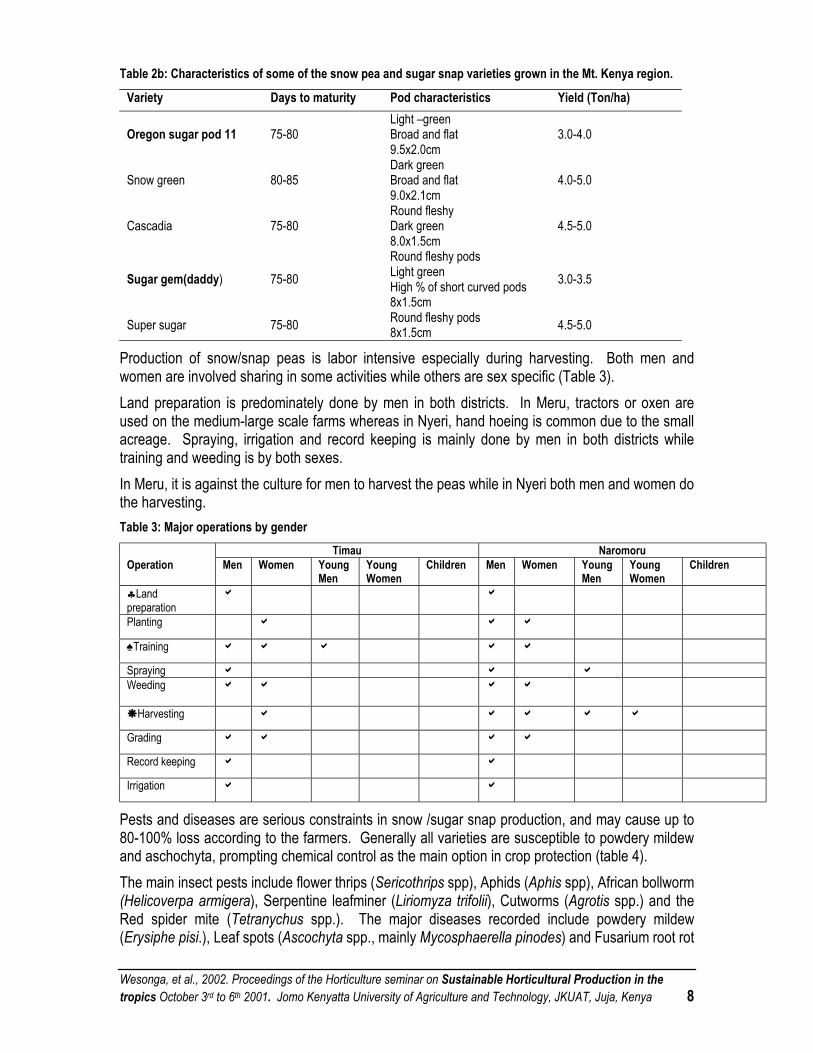
Production of snow/snap peas is labor intensive especially during harvesting. Both men and women are involved sharing in some activities while others are sex specific (Table 3). Land preparation is predominately done by men in both districts. In Meru, tractors or oxen are used on the medium-large scale farms whereas in Nyeri, hand hoeing is common due to the small acreage. Spraying, irrigation and record keeping is mainly done by men in both districts while training and weeding is by both sexes. In Meru, it is against the culture for men to harvest the peas while in Nyeri both men and women do the harvesting. Table 3: Major operations by gender
Timau Naromoru Operation Men Women Young
Men Young Women
Children Men Women Young Men
Young Women
Children
♣Land preparation
a a
Planting a a a
♠Training a a a a a
Spraying a a a Weeding a a a a
Harvesting a a a a a
Grading a a a a
Record keeping a a
Irrigation a a
Pests and diseases are serious constraints in snow /sugar snap production, and may cause up to 80-100% loss according to the farmers. Generally all varieties are susceptible to powdery mildew and aschochyta, prompting chemical control as the main option in crop protection (table 4). The main insect pests include flower thrips (Sericothrips spp), Aphids (Aphis spp), African bollworm (Helicoverpa armigera), Serpentine leafminer (Liriomyza trifolii), Cutworms (Agrotis spp.) and the Red spider mite (Tetranychus spp.). The major diseases recorded include powdery mildew (Erysiphe pisi.), Leaf spots (Ascochyta spp., mainly Mycosphaerella pinodes) and Fusarium root rot

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 9
(Fusarium spp.). However, the most serious problem is "pimple" on pods popularly known as “pod pitting.” Root knot nematodes (Meloidogyne spp) were observed in some farms and were found to cause considerable crop losses (up to 100% on 2 farms) . Table 4: Pests of Snow peas and current farmers’ management strategies
PEST CONTROL 1. Flower thrips (Sericothrips spp.) Karate, Bulldock, Brigade (Pyrethroids),
Malathion, Dimethoate (Organophosphates) 2. Aphids (Aphis spp.) Dimethoate, karate, Bulldock 3. Serpentine leafminer (Liriomyza trifolii) Evisects, Trigard 4. African bollworm (Helicoverpa armigera) Thuricide HP, Dipel 2X 5. Cutworm (Agrotis spp.) Brigade, Dimethoate 6. Red spider mite (Tetranychus spp.) Brigade, Thiovit 7. Powdery mildew (Erysiphe spp.) Baycor, Thiovit, Wetsulf Anvil 8. Leaf spot (Ascochyta spp.) Milraz, Kocide DF 9. Fusarium root rot, Root knot nematodes (Meloidogyne spp.) Uprooting the infected plants. Furadan 5G
Post-harvest The farmers are aware of the right time to commence harvesting-usually 75-80 days after sowing. They use plastic containers, old baskets (ciondos)and old lessos made into containers. The pods are then emptied into plastic crates and nylon bags. The small scale farmers generally do not have any sheds to sort and grade their produce. However, large scale farmers have the shade nets for this purpose. Initial sorting is done at the farm to remove the malformed, diseased and mechanically damaged pods. In some farms the charcoal coolers are used to pre-cool the produce from the fields. The Everest Exporters Enterprise limited has a cold storage facility that serves its contracted farmers only. The non-contract farmers deliver their produce to collection centers where they sell to brokers. Table 5: Major causes of post-harvest losses on the farms
Cause Estimated loss (%) Diseases –Powdery mildew, aschochyta 10 – 20 Pimple disorder 30 – 90 Insect damage (thrips, bollworms) 30 – 60 Mechanical damage 5 – 10 Overgrown, undersized and twisted/malformed pods 5 Damage by birds 2 Overall loss estimates 40 – 90
CHALLENGES/CONSTRAINTS IN PRODUCTION OF SNOW PEAS AND SUGAR SNAPS INSECT PESTS AND DISEASES High cost of chemicals and their unavailability The maximum residue level requirement by the market. Pod pimple disorder.
MARKETING Fluctuating prices Exploitation by brokers

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 10
High post-harvest loses Dishonoring of the contracts
PLANTING MATERIAL Poor seed quality- (mixed with garden peas and poor germination) High cost of the seeds.
CULTURAL MANAGEMENT Inadequate irrigation water Shortage of labor Inadequate information on: nutritional requirements and plant density.
RESEARCH OPPORTUNITIES Develop appropriate agronomic packages for the crop. Introduce and screen new varieties for resistance to major diseases and insect pests. Develop integrated crop and pest management packages. Develop bio-pesticides for the control of major pests and diseases. Establish the causal agent of pod pimple disorder and its control. Develop post harvest packages to reduce losses.
- Technology transfer on proper harvesting and proper handling of the produce. - Appropriate means of pre-cooling the produce at the farm to reduce losses due to wilting. - Value addition/product development. - Assess the economic viability of snow pea and sugar snap production.
References KARI 1996, priority setting workshop for Horticultural crops. HCDA 1993-2000, Export Statistics for Horticultural crops .
POST HARVEST HANDLING OF CUT FLOWERS Theophilus M. Mutui Department of Horticulture, Moi University, P.O.Box 1125, Eldoret, Kenya, Email: [email protected]
ABSTRACT Post harvest handling procedures of cut flowers involves those activities that are performed from the time they are harvested to the final stages within marketing system. Conscientious attention to flower production is of no benefit if the grower's post harvest handling practices are improper. However, post harvest handling practices of horticultural produce are generally poorly developed, not only in Kenya but this is the situation in the whole of sub-Saharan region. The process of harvesting, packaging and transportation often results in substantial mechanical damage and alteration of the gravitational orientation of the produce. Additionally, cut flowers are frequently subjected to substantial physical pressure, altered temperature regime and undesirable gaseous environment. Overall, a living plant material is typically subjected to very harsh treatment during its post harvest life. After incurring high costs during production, it is only sensible that growers learn a few fundamentals of post harvest handling so as to maintain or even increase the value of their crops and reduces post harvest losses. It is estimated that about 20 % of cut flower losses occur during harvest, handling, storage, transportation, wholesaling and retailing. These losses are excessive and can be reduced by paying greater attention to careful handling that more often than not, make the difference between profit and loss. Therefore this paper describes post harvest handling practices and procedures that spans from harvesting, grading, sanitation, use of preservatives, better temperature management in storage rooms and marketing channels, standardization and quality control during the marketing process to meet the consumer's aesthetic satisfaction.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 11
INTRODUCTION Status The history of cut flower production in Kenya dates back to 1960's or early 1970's when former British settlers introduced them. These years saw the export of cut flowers from Lake Naivasha and this has grown over the years. Kenya has an advantage over many countries due to her geographical location and Agro-climatic diversity, which allows for a year-round production of a wide range of cut flowers (Mutui, 1999). Cut flower production in Kenya is done by a few large scale growers who account for 80 % of total exports while 20 % is produced by numerous medium to small scale growers concentrated around Nairobi city and is geared towards export market, although there is a minor element of local consumption in big cities (Mutui, 1999). Floriculture is the science and practice of growing, harvesting, storing, designing, marketing and exporting of foliage and flowering plants as cut or container grown plants (Cathey, 1994). Floriculture is labour intensive and contributes the highest value: volume ratio within the fresh produce sector of the horticulture industry. Hence, it creates employment opportunities for rural youth and women and generates income since income per unit area is high compared to other agricultural produce (Mutui, 1999). In 1999, the volume and value of just the floriculture segment stood at 36,992 tons valued at Ksh 7.24 billion. Expressed as a percentage of total fresh produce exports (98,964 tons valued at Ksh. 14.204 billion), cut flowers accounted for 37.38 % by volume and 50.94 % by value, respectively (HCDA, 1999). The total world imports of cut flowers are estimated at around $ 4,770 million. The European Union (EU) market accounts for some 75-80 % of the total world import trade (White, 1999). Kenya is the leading fresh cut flower exporting country among the African, Caribbean and Pacific (ACP) countries and accounts for 60 % of European Union (EU) imports (Kiptum, 1998). It is the world's fourth supplier of cut flowers to the EU bloc due to her reputation in supplying consistently top quality cut flowers. The principal cut flowers for export are roses, carnations, statice, Alstroemeria, cut foliage, solidaster, eryngium, chrysanthemums, carthamus and ammi majus (Kiptum, 1998). Germany is by far the largest single flower import market with imports in 1997 estimated at $ 1,769 million. This is because over 90 % of the German households buy flowers on regular basis (White, 1999). In 1996, Germany was the leading importer of Kenyan cut flowers accounting for 30.3 % of all EU imports (Kiptum, 1998). On the other hand, the Netherlands dominates the world export trade with an estimated market share of 65 % (White, 1999). The demand for cut flowers is highest during winter season (September to March) when flower production in Europe is expensive due to supplementary heating and lighting of greenhouse. Also the demand is influenced by special occasions like Valentine day, Mother's day, Christmas and New Year holidays (Mutui, 1999). Overview Post harvest physiology is the division of plant science that deals with functional processes in the plant material after it has been harvested (Kays, 1991). It is concerned with plants or plant parts that are handled and marketed in the living state including cut flowers, fruits and vegetables to mention but a few. Post harvest physiology deals with the time period from harvest or removal of the plant part from its normal growing environment to the ultimate utilization, deterioration or death. For cut flowers and cut foliage, their utilization involves displaying and maintaining them for decorative purposes. When pre harvest and harvesting factors have a direct influence on post harvest responses, they are also considered to be vital components of the complete post harvest picture (Kays, 1991).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 12
Pre harvest factors such as optimum growing conditions, proper nutrition and high light intensity are important to post harvest flower quality since they determine the content of starch or sugar stored in the stems, leaves and petals that are necessary for respiration and for maintenance of turgidity during flower development. High quality flowers should have high total solid matter content as contrasted with soft blooms that usually develop under conditions of forced growth (Evans and Reid, 1991). Cut flowers compared with flowers in the field or greenhouses are from a physiological point of view in an unfavourable, precarious conditions (White, 1999). Cut flowers lack means to excrete waste materials, lack uptake of water and nutrients via the roots and sufficient light to assimilate these nutrients to carbohydrates, proteins, hormones and other basic components. Consequently, cut flowers are highly perishable in nature. This coupled with poor infrastructure, unavailability of market information and post harvest handling facilities leads to a considerable loss (Kiptum, 2001). Post harvest handling procedures of horticultural produce are generally poorly developed not only in Kenya but also in the whole of sub-Saharan region (Mutui, 1999). To address this problem, the Government of Kenya has recently (2001) commissioned Horticultural Produce Handling Facilities Project (HPHFP) in line with the implementation of a strategy to address challenges facing the horticultural export industry. The objective of this project is to reduce post harvest losses through rapid removal of field heat, providing a cool-chain link right from harvest to the Auction Center in Nairobi as well as to regulate the market information flow and prices of horticultural produce. Additionally, farmers will be trained on improved post harvest handling methods such as grading, packaging and proper handling of their produce (Kiptum, 2001). A study by FAO approximates post harvest loss of horticultural produce at about 40 % (Kiptum, 2001). Additionally, it is estimated that about 20 % of cut flower losses occurs during marketing as they pass through a number of links in the producer / consumer chain. Losses occur during harvest, handling, storage, transportation, wholesaling and retailing. These losses are excessive and can be reduced by greater attention to careful handling, better temperature management, sanitation and use of preservatives (Hardenburg et al., 1986).
PRACTICES AND TECHNIQUES Harvesting Great care must be exercised during harvesting and field handling to minimize mechanical injuries. Damaged plant tissues produce a lot of ethylene (a gaseous plant hormone) that hastens the onset of senescence. Rough handling of cut flowers results in bruises and injuries that later shows up as brown and black patches making the blooms unattractive and unsaleable (Hardenburg et al., 1986). Additionally, they serve as avenues for microbes that cause spots and rots. High temperatures and relative humidity that are prevalent in the tropical climate favour these microbes. Most successful growers prepare the flowers for market carefully so that the best possible price may be realized. This starts with cutting the flowers at the right stage of maturity. There is a common tendency to cut flowers too soon when the market demand is good and too late when the demand is low (Mutui, 1999). The time of the day when harvesting is done has a bearing on the quality of cut flower. They should be harvested early in the morning when the temperatures are low and plant water status is high (Evans and Reid, 1991). Roses are harvested in the morning and later in the afternoon due to their rapid growth rate. Harvesting is done by hand using sharp blades or knives. At no time should flowers be placed on the bare ground to avoid fungal infection. The cutting tools should be sterilized before using them to avoid transmission of viral diseases or better still use of disposable

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 13
razor blades. However, this increases the cost of production (Evans and Reid, 1991; Hardenburg et al., 1986). The position on the flower stem where the cut is made is very important. In Alstroemeria, stems left uncut or cut through the blanched area of the stem absorbs less holding solution than stems cut through a green tissue (Healy and Lang, 1989). For chrysanthemums, stems should be cut at least 4 inches above the soil line to avoid taking woody plant tissues. In roses, the cut is normally made so as to leave 2 five-foliate leaves below the cut. Cuts that leave only trifoliate leaves may result in weak breaks that may produce blind shoots. After harvesting, the flowers should be place in the shade as soon as possible and if not packed immediately, they should be placed in good quality water and transported to the packing shed or cooler places to remove field heat. Damage through multiple handling can be reduced if all the operations are done in the field. Ideally flowers should be sorted, graded and bunched before being placed in water and transferred to cool-room (Evans and Reid, 1991). Sorting and Grading Cut flowers should not be sorted and graded while placed on the ground but should be placed on sorting tables and be protected from direct sunlight when sorting to discard any weak, diseased, short and malformed stems (Hardenburg et al., 1986). The lower and dirty leaves are removed during cleaning to avoid contamination from bacteria and fungi present in the diseased material. Grading can enhance the quality of cut flowers and fetch higher prices over the cut flowers of the same quality but poorly graded. Flowers must be graded so that those of the same kind and quality are bunched together. The grading standards are based on size of flower, length of stem, strength of stem, straightness of stems, freedom from defects & diseases, maturity, uniformity and foliage quality especially in Alstroemeria (Laurie et al., 1989). Bunching The graded cut flowers are bunched into 10, 20 or 25 blooms depending on the type of the flower and market preference. Bunches are held together by elastic bands or strings and are frequently sleeved immediately to separate bunches, protect the flower heads, prevent tangling and identify the product and grower through labeling. The current trend in the international market is to have a bunch of different kinds of flowers based on the customer's specification, the so-called value added "mixed bouquets" for specific niche market. This saves time that would have otherwise been used in buying different kinds of flowers, sorting and flower arrangements, leave alone the cost involved. The wrapping for the bunch of flowers should provide maximum protection and display. Usually, clear polyethylene (perforated, unperforated and blister) are used since they provide protection, in addition to the flowers being attractively displayed (Evans and Reid, 1991). Pretreatment Pretreatment solutions usually contain sugar (sucrose) as a source of energy, a biocide to inhibit the growth of microorganisms and help prevent bacterial plugging of water-conducting tissues. Additionally, they contain acidifying agent (citric acid) to reduce the pH to 3.0-3.5 that in turn improves the water uptake (Evans and Reid, 1991; Hardenburg et al., 1986). Mutui (1999) found holding Alstroemeria cut flowers in 2.5, 5.0 or 7.5 mg/litre GA4+7 had no effect on the number of days to full opening of the primary florets. Cut stems held at 10.0 mg/litre GA4+7 delayed flower senescence, increased days to 50 % petal fall, leaf nitrogen, chlorophyll, and water content. In the second experiment, 10 mg/litre GA4+7 decreased the dry weight of the leaves at 14 and 21 days after harvest (Mutui, 1999). The cultivar difference was highly significant. `Yellow King' had significantly longer vase life and leaf quality retention than `Marina'. Mutui (1999)

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 14
suggested that 10 mg/litre GA4+7 has the potential to be used as a commercial Alstroemeria flower preservative in order to delay senescence, prolong vase life and post harvest quality of cut flowers. Pulsing cut flowers with Silver thiosulphate (STS) is essential for the flowers sensitive to ethylene such as Sweet Pea, Carnation, Alstroemeria and Snapdragon. Florissant 100R is a commercial preservative that contains STS to counteract the side effects of ethylene. Cut flowers are conditioned by recutting the stems (removal of about 2 cm) and placing them in vases with warm water 38oC to 43oC in 4oC storage rooms for 6 to 12 hours (Evans and Reid, 1991; Hardenburg et al., 1986). Nonmetallic (plastic) containers should be used for holding preservative solutions to avoid any chance of preservative constituents being inactivated by metallic ions. Water cans, buckets, vases, storage racks and rooms need regular cleaning with a good detergent or soap containing a disinfectant to keep microorganisms under control. A chlorine rinse or soak of containers is very effective (Evans and Reid, 1991; Hardenburg et al., 1986). Water quality has an important effect on the keeping quality of cut flowers and decorative foliage. Ideally, the water should contain little salts. Deionised or distilled water is often better than tap water with or without preservatives. Packaging The sales potential for some of the best grown flowers can be ruined simply by the way they are presented for marketing. Bleached white cartons, which are long, flat and easily labeled, are the best. This reduces the depth of flowers in the box, thus reducing mechanical damage. To avoid longitudinal slip of flowers, some packers use one or more cleats. High value specialty flowers (anthurium, ginger, strelitzia, orchids etc.) are packed in various ways to minimize friction damage during transportation (Evans and Reid, 1991). Snapdragons and gladioli are sensitive to gravity and must be stored and shipped upright, to prevent the spikes from exhibiting geotropic bending (curvature) and thus become unsaleable. Low temperature storage reduces but does not prevent bending of horizontally stored gladioli (Evans and Reid, 1991; Hardenburg et al., 1986). Storage Cut flowers have high surface to volume ratio hence they attain the surrounding air temperature fast. Proper temperature control during storage will maintain the inherent high quality of flowers. The most important part of temperature management is to ensure that the flowers are cooled immediately after cutting to remove field heat and after packing by transferring them to a cold storage room. This also reduces the incidences of fungal infections (Mutui, 1999). Therefore, it is mandatory that cut flowers should be stored for only brief periods (for accumulation of enough stocks to warrant shipment or meet high demand as in holidays). Forced air-cooling is the most commonly used method. Cold air is passed through the boxes via end holes or closeable flaps after packaging. Air may be sucked or blown through the boxes with equal success, hence the flowers should be packed in such a way that air could flow right through the box. Packaging of ice with flowers is only effective if the ice packs are placed so as to intercept heat entering the boxes. The optimum temperature for storage of almost all cut flowers is close to 0oC. However, care should be taken when handling tropical flowers like Anthurium, Strelitzia, Cattleya Orchids, Poinsettia and Ginger that are injured by temperatures below 10oC. Usually, the relative humidity is maintained at 90-95 % in the storage rooms to reduce water loss through transpiration (Evans and Reid, 1991; Hardenburg et al., 1986).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 15
Transportation The flowers should be transferred from the cold room and directly loaded into a refrigerated or insulated truck and transported directly to the airport for final shipment. The cooling chain should be maintained at 3-5oC from the grower to the importer. Usually, the flower growers have special arrangements with airlines to ensure availability of cargo space to avoid any delays in airlifting the blooms to Europe. Quality Control At Jomo Kenyatta International Airport (JKIA), the blooms are subjected to thorough inspection by the Kenya Plant Health Inspectorate Service (KEPHIS) inspectors. This is done to ensure that they meet the stringent quality requirements demanded in the world market (Mutui, 1999). Standards have been established for each kind of flower since different qualities are involved. Quality attributes includes: freshness, right stage of development (maturity), uniformity, not diseased or malformed, stem length, strength, straightness, bunch-size, fragrance, sentiment, form, floret development in the spikelet like tuberose and snapdragon, retention of flower colour, lateral bud development especially in spray carnations and maintenance of photosynthetically active foliage (green leaves) as in Alstroemeria, Lilies and Euphorbia fulgens (Hofman, 1988). Marketing Marketing encompasses packaging, pricing and product promotion. Quality is a measure by which a product or service is adapted to the needs, wishes, behaviour and even inconsistent behaviour of the consumers (White, 1999). Proper packaging in terms of design, clarity and truthfulness in labeling can enhance the value of flowers. This has to do with how attractive the package is to the eye the so-called "eye appeal." Lastly, cut flowers are high value products that can easily withstand high freight charges and still sale at a very competitive price. All in all, the most important cut flower quality test is its vase life at the ultimate consumer's destination (Laurie et al., 1980). The Kenya Flower Council in collaboration with Horticultural Crops Development Authority (HCDA) and Fresh Produce Exporters Association of Kenya (FPEAK) has been coordinating participation of private sector to floriculture trade fairs in Holland, Germany and London. The benefits derived from these fairs include advertisements, evaluation of local markets, gathering of market information on competitions ("Market Intelligence") as well as identifying and meeting new potential customers and market niches.
CONCLUSION The Kenyan flowers have become the first choice of European buyers who go for the highest standards of quality and practice. However, every year a percentage of flower consumers in Europe, albeit small get disappointed by their purchases. Therefore it is prudent that the quality of cut flowers should be high and sufficient quantities produced to satisfy demand in order to win and maintain the European Union market. Usually, the consumers want flowers that are fresh, of high quality and that last for a reasonable period. Therefore, cut flowers should be properly handled at all levels to avoid bruising and crushing that will shorten their storage life and reduce their marketability. Post harvest handling procedures are critical in order to achieve high quality produce that meets the consumer's aesthetic satisfaction, so as to ensure that Kenya remains a world leader in the cut flower industry. Consequently, it is only those flowers that are treated with great care can survive the harsh post harvest conditions and give the consumer the beauty and pleasure they paid for.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 16
References Cathey, H. M. 1994. Floriculture. In: Encyclopaedia of Agricultural Science, Vol. 2. C.J. Arntzen
and E. M. Ritter (eds.). Academic Press, San Diego. pp. 205-211. Evans, R.Y. and M. S. Reid. 1991. Post harvest care and handling of greenhouse and field-grown cut
flowers. Ornamentals Northwest Seminars, Portland. pp. 1-19. Hardenburg, R.E., Watada, A.E. and C.Y. Wang. 1986. The commercial storage of fruits, vegetables,
Florist and nursery stock. U.S. Dept. of Agric. Handbook No. 66 (revised). pp. 75-88. HCDA. 1999. Export statistics figures for fresh fruits, vegetables and cut flowers from Kenya by air
and sea [ by volume (MT) and value (KSH)]. Nairobi, Kenya. Healy, W. and D. Lang. 1989. Post harvest handling of Alstroemeria. Hortscience 24(4): 641-643. Hofman, N.C. 1988. The importance of preshipment treatment. International Floriculture Seminar,
Amsterdam. Pathfast Publishing, Essex, England. Pp109-115. Kays, S.J. 1991. Post harvest physiology of perishable plant products. Van Nostrand Reinhold,
NY. 532 pp. Kiptum. B.K. 1998. Marketing of cut flowers. HCDA Horticulture news 20: 6-7. Kiptum. B.K. 2001. An Overview of Horticulture Industry in Kenya including Financing of
Production, Marketing and Export. A paper presented in a Public Forum entitled, "The Potential of Horticultural Production in Rift Valley and Western Kenya Provinces to Exploit the Existing Infrastructure for Domestic and Export Markets" organized by the Faculty of Agriculture, Departments of Horticulture and Agricultural Marketing & Cooperatives of Moi University at Sirikwa Hotel, Eldoret on 30th March.
Laurie, A., D.C. Kiplinger and K.S. Nelson. 1980. Commercial flower forcing. 8th Edition. Mc Graw-Hill, Inc.
Mutui, T .M. 1999. The effect of benzyladenine and gibberellins on the post harvest physiology of Alstroemeria (Alstroemeria aurantiaca) cut flowers. M.Sc. Thesis. University of Nairobi, Kabete. 138 pp.
White, R. 1999. The increasing importance of the supermarkets and other factors in marketing florals. In: Marketing horticultural produce: The changing scene. Roger White & Associates, UK. Proceedings of Hortec 1999 Seminar, Nairobi, 4. pp.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 17
INTERACTIONS IN PLANT DISEASE EPIDEMICS Hau, B. Institute of Plant Diseases and Plant Protection, University of Hannover, Herrenhaeuser Str. 2, D-30419 Hannover, Germany, e-mail: [email protected]
Abstract The simultaneous occurrence of two or more pathogens or pests on the same host is frequent in many production systems, especially in tropical crops. Dealing only with plant diseases, it can be expected that the multiple infections of two pathogens will lead to interactions of the diseases, which may alter the occurrence and the speed of epidemics. In the talk, the disease dynamics of interacting diseases are explained by means of mathematical models using two examples from literature. In the first example, representing an antagonistic interaction, the disease progression of two tobacco virus diseases (TEV and TVMV) is modelled using the Lotka-Volterra competition equations (MADDEN et al., 1987). The speeds of both epidemics, given by the apparent infection rates, as well as the maximum disease levels are reduced in the interaction situation. As a second example, the interaction in disease dynamics of two wheat leaf diseases (Septoria glume blotch and powdery mildew) is modelled by coupled differential equations (WEBER 1996). The Septoria disease lowers the infection rate and the maximum severity of mildew, while mildew raises the epidemic speed and the attainable disease level of glume blotch. Therefore, if both diseases occur together, neither their dynamics, nor the influence of external factors can be understood by neglecting the interspecific interactions. As host growth can change the dynamics of diseases and, on the other hand, diseases affect host growth, the mutual effects of the host plant and the diseases must also be taken into consideration. Without understanding the interactions between host dynamics and progression of diseases, no appropriate yield and yield loss predictions can be achieved. Reliable predictions, however, are essential elements in decision-making in plant protection, needed to reduce the application of pesticides, especially in sustainable production systems.
DIAMONDBACK MOTH, PLUTELLA XYLOSTELLA (L.), ON PEAS IN KENYA: IMPACT OF THE HOST SHIFT ON THE PEST AND ITS PARASITOID Löhr, B. and Rossbach, A. Plant Health Division, International Centre of Insect Physiology and Ecology, P.O. Box 30772, Nairobi, Kenya, e-mail: [email protected]
Abstract Diamondback moth (DBM) was detected in 1999 on sugar snap peas in the export vegetable growing area south of Lake Naivasha in the Rift Valley Province of Kenya. The pea strain (DBM-P) was compared in laboratory studies to the normal crucifer strain (DBM-C). While DBM-P performed equally well on kales and peas, DBM-C survival on peas was only 2.4%. Larval development of DBM-C on peas took five days longer than on kale and larval growth was severely reduced. Pupal weights of DBM-C on pea (3.8mg) were significantly lower than those of DBM-P (4.6MG) and those of both strains on Kale (5.7 and 5.3mg, respectively). Crosses between the strains in both directions were fertile and egg hatch was similar to hatching rates of a laboratory culture on crucifers. The influence of the host shift on Diadegma mollipla, one of the most frequent parasitoids of DBM in Kenyan highlands, was also investigated. DBM larvae on peas as hosts were preferred and parasitation rates were higher, no matter whether the parasitoid population originated from cabbage or pea. When the two host plants were offered in a choice situation, the level of parasitation was as low as when cabbage was offered alone. D. mollipla developed equally well on both hosts, but D. mollipla from cabbage had a longer developmental time on DBM larvae on peas. Larval growth was also dependent on the host plant. Parasitoid cocoons from cabbage were significantly heavier than from peas and there was no difference between the pupal weight of parasitized and unparasitized larvae.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 18
PHAEORAMULARIA LEAF AND FRUIT SPOT OF CITRUS: A MAJOR THREAT TO CITRUS PRODUCTION IN TROPICAL AFRICA, PRESENT STATUS AND FUTURE PERSPECTIVES Mohammed Yesuf Ethiopian Agricultural Research Organization (EARO), Melkassa Agricultural Research Center, P. O. Box 436, Nazareth, Ethiopia, E-mail: [email protected]
Abstract Leaf and fruit spot of citrus caused by a fungus Phaeoramularia angolensis (Cavalho and Mendes) was first reported in Angola and Mozambique in 1952. P. angolensis is now spread to 21 countries, to the south of Sahara, including Yemen in the middle east. This disease can attack all citrus species, grapefruit and sweet orange being the most susceptible. P. angolensis attack leaves, fruits, young shoots and stems. The destructive nature of this disease is severe leaf and fruit drop die-back of young shoots and premature ripening of fruits. Currently, among the recorded diseases of citrus in Tropical Africa, Phaeoramularia leaf and fruit spot is considered the most devastating. In most tropical African countries, where P. angolensis is reported, a total yield loss of 50-100 % is quite common. A complete crop failure has also been reported in severely attacked citrus orchards in Ethiopia mainly on grapefruit and sweet orange varieties. Different management options have been and being advocated in different African countries for the control of this destructive disease. Despite the very high importance and potential threat on citrus production, the available information of this economically important disease is scattered and very limited. The biology and survival mechanism of the pathogen is not yet properly understood. Therefore, based on the available information and experience elsewhere, various aspects of P. angolensis are reviewed, in a view it could direct future collaborative research efforts and serve as source of information for researchers, citrus growers, quarantine services and policy makers as well.
Introduction The citrus industry in Africa is of recent development and attempts have been made to steadily expand citrus production in African agriculture. Production of citrus in Africa constitutes 6% of the total citrus production in the world (Seif 1993). Citrus is mainly a sub-tropical fruit, and among others the major citrus producing countries in Africa are Egypt, Algeria, Morocco, South Africa and Tunisia. These countries are not typical of Tropical Africa since they have in most cases Mediterranean type climate. In most Tropical African countries citrus is planted both by small-scale farmers and commercial or state enterprises for both local consumption and export purposes. In Ethiopia citrus production was started mainly by expatriates. The near Mediterranean climate of the plateau region of Ethiopia creates conducive condition to produce high quality citrus in the country. Among the widely grown citrus species in Ethiopia, sweet orange (Citrus sinensis), lemon (C. limon), mandarin (C. reticulata), grape fruit (C. paradisa) are the most important ones. Citrus in Tropical Africa plays an important role as source of income for small-scale farmers, raw material for citrus industries and nutritional standard for the people. The attempt to increase citrus production in the tropics has not showed too much progress due to several biotic stresses, like diseases and insect pests. Among the recorded diseases so far, leaf and fruit spot of citrus caused by a fungus Phaeoramularia angolensis (cavalho and Mendes) is found to be the most destructive to citrus production in Tropical Africa. Despite the very high importance of P. angolensis as a major threat to citrus production in Tropical Africa, the available information is very scattered and limited.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 19
Therefore, this paper attempts to organize and review the available information with a view it could serve as source of information to researchers, citrus growers, quarantine personnel and policy makers. Furthermore, it could suggest future research areas and collaboration on citrus pathology.
History and distribution Leaf and fruit spot of citrus was first reported in Angola and Mozambique by Decavalho and Mendes in 1952. Within a short period of time the disease has rapidly spread to northwards south of Sahara (Meyonga 1971, Brun 1972, Emechebe 1981, Seif and Whittle 1984) and also eastern part of Africa to Uganda (Kirk 1986), Kenya (Seif and Whittle 1984), Comoros (Aubert 1986) and Ethiopia (Eshetu 1995). This disease has also been reported in Yemen (Kirk 1986). In Ethiopia, the symptom of leaf and fruit spot of citrus was first observed in 1988 to the southern part of the country near to the Kenya border (Yimenu 1993). Latter the disease has been widespread to the south and southwest part of the country, and the causal fungus has been isolated and confirmed to be Phaeoramularia angolensis (IMI, Eshetu 1995). All citrus growing areas of south and south west of Ethiopia (Jimma, Illubabor, Sidamo and Gurage zone) are severely affected by this disease (Eshetu 1995, Yimenu 1993). Latter the occurrence of P. angolensis was confirmed in the north west part of Ethiopia, where citrus plantation is being promoted in collaboration with Governmental and non Governmental organizations (Mohammed 2001). Table 1. Chronology of occurrences of P. angolensis in different countries.
____________________________________________________________ Country Year References _______________________________________________________________________________ Angola 1952 Decavalho and Mendes 1952 Mozambique 1952 Decavalho and Mendes 1952 Zaire 1966 Brun (1972) Central African Republic 1968 Brun (1972) Cameroon 1969 Menyonga (1971) Gabon 1971 Oberti (1971) Congo 1971 Brun (1972) Cotedivoire 1972 Brun (1972) Togo 1972 Brun (1972) Zambia 1973 IMI, Kirk (1986) Nigeria 1978 IMI, Emechebe (1981) Burundi 1980 IAPS (1985) Zimbabwe 1982 Maramba (1982 Uganda 1983 IMI, Kirk (1986) Kenya 1984 Seif and whittle (1984c) Comoros 1985 Aubert (1986) Yemen 1986 Kirk (1986) Tanzania 1990 NARL Ethiopia 1990 IMI, Eshetu (1995) Republic of Guinea - Kuate (1998) Rwanda - Kuate (1998) Source: Seif and Hillocks 1993, Eshetu 1995, Kuate 1998

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 20
Symptoms and nature of damage Phaeoramularia leaf and fruit spot can affect all citrus species including orange, grapefruit, lemon, lime and mandarin. This disease seriously affects citrus leaves and fruits causing several and various sizes of spots and lesions. On leaves the fungus produces circular spots, which often coalescing up to 10 mm in diameter producing light brown or grayish center during the dry season, and become gray to black when the fungus sporulate during the onset of rain. In most cases the lesions are surrounded by a prominent yellow halo. After some time the center of the lesion teared out, creating a shot-hole effect. Heavy attack on leaves may also spread to young branches and cause general necrosis (Kuate 1998). In Ethiopia a complete crop failure has been observed on sweet orange varieties, especially on citrus orchards owned by small-scale farmers. On young fruits symptoms often create a type of hyperplasia producing tumor like lesions surrounded by yellow halo. A mass of gray fungal sporulation can be observed at the early stage of young fruits. Lesion on mature fruits produces sunken brown center. Diseased fruits are hard, juiceless, unmarketable and even not good for local consumption. The loss of marketable fruit yield due to P. angolensis ranges from 50-100% (Seif 1995, Kuate 1998).
Causal organism The causal fungus of this disease was first identified as Cercospora angolensis Cavalho and Mendes in 1953. Recently it was noted that conidia are solitary or catenate borne in simple or branched chains. The fungus was then renamed as Phaeoramularia angolensis (Anonyme 1986). Conidiophores macronematous, mononematous and fasciculate or forming loose synnemata 12-45µm wide. Conidia acrogenous, becoming acropleurogenous, solitary or catenate, borne in simple branched chain of 2-4 conidia, cylindrical to narrowly obclavate rounded at the apex. The fungus can grow well and sporulate properly on artificial culture media such as orange fruit peel extract agar, orange leaf extract agar (Emechebe 1981), V-8 juice agar, mycophyl agar (Ndzoumba 1985) and potato dextrose agar (Kuate and Foure 1988, Siboe1988, White side 1989). P. angolensis was found to be slow grower on potato carrot agar, malt agar and carrot juice-PDA incubated at 25°C under continuous light (Seif and Hillocks 1993).
Economic significance Although Phaeoramularia leaf and fruit spot of citrus was first observed on sweet orange, it can also attack various citrus species including grapefruit, mandarin, lemon, lime and sour orange. The most devastating nature of this disease on various citrus species is mainly premature abscission of young fruits and leaves, and development of spots and lesions, which make the fruit unmarketable. A yield loss of 50-100% is very common in countries affected by P. angolensis (Menyonga 1971, Seif and Kungu 1989, Kuate 1998). Small-scale citrus growers in Ethiopia are forced to abandon their orchard, and substitute by other crops mainly due to severe infection by leaf and fruit spot of citrus.
Biology of the fungus Despite its importance, biology of P. angolensis is little known, and its populations have not yet properly characterized. Differences in response to the disease, both among citrus species and within citrus cultivars suggested that certain degree of genetic variation in the fungus is possible (Ragazzi 1997).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 21
The survival mechanism of this fungus under natural conditions is still unknown. Physiological specialization is also not yet understood, and no telemorph has been reported so far. Pure P. angolensis culture can be obtained from young and freshly infected leaves and fruits. After few weeks of infection the whole lesion can be overgrown by other secondary invaders like Colletothricum and Phoma spp., which makes the identification of the pathogen difficult.
Host susceptibility Susceptibility to P. angolensis varies within citrus species and cultivars. Grapefruit (Citrus paradisi) and sweet orange (C. sinensis) are highly susceptible, while Mandarin (C. reticulata), lemon (C. limon) and tahiti lime (C. latifolia) are less susceptible (Kuate 1998, Seif 1996). In our preliminary observation severity of the disease was also vary to different sweet orange cultivars, and Valencia was found to be the most susceptible, where as Washington navel showed relatively better tolerance at Ghibe citrus farm. A complete crop failure has been also observed under farmer conditions especially on unbuded locally grown citrus trees.
Epidemiology Detailed studies on inoculum sources and disease cycle are lacking. However, leaf lesions were found to be the main source of inoculum of P. angolensis, while fruit lesions serve as subsidiary inoculum source. Citrus fruits are considerably resistant to P. angolensis with age and become easily attacked by the disease when exposed to wet condition and high humidity during the active growth stage of the plant. The disease is transmitted by air-borne conidia or infected planting materials (Kuate 1998). Fruits can be attacked throughout its development, where as leaves no longer attacked by the disease after 5-8 weeks of age (Seif and Hillocks 1996).
Disease management In Kenya, Seif and Kungu (1989, Seif 1996) recommend to use sanitation measures such as collection and destruction of affected leaves, fruits and shoots, and cultural practices like planting of windbreaks around the citrus orchard, judicious pruning of affected shoots and discouraging of inter-planting in affected orchards with mature producing trees. Series of fungicides have also been screened in Cameroon, and Perenox, Benlate and Benzimidazoles were recommended against P. angolensis in West Africa (Rey et al. 1989), (Kuate 1998). Furthermore, application of Benlate by alternating with Copper-based fungicides was also recommended in Kenya. In Ethiopia, sanitation measures have been widely used by citrus growers to reduce inoculum buildup. Fungicidal treatment with Chlorathalonil and Kocide 101 was also recommended (Eshetu 1999). Some of the recommended fungicides like Benzimidazoles and Chlorothalonil are expensive and beyond the reach of small-scale farmers in many citrus growing countries of Tropical Africa. Furthermore, chemical application on citrus trees is difficult for most small-scale farmers due to lack of application equipment like motorized sprayer.
Future recommendation Based on the opinion of different authors, leaf and fruit spot of citrus is found to be the most important production threat to citrus production in most tropical African countries. Detailed and well-organized research on the biology, factors of epidemics and sustainable management are important research areas to be focused. The following recommendations are proposed for effective and sustainable management of leaf and fruit spot of citrus in Tropical Africa:
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 22
Training at all levels on citrus pathology Detailed study on the biology and pathogen variability of P. angolensis Strengthening disease management strategies towards integrated disease management Strengthening the research-extension system Establish regional collaboration efforts on research and germplasm exchange Establish strong quarantine regulation Create awareness to policy makers on the economic importance of the disease
Acknowledgement The author is very grateful to Dr. Seif Abdurabi Abdu for his keen assistance in supplying microscopic pictures of P. angolensis and photocopies of journal articles.
Figure 1. Characteristic symptoms of Phaeoramularia leaf and fruit
spot of citrus on sweet orange.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 23
Figure 3. Distribution of Phaeoramularia leaf and fruit spot of citrus in Ethiopia
Figure 4. Distribution of Phaeoramularia leaf and fruit spot of
citrus in Africa and the Middle East
Key to the map 1. Angola 2. Burundi, 3. Cameroun, 4. C. African Republic, 5. Comoros, 6. Congo, 7. Ivory Coast, 8. Ethiopia, 9. Gabon, 10. Kenya, 11. Mozambique, 12. Nigeria, 13. Rwanda, 14. Rep. Of Guinea, 15. Tanzania, 16. Togo, 17. Uganda, 18. Yemen, 19. Zaire, 20. Zambia, 21 Zimbabwe

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 24
References Anonyme 1986. Description of pathogenic fungi and bacteria. Kew surrey: CMI No. 841. Brun, J. 1972. La Cercosporiose des agrumes provoquee par cercospora angolensis. Fruits,
27 (7-8), 539-541. Decavalho, T. and Mendes, O. 1952. Una Cercosporiose em citrinose Mocambique 72, 8pp. Emechebe, A. M. 1981. Brown spot of citrus caused by Phaesariopsis sp. Annals of applied
biology, 97, 257-262. Eshetu D. (1996). Leaf and fruit spot: A new disease of citrus in Ethiopia. In the proceedings of
the seventh annual conference of the crop science Society of Ethiopia, 27-28 April 1995, Addis Abeba, Ethiopia, pp. 215-221.
Eshetu, D. 1999. Occurrence, prevalence and control methods of Phaeoramularia leaf and fruit spot disease of citrus in Ethiopia. Fruits, Vol. 54 (4), 225-232.
Inter-African phytosanitary commission 1985. Cercospora angolensis Cavalho and Mendes, distribution maps of major crop pests and diseases in Africa, No. 13.
Kirk, P. M. 1986. Phaeoramularia angolensis. Mycopathologia, 94, 177-178. Kuate, J. and Foure E. 1988. La Cercosporiose des agrumes (Cercospora
angolensis).Contribution a letude epidemiolologique dans la zone ecologique de Dschang. Fruits, 43 (10), 559-667.
Kuate, J. 1998. Citrus leaf and fruit spot disease caused by Phaeoramularia angolensis Cahiers Agricultures 7: 121-129.
Maramba, P. 1982. New disease threatens citrus. The farmer, October 25, p 27. Menyonga, G. M. 1971. Cercospora fruit and leaf spot disease of citrus in Cameroon.
Response to four fungicidal treatments. OAU/IAPSC. Document 71/28. Mohammed, Y. 2001. Occurrence of Phaeoramularia leaf and fruit spot of citrus around north
west part of Ethiopia, Field trip report. Ndzoumba, B. 1985. Inoculation experimentales de Cercospora angolensis sur jeunes
plantules d’ agrumes. Fruits, 40 (3), 191-195. Ragazzi, A. 1997. Characterization of Phaeoramularia angolensis isolates from citrus spp. in
Angola by vegetative compatibility tests. Journal of plant diseases and protection 104 (1) 29-35.
Rey et al. 1988. Sensibilite varietale a la Cercosporiose et premiers resultats des tests fongicides dans la province du centre, Cameroun. R.A. 88, IRFA, Agrumes, Doc. No. 20.
Seif, A. A. and whittle A.M. 1984. Diseases of citrus in Kenya. FAO plant protection bulletin 32 (4), 122-127.
Seif et al. 1989. Cercosporiose of citrus in Kenya. In proceedings of the Asian citrus rehabilitation conference, 4-14 July 1989, Malang, Indonesia.
Seif, A. A. and Kungu J.N. 1989. The current status of Cercosporiose of citrus in Kenya. In proceedings of the second conference of the Kenya Agricultural Research Institute, 4-7 September 1989. Pan African Hotel, Nairobi, Kenya.
Seif, A. A. 1993. Phaeoramularia fruit and leaf spot of citrus with special reference to Kenya. International Journal of pest management, 39 (1), 44-55.
Seif, A. A. 1995. Phaeoramularia fruit and leaf spot of citrus in Kenya. Information bulletin No. 15, Kenya Agricultural Research Institute.
Seif, A.A. 1996. Integrated Pest Management for citrus: an option for Kenya. In the Proceedings of the 1st Biennial crop protection conference27-28 March, Nairobi, Kenya, p. 31-39
Seif, A.A. and Hillocks R. J. 1996. Epidemiology of fruit and leaf spot of citrus caused by Phaeramularia angolensis in Kenya: an overview. Proc. Int. Citriculture, 338-339.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 25
Seif, A. A. and Hillocks, R. J. 1998. Some factors affecting infection of citrus by Phaeoramularia angolensis. Journal of Phytopathology 146, 385-391.
Siboe, G.M. 1989. Morphological and biochemical inter-relationship amongst some isolates of Cercospora from Kenya, Ph.D. Thesis, University of Reading, England.
Whiteside, J. O. 1989. Cercospora leaf and fruit spot. In compendium of citrus diseases ( J. O. Whiteside, S.M. Garnsey and L.W. Timmer, (Eds.) (St Paul, Minnesota: American Phytopathological Society), p. 13.
Yimenu, J. 1993. Preliminary surveys report on leaf spot disease of citrus in Sidamo. Coffee Development Authority.
RAPHANUS RAPHANISTRUM L AS A DIVERSIONARY HOST OF BREVICORYNE BRASSICAE L AND BAGRADA CRUCIFERARUM KIRK. J.A. Makwali, F.M.E. Wanjala, and B.M. Khaemba
Abstract The study reported here aimed at depicting the role that the annual weed, Raphanus raphanistrum L could possibly play as a diversonary host to deter two major pests of cabbages namely: the aphid Brevicoryne brassicae L and the Bagrada bug, Bagrada cruciferarum kirk in Kenya. The effects of the aphid B. brassicae were made on stands and rates of weed depletion in plots of 3 x 3, 6 x6 and 10 x 10 m assessed by estimating the level of infestation on entire plants, dry matter (DM) reduction and the incidence of it-aphid-on all above ground parts of the weed. The aphid infested upto 39% of entire stands of the weed by removing 15 - 29% of DM during feeding. This spanned over a period of 4 weeks. The aphid preferred the flowers to other parts of the plant. Different observations were made to understand the developmental biology of B. cruciferarum showed that both nymphal and adult stages of the bug fed on the foliage of the weed. This led to drying up and wilting of mature plants. The females of the Bagrada bug were highly fecund and produced on average 136.8 ± 0.21 eggs in 17 days which initiated control effects on plants on hatching after 17 days. The bug produced five nymphal instars which fed continuously for a period of 57 days (≈≈≈≈ 2 months). Additional studies are being undertaken to assess the yield potential that cabbage attains when protected for 2 months via this technique of using the weed as host to the two pests. When presented with cabbage hosts under a variable number of cultural practices.
Introduction Brassica oleracea var. capitata L., Brassica oleracea var botrytis, Brassica oleracea L. sub sp. Alboglabra and Brassica oleracea L. sub sp acephala constitute the main cruciferous (Brassica) vegetables in Kenya. They are commonly referred to as cabbage, cauliflower, kale (sukuma wiki) and collard, respectively (Masinde et al., 1999). The production of these crops is greatly hampered by insect pests. The most important pests of Brassicas in Kenya comprise the diamond back moth (DBM) Plutella xylostella L., the cutworms, armyworms, cabbage webworms, the aphid, Brevicoryne brassicae L. and the Bagrada bug, Bagrada cruciferarum kirk, among others. The DBM, the B. brassicae and B. cruciferarum, (Talekar and Griggs, 1992; this study in part) feed on numerous plants that are considered as weeds besides the cultivated hosts. The host range is however limited to plant species that contain mustard oils and glucosides which act as specific feeding and oviposition stimulants (Talekar and Griggs 1992). Raphanus raphanistrum L. cruciferae; Brassica is an annual weed that attains a height of upto one metre in Kenya (Ivens 1967). Dense monospecific stands of the weed prevail at altitudes of 2000m above sea level and reduce crop yields for which herbicide use is often recommended. Herbicide use is largely palliative and uneconomic as the weed seeds

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 26
profusely and has a high regenerative capacity. In this respect a biological control strategy would seem appropriate to compliment herbicide use. Such a strategy if feasible would qualify as an integrated control since the weed by attracting B. brassicae and B. cruciferarum would be a diversionary host for the two cabbage pests which in turn by their feeding on the plant would be complimentary to the control of the weed plant. R. raphanistrum, although alien to Kenya, is susceptible to the aphid, B. brassicae and the bug, B. crudiferarum. Both the aphid and the bug use the weed as an alternative host with which they associate throughout the year. The objective of this study therefore was to determine the field efficiency of B. brassicae and B. cruciferarum in controlling R. raphanistrum in the Kenya highlands. This was done by determining the prevalence and effects of the pest stages on the weed and by also assessing their ability to suppress the weed to which they had been diverted.
Literature Review Patch (1938) listed the hosts on which aphids could be reared in large numbers. Johnson (1958) identified the causes of aphid aggregation on plants. Hughes (1962) described the life cycle of B. brassicae in Australia. The species B. brassicae and B. cruciferarum, have been recorded from East Africa, Afghanistan, Pakistan, Sri Lanka, India and South-East Asia (Hill, 1983). Both pest species suck the sap and are associated with drying up and wilting of the attacked plants. Elsewhere, a number of biological agents have been studied to determine their efficacy against weeds. Blossey and Schroeder (1991) studied and screened the potential biological control agents of purple loose strife (Lythrum salicaria L) in Europe. This yielded the most promising agents, some of which were put to commercial application. The control of B. cruciferarum has hitherto emphasised clean weeding as the only other alternative to insecticide application. In this study, incidence of the two pests on the weed was deemed desirable as it represented the ability of the aphid and the bug to defoliate the noxious weed and it also represented an indirect way of establishing the diversionary ability of the weed that could realign damage off the cabbage crop. This is what was quantified.
Materials and Methods The study was conducted at Moi University, Chepkoilel campus. This is in Uasin Gishu district of Rift Valley province approximately 9Km North-East of Eldoret (380 150E latitude, 00 150N longitude, altitude 2000 - 2500m). Effects of B. brassicae This part of the study was done in plots of 3 x 3, 6 x 6 and 10 x 10m with dense stands of the weed. Infestation by the cabbage aphid resulted from nearby cabbage plots. Prevalence of the aphids on the weed was determined from cut lengths of 1, 2, 3, and 4 cm and then by washing them off with ethanol and counting the numbers in their entirely using tally counters. The affinity for the plant was then expressed as number of aphids per cm length of the weed. Vernier calipers were used to determine the sizes of aphids and categorise them as either nymphs (1.0 - 1.5mm) and adults (1-8 - 2.0 mm). They were further categorised as apterate or alate forms. The effects of the aphid on the weed were assessed from the cleaned cuttings taken approximately from mid canopies of attacked and unattacked weeds. The cuttings were labelled and oven dried overnight until all moisture had been removed. Their dry matter was estimated by weighing on a sartory balance accurate to 1mg and expressed as the weight of the attacked as a percentage of the unattacked cutting.
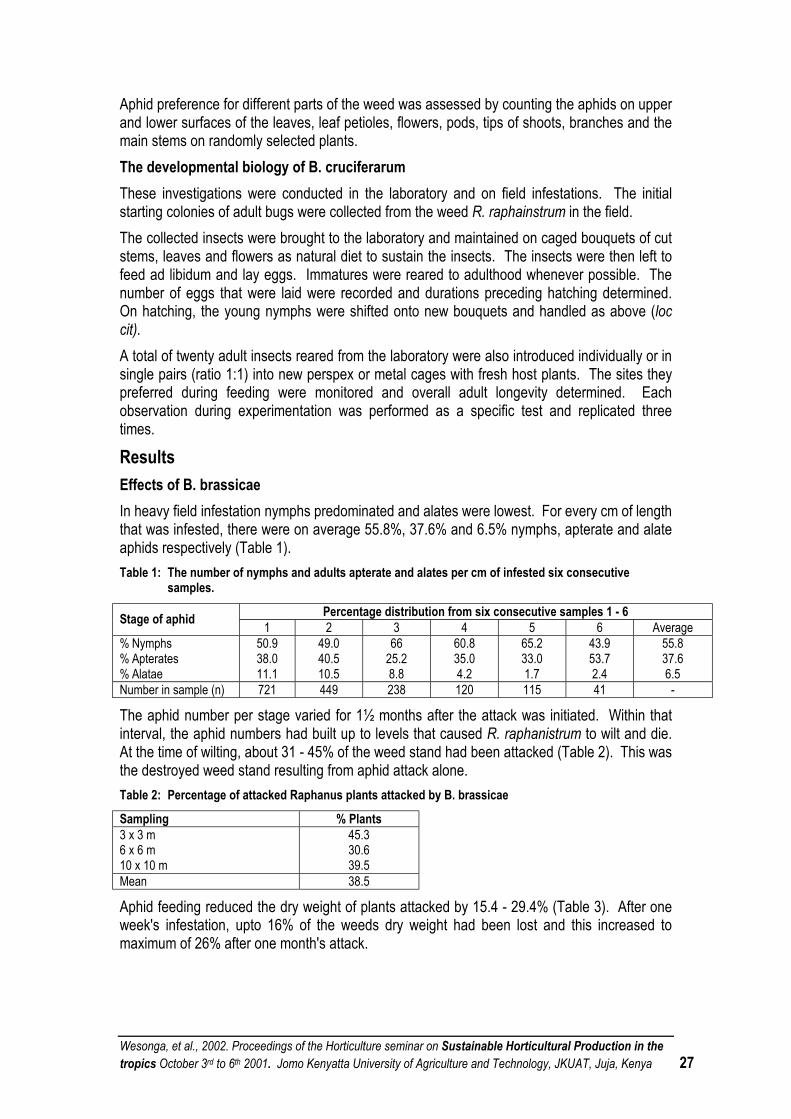
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 27
Aphid preference for different parts of the weed was assessed by counting the aphids on upper and lower surfaces of the leaves, leaf petioles, flowers, pods, tips of shoots, branches and the main stems on randomly selected plants. The developmental biology of B. cruciferarum These investigations were conducted in the laboratory and on field infestations. The initial starting colonies of adult bugs were collected from the weed R. raphainstrum in the field. The collected insects were brought to the laboratory and maintained on caged bouquets of cut stems, leaves and flowers as natural diet to sustain the insects. The insects were then left to feed ad libidum and lay eggs. Immatures were reared to adulthood whenever possible. The number of eggs that were laid were recorded and durations preceding hatching determined. On hatching, the young nymphs were shifted onto new bouquets and handled as above (loc cit). A total of twenty adult insects reared from the laboratory were also introduced individually or in single pairs (ratio 1:1) into new perspex or metal cages with fresh host plants. The sites they preferred during feeding were monitored and overall adult longevity determined. Each observation during experimentation was performed as a specific test and replicated three times.
Results Effects of B. brassicae In heavy field infestation nymphs predominated and alates were lowest. For every cm of length that was infested, there were on average 55.8%, 37.6% and 6.5% nymphs, apterate and alate aphids respectively (Table 1). Table 1: The number of nymphs and adults apterate and alates per cm of infested six consecutive
samples.
Percentage distribution from six consecutive samples 1 - 6 Stage of aphid 1 2 3 4 5 6 Average % Nymphs % Apterates % Alatae
50.9 38.0 11.1
49.0 40.5 10.5
66 25.2 8.8
60.8 35.0 4.2
65.2 33.0 1.7
43.9 53.7 2.4
55.8 37.6 6.5
Number in sample (n) 721 449 238 120 115 41 -
The aphid number per stage varied for 1½ months after the attack was initiated. Within that interval, the aphid numbers had built up to levels that caused R. raphanistrum to wilt and die. At the time of wilting, about 31 - 45% of the weed stand had been attacked (Table 2). This was the destroyed weed stand resulting from aphid attack alone. Table 2: Percentage of attacked Raphanus plants attacked by B. brassicae
Sampling % Plants 3 x 3 m 6 x 6 m 10 x 10 m
45.3 30.6 39.5
Mean 38.5
Aphid feeding reduced the dry weight of plants attacked by 15.4 - 29.4% (Table 3). After one week's infestation, upto 16% of the weeds dry weight had been lost and this increased to maximum of 26% after one month's attack.
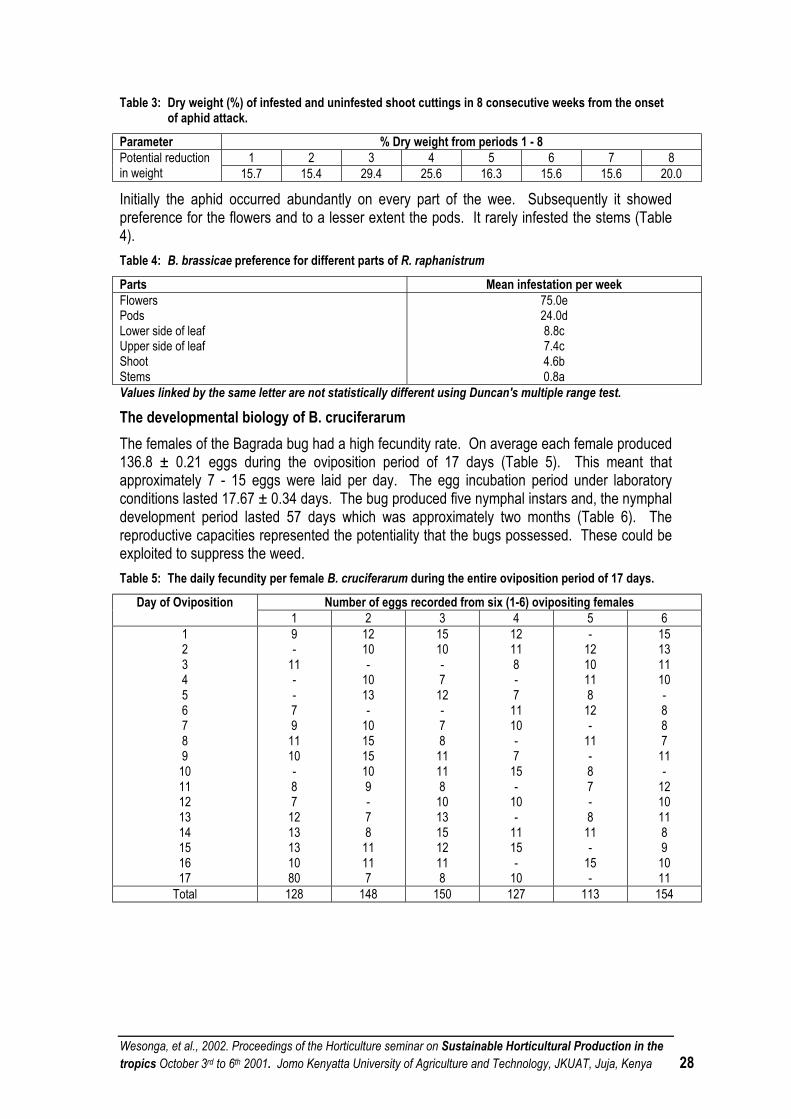
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 28
Table 3: Dry weight (%) of infested and uninfested shoot cuttings in 8 consecutive weeks from the onset of aphid attack.
Parameter % Dry weight from periods 1 - 8 1 2 3 4 5 6 7 8 Potential reduction
in weight 15.7 15.4 29.4 25.6 16.3 15.6 15.6 20.0
Initially the aphid occurred abundantly on every part of the wee. Subsequently it showed preference for the flowers and to a lesser extent the pods. It rarely infested the stems (Table 4). Table 4: B. brassicae preference for different parts of R. raphanistrum
Parts Mean infestation per week Flowers Pods Lower side of leaf Upper side of leaf Shoot Stems
75.0e 24.0d 8.8c 7.4c 4.6b 0.8a
Values linked by the same letter are not statistically different using Duncan's multiple range test.
The developmental biology of B. cruciferarum The females of the Bagrada bug had a high fecundity rate. On average each female produced 136.8 ± 0.21 eggs during the oviposition period of 17 days (Table 5). This meant that approximately 7 - 15 eggs were laid per day. The egg incubation period under laboratory conditions lasted 17.67 ± 0.34 days. The bug produced five nymphal instars and, the nymphal development period lasted 57 days which was approximately two months (Table 6). The reproductive capacities represented the potentiality that the bugs possessed. These could be exploited to suppress the weed. Table 5: The daily fecundity per female B. cruciferarum during the entire oviposition period of 17 days.
Number of eggs recorded from six (1-6) ovipositing females Day of Oviposition 1 2 3 4 5 6
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17
9 -
11 - - 7 9
11 10 - 8 7
12 13 13 10 80
12 10 -
10 13 -
10 15 15 10 9 - 7 8
11 11 7
15 10 - 7
12 - 7 8
11 11 8
10 13 15 12 11 8
12 11 8 - 7
11 10 - 7
15 -
10 -
11 15 -
10
- 12 10 11 8
12 -
11 - 8 7 - 8
11 -
15 -
15 13 11 10 - 8 8 7
11 -
12 10 11 8 9
10 11
Total 128 148 150 127 113 154

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 29
Table 6: Durations for immature stages of B. cruciferarum on R. Raphanistrum
Stage Duration (days ± S.E) Egg incubation First nymphal instar Second nymphal instar Third nymphal instar Fourth nymphal instar Fifth nymphal instar
17.67 ± 08.34 6.67 ± 0.34 8.17 ± 0.34 9.00 ± 0.34 7.83 ± 0.34 8.58 ± 0.34
Total nymphal development period 57.0 days
It was observed that both nymphs and the adults fed by sucking juices from the foliage and floral parts of the plant. When the growing points of the young plants were attacked, they reacted by withering and wilting before they completely dried up. Damaged leaves that survived were comparatively smaller in size and malshaped as compared to the normal ones. The attack of the pest on flower buds and young flowers led to their premature abscission. Similarly its attack on very young pods led to their complete arbotion. Thus the pest curtailed the overall reproductive capacity of the R. Raphanistrum plants.
Discussion The mode and rate of reproduction are outstanding features that make both B. brassicae and B. cruciferarum potential useful indigenous biological control agents for R. raphanistrum in Kenya. R. raphanistrum is a definite diversionary host to the aphid and bug rather than a mere alternative host. The weed attracted and supported upto 228 nymphs of the aphid per cm2 of the infested weed surface. When a weed species is susceptible to an insect species, it ultimately wilts and fails to seed in the presence of an effective insect agent (Miller et al., 1988). Preference for flowers, which account for seed production, implied that B. brassicae was an effective agent. Many aphid species are attracted to their hosts during periods of luxuriant vegetative growth (Sticher, 1971). R. rhaphanistrum had a rapid growth with succulent parts over a period of 1½ months. This led to an appropriate time when the species was most susceptible. Many aphid species are able to synchronise the chemical content of the hosts and the plant parts they feed upon (Crawley, 1983). The aphid was capable of controlling up to 38.5% of an entire stand of weed. This was substantial and represented the efficacy that B. brassicae attained during its parasition of the weed alone. Why the aphid could not feed on the weed in its entirety is not yet known. However, the overall efficiency for field based weed control with biological agents are always variable because of interactions between the insect agents and the plant hosts they prefer (Crawley, 1983). Variability is highest in situations where the agents involved are indigenous (Schroeder, 1985). It was evident that both adults and nymphs of B. cruciferarum fed on the foliage of R. raphanistrum, whose leaves briskly wilted and thereafter dried up. Young plants often died completely. The two stages sucked the vital sap and devitalised the pods and thereby adversely affected the seed formation. This is one way by which typical herbivores deplete the vigour of their hosts and thus render control (Hill, 1983). What characterised this study was the fact that the two pests did not co-exist on the same host plants at ago. This did not imply any specificity of agents to weeds since, this was not expected. Most successful weed control are invariably host specific (Jeanneret and Schroeder, 1992). The pest-weed association would seem to be intense where the insect is being imported to control the weed species. This was not the case in this study. However, there is not evidence

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 30
to suggest why the intensity should not apply to a situation where an exotic weed is confronted by local biological control agents, as was the case with B. brassicae and B. cruciferarum when analysed and evaluated separately for their efficacy on R. raphanistrum. The two insects can possibly compliment each other very effectively.
Acknowledgement We are grateful to the technical staff of the department of Zoology for the enthusiasm they showed in this work during its execution.
References Blossey, B and Schroeder, D. (1991). Study and screening of potential biological control of
purple loose strive (Lythrum salciera L). Final report (11BC) Delemont - 41pp. Crawley, M (1983). Herbivory. The dynamics of animal plant interactions. Blackwell
publications Oxford 430 pp. Hill, D.S. (1983). Agricultural insect pests of the tropics and their control 634pp. Hughes, R.D. (1962). Population dynamics of the cabbage aphid, Brevicoryne brassicae. J.
anim. Ecol. 32:393-423. Ivens, G.W. (1967). East African weeds and their control. 147pp. Jeanneret, P. and Schroeder, D. (1992). Biology and host specificity of Mecinus janthinus
Germar. Biocontrol science and Technology 2, 25 - 34. Johnson, B.L. (1958). Factors affecting locomotion and settling responses of alate aphids.
Anim. Behaviours, 6:9-26. Masinde, P.W., Masuda, Agong, S.G. and Sugiura, H. (1990). Premature flowering of collard
(Brassica oleracea var. Oleracea) as influenced by the genotype and season in Kenya. In Agong et al (eds) proceedings of 2nd National Horticulture seminar on the way for Horticulture Towards Kenya's Industrialization 59 - 70.
Patch, E.M. (1938). Food-plant catalogue of the aphids of the world including the phylloxeridae. Maine Agric. Exp. Sta Bull 393: 431pp.
Schroeder, D. (1985). The search for effective biological control agents in Europe. I. Diffuse and spotted knapweed. Proc. Vi Int. Symp. Biol. Contr. Weeds 19 - 25 August 1984. Vancouver, Canada, Delfose Es. (ed.) Agric. Can. Pp. 103-19.
Sticher, O. (1971). Isolation of antirrhinoside from Linaria vulgaris. Phyto chemistry 10, 1974 - 1975.
Talekar, N.S. and Griggs, T.D. (1992). Management of diamondback moth and other crucifer pests: Proc. 2nd Int. workshop Shauhia, Taiwan. Asian vegetable Research and Development Centre, 603pp
TOWARDS SUSTAINABLE CROP PROTECTION THROUGH UTILISING FAVOURABLE CULTURAL PRACTICES: POTENTIAL IN VEGETABLE CROPS Sithanantham, S.; Agong, S.G.; Matoka, C.; Raini, R. and Makatiani, J.
Abstract This paper focuses on cultural practices as a promising research area towards sustainable crop protection in tropical vegetable crops. The potential for reducing the infestation by diamondback moth (Plutella xylostella) and aphids on cabbage crop by companion cropping with more susceptible host plants as trap crops has been demonstrated in India (Karnataka), USA (Florida) and South Africa. Promising trap crops identified till recently include mustard and collards. Recent evaluations initiated at ICIPE included such promising trap crops (as “pull crops”) and another group of plants that are likely to be repulsive to the onset of pest infestations (and so could possibly act as “push crops”). In a field trial conducted during 2000, cabbages planted along with non-favourable host plants like Cleome and

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 31
coriander (in a ratio of 2:1) showed distinct reduction in infestation by P. xylostella comparable to the levels achieved under B.t. sprays in the sole cabbage crop. Laboratory choice tests also showed that these two plants are not preferred for oviposition by moths of P.xylostella. Follow-up studies are being undertaken to verify the relative potential of beneficial promising companion (push-pull) crops, as adapted to the two major crops production ecologies –highlands (cabbage) and lowlands (kale). Attention is also being given to the potential effect of these companion crops on other pests and diseases. The scope for optimising the row ratios and for adjusting the planting geometry so as to maximise the benefits of companion cropping are being explored. Past research has shown that selected non-host plants like African Marigold (Tagetes) when planted in rotation in intensive vegetable cropping systems can help suppress the nematode populations in the soil. This option is indeed very relevant to our current preference to avoid or minimise the use of chemical nematicides in these cropping systems. On-station/on-farm demonstration of the direct benefits of promising rotation crops in suppressing the soil nematode populations, should be influenced to favourably influence the attitude of vegetable farmers towards adopting them. The two promising cultural practices discussed – crop rotation and companion cropping – could offer a window of opportunity to the vegetable growers in the tropics whose main constraints have been arthropod pests and soil nematodes. It is recommended that suitable applied and adaptive research on these aspects be supported so as to contribute to sustainable reduction in pesticide use and so pave way for sustainable crop protection in the target vegetable production systems.
POTENTIAL FOR ENHANCING THE IMPACT OF AUGMENTATION BIOCONTROL OF TWO KEY CATERPILLAR PESTS ON IMPORTANT VEGETABLE CROPS IN KENYA Sithanantham, S.; Maniania, N.K.; Osir, E.O. and Baya, J.
Abstract Helicoverpa armigera (Hbn.) and Plutella xylostella are two major lepidopteran pests of vegetable crops in Kenya. Vegetable farmers are in dire need of safer alternatives to the use of chemical pesticides as means of sustainable crop protection. On-going research at ICIPE on utilising augmentation biocontrol agents against these pests is illustrated with focus on enhancing their potential impact. For biocontrol of H armigera, several native strains of the baculovirus (NPV) have been assembled and bioassayed. Results indicate the potential of this virus to cause substantial mortality. The need for dosage adjustments to impact upon different larval instars have been also ascertained. Large-scale field evaluations for efficacy and product registration are being planned. A regional network project has been recently launched for better understanding of the diversity among native trichogrammatid egg parasitoids of H. armigera as a basis for utilising promising taxa in augmentation biocontrol of this pest. It is proposed to establish a pilot mass production unit for trichogrammatids as means of creating awareness about the potential local availability and utility of these agents. For biocontrol of P xylostella a new product. "Green guard" a Bacillus thuringiensis var kurstaki (Bt.k) - based biopesticide from China has been evaluated in the field. Its efficacy is comparable to other Bt.k products available in the Kenyan market (e.g Dipel and Thuricide). Different ecological populations of this target pest have been found to vary in their susceptibility to B.t. products. Further research to ascertain and elucidate the interactions in diversity between the pest and the pathogen is being pursued. Surveys have also been undertaken in Kenya resulting in isolation of several naturally occurring strains of Bt.. Metarhizium anisopliae and granulosis virus. Evaluation of these three groups of pathogens against DBM is in progress. For enhancing the potential impact of the promising biocontrol agents, it is important to strengthen and widen the existing research collaboration with the national partner institutions as well as with advanced research centers, besides bringing about greater awareness among consumers as well as motivating private entrepreneurs to undertake commercial production. ICIPE is keen to pursue these initiatives with interested partners.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 32
HOST PLANT RESISTANCE IN OKRA (ABELMOSCUS ESCULENTUS (L) MOENCH:MALV ACEAE) VARIETIES. E. W. Wanjal2, R.O. Marangal, R. Hallett2 1 Jomo Kenyatta University, Department of Zoology P .0 Box 62000, Nairobi, Kenya Email: [email protected] 2 University of Guelph, Canada. Corresponding author: R. O Maranga
Abstract Studies were initiated to investigate the potential of utilizing pest resistant varieties of okra in small-scale farms to reduce the dependency on pesticides and also preserve the quality of our environment. The specific objectives of the study were; to evaluate the response of okra to infestations by insects in the field and to determine seasonal insect population changes in okra varieties; to determine the presence and / or absence of varietal resistance and / or susceptibility in okra varieties, some of which are sold by seed companies and grown locally; to establish the chemical basis of resistance, if any and to establish behavioural and physiological responses of the insects to phytochemicals in laboratory bioassays. Eight okra varieties (Abhay Clemson spineless, Pharbhani, Anamika, Dwarf green, Louisiana velvet, Pusa sawani and Kangwani) were planted in a randomized block design in a plot measuring 25mx 19m. This plot was then subdivided into six replicates measuring 8mx4m and each variety was planted on a plot measuring 2mx2m with a 1m boundary between plots. Three of the replicates were given a blanket insecticide treatment three weeks after germination while the other three replicates were left untreated. All the replicates were given fungicide sprays four weeks after germination. Experiments were first conducted at JKUAT and Sagana for two seasons (i.e long and short rains). In a farmer participatory approach, farmers from different okra growing areas were supplied with the various okra varieties which they planted in a split plot design with treatments as the main plots and varieties as the sub plots. Half of the plots were given insecticide treatment three weeks after germination while the other half remained untreated. All varieties were given fungicide spray four weeks after germination. Normal agronomic practices were observed in all the plots. Farmers were instructed on how to recognize pests, pest damage, evaluation of quantity, quality and marketability of the different varieties. Extraction and identification of the chemical components of the most resistant and two most susceptible varieties are to be done and behavioural responses of insects to the extracted chemicals established in laboratory bioassays. Identified chemical compounds will be used in field trials as pest attractants or repellants. It is hoped that these compounds will be packaged commercially and made available to okra farmers to be used in pest management programmes. Some okra varieties have been identified to have some degree of resistance and / or susceptibility to the cotton aphid and other pests. Varieties 2 and 7 were susceptible while 4 and 8 showed some degree of resistance. Crude extracts of volatile and non-volatile okra leaf compounds have given an indication that both may be involved in host finding and recognition by the cotton aphid. In orientation responses, aphids were attracted to extracts of susceptible varieties more than to the control and resistant varieties. Elucidation of the compounds extracted are in progress at the University of Guelph, Canada. The participatory research is in progress and since its inception in March, 2001, the farmers have shown great interest in the work and most of those who have been involved have been very co-operative, participating effectively in the research. They have learnt to identify the major pests of okra and have been able to identify and appreciate the resistant varieties on their own. The farmers have been able to market the produce from their farms which has given them some earning and they appreciate that if they increase the scale, they can depend on okra for income. Key words: Host plant resistance, Bioassays, Okra varieties, Phytochemicals
Introduction Export vegetables are emerging as important source of income for many small scale farmers in Africa (ICIPE, 1997). In East Africa, okra (Abelmoscus esculentus) is among the most important export vegetables. In Kenya an estimated 300,00 families earn their income through cultivation and marketing of vegetable crops (Sithanatham et al., 1998). There has been notable increase in the production off and export of Asian vegetables in Kenya since 1996, okra being one of them. Between 1995 and 1996, a total of 6,183 tonnes of okra valued at USS 83,453 were produced at Kibwezi region of Eastern Province, which is relatively a semi-arid

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 33
area. This production is likely to increase as farmers realize the okra potential in foreign exchange earning. A major constraint in vegetable production is the effective prevention and control of pests and diseases, which cause high, yield losses (Tindall, 1983). Vegetable losses due to pests can be as high as as 25%, which can result in severe income loss to small-scale farmers. Avoidable loss in okra due to pests has been placed at about 54% (Chaudhary and Dadheeck, 1989). Okra is attacked by large number of insect pests, which reduce quality and yield. Ewete, (1978), reported that one of the factors limiting yield of okra fruits in the tropics is the range of insect pests associated with the crop. These pest include sapsuckers, leaf eaters, leaf and fruit borers, leaf rollers, flower feeders and leaf miners. Among the sapsuckers are the nymphs and adults of the cotton aphid, Aphis gossypii that cause leaf distortion and cupping. In severe infestations they may debilitate the plant and may also be serious vectors of viral diseases (Tindall, 1983). White flies, Bemisia tabaci transmit the yellow vein mosaic virus, which is the most serious disease of okra in India (Narayan et al, 1986) while green leafhoppers Empoasca spp. inhibit food translocation through their feeding (Hill, 1983). Others include cotton stainers Dystercus spp. and the cottonseed bug Oxycarenus hyalinipennis. The leaf eaters include beetles such as Lagria villosa and the flea beetle Nisotra spp. that feed on outer layers of leaf tissues damaging cotyledons. Spiny bollworms Earias spp.bore terminal shoots of young plants causing death of the tip, which results in the subsequent development of side shoot (Tindall, 1983). Young capsules and flower buds wither and fall after being bored into (Hill and Walter, 1994). Helicoverpa (Heliothis) armigera bores clean circular holes in fruits and flower buds. Leaf rollers include the cotton leaf roller Sylepta derogata larvae that feed inside the rolled leaf. Environmental friendly methods for pest management are being sought and Host Plant resistance (HPR) has been identified as one of the most economical and environmentally safe method for pest control. Painter (1951) defined HPR as the relative amount of heritable qualities possessed by the plant, which influence the ultimate degree of damage done by pests in the field. Russell (1978), defined resistance as any heritable characteristics of a host plant, which lessen the effect of parasitism. Expression of resistance is determined by the inherent ability of a plant and involves only plant characteristics and is not part of an insect-plant interaction. Plants contain a large number of substances, which have their primary use as a means of defense against natural enemies (Fraenkel, 1959). With the European Union setting limits of maximum residue levels especially for horticultural products, emphasis must be placed on non-chemical means of control of pests in okra and HPR is one such methods. Farmers must be able to identify pests and control them before they reach economic injury levels. This way other means of pest control other than chemicals may be used prior to harvest to reduce residue. Host Plant Resistance is seen to be a sustainable approach to pest management and varietal trials of different okra plants to insect pests are essential. This is an attempt to identify the response by okra to pests in order to determine resistance and/or susceptibility. Convenient, safe and inexpensive plants identified would be recommended to growers to avoid losses due to pests. Genes responsible for resistance may be identified and incorporated into high yielding susceptible cultivars. Identification of physical and morphological characteristics of resistant varieties and evaluation of their phytochemicals may lead to the introduction of resistance traits to preferred varieties. Also phytochemicals from susceptible varieties may be used for mass trapping of insects in the field. Hence identification of resistant varieties of high yield will provide a convenient, economical and environmetally safe method of control to the farmer.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 34
General objective To reduce dependency on pesticides hence reducing costs and preserving the quality of our environment through the use of resistant varieties.
Specific objectives 1. To evaluate the response of okra to infestations by insects in the field and to determine
seasonal insect population changes in okra varieties 2. To determine the presence and/ or absence of varietal resistance and/or susceptibility in
okra varieties, some of which are sold by seed companies and grown locally. 3. To evaluate the physical and morphological characteristics of varieties that may
demonstrate resistance and/or susceptibility 4. To establish the chemical basis of resistance, if any 5. To establish behavioural and physiological responses of the insects to phytochemicals in
laboratory bioassays
Expected outputs 1. Levels of insect infestations and at what time of the season to expect them will be
determined 2. Varieties with resistant and/or susceptibility to pests will be established 3. Physical characters that contribute to resistance or susceptibility in okra will be elucidated 4. Plant chemicals that may protect plants from pest infestations or attract pests 5. will be determined 6. Responses of plants to extracted plant chemicals as attractants or repellants will be
determined
Method Eight okra varieties (Abhay Clemson spineless, Pharbhani, Anamika, Dwarf green, Louisiana velvet, Pusa sawani and Kangwani) were planted in a randomized block design in a plot measuring 25mx19m. This plot was then subdivided into six replicates measuring 8mx4m and each variety was planted on a plot measuring 2mx2m with a 1 m boundary between plots. Three of the replicates were given a blanket insecticide treatment three weeks after germination while the other three replicates were left untreated. All the replicates were given fungicide sprays four weeks after germination. Experiments were first conducted at JKUA T and Sagana for two seasons (i.e long and short rains). In a farmer participatory approach, farmers from different okra growing areas were supplied with the various okra varieties, which they planted in a split plot design with treatments as the main plots, and varieties as the sub plots. Half of the plots were given insecticide treatment three weeks after germination while the other half remained untreated. All varieties were given fungicide spray four weeks after germination. Normal agronomic practices were observed in all the plots. Farmers were trained on how to recognize pests, pest damage, evaluation of quantity, quality and marketability of the different varieties. Extraction and identification of the chemical components of the most resistant and two most susceptible varieties are to be done and behavioural responses of insects to the extracted chemicals established in laboratory bioassays. Identified chemical compounds will be used in field trials as pest attractants or repellants. It is hoped that these compounds will be packaged commercially and made available to okra farmers to be used in pest management programmes.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 35
Preliminary results Preliminary field experiments to establish the mechanisms of resistance to cotton aphid, Aphis gossypii, which had been earlier, identified as an economic pest were carried out in green house. Some varieties of okra were identified to have some degree of resistance and / or susceptibility to the cotton aphid and other pests. Varieties 2(Spineless clemson) and 7(Pusa sawani) were susceptible while 4(Anamika) and 8(Kangwani) showed some degree of resistance. Experiments on physical and morphological characteristics were carried out and feeding and orientation bioassays with okra leaves with trichome removed and those with trichomes did not give any correlation between aphid numbers and the number of trichomes. This may be an indication that trichomes may not play an important role in aphid attack in okra plants.
Chemical basis for resistance Crude extracts of volatile and non-volatile okra leaf compounds have given an indication that both may be involved in host finding and recognition by the cotton aphid. In orientation responses, aphids were attracted to extracts of susceptible varieties more than to the control and resistant varieties. Elucidation of the compounds extracted are in progress.
Impact Short term effect in adoption: Participatory research with farmers is in progress in Matuu and Maragwa. Matuu is a very promising area for okra farming since the soil and prevailing weather conditions are suitable with plenty of water for irrigation. Horticultural farming is a major income earner for farmers in this area and they have a good understanding of the conventional agronomical practices. Since this participatory research was initiated in March, 2001, the farmers have shown great interest in the work and most of those who have been involved have been very co-operative, participating effectively in the research. They have learnt to identify the major pests of okra and have been able to identify and appreciate the resistant varieties on their own. The farmers have been able to market the produce from their farms, which has given them some earning, and they appreciate that if they increase the scale, they can depend on okra for income. It is hoped that the extracts from the resistant varieties of okra will be used on the high yielding susceptible varieties as pest repellants so that okra can be grown without the use of chemical pesticides. Studies to this effect are going on and are expected to be concluded soon after which the products will be subjected to trials in farmers' fields. Farmers are eagerly waiting for this product. Long term effects: In the long run, it is expected that farmers will save money currently used on pesticides and
the environment will be saver for all of us. Higher yields are expected with less damage from pests as a result of the use of the
products from the resistant varieties as well as the growing of the identified resistant varieties.
Income for the farmers will improve and so will their general social welfare.
Extension programme: The growing and use of okra will be encouraged in an extension programme that has already been established in these areas through the participating farmers and extensions officers. An outreach programme through the local administration whose groundwork has been done and seems to be very promising is planned. These programmes are designed to create awareness

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 36
among farmers especially that okra is a high income earner both locally and as an export crop and can alleviate poverty. That the use of the resistant varieties will reduce dependency on chemical pesticides, which will save the much-needed money currently, used for the purchase of chemical pesticides. That the use of the resistant varieties and their products will eliminate the consumption of chemical pesticides as residues in produce and hence improve their health and the marketing of their produce in international markets. It is hoped that the use of these resistant varieties of okra and their products for pest management will be spread to other parts of the country to be grown by resource poor farmers. Targeted areas include Kibwezi, an area with poor resource farmers but has a high potential for okra farming. The extension programme that has been started forms a very important component of this work as it forms the avenue of technology transfer and adoption.
References Chaudhary,H.R. and Deedhack, L.N. (1989) Incidence of insects attacking okra and the
avoidable losses by them. Annals of arid zone 28: 305-307. Frankel,G.S. (1959) the raison d'etre of secondary plant substances. science 129:1446-4170 Hill, D. and Waller, J (1994) Pests and diseases of tropical crops, pp.10-134. Cambridge
University press, London Hill, D. (1983) Agricultural insect pests of the tropics and their control, pp. 96-134 Cambridge
University press, London Sithanantham, S., Lako, G., Nyarko, K. and Bob, M. (1998) Improved pest management
approaches for Africa; A thematic study. ICIPE/IF AD Publication, Nairobi, Kenya Tindall, H. (1983) Vegetables in the tropics. Oxford University press, London pp 90-145 Painter, R.H. (1951) Insect resistance in plant crops. Mcmillan, New York Russel, G.E. (1978)
Plant breeding for pest and disease resistance. Butterworths, London pp485
Discussion Q: Are you aware of the PRA studies in eastern and central regions? These studies should help in selecting farmers for the purpose of your experiments A: We will involve farmers from various locations in the long run
SOME ASPECTS OF SUSTAINABLE TOMATO PRODUCTION Fricke, A. Institute of Vegetable and Fruit Science, University of Hannover, Herrenhaeuser Str. 2, D- 30419 Hannover, Germany, e-mail: [email protected]
Abstract Tomato is a vegetable with a high demand. The consumers appreciate taste, nutritional value and the broad possibilities of using tomatoes in the nutrition. Especially because of its high needs concerning water and nutrients a sustainable tomato production system is necessary. With tomato a lot of work is done concerning resistances. But still there are problems with virus diseases, which make it necessary to fight against aphids. Also red spider mite and white fly make problems. So here a protected cultivation (foliar cover, nets) can help to reduce the use of insecticides and enables the use of beneficial insects. Also, many fungal and some bacterial diseases can be controlled by resistances. But still there is a lot to do in breeding. The right choice of the variety in the respect of resistances and climatic condition and the possibility of grafting are helping here to be as environmental friendly as possible.To overcome the negative impacts on the soil quality (compression, erosion) mulch materials have to be used. Concerning fertilisation the uptake of nutrients is well known. To supply nutrients according to demand should not be a problem if mineral nutrients are used. If only organic fertilisation is available, knowledge of the mineralisation has to be available to avoid leaching of nutrients.To produce a high quality tomatoes have to be irrigated. There are two factors to take into account: the amount of water and the water quality. Important are not only sufficient but also the constant availability of water. Here the challenge is to

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 37
develop irrigation management systems with a high water use efficiency. This efficiency is also closely related to the irrigation system. Using drip irrigation and mulching materials are preconditions to reach this aim. The development of irrigation models, which will quantify the effects of different water supply, can help to improve the irrigation management.
Discussion Q: How does fresh market tomato production cause weed problem ? A: Because of its continuity and weeds are best controlles by herbicides to avoid soil compression Q: How is tomato grafting done to control disease? A: there are rootstocks, which are for pest and disease resistance and cold temperature tolerance. Q: Based on your presentation, you have developed tomato varieties, which are resistant to Fusarium, Phytophthora, and nematodes. Are they easily available in the market? A: All resistant varieties re commercially readily available, the seed are available from Dutch seed producers easily. They are all F1 and high in price. Q: Could you comment on the role of seed production as an important component of sustainable tomato production especially in the common farmers conditions? A: Seed quality is very important factor for successful crop. This is true on one hand for the percentage germination and on the other for the value of the variety according to climatic adaptation. So, to produce seeds of varieties which fulfil the conditions above would be an important factor for sustainable production. The use of seed materials which quality (esp. resistant) is not known can cause a lot of problems during growth and therefore needs a lot of pesticides
RESPONSE OF TOMATO (LYCOPERSICON ESCULENTUM L.) GENOTYPES TO INDUCED SALT STRESS Agong, S. G. Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P .O. Box 62000 Nairobi, Kenya
Abstract Thirteen tomato genotypes were subjected to salt treatment under hydroponics and their responses monitored in a set of two experiments with the objective of advancing them as potential salt tolerant tomato scion and/or rootstocks. Salt applications ranged from 0 to 2% NaCl with the resultant EC values of 1.4 to 37 dS m-l, respectively. Genotypes were cultured in the experimental solutions for up to four weeks in a three replicate completely randomized design in the greenhouse. Significant genotypic and/or salt treatment effects were registered on plant height, leaf green meter value and area, dry matter yield, Na.+. and Cl- accumulation in tomato tissues. Salt treatment at 2% NaCl stimulated chlorophyll production per unit leaf area but caused severe depression on dry matter yield and leaf area. These results revealed that some tomato genotypes consistently showed superior biological activity at higher salinity and others exhibited greater shift in the shoot: root ratio based on dry matter biomass production thus displaying relatively greater adaptation to salt stress. Two tomato genotypes ('Siozawa' and 'Gambaru Ne-3') displayed superior performance based on these data.
Discussion Q. What specifically is the potential of nutritional breeding in vegetable in improvement? Is there some example of characters of interests which can be obtained through mutation?

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 38
A: Considering crops like spider plant, it has been observed that existing population show little geneotypic variation and this may not support an elaborate breeding programme. There is then need to increase the existing variability by use of artificial means such as mutations induction. Q: What is the role of African indigenous vegetable in the future sustainable vegetable production in Kenya? A: We should promote indigenous vegetables in Kenya just us we do for exotic vegetables. Exotic vegetables are high value but need high level of inputs. Thus the indigenous vegetables would be important in sustainable vegetable production Q: Were you able to establish if the sodium is compartmentalised in bascules as a way of salt tolerance mechanism. A: in this current study I was not able to establish the cellular depositions of sodium within the leaves and the stems, however, in the parallel studies elsewhere it has been confirmed that most sodium is compartmentalised within the vacuoles of the tomato scion Q: there were considerably higher levels of Na+ and Cl- ions accumulation in the leaves, how did this affect the photosynthetic capacity of the leaves? Q.: How was the assimilates partitioning towards the fruit development? Since the fruit is the economic part of the plant? A: photosynthetic capacity was definitely reduced as confirmed by the reduced dry matter accumulation under the salt stress condition. A: These study was limited to seedling (4 weeks of age) otherwise elsewhere Agong et al. (1997, 2000) confirmed that most assimilates are redirected into the fruit formation and development as a survival strategy under salt stress
GENETIC TRANSFORMATION OF TOMATO LINES FOR RESISTANCE OF CUCUMBER MOSAIC VIRUS Kusolwa, P. Department of Crop Science and Production, Sokoine University of Agriculture, P .0. Box 3005, Morogoro, Tanzania.
Abstract This study was conducted with the purpose to develop transgenic tomato plants resistant to cucumber mosaic virus (CMV). Three tomato varieties were used for invitro genetic transformation via A. tumefaciens using a virus coat protein (CP) mediated resistance from two CMV strains (I-17F and R). In vitro regenerated plants resistant to kanamycine were screened by PCR analysis and western blot techniques to reveal the presence and expression of the transgenic coat protein DNA sequence respectively. Early analyses were effected on in vitro regenerated plants and later, on few weaned plants kept under the green house conditions. Few transgenic lines expressing the presence of CP were obtained. Furthermore, these transgenic lines were tested for resistance to CMV by inoculation with the two corresponding virus strains. The infection was very heterogenous and unsuccessful for the majority of the plants, this did not allow for judgement of the actual potential resistance of transgenic lines. Tests by DAS-ELISA on the extracts of both inoculated leaves and systemic leaves (subsequent leaves) showed high optical densities for the transgenic plants proving the presence of endogenous transgenic viral coat protein. The few infected non transgenic lines showed high optical densities in the same way as transgenic plants but with visual symptoms of CMV. Plants of Rl generation were produced by self pollinating each individual transgenic lines. Further resistance tests need to be conducted on these lines.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 39
EFFECTS OF DURATION OF SOAKING AND CONCENTRATION LEVEL OF SALICYLIC ACID ON SEED GERMINATION OF CABBAGE (Brassica oleracea var. capitata L.), CUCUMBER (Cucumis sativus L.) AND TOMATO (Lycopersicon esculentum Mill.) K’Opondo, F.B.O.1, E.O. Auma1*, P.W. Mathenge1 and S.P.C. Groot2 1Deparment of Crop Production and Seed Technology, Moi University, P.O.Box 1125, Eldoret, Kenya. 2Plant Research International, P.O.Box 6700 AA, Wageningen, The Netherlands. *Corresponding author: Tel- 0321-63212 Fax- 0321-63257 E-mail- [email protected]
Abstract The aim of the research was to investigate the effects of duration of soaking and concentration level of salicylic acid (SA) on seed germination of cabbage, cucumber and tomato. The planting materials of this study were F1 hybrid seeds of a cabbage cultivar “Bartolo”, cucumber cultivar “Pyralis” and tomato cultivar “Carmello”, all from seed companies in the Netherlands. In the first approach the seeds were soaked in various concentrations of SA for 24 hours, after which they were rinsed, blotted and dried. Four replicates each of 50 seeds of cabbage and tomato, and 25 seeds of cucumber were germinated. In the second approach four replicates each of 50 seeds of cabbage and tomato, and 25 seeds of cucumber were germinated on four layers of filter paper soaked continuously with various concentrations of SA. The data collected were processed using computer software package called SeedCalculator developed at the Plant Research International and comparisons of data means were performed with student t-Test. With cabbage, percentage maximum germination (Gmax) was higher for continuous soaking compared to 24-hour soaking while at higher SA levels Gmax was lower. In the case of tomato Gmax was higher for 24-hour soaking at lower SA levels, and lower for continuous soaking at increasing levels of SA. Continuous soaking had lower Gmax at increasing SA levels compared to 24-hour soaking. Time to attain 50% germination (T50) with 24-hour soaking was shorter at lower levels of SA in the case of cabbage and longer at increasing levels of SA. On the other hand continuous soaking had longer T50 at all levels of SA. For cucumber longer T50 was obtained with continuous soaking for all SA levels compared to 24-hour soaking. In the case of tomato 24-hour soaking had shorter T50 at lower levels of SA. With continuous soaking longer T50 was obtained at increasing SA levels compared to 24-hour soaking. Mean germination time (MGT) for cabbage was shorter at lower SA levels in the case of 24-hour soaking and longer at increasing levels of SA for continuous soaking. MGT for continuous soaking was shorter at higher SA levels and longer at lower levels, compared to 24-hour soaking. With cucumber longer MGT was obtained for continuous soaking at all SA levels compared to 24-hour soaking. For tomato MGT was shorter with 24-hour soaking at lower levels of SA and longer for continuous soaking at increasing levels. MGT for continuous soaking was longer at increasing levels of SA in the case of tomato, compared to 24-hour soaking. In conclusion the results obtained indicate that generally 24-hour soaking showed better germination performance than continuous soaking; lower concentration of SA gave better germination performance than higher concentration and cucumber behaved differently from cabbage and tomato. Key words: continuous soaking, primed, 24-hour soaking.
Introduction Seed quality is defined as “the sum of those properties that determine the potential level of activity and performance of the seed lot during germination and seedling emergence” (Perry 1978). These properties are related to parental genetic characters, ecological factors and management practices during seed production. A seed lot may have high germination capacity under the laboratory conditions, but may produce reduced number of normal seedlings under field conditions. Such differences normally lead to losses resulting from waste of plant material and inefficient use of facilities and labour. Losses of such nature are particularly high in the labour- and cost-intensive cultivation of vegetables. New methods are therefore developed to improve seed quality for expensive vegetable seeds such as cabbage, cucumber and tomato. In practice there is always need for the recognition and the selection of seeds of higher physiological quality, and sometimes presowing treatments can distinctly improve the

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 40
performance of such seeds. In an attempt to increase the uniformity and speed of germination, and to enable seeds to germinate under stress, seeds may be “primed” or “osmoconditioned” (Giesenberg and Stewart 1986). The process does not result in radicle emergence, so seed can be dried back and stored until planting. Coolbear et al. (1980) reported that both duration of soaking and the concentration of the osmoticum must be considered in order to determine optimum conditions. Green (1980) compared the use of primed tomato seeds, with other techniques of crop establishment and found out that in controlled environments the use of primed seed hastened emergence and increased the final percentage emergence in comparison with untreated seeds. On a field scale similar trends were observed and earlier ripe fruits were obtained. Salicylic acid (SA) is a natural plant growth regulator and has been suspected to play a role in defense response in plants, including the induction of plant resistance genes. Mills and Wood (1984) found that the pre-treatment of cucumber plants with salicylic, acetyl salicylic or polyacrylic acids induced local, and to a lesser extent systemic resistance to subsequent infection with Colletrotrichum lagenarium. From studies carried out by Shakirova and Bezrukova (1997) it was reported that exogenous treatment of wheat (Triticum aestivum) seeds with SA at 0.1 mM increased the rate of seed germination at various degrees of medium salinization (0.5-1.5% NaCl). Stitcher et al. (1997) report that SA can be phytotoxic, but when applied exogenously at optimal levels it can induce systemic resistance genes without lesion formation.
MATERIALS AND METHODS The research was conducted in the research laboratories of the Department of Plant Development and Reproduction, Plant Research International in the Netherlands, during the period of 11th June to 9th November, 1999. The planting materials were F1 hybrid seeds of a cabbage cultivar “Bartolo”, cucumber cultivar “Pyralis” and tomato cultivar “Carmello”, all obtained from seed companies in the Netherlands. Salicylic acid solution was prepared from sodium salt of salicylic acid (C7H5NaO3). A completely randomized design (CRD) was used and the various treatments were assigned at random in the germination cabinet. In the first approach seeds were put in stainless steel tea strainers made of fine wire mesh and steeped in 0,0, 0.1, 0.3. 1.0, 3.0 and 10.0 mM SA solutions in beakers. Seeds were soaked in the SA solutions for 24 hours. Seeds were then rinsed in running tap water for 5 minutes and blotted for a further 5 minutes, to remove adhering SA solution and water. Seeds were then dried back to 20 0C/32% relative humidity (RH) for 3 days in the dark. Four replicates each of 50 seeds of cabbage and tomato, and 25 seeds of cucumber were germinated on four layers of filter paper wetted with tap water, and placed in covered Petri dishes. The Petri dishes were arranged in covered plastic germination boxes then placed in a germination cabinet at 20 0C. The cabinet received 8 hours of light per day. Germination test was done according to International Seed Testing Association (ISTA) (1996). Germination counts were recorded daily up to the 4th day. In the second approach, four replicates each of 50 seeds of cabbage and tomato, and 25 seeds of cucumber were germinated on four layers of filter papers continuously soaked in 0.0, 0.1, 0.3, 1.0, 3.0 and 10.0 mM SA solutions and placed in covered Petri dishes. The Petri dishes were arranged in covered plastic germination boxes then placed in a germination cabinet at 20 0C. The cabinet received 8 hours of light per day. Germination test was similarly done according to ISTA (1996). Germination counts were recorded daily up to the 4th day. Data were taken on the following parameters: percentage maximum germination (Gmax), time in days to attain 50% maximum germination (T50) and mean germination time in days (MGT).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 41
The data collected were processed using computer software package called SeedCalculator developed at the Plant Research International, and comparisons of data means were performed with student t-Test.
Results As shown in Table 1, with 24-hour soaking percentage maximum germination (Gmax) was significantly lower (p=0.05) at 3.0 and 10.0 mM SA for cabbage; not significantly different (p=0.05) at all SA levels for cucumber, and significantly higher (p=0.05) at 0.1 and 3.0 mM SA for tomato. With continuous soaking Gmax was significantly lower (p=0.05) for cabbage at 10.0 mM SA; not significantly different (p=0.05) at all SA levels for cucumber, and significantly lower (p=0.05) at 0.3, 1.0, 3.0 and 10.0 mM SA for tomato. Gmax for continuous soaking was significantly higher (p=0.05) than for 24-hour soaking at 3.0 and 10.0 mM SA for cabbage; not significantly different (p=0.05) from 24-hour soaking at all SA levels for cucumber, and significantly lower (p=0.05) at 0.3, 1.0, 3.0 and 10.0 mM SA for tomato.
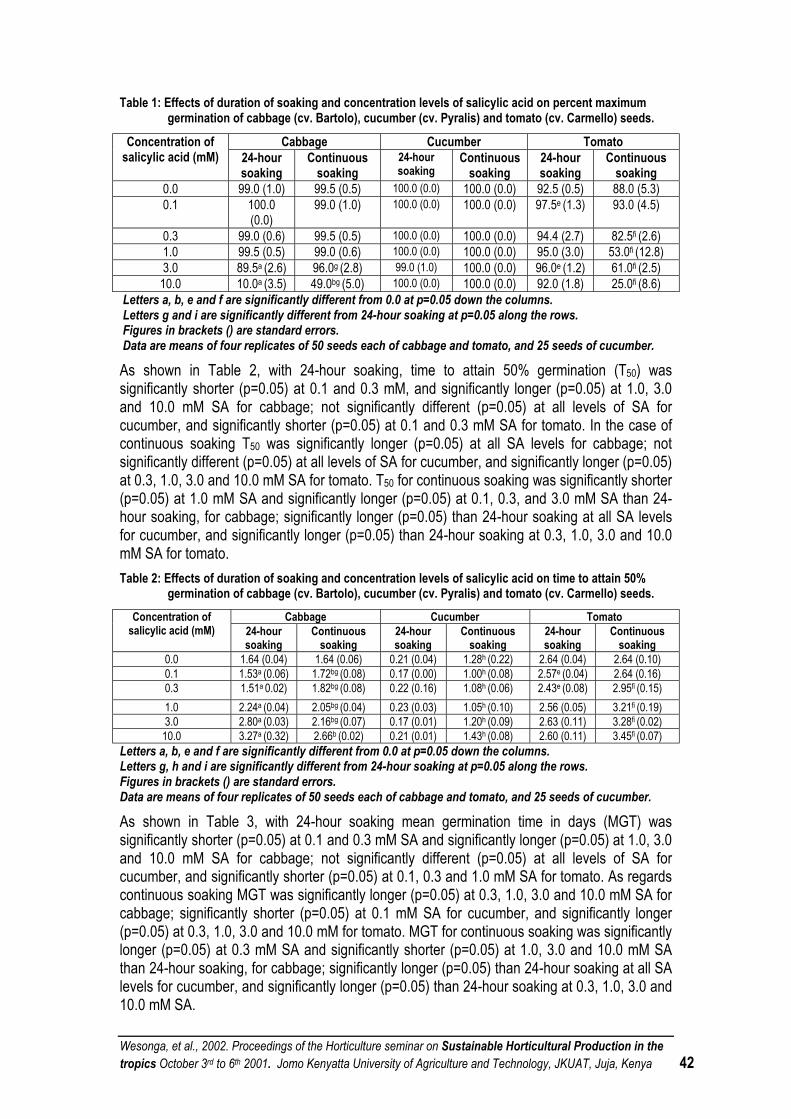
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 42
Table 1: Effects of duration of soaking and concentration levels of salicylic acid on percent maximum germination of cabbage (cv. Bartolo), cucumber (cv. Pyralis) and tomato (cv. Carmello) seeds.
Cabbage Cucumber Tomato Concentration of salicylic acid (mM) 24-hour
soaking Continuous
soaking 24-hour soaking
Continuous soaking
24-hour soaking
Continuous soaking
0.0 99.0 (1.0) 99.5 (0.5) 100.0 (0.0) 100.0 (0.0) 92.5 (0.5) 88.0 (5.3) 0.1 100.0
(0.0) 99.0 (1.0) 100.0 (0.0) 100.0 (0.0) 97.5e (1.3) 93.0 (4.5)
0.3 99.0 (0.6) 99.5 (0.5) 100.0 (0.0) 100.0 (0.0) 94.4 (2.7) 82.5fi (2.6) 1.0 99.5 (0.5) 99.0 (0.6) 100.0 (0.0) 100.0 (0.0) 95.0 (3.0) 53.0fi (12.8) 3.0 89.5a (2.6) 96.0g (2.8) 99.0 (1.0) 100.0 (0.0) 96.0e (1.2) 61.0fi (2.5)
10.0 10.0a (3.5) 49.0bg (5.0) 100.0 (0.0) 100.0 (0.0) 92.0 (1.8) 25.0fi (8.6) Letters a, b, e and f are significantly different from 0.0 at p=0.05 down the columns. Letters g and i are significantly different from 24-hour soaking at p=0.05 along the rows. Figures in brackets () are standard errors. Data are means of four replicates of 50 seeds each of cabbage and tomato, and 25 seeds of cucumber.
As shown in Table 2, with 24-hour soaking, time to attain 50% germination (T50) was significantly shorter (p=0.05) at 0.1 and 0.3 mM, and significantly longer (p=0.05) at 1.0, 3.0 and 10.0 mM SA for cabbage; not significantly different (p=0.05) at all levels of SA for cucumber, and significantly shorter (p=0.05) at 0.1 and 0.3 mM SA for tomato. In the case of continuous soaking T50 was significantly longer (p=0.05) at all SA levels for cabbage; not significantly different (p=0.05) at all levels of SA for cucumber, and significantly longer (p=0.05) at 0.3, 1.0, 3.0 and 10.0 mM SA for tomato. T50 for continuous soaking was significantly shorter (p=0.05) at 1.0 mM SA and significantly longer (p=0.05) at 0.1, 0.3, and 3.0 mM SA than 24-hour soaking, for cabbage; significantly longer (p=0.05) than 24-hour soaking at all SA levels for cucumber, and significantly longer (p=0.05) than 24-hour soaking at 0.3, 1.0, 3.0 and 10.0 mM SA for tomato. Table 2: Effects of duration of soaking and concentration levels of salicylic acid on time to attain 50%
germination of cabbage (cv. Bartolo), cucumber (cv. Pyralis) and tomato (cv. Carmello) seeds. Cabbage Cucumber Tomato Concentration of
salicylic acid (mM) 24-hour soaking
Continuous soaking
24-hour soaking
Continuous soaking
24-hour soaking
Continuous soaking
0.0 1.64 (0.04) 1.64 (0.06) 0.21 (0.04) 1.28h (0.22) 2.64 (0.04) 2.64 (0.10) 0.1 1.53a (0.06) 1.72bg (0.08) 0.17 (0.00) 1.00h (0.08) 2.57e (0.04) 2.64 (0.16) 0.3 1.51a 0.02) 1.82bg (0.08) 0.22 (0.16) 1.08h (0.06) 2.43e (0.08) 2.95fi (0.15) 1.0 2.24a (0.04) 2.05bg (0.04) 0.23 (0.03) 1.05h (0.10) 2.56 (0.05) 3.21fi (0.19) 3.0 2.80a (0.03) 2.16bg (0.07) 0.17 (0.01) 1.20h (0.09) 2.63 (0.11) 3.28fi (0.02) 10.0 3.27a (0.32) 2.66b (0.02) 0.21 (0.01) 1.43h (0.08) 2.60 (0.11) 3.45fi (0.07)
Letters a, b, e and f are significantly different from 0.0 at p=0.05 down the columns. Letters g, h and i are significantly different from 24-hour soaking at p=0.05 along the rows. Figures in brackets () are standard errors. Data are means of four replicates of 50 seeds each of cabbage and tomato, and 25 seeds of cucumber.
As shown in Table 3, with 24-hour soaking mean germination time in days (MGT) was significantly shorter (p=0.05) at 0.1 and 0.3 mM SA and significantly longer (p=0.05) at 1.0, 3.0 and 10.0 mM SA for cabbage; not significantly different (p=0.05) at all levels of SA for cucumber, and significantly shorter (p=0.05) at 0.1, 0.3 and 1.0 mM SA for tomato. As regards continuous soaking MGT was significantly longer (p=0.05) at 0.3, 1.0, 3.0 and 10.0 mM SA for cabbage; significantly shorter (p=0.05) at 0.1 mM SA for cucumber, and significantly longer (p=0.05) at 0.3, 1.0, 3.0 and 10.0 mM for tomato. MGT for continuous soaking was significantly longer (p=0.05) at 0.3 mM SA and significantly shorter (p=0.05) at 1.0, 3.0 and 10.0 mM SA than 24-hour soaking, for cabbage; significantly longer (p=0.05) than 24-hour soaking at all SA levels for cucumber, and significantly longer (p=0.05) than 24-hour soaking at 0.3, 1.0, 3.0 and 10.0 mM SA.
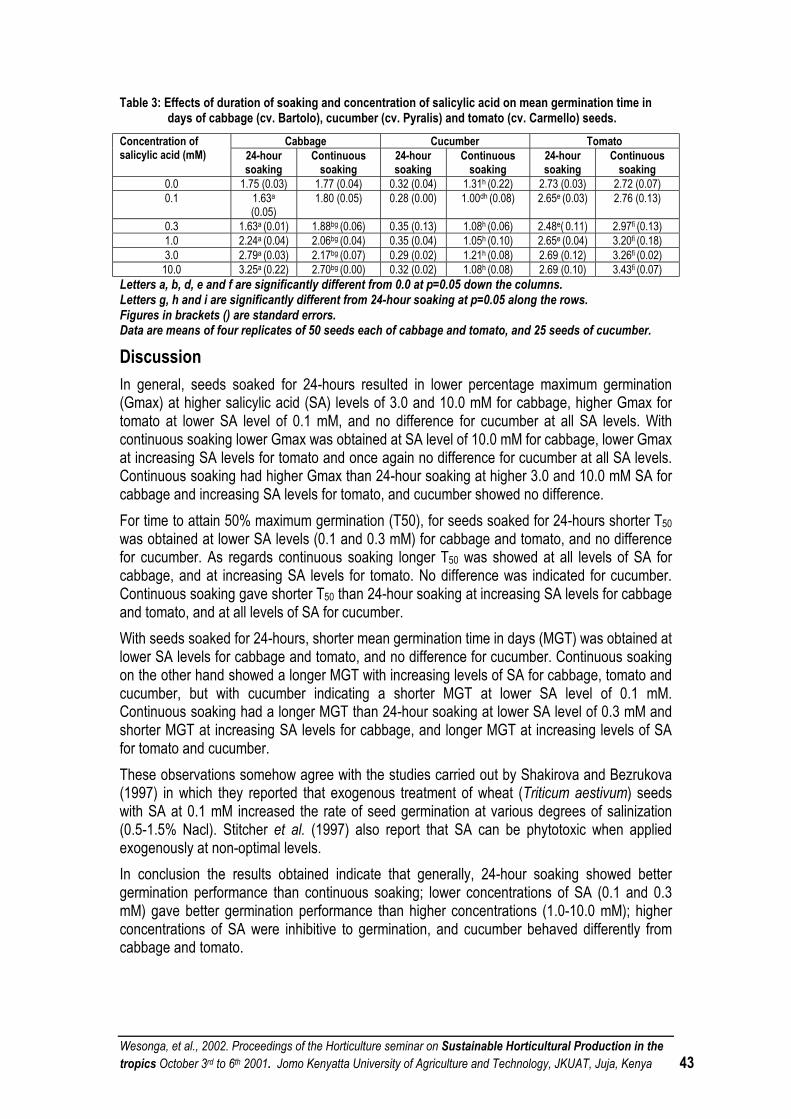
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 43
Table 3: Effects of duration of soaking and concentration of salicylic acid on mean germination time in days of cabbage (cv. Bartolo), cucumber (cv. Pyralis) and tomato (cv. Carmello) seeds.
Cabbage Cucumber Tomato Concentration of salicylic acid (mM) 24-hour
soaking Continuous
soaking 24-hour soaking
Continuous soaking
24-hour soaking
Continuous soaking
0.0 1.75 (0.03) 1.77 (0.04) 0.32 (0.04) 1.31h (0.22) 2.73 (0.03) 2.72 (0.07) 0.1 1.63a
(0.05) 1.80 (0.05) 0.28 (0.00) 1.00dh (0.08) 2.65e (0.03) 2.76 (0.13)
0.3 1.63a (0.01) 1.88bg (0.06) 0.35 (0.13) 1.08h (0.06) 2.48e( 0.11) 2.97fi (0.13) 1.0 2.24a (0.04) 2.06bg (0.04) 0.35 (0.04) 1.05h (0.10) 2.65e (0.04) 3.20fi (0.18) 3.0 2.79a (0.03) 2.17bg (0.07) 0.29 (0.02) 1.21h (0.08) 2.69 (0.12) 3.26fi (0.02) 10.0 3.25a (0.22) 2.70bg (0.00) 0.32 (0.02) 1.08h (0.08) 2.69 (0.10) 3.43fi (0.07)
Letters a, b, d, e and f are significantly different from 0.0 at p=0.05 down the columns. Letters g, h and i are significantly different from 24-hour soaking at p=0.05 along the rows. Figures in brackets () are standard errors. Data are means of four replicates of 50 seeds each of cabbage and tomato, and 25 seeds of cucumber.
Discussion In general, seeds soaked for 24-hours resulted in lower percentage maximum germination (Gmax) at higher salicylic acid (SA) levels of 3.0 and 10.0 mM for cabbage, higher Gmax for tomato at lower SA level of 0.1 mM, and no difference for cucumber at all SA levels. With continuous soaking lower Gmax was obtained at SA level of 10.0 mM for cabbage, lower Gmax at increasing SA levels for tomato and once again no difference for cucumber at all SA levels. Continuous soaking had higher Gmax than 24-hour soaking at higher 3.0 and 10.0 mM SA for cabbage and increasing SA levels for tomato, and cucumber showed no difference. For time to attain 50% maximum germination (T50), for seeds soaked for 24-hours shorter T50 was obtained at lower SA levels (0.1 and 0.3 mM) for cabbage and tomato, and no difference for cucumber. As regards continuous soaking longer T50 was showed at all levels of SA for cabbage, and at increasing SA levels for tomato. No difference was indicated for cucumber. Continuous soaking gave shorter T50 than 24-hour soaking at increasing SA levels for cabbage and tomato, and at all levels of SA for cucumber. With seeds soaked for 24-hours, shorter mean germination time in days (MGT) was obtained at lower SA levels for cabbage and tomato, and no difference for cucumber. Continuous soaking on the other hand showed a longer MGT with increasing levels of SA for cabbage, tomato and cucumber, but with cucumber indicating a shorter MGT at lower SA level of 0.1 mM. Continuous soaking had a longer MGT than 24-hour soaking at lower SA level of 0.3 mM and shorter MGT at increasing SA levels for cabbage, and longer MGT at increasing levels of SA for tomato and cucumber. These observations somehow agree with the studies carried out by Shakirova and Bezrukova (1997) in which they reported that exogenous treatment of wheat (Triticum aestivum) seeds with SA at 0.1 mM increased the rate of seed germination at various degrees of salinization (0.5-1.5% Nacl). Stitcher et al. (1997) also report that SA can be phytotoxic when applied exogenously at non-optimal levels. In conclusion the results obtained indicate that generally, 24-hour soaking showed better germination performance than continuous soaking; lower concentrations of SA (0.1 and 0.3 mM) gave better germination performance than higher concentrations (1.0-10.0 mM); higher concentrations of SA were inhibitive to germination, and cucumber behaved differently from cabbage and tomato.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 44
References Coolbear, P., D. Grierson and W. Heydecker (1980). Osmotic pretreatment and nucleic acid
accumulation in tomato seed (Lycopersicon esculentum). Seed Sci. Technol. 8: 289-303.
Giesenberg C. and K. Stewart (1986). In The Tomato Crop: A Scientific basis for improvement, eds. J.G. Atherton and J. Rudich. Chapman and Hall, London, New York, pp. 552-557.
Green, J.C. (1980). Plant density and crop establishment studies with tomato. Acta Hort. 400: 129-135.
Mills, P.R. and R.K.S. Wood (1984). The effects of polyacrylic acid, acetylsalicylic acid and salicylic acid on resistance of cucumber to Colletotrichum lagenarium. Phytopathologische Zeitschroft 111: 209-216.
Perry, D.A. (1978). Report of the vigour test committee 1974-1977. Seed Sci. & Technol. 6: 159-181.
Shakirova, F.I. and M.V. Bezrukova (1997). Induction of wheat response against environmental salinization by indolylacetic acid. Izvestiya Akademic Nauk Seviya Biologicheskaya (Moscow) 0 (2): 149-153.
Stitcher, L., B. Mauch-Mani and J.P. Me’traux (1997). Systemic Acquired Resistance. Annual Rev. Phytopathol. 35: 235-270.
Discussion Q: Have you compared other seed priming chemicals such as PEG to salicyclic acid and how do they compare? A: Yes; this is just but one of the studies I carried out But one other study I carried out I used PEG without salcyclic acid (SA). in one particular instance, I studied seed storability by using PEG alone, PEG + SA and SA alone: the results obtained indicated that SA used alone gave higher storability in terms of the germination parameters measured. Q Can these methods be recommended for other crops? A: It is worth trying with other crops in Kenya and even other Kenyan varieties of tomato, cabbage etc Q: Are the germination percentages of the three vegetable low in Kenya. If not what was the rational of carrying out the study? A: In priming the germination processes are initiated in the seed, but these are stopped just before the radical protrusion. The seeds may be planted soon thereafter or can be Kept/stored to be used later. When planted they will germinate immediately and even have a faster speed of growth. In Kenya, some seeds are soaked to enhance germination, but since the radicle normally come out, such seeds have to be planted immediately. They cannot be kept/stored to be panted later lest they die.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 45
EFFECT OF IRRIGATION MANAGEMENT ON YIELD, WATER AND NITROGEN USE OF OKRA (ABELMOSCHUS ESCULENTUS L.) P.G. Home Department of Biomechanical and Environmental Engineering, Jomo Kenyatta University of Agriculture and Technology, P.O. Box 62000, Nairobi, Kenya. E-mail: [email protected]
Abstract Production of fresh vegetables usually calls for application of large amounts of irrigation water and fertilizer nitrogen (N). Combined application of high rates of water and N leads to excessive leaching of nitrate nitrogen, making most of it unavailable to the plants. Very few studies have been conducted to investigate the combined effect of irrigation method and scheduling on the yield, N-use (based on N balance in the root zone) and N uptake (based on N content of plant material) of vegetable crops, especially Okra. A field experiment was conducted in a coarse textured lateritic soil (Haplustalf) planted to Okra to investigate the effect of method and scheduling of irrigation on yield attributes. Three methods of irrigation: sprinkler, furrow and basin and three irrigation treatments scheduled at 15, 30, 45 and 60% maximum allowable depletion (MAD) of available soil water were studied. The results of the study revealed that on sandy loam lateritic soil planted to Okra crop, maximum fresh fruit yield, nitrogen uptake and nitrogen uptake efficiency of Okra were obtained with sprinkler irrigation scheduled at 30% MAD, whereas the maximum nitrogen use efficiency was obtained with furrow irrigation scheduled at 15% MAD. Irrigation schedules with 60% MAD resulted in the minimum fresh fruit yield, nitrogen use, nitrogen uptake as well as the least nitrogen use and uptake efficiencies irrespective of irrigation method. Thus, scheduling irrigation at 30% MAD under sprinkler method of irrigation was found to be the best for okra crop in sandy loam soil. Key words: Irrigation method, scheduling, deep percolation, leaching of nitrate, crop yield.
Introduction The world food production is highly dependent on irrigated agriculture. In the irrigated agriculture, there has been a progressive an intensified use of increasing quantities of water and chemical fertilizers. Currently there are about 250 million hectares of irrigated land worldwide. Although this constitutes only 17% of the total agricultural land, it produces 36% of the world’s total food requirement. Despite high levels of productivity, irrigated lands are posed with the concomitant problems of high losses of nitrogen through leaching of nitrate-nitrogen, waterlogging, and salinisation. Proper irrigation management is a major factor in the efficient uptake and utilization of applied plant nutrients, and especially so for the highly soluble nitrogenous fertilizers (Ongley, 1996; Pereira, 1999). Irrigation schedules that maintained soil moisture near the field capacity significantly enhances the yield and N uptake, especially for vegetables as well as field crops (Tisdale et al., 1990, Forster et al., 1995). Chartzoulakis and Doros (1995) found that for vegetables, the maximum water use as well as yield were obtained when soil water potential was maintained higher than –20 kPa, and there was a drastic yield reduction when irrigation was scheduled at 45% of the potential evapotranspiration. Hence, Scientific management of water and fertilizer inputs thus becomes imperative in the conservation and protection of the precious land and water resources. Among the chemical fertilizers nitrogen (N) is the most abundantly used crop nutrient. This is especially so in case of vegetable crops where the interest is on the fresh harvest. In quest of augmenting production, there has been a tendency to adopt high application rates of N and irrigation water, often together. Such combined application of high rates of water and N leads to excessive leaching of NO3-N, making most of it unavailable to the plants and contaminating the ground water. An estimated 25-50% of applied N is lost by leaching (Ongley, 1996). The amount of nitrogen lost through leaching is dependent on water and fertilizer management strategies adopted in the farming system. Irrigation management is even of greater importance than N management in reducing the leaching loss of N. It has been found that in irrigated
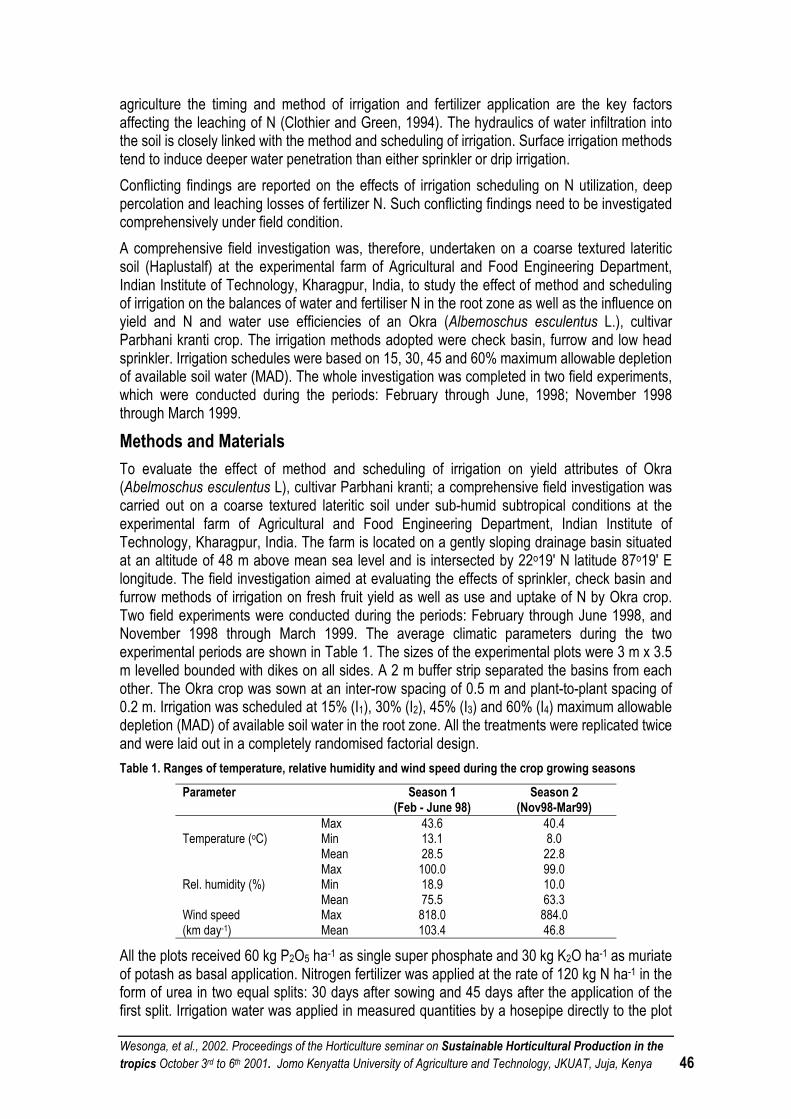
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 46
agriculture the timing and method of irrigation and fertilizer application are the key factors affecting the leaching of N (Clothier and Green, 1994). The hydraulics of water infiltration into the soil is closely linked with the method and scheduling of irrigation. Surface irrigation methods tend to induce deeper water penetration than either sprinkler or drip irrigation. Conflicting findings are reported on the effects of irrigation scheduling on N utilization, deep percolation and leaching losses of fertilizer N. Such conflicting findings need to be investigated comprehensively under field condition. A comprehensive field investigation was, therefore, undertaken on a coarse textured lateritic soil (Haplustalf) at the experimental farm of Agricultural and Food Engineering Department, Indian Institute of Technology, Kharagpur, India, to study the effect of method and scheduling of irrigation on the balances of water and fertiliser N in the root zone as well as the influence on yield and N and water use efficiencies of an Okra (Albemoschus esculentus L.), cultivar Parbhani kranti crop. The irrigation methods adopted were check basin, furrow and low head sprinkler. Irrigation schedules were based on 15, 30, 45 and 60% maximum allowable depletion of available soil water (MAD). The whole investigation was completed in two field experiments, which were conducted during the periods: February through June, 1998; November 1998 through March 1999.
Methods and Materials To evaluate the effect of method and scheduling of irrigation on yield attributes of Okra (Abelmoschus esculentus L), cultivar Parbhani kranti; a comprehensive field investigation was carried out on a coarse textured lateritic soil under sub-humid subtropical conditions at the experimental farm of Agricultural and Food Engineering Department, Indian Institute of Technology, Kharagpur, India. The farm is located on a gently sloping drainage basin situated at an altitude of 48 m above mean sea level and is intersected by 22o19' N latitude 87o19' E longitude. The field investigation aimed at evaluating the effects of sprinkler, check basin and furrow methods of irrigation on fresh fruit yield as well as use and uptake of N by Okra crop. Two field experiments were conducted during the periods: February through June 1998, and November 1998 through March 1999. The average climatic parameters during the two experimental periods are shown in Table 1. The sizes of the experimental plots were 3 m x 3.5 m levelled bounded with dikes on all sides. A 2 m buffer strip separated the basins from each other. The Okra crop was sown at an inter-row spacing of 0.5 m and plant-to-plant spacing of 0.2 m. Irrigation was scheduled at 15% (I1), 30% (I2), 45% (I3) and 60% (I4) maximum allowable depletion (MAD) of available soil water in the root zone. All the treatments were replicated twice and were laid out in a completely randomised factorial design. Table 1. Ranges of temperature, relative humidity and wind speed during the crop growing seasons
Parameter Season 1 (Feb - June 98)
Season 2 (Nov98-Mar99)
Max 43.6 40.4 Min 13.1 8.0
Temperature (oC)
Mean 28.5 22.8 Max 100.0 99.0 Min 18.9 10.0
Rel. humidity (%)
Mean 75.5 63.3 Max 818.0 884.0 Wind speed
(km day-1) Mean 103.4 46.8
All the plots received 60 kg P2O5 ha-1 as single super phosphate and 30 kg K2O ha-1 as muriate of potash as basal application. Nitrogen fertilizer was applied at the rate of 120 kg N ha-1 in the form of urea in two equal splits: 30 days after sowing and 45 days after the application of the first split. Irrigation water was applied in measured quantities by a hosepipe directly to the plot

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 47
under check basin and furrow irrigation and by four micro-sprinklers per plot under sprinkler irrigation, in order to apply the estimated amount of water for replenishing the root zone to field capacity as soon as the predefined MAD limit was attained. A water meter was used to measure the amount of water applied. For water and N balance studies, soil moisture was monitored using the Hydroprobe, gypsum based Soil moisture sensors as well as by gravimetric measurements. Soil samples were collected regularly at two weeks intervals and also a day before and 2-3 days after irrigation at 15, 30, 50, 75 and 90 cm depths from the ground surface for the determination of soil moisture. Same soil samples were utilized for the determination of water soluble NO3-N and NH4-N. The concentration of water soluble NH4-N and NO3-N in the soil samples were estimated with the help of an ORION Ion meter using specific ion electrodes for NH4-N and NO3-N. The N content in the plants was determined by the Kjeldahl digestion method (Jackson, 1973). To estimate the water and N balance components, the magnitude of deep percolation (DP) rate was estimated from the hydraulic conductivity of the soil as a function of water content at 60 cm depth estimated using van Genuchten (1980) relation derived from the soil moisture characteristics and assuming a unit hydraulic gradient. The leaching loss of N was calculated from the DP and the average concentration of NO3-N in the leachate.
Results The effect of method and scheduling of irrigation on deep percolation (DP) and evapotranspiration (ET) is presented in Tables 2 and 3 respectively for experiment 1 and experiment 2. The data shows that irrigation schedules significantly influenced DP and ET with the wet regime (I1 and I2) resulting in higher values of DP and ET than the dry regime (I3 and I4) during both the experiments. The differences in DP were more apparent in experiment 2 where there was minimal interference of rainfall. The DP when irrigation was scheduled at 15% MAD was almost twice that at 30% MAD schedule. The DP at 60% MAD schedule was only slightly lower (maximum difference of 3.6 mm for furrow irrigation) than that at 30% MAD irrigation schedule (Table 3). During experiment 1 as well, DP was also highest at 15% MAD, but in this case DP at 30% MAD was about 88% of that at 15% MAD. In general DP decreased with the increase in allowed maximum depletion. The impact of method of irrigation on DP was not significant at 5% level during experiment 2, but during experiment 1 significantly higher DP was observed in furrow irrigated than sprinkler or basin irrigated fields. Sprinkler irrigated plots had the least DP. Evapotranspiration (ET) was significantly influenced by irrigation scheduling during both the experiment. During experiment 2, seasonal ET at 60% MAD schedule was on an average 50 mm lower than ET under 30% MAD schedule. Incidentally, ET at 15% MAD was slightly lower than at 30% MAD. However, irrigation method did not significantly influence Okra crop evapotranspiration.
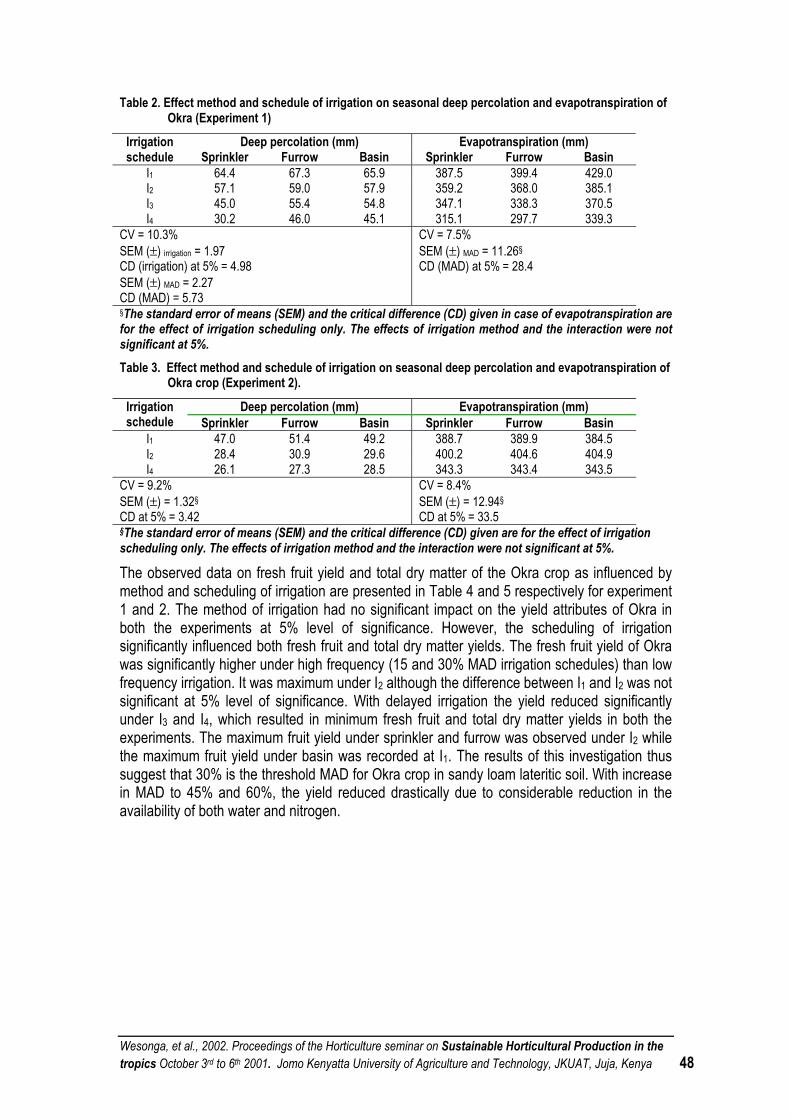
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 48
Table 2. Effect method and schedule of irrigation on seasonal deep percolation and evapotranspiration of Okra (Experiment 1)
Deep percolation (mm) Evapotranspiration (mm) Irrigation schedule Sprinkler Furrow Basin Sprinkler Furrow Basin
I1 64.4 67.3 65.9 387.5 399.4 429.0 I2 57.1 59.0 57.9 359.2 368.0 385.1 I3 45.0 55.4 54.8 347.1 338.3 370.5 I4 30.2 46.0 45.1 315.1 297.7 339.3
CV = 10.3% SEM (±) irrigation = 1.97 CD (irrigation) at 5% = 4.98 SEM (±) MAD = 2.27 CD (MAD) = 5.73
CV = 7.5% SEM (±) MAD = 11.26§ CD (MAD) at 5% = 28.4
§The standard error of means (SEM) and the critical difference (CD) given in case of evapotranspiration are for the effect of irrigation scheduling only. The effects of irrigation method and the interaction were not significant at 5%. Table 3. Effect method and schedule of irrigation on seasonal deep percolation and evapotranspiration of
Okra crop (Experiment 2).
Deep percolation (mm) Evapotranspiration (mm) Irrigation schedule Sprinkler Furrow Basin Sprinkler Furrow Basin
I1 47.0 51.4 49.2 388.7 389.9 384.5 I2 28.4 30.9 29.6 400.2 404.6 404.9 I4 26.1 27.3 28.5 343.3 343.4 343.5
CV = 9.2% SEM (±) = 1.32§ CD at 5% = 3.42
CV = 8.4% SEM (±) = 12.94§ CD at 5% = 33.5
§The standard error of means (SEM) and the critical difference (CD) given are for the effect of irrigation scheduling only. The effects of irrigation method and the interaction were not significant at 5%.
The observed data on fresh fruit yield and total dry matter of the Okra crop as influenced by method and scheduling of irrigation are presented in Table 4 and 5 respectively for experiment 1 and 2. The method of irrigation had no significant impact on the yield attributes of Okra in both the experiments at 5% level of significance. However, the scheduling of irrigation significantly influenced both fresh fruit and total dry matter yields. The fresh fruit yield of Okra was significantly higher under high frequency (15 and 30% MAD irrigation schedules) than low frequency irrigation. It was maximum under I2 although the difference between I1 and I2 was not significant at 5% level of significance. With delayed irrigation the yield reduced significantly under I3 and I4, which resulted in minimum fresh fruit and total dry matter yields in both the experiments. The maximum fruit yield under sprinkler and furrow was observed under I2 while the maximum fruit yield under basin was recorded at I1. The results of this investigation thus suggest that 30% is the threshold MAD for Okra crop in sandy loam lateritic soil. With increase in MAD to 45% and 60%, the yield reduced drastically due to considerable reduction in the availability of both water and nitrogen.
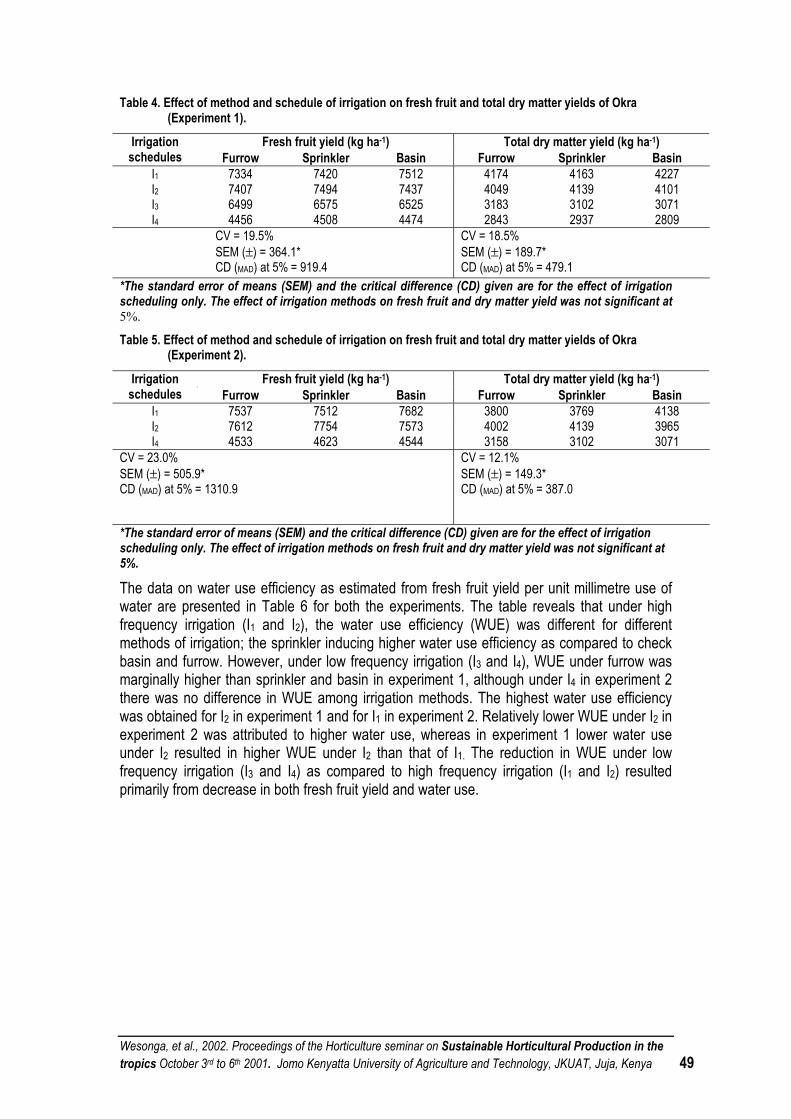
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 49
Table 4. Effect of method and schedule of irrigation on fresh fruit and total dry matter yields of Okra (Experiment 1).
Fresh fruit yield (kg ha-1) Total dry matter yield (kg ha-1) Irrigation schedules Furrow Sprinkler Basin Furrow Sprinkler Basin
I1 7334 7420 7512 4174 4163 4227 I2 7407 7494 7437 4049 4139 4101 I3 6499 6575 6525 3183 3102 3071 I4 4456 4508 4474 2843 2937 2809
CV = 19.5% SEM (±) = 364.1* CD (MAD) at 5% = 919.4
CV = 18.5% SEM (±) = 189.7* CD (MAD) at 5% = 479.1
*The standard error of means (SEM) and the critical difference (CD) given are for the effect of irrigation scheduling only. The effect of irrigation methods on fresh fruit and dry matter yield was not significant at 5%.
Table 5. Effect of method and schedule of irrigation on fresh fruit and total dry matter yields of Okra (Experiment 2).
Fresh fruit yield (kg ha-1) Total dry matter yield (kg ha-1) Irrigation schedules Furrow Sprinkler Basin Furrow Sprinkler Basin
I1 7537 7512 7682 3800 3769 4138 I2 7612 7754 7573 4002 4139 3965 I4 4533 4623 4544 3158 3102 3071
CV = 23.0% SEM (±) = 505.9* CD (MAD) at 5% = 1310.9
CV = 12.1% SEM (±) = 149.3* CD (MAD) at 5% = 387.0
*The standard error of means (SEM) and the critical difference (CD) given are for the effect of irrigation scheduling only. The effect of irrigation methods on fresh fruit and dry matter yield was not significant at 5%.
The data on water use efficiency as estimated from fresh fruit yield per unit millimetre use of water are presented in Table 6 for both the experiments. The table reveals that under high frequency irrigation (I1 and I2), the water use efficiency (WUE) was different for different methods of irrigation; the sprinkler inducing higher water use efficiency as compared to check basin and furrow. However, under low frequency irrigation (I3 and I4), WUE under furrow was marginally higher than sprinkler and basin in experiment 1, although under I4 in experiment 2 there was no difference in WUE among irrigation methods. The highest water use efficiency was obtained for I2 in experiment 1 and for I1 in experiment 2. Relatively lower WUE under I2 in experiment 2 was attributed to higher water use, whereas in experiment 1 lower water use under I2 resulted in higher WUE under I2 than that of I1. The reduction in WUE under low frequency irrigation (I3 and I4) as compared to high frequency irrigation (I1 and I2) resulted primarily from decrease in both fresh fruit yield and water use.
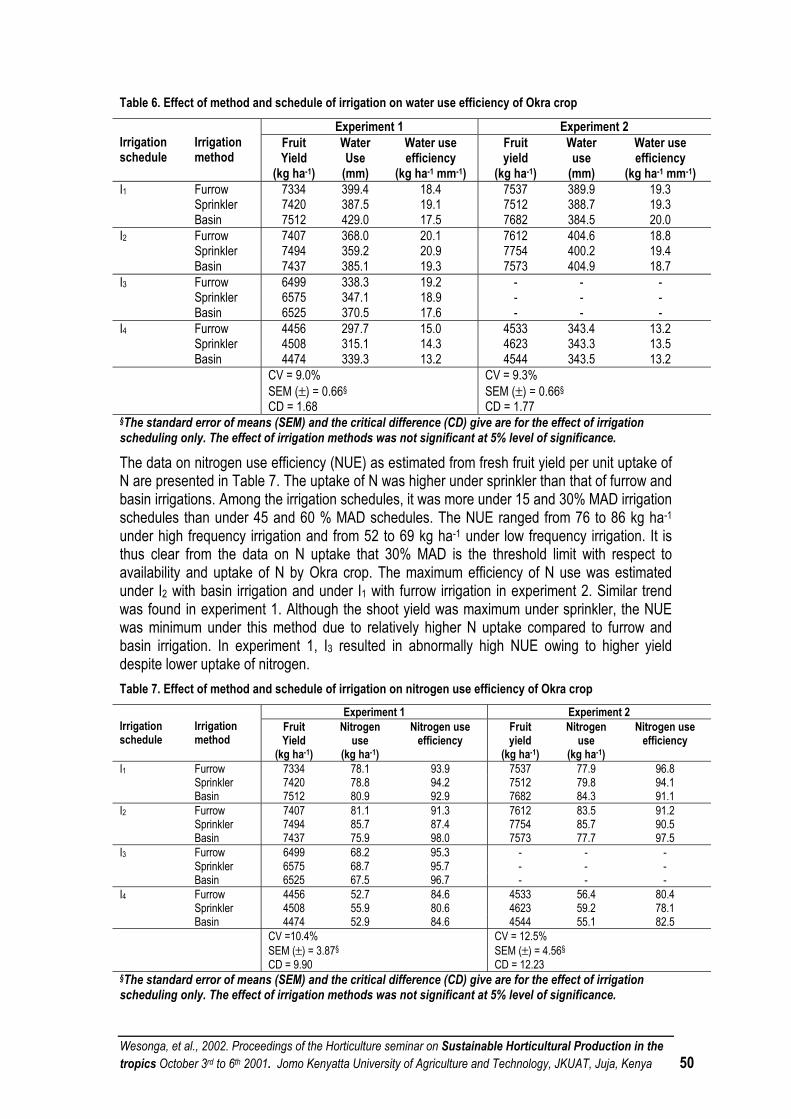
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 50
Table 6. Effect of method and schedule of irrigation on water use efficiency of Okra crop
Experiment 1 Experiment 2 Irrigation schedule
Irrigation method
Fruit Yield
(kg ha-1)
Water Use
(mm)
Water use efficiency
(kg ha-1 mm-1)
Fruit yield
(kg ha-1)
Water use
(mm)
Water use efficiency
(kg ha-1 mm-1) I1 Furrow 7334 399.4 18.4 7537 389.9 19.3 Sprinkler 7420 387.5 19.1 7512 388.7 19.3 Basin 7512 429.0 17.5 7682 384.5 20.0 I2 Furrow 7407 368.0 20.1 7612 404.6 18.8 Sprinkler 7494 359.2 20.9 7754 400.2 19.4 Basin 7437 385.1 19.3 7573 404.9 18.7 I3 Furrow 6499 338.3 19.2 - - - Sprinkler 6575 347.1 18.9 - - - Basin 6525 370.5 17.6 - - - I4 Furrow 4456 297.7 15.0 4533 343.4 13.2 Sprinkler 4508 315.1 14.3 4623 343.3 13.5 Basin 4474 339.3 13.2 4544 343.5 13.2 CV = 9.0%
SEM (±) = 0.66§ CD = 1.68
CV = 9.3% SEM (±) = 0.66§ CD = 1.77
§The standard error of means (SEM) and the critical difference (CD) give are for the effect of irrigation scheduling only. The effect of irrigation methods was not significant at 5% level of significance.
The data on nitrogen use efficiency (NUE) as estimated from fresh fruit yield per unit uptake of N are presented in Table 7. The uptake of N was higher under sprinkler than that of furrow and basin irrigations. Among the irrigation schedules, it was more under 15 and 30% MAD irrigation schedules than under 45 and 60 % MAD schedules. The NUE ranged from 76 to 86 kg ha-1 under high frequency irrigation and from 52 to 69 kg ha-1 under low frequency irrigation. It is thus clear from the data on N uptake that 30% MAD is the threshold limit with respect to availability and uptake of N by Okra crop. The maximum efficiency of N use was estimated under I2 with basin irrigation and under I1 with furrow irrigation in experiment 2. Similar trend was found in experiment 1. Although the shoot yield was maximum under sprinkler, the NUE was minimum under this method due to relatively higher N uptake compared to furrow and basin irrigation. In experiment 1, I3 resulted in abnormally high NUE owing to higher yield despite lower uptake of nitrogen. Table 7. Effect of method and schedule of irrigation on nitrogen use efficiency of Okra crop
Experiment 1 Experiment 2 Irrigation schedule
Irrigation method
Fruit Yield
(kg ha-1)
Nitrogen use
(kg ha-1)
Nitrogen use efficiency
Fruit yield
(kg ha-1)
Nitrogen use
(kg ha-1)
Nitrogen use efficiency
I1 Furrow 7334 78.1 93.9 7537 77.9 96.8 Sprinkler 7420 78.8 94.2 7512 79.8 94.1 Basin 7512 80.9 92.9 7682 84.3 91.1 I2 Furrow 7407 81.1 91.3 7612 83.5 91.2 Sprinkler 7494 85.7 87.4 7754 85.7 90.5 Basin 7437 75.9 98.0 7573 77.7 97.5 I3 Furrow 6499 68.2 95.3 - - - Sprinkler 6575 68.7 95.7 - - - Basin 6525 67.5 96.7 - - - I4 Furrow 4456 52.7 84.6 4533 56.4 80.4 Sprinkler 4508 55.9 80.6 4623 59.2 78.1 Basin 4474 52.9 84.6 4544 55.1 82.5 CV =10.4%
SEM (±) = 3.87§ CD = 9.90
CV = 12.5% SEM (±) = 4.56§ CD = 12.23
§The standard error of means (SEM) and the critical difference (CD) give are for the effect of irrigation scheduling only. The effect of irrigation methods was not significant at 5% level of significance.
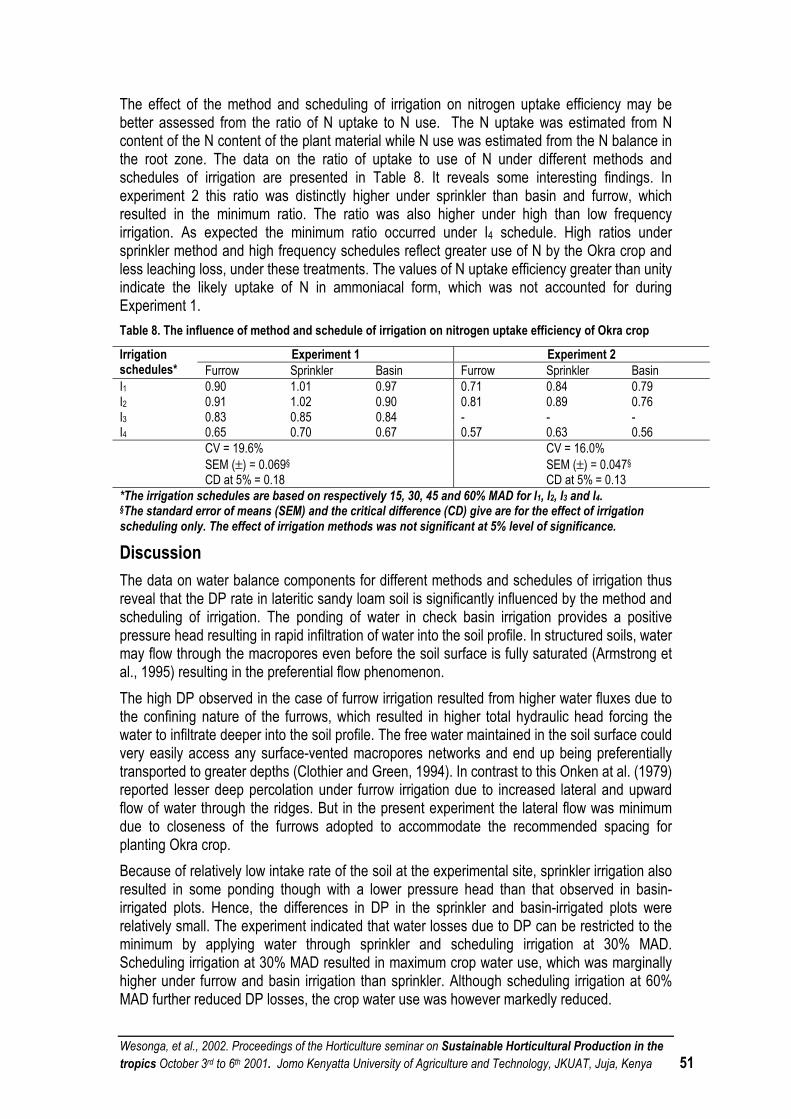
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 51
The effect of the method and scheduling of irrigation on nitrogen uptake efficiency may be better assessed from the ratio of N uptake to N use. The N uptake was estimated from N content of the N content of the plant material while N use was estimated from the N balance in the root zone. The data on the ratio of uptake to use of N under different methods and schedules of irrigation are presented in Table 8. It reveals some interesting findings. In experiment 2 this ratio was distinctly higher under sprinkler than basin and furrow, which resulted in the minimum ratio. The ratio was also higher under high than low frequency irrigation. As expected the minimum ratio occurred under I4 schedule. High ratios under sprinkler method and high frequency schedules reflect greater use of N by the Okra crop and less leaching loss, under these treatments. The values of N uptake efficiency greater than unity indicate the likely uptake of N in ammoniacal form, which was not accounted for during Experiment 1. Table 8. The influence of method and schedule of irrigation on nitrogen uptake efficiency of Okra crop
Experiment 1 Experiment 2 Irrigation schedules* Furrow Sprinkler Basin Furrow Sprinkler Basin I1 0.90 1.01 0.97 0.71 0.84 0.79 I2 0.91 1.02 0.90 0.81 0.89 0.76 I3 0.83 0.85 0.84 - - - I4 0.65 0.70 0.67 0.57 0.63 0.56 CV = 19.6%
SEM (±) = 0.069§ CD at 5% = 0.18
CV = 16.0% SEM (±) = 0.047§ CD at 5% = 0.13
*The irrigation schedules are based on respectively 15, 30, 45 and 60% MAD for I1, I2, I3 and I4. §The standard error of means (SEM) and the critical difference (CD) give are for the effect of irrigation scheduling only. The effect of irrigation methods was not significant at 5% level of significance.
Discussion The data on water balance components for different methods and schedules of irrigation thus reveal that the DP rate in lateritic sandy loam soil is significantly influenced by the method and scheduling of irrigation. The ponding of water in check basin irrigation provides a positive pressure head resulting in rapid infiltration of water into the soil profile. In structured soils, water may flow through the macropores even before the soil surface is fully saturated (Armstrong et al., 1995) resulting in the preferential flow phenomenon. The high DP observed in the case of furrow irrigation resulted from higher water fluxes due to the confining nature of the furrows, which resulted in higher total hydraulic head forcing the water to infiltrate deeper into the soil profile. The free water maintained in the soil surface could very easily access any surface-vented macropores networks and end up being preferentially transported to greater depths (Clothier and Green, 1994). In contrast to this Onken at al. (1979) reported lesser deep percolation under furrow irrigation due to increased lateral and upward flow of water through the ridges. But in the present experiment the lateral flow was minimum due to closeness of the furrows adopted to accommodate the recommended spacing for planting Okra crop. Because of relatively low intake rate of the soil at the experimental site, sprinkler irrigation also resulted in some ponding though with a lower pressure head than that observed in basin-irrigated plots. Hence, the differences in DP in the sprinkler and basin-irrigated plots were relatively small. The experiment indicated that water losses due to DP can be restricted to the minimum by applying water through sprinkler and scheduling irrigation at 30% MAD. Scheduling irrigation at 30% MAD resulted in maximum crop water use, which was marginally higher under furrow and basin irrigation than sprinkler. Although scheduling irrigation at 60% MAD further reduced DP losses, the crop water use was however markedly reduced.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 52
The data on yield and use efficiencies of water and N of Okra crop clearly revealed that the irrigation schedules that maintained soil moisture near the field capacity significantly enhanced the yield and N uptake. The extreme drying observed in the fields under I4 might have lessened the root activity, and reduced the N uptake by the plant roots (Humphreys et al., 1989). The yield data thus indicate that for the coarse textured lateritic soil, the maximum nitrogen uptake efficiency and fruit yield of Okra can be attained by scheduling irrigation at 30% MAD with sprinkler.
Conclusions On the basis of the results of the study, the following conclusions were drawn. i) On sandy loam lateritic soil planted to Okra crop, furrow irrigation results in the
maximum deep percolation loss while the minimum deep percolation occurs under sprinkler irrigation.
ii) Low volume and high frequency irrigation scheduled at 15% maximum allowable depletion (MAD) of available soil water results in higher deep percolation loss than the low frequency high volume irrigation timed at 60% MAD, particularly under surface irrigation methods.
iii) Maximum use of water and N by Okra crop can be obtained by sprinkler irrigation scheduled at 30% MAD, which appears to be the threshold limit for scheduling irrigation based on MAD irrespective of method of irrigation.
iv) In correspondence to the trends of water and N uses, the maximum fruit yield of Okra is attainable by scheduling irrigation at 30% MAD with sprinkler.
References: Clothier, B.E. and Green, S.R., 1994. Root zone processes and the efficient use of irrigation
water. Agric. Water Manage., 25: 1-12. Chartzoulakis, K. and N. Doros. 1995. Water use and yield of greenhouse grown eggplant
under drip irrigation. Agric. Water Manage., 28: 113-120. Foster, E.F., Pajarito, A. and Acosta-Gallegos, J., 1995. Moisture stress impact on nitrogen
partitioning, nitrogen remobilization and nitrogen use efficiency in beans (Phaseolus vulgaris). J. Environ. Qual., 24: 27-37.
Humphreys, E., Muihead, W.A., Melhuish, F.M., White, R.G., Blackwell, J. and Chalk, P.M., 1989. The growth and nitrogen economy of rice under sprinkler and flood irrigation in South East Australia: III. 15 N balance. Irrig. Sci., 10: 281-292.
Jackson, M.L., 1973. Soil chemical analysis. Indian Reprint, Prentice Hall of India Ltd, New Delhi.
Onken, A.B., Wedt, C.W., Wilkf, O.C., Hargrove, R.S., Bausch, W. and Barnes, L., 1979. Irrigation system effects on applied fertilizer nitrogen movement in soil. Soil Sci. Soc. Am. J., 43: 367-372.
Pereira, L.S. 1999. Higher performance through combined improvements in irrigation methods and scheduling: a discussion. Agric. Water Manage. 40: 153-169.
Tisdale, S.L., Nelson, W.L. and Beaton, J.D., 1990. Soil fertility and fertilizers. 4th Edition. MacMillan Publishing Company. MacMillan Inc.
van Genuchten, M. Th., 1980. A closed form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J., 44: 982-898.
Discussion Q: What type of fertiliser was used I the study A: Urea Q: How did you calculate water use efficiency? - Stuetzel

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 53
A: Water use efficiency was calculated from the amount of N-use and total dry biomass (Fruit + shoot) Q: What was the N application level at the different MADS Treatments? Q: The soil water content in the soil was measured at what soil level? A: N application was uniform for all treatment at 120kg N/ha A: Soil water content was measure at 15 cm intervals to a maximum depth of 100cm. The root zone was 60 cm, Q: What water volumes were used for irrigation in the various MAD irrigation schedules? A: 15% MAD – 20 mm 30% MAD – 40 mm 45% MAD – 60 mm 60% MAD – 80 mm
GENETIC IMPROVEMENT OF VEGETABLE CROPS FOR SUST AINABLE HORTICULTURAL PRODUCTION Stephen G. Agong
Abstract Assessment of the genetic material at the disposal of the farmer, plant breeder, agronomist and geneticist is a primary requirement for a focused varietal improvement strategy. Fundamentally, three components of plant improvement are envisaged namely: yield, quality and adaptability. These components under the generalised theme 'Triology' in plant breeding translate via genetic adjustment into the final phenotype. The genetic structure of a vegetable crop therefore, accounts for about 40% of the expressible phenotypic value of the crop. In many vegetables grown in the tropics, the key questions that arise in terms of the genetic improvement oscillates around the: i. Yield and quality of the edible portion (fruit leaf, root, bulb, pod, seeds, and stem) ii. Adaptability (biotic-pathogens, pests and weeds; abiotic-saline, aluminium toxicity, water stress,
heat/cold) Vegetable provides a majority of the minerals and vitamins of the dietary composition in the tropical region. Therefore, both yield and quality must be addressed in the genetic improvement of the crops. Most vegetables grown in the tropic are exotic with limited adaptability to the tropical regions. Extensive genetic improvement of these crops is therefore necessary before economic utilisation is possible. On the other hand, the traditional vegetable varieties have inherent potential for direct cultivation but with limited genetic potential. Largely, conventional as well as non-conventional approaches have been used in an attempt to improve the vegetables for yield, quality and adaptability. However, non-participatory approaches are destined to fail and do offer no sustainability in horticultural production. Farmers should therefore be directly involved in the choice/selection of variety. Use of chemical for crop protection in horticulture has limited scope and we have to fall back on the resistant crop varieties but to an extent sacrifice on quality upon achievement of economic yield.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 54
PROPAGATION OF KEI APPLE (Dovyalis caffra Warb.); TPRI nursery Experience
Elvania Bujulu and Julius E. Mkenda
Tropical Pesticide Research Institute (TPRI), P.O. Box 3024, Arusha Tanzania, Email: [email protected] , Tel. 255 278813-15
Abstract Kei apple (Dovyalis caffra Warb.) propagation at TPRI started as an idea to produce seedlings for generating funds through sales. The demand of live fencing (hedging) in northern regions of Tanzania is now very high and still increasing. Despite its growing demand, D. caffra.is sparsely known to many of those enjoying its beautiful and efficient fencing shrub. From 1999 to July 200 1 the demand of D. caffra for fencing and as an ornamental has been increasing remarkably. Procedures for developing the seedling at the nursery bed and ultimately the mature hedge are addressed here for further propagation of the plant, not only for the present uses as live fence and fruits, but also for future uses as a source of herbicide Keywords: Kei apple (Dovyalis caffra), propagation, seedlings, live fence
Introduction Kei apple seedling raising project at TPRI: The Kei apple (Dovyalis caffra Warb.) seedling raising at TPRI nursery was started as part of small pioneer production project in 1999. The seedlings are produced for sale as part of income generation. The market is expanding due to the fact that in northern regions of Tanzania there

Wesongtropics
is a higher local demand for live fencing (hedging) especially D, caffra due to its thorny and disease resistance nature, Despite its potential values, the inside information about D. caffra is sparsely known to many of those enjoying it as a live fencing shrub. Procedures for developing the seedling at the nursery bed and ultimately the mature hedge need to be addressed for further propagation of the plant, not only for the present uses as food and fencing requirements, but also for future uses as source of herbicide. Currently- scientists at TPRI are working on the possibility of using Kei apple as a source of herbicide. This paper describes all aspects involved in production of D. caffra from propagation to mature shrubs forming hedges (live fences). General Information about D. Caffra and its Production at TPRI Description Kei Apple, D. caffra (Family Flacourtiaceae ), originates from South Africa around the Eastern Cape, Kaffraria and Natal. It is a thorny evergreen shrub growing to a height of 30ft (9m) with a
spread of 25ft (7.5m). It has many sharp spines l-3in (2.5- 7.5cm), though it is often entirely spineless if not trimmed. The leaves are often clustered on short spurs l-3in (2.5-7.5cm) long, glossy and short petioles. Pale-yellow male and female flowers are usually borne on separate trees. The aromatic fruit is oblate or nearly round, 1-11/2 in (2.5-4cm) diameter, with bright yellow, smooth but minutely downy, somewhat tough skin and mealy, apricot - textured, juicy, highly acidic flesh. There are 5 - 15 seeds arranged in double rings in the center.
Fig 1: T
he Kei apple bush (Dovaylis caffra) with fruits(Morton J, 1987)Fig 2: Kei apple mature shrub with yellow ripe fruits at TPRI, Arusha. The sharp spines can be clearly seen (Photo: Elvania Bujulu, March 2001
a, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 55

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 56
Climate The Kei apple, a drought and wind resistant plant is a subtropical shrub and thrives well at and above 2,600ft (800m) a.s.l. Arusha, Tanzania being in one of subtropical regions favors the growth of Kei apple. TPRI is found in Arusha at 1433 m a.s.l. (030 20'S; 36037E). Soil The Kei apple does well in almost any type of soil that do not have high water table- It is extremely drought resistant and tolerates saline soil and salt spray. It grows well on rich soil with high organic matter content. Propagation Propagation of Kei apple is normally by seeds. Seeds germinate readily when fresh. Seedlings mature and begin to bear fruits in 4 or 5 years. Dried seeds are normally soaked before sowing so as to soften the outer hard fibrous coat. It was proved by the authors that when soaked, the D caffra seeds take less time for the seedling to emerge after sowing. It also minimizes the need for constant watering during the dry season, and the use of labour and irrigation water. The same has been recorded by the Vegetable Research & Development Center 1971 (AVRDC. 1990). Season Kei Apple seedling can be raised throughout the year if water for irrigation is available. Generally the plants bloom in spring and the fruits ripen from August to October. In Arusha, Tanzania fruits were found even to the end of March at TPRI (Fig 2).
SEEDLING RAISING PROCESS AT TPRI Seed preparation
Fruit collection When D. caffra fruits become mature they ripen and normally drop down. Early harvest necessitate picking, otherwise they are to be collected by hand from the ground. At TPRI there are big mature bushes of more than 10 years of age.
These are the ones, which are being used as mother plants in propagation of seedlings in this project. When necessary some fruits are collected from other mature bushes outside TPRI compound.
Fruit fermentation, seed cleaning/washing and drying Collected fruits are put in sisal sacks. They are flushed with some water and left for two days to ferment. They are then squeezed by hand to remove the outer skin in order to release the seeds. They are then washed, cleaned and dried on Hessian cloth in natural sun for about three days. The dried seeds are stored in dry clean containers ready for sowing.
Fig 3. D. caffra fruit collection at TPRI National Plant Quarantine Services Station hedge (Photo: Elvania Bujulu, March 2001

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 57
Fig 5: Fresh D. caffra seeds drying on Hessian cloth (Photo: Elvania Bujulu, March 2001
Fig 4: Hand squeezing the seed from the fermented fruits after two days before washing (Photo: Elvania Bujulu, March 2001

Seedbed preparation Seedbeds, 0.5 x 2m are prepared after ploughing well the land. One bucket (20lt capacity) of farmyard manure (FYM) is added and mixed thoroughly with soil in every seedbed. They are then irrigated and left for at least one day in order to enhance compatibility before sowing.
Sowing Seeds are sown in rows by drilling method. The distance between rows is normally 12cm. After sowing the beds are covered with a mulch to retain moisture and to enhance germination. This mulch is removed immediately when the shoots emerge from the soil. After sowing the seeds, the seedbed is irrigated daily depending on daily weather conditions. This process continues to the time of transferring the seedlings to the pots. This is done when the seedlings have developed the 5th leaf.
Fig 7: Drilling the seeds on seedbeds (Photo: Elvania Bujulu, March 2001
Wt
Fig 6. Dried D. caffra seeds (Photo: Elvania Bujulu, March 2001
esonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the ropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 58

Wt
Potting Polythene tube preparation Rolls of black polythene tubes (diameter 12cm) are cut into tubes of 12cm height. Soil mixture At TPRI, forest soil is mixed with farmyard manure (FYM) at a ratio of 2:3 (FYM: soil). The mixture is sieved by using coffee wire mesh in order to remove large particles and other forest debris before filling it into the tubes (pots). It is then slightly watered to make it wet for easy potting. Seedling transfer from the seedbed to the pots (poly-tubes) When the seedlings have developed the 5th leaf they are transferred from the bed to the
polythene pots containing wet soil mixture explained above. A small hole is made at the center of the soil mass into which the seedling is planted. Watering The potted seedlings are put under the shade and watered every day until when they are taken to the field for transplantation.
F
Fig 9: Filling the polytubes with soil making the holes and transplanting the seedling (Photo: Elvania Bujulu, March 2001
esonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the ropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 59
ig 9: Potted seedlings arranged under the shade (Photo: Elvania Bujulu, March 2001

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 60
USES OF KEI APPLE Hedging Kei apple is widely used as an effective fruiting hedge that is almost goat and some other animals proof. The thorny hedge is ideal in discouraging thieving (Epenhuijsen, 1976). As already stated, in Tanzania the plant is widely planted in the northern regions of Arusha and Kilimanjaro as a live fence. Hedge plants are set at 0.33m apart and are trimmed twice a year. The trimming height depends on individual's decision. The hedge is planted around residential houses, animal sheds, and farm boundaries and as a decoration on various compounds. Food Food Value The fruit is too acidic for eating out-of-hand even when fully ripe. It is best cut in half. peeled, seeded, sprinkled with sugar and allowed to stand for a few hours before serving as dessert or in fruit salads. The halves can stand only a few minutes of cooking before they turn into sauce. Simmered briefly into syrup, they make excellent shortcakes. Kei apple are customarily made into jam and jelly, and when semi ripe into pickles (Morton, 1987). It is reported that fresh ripe fruits contain 83 mg ascorbic acid per l00 g and 3.7% pectin. Scientists in Egypt have reported 15 amino acids: alanine, 0.41 %; arginine, 0.36%; aspartic acid, 0.96%; glutamic acid, 2.00%; glycine, 0.39%; histidine, 0.10%; isoleucine, 0.25%; leucine, 0.75%; lysine, 0.36%; methionine + valine, 0.28%; phenylalanine, 0.40%; proline. trace; serine, 0.48%; threonine, 0.34% (Nerd & Aronson, 1994). Morton, (1987) reports further that in some other growing areas the fruit contains up to 117mg ascorbic acid (vitamin C) per 100 gm of fruit, (baobab has around 213 mg/100gm). As each Kei apple fruit weighs about 15 grams, each small (about walnut size) fruit provides around 17 grams of vitamin C - four times more than an apricot fruit, a fruit twice the Kei apples' weight! The recommended adult daily allowance' of vitamin C is 60 mg, and would be supplied by only 4 of these small fruits. It is more likely that gatherers would have eaten at least double this number every day in the fruiting season. Herbicide Kei apple exhibits allelopathy properties. This means that its roots excrete growth inhibitors, which prevent the occurrence of other plants in its vicinity. Investigators in Egypt have demonstrated that the roots. stem and fruit, but not the leaves and branches, possess antibiotic properties (Morton, 1987).
KEI APPLE SEEDLING PRODUCTION AT TPRI AS A PIONEER PROJECT As stated before, Kei apple seedlings project is one of the production segments ran by Farm Management Section among others at TPRI. Since the beginning of Kei apple production at TPRI, 28,000 seedlings were produced (Fig. 10) between January 1999 and July 2001. It is expected that during the year 2001 at least 20,000 D. caffra seedlings will be produced and sold. Since the demand of D. caffra seedlings for live fencing is increasing in Tanzania especially in Arusha and Kilimanjaro regions it is obvious that there is a big potential and a need to inform the community more about the plant specifically on its benefits. Figure 10 indicates a remarkable increase of production, which is a result of growing demand.
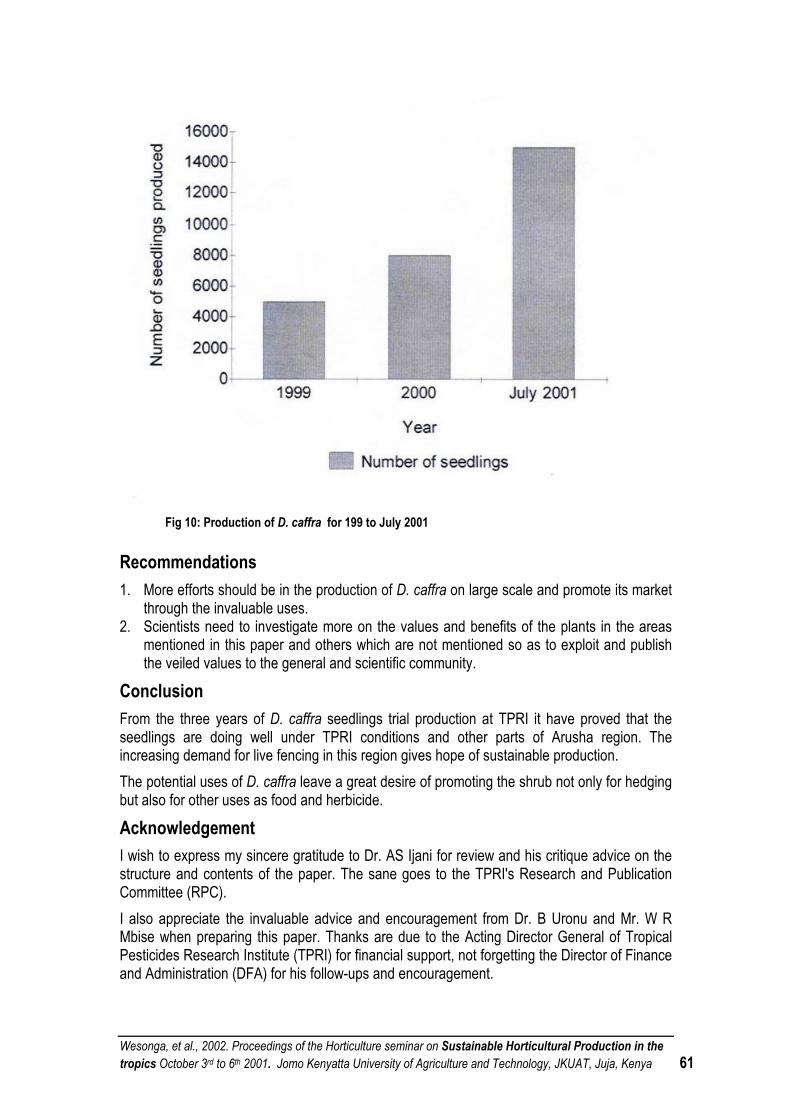
Wesonga, et al., 2tropics October 3
Recommend1. More effor
through the2. Scientists
mentionedthe veiled
Conclusion From the threeseedlings are increasing demThe potential ubut also for oth
AcknowledgI wish to exprestructure and cCommittee (RPI also appreciaMbise when pPesticides Resand Administra
Fig 10:
Production of D. caffra for 199 to July 2001002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 61
ations ts should be in the production of D. caffra on large scale and promote its market invaluable uses.
need to investigate more on the values and benefits of the plants in the areas in this paper and others which are not mentioned so as to exploit and publish values to the general and scientific community.
years of D. caffra seedlings trial production at TPRI it have proved that the doing well under TPRI conditions and other parts of Arusha region. The and for live fencing in this region gives hope of sustainable production. ses of D. caffra leave a great desire of promoting the shrub not only for hedging er uses as food and herbicide.
ement ss my sincere gratitude to Dr. AS Ijani for review and his critique advice on the ontents of the paper. The sane goes to the TPRI's Research and Publication C). te the invaluable advice and encouragement from Dr. B Uronu and Mr. W R
reparing this paper. Thanks are due to the Acting Director General of Tropical earch Institute (TPRI) for financial support, not forgetting the Director of Finance tion (DFA) for his follow-ups and encouragement.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 62
Finally, my heartfelt gratitude to my husband Dr. Joel Bujulu for his lovely encouragement and moral support.
References Epenhuijsen C. W. van, 1976). Deciduous fruits in Tanzania. Ministry of Foreign affairs.
International Technical Assistance Department; The Hague, Netherlands. Mortan, J. (1987). Kei Apple p. 315 -319. In: Fruits of warm climates. Julia F Mortan, Miami, FL. Nerd, A., J. A. Aronson (1994). Introduction and domestication of rare and wild fruit and nut
trees for desert areas. WANATCA Yearbook 18: 42-53. Dep. Life Sci., Ben-Gurion Univ. Negev, PO Box 1025, Beer-Sheva 84110, Israel
Discussion Q: What is the attitude of people towards from in Tanzania? A: The people respond positively Q. Are seeds planted directly from seed? A: Yes Q: For how long can the seed be stored after extraction A: Not known Q: What is the possibility of planting seed straight after extracting them from the fruit without processing (e.g. without fermenting them)? A: This is possible Q: How long may the seeds be remain viable in during storage and is there a special way of storing the seeds for say one more year (A.O. Auma) A: This is not yet known
SELECTION OF MACADAMIA ROOTSTOCKS LEAST SUSCEPTIBLE TO CHLOROSIS AND NECROSIS S. Ngugi1, J. Mbakal, L. Gitongal, Wepukhulul and L.S. Wamocho2 1 Research Scientists, National Horticultural Research Center Thika 2 Senior Lecturer, Jomo Kenyatta University of Agriculture and Technology
Abstract Chlorosis is one of the major factors limiting the production of healthy macadamia seedlings affecting about 40% rootstocks annually. Seedlings (4-6 hard leaves) of six macadamia varieties; KMB-3, MRG-25, KRG-3, KRG-4, KRG-15 and EMB-1 were transplanted into polybags containing 2 kgs of media consisting of soil: manure: sand (10:3:1). Except for KMB-3 which is a hybrid (integrifolia * tetraphylla) the rest of the varieties are integrifolia. The varieties were screened for susceptibility to chlorosis and necrosis. From the results, KMB-3 variety had the best germination % (44 %) and the highest growth vigour. The varieties least susceptible to chlorosis were KMB-3 and EMB-1 while KRG-3, 4, 15 was the most susceptible. The highest level of necrosis was observed in KRG-3 with the rest showing moderate levels. This study showed that KMB-3 is a good source of rootstock material with least susceptibility to chlorosis and necrosis.
Introduction Macadamia is the most important nut in Kenya. It is grown by over 200,000 small scale farmers. The production is concentrated in central, Eastern and Rift Valley provinces with great Potential in Western and Nyanza Provinces (MALD, 1997).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 63
Shortage of grafted planting materials continue to be a constraint in macadamia production (MALD, 1997). The existing nurseries in the country supply about 100,000 seedlings against an estimated annual demand of 500,000 seedlings. Chlorosis is one of the major factors limiting the production of healthy macadamia seedlings. About 40% (of the rootstocks are affected annually. Once chlorosis has developed. It is extremely difficult to correct, even though the initial cause is removed. In most cases, chlorosis in macadamia is inherited and it is not due to lack of iron in the soil (Westree et al. 1974). A major way of alleviating the problem is by use of rootstocks resistant to chlorosis. This study was set up to investigate the susceptibility to chlorosis of six macadamia varieties; KMB-3, KRG-3, KRG-4, KRG-15, MRG-25 AND EMB-1 as rootstocks.
Materials and methods The study was carried out in the nursery at the Horticultural Development Project, Thika. The seed nuts of. six macadamia varieties; KMB-3, KRG-3, KRG-4, KRG-15, MRG-25, and EMB-1 were obtained from the mother trees. The seeds were planted on 8th July, 1999 in sand beds and watered regularly when the seedlings attained 4-6 hard leaves they were transplanted into po1lybags Containing 2 kgs of transplant media. The media composition was topsoil, farmyard manure and sand in the ratio of 10:3: 1 (v/v). A compound fertilizer (17:17:17) was added to the mixture at the rate of300g for every 10 wheelbarrows of soil and mixed thoroughly. At transplanting, 20 seedlings of each variety with four replications were used resulting in 80 seedlings per variety. They were arranged on drain beds in a completely randomised design. The seedlings were kept under light shade (40-50 % light reduction) for two months before being transferred to the open area. Watering of the seedlings was done on a regular basis. In the open al-ea the seedlings were sprayed with benlate and folimat for the control of pests only when necessary. The seedlings were topdressed once with Calcium Ammonium Nitrate (CAN) in the fOU11h month after transplanting. The following data was collected in the course of the experiment: 1. Germination % 2. Degree of chlorosis On a scale 1-4 as follows:
Non chlorotic Slightly chlorotic (only new shoot) Medium chlorotic (upper half yellow (3-5 leaves)) Chlorotic - entire seedling yellow
3. Degree of necrosis. On a scale 1 -4 as follows:
Non necrotic 2.Slightly necrotic (only new shoot) 3.Medium necrotic (upper half yellow. (3-5 leaves)) 4.Necrotic - entire seedling yellow
Results and discussion The results of the germination % are shown on table 1. As can be observed from the table KMB-3 had the highest germination % with KRG-15 having the lowest. It was not clear why this was so given that the mother tree of KRG-15 was one of the best among the varieties.
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 64
Table 1 Germination % of macadamia seedlings Variety Germination %
Variety Germination %
KME-3 44 KRG-3 37 KRG-4 6.8
KRG-15 4 MRG-25 35 EMB-1 11
The expression of chlorosis in the macadamia varieties is shown on Table 2. As can be observed from the table the six varieties were susceptible to chlorosis to different degrees. For the first five months after transplanting only KRG-15 variety expressed very slight chlorosis. For the next three months KRG-3, KRG-4 and KRG-15 had the highest levels of chlorosis which were significantly different from the other varieties. At the time of grafting however, KRG-3 and KRG-4 had the highest expression that was significantly different from the varieties that had the least. The results of the degree of necrosis are recorded on Table 3. From the table it can be observed that KMB-3 and KRG-15 had significantly lower levels of necrosis over time as compared with the other varieties;. The highest expression was observed in KRG-3 and MRG-25 in the 7th month after transplanting. From the preliminary results KMB-3 a hybrid appears promising as a source of rootstock material as it had the highest germination %, and was least susceptible to both chlorosis and necrosis. Table 2:The expression of chlorosis in macadamia seedlings
Variety Month 3
Month 4
Month 5
Month 6
Month 7
Month 8
Month 9
KMB-3 1.01B 1.00C 1.11A 1.35BA 1.39B 1.31B 1.20C KRG-3 1.0lB 1.00B 1.03C 1.31BAC l.54A I 1.58A 1.58A KRG-4 1.00B 1.00B 1.05BC 1.38BA 1.63A 1.53A 1.56A KRG-15 1.04A 1.08A 1.08BA 1.4A 1.63A 1.58A 1.31BC MRG-25 1.00B 1.00B 1.03C 1.28BC 1.55A 1.48A 1.39B EMB-1 1.01B 1.00B 1.01C 1.21C 1.35B 1.33B 1.21C The data collection started the 3rd month after transplanting . Figures having the same letter down the column are not significantly different at alpha = 0.5 according to Sceffe’ s mean separation test Table 3:The expression of necrosis in macadamia seedlings
Variety Month 3
Month 4
Month 5
Month 6
Month 7
Month 8
Month 9
KMB-3 1.10A 1.08BC 1.09C 1.30D 1.74C 1.40C 1.44C KRG-3 1.08BA 1.16A 1.31A 1.90A 2.19A 1.88B 1.98A KRG-4 1.03C 1.01DC 1.19B 1.4875C 1.85BC 1.84B 1.94BA KRG-15 1.04BC 1.10BA 1.16CB 1.2125D 1.36D 1.39C 1.41C MRG-25 1.00C 1.00D 1.15BC 1.7375B 2.08A 2.00A 1.94BA EMB-1 1.09A 1.IIBA 1.29A 1.75B 2.04BA 1.90BA 1.85B The data collection started the 3rd month after transplanting Figures having the same letter down the column are not significantly different at alpha = 0.5 according to Sceffe's mean separation test
Areas for further study Establish the cause of the necrosis and chlorosis in the macadamia seedlings. Investigate possibilities of increasing the germination % of the macadamia seed nuts.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 65
References Ministry of Agriculture, Livestock Development and Marketing. Horticultural Division, Annual
Report, pp. 3, 1997. Westree, N.E; A.V. Snider; L. Johnson, L.E. James. 1974 Answers to 25 commonly asked
questions on macadamia. Calif. Macadamia Soc. Year book 20:56-62.
Acknowlegements This project has been made possible by funds from Agricultural Research Fund (ARF)
Discussion Q. What is the cause of chlorosis and necrosis Macadamia seedlings? M.E. Omunyu A. Still unknown and investigations are going on Could you also examine the root mass for various varieties; this may be linked to changes in chlorotic status etc How about nutrient status of seedlings i.e. N, Fe, etc? How safe is it to narrow down from six to one variety based on ‘relative’ low incidence? A. We have started with six; we are adding more; this is only a beginning
SNAP BEAN IMPROVEMENT IN KENYA Odindo, A.O. and Rheenen, H.A.van
Abstract Seeds of several locally produced snap bean varieties are available in Kenya for the production of green young pods. For instance, various seed companies market the variety Monel. However, seed of most of the cultivars presently grown for export is imported and the imported seed is expensive. A sample of such imported seed when grown in Eldoret, appeared to be susceptible to anthracnose and rust and is suspected to be susceptible to bean common mosaic virus. The plants seemed to have good yield potential, but were short in stature with little pod clearance from the ground. This seemed to suggest that a variety improvement programme might be useful. The objectives being to develop disease resistant varieties of high yield potential and good pod quality and to stimulate local seed production. Germplasm was collected from the National Horticultural Research Centre, Thika, and the National Gene Bank of Kenya, Muguga. Parents were selected and crosses were made in 1998. The breeding programme has reached the F4 progeny testing stage, with progenies from four populations. Preliminary observations of the F4 progeny show clear differences in vigour between spreader rows, the control and progeny rows. Single plant selections will continue in the F4 generation, and the intention is to bulk the best progenies for yield, resistance and quality testing.
Introduction The export of green pods of snap beans is important and snap bean production by smallholder farmers is remunerative. KARI (1993) observes that farmers who grow snap beans for export may experience problems of non-availability of good quality seed when required Seeds of several locally produced snap bean varieties are available in Kenya for the production of green young pods. For example, various seed companies market Monel, which is an old variety. More recently, other varieties such as Amy and Julia, are being imported into the country. This seed is very expensive. For example, 1kg of such an imported variety purchased in Eldoret cost Ksh 890 (about US$ 11.40) per Kg. When planted in the field, the variety appeared to be susceptible to anthracnose and rust, and is suspected to be susceptible to the bean common mosaic virus (BCMV). The plants seemed to have good yield potential, but were short in stature with little pod clearance from the ground.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 66
These observations suggest that a snap bean variety development programme in Kenya may be useful. The objectives would be to develop disease resistant varieties of high yield potential and good pod quality. The programme could also stimulate local production of good quality snap bean seeds.
Materials and Methods Choice of parents Germplasm was collected from the National Gene Bank of Kenya, Muguga and the National Horticultural Research Station, Thika in 1997. The parents consisted of CPST-B8, which has resistance to anthracnose and bean common mosaic virus and has field resistance to rust. CPST-B40, which is resistant to halo blight. Amy (CPST-B34) and Samantha (CPST-B39), french bean varieties from Regina Seeds, selected for their high yield potential. Method of breeding Initial crossing was done in 1998, between CPST-B34 x CPST-B8 and CPST-B39 x CPST-B40 in pots (figure 1). The F1 of (CPST-B34 x CPST-B8) was crossed with CPSTB-40 to a three-way cross. Both single crosses (CPST-B34 x CPST-B8) and (CPST-B39 x CPST-B40) were crossed to form a four-way cross. These crosses were all planted in pots to produce F1 generation, which were then advanced to the F2. The two single crosses were planted in plots in the field, while 150 plants each of the three-way cross and the four-way cross were planted in pots. Single plant selection started in the F3 generation. The four crosses were sown in the field in plots with strips of spreader row plants at regular intervals. CPST-B34 and CPST-B40 were used for sowing the spreader rows. Selection was based on desired pod shape, colour and plant health. Diseased plants especially those with rust and BCMV were selected to be used later in spreader rows. The F4 progenies were planted in 6mx6m blocks of plot of 20 rows. Each row was 2m. Spacing between the rows was 50cm and within the rows 20cm. Disease spreader rows of CPST-B40 were sown around and within the trial. Four rows of CPST-B34 were included in each block to serve as control.
Results and Discussion 1012 and 157 single plants of (CPST-B34 x CPST-B8) and (CPST-B39 x CPST-B40) were selected respectively from the F3 generation. For the three-way cross, (CPST-B34 x CPST-B8) x CPST-B40 and the four-way cross (CPST-B34 x CPST-B8) X (CPST-B39 x CPST-B40), 108 and 136 single plants were selected. Preliminary observations of the F4 progeny show clear differences in vigour between spreader rows (CPST-B40) the control (CPST-34) and progeny rows. Single plant selections will continue in the F4 generation, and the intention is to bulk the best progenies for yield, resistance and quality testing.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 67
SNAP BEAN IMPROVEMENT IN KENYA Breeding scheme for the development of a new snap bean variety
CPST-B34 X CPST-B8 CPST-39 X CPST-B40
(CPST-B34 X CPST-B8) X CPST-B40 (CPST-B34X CPST-B8) X (CPST-B39X CPST-B40)
F1 (Pots)
F2 (Plots, Pots)
F3(Single plant selection)
F4 (Progeny testing)
References Allard RW. 1960. Principles of Plant Breeding. New York and London: John Wiley and Sons
Inc. Kenya Agricultural Research Institute. 1993. Annual Report. Nairobi, Kenya. KARI
Discussion Q. Have you involved the private sector (seed merchants and exporter) in developing the new varieties? A. Up to this level not yet, but we plan to approach them before we release the new variety Comment. It is necessary to involve the farmers and companies dealing with snap beans at the onset to ensure successful uptake of the product at the end of the study. M.E. Omunyu

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 68
EFFECT OF EXPLANT TYPE ON GUS GENE EXPRESSION AND SHOOT REGENERATION FROM BRUSSELS SPROUTS AFTER EXPOSURE TO AGROBACTERIUM TUMEFACIENS. Lucy Gitonga1 and Graham Collins2
1Research scientist, Kenya Agricultural Research Institute, National Horticultural Research Center, Kenya 2Senior Lecturer, Adelaide University, Dept of Horticulture, Viticulture and Oenology, Australia
Abstract Brussels sprouts (Brassica oleracea var. gemmifera) is a highly nutritious vegetable containing high levels of vitamins B1, B2, C, minerals and folic acid. It also contains sulfur-containing compounds - glucosinolates, that have been associated with protection against some forms of cancer and also protective mechanisms against some crop pests and diseases. However, consumers have implicated these compounds with the characteristic bitter taste that results in a low preference score for Brussels sprouts. Conventional selection breeding may result in milder tasting cultivars but may also result in lower levels
of these beneficial compounds. The gene for thaumatin, an intensely sweet protein if transformed into Brussels sprouts, may have the potential to mask the bitter taste and hence improve consumption of this highly nutritious vegetable. This therefore calls for development of an efficient gene transfer system. As part of this study, different types of explants namely stem (hypocotyl) sections, leaf petioles and leaf sections were evaluated on their response to in vitro exposure to Agrobacterium tumefaciens already transformed with the reporter gene GUS (ββββ-glucuronidase). Explants were inoculated with the bacteria for 10 min and cultured on MS media for 1 wk at 22°°°°C under a 16 hrs light/8 hrs dark photoperiod (40µµµµmols.m-2.s-1). GUS gene expression was histochemically scored as presence and number of blue spots, while shoot regeneration frequency was scored after 4 wk. Leaf sections had significantly higher GUS expression (44.8%) than hypocotyl sections (6.6%) and leaf petioles (0.0%) at 5% level. However, petioles had significantly higher shoot regeneration (80.0%) than leaf sections (45.8%) but not significantly higher than hypocotyl sections (65.8%) at the 5 % level. This study showed that Agrobacterium tumefaciens -mediated transformation could be used to transform the thaumatin gene into Brussels sprouts. However, explants that expressed highest GUS had lowest shoot regeneration and vise versa indicating a need for further research into methods that will tend to combine transformation competence and totipotency of plant cells in order to recover transformed shoots. This study also opens an avenue for similar research into improving other vegetables with antinutritional but otherwise useful compounds.
Introduction Various types of explants have been used in the transformation of Brassica species. Radke et al. (1992) used hypocotyl explants of B. rapa to develop a routine transformation procedure with A. tumefaciens, although regeneration could be obtained from either cotyledon, leaf or root segments. Hypocotyl explants have been used in the transformation of B. juncea (Barfield and

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 69
Pua, 1991), B. rapa (Takasaki et al., 1997) and B. campestris (Xiang et al., 2000). In the transformation of B. napus, hypocotyls (Radke et al., 1988; De Block et al., 1989) as well as stem internodes (Fry et al., 1987), and cotyledons (Moloney et al., 1989) have been used as explants in Agrobacterium-mediated transformation procedures. Within B. oleraceae a wide range of explants have been used for transformation and regeneration of transgenic plants. For Brussels sprouts, leaf petioles (Hamada et al., 1989; Hosoki and Kigo, 1994) were used for transformation with A. rhizogenes. Recently, hypocotyl explants (Gu, 1998) were used in a study of transformation with A. tumefaciens, but with low success. It is hence important to establish the type of explant that would produce the highest transformation and regeneration capacity for efficient transformation of Brussels sprouts.
Objectives 1. 1. To test the frequency of GUS reporter gene expression in explants of hypocotyl, leaf
sections and leaf petioles of Brussels sprouts exposed to A. tumefaciens 2. 2. To investigate shoot regeneration capacities from the three types of explants.
Materials and Methods Plant material Two commercial cultivars of Brussels sprouts, Troika and Winter Pick, were tested in this experiment. Troika is high yielding with good disease resistance while Winter Pick is an early maturing hybrid that produces firm green heads that retain color and flavor for a long time after maturity. Seeds of both cultivars were surface sterilized in 70% ethanol for 2 min and rinsed twice with sterile MQ water. They were then washed for 30 min on an orbital shaker in 25% Milton bleach (providing 0.25% available chlorine) with 1-2 drops of Tween 20 and then thoroughly rinsed 5 times with sterile MQ water. Excess water was removed by blotting the seeds on sterile filter paper and the seeds were then aseptically germinated for 7 d on MS (Murashige & Skoog, 1962) containing half rate MS macro salts, 30 g/L sucrose and solidified with 4g/L phytagel (Sigma Chemical Co., St. Louis). Explant preparation Hypocotyl sections (0.5-0.7 cm) were aseptically excised starting at 0.5 cm from each end. Leaf petiole sections (0.5-1.0 cm) were excised leaving about 0.2 cm from both the distal and the proximal end. Leaf sections (discs) were aseptically excised leaving about 0.5 cm from both the distal and proximal ends. After removing the leaf edges, sections 0.4 cm wide and 1.0 cm long were cut across the mid rib. The target sites for transformation by the Agrobacteria were the cut surfaces. Culture media All media used in this experiment was based on Murashige and Skoog (1962). Stock solutions were prepared using MQ water and stored at 4°C. Sucrose at 30 g/l was used. After addition of all MS components and growth regulators, pH was adjusted to 5.7. Medium was sterilized by autoclaving at a steam pressure of 103.4 kPa (15 p.s.i.) at 121°C for 20 min. Silver nitrate and antibiotics were added after autoclaving. Phytagel (Sigma Chemicals Co. St. Louis) was used for gelling. Agrobacterium strain and plasmid Agrobacterium tumefaciens strain LBA 4404 (Hoekima et al., 1983) was used for the development of the transformation procedure that could later be employed for transformation

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 70
with the thaumatin gene. This strain carries a disarmed binary vector containing a p35SGUS intron. The plasmid contains the NPT11 and the GUS genes in the T-DNA region. The NPT11 gene encodes for neomycin phosphotransferase for kanamycin resistance and hence is used as a marker gene to select transformed tissues growing on medium supplemented with kanamycin antibiotic (Draper and Scott, 1991). The GUS gene encodes for β-glucuronidase. This enzyme breaks down a number of β-glucuronides to compounds that turn blue after oxidation (Jefferson, 1987) and hence acts as a good reporter gene allowing successful gene transfer to be detected in the receiver cells at a very early stage. Transformation procedure Explants were aseptically inoculated with A. tumefaciens strain LBA 4404 (OD600 = 0.300) for 10 min and excess bacteria were removed by blotting on sterile filter paper. Co-cultivation of explants together with the Agrobacterium was carried out for 2 d in continuous darkness at 21±1°C on sterile filter paper saturated with MS medium supplemented with 0.1 mg/L 2,4-D and 2.0 mg/L BAP in 90 x 14 mm petri dishes sealed with two layers of parafilm. The explants were then washed 4 times for 5 min each with MS medium supplemented with 150 mg/L cefotaxime and, after blotting on sterile filter paper, cultured on MS medium supplemented with 2 mg/L BAP, 5 mg/L AgNO3, 150 mg/L cefotaxime and solidified with 7 g/L phytagel for 7 d at 24±1°C under a 16 hr light/8 hr dark (40 µmol.m-2.s-1) photoperiod. Some explants of each type were randomly selected and tested for GUS expression, and the remaining explants were cultured on similar medium supplemented with 50 mg/L kanamycin. After a further 7 d, explants were again tested for GUS expression. Histochemical GUS assay Samples of explants were dipped in 300-500L X-Gluc solution (GUS substrate reaction buffer) (Progen Industries Ltd.) and incubated overnight at 37°C. Chlorophyll was removed by replacing the X-Gluc solution with an equal volume of 70% ethanol and soaking for several hours. Samples were then observed under a dissecting microscope for the presence of blue spots. Statistical analysis Experiments were laid out in a completely randomized design (CRD). Each treatment was replicated 5-7 times for leaf and hypocotyl explants with 12-21 explants per plate and 2-3 times for petiole explants with 11-13 explants per plate. Analysis of variance (ANOVA) was done using GENSTAT 5 (Release 4.1) with a significance of difference levels of P<0.05 and separation of means using the least significant difference (L.S.D).
Results GUS expression Blue spots indicating GUS expression, were found on callus on the cut ends of hypocotyls. No blue spots were observed on the epidermis along the entire length of the hypocotyl sections, while for the leaf sections, blue spots were found both on the cut edges and also interspersed within the leaf tissue and sometimes on both sides of the leaf section. No GUS expression was observed in petiole sections. GUS expression in hypocotyl and leaf explants of cultivar Winter Pick is shown in Fig 1 (a) and (b) respectively.
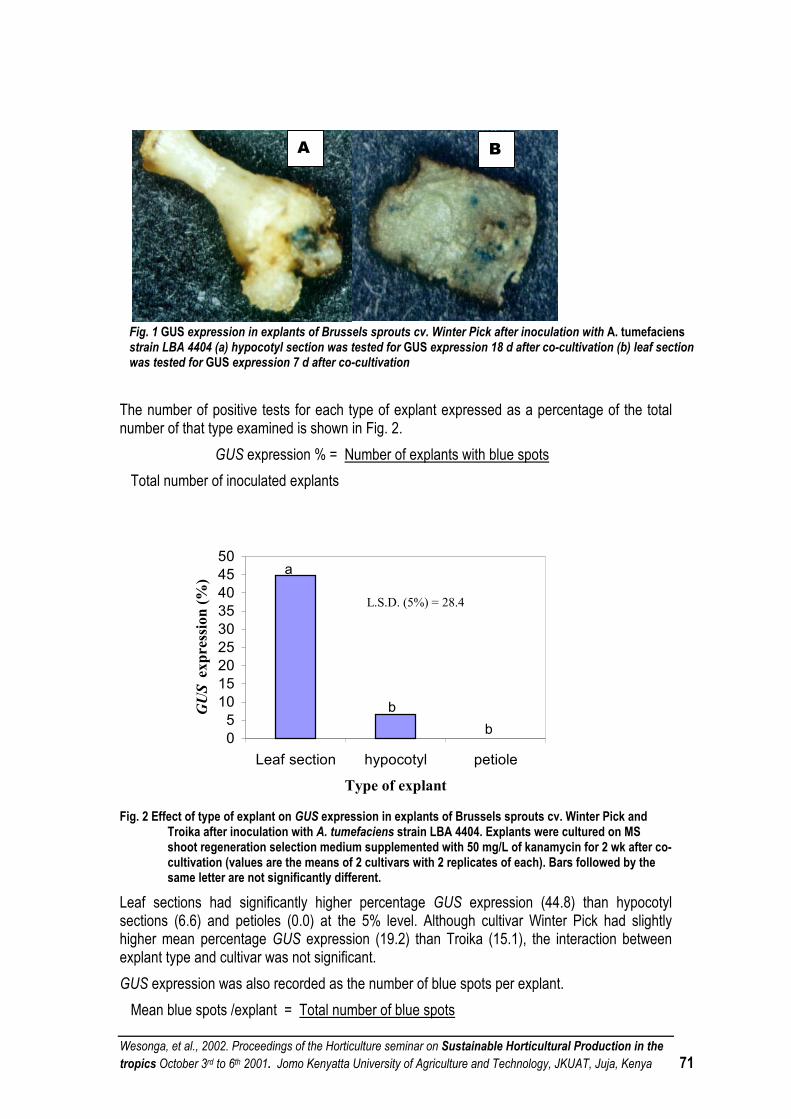
Wt
Tn
F
LsheG
B A
Fig. 1 GUS expression in explants of Brussels sprouts cv. Winter Pick after inoculation with A. tumefaciens strain LBA 4404 (a) hypocotyl section was tested for GUS expression 18 d after co-cultivation (b) leaf section was tested for GUS expression 7 d after co-cultivation
esonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the ropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 71
he number of positive tests for each type of explant expressed as a percentage of the total umber of that type examined is shown in Fig. 2.
GUS expression % = Number of explants with blue spots Total number of inoculated explants
ig. 2 Effect of type of explant on GUS expression in explants of Brussels sprouts cv. Winter Pick and Troika after inoculation with A. tumefaciens strain LBA 4404. Explants were cultured on MS shoot regeneration selection medium supplemented with 50 mg/L of kanamycin for 2 wk after co-cultivation (values are the means of 2 cultivars with 2 replicates of each). Bars followed by the same letter are not significantly different.
eaf sections had significantly higher percentage GUS expression (44.8) than hypocotyl ections (6.6) and petioles (0.0) at the 5% level. Although cultivar Winter Pick had slightly igher mean percentage GUS expression (19.2) than Troika (15.1), the interaction between xplant type and cultivar was not significant. US expression was also recorded as the number of blue spots per explant.
Mean blue spots /explant = Total number of blue spots
05
101520253035404550
Leaf section hypocotyl petiole
Type of explant
GU
S e
xpre
ssio
n (%
)
a
bb
L.S.D. (5%) 28
L.S.D. (5%) = 28.4
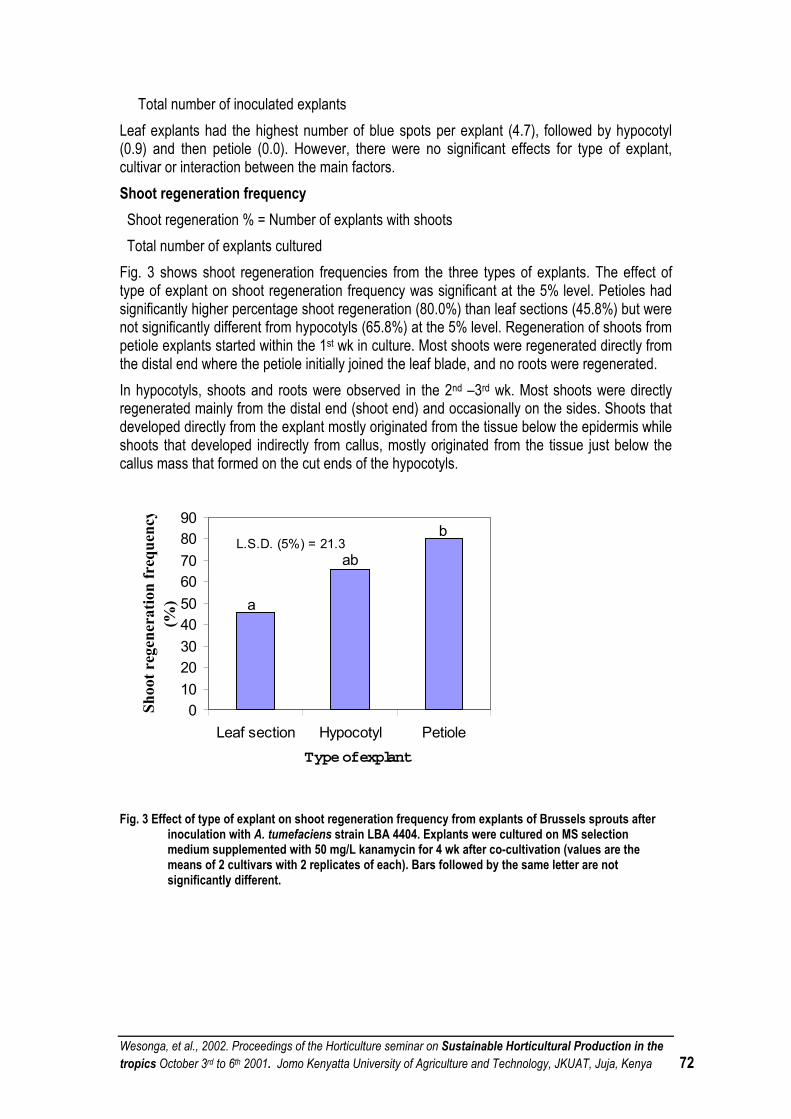
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 72
Total number of inoculated explants Leaf explants had the highest number of blue spots per explant (4.7), followed by hypocotyl (0.9) and then petiole (0.0). However, there were no significant effects for type of explant, cultivar or interaction between the main factors. Shoot regeneration frequency Shoot regeneration % = Number of explants with shoots Total number of explants cultured Fig. 3 shows shoot regeneration frequencies from the three types of explants. The effect of type of explant on shoot regeneration frequency was significant at the 5% level. Petioles had significantly higher percentage shoot regeneration (80.0%) than leaf sections (45.8%) but were not significantly different from hypocotyls (65.8%) at the 5% level. Regeneration of shoots from petiole explants started within the 1st wk in culture. Most shoots were regenerated directly from the distal end where the petiole initially joined the leaf blade, and no roots were regenerated. In hypocotyls, shoots and roots were observed in the 2nd –3rd wk. Most shoots were directly regenerated mainly from the distal end (shoot end) and occasionally on the sides. Shoots that developed directly from the explant mostly originated from the tissue below the epidermis while shoots that developed indirectly from callus, mostly originated from the tissue just below the callus mass that formed on the cut ends of the hypocotyls.
Fig. 3 Effect of type of explant on shoot regeneration frequency from explants of Brussels sprouts after
inoculation with A. tumefaciens strain LBA 4404. Explants were cultured on MS selection medium supplemented with 50 mg/L kanamycin for 4 wk after co-cultivation (values are the means of 2 cultivars with 2 replicates of each). Bars followed by the same letter are not significantly different.
0102030405060708090
Leaf section Hypocotyl PetioleType of explant
Shoo
t reg
ener
atio
n fr
eque
ncy
(%)
L.S.D. (5%) = 21.3
a
ab
b

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 73
Leaf sections produced callus and roots within the 1st wk but shoots developed in the 3rd-4th wk. Most shoots were regenerated indirectly from callus with occasional direct shoots. Shoots that were regenerated directly from the explant were mostly dark green while those that regenerated indirectly from callus were mostly pale green. Shoots first appeared from the callus around the mid rib and other major veins before regenerating from callus at the cut edges (Fig. 4).
Fig. 4 Direct and indirect shoot regeneration from the mid rib of leaf explant of Brussels sprouts cv. Winter Pick after inoculation with A. tumefaciens strain LBA 4404. Explants were cultured on MS selection medium supplemented with 50 mg/L kanamycin for 5 wk after co-cultivation (a) shoot regenerating indirectly from callus (b) shoot regenerating directly from the explant.
Discussion In the present study, leaf sections expressed the highest GUS activity (44.8%) followed by hypocotyl (6.6%) and then leaf petioles (0.0%), indicating that they probably secreted higher levels of asetosyringone, attracting more Agrobacterium and hence more transformation events. Chabaund et al. (1988) also obtained higher tranformation effiency of 36% in leaf explants compared to 9% in petioles in the transformation of alfalfa with A. tumefaciens. They also reported that leaf tissue was more amenable to transformation than hypocotyl and petiole tissues perhaps due to the fact that wounding was easier on leaves than it was in other explant types. Highest shoot regeneration was obtained from petiole sections (80.0%) followed by hypocotyl sections (65.8) and then leaf section (45.8%). Radke et al. (1992) also reported differences in regeneration capacities of different explant types of B. rapa inoculated with A. tumefaciens strain EHA 101. They obtained up to 36% from petiole cross-sections, 15% from cotyledon and leaf explants and 1% from root segments. These findings and the results of the present study indicate that different types of explants have different regeneration capacities and they vary with different plant genotypes. High shoot regeneration frequency increases chances of recovery of transformed shoots. However, the results of this study showed that leaf petioles had highest shoot regeneration but lowest GUS expression while leaf sections had lowest shoot regeneration but highest GUS expression, posing a problem of combining high shoot regeneration with high GUS expression and hence low transformation frequency. Draper and Scott (1991) indicated that in practice, it often seems difficult to combine competence to transformation and totipotence of the plant cell. Mode of shoot regeneration From leaf sections shoots were mostly indirectly regenerated through callus that developed on cut ends but most blue spots were observed within leaf tissue. For hypocotyls, shoots were directly regenerated from distal ends without intervening callus but blue spots were observed at

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 74
proximal end, while for petioles, shoots were directly regenerated from the distal end and no blue spots were obtained. It is possible that in the present study, the meristematic cells competent to regenerate were situated away from the cut surface and were either not accessible to the Agrobacterium or were not competent physiologically to be transformed. Conclusions and future research Brussels sprouts are amenable to transformation with A. tumefaciens strain LBA 4404. Leaf sections are better explants than petioles and hypocotyls. High GUS expression can be obtained, they are easy to manipulate and the number of explants obtained per plant is high. However, shoot regeneration from transformed tissues is still low.It is suggested that in in vitro techniques of transformation with A. tumefaciens, shoot regeneration from transformed tissues of leaf sections of Brussels sprouts be improved before transformation with the thaumatin gene is done to improve the bitter taste. Other in planta techniques such as the floral dip and vacuum infiltration methods should also be tried for Brussels sprouts. These methods have already been found to be successful for Arabidopsis thaliana (Mengiste et al., 1997).
Acknowlegdements 1. Government of Australia through AUSAID for provision of full scholarship 2. Government of Kenya through KARI for provision of study leave 3. Family and friends for their support and encouragement
References Barfield, D.G., Pua, E-C. (1991) Gene transfer in plans of Brassica juncea using Agrobacterium tumefaciens-mediated transformation. Plant Cell Reports 10: 308-314
Chabaund, M., Passiatore, J.E., Cannon, F. and Buchanan-Wollaston, V. (1988) Parameters affecting the frequency of kanamycin resistant alfalfa obtained by Agrobacterium tumefaciens -mediated transformation. Plant Cell Reports 7: 512-516
De Block, M., De Brouwer, D., Tenning, P. (1989) Transformation of B. napus and B. oleraceae using Agrobacterium tumefaciens and the expression of bar and neo genes in transgenic plants. Plant Physiology 91: 694-701
Draper, J. & Scott, R. (1991) Gene transfer to plants; Plant genetic Engineering. Glasgow, NY pp 40-77
Fry, J., Barnason. A. and Horsch, R. B. (1987) Transformation of B. napus with A. tumefaciens based vectors. Plant Cell Reports 6: 321-325
Gu, X. (1998) Tissue and culture and transformation of Brussels sprouts. Thesis for the degree of Master of Agricultural Biotechnology in Horticulture. Faculty of Agriculture and Natural Resource Sciences, University of Adelaide
Hamada, M., Hosoki, T., Kusabiraki, Y. and Kigo, T. (1989) Hairy root formation and plantlet regeneration from Brussels sprouts (Brassica oleraceae var. gemmifera Zenk) mediated by A. rhizogenes. Plant Tissue Culture Letters 6: 130-133
Hoekima, A., Hirsch, P.R., Hooykaas, PJ.J. and Schliperoot, R.A. (1983) A binary plant vector strategy based on separation of vir- and T-region of the Agrobacterium tumefaciencs Ti-plasmid. Nature 303: 179-180
Hosoki, T. and Kigo, T. (1994) Transformation of Brussels sprouts (Brassica oleraceae var. gemmifera Zenk) by Agrobacterium rhizogenes harbouring a reporter, beta glucuronidase gene. Journal of the Japanese Society for Horticultural Science 63: 589-592
Jefferson, R.A. (1987) Assaying chimeric genes in plants: the gus gene fusion system. Plant Molecular Biology Reporter 5: 387-405
Mengiste, T., Amedo, P. and Paszkowski, J. (1997) High efficiency transformation of Arabidopsis thaliana with a selectable marker gene regulated by the T-DNA 1' promoter. Plant Journal 12(4): 945-948

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 75
Moloney, M.M., Walker, J.M. and Sharma, K.K. (1989) High frequency transformation of B. napus using Agrobacterium vectors. Plant Cell Reports 8; 238-242
Murashige, T. & Skoog, F. (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia of plantarum 15: 473-497
Radke, S.E., Turner, J.C., Faccioti, D. (1992) Transformation and regeneration of Brassica rapa using Agrobacterium tumefaciens. Plant Cell Reports 11: 499-505
Radke, S.E., Andrews, B.M., Molony, M.M., Crouch, M.L., Kridl, J.C. and Knauf, V.C. (1988) Transformation of B. napus L using Agrobacterium tumefaciens: developmentally regulated expression of a re-introduced napin gene. Theoretical Applied Genetics 75: 685-694
Takasaki, T., Hatakeyama, K., Ojima, K., Watanabe, M., Toriyama, K. and Hinata, K. (1997) Factors influencing Agrobacterium-mediated transformation of Brassica rapa L. Breeding Science 47: 127-134
Xiang, Y. Wong, W.K.R., Ma, M.C. and Wong, R.S.C. (2000) Agrobacterium-mediated transformation of Brassica campestris ssp. Parachinensis with synthetic Bacillus thuringiensis cry A(b) and cry A(c) genes. Plant Cell Reports 19: 251-256
Discussion Q. Glucosinalates cause nutritional disorders; if you husk their testa and increase consumption, won’t this be dangerous to consumer? A: Though the glucosinolates are harmful to consumers, they are vital for plant defences, hence a balance need to be struck so that the amounts are not harmful to human
THE EFFECT OF PREPARATION METHODS ON THE NUTRITIONAL QUALITY OF SOME VEGETABLES CONSUMED IN KENYA. A. 0. Makokha and D.K. Kebenei, Department of Food Science and Post-harvest Technology, Jomo Kenyatta University of Agriculture and Technology, P.O. Box 62000, Nairobi, Kenya. E-mail. [email protected]
Abstract Most of the local vegetables eaten in Kenya are prepared for consumption using various methods that includes boiling. Such preparation methods result in the loss of some nutrients, particularly vitamins. The most labile of these vitamins is ascorbic acid. This study aimed at assessing the effect of cooking on ascorbic acid loss in seven local vegetables. These vegetables were Brassica oleracea, Corchorius olitorius, Amaranthus spp., Clotoraria breviden, Solanum nigrum, Brassica napus, and Vigna unguiculata. The ascorbic acid in the vegetables was determined after 0, 5, 10, 15, 20, 25 and 30 minutes of boiling. The ascorbic acid loss ranged from a minimum of 12.4% in B. oleracea boiled for 5 minutes to a maximum of 78.4% in C. breviden boiled for 30 minutes. The ascorbic acid loss after 5 minutes, the shortest boiling time in Amaranthus spp., V. unguiculata, B. oleracea, C. breviden, B. napus, C. olitorius and S. nigrum was 33.3%, 35.7%, 12.4%,48.5%, 14.5%, 18.5% and 34.2% respectively. After 30 minutes, the longest boiling time, the respective ascorbic acid loss was 66.7%, 75.3%, 56.6%, 78.4%, 66.1%, 57.4% and 73.7% respectively. During both boiling periods, B. oleracea consistently lost the least ascorbic acid, while C. breviden had the highest loss. For all varieties of vegetables, ascorbic acid loss increased with boiling time. However, the rate of loss differed among the vegetable varieties. The total vitamin loss was also influenced by the time required to cook the vegetable to adequate flavour and texture acceptability.
Introduction A wide range of leafy vegetables is consumed in Kenya. Juma (1989) and Maundu et al (1993) reported that about 220 different traditional leafy vegetables (TLV) are consumed in the country. This wide range of TL V is in accordance with the great diversity of communities and varied agro ecological zones found in the country (Maundu et al, 1999). These vegetables have in the past contributed significantly to the nutritional requirements of the Kenyan communities.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 76
Among the advantages of these TLVs over modern exotic ones is that they are easily and cheaply available. Their cultivation does not usually involve the use of expensive inputs such as fertilizer and pesticides. Some usually grow uncultivated (wildly). They are also generally resistant to drought, and to pests and diseases. They also have long harvesting periods. TL V such as Vigna unguiculata, Amaranthus spp., Solanum nigrum and Corchorus olitorius may be harvested severally as required for up to four months. Others such as Basella alba are perennial (Chweya, 1994). These vegetables are a valuable source of micronutrients, particularly in the rural areas. They provide most of vitamins C, A and some of the B vitamins. They are also an important source of essential minerals and dietary fibre (Mnzava, 1997). The content of these nutrients in the fresh vegetables have consistently been reported to be high by several authors ( e.g. Mathooko and Imungi, 1994; Imungi, 1989; Imungi and Potter, 1983). The main method of preparation of these TLV for consumption involves boiling in unspecified amounts of water. But most of the vitamins are heat labile, though the extent of susceptibility to heat varies among different vitamins. The maximum cooking losses of some vitamins are shown in Table 1. These losses not only include those due to heat, but some vitamins are also unstable in the presence of oxygen, while most are also sensitive to pH conditions. Water-soluble vitamins (vitamins C and the B group) may undergo some considerable leaching. Hence the most severe losses during cooking usually occur in this group of vitamins. Table 1: Stability of some vitamins during cooking (Ibekoronye and Ngoddy, 1985).
Vitamin Maximum cooking loss (%)
Ascorbic acid (C) 100 Folic acid 100 Thiamine (B1) 80 Riboflavine (B2) 75 Biotin 60 Tocopherol 55 Pantothenic acid 50 Pyridoxine (B6) 40 Carotene (provitamin A) 30
Traditional home preparation of vegetables is primarily aimed at making them more palatable and improving their flavour. There is little attention paid to the nutritional value. The boiling periods employed in the preparation of the different TL V may therefore cause considerable losses in the vitamins. But there is little documented information on the extent of vitamin loss in the different TLV. The main objective of this study was therefore to determine the loss of vitamin C during the cooking of some popular TLV. The study determined the loss of the vitamin in the different vegetables after various boiling periods, and also after the period that the vegetable took to be adequately cooked to palatability.
Materials and Methods Samples of seven different species of vegetables, all four weeks old were obtained from the university farm at Jomo Kenyatta University of Agriculture and Technology. These vegetables were (common name in brackets): Brassica napus (Rape); Brassica oleracea (Kale); Vigna unguiculata (Cowpea); Corchorus olitorius (Mrenda/Murere); Crotolaria brevidens (Mito); Amaranthus spp. (Pigweed) and Solanum nigrum (Black nightshade).The vegetables were harvested and packed in polythene bags. They were immediately transferred to the laboratory for ascorbic acid determination.
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 77
The leaves of :V. unguiculata, C. olitorius, Amaranthus spp, S. nigrum and C breviden were weighed, cooked and analysed whole. Those of B.oleracea and B. napus, which are relatively bigger, were cut into 2 cm2 pieces. Samples weighing l00g of each variety were placed in 700 ml of boiling water and boiled for 5, 10, 15, 20, 25 or 30 minutes. Ascorbic acid analysis of the fresh and boiled samples was done according to the 2, 6-dichlorophenolindophenol colorimetric method (AOAC, 1984). Palatability of the vegetables was determined subjectively by mouth feel of experienced consumers. All the laboratory determinations were done in triplicate. Analysis of data was done using the analysis of variance and also descriptively.
Results and Discussion Table 2: Effect of boiling time on the rate of loss of ascorbic acid in the vegetables
Percentage loss in ascorbic acid with cooking time (minutes) Vegetable variety
Ascorbic acid in fresh
vegetable (mg/100g)
5 10 15 20 25 30
Amaranthus 107.8a* 33.3ab 48.9bc 53.3ab 55.6bc 62.2ab 66.7ab V. unguiculata 106.4a 35.7ab 40.6ab 61.9b 66.6bc 73.0b 75.3b B. Oleracea 104.0a 12.4a 41.7ab 51.3ab 53.9bc 56.3ab 56.6a C. breviden 101.9a 48.5c 62.3c 65.3b 72.2c 78.2b 78.4b B. napus 104.5a 14.5a 21.8a 32.7a 45.7ab 58.3ab 66.1ab C. olitorus 130.1b 18.5a 27.8ab 29.6a 37.0a 48.1a 57.4a S. nigrum 117.7ab 34.2ab 50.0bc 57.8b 60.5bc 68.5ab 73.7ab Mean 110.3±9.35 28.2±13.3 41.91±3.76 50.3±13.93 55.9±12.02 63.5±10.40 67.7±8.57
* values in the same column followed by the same letter(s) are not significantly different (P<0.05) from each other. They differ significantly (P<0.05) with values that do not have same letter(s).
The ascorbic acid loss during boiling of the different vegetables for 5, 10, 15, 20, 25 and 30 minutes is shown in Table 2. The minimum ascorbic acid content in the fresh vegetables was 110.3mg/100g. The mean content of 101.9mg/100g was observed in C. breviden, while the maximum value of 130.1mg/100g was observed in C. olitorius. These values are similar to those reported by other workers (e.g. Maundu et al, 1999). Apart from C.olitorius, which had higher ascorbic acid than the rest of the vegetables (P<0.05), there was no significant difference in the ascorbic acid content in the other fresh vegetables. The loss in ascorbic acid increased with increasing boiling time in all vegetables (Figure l). The mean percent loss of ascorbic acid increased from 28.2% after 5 minutes to 67.7% after 30 minutes. However, the degree of loss was not similar in the different vegetables. At each boiling period, there were significant differences in the percent loss of the vitamins among the different vegetables (P<0.05). C. breviden consistently experienced the highest loss of the vitamin at all the boiling periods (P<0.05). On the other hand, C. olitorius was among the vegetables, which consistently lost the least vitamin Cat the various boiling periods. After 30 minutes, B. oleracea and C.olitorius had experienced the lowest loss of the vitamin while C. breviden and V. unguiculata had the highest loss. The results imply that some vegetables retain ascorbic acid better than others do during cooking. B. oleracea and C. olitorius had a higher retention of ascorbic acid than C. breviden and V unguiculata. Given the importance of these vegetables as a source of vitamins, and that the main loss of these vitamins is through cooking, then further investigation to confirm these
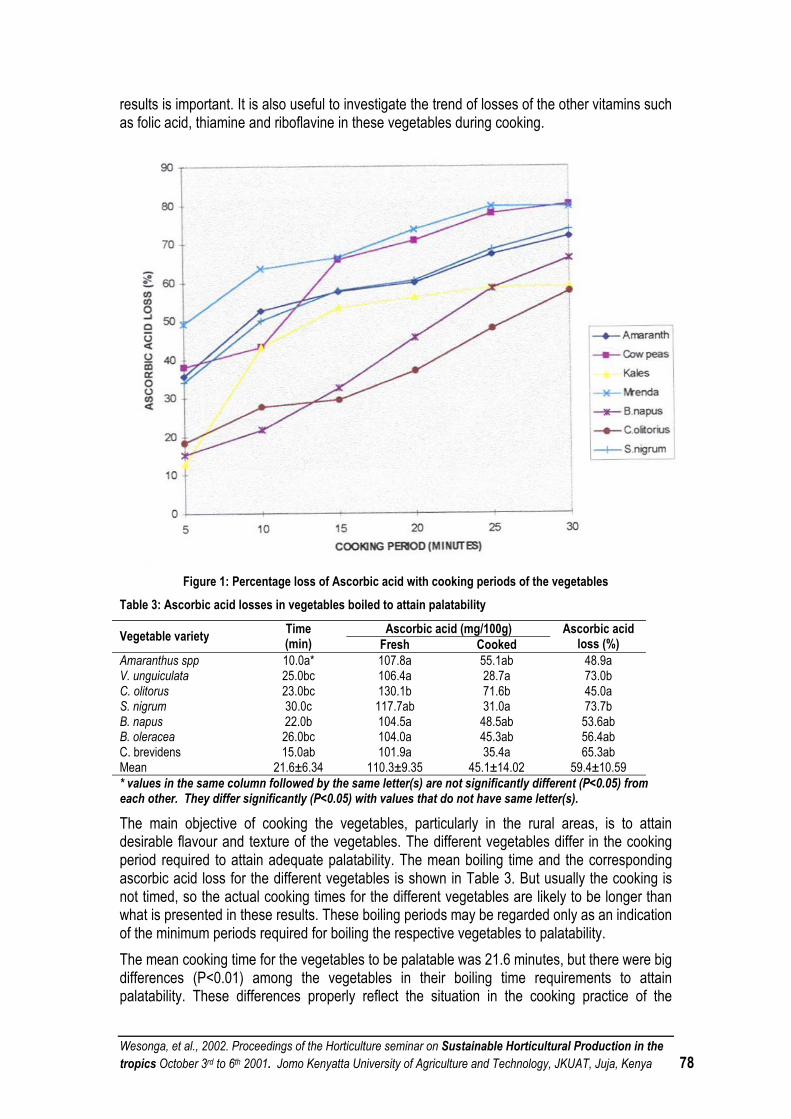
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 78
results is important. It is also useful to investigate the trend of losses of the other vitamins such as folic acid, thiamine and riboflavine in these vegetables during cooking.
Figure 1: Percentage loss of Ascorbic acid with cooking periods of the vegetables
Table 3: Ascorbic acid losses in vegetables boiled to attain palatability
Ascorbic acid (mg/100g) Vegetable variety Time (min) Fresh Cooked
Ascorbic acid loss (%)
Amaranthus spp 10.0a* 107.8a 55.1ab 48.9a V. unguiculata 25.0bc 106.4a 28.7a 73.0b C. olitorus 23.0bc 130.1b 71.6b 45.0a S. nigrum 30.0c 117.7ab 31.0a 73.7b B. napus 22.0b 104.5a 48.5ab 53.6ab B. oleracea 26.0bc 104.0a 45.3ab 56.4ab C. brevidens 15.0ab 101.9a 35.4a 65.3ab Mean 21.6±6.34 110.3±9.35 45.1±14.02 59.4±10.59 * values in the same column followed by the same letter(s) are not significantly different (P<0.05) from each other. They differ significantly (P<0.05) with values that do not have same letter(s).
The main objective of cooking the vegetables, particularly in the rural areas, is to attain desirable flavour and texture of the vegetables. The different vegetables differ in the cooking period required to attain adequate palatability. The mean boiling time and the corresponding ascorbic acid loss for the different vegetables is shown in Table 3. But usually the cooking is not timed, so the actual cooking times for the different vegetables are likely to be longer than what is presented in these results. These boiling periods may be regarded only as an indication of the minimum periods required for boiling the respective vegetables to palatability. The mean cooking time for the vegetables to be palatable was 21.6 minutes, but there were big differences (P<0.01) among the vegetables in their boiling time requirements to attain palatability. These differences properly reflect the situation in the cooking practice of the

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 79
vegetables. Some vegetables such as Amaranthus spp., which required only ten minutes of boiling to become palatable, are usually cooked for a much shorter time than others such as Solanum nigrum, which was heated for thirty minutes to become palatable. These differences in the requirement of boiling time will also affect the ascorbic acid loss during cooking. For instance, Amaranthus spp., which was boiled for only 10 minutes to attain adequate palatability, experienced a loss of 48.9% of the ascorbic acid. On the other hand, V unguiculata and S.nigrum, which were heated for 25 and 30 minutes respectively experienced a relatively higher loss of 73.0 and 73.7%. Further work involving a survey of vegetable consumers, to establish an estimate of the actual cooking times in households will be useful. The recommended daily requirement for ascorbic acid is 60 mg for an adult (F AO, 1988). Considered on fresh weight basis, 100g of each of the seven vegetables in this study would be enough to supply the required vitamin per day. But after boiling for 30 minutes, the same quantity of any of the vegetables will not be able to supply sufficient quantities of the vitamin. Unfortunately, most of the food composition data give information about the vitamins available in fresh vegetables, and not the cooked ones. Such data may therefore be misleading as an indication of the dietary vitamins obtained from vegetables in cooking conditions involving boiling.
Conclusions and Recommendations Though boiling results in loss of vitamin C in all vegetables, the rate of loss of the vitamin is higher in some vegetables under the same cooking conditions than in others. Under such conditions, the vegetables that lose the vitamin at a lower rate will retain higher levels of the vitamin. The different vegetables will also vary in their vitamin C retention because they have different cooking periods required to attain acceptable palatability. The vitamin loss is generally higher in the vegetables requiring longer cooking period to attain palatability than in those that take a shorter period of cooking. There is a need to investigate further the cooking conditions used by the Kenyan communities for local vegetables, and the effect of these conditions on the loss or retention of labile nutrients. These cooking conditions may vary among communities, or due to geographical location. There is also need to investigate the changes taking place in other labile nutrients, particularly the B group of vitamins such as folic acid and riboflavine during this preparation of vegetables. Further work is also required to determine the basis for the differences in the rate of loss of ascorbic acid among the vegetables.
References AOAC (Association of Official Analytical Chemists), 1984. Official methods of analysis of the
Association of Official Analytical Chemists. 14th ed. Washington DC, USA. Chweya, J. 1994. Potential for Agronomic Improvement of Indigenous Plant Germplasm in
African Agriculture - A Case Study of Indigenous Vegetables in Kenya. In Safeguarding the Genetic basis of Africa's Traditional Crops. Proceedings ofa CTA/IPGRI/KARI/UNEP seminar, 5-9 October 1992, Nairobi, Kenya (putter, A. ed.). Pg.105-113.
F AO (Food and Agriculture Organization of the United Nations), 1988. Requirements of vitamins. Report of a joint F AO/WHO Expert Consultation, Rome Italy.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 80
Ihekoronye, A.I. and Ngoddy, P.O. 1985. Integrated Food Science and Technology for the Tropics. Macmillan International College Editions. Pg. 91.
Imungi, J. K. 1989. The role of indigenous vegetation in human nutrition in Kenya. Paper presented at the Indigenous Vegetation Experience Sharing Forum, held at the Jomo Kenyatta University College of Agriculture and Technology, Juja.
Imungi, J. K. and Potter, N. N. 1983. Nutrient contents of raw and cooked cowpea leaves. J. Food Sci. 48: 1252
Juma, C. 1989. Biological Diversity and Innovation: Conserving and utilizing genetic resources in Kenya. ACTS Research Series No. 3:35-40.
Mathooko, F.M., Imungi, J.K. 1994. Ascorbic acid changes in three indigenous Kenyan leafy vegetables during traditional cooking. Ecol. Food Nutrition 32:239-245.
Maundu, P.M., Kabuye, C.H.S. and Chweya, J.A. 1993. Proceedings of the Indigenous Food Plants Workshop. National Museums of Kenya, Nairobi.
Maundu, P.M., Njiro, E.I., Chweya, J.A., Imungi, J.K., Sewe, E.N. 1999. Kenya in Biodiversity of Traditional Leafy Vegetables (Chweya, J.A. and Eyzaguirre, P.B. eds.) IPGRI. Pg. 51-84.
Mnzava, N .A. 1997. Comparing nutritional values of exotic and indigenous vegetables in African Indigenous Vegetables. Proceedings ofNRI/IPGRI/CPRO workshop, 13-28 January 1997, Limbe, Cameroon (R. Scippers and L. Budds, eds.). ODA, UK. Pg. 70-75
Discussion Q. Did you consider the factors like addition of milk salt etc in relation to how they affect vitamin content after boiling? A. This was not done but is important and need to be assessed. Q. Which equipment was used for boiling; pot or sufuria? A. The boiling was done in a sufuria (pan) Q. Does the intensity of heating affects the loss of nutrients - Buigut S.K. A. We used only boiling water; we did not measure the temperature
PRINCIPLES OF SUSTAINABLE NUTRIENT MANAGEMENT IN VEGETABLE PRODUCTION Stützel, H. Institute of Vegetable and Fruit Science, University of Hannover, Herrenhäuser Str. 2, D-30419 Hannover, Germany e-mail: [email protected]
Abstract Sustainable nutrient management requires in the steady-state situation of soil fertility (i.e. if soil fertility has reached its optimum) a balance between nutrient input and output. This is particularly important for nutrients that can easily be carried by water out of the rooting zone. In vegetables such a balance is difficult to achieve since the production of good market qualities together with the mostly limited root system requires high nutrient supply. Since vegetables are, unlike arable crops, harvested at full growth, they leave appreciable amounts of mineral nutrients in the soil behind. Moreover, many vegetable crops leave appreciable amounts of nutrients in their harvest residues behind which readily decompose. To achieve an input-output balance requires optimization of nutrient management on several scales: On the crop level, the minimum quantity to achieve maximum yield and quality has to be determined for given conditions and yield expectations. The empirical approach has very successfully been pursued for nitrogen requirement in Germany and has lead to very reliable recommendation systems. This approach is based on the relationships between nitrogen supply at given stages and maximum yield. In Hanover, we

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 81
are working on a method to determine the nitrogen strategy based on the physiology of nitrogen in the plant. The two major impacts of nitrogen on productivity are through the role of proteins in photosynthesis and the fact that proteins require energy for their turnover, i.e. cause maintenance respiration. Model studies show that the nitrogen requirement depends on the light intensity, which influences photosynthesis, and temperature, which determines respiration. On the cropping systems level it is important to include crops in the rotation which are able to capture the nitrogen left behind by the vegetables and released by mineralization of the residues. Important traits of such catchcrops are high growth rates and deep rooting systems. In areas with high rainfall growing such crops in mixtures with vegetables seems to be particularly advantageous.
RATIONALIZATION OF NUTRITIONAL REQUIREMENTS FOR SUSTAINABLE COFFEE PRODUCTION IN KENYA Bunyasi, S. W. and Mburu, J. N. Coffee Research Foundation P.O. Box 4 Ruiru
Abstract The aim of the coffee farmer is to maximise profit on a sustainable basis. To achieve this, he must manage his coffee trees efficiently for maximum output of high quality coffee fruits. This is majorly achieved if the coffee tree receives optimum nutritional conditions for the current season's production and ensuring that the tree is able to bear high yield in the subsequent season. This has been elusive to the Kenyan farmer as evidenced by the decline in the coffee production from national production of 130,000 tonnes in 1987/88 to about 56,000 tonnes in 1997/98. This is due to various factors of which decline in soil fertility on small scale farms and nutrient imbalance in large-scale farms contribute a major part. The paper presents results of a soil fertility survey carried out in six major coffee growing districts in Kenya in the year 2000. The survey showed excessive exchangeable acidity (> 0.5 m.e/100g soil), deficiency in Calcium (<1.6 m.e/100g soil), Magnesium (< 2.2 m.e/100g/soil), Zinc (<10 ppm) and Boron (<40ppm) in small scale farms. In the large-scale farms, there was excessive phosphorus (>100ppm) and Zinc and Boron deficiency. To reverse the declining coffee production and quality, the paper singles out four physiological stages in the coffee growth cycle as being critical in the nutrient management. These are floral initiation, flowering, berry development and ripening. It further suggests the nutrient requirements for each stage so that cumulatively, the farmer optimizes the yield and quality.
AFRICAN INDIGENOUS VEGETABLES-OPPORTUNITIES AND CONSTRAINTS Mary O. Abukutsa Onyango) Department of Horticulture, Maseno University, P.O. Private Bag, Maseno. Kenya, Tel. + 254-35-51622, 51179; Fax: + 254-35-51153, 51221; E-mail: [email protected]
Introduction Poverty, which comprises of food, shelter and clothing among other aspects is a major developmental problem in many developing countries. .In Kenya poverty remains a pervasive national problem presenting formidable challenges, which call for urgent action (GOK, 1999). According to surveys carried out by the Ministry of Planning and National Development, 50% of Kenya’s rural population are living below the poverty line. The central objective of the Kenyan national food security policy as stated in the sessional paper number 4 of 1981 is to ensure that adequate supply of nutritionally balanced food is available in all parts of the country at all times (Anon, 1981). The policy advocates the use of indigenous food sources to achieve this objective although for along time this has not been the case. (MOA, 1989, 1998).. According to Kenya government’s recent sessional paper, poverty alleviation is one of the main areas the government would like to address urgently. Among the natural resources that Kenya is endowed with include water, human, land, climate, and bio-diversity..Most of the agricultural bio-diversity has not been exploited in poverty alleviation strategies .Vegetables are considered

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 82
by many people in Kenya as just a by-product to add some flavour but are rarely considered as a main source of food The terms indigenous and traditional have been used interchangeably in reference to vegetables, however, Maundu (1997) has distinguished the two terms. Indigenous vegetables are those whose natural home is known to be in a specified region in this case Africa and traditional ones as indigenous or exotic species which due to long use have become part of the culture. The emphasis in this paper will be on African indigenous vegetables although some traditional ones will be included in the discussion. Studies carried out in various parts of Africa indicate that indigenous vegetables play a highly significant role in food security of the under-privileged in both urban and rural settings (Schippers, 1997). However, little is known about them and that many local vegetables found in sub-Saharan Africa are hardly domesticated. The major constraints facing the farming communities include lack of good quality seed and lack of information on these crops. Indigenous vegetables play an important role in income generation and subsistence. Recent surveys carried out in Western Kenya markets provided evidence that indigenous vegetables offer a significant opportunity for the poorest people to earn a living as producers and traders without requiring large capital investiments (Schippers, 1997). Indigenous vegetables provide important source of employment for those outside the formal sector in peri-urban areas in many African cities because of their generally short, labour intensive production systems, low levels of purchase input use and high yields (Schippers, 2000). Commercial growers can make a lot of money from the fast growing leafy vegetables now seen around many large cities. For many years, these vegetables have been neglected by researchers, policy makers, agriculturalists and funding agencies.These vegetables are facing extinction and there is a great need of not only conserving their germplasm but also improving their production. The potential that these crops have in food security and poverty alleviation has yet to be exploited.
Objectives of the paper: Highlight the value and potentials of African indigenous vegetables Highlight some of the research findings on African indigenous vegetables Identify major constraints of production and possible solutions Identify gaps for future research in African indigenous vegetables. .
Some identified African indigenous vegetables in Kenya Kenyans have traditionally made use of edible leaves of species growing wild as weeds. These plants are well known to the rural people and are often planted in home gardens. These vegetables are likely to be well adapted to harsh climatic conditions and disease infestation and are easier to grow.. These vegetables can be classified as cultivated, semi-cultivated and un-cultivated species. Some work done in Kenya has identified some of the indigenous leafy vegetables, which could be developed as high value cultivated crops (Table 1).
The value and potentials of African Indigenous Vegetables Nutritive value of African indigenous vegetables Limited research also indicates that these vegetables have high nutritive value. They contain high levels of minerals especially calcium, Iron and Phosphorus, Vitamins and Proteins (Chweya, 1985, Imbamba, 1973). In most cases the mineral and vitamin contents supersedes that found in popular exotic vegetables like cabbage, spinach and lettuce. African indigenous vegetables are therefore a valuable source of nutrition in rural areas where they contribute substantially to protein, mineral and vitamin intake (Mnzava, 1997). They are compatible to use

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 83
with starchy staples and represent a cheap but quality nutrition to the poor sector of the population in both urban and rural areas where malnutrition is widespread. Healthy people need a balanced diet consisting not just of starches but also proteins and minerals. People often consider vegetables to be of limited importance, mainly because they are not aware of the nutritive value. This is a clear misconception because the human body needs major minerals like iron, calcium, phosphorus, magnesium as well as trace elements and micronutrients that are essential for the health of the people, especially vitamins such as ß-carotene and ascorbic acid. a). Vitamins A Vitamin A is obtained from plants as carotene, which is broken down by oxidative processes to give the vitamin. Deficiency of vitamin A, results in night blindness where the light receptors sensitive to dim light do not function normally. TABLE 1. SOME IDENTIFIED KENYAN LEAFY VEGETABLE PLANTS ________________________________________________________________________________ COMMON NAME BOTANICAL NAME LOCAL NAME ________________________________________________________________________________ AMARANTHACEAE Amaranths Amaranthus blitum/lividus terere/libokoi/ododo Amaranthus cruentus Amaranthus dubius CAPPARACEAE Spiderplant Cleome gynandra Thageti/Tsisaka/Dek Gynandropsis gynandra CRUCIFERAE African kale Brassica carinata Kanjila/kadhira LEGUMINOSAE Sunnhemp Crotalaria brevidens kamusuusuu//Emiro/Mito Crotalaria ochroleuca Cowpea leaves Vigna unguiculata thoroko/Likhubi/Alot-bo SOLANACEAE African nightshades Solanum scabrum managu/Lisutsa/Osuga Solanum villosum Solanum eldoretii Solanum americanum TILIACEAE Jute Mallow Corchorus olitorius Mlenda/Murere/Apoth _________________________________________________________________________ Source: Imbamba, 1973, Chweya, 1985, Olembo et al, 1995, Maundu et al, 1999, Schippers, 2000.
Some of the identified indigenous plants would provide much more carotene than cabbage (Brassica oleraceae var capitata), which is commonly eaten in Kenya (Table 2) although the value for Kale (Brassica oleracea var acephala) is equally high. b). Vitamin C Vitamin C is obtained from vegetables as ascorbic acid or the acid-oxidized from dehydro-ascorbic acid. Its deficiency results in scurvy. From table 2 it can be noted that african indigenous vegetables have relatively higher levels than the exotic vegetables.
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 84
c). Iron Iron is important in the structure and function of the red blood cells. Lack of it results in anaemia, a common health problem in many developing tropical countries. Green leaves may provide some iron as shown in table 2. These indigenous vegetables may also be more superior in providing iron than the exotic leafy vegetables. d). Calcium Calcium is essential in the formation of bones. For growing children milk is the best source of calcium. Unfortunately, in developing tropical countries including Kenya, milk is rarely available after breast feeding ends. Green leafy vegetables may be very good sources of calcium (table 2). Table 2 also shows that the indigenous leafy vegetables may be superior to the exotic ones in their calcium content. TABLE 2: Nutrient contents of 100 g fresh weight edible portion of some indigenous and exotic leaf y
vegetables
_______________________________________________________________ Protein Calcium Iron Vit. A VitC (%) (mg) (mg) (mg) (mg) ___________________________________________________________________________________ Indigenous /Traditional Vegetables Amaranths 4.0 480 10 10.7 135 Spiderplant 5.1 251 19 8.7 144 A.Kale 3.5 --- --- --- 197 Sunnhemp 8.8 270 4 7.0 100 Cowpeas 4.6 152 39 5.7 87 Nightshades 4.7 291 12 8.8 131 Jute mallow 4.5 270 7.7 6.4 187 Exotic vegetables Kale -- 187 32 7.3 93 Cabbage 14 44 -- 1.2 33 Lettuce 12 62 2.2 0.04 18 Spinach 23 93 32 5.1 28 ___________________________________________________________________________________ Source :Imbamba, 1973, Adams and Richardson, 1977, Gomez, 1981, 1982, Imungi and Porter, 1983, Maundu et al, 1999.
e)Protein Protein is often deficient in diets in developing countries. This is especially so for nursing and expectant mothers, weanlings and pre-school children. In Kenya the production and consumption of grain legumes, which are not always available to the rural population, are encouraged but there is need to look for other foods that could supply protein. It has been shown that green leaves can provide as much protein as grain legumes and animals (Oke, 1973). Table 2 shows the protein content of the identified indigenous and other leafy vegetables consumed in Kenya

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 85
Health Most of these vegetables have been reported to have medicinal properties (Kokwaro, 1976, Olembo et al, 1995, Schippers, 2000), spiderplant has been reported to aid constipation and can facilitate birth and African nightshades have been reported to cure stomachache. Medicinal properties of some of the identified indigenous vegetables are shown in table 3. TABLE 3: Medicinal properties of indigenous vegetables
_____________________________________________________________ Vegetable local name Ailment treated _________________________________________________________________________________ Cleome gynandra Dek/Tsisaka Constipation. Facilitates birth Solanum spp Osuga/Lisutsa Stomach-ache Crotalaria spp Mitoo Stomach-ache Basela alba Indelema Stomachache and constipation Colocasia esculenta nduma Cure fast heartbeat Corchorus olitorius Mlenda Constipation _________________________________________________________________________________ Source: Olembo et al, 1995, Maundu et al, 1999, Schippers, 2000.
The little information available on the mode of preparation suggests that the presence of undesirable chemical compounds in these potential crops cannot be overruled. Tropane alkaloids constitute one of the distinctive groups of secondary metabolites of Solanaceae and many plants containing them have long been utilized for their medicinal, hallucinogenic and poisonous properties. Many plants belonging to the genus Solanum contain steroidal glycoalkaloids, which are known to exhibit in vitro toxicity to a wide variety of microorganisms and animals. This could be the reason why pest and disease incidences in some indigenous vegetables are less prevalent. Cleome gynandra, Solanum nigrum and Crotalaria brevidens have been reported to contain phenolic compounds and oxalates. Further research needs to be done to elucidate the medicinal properties of these African indigenous vegetables. Food Security and Income Generation The African Indigenous Vegetables can play an important part in food security, especially during periods of scarcity, they can be preserved and used to fill in the relish gap (Mnzava, 1997) and can be important household food security. Some leaves could be stored for upto 2 years when dried. One of the traditional ways of preserving vegetables is by sundrying or air drying and then stored. However, much valuable food is lost through damage by insects, rats, mice and other vermin, also as a result of high humidity, which encourages growth of fungi and bacteria. Further research is needed to utilise this opportunity and look into ways of improved storage using low cost methods including sun drying and packing in containers. Socio-economic survey was carried out on three markets in Western Kenya, namely Kakamega municipal market, Chavakali and Kiboswa on African Indigenous in June 1996 and June 2001. This was done in a period of seven days using a questionnaire where 400 traders were interviewed. The survey revealed the following: African indigenous vegetables are important commodities in household food security during
periods of scarcity. Leaves preserved traditionally by sun or air-drying can be stored for a period of two years. African indigenous vegetables can generate income and provide employment
opportunities. 70% of the farmers and traders interviewed were women
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 86
In certain cases especially in the smaller markets 95 % of the traded vegetables were African indigenous vegetables
There was a high demand for these vegetables in urban centres. In one of the markets 9% of the African Vegetables were being packed and transported to Nairobi.
Many of the vegetables may require low level purchase inputs Agronomic advantages a) Seed Production Seed production is one of the major problems in the vegetable industry in many tropical regions because most of the vegetables promoted are cool season biennial types which cannot produce seeds under tropical conditions. The African Indigenous Vegetables have an advantage over these exotic ones in that they can produce large quantities of seed within one season. The seed yield of six African Indigenous vegetables bulked at Maseno University is shown in table 4.. It was observed that seed yields were over 600 kg per hectare for all the vegetables bulked. The seeds if well selected, improved, and multiplied would readily be available to the farmers. These seeds however, need to be processed properly to maintain high seed quality TABLE 4: Seed yield of six indigenous vegetables
Name of Vegetable Seed yield (Kg/ha)
Amaranthus hybridus (Amaranths) Cleome gynandra (spiderplant) Crotalaria brevidens (sunnhemp) Vigna unguiculata (cowpeas) Solanum nigrum (Nightshades) Corchorus olitorius (Jute mallow)
920 700 750 700 700 620
Source: Onyango et al, 1999.
b) Response to fertilisers and manures The majority of farmers in the Kenya get leaf yields of 1-2 tons per hectare, yet research results indicate that yields up to 40 tons/ha are possible (Nekesa, 1997, Onyango et al, 1999).Far less researched are basic agronomic issues such as differentiating a number of varieties, the fertiliser regime and the possible use of readily available organic sources of manure, identify the best planting distance, cultivation techniques and post-harvest issues. Chweya, (1997) reported that Cleome gynandra responds well to inorganic fertilisers and farmyard manure. Most of the rural farmers cannot afford the inorganic fertilisers. This calls for the development of appropriate technologies that farmers can afford. Tithonia (Tithonia diversifolia) popularly known as Mexican sunflower, wild sunflower or bitter flower, which belongs to the family compositae, is a perennial shrub that proliferates around farms. Green and leaf biomass of tithonia has high contents of nitrogen (4%), phosphorus (0.7%) and potassium (4.1%) on dry matter basis (Jama et al, 2000). The leaves can be harvested and incorporated into soil to replenish the soil nutrients (ICRAF; 1996). This species is abundant in Western Kenya usually in untilled hedgerows around farms. Tithonia is a fast growing plant producing high biomass within a short period and can be propagated from both seed and vegetative cuttings. Biomass from Tithonia has been used as mulch in maize (Zea mais,L) and has been shown to improve crop yields in terms of size and quality of cobs, however, this manure has not been tried on vegetables. There is very little work reported in the use of organic and inorganic sources and their combination in the production of some of the traditional vegetables. There is a great need to promote the production of these indigenous vegetables that would play an important role in poverty alleviation and the general development
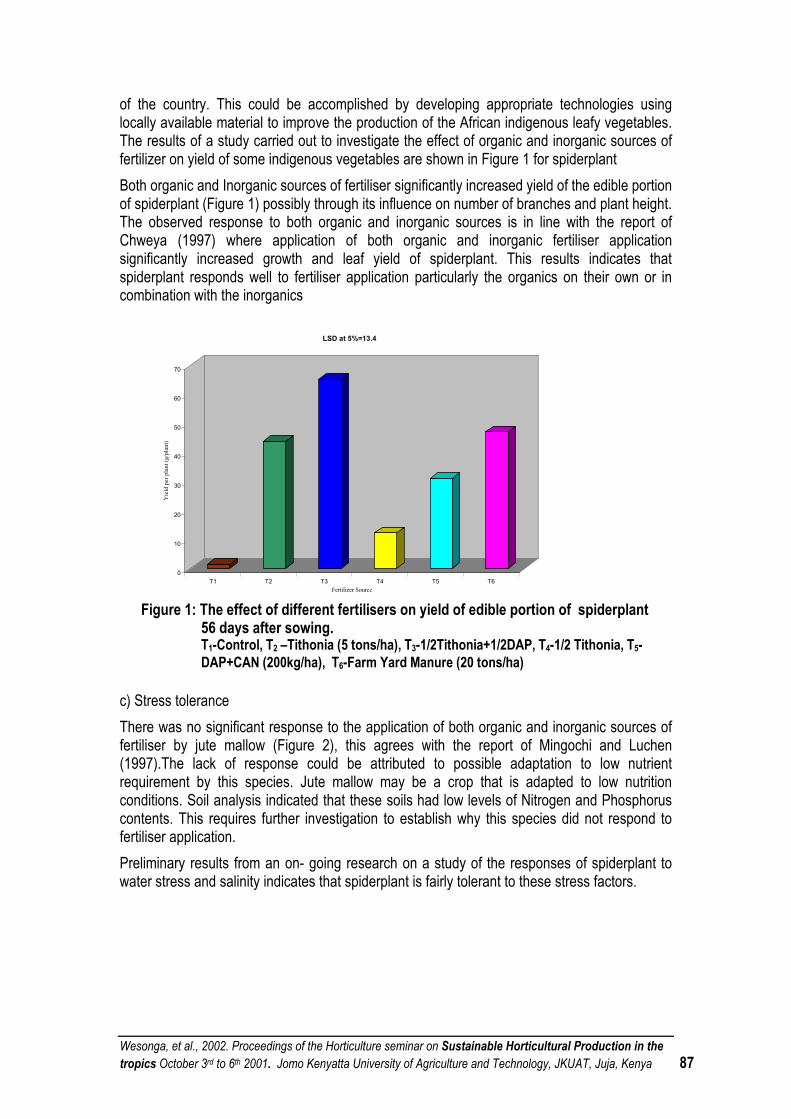
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 87
of the country. This could be accomplished by developing appropriate technologies using locally available material to improve the production of the African indigenous leafy vegetables. The results of a study carried out to investigate the effect of organic and inorganic sources of fertilizer on yield of some indigenous vegetables are shown in Figure 1 for spiderplant Both organic and Inorganic sources of fertiliser significantly increased yield of the edible portion of spiderplant (Figure 1) possibly through its influence on number of branches and plant height. The observed response to both organic and inorganic sources is in line with the report of Chweya (1997) where application of both organic and inorganic fertiliser application significantly increased growth and leaf yield of spiderplant. This results indicates that spiderplant responds well to fertiliser application particularly the organics on their own or in combination with the inorganics
0
10
20
30
40
50
60
70
Yie
ld p
er p
lant
(g/p
lant
)
T1 T2 T3 T4 T5 T6Fertilizer Source
LSD at 5%=13.4
Figure 1: The effect of different fertilisers on yield of edible portion of spiderplant 56 days after sowing. T1-Control, T2 –Tithonia (5 tons/ha), T3-1/2Tithonia+1/2DAP, T4-1/2 Tithonia, T5-DAP+CAN (200kg/ha), T6-Farm Yard Manure (20 tons/ha)
c) Stress tolerance There was no significant response to the application of both organic and inorganic sources of fertiliser by jute mallow (Figure 2), this agrees with the report of Mingochi and Luchen (1997).The lack of response could be attributed to possible adaptation to low nutrient requirement by this species. Jute mallow may be a crop that is adapted to low nutrition conditions. Soil analysis indicated that these soils had low levels of Nitrogen and Phosphorus contents. This requires further investigation to establish why this species did not respond to fertiliser application. Preliminary results from an on- going research on a study of the responses of spiderplant to water stress and salinity indicates that spiderplant is fairly tolerant to these stress factors.
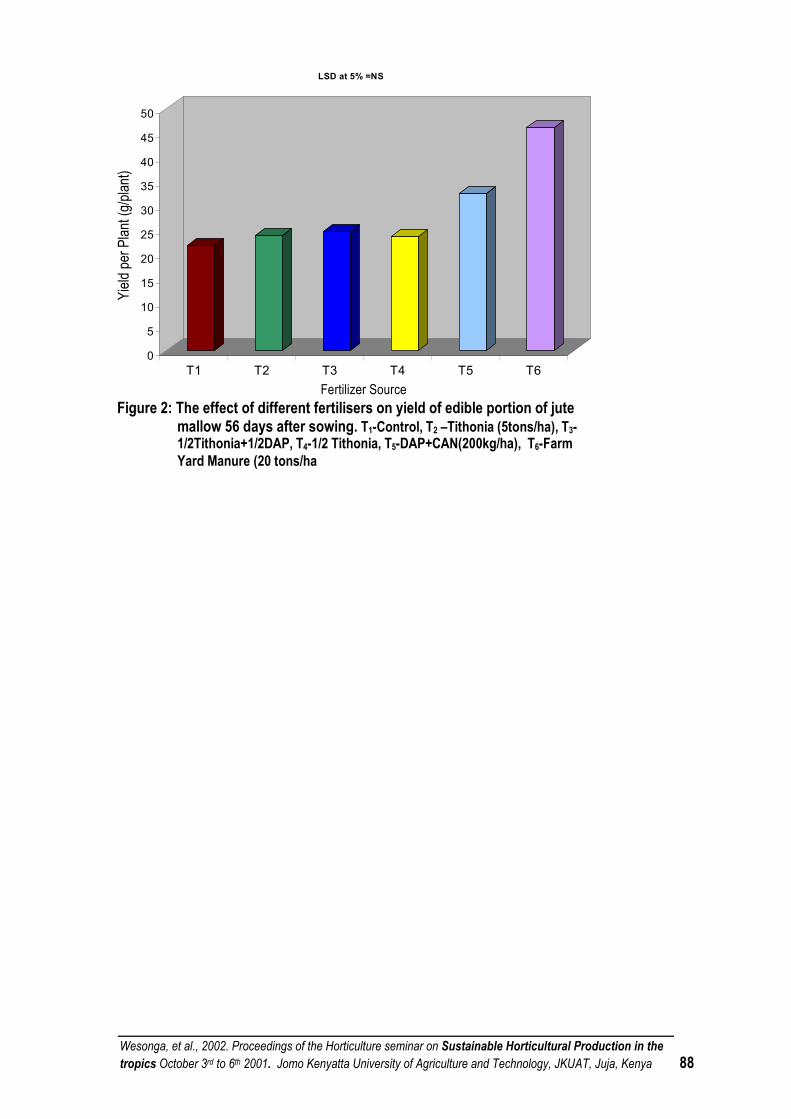
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 88
0
5
10
15
20
25
30
35
40
45
50Yi
eld pe
r Plan
t (g/p
lant)
T1 T2 T3 T4 T5 T6Fertilizer Source
LSD at 5% =NS
Figure 2: The effect of different fertilisers on yield of edible portion of jute mallow 56 days after sowing. T1-Control, T2 –Tithonia (5tons/ha), T3-1/2Tithonia+1/2DAP, T4-1/2 Tithonia, T5-DAP+CAN(200kg/ha), T6-Farm Yard Manure (20 tons/ha

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 89
Major Constraints in the production of African Indigenous vegetables Lack of awareness of the merits and opportunities of African Indigenous Vegetables Lack of seed in terms of availability, seed quality and choice of varieties Lack of technical production packages and access to information Lack of organised and effecient marketing channels
Possible interventions in improving production of African Indigenous Vegetables Publicity campaign and sensitization of the merits and opportunities of African Indigenous
vegetables Identify the preference of consumers, producers and traders of the various African
Indigenous Vegetables. Conduct participatory landrace enhancement of African Indigenous vegetables Encourage the establishment of village based banks with Universities and Researchers
providing technical backstopping. Raise the status of cultivated and semi-cultivated species to commercial Horticultural
commodity by development of appropriate agronomic packages through on station and on farm research
Urge training institutions to include African indigenous vegetables in the curricula.
Conclusions and recommendations African indigenous vegetables are excellent sources of essential micro-nutrients, have
medicinal properties and can provide quality and affordable sources of foods to most people in both rural and urban areas.
The development of appropriate and affordable agronomic technologies can increase production of African indigenous vegetables this would not only improve the nutrition of the rural poor but also enable them earn some income from the surplus and this would improve the livelihoods and create employment
The potential for African Indigenous Vegetables in improving the livelihoods of the rural communities exists and can be exploited when the suggested research areas are addressed
Research on African Indigenous vegetables is likely to lead to new findings and not only contribute to science but also to people’s food security and general well being
Suggestions for further research Collection of germplasm-It is essential to make extensive and systematic collection of
germplasm, especially from more isolated areas and from people’s home gardens Screening:The germplasm needs to be screened by scientists with the help of producers
and consumers to identify desirable characteristics. Selected material should then be purified, leading to the development of uniform varieties. Non-selected materials with distinctly different characteristics should be preserved in gene banks for later use. Such material could be of value when searching for tolerance or resistant stress factors
Seed multiplication:Multiplication of seed from a number of varieties allowing farmers or traders to choose which ones suit them most
Development of new varieties: Combining desirable characteristics or elimination of less desirable ones is usually possible by means of simple crossing
Agronomic research: Identifying the major technical constraints facing farmers of indigenous vegetables and carrying out research to overcome these constraints.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 90
Technical package: Development of appropriate technical advisory packages dealing with agronomic issues i.e. nursery, spacing and fertilisation, use of tithonia, pest and disease control and with market and post harvest related issues. Farmers will also benefit from provision of reliable market information.
Plant analyses: Comprehensive leaf and tissue analyses for nutrients, anti-nutrient factors, active ingredients for medicinal purposes and insect repellent properties.
Acknowledgement We gratefully acknowledge the financial support from AHI (African Highland Initiative) , Maseno University and IPGRI (International Plant Genetic Resources Institute)
References Adams, C.A. and Richardson, M. 1977. Nutritive value of foods. USDA, Home and Garden
Bulletin No. 72. Anon, 1981. Sessional paper No 4 of 1981, on National food Security. MOA 1989. Horticultural production guidelines, Farm management handbook of Kenya vol. V.
Ministry of Agriculture. Kenya. MOA, 1998. District Annual reports. Ministry of Agriculture. Kenya. ICRAF; 1996. Biomass transfer technology for soil fertility improvement in Western Kenya.
Annual Report, ICRAF; 1996. Chweya, J.A. (1985). Identification and nutritional importance of Indigenous green leaf
vegetables in Kenya.Acta Horticulturae:153: 99-108. Chweya, J.A. (1997). Genetic enhancement of indigenous vegetables in Kenya. In: Traditional
AfricanVegetables. Promoting the conservation and use of under-utilised and neglected crops. 16. Guarino, L. editor. Proceedings of the IPGRI International workshop on genetic Resources of Traditional Vegetables in Africa: Conservation and Use, 29-31 August 1995, ICRAF-HQ, Nairobi, Kenya. Institute of Plant Genetic and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute, Rome, Italy. Pp 86-95
GOK, 1999. National Poverty Eradication plan 1999-2015. Gomez, M.I. 1981. Carotene content of some green leaf vegetables of Kenya and effects of
dehydration and storage on carotene retention. Journal of plant and food 3:231-244. Gomez, M.I. 1982.The evaluation of fruit and vegetable resources in the Machakos district in
relation to seasonal deficits and micro-nutrient deficiencies. Technical Report IDRC 39. Imbamba, S.K. 1973. Leaf protein content of some Kenya vegetables. East African Agriculture
and Forestry Journal 38:246-251. Imungi, J.K. and Porter, N.N. 1983. Nutrient contents of raw and cooked cowpea
leaves.Journal of food science. 48:1252-1254. Kemei, J.K., R.K.Wataaru and E.N. Seme. 1997. The role of National Genebank of Kenya in
the collecting, characterization and conservation of traditional vegetables. In: Traditional African Vegetables. Promoting the conservation and use of underutilized and neglected crops. 16. Guarino, L. editor. Proceedings of the IPGRI International workshop on genetic Resources of Traditional Vegetables in Africa: Conservation and use, 29-31 August 1995, ICRAF-HQ, Nairobi, Kenya. Institute of Plant Genetic and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute, Rome, Italy: pp. 78-85
Kokwaro, J.O. (1976). Medicinal Plants of East Africa.East African Literature Bureau. Kampala, Nairobi, Dar es salaam: pgs 384.
Maundu, P.M.1997. The status of traditional vegetable utilization in Kenya. In: Traditional African Vegetables. Promoting the conservation and use of underutilized and

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 91
neglected crops. 16. Guarino, L. editor. Proceedings of the IPGRI International workshop on genetic Resources of Traditional Vegetables in Africa: Conservation and Use, 29-31 August 1995, ICRAF-HQ, Nairobi, Kenya. Institute of Plant Genetic and Crop Plant Research, Gatersleben/ International Plant Genetic Resources Institute, Rome, Italy: pp.66-75
Maundu, P.M., G.W. Ngugi and Kabuye, C.H. (1999) Traditional Food Plants of Kenya.Kenya Resource Centre for Indigenous Knowledge, National Museums of Kenya: pgs 270.
Mingochi D.S and S.W.S. Luchen (1997). Traditional vegetables in Zambia:genetic resources, cultivation and uses. In: Traditional African Vegetables. Promoting the conservation and use of underutilized and neglected crops. 16. Guarino, L. editor. Proceedings of the IPGRI International workshop on genetic Resources of Traditional Vegetables in Africa: Conservation and Use, 29-31 August 1995, ICRAF-HQ, Nairobi, Kenya. Institute of Plant Genetic and Crop Plant Research, Gatersleben International Plant Genetic Resources Institute, Rome, Italy:pp. 136-141
Mnzava, N.A. (1997). Vegetable crop diversification and the place of traditional Species in the tropics. In: Traditional African Vegetables. Promoting the conservation and use of underutilized and neglected crops. 16. Guarino, L. editor. Proceedings of the IPGRI International workshop on genetic Resources of Traditional Vegetables in Africa: Conservation and Use, 29-31 August 1995, ICRAF-HQ, Nairobi, Kenya. Institute of Plant Genetic and Crop Plant Research, Gatersleben International Plant Genetic Resources Institute, Rome, Italy:pp. 1-15
Nekesa,P. and B. Meso.1997. Traditional African vegetables in Kenya: production, marketing andutilization. In: Traditional African Vegetables. Promoting the conservation and use of underutilized and neglected crops. 16. Guarino, L. editor. Proceedings of the IPGRI International workshop on genetic Resources of Traditional Vegetables in Africa: Conservation and Use, 29-31 August 1995, ICRAF-HQ, Nairobi, Kenya. Institute of Plant Genetic and Crop Plant Research, Gatersleben International Plant Genetic Resources Institute, Rome, Italy:pp. 98-103.
Oke, O.L. (1973). Leaf protein research in Nigeria: A review of Tropical Science. 15:139-155 Olembo, N.K., S.S. Fedha and E.S. Ngaira (1995). Medicinal and Agricultural Plants of
Ikolomani Division, Kakamega District. ISBN 9966-9893-1-5:pgs 107. Onyango, M.O.A., J.C. Onyango and B.Jama (1999). Traditional Crops in Western
Kenya:Adaptive Technologies for Increased Production.IRPS. Maseno University , Reprint Series No. 3.
Schippers, R.R. 1997. Workshop proceedings of African indigenous vegetables.January 13-18, 1997. Limbe, Cameroon, ODA, IPGRI&NRI. UK & KENYA
Schippers, R.R. 2000. African indigenous vegetables an overview of the cultivated species. Chatham, UK. Natural Resources Institute /ACP-EU Technical Centre for Agricultural and rural Cooperation.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 92
INFLUENCE OF WATER LEVEL AND GENOTYPE ON TRANSPIRATION OF SPIDER PLANT AND BLACK NIGHTSHADE IN POT EXPERIMENTS Masinde P.W2., H. Stuetzel1 and S.G. Agong2 1. Institute of vegetable production, Hannover University, Germany 2. Department of Horticulture, JomoKenyatta University of Agriculture & Technology
Abstract A screening trial was conducted to establish the response of transpiration of selected genotypes of spider plant (Gynandropsis gynandra (L.) Briq) and black nightshade (Solanum nigrum L.) under well-watered and droughted conditions. There were five spider plant genotypes (wild types from Makueni and Kitui, landraces from Bungoma and Mombasa and a commercial type from Kenya seed), and four genotypes of black nightshade (landrace from Bungoma, wild types from Makueni and Meru and commercial type from Kenya seed company). The experiment was conducted at the JomoKenyatta University of Agriculture & Technology farm (March-June, 2001). Sowing was done directly in 20 litre pots. Temperature and light were not controlled. The fraction of transpirable soil water (FTSW) was used as the stress index. Genotypic differences were observed in the cumulative transpiration under well-watered conditions for both crop species. The ranking of cumulative transpiration from highest to lowest was Bungoma landrace/Commercial type, Makueni/Kitui wild types and lastly Mombasa landrace for spider plant and Bungoma landrace/Commercial type, Meru and Makueni wild types for black nightshade. Under droughted conditions, the Mombasa landrace had higher FTSW at all dates compared to the other genotypes of spider plant. In the case of black nightshade, the wild types from Makueni and Meru maintained higher FTSW at all dates compared to the Bungoma landrace and the commercial type. At the same time the wild types took longer to reach the end point. The normalized transpiration ratio (NTR) declined below 1.0 at FTSW range of 0.42-0.68, for the spider plant genotypes. However, the differences were not significant. For black nightshade, the range was 0.34-0.80 and the differences were significant. Key words: Traditional vegetables, spider plant, black nightshade, transpiration
Introduction Spider plant (Gynandropsis gynandra (L.) Briq) and black nightshade (Solanum nigrum L.) are among the most important traditional vegetable crops in Kenya. They are grown widely in Western and Coast provinces, predominantly in home gardens (MOALD&M, 1997). These crops form a substantial part of the diets of most low and middle level income earners in rural Kenya thereby earning the growers more income (Chweya, 1997). Moreover, these crops are grown and consumed as leafy vegetables in most parts of Africa and South East Asia (Edmonds and Chweya, 1997). Although there are no known botanical varieties or cultivars for both spider plant and black nightshade, genetic variation has been reported (Chweya, 1997; Edmonds and Chweya, 1997). Thus, there exists various variants and landraces in the areas in which these crops are utilized. To date there are no clear agronomic guidelines with respect to watering management of these key traditional vegetable crops in Kenya (MOALD&M, 1997). Water is recorded as the most limiting constraint in garden vegetable production. Consequently, we found it necessary to investigate the response of these two crop species to different water management. Production of spider plant and black nightshade in Kenya is mainly rainfed. In general, there are two seasons in Kenya, long rains (March-July) and short rains (October-December). This rainfall is unreliable making irrigation appropriate in solving this problem of erratic rains. However, water supply is limited and cost of irrigation is exorbitant for the small scale farmers. Furthermore detailed studies on the water relations of these crops are necessary in order to establish their responses to different water regimes. The objective of this screening was establish the reaction of various genotypes of two indigenous vegetable crops with respect to

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 93
changes in transpiration under well watered and droughted conditions. Fraction of transpirable soil water was used as the stress index.
Materials and Methods Experimental Site and Plant Materials The screening was conducted in plastic house at the JomoKenyatta University farm between March and June 2001. There were five types of spider plant and four types of black nightshade (Table 1). Table 1. Summary of screening experiment for both spider plant and black nightshade
Spider Plant Factor level details Plant type 5 1. 1 ‘Purple stem and purple petiole’ landrace (from Bungoma-
Western Kenya) 2. 2 wild types, ‘purple stems and purple petioles’ (Makueni and Kitui –Eastern Kenya) 3. 1 ‘Purple stem and purple petiole’ commercial line (Kenya seed company) 4. 1 ‘Green stem and green petioles’ landrace (from Mombasa-Coastal Kenya, sea level)
Water 2 droughted-no water after start of water treatments watered-watering to replace transpiration loss after start of water treatments,
Black Nightshade Factor level details Plant type 4 1 landrace ( Bungoma-Western Kenya)
2 wild types (Makueni and Meru –Eastern Kenya) 1 commercial line (Kenya seed company)
Water level 2 Watered and droughted
Pot Preparation and Planting 20 Litre plastic pots of 20 cm diameter were used. The pots were filled with red soil which had been thoroughly mixed, and weighed. There were 93 pots all with the same amount of soil. Each plant type had 10 pots (5 watered and 5 droughted), giving a total of 90 pots with three blanks. Pot capacity was determined by watering three pots thoroughly and allowing them to drain overnight while covered by polythene paper to prevent any evaporation. The pots were weighed (W1). Soil from these pots was sampled and weighed (W2), dried at 105°C for 48 hours and weighed again (W3). Pot capacity was calculated as:
W3)(W2W2W1yPotcapacit −×=
Seeds were sown directly in pots on 24th March 2001. They were well watered and covered by a polythene paper. Watering was done in the morning, afternoon and evening each day. Measurements Transpiration was measured by weighing pots at two days interval. The difference in pot weight was taken as the transpirational water loss. Transpiration ratio was calculated as:
plants wateredoftion transpiraAverageplants stressed ofion Transpirat(TR) ratioion Transpirat =

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 94
A second normalization was done so that the TR rate of each plant was centred on a value of 1.0 when the soil water content in each pot was high. First, a mean TR was calculated for each pot for the first 5 days from start of water treatments. Then the daily TR for each pot was divided by the mean TR to give a daily normalized transpiration ratio, thus:
mean
daily
TRTR(NTR) ratiotion transpiraNormalized =
Available soil water was expressed as the fraction of transpirable soil water (FTSW) for each pot in the droughted plants. Daily FTSW for each pot was calculated as:
Pot Weight FinalPot Weight Intialpot Weight Finalpot weightDaily FTSWDaily
−−=
The end point of the drought period was when the transpiration of stressed plants was about 10 % of that of watered plants. At this point, plants were harvested and divided into blades, petioles, stem and roots. Leaf area of blades was measures using leaf area meter (Model LI-3100). The plants were then dried at 70°C for 72 hours and weighed. Harvesting was done only for spider plant. Black nightshade plants were instead rewatered and maintained for seed production. Data analysis was done by costat computer package, and means were separated by LSD. The relationship between normalized transpiration ratio (NTR) and fraction of transpirable soil water (FTSW) was developed using both logistic function:
FTSW))*Bexp(*A1/(1NTR −+= ,
and plateu linear regression: NTR=1+A*(FTSW-B) if FTSW<B and NTR=1 if FTSW>B.
Using non linear procedure of SAS (SAS, 1999). r2 values were calculated as: r2=1-SSE/CSE where, SSE is the sum of squares of the residual, CSE is the corrected sum of squares.
Results Effect of Water Level on Transpiration of Spider Plant and Black Nightshade Cumulative transpiration of well watered plants showed significant differences between plant types for both crops (Fig. 1a). For spider plant, the Mombasa landrace had the lowest cumulative transpiration at all dates, while the Bungoma landrace had the highest in the early stages of the experiment. The commercial type had the highest in the later stages of the experiment. For black nightshade, the wild type from Makueni had the lowest cumulative transpiration, while the Bungoma landrace and the commercial type had the highest (Fig. 1b). The decline in FTSW was generally slower in the Mombasa landrace for spider plant (Fig. 2a) and in the wild types from Makueni and Meru for black nightshade (Fig. 2b). The time taken to reach the point at which the transpiration of the droughted plants was 10 % of the watered plants was significantly different between black nightshade types with the wild types taking longer to reach the end point. Spider plant types showed no differences in the time taken to the end point.
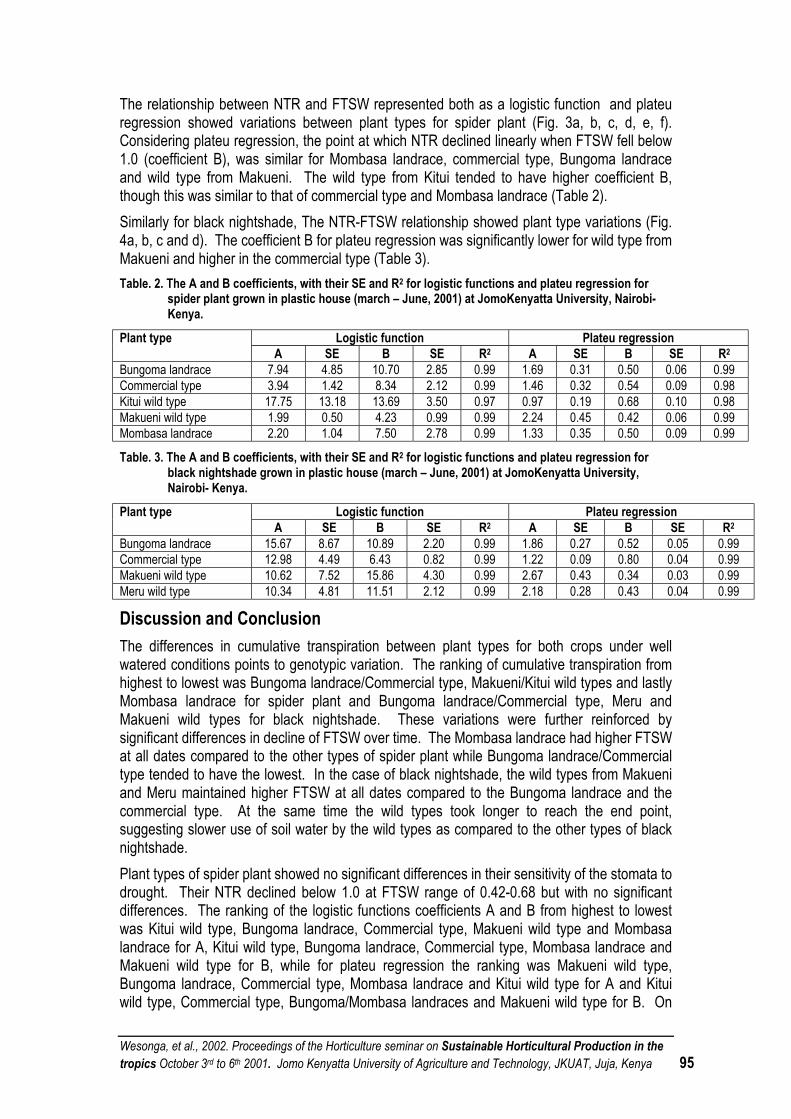
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 95
The relationship between NTR and FTSW represented both as a logistic function and plateu regression showed variations between plant types for spider plant (Fig. 3a, b, c, d, e, f). Considering plateu regression, the point at which NTR declined linearly when FTSW fell below 1.0 (coefficient B), was similar for Mombasa landrace, commercial type, Bungoma landrace and wild type from Makueni. The wild type from Kitui tended to have higher coefficient B, though this was similar to that of commercial type and Mombasa landrace (Table 2). Similarly for black nightshade, The NTR-FTSW relationship showed plant type variations (Fig. 4a, b, c and d). The coefficient B for plateu regression was significantly lower for wild type from Makueni and higher in the commercial type (Table 3). Table. 2. The A and B coefficients, with their SE and R2 for logistic functions and plateu regression for
spider plant grown in plastic house (march – June, 2001) at JomoKenyatta University, Nairobi- Kenya.
Logistic function Plateu regression Plant type A SE B SE R2 A SE B SE R2
Bungoma landrace 7.94 4.85 10.70 2.85 0.99 1.69 0.31 0.50 0.06 0.99 Commercial type 3.94 1.42 8.34 2.12 0.99 1.46 0.32 0.54 0.09 0.98 Kitui wild type 17.75 13.18 13.69 3.50 0.97 0.97 0.19 0.68 0.10 0.98 Makueni wild type 1.99 0.50 4.23 0.99 0.99 2.24 0.45 0.42 0.06 0.99 Mombasa landrace 2.20 1.04 7.50 2.78 0.99 1.33 0.35 0.50 0.09 0.99
Table. 3. The A and B coefficients, with their SE and R2 for logistic functions and plateu regression for black nightshade grown in plastic house (march – June, 2001) at JomoKenyatta University, Nairobi- Kenya.
Logistic function Plateu regression Plant type A SE B SE R2 A SE B SE R2
Bungoma landrace 15.67 8.67 10.89 2.20 0.99 1.86 0.27 0.52 0.05 0.99 Commercial type 12.98 4.49 6.43 0.82 0.99 1.22 0.09 0.80 0.04 0.99 Makueni wild type 10.62 7.52 15.86 4.30 0.99 2.67 0.43 0.34 0.03 0.99 Meru wild type 10.34 4.81 11.51 2.12 0.99 2.18 0.28 0.43 0.04 0.99
Discussion and Conclusion The differences in cumulative transpiration between plant types for both crops under well watered conditions points to genotypic variation. The ranking of cumulative transpiration from highest to lowest was Bungoma landrace/Commercial type, Makueni/Kitui wild types and lastly Mombasa landrace for spider plant and Bungoma landrace/Commercial type, Meru and Makueni wild types for black nightshade. These variations were further reinforced by significant differences in decline of FTSW over time. The Mombasa landrace had higher FTSW at all dates compared to the other types of spider plant while Bungoma landrace/Commercial type tended to have the lowest. In the case of black nightshade, the wild types from Makueni and Meru maintained higher FTSW at all dates compared to the Bungoma landrace and the commercial type. At the same time the wild types took longer to reach the end point, suggesting slower use of soil water by the wild types as compared to the other types of black nightshade. Plant types of spider plant showed no significant differences in their sensitivity of the stomata to drought. Their NTR declined below 1.0 at FTSW range of 0.42-0.68 but with no significant differences. The ranking of the logistic functions coefficients A and B from highest to lowest was Kitui wild type, Bungoma landrace, Commercial type, Makueni wild type and Mombasa landrace for A, Kitui wild type, Bungoma landrace, Commercial type, Mombasa landrace and Makueni wild type for B, while for plateu regression the ranking was Makueni wild type, Bungoma landrace, Commercial type, Mombasa landrace and Kitui wild type for A and Kitui wild type, Commercial type, Bungoma/Mombasa landraces and Makueni wild type for B. On

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 96
the other hand, plant types of black nightshade had significant differences in their sensitivity of the stomata to drought. The commercial type was most sensitive starting to close the stomata at FTSW of 0.80, followed by Bungoma landrace at 0.52, while the wild types were less sensitive, starting to close the stomata at 0.43 for Meru type and 0.34 for Makueni type. The NTR – FTSW relationship for both spider plant and black nightshade followed a similar pattern as that reported in many other crops. However, the FTSW values at which the NTR declined below 1 for spider plant and both types of black nightshade were relatively higher. In maize FTSW values of 0.2 (Muchow and Sinclair, 1991), 0.38 – 0.60 for a range of hybrids (Ray and Sinclair, 1997), and 0.31 (Ray and Sinclair, 1998) have been found. Sinclair and Ludlow (1986) reported that for grain legumes, soyabean, cowpea, blackgram and pigeon pea, a decrease in NTR occurred when FTSW fell below 0.3 while for field pea, a similar decrease NTR occurred at FTSW value below 0.4 (Lecoeur and Sinclair, 1996). Thus, the results in this study suggest that transpiration in spider plant and black nightshade is relatively more sensitive to drought. Cumulative transpiration and decline of FTSW over time formed the basis of selecting two types of spider plant for the field experiment since the other traits measured showed no significant differences. On this basis, the Mombasa landrace was selected as a type that used water slowly while Bungoma landraces a type that used water much faster. The selection for black nightshade was based on the NTR-FTSW relationship. In this case, Commercial type was selected as the most sensitive type starting to close the stomata at FTSW of 0.80 while Makueni wild type was less sensitive starting to close the stomata at FTSW of 0.34. However, the seed of Makueni wild type was not enough for field experiment, hence the Meru wild type which was the second less sensitive, starting to close the stomata at FTSW of 0.43, was selected.
Aknowledgement We wish to thank the Germany Academic Exchange Service (DAAD) for sponsoring this study, and also the department of Horticulture, JomoKenyatta University of Agriculture and Technology for for providing the necessary materials and logistic support during the study.
References Chweya, J. A. 1997. Genetic enhancement of indigenous vegetable in Kenya. In: Guarino L.,
(ed.), Traditional African vegetables. Promoting the conservation and use of underutilized and neglected crops. 16. Proceedings of the IPGRI international workshop on genetic resources of traditional vegetables in Africa: Conservation and use, 29-31 August 1995, ICRAF-HQ, Nairobi, Kenya). Institute of plant genetics and crop plant research, Gatersleben/International plant genetic resources institute, Rome, Italy, pp. 86-95.
Edmonds, J. M., Chweya J. A., 1997. Black nightshades Solanum nigrum L. and related species. Promoting the conservation and use of underutilized and neglected crops. 15. Institute of plant genetics and crop plant research, Gatersleben/international plant genetic resources institute, Rome, Italy, pp. 77-79.
Lecoeur, J., Sinclair T.R., 1996. Field pea transpiration and leaf growth in response to soil water deficits. Crop Sci. 36:331-335.
MOALD & M. 1997. Ministry of agriculture, livestock development and marketing, Horticulture division, Annual report pp. 7-8, 34.
Muchow, R.C., Sinclair T.R., 1991. Agroclimatology and modeling: Water deficit effects on maize yields modeled under current and greenhouse climates. Agron. J. 83:1052-1059.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 97
Ray, J.D., Sinclair T. R., 1998. The effect of pot size on growth and transpiration of maize and soyabean during water deficit stress. J. Expt. Bot. 49:1381-1386.
Ray, J.D., Sinclair T.R., 1997. Stomatal closure of maize hybrids in response to drying soil. Crop Sci. 37:803-807.
SAS Institute Inc., 1999. SAS/STAT Users guide. Sinclair, T.R., Ludlow M. M., 1986. Influence of water supply on the plant water balance of four
tropical grain legumes. Aust. J. Plant Physiol. 13:329-341.
Discussion Q: Which species of solanum were you working with (S. americani scrabrum or villisum, eldorlii) A: The classification is still not clear. I obtained the gerplasm from Genebank, under the name Solanum nigrum but I still have to get a correct classification Q. With transpiration stress for the various species LAI should be looked into, was this considered? In other words are there significant differences in LAI under the control, if there was any control treatment? F.K. Lenga
LABORATORY METHODS IN SCREENING CROPS FOR ALUMINIUM TOLERANCE Mugai, E.N. and Agong, S.G Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P .O. Box 62000 Nairobi, Kenya
Abstract Aluminium (Al) toxicity is the main soil factor inhibiting crop growth in acid soils with pH below 5.0. The growth inhibition is through reduced root growth, which in turn reduces the plant’s ability to take up water and nutrients. Although acid soils in Kenya occupy a small proportion of the total land mass (~10 %), this small area is the most agriculturally productive because of its higher precipitation. The reclamation of acid soils through techniques like liming is beyond the reach of many growers due to either financial or technological constraints. The most feasible alternative would be to fit crops to the soils through selection of adaptable crops. Selection of such crops would also be a primary step in breeding programmes in production acid soil-tolerant crops. Moreover most lucrative horticultural crops have been bred in alkaline soils of northern latitudes and are likely to be Al sensitive. There is little knowledge on the Al tolerance levels of most of the existing horticultural crops while progress to ameliorating the problem is hampered by lack of capacities in techniques of Al tolerance screening. This paper introduces some simple laboratory methods of Al tolerance screening and at the same time discusses the applicability for such technique under particular conditions.
POSSIBLE ROLE OF DAY LENGTH ON GRAPEVINE PERFORMANCE IN THE TROPICS Ndung'u, C. K.1; Okamoto G.2.; and Hirano K.2 1Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P. O. Box 62000 Nairobi, Kenya. 2Faculty of Agriculture, Okayama University, 1-1-1 Tsushima-naka, Okayama 700, Japan.
Abstract Production of temperate fruits in the tropical regions faces problems of erratic budbreak, poor bloom, and delayed foliation, resulting in reduced yields and poor quality fruits. Lack of chilling to release dormancy is considered the major limiting factor. However, lack of chilling has been shown to be compensated for by other factors such as water deficit stress, defoliation, pruning and training practices, and dormancy breaking chemicals. This study was designed to test the response of Kyoho grapevines to daylength, and whether a 12-hour daylength as prevails in the tropics might pose short day conditions that could limit budbreak and flowering. Vines were grown under 10, 12, 14, or 16-hour photoperiod. Short day treatments

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 98
were induced by covering the vines with opaque reflective polythene sheets, while long day treatments were induced by supplying extra hours of artificial light with 40-watt white fluorescent lamps. Photoperiod treatments were given for 3 months from anthesis to harvest. Short day conditions induced early cane ripening (periderm development) and berry maturation (red colour development). Lateral shoot growth was also less vigorous under short days. Accumulation of starch and nitrogen in shoot tissues was enhanced under short day conditions. Percent budbreak was lower, and numbers of clusters per shoot were fewer under short day than long day conditions. The results indicate a 12-hr daylength might pose short day conditions with significant effects on growth and development of grapevines in the tropics.
STORAGE AND PROCESSING POTENTIALS FOR HORTICULTURAL PRODUCE IN KENYA D. Shitanda Department of Biomechanical and Environmental Engineering Jomo Kenyatta University of Agriculture and Technology P.O. Box 62000 Nairobi, Kenya. Tel. 0151-52029, Email: [email protected]
Abstract Horticultural farming is gaining a lot of fame in Kenya and is now a among the leading foreign exchange earners. The produce which mainly include fruits and vegetables have high value but poor storage quality resulting in tremendous losses from the time of harvest up to the time they are marketed. Currently more effort is being directed towards the production and marketing of raw horticultural produce in foreign countries. However, little emphasis has been put on the storage, processing and local use of such produce. These aspects are very central when considering the product quality and net returns. Thus, there is need for them to be highlighted and greater attention given to them especially in the area of research. This is because storage helps to maintain quality, allows for market rescheduling and reduces losses. Processing on the other hand transforms the produce into products that are more valuable, easy to handle, use, and store. It also minimize the astronomical losses encountered during the peak season apart from ensuring continuous supply of similar quality products during off season. This paper therefore presents various storage and processing techniques that can be accessed buy the local producers and manufacturers in Kenya.
Introduction Kenya has an area of about 550,000 km2, of which over 70% is arid or semi arid. Despite the above limitation, Kenya's economy depends heavily on agriculture. Since independence in the early 60s, the country has depended on the export of coffee and tea for most of its foreign earnings. However, with time, tourism has also developed to be one of the major foreign exchange earners despite the high risks of political instability. With the drastic fall in world prices of coffee and tea, there has been a steady shift from perennial crop farming to horticultural farming with tremendous success. Horticultural farming has therefore optimized the use of the scarce arable land earning the country over US$0.2 billion in foreign exchange. It is now the second foreign exchange earner after tea (Nation News Paper, 2001). Most of the produce are marketed in Europe and the United States of America. Development and marketing of horticultural produce in Kenya has been under the Horticultural Crop Development Authority (HCDA) since the 1960s. The Board has put emphasis basically on the promotion of production and marketing of the crops although there are now few developments in the area of storage. Suggestions are however being made by the Fresh Produce Exporters Association of Kenya (FPEAK) that HCD should only act as an advisory board instead of being involved in the production. As farmers continue to increase their acreage with the increase international demands for Kenyan produce, the biggest challenge has been the sustainability of the production under the unpredictable world market economic trends and the stringent quality standards demanded by the buyers. Sustainability can be
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 99
attained through diversification of horticultural produce products and development of effective and efficient handling systems. The issue of quality especially for the Minimum Residue Levels (MRL) posses great challenge when considering that diseases and insect attacks are a major threat in the tropics. Currently there are no other effective non-chemical control methods for the same. The objective of this paper is therefore to highlight the potentials available for storage and processing of horticultural produce in Kenya so as to boost internal demand and minimize dependency on external markets.
Horticultural Production in Kenya Kenya produces a wide range of horticultural produce due to its conducive tropical climate. With the use of green houses, and irrigation production is be carried out all the year round with minimal climatic constraints. Some of the commonest crops produced are given in below.
Flowers Mangoes French beans Bananas Cucumbers Plums Paw paw Avocado Curry Pears Carrots Cabbage Tomatoes Pineapples Oranges Apples Radish Kales Grapes Onions Tangerine Coconuts Passion fruits
Objective Storage Storage of horticultural produce is very important since they loose their quality very fast and production occurs far from the market. Without effective and efficient storage systems, losses can be astronomical making the whole production processes uneconomical (Odogola, 1994). Basically, the reasons for storage include;
Continuity of supply during off seasons Handling of over production Sustainability and continuity of farm operations Reduction of field losses Reduction of quality losses Stabilization of market prizes
Currently in Kenya, cold storage is the main form of storage being used especially for high value produce. The storage is done at the source, during transportation and at the destination. The modern cold storage systems are however very expensive especially when considering the fact that electricity tariffs in Kenya are among the highest in the world. Also less than 30% of the country has access to electricity supply making it impossible to use modern cold storage systems especially in the rural areas. Continuous supply of electricity is also not guaranteed especially during the dry season when the water levels in the hydroelectric dams are low.
Percentage Crop Losses In Developing CountriesHorticultural crop losses in developing countries are generally very high due to the poorly developed infrastructure and scarce knowledge on the management of production systems. The situation is very critical in the tropics where people now depend on less than US$ 1 per day. Table 1 below gives some approximate figures for crop losses in developing countries. The losses occur mainly in the field, during handling and in storage.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 100
Table 1. Percentage Crop Losses in Developing Countries
Source: Bakker-Arkema, 1999
During storage, it is important to control the storage environment to ensure effective preservation of the stored produce. Some of the control parameters that are critical in modern storage systems include; Temperature Moisture and humidity Air velocity Lighting Oduor Pressure
Since the modern storage systems are expensive for the Kenyan farmer, there are other cost effective and simple systems that can be easily adapted for horticultural crops. They include; Charcoal coolers Clamps Box stores Underground pits Bins Crips
Figure 1 below shows an improved natural ventilated onion store that can be constructed easily on most Kenyan farms at low cost using readily available materials.
Apples 14 Avocados 43 Bananas 20 – 80 Cabbage 37 Carrots 44 Cassava 10 – 25 Cauliflower 49 Citrus 20 – 95 Grapes 27 Lettuce 62 Papaya 40 – 100 Plantain 35 – 100 Potatoes 5 – 40 Onions 16 – 35 Raisins 20 – 95 Stone fruit 28 Sweet potatoes 35 – 95 Tomatoes 5 – 50 Yams 10 - 60
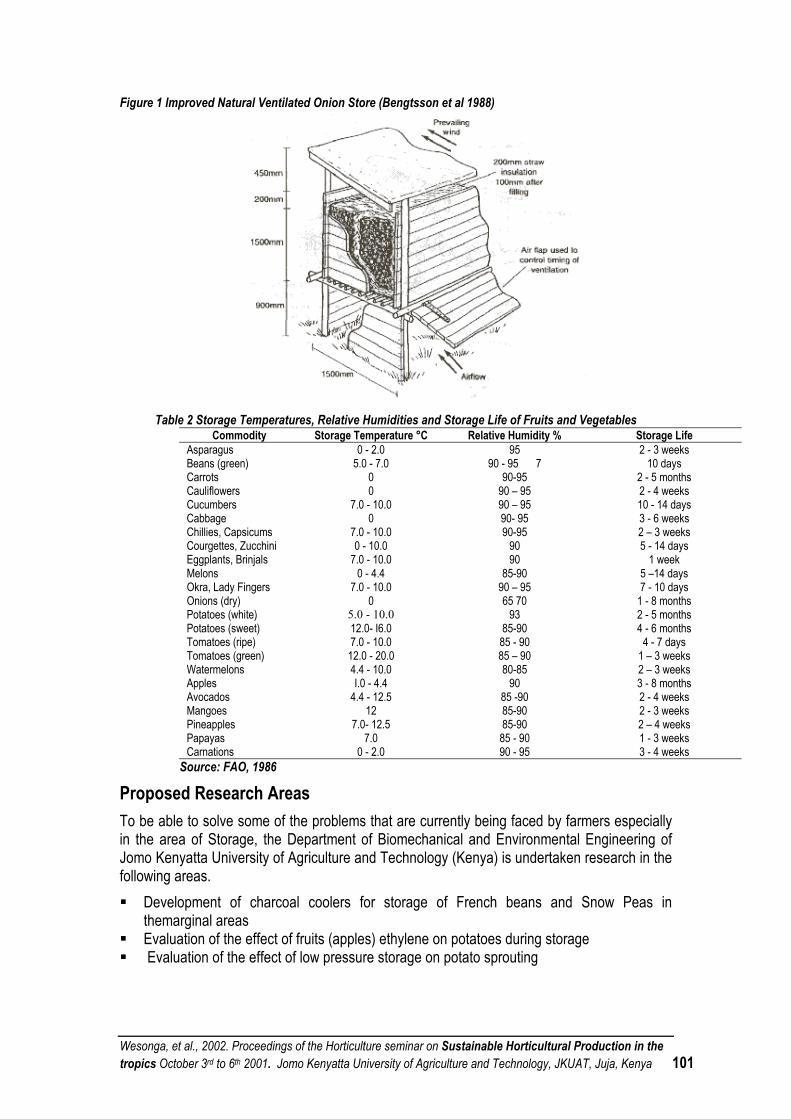
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 101
Figure 1 Improved Natural Ventilated Onion Store (Bengtsson et al 1988)
Table 2 Storage Temperatures, Relative Humidities and Storage Life of Fruits and Vegetables
Commodity Storage Temperature °C Relative Humidity % Storage Life Asparagus Beans (green) Carrots Cauliflowers Cucumbers Cabbage Chillies, Capsicums Courgettes, Zucchini Eggplants, Brinjals Melons Okra, Lady Fingers Onions (dry) Potatoes (white) Potatoes (sweet) Tomatoes (ripe) Tomatoes (green) Watermelons Apples Avocados Mangoes Pineapples Papayas Carnations
0 - 2.0 5.0 - 7.0
0 0
7.0 - 10.0 0
7.0 - 10.0 0 - 10.0
7.0 - 10.0 0 - 4.4
7.0 - 10.0 0
5.0 - 10.0 12.0- I6.0 7.0 - 10.0 12.0 - 20.0 4.4 - 10.0 I.0 - 4.4
4.4 - 12.5 12
7.0- 12.5 7.0
0 - 2.0
95 90 - 95 7
90-95 90 – 95 90 – 95 90- 95 90-95
90 90
85-90 90 – 95 65 70
93 85-90 85 - 90 85 – 90 80-85
90 85 -90 85-90 85-90 85 - 90 90 - 95
2 - 3 weeks 10 days
2 - 5 months 2 - 4 weeks 10 - 14 days 3 - 6 weeks 2 – 3 weeks 5 - 14 days
1 week 5 –14 days 7 - 10 days
1 - 8 months 2 - 5 months 4 - 6 months
4 - 7 days 1 – 3 weeks 2 – 3 weeks 3 - 8 months 2 - 4 weeks 2 - 3 weeks 2 – 4 weeks 1 - 3 weeks 3 - 4 weeks
Source: FAO, 1986
Proposed Research Areas To be able to solve some of the problems that are currently being faced by farmers especially in the area of Storage, the Department of Biomechanical and Environmental Engineering of Jomo Kenyatta University of Agriculture and Technology (Kenya) is undertaken research in the following areas. Development of charcoal coolers for storage of French beans and Snow Peas in
themarginal areas Evaluation of the effect of fruits (apples) ethylene on potatoes during storage Evaluation of the effect of low pressure storage on potato sprouting

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 102
Processing It’s a very important postharvest operation, which involves material handling and transformation aimed at meeting specified needs. Some of this needs include: - Increased storageability Diversified product forms High quality products Ease of handling materials and products Refined raw materials for secondary operationsDuring the processing, some of the
critical factors to consider include cost of the final product and operation, acceptability and nutritive value of the final product, availability of market and the environmental impact of the processing operation. Based on the above factors, the following processing operations can be carried out on the horticultural produce (Fellows, 1997).Drying Heat treatment Chemical treatment Freezing Dehydration Irradiation Solid-liquid extraction Fermentation Size reduction
Drying, which is one of the very common processing operations has been done for centuries (Brooker et al, 1992). It can be done simply and on small or large scale in any part of the country using the locally available resources. The modes of drying that can be used for horticultural produce include solar drying, freeze-drying, vacuum drying, air-drying and evaporation. Since solar radiations are abundant in the tropics (Lindley, et al, 1996), this makes it a very viable mode of drying. Solar driers can be used to facilitate the drying through concentration of the solar radiation (Hankins, 1995). Apart from the abundant, solar drying has also been found to improve quality of crops like coffee and rice. Air-drying can also be carried out in the rural area using ambient air or fuel-powered dryers (Figure 2). The fuels readily available include firewood, charcoal, maize cobs, and other plant wastes.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 103
Figure 2 Solid Fuel Rotary Dryer (Bakker-Arkema et al 1999)
Frit Juice Extraction Although fruit juice extraction is a very common processing operation in the developed countries, it has not taken root in Kenya where most of the horticultural produce are eaten or sold raw. The processing can be done on small scale or large scale by use of blanchers, fruit extractors, clarifiers, filters, mixers, sterilizers, pasteurization, and evaporation among others (Brennan, et al 1976; Bakker-Arkema, et al 1999). In Kenya, there is a very high potential for fruit juice extraction especially when considering the abundant and different fruit varieties available in the country. There is therefore great need to make use of the readily available fruits and vegetables like mangoes, oranges, apples, guavas, plums, paw paw, cashew nut fruit, carrots, tomatoes, passion fruits, and grapes among others. It is very important to note that most of the traditional fruits and vegetables have great potential for use and should be promoted hand in hand with the others.
Proposed Research Areas As a contribution to the promotion of processing of horticultural produce in Kenya, research is being carried at Jomo Kenyatta University of Agriculture and Technology, Department of Biomechanical and Environmental Engineering on the drying of the following fruits and vegetables. - Kales - Avocados - Bananas - Cabbage - Mangoes - Cashew nut fruits - Drying of cooked soybean - Amaranthus (Traditional vegetable) The venture into the drying of fruits and vegetables is due to the increased demand for dried products especially in the developed countries. Effort should therefore be made to encourage the use of dried products both locally and for export.
Conclusion Processing and storage of both conventional and non-conventional horticultural produce has great potential in developing counties especially Kenya. There is therefore need to intensify research in the highlighted areas so as to improve productivity, and quality and diversify the products for a much wider market both locally and internationally. This will minimize

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 104
dependency on raw materials for export, increase consumer variety, create more job opportunities through development of local industries boost local demand and give the country a competitive advantage on the international market
Reference Bakker-Arkema, F.W., Debaerdemaeker, J., Amirante, P., Ruiz-Altisent, M. and Studman, C.J.
(1999). CIGR Handbook of Agricultural Engineering. Agricultural Processing Engineering. Published by the American Society of Agricultural Engineers, USA.
Brennan, J.G., Butters, J.R., Cowell, N.D. and Lilly, A.E.V. (1976)Food Engineering Operations. Second Edition. Applied Science Publishers Ltd, London.
Brooker, D.B., Bakker-Arkema, F.W. and Hall, C.W. (1992). Drying and Storage of Grains and Oilseeds. Van Nostrand Reinhold publisher, New York.
Fellows, P. (1997). Traditional Foods. Processing for Profit. Intermediate Technology Publication, London, UK.
Hankins, M. (1995). Solar Electricity Systems for Africa. Published by Commonwealth, Science Council, London – England.
Lindley J.A. and Whitaker, J.H. (1996). Agricultural Buildings and Structures. Agricultural Processing Engineering. American Society of Agricultural Engineers, USA.
Nation News Paper (2001). Horticulture Players Fight New Law. Tuesday, September, Nairobi - Kenya
Odogola, W.R. (1994). Postharvest Management and Storage of Legumes. AGROTEC, UNDP/OPS Regional Programme, RAF/92/R51. Harare, Zimbabwe.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 105
COMPARATIVE STUDY OF BANANA (MUSA SPP.) PROPAGATION METHODS AT FARM LEVEL IN KENYA Macharia, I.1; Eckert M.V.2; Agong S.G.1 and Löhr, B.2
1Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P .O. Box 62000 Nairobi, Kenya 2Plant Health Division, International Centre of Insect Physiology and Ecology, P.O. Box 30772, Nairobi, Kenya, e-mail: [email protected]
Abstract Banana is one of the common fruits grown in Kenya. There are various methods of propagation but the recent one is by use of Tissue Culture (TC) technology. However most of the farmers in Kenya have not adopted this new technology. The main reasons have been identified as issues, but not quantified. There is also lack of information on opportunities and constraints related to this technology at farm level as compared to convectional suckers. The aim of designing this study was to determine the significance of TC banana and quantifying factors, which influence their adoption in Kenya. This will help to come up with a redefined extension package that will be used by extension officers in creating awareness and improving the information flow among the banana growers. The study was conducted in Runyenjes Division (Embu District), Suneka Division (Kisii District) and Soy Division (Keiyo District). Primary and secondary data were collected. Primary data were collected through participatory Rural Appraisal (Group discussions and interviews), field measurements and observations. Secondary sources of data included vital statistics of these Districts and monthly reports. The importance of tissue culture and convectionally produced banana was evaluated as a sources of income, their viability at farm level and yield performance. Farmers knowledge and knowhow of this technology was also examined. Average gross margin per banana per year derived from TC banana in Suneka, Runyenjes, and Soy were Kshs. 313, Kshs. 276 and Kshs. 144 respectively when compared to Kshs. 217, Kshs. 142 and Kshs. 113 for convectional suckers. This indicated that TC banana could function as a major source of income if adopted by all farmers in Kenya. It was also found out that tree crops contribute 26% in Suneka, 22% in Runyenjes and 11% in Soy. TC accounts for over 21% respectively towards this. The study found out that several factors, which include non-accessibility, lack of appropriate knowhow and low farm gate prices for bananas have limited adoption significantly. Farmers were found adopting the TC suckers instead of the plantlets. To reverse the persistent scarcity of adoption of plantlets, the study recommends a redefined extension approach, which can promote adoption. Creation of appropriate awareness, accessibility of plantlets at farm level and finding valuable market e.g. international market are suggested as necessary step for realization of benefits of TC banana. Hence effort should be made to back the extension officers in disseminating the information. It can also be sensible to collapse extension and social economic research into one body thus converting extension officer to social-economic researchers participatory development facilitators. This would greatly reduce the lag from the time information is generated, disseminated and used. Possibilities of establishing divisional deports for the TC plantlets should be sought out to give farmer access by relevant non-governmental organizations (NGO’s). Due to re-infection at farm level possibilities of breeding for resistance to these TC banana is also worthy researching. In conclusion, although the technology has been in Kenya since 1995 there is low adoption due to more people being unaware and non-accessibility of these plantlets at farm level. The significance of this study shows that TC banana to all farmers can greatly reduce poverty due to the higher income portrayed by gross margin per banana.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 106
FARMING CHARACTERISTICS AND CONSTRAINTS AMONG SMALL SCALE FRENCH BEAN (PHASEOLUS VULGARIS) FARMERS IN MWEA, KENYA. A.O. Makokha, F.K. Ombwara, L.S. Wamocho, J.M. Wesonga and P. Sichangi Jomo Kenyatta University of Agriculture and Technology, P.O. Box 62000, Nairobi, Kenya E-Email: [email protected]
Abstract Horticultural produce is the second most important commodity in export value in Kenya. A survey was done in Mwea division, Kirinyaga district to elicit information among small scale French bean (Phaseolus vulgaris) farmers about their farming practices and problems. Most respondents had farm sizes of less than five acres. Their entire French bean crop was meant for export. The beans were planted throughout the year under intensive production involving use of fertilizer, irrigation and pesticides. The French bean variety Amy was the most widely grow-n in the area. Other varieties grown included Samantha, Julia and Paulista. Pests and diseases of French beans were among the major problems experienced by the farmers. The main pests included red spider mite, bean fly, thrips and white fly. The main disease affecting the French beans was leaf rust. Other problems included lack of capital and credit facilities, fluctuation in demand, and difficult accessibility to the market.
Introduction Horticulture has been one of the most rapidly growing economic activity in the agricultural sector in Kenya. It offers employment to about two million Kenyans and also provides food. It therefore helps alleviate poverty and provide food security. By the year 2000, horticultural produce was the second leading export item in terms of value. It earned the country about 14 billion Kenya shillings (KES) (CBS, 2001). Out of this amount, about 5.5 billion KES was earned from the sale of vegetables, 7.3 billion KES from the sale of cut flowers and the rest from the sale of fruits. However, the Export market, where the European Union accounts for about 85% (CBS, 1999), is very competitive. The consumer and market demand high quality products. These quality requirements are in addition to the government and trade regulations such as those governing the use of pesticides. The quality requirements are in turn impacted upon by other factors. These include the variety planted, cultural practices, harvesting and post-harvest handling and preservation. All these aspects can be addressed by application of improved technologies. Horticultural export trade involves mainly cut flowers, vegetables and fruits. Cut flowers are mainly produced by large-scale enterprises. The export of fruits has been dominated by the pineapples from the Del Monte (formally Kenya Canners) company. It is in vegetable production and export trade where the growth has been fuelled by the participation of many new small-scale farmers. About 60% of the horticultural enterprises are run by small-scale enterprises. French beans (Phaseolus vulgaris) account for more than half of the value of vegetable exports, or about one quarter of the total horticultural exports. These French beans are the immature green pods of Phaseolus vulgaris. They are grown mainly in Central and Eastern Provinces. Mwea Division, Kirinyaga District in Central Province is one area where there is a high concentration of small-scale farmers of this crop. Within this context, this study therefore aimed at establishing the farming practices and problems experienced by the small-scale farmers involved in production and trade of French beans in Mwea Division, Kirinyaga District.
Methodology A survey was carried out among small scale French bean farmers in the two Locations of Kangai and Nyangati in Mwea Division, Kirinyaga district. This division was chosen for the survey because it has a relatively large concentration of small-scale farmers who are engaged in French bean farming. The two locations had the highest number of the farmers engaged in

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 107
French bean farming among the six locations in the division. In the two locations, about 1100 households were engaged in French bean farming. A sample of 100 farmers was interviewed. Sampling was done by choosing every 10th home along a road transect within the divisions. Where the l0th home did not have a respondent, the next nearest home to it with a respondent was chosen. Information about the farming practice and problems was obtained using structured questionnaires, observations and participatory discussions. The data obtained from the information was analysed descriptively.
Survey results and discussion Household Characteristics In both Kangai and Nyangati locations most of the respondents (farmers) were male. They were also the household heads. Their ages ranged from 25 to 60 years. Most of the farmers were educated up to primary level, with only a few educated up to secondary level. All the farmers were resident on their farms. Their major occupation, and major source of income was farming. There were three distinct types of land tenure systems among the French bean farmers in the division. There were those who had inherited (38.5%), purchased (30.7%) or rented (30.8%). Most farmers (68.2%) had less than two acres under French beans, while the remaining, 31.2%, planted between two to five acres of French beans. The acreage under the beans ranged from a minimum of less than half an acre to a maximum of six acres. On the average, about 50% of the farmers’ land was used for French bean cultivation. Generally, farm sizes in the Division were small. The division had a high population density of 246 people per square kilometre. Most of the farmers in the division (55%) started French bean cultivation between 1990 and 1995. One quarter of the farmers had started French bean cultivation between 1983 and 1989, while 25% started French bean cultivation after 1996. It is the main cash crop grown by these farmers. Other cash crops grown in by the farmers were tomatoes, bananas and rice in Kangai location and maize, tomatoes, bananas and vegetables in Nyangati location. The main food crops in both locations were maize and beans. Cultural Practices In both locations planting and harvesting of French beans was carried out continuously throughout the year. All the farmers applied fertilizer to their crop. Over half of them (55.1%) applied both inorganic and organic (farmyard) manure. Of the rest, 27.3% applied only inorganic fertilizer while 9% applied only organic fertilizer. The majority of the farmers (72.7%) in the division intercropped their French beans with other crops. The rest (27.3%) practised monocropping. In Kangai location only inter-cropping was practised while in Nyangati location both mono cropping and inter-cropping were practised, with mono cropping dominating. In both locations the farmers practised irrigation. The type of irrigation practised was furrow / canal irrigation. But the source of irrigation water differed in the two locations. In Kangai location canal water was the one used for irrigation, while in Nyangati location most of the farmers relied directly on river water for irrigation. The main source of power for irrigation in Kangai location was gravity with only a few relying on petrol pumps. But in Nyangati location the farmers relied on petrol pumps to get water from the river to their farms.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 108
Slightly less than half of the farmers in the two locations (40.9% ), experienced problems in obtaining seeds for their French bean crop. The main source of these seeds was purchasing from the stockist (36.4%). Some farmers sourced their seeds from their previous crops (18.2%) and some from other farmers (13.2%). There is also a proportion of farmers (22.7%) that obtained their seeds from their produce buyers. The majority of the farmers (68.2%) reported that they knew the seed rate requirements for planting, while 31.8% did not. The most common variety of French bean grown in the two locations was Amy. This was grown by 44.1 % of the farmers. The other varieties grown were Samantha (20.6%), Julia (17.6%), and Paulista (14.7%). The reasons for choosing the varieties, which they grew, differed among farmers. The majority of farmers (54.2%) choose to cultivate a certain variety because of market demand. Other farmers, 33.3%, cultivate their varieties because of high yield, while 8.3% of the respondents chose their variety because of resistance to pests and diseases. The source of labour in French bean cultivation in the two locations is both from the family and hired. There were clear gender considerations in labour requirements according to farming activity. Land preparation and spraying of pesticides was mainly done by men. On the other hand, weeding and harvesting was done mainly by women. The most common implement used in land preparation was the ox-plough. Some of the farmers with very small acreage used jembes. The implement used for weeding was mainly the panga. Fertilizer application was mainly by hand. Harvesting was also done by hand. Over half of the farmers (59% ), received some form of extension services while the rest, 41% did not. Of those who received the services, 55.5% reported that they received the services from the Ministry of Agriculture or Kenya Agriculture Research Institute (KARl) staff. The rest (44.5%) received the services from firms that had contracted to purchase their produce. The distribution of the services differed in the two divisions. In Nyangati location almost all the farmers got extension services. But in Kangai location only a few of the farmers benefited from extension services while the majority did not. The majority of farmers (63.6%) kept records of their key farming activities, while the rest (27.3%) did not. For those who were contracted to supply their produce to specific firms, these records were frequently inspected by those firms to ensure that the farming practices were in conformity with the market demands. The implication for those who did not keep records was that would may not remember when certain key activities were last performed or when they would be due next. Pests and Diseases The farmers in the two locations experienced a serious problem of pests and diseases. Most farmers reported the presence of several pests and diseases on their farms. The most widely occurring pest was the red spider mite (Tetranychus urticae). This pest, which was reported by 72.3% of the respondents, damages the bean leaves, which then turn silvery and brownish in colour. Infested leaves have cobwebs on the lower surface (MOA, 2000). The second most widely distributed pest was the bean fly. This fly may belong either to the Ophiomyia or Phorbia genus (MOA, 2000). It was a reported to be a problem by 45.5% of the farmers. The adult is a small two-winged insect, which can be seen resting on leaves where it lays the eggs. The damage is caused by the larvae, which eat into the stem. The larvae also feed on the cotyledons of seedlings before or after emergence. Affected plants are yellow, stunted and stems are cracked at the soil level. The other serious pest problem is the bean thrips (Megalothrips spp.). It was reported by 36.4% of the respondents. The damage is caused by the nymphs and adults that eat and damage the bean flowers. The white fly (Bemisia tabaci) was another important pest that was reported by the farmers. It was reported to be a problem

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 109
by 31.8% of the respondents. Cutworms (Agrotis spp.) were another important pest problem that was reported by 13.6% of the respondents. These cause damage by cutting stems of young plants below or above the soil level, and also feed on plant foliage. Other pests of less importance, whose occurrence was reported by less than 10% of the farmers included aphids (Aphis fabae). These cluster on stems, leaves and bean pods. They suck plant sap and cause the plants to stunt. Similarly, crickets were also reported to be a problem by some farmers. The most important disease problem in the two locations was leaf rust. The disease is caused by the fungus Uromyces appendiculatus. It was reported to be a problem by 50% of the farmers. The symptoms of the disease are white spots on the lower leaf surface, which turn red to dark brown after a few days (MOA, 2000). Other diseases, which were reported included, blight. It was reported by 9.1% of the farmers. Bacterial blight in French beans is usually caused by Pseudomonas phaseolicola and Xanthomonas phaseoli. Symptoms of the disease include drying of leaf margins, yellowing and water soaked pods. Another disease that was reported with the same frequency as the blight was Black spot. These could either have been angular leaf spot, a fungal disease caused by Phaeoisareopsis griseola, or Anthracnose, another fungal disease caused by Colletotrichum lindemuthanium. The major control measure against pests and diseases in both locations was pesticide application. All the farmers use pesticides to combat pests and diseases inflicting their French bean crop. A wide variety of chemicals were applied, with the most widely used being that of the brand name dimethoate. Other brands of pesticide included plantvax, karate, omite, sherpa, thiovit, anvil, copper, decis, danadim, benlate and caprocaffaro. The frequency of application of pesticide spray was either once every seven days or once every two weeks. Contracted farmers applied the pesticide according to the instructions of the buyers of their produce. Other farmers applied the pesticides according to the instructions of extension workers from the Ministry of agriculture There was also use of local methods to combat the pests an example of which was use of an ash-based concoction. Post Harvest Handling Most of the farmers from both locations did not have access to cold storage facilities where they could keep their beans after harvest and before sale. Partly because of this reason, there was no storage of the produce after harvest. The produce was taken for sale immediately after harvest. Failure for the produce to be purchased implies that the produce would go to waste, as the beans deteriorate very rapidly in quality if there are no cold storage facilities for their storage. The majority of the farmers did not experience any post harvest losses due to poor storage though a few in each location did. The unsold produce was mainly thrown away in both locations. A very small fraction of the unsold produce was used either for home consumption or for feeding animals. In view of this situation, a big cold storage facility had been constructed near Mwea town. But the modalities of how it was to be used by farmers had not yet been agreed upon. Most of the farmers (72.7%) delivered their produce to buyers at specified purchasing centres. For the rest, the produce was collected from their farms. Only a small proportion of farmers (18.2%) used special delivery vehicles to transport their produce to the purchasing centres. Some (27.3%) used any type of vehicle. The rest used bicycles, cattle or carried the produce on foot.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 110
The beans were packaged in different ways after harvest. Some farmers (38.1 %) packaged the beans in open crates. This is the recommended way of packaging the beans. Others (14.3%) packaged in cartons while 28.6% used polythene bags. There were also a few farmers (9.5%) who transported their produce in bulk. Sales and Marketing The entire French bean crop in the two locations was meant for export. But there were market demand fluctuations affecting sale of the French beans in the export market. There was a distinct season of high market demand from October to April, with a short break during late December and Early January. This high market demand season generally coincide with the winter period in Europe, where most of the French beans are exported. Then there is limited production of fresh vegetables there, leading to higher demand for imported vegetables. During this high demand period, virtually all the beans produced by the farmers were sold for export. During this period farmers were offered 30 to 70 KES per kilogram of beans. They did not experience problems with finding buyers of their produce during this period. After the high season, there follows a season of low market demand, which usually starts in May and lasts up to September. This period coincides with the summer period in Europe. There is more production of fresh vegetables in Europe then, leading to lower demand for imported vegetables. Farmers were offered a much lower price of between KES 10 to KES 40 per kg of beans. Some farmers also experienced problems in getting buyers of their produce. Half of the farmers sold their beans to exporting firms with whom they had a contract. In Kangai location, most of the farmers had a contract with a firm called Frigoken. In Nyangati location, the farmers had contracts with a wider variety of firms. These included Avenue, Sunrise, Homegrown, Everest, Green Venture, and East African Growers. The other farmers sold their produce mainly to middlemen. Those who had contracts with firms were usually assured of a market for the agreed quota of their beans. The prices at which they sold their beans for a certain period would also be agreed upon between the two parties. They were hence shielded from market fluctuations in demand and price of the beans that were experienced by those who sold their produce to middlemen. There were complaints of exploitation by middlemen, particularly during the low demand season. The farmers who sold to middlemen were then forced to accept very low prices for their produce. Less than half of the farmers (40.9%) belonged to some form of co-operative. The rest did not belong to any co-operative. Such farmers may find it more difficult to bargain as a group in purchasing inputs or in the sale of their produce. Main Problems Other than the problem of pests and diseases, there are several other problems, which affect the French bean farmers in the two locations. Lack of capital and inaccessibility to capital were cited by 50.7% of the farmers as major problems affecting their French bean farming. Poor prices and low produce demand, particularly during the low demand season were cited by 22.2% of the farmers as major problems. Other problems which were reported, but each affected less than 10% of the farmers were poor roads and lack of suitable transport for the produce to the market. Other serious problems that were inferred included inadequate extension services. Only about half the farmers had access to regular extension services. Yet the farmers grow a crop which is exclusively meant for export. There are stringent quality requirements affecting various aspects such as pesticide application and maturity at harvest. It may be difficult for the farmers to meet these requirements without access to extension services.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 111
The other problem that was deduced from the respondents was the poor post-harvest handling. It was observed that all the farmers have no access to cold storage facilities. These are essential if any storage or preservation of the beans has to be done at the farm level. Such preservation can also help the farmers to minimize loss of produce that is not immediately sold.
Conclusions Small-scale French bean farmers in Mwea Division practise intensive production using irrigation, fertilizer and pesticides. They produce the beans throughout the year. There is high market demand of their produce from October to April, and relatively low demand from May to September. The main French bean variety grown is Amy. The main problems experienced by these farmers are infestation of their beans with pests and diseases, lack of capital and credit facilities, lack of cold storage facilities and limited technical know how to enable them meet the necessary quality requirements.
Acknowledgement We acknowledge with gratitude the financial support for this study from the African Technology and Policy Studies Network (ATPS).
References CB S (Central Bureau of Statistics), 200 I. Economic Survey 2001. Ministry of Finance and
Planning. Republic of Kenya. CBS (Central Bureau of Statistics), 1999. Statistical Abstract 1999. Ministry of Finance and
Planning. Republic of Kenya. MOA (Ministry of agriculture and Rural Development), 2000. Local and Export Vegetables
Growing Manual.. Horticulture Division. Republic of Kenya. p.82 - 87.
PARTICIPATORY PROMOTION OF HORTICULTURE PRODUCTION IN RIFT VALLEY AND WESTERN KENYA Omunyin* M.E.; Anjichi, V.; Omami, E.; Odhiambo, M.O.1 and Kamar M.J.2 Department of Horticulture, Moi University - Chepkoilel Campus, P.O. Box 1125, Eldoret, Kenya. 1Department of Agricultural Marketing and Cooperatives, F aculty of Agriculture, Moi University. 2Campus Administration, Moi University - Chepkoilel Campus. * Corresponding Author: Michael E. Omunyin E-mail: [email protected]
Abstract The horticultural sub sector is important in the Kenyan economy; however, its potential in the Rift Valley and Western Kenya has not been exploited to the full in terms of domestic and export production. A strategy to exploit horticulture production in Rift Valley and Western Kenya has been evolved as a mechanism for poverty reduction. Elements of the strategy include: assessment of both the potential of horticulture production in the region and farmers' needs; and packaging of identified production and marketing constraints in form of outreach programs and action plans with farmers' participation. Following the consultative process with stakeholders, outreach programs are then implemented through participatory and adaptive community involvement targeting poverty in the process. The key players in this approach include farmers, community based organizatios (CBOs), non-governmental organizations (NGOs), the private sector and the government and its development partners. Recently a public forum on the potential of Horticulture production in Rift Valley and Western Kenya was held in Eldoret attended by farmers and other stakeholders. It was stressed that Moi University, Faculty of Agriculture (MUFA) has an important role to play in the development of Horticulture in the region. Such roles include training endeavours, research and extension and strong University-Industry Linkages. Also pointed out was the fact that although potential exists for increased production of fruits, vegetables and cut flowers, farmers face various constraints, which must be addressed for the potential to be realised.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 112
Moi University and its collaborators embarked on a study to assess the potential of horticulture sub sector in the region starting in the Kerio Valley. This paper presents some highlights of the horticulture forum and preliminary results of the study in some Kerio Valley districts.
SOCIO-ECONOMIC ANALYSIS OF THE CONSTRAINTS TO ADOPTION OF COMMERCIALIZED SMALLHOLDER HORTICULTURAL PRODUCTION SYSTEM IN THE RIFT VALLEY REGION: A CASE OF NANDI AND UASIN GISHU DISTRICT OF KENYA Kipsat, M. J1.; 2Korir, M. K. and Mutai, J. K. Corresponding address: Moi University, Faculty of Agriculture, Department of AMC, P .0. Box 1125, Eldoret).E mail: [email protected] 1M.Phi. Student, Depart. in Agricultural Resource Economics & Marketing (ARE & M) 2 Former DAO, Uasin Gishu District and currently. M. Phi. Student Depart. of ARE&M 3 District Crops Officer, Nandi.
Abstract Despite the importance of the horticultural sub-sector in Kenyan economy, its potential in the Rift Valley has not been fully exploited in terms of domestic and export production. A lot has been done on promoting production and marketing of horticultural products in the region and this has resulted in increased output over the years. Statistics show an 8.45% increase in area under horticultural production between 1995 and 2000 (rising from 30,000 ha in 1995 to 32,771 ha in 2000) in Rift valley Province. Similarly, there were 6.33 % increase in volume (395,028 tons compared to 370,000 tons) and 16.675 increases in value (Ksh. 7.2 billion compared to 6 billion) over the 2 periods in the region (MoA, 2001). A potential for further increase in production exists through increase in production per unit area and increase in area under production. Furthermore the Eldoret International Airport could boost. horticultural export production in the North Rift and Western Kenya regions. The airport facilitates horticultural exports by housing export regulatory firms including HCDA; KEPHIS, Export Promotion Council and the Ministry of Agriculture personnel. Seminars have been organized locally, regionally and internationally to sensitize farmers, investor and other stakeholders on the services offered by the airport. However not much has been done to determine the constraints to adoption of commercialized horticultural production in the smallholder farms in the region. In this study a binomial logit model was used to analyze the influence of farm and farmer characteristics, and the institutional aspects postulated to affect adoption of commercialized horticultural production system among the resource poor farmers in the North Rift region of Kenya. Data were collected from a random sample of 200 farmers selected from Nandi and Uasin Gishu Districts. Primary and secondary data were used. Binomial logit regression was used in data analysis. The regression analysis indicated that among the 10 factors postulated to affect adoption of commercial horticultural production system, six were statistically significant at 95% confidence level. These were gender, availability of credit, access to good road transport to marketing outlet and education level of head of household that positively affected adoption. Others that affected adoption significantly but negatively were farm size and age of the head of the household. In conclusion therefore, targeting women is important in the promotion of commercialized smallholder horticulture production in the North Rift region. Small-scale resource poor farmers should also be encouraged to venture into the horticulture industry through improvement of road transport, access to credit and education of farmers on the importance of horticulture in improving their cash incomes and overall national economic growth.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 113
INCIDENCE OF STRIGA HERMONTHICA INFESTATION DEPTHS ON STRIGA INFECTION INDICES AND MAIZE YIELD COMPONENT Babalola, O.O.1; Odhiambo, G.D.2 and Odulaja, A.1,3 1Department of Botany and Microbiology, University of Ibadan, Ibadan Nigeria 2Kenya Agricultural Research Institute, Kisumu, Kenya 3International Centre of Insect Physiology and Ecology, Nairobi, Kenya. [email protected]
Abstract In different results of S. hermonthica emergence, the main causes to different dates of emergence are infestation depths, accurate understanding of artificial infestation depth can identify gaps in our knowledge in evaluating crop germplasm during artificial infestation. The design was Completely Randomised Design with five replications and the treatment consisted of artificial soil infestation of 50 mg/plot S. hermonthica seeds at depths of 0cm, 5cm, 10cm, 15cm, 20cm, 25cm, 30cm from soil surface (pot rims) and a control with no S. hermonthica infestation. Maize seeds were planted 14 days after infestation and agronomic data were collected. At 77 days after sowing, the harvesting of the experiment was done. The data were analyzed using the Genera linear model procedures of SAS, 1998. Results showed that correspondence between raw and transformed data for emergence counts were good. S. hermonthica emergence time increased with infestation depth with the exception of 0cm infestation zone. Earliest emergence time (31.6 DAP) was recorded at 5cm depth level of infestation. Interactions between time and infestation depth for S. hermonthica were highly significant P<0.0001). Early Striga counts taken provided more evidence for difference among 5cm and 10cm depth. Striga infestation zone of 5cm produced the highest visual rating of striga growth (3.4), visual damages symptoms (7.6), and striga fresh weight (21.80g); smallest plant height (51.36cm), stover dry weight 11.67cm) and root dry matter. At 30cm depth no germination was observed. Infestation zone of 25cm had reduced germination. With the exception of 0cm zone, increasing Striga infestation depth increases plant height (cm) and stover dry weight (g) significantly. Among the traits studies, extremely significant (p<0.0001) are the visual rating of striga growth, visual damage symptoms and the stover dry matter. The un-infested control produced on average 35.41% greater yield than the mean of the infested pots (0-30cm soil zone). The information could be exploited in channeling of water contaminated with S. hermonthica seed and burying of waste soils to further prevent build up of the seed bank.
ROLE OF ORGANIC SOIL AMENDMENTS IN THE MANAGEMENT OF ROOT-KNOT NEMATODES ON OKRA Waceke, J. W. Botany Dept. Kenyatta University, P. O. Box 43844, Nairobi-KENYA E-mail:[email protected] or [email protected]
Abstract Amendment of soil with organic materials is a promising cultural practice in the management of plant parasitic nematodes. Effects of locally available materials namely, chicken manure, goat manure, kale leaves, N-P-K, maize stalks, sawdust and bean stems on control of root-knot nematodes on Okra were evaluated under glasshouse and field conditions. The organic and inorganic materials were incorporated into the soil 14 days before planting and at planting time, respectively, at the rate of 10 and 2g per 1kg of soil, respectively. Results revealed that chicken manure, goat manure, kale leaves, N-P-K, maize stalks and sawdust suppressed root-knot nematodes by up to 75.7, 68.9, 67.2, 90.3, 67.2 and 22.1%, respectively. Bean stems did not suppress pathogenic effects of root-knot nematodes as was revealed by poor plant performance and high disease severity. Gall indices of up to 3.4 on a 0-4 gall rating scale was recorded from plants grown in soils amended with bean stems. Key words: organic amendments, root-knot nematodes, okra
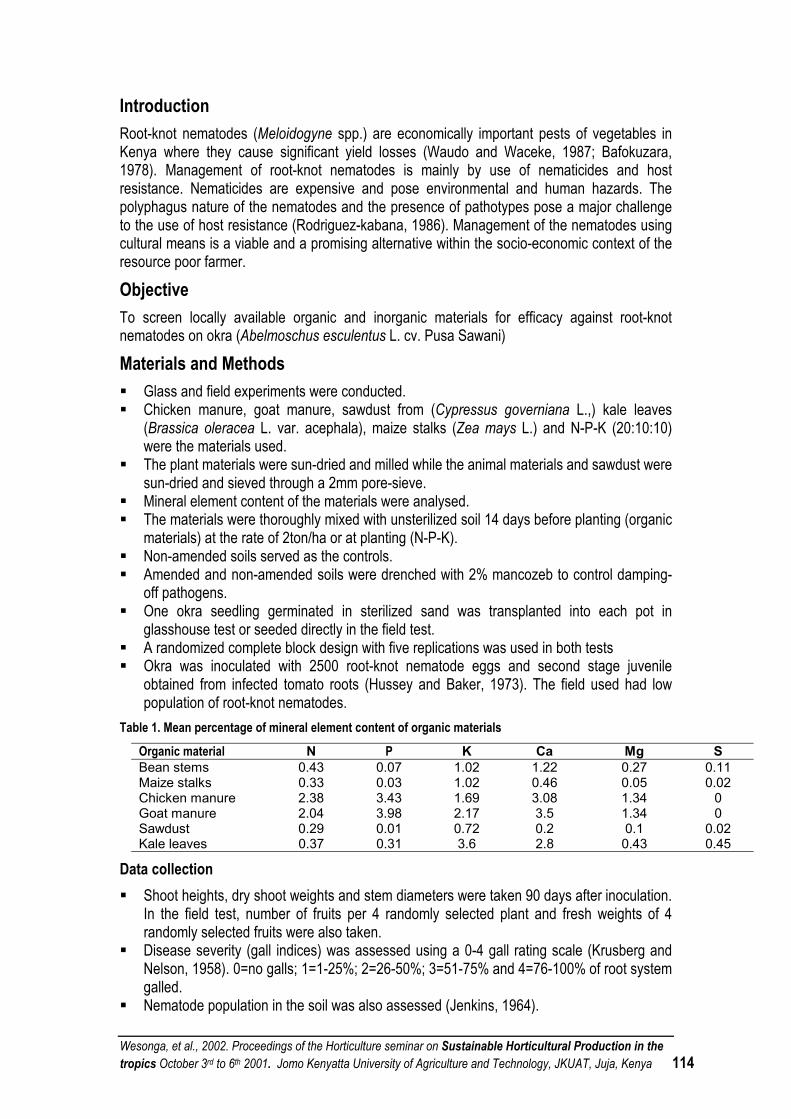
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 114
Introduction Root-knot nematodes (Meloidogyne spp.) are economically important pests of vegetables in Kenya where they cause significant yield losses (Waudo and Waceke, 1987; Bafokuzara, 1978). Management of root-knot nematodes is mainly by use of nematicides and host resistance. Nematicides are expensive and pose environmental and human hazards. The polyphagus nature of the nematodes and the presence of pathotypes pose a major challenge to the use of host resistance (Rodriguez-kabana, 1986). Management of the nematodes using cultural means is a viable and a promising alternative within the socio-economic context of the resource poor farmer.
Objective To screen locally available organic and inorganic materials for efficacy against root-knot nematodes on okra (Abelmoschus esculentus L. cv. Pusa Sawani)
Materials and Methods Glass and field experiments were conducted. Chicken manure, goat manure, sawdust from (Cypressus governiana L.,) kale leaves
(Brassica oleracea L. var. acephala), maize stalks (Zea mays L.) and N-P-K (20:10:10) were the materials used.
The plant materials were sun-dried and milled while the animal materials and sawdust were sun-dried and sieved through a 2mm pore-sieve.
Mineral element content of the materials were analysed. The materials were thoroughly mixed with unsterilized soil 14 days before planting (organic
materials) at the rate of 2ton/ha or at planting (N-P-K). Non-amended soils served as the controls. Amended and non-amended soils were drenched with 2% mancozeb to control damping-
off pathogens. One okra seedling germinated in sterilized sand was transplanted into each pot in
glasshouse test or seeded directly in the field test. A randomized complete block design with five replications was used in both tests Okra was inoculated with 2500 root-knot nematode eggs and second stage juvenile
obtained from infected tomato roots (Hussey and Baker, 1973). The field used had low population of root-knot nematodes.
Table 1. Mean percentage of mineral element content of organic materials
Organic material N P K Ca Mg S Bean stems 0.43 0.07 1.02 1.22 0.27 0.11 Maize stalks 0.33 0.03 1.02 0.46 0.05 0.02 Chicken manure 2.38 3.43 1.69 3.08 1.34 0 Goat manure 2.04 3.98 2.17 3.5 1.34 0 Sawdust 0.29 0.01 0.72 0.2 0.1 0.02 Kale leaves 0.37 0.31 3.6 2.8 0.43 0.45
Data collection Shoot heights, dry shoot weights and stem diameters were taken 90 days after inoculation.
In the field test, number of fruits per 4 randomly selected plant and fresh weights of 4 randomly selected fruits were also taken.
Disease severity (gall indices) was assessed using a 0-4 gall rating scale (Krusberg and Nelson, 1958). 0=no galls; 1=1-25%; 2=26-50%; 3=51-75% and 4=76-100% of root system galled.
Nematode population in the soil was also assessed (Jenkins, 1964).
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 115
Data Analysis Data was subjected to Analysis of Variance (ANOVA). Regressions between plant growth parameters and disease severity were done. Means were separated using Duncan’s Multiple Range Test.
Results Significant differences were detected in shoot heights, dry weight and stem diameters in
the glasshouse test but not in the field tests. Table 2: Mean1 shoot heights (SH), stem diameters (SD), dry shoot weights (DSW), gall indices (GI) and
number of nematodes (NN), 90 days after inoculation, glasshouse test
Treatment SH SD DSW GI2 NN/300c.c Bean stems 3.96ab3 00.52c 4.26bc 3.40d 363.60d Maize stalks 3.85ab 0.35a 3.60b 2.00c 230.20bc Chicken manure 8.42d 0.86a 7.22e 0.80ab 146.80b Goat manure 5.96bc 0.61d 5.97d 1.40bc 183.00bc Sawdust 2.40a 0.32a 2.92a 0.20a 66.20a Kale leaves 4.14ab 0.48bc 3.76bc 1.60bc 208.40bc N-P-K 2.96a 0.43b 3.35ab 2.20c 265.00c Control 4.78ab 0.52c 4.62c 3.80d 459.60e 1 Mean of five replicates 2 Gall index based on a 0-4 gall rating scale (Krusberg and Nelson, 1958). 0=no galls; 1=1-25%; 2=26-50%; 3=51-75% and 4=76-100% of root system galled. 3 Figures followed by the same letter(s) are not significantly different (P>0.05) with Duncan’s Multiple Range Test. Highly significant differences (P<0.001) were, however, detected in disease severity (Gall
indices) and number of nematodes in the soil both in glasshouse and field tests. Plants grown in soils amended with chicken manure, and goat manure were in most case
the tallest, the heaviest and had the lowest disease severity and nematode population in the soil
Plants grown in the sawdust and maize stalk- amended soils were the shortest, smallest and lightest.
Sawdust amended soils supported plants with moderately low disease severity and nematode population while maize-stalk amended soils supported plants with the highest disease severity and nematode population in the soil
Plant performance, disease severity and nematode population in bean-stem, kale leaf or N-P-K amended soils were moderately high.
Significant direct relationships occurred between gall indices and plant growth parameters. Chicken manure, goat manure, kale leaves, N.P.K., maize stalks and sawdust suppressed root-knot nematodes by up to 75.5% (r2 = 0.757), 68.9% (r2 = 0.689), 67.2% (r2 = 0.672), 90.3% (r2 = 0.903), 67.2% (r2 = 0.672) and 22.1% (r2 = 0.221), respectively.
Bean stems did not suppress root-knot nematodes as was revealed by poor plant performance and high disease severity (Gall index of up to 3.4).
Root-knot nematodes reduced plant performance of untreated plants by up to 10.9% (r2 = 0.903).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 116
Table 3: Mean1 shoot heights (SH), stem diameters (SD), number of fruits per plant (NFP), fresh fruit weight (FFW), dry shoot weights (DSW), gall indices (GI) and number of nematodes (NN), 90 days after inoculation, field test
Treatment SH SD NFP FFW DSW GI2 NN/300cc Bean stems 40.79 1.10 4.07 72.31 70.88 1.48c3 194.41bc Maize stalks 30.08 0.96 4.00 58.11 30.96 3.39d 357.63d Chicken manure 32.04 1.00 3.80 60.86 30.14 0.15a 58.91a Goat manure 41.16 1.05 5.3 76.90 45.90 0.32a 86.72ab Sawdust 35.56 0.99 3.15 68.46 39.90 1.07b 123.46abc Kale leaves 53.25 1.06 4.82 66.10 40.30 1.55c 230.32d N-P-K 44.48 1.30 5.27 70.71 45.22 1.32bc 169.32bcd Control 42.95 0.91 3.80 84.75 28.18 0.35a 97.00ab P=0.05 NS4 NS NS NS NS
1 Mean of five replicates 2 Gall index based on a 0-4 gall rating scale (Krusberg and Nelson, 1958). 0=no galls; 1=1-25%; 2=26-50%; 3=51-75% and 4=76-100% of root system galled. 3 Figures followed by the same letter(s) are not significantly different (P>0.05) with Duncan’s Multiple Range Test. 4No significant differences detected among the treatments
Discussion The relatively low nematode population, disease severity and superior plant performance
associated with chicken or goat manure-amended soils could be due; -to sub-lethal or lethal effects of high temperature during the initial phase of decomposition (Hoitink and Fahy, 1986). The infective stage of root-knot nematode, the J2, is very sensitive to temperature changes (Dusenberg, 1987).
-toxic effects of ammonia produced by low C:N ratio materials (Hoitink and Fahy, 1986). Ammonia inhibits hatching of nematode eggs
-improved nutritional status of the plant hence improved host resistance as the manure are rich nutrient sources
-increase in microbial populations in favour of antagonists (Jatala, 1986; Mankau, 1980). The low nematode population, disease severity and plant growth in soils amended with
sawdust might be due N deficiency resulting from N immobilization favoured by high C:N ratio (30:1). Sawdust has a 400:1 C:N ratio (Tisdale et al., 1985).
The high disease severity, nematode population in the soil and poor plant growth associated with beans or maize stalk-amended soils could be due to poor nutrient supplies by the decomposing organic materials
The heavily galled root systems, the high nematode populations and the superior plant performance associated with kale leaf or N-P-K-amended soils might be attributed to high potassium (Table 1) which is reported to improve plant growth and favour root-knot nematode oviposition and rate of development (Marks and Sayre, 1964).
Acknowledgement
I thank Kenyatta University for providing research facilities and the Ministry of Education, Kenya for financing the project.
References Bafokuzara, M. D. 1978. Population dynamics of nematode parasites and evaluation of
damage by Meloidogyne javanica to some common vegetables in Kenya Highlands. M.Sc. thesis, Department of Zoology, University of Nairobi.
Dusenberg, D. B. 1987. Behavioral responses of Meloidogyne incognita to temperature and carbon dioxide. Journal of Nematology 19, 231.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 117
Hoitink, H. A. and Fahy, P. C. 1986. Basis for the control of soil-borne plant pathogens with composts. Annual Review of Phytopathology 24, 93-114.
Hussey, R. S. and Baker, K. R. 1973. A comparison of methods of collecting inocula of Meloidogyne species including a new technique. Plant Disease Report, 57, 1025-1028.
Jatala, P. 1986. Biological control of plant parasitic nematodes. Annual Review of Phytopathology 24, 453-489.
Jenkins, W. R. 1964. A rapid centrifuge-flotation technique for separating nematodes from soil. Plant Disease Report 48, 692.
Krusberg, L. R. and Nelson, L. W. 1958. Pathogenesis of root-knot nematodes to Porto Rico variety of sweet potato. Phytopathology 48, 30-39.
Marks, C. F. and Sayre, R. M. 1964.The effects of potassium on the rate of development of root-knot nematodes Meloidogyne incognita, Meloidogyne javanica and Meloidogyne hapla.. Nematologica 10, 323-327.
Rodriguez-kabana, R. 1986. Organic and inorganic amendments to soil as nematode suppressants. Journal of Nematology 18, 129-135.
Tisdale, S. L., Nelson, W. L. and Beaton, J. D. 1985. Soil fertility and fertilizers. New York. MacMillan.
Waudo, S. W. and Mbugua, J. Waceke. 1987. Responses of spinach (Spinach oleracea L.) to Meloidogyne incognita. East African Agriculture and Forestry Journal, 52, 260-266.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 118
CURRENT FRENCH BEAN PESTS AND DISEASE MANAGEMENT AT MWEA-TEBERE, CENTRAL KENYA Nderitu, J. H.; Olubayo, F.; Waturu, C. N.; Aura, J. and Kasina, J. University of Nairobi, P.O. Box 30197, Nairobi, Kenya.
Abstract A formal survey was carried out in 104 farms during January and February 2001 to identify the major pests and diseases, and use of pesticides on French bean (Phaseolus vulgaris L.) production at Mwea-Tebere, Central Kenya. The main insect pests identified were, in order of importance, bean stem maggot (Ophiomyia spp ), red spider mites (Tetranychus spp), bean flower thrips (Megalurothrips sjostdeti Trybom and Frankliniella occidentalis Pergande), spiny brown bugs (Acanthomia spp.), whiteflies (Bemisia tabaci Gen), podborers and bean aphid (Aphis fabae Scopoli). The main diseases were rust (Uromyces appendiculatus) and bean root rots. Insect pests and diseases caused over 50% losses when not controlled. Pesticides were the sole insect pest and disease control method. Twenty-five pesticides were currently used to control insect pests and diseases compared to thirty-five pesticides used in 1993. Most farmers (80.9%) applied pesticides once per week and over ten sprays applications were done per French bean crop. Ten main pesticides used for control of pests, in order of importance, were L-cyhalothrin (Karate), Alpha-cypermethrin (Fastac), Rogor E (Dimethoate), Cypermethrin (Polytrin), cypermethrin (Sherpa), Deltameltamethrin (Decis), Propargite (Omite), Bifenthrin (Brigade), Tetradifon (Tedion) and Diazinon. The Main fungicides for control of diseases, in order of importance, were Sulfur (Thiovit), Copper oxychloride (copper), Propineb (Antracol), Penconazole (Pencozeb), Oxycaboxin (Plantvax), Hexaconazole (Anvil), Dimethomorpho+Mancozeb (Acrobat) and Mancozeb (Dithane M-45). These pesticides are of low toxicity ratings and farmers do not expect to exceed Maximum Residue Levels (MRLs) on fresh pod produce if the recommended pre-harvest intervals of these pesticides are observed. The shift to use of pesticides of low or no residues on the produce will enable the farmers to continue to export French beans to the European market.
Introduction French beans (snap beans) is a major vegetable crop grown in Kenya mainly for export market as fresh green pods. It accounts for volume of more than 20% of horticultural export (Kibanga, 1996). The national production levels have been on increase year after year. In 1997, 1998 and 1999,the production figures were 16,510, 23,60 and 30,680 tonnes, respectively (Anon, 1999). The fresh green pods are important as they contribute to human nutrition by providing particularly ascorbic acid, proteins, fibre and minerals (particularly iron). French beans is normally grown by small-scale farmers who have small land holdings. These growers seek to maximize their yields and satisfy the aesthetic values of the market through intensive management regimes, with irrigation and rigid crop protection practices, which rely heavily on chemical pesticides (Nderitu et al, 1997). Survey carried out by Nderitu et al (1997) in Mwea-Tabere found that farmers were using 35 pesticides regularly and employed up to 15 foliar sprays during a single crop cycle. A survey was conducted to describe and evaluate the extent of current pests and pesticides use in the French bean production at Mwea-Tabere, central Kenya.
METHODOLOGY The survey was carried out at Mwea-Tabere in Central Kenya. The area is classified as lower midland (LM4) zone with a mean temperature range of 22.0-22.2 °c (Jetzold and Schmidt, 1983). The soil is mainly black cotton type. The first rains normally starts at the end of March and the second rains start end of October. The French beans are grown throughout the year but peak production, aimed at the export market, is between September and April.
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 119
A formal survey by use of questionnaire was carried out during January and February 2001 in 104 farms. Farmers were selected systematically, with a distance of a quarter Kilometer between farms. The survey was carried out primarily to identify the major insect pests and disease loses they cause, and pesticides used by farmers on French bean production. This covered the types of pesticides used, frequency of application, use of appropriate or recommended pesticides for specified insect pests and diseases
Results Among the crops grown in Mwea-Tabere, French beans, was the first priority crop.Various production agronomic activities are carried out at different times of the French bean crop season as summarized in Table 1. Most farmers planted in the month of December, planted two crops per season with two weeks interval. Table 1: French bean production practices
Activity Percent farmers No. of Plantings 2 (40.0%), 1 (24.2%) 3 (23.2%), 4 (9.5%), 5 (3.2%) Planting interval (weeks) 29(1.55), 3 (24.2%), 1 (2.2%), 6 (1.1%), 7 (1.1%), 9 ( 1.1%) Month of planting December 958.9%), January (30.5%), November (8.4%), September (1.1%) Key: ( )--- percent farmers
Farmers listed pests and diseases at each stage of development of French bean crop as summarized in Table 2. Table 2: French bean insect pests and diseases at each developmental stage of he bean crop
Pests At seedling Before flowering
During flowering
After flowering Near maturity
Insects/mites Bean fly, white fly, mites, cutworms, crickets
Thrips, white fly, mites, aphids, caterpillars, bean fly, cutworms, brown bugs
Thrips, aphids, mites, caterpillars, bean fly, white fly, brown bugs
Caterpillars, thrips, aphids, pod sucking bugs, mites, pod borers, white fly, bean fly
Caterpillars, mites, pod sucking bugs, thrips, pod borers, bollworms, white fly, aphids
Diseases Bean root rot, halo blight, rust
Rust, halo blight, leaf spot
Rust, halo blight, leaf spot
Rust, halo blight, leaf spot
Rust, halo blight, leaf spot
Farmers listed the most important pests and diseases caused by the insect pests and diseases (table 40. They said that both pests and diseases caused more than 50% loss to the final crop yield. Table 4: yield loss due to pests and diseases
% Yield loss Pests Diseases 1-25 1.1 1.1
25-50 4.4 7.7 >50 94.5 91.2
Of the various control measures undertaken to control the French bean pests and diseases, farmers were able to rank them as to how often they relied on each (Table 5). All the farmers used pesticides for disease and pest management. Table 5: Control methods used by the farmer for insect pests and diseases
Control method Rank order of importance Chemical 1 Cultural 2 Varietal 3 Physical/mechanical 4 Biological 5
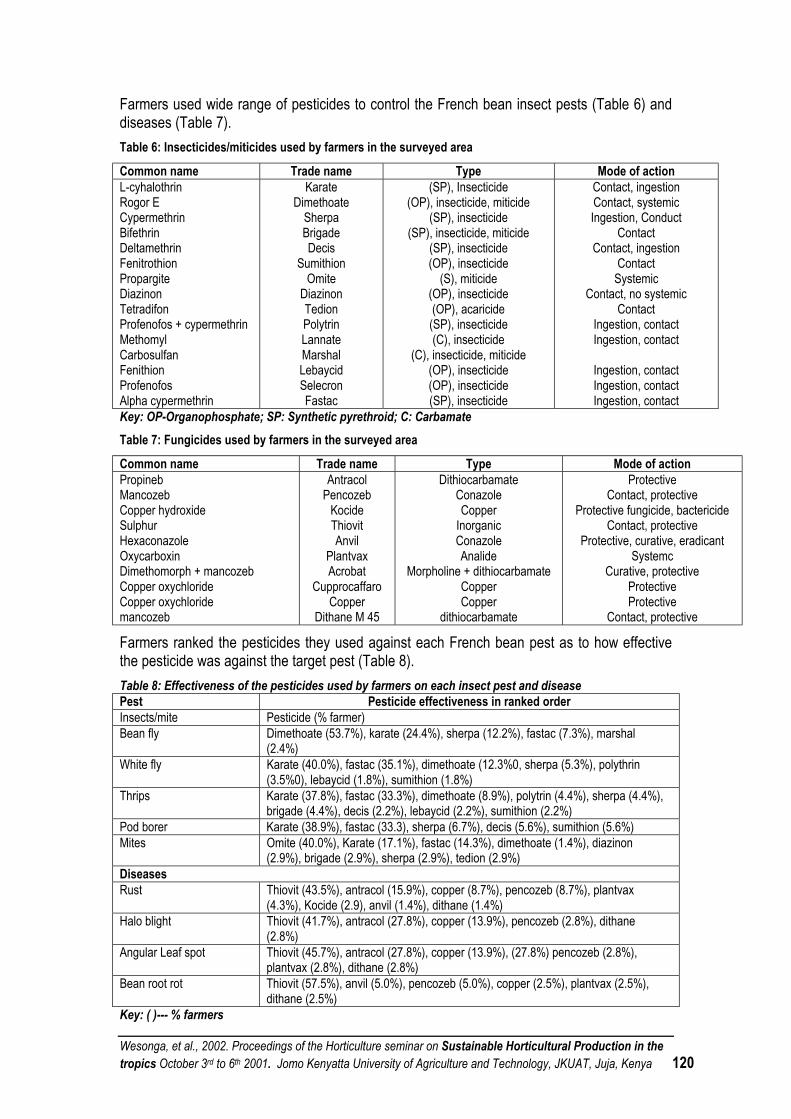
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 120
Farmers used wide range of pesticides to control the French bean insect pests (Table 6) and diseases (Table 7). Table 6: Insecticides/miticides used by farmers in the surveyed area
Common name Trade name Type Mode of action L-cyhalothrin Rogor E Cypermethrin Bifethrin Deltamethrin Fenitrothion Propargite Diazinon Tetradifon Profenofos + cypermethrin Methomyl Carbosulfan Fenithion Profenofos Alpha cypermethrin
Karate Dimethoate
Sherpa Brigade Decis
Sumithion Omite
Diazinon Tedion Polytrin Lannate Marshal Lebaycid Selecron Fastac
(SP), Insecticide (OP), insecticide, miticide
(SP), insecticide (SP), insecticide, miticide
(SP), insecticide (OP), insecticide
(S), miticide (OP), insecticide (OP), acaricide (SP), insecticide (C), insecticide
(C), insecticide, miticide (OP), insecticide (OP), insecticide (SP), insecticide
Contact, ingestion Contact, systemic Ingestion, Conduct
Contact Contact, ingestion
Contact Systemic
Contact, no systemic Contact
Ingestion, contact Ingestion, contact
Ingestion, contact Ingestion, contact Ingestion, contact
Key: OP-Organophosphate; SP: Synthetic pyrethroid; C: Carbamate Table 7: Fungicides used by farmers in the surveyed area
Common name Trade name Type Mode of action Propineb Mancozeb Copper hydroxide Sulphur Hexaconazole Oxycarboxin Dimethomorph + mancozeb Copper oxychloride Copper oxychloride mancozeb
Antracol Pencozeb
Kocide Thiovit Anvil
Plantvax Acrobat
Cupprocaffaro Copper
Dithane M 45
Dithiocarbamate Conazole Copper
Inorganic Conazole Analide
Morpholine + dithiocarbamate Copper Copper
dithiocarbamate
Protective Contact, protective
Protective fungicide, bactericide Contact, protective
Protective, curative, eradicant Systemc
Curative, protective Protective Protective
Contact, protective
Farmers ranked the pesticides they used against each French bean pest as to how effective the pesticide was against the target pest (Table 8). Table 8: Effectiveness of the pesticides used by farmers on each insect pest and disease Pest Pesticide effectiveness in ranked order Insects/mite Pesticide (% farmer) Bean fly Dimethoate (53.7%), karate (24.4%), sherpa (12.2%), fastac (7.3%), marshal
(2.4%) White fly Karate (40.0%), fastac (35.1%), dimethoate (12.3%0, sherpa (5.3%), polythrin
(3.5%0), lebaycid (1.8%), sumithion (1.8%) Thrips Karate (37.8%), fastac (33.3%), dimethoate (8.9%), polytrin (4.4%), sherpa (4.4%),
brigade (4.4%), decis (2.2%), lebaycid (2.2%), sumithion (2.2%) Pod borer Karate (38.9%), fastac (33.3), sherpa (6.7%), decis (5.6%), sumithion (5.6%) Mites Omite (40.0%), Karate (17.1%), fastac (14.3%), dimethoate (1.4%), diazinon
(2.9%), brigade (2.9%), sherpa (2.9%), tedion (2.9%) Diseases Rust Thiovit (43.5%), antracol (15.9%), copper (8.7%), pencozeb (8.7%), plantvax
(4.3%), Kocide (2.9), anvil (1.4%), dithane (1.4%) Halo blight Thiovit (41.7%), antracol (27.8%), copper (13.9%), pencozeb (2.8%), dithane
(2.8%) Angular Leaf spot Thiovit (45.7%), antracol (27.8%), copper (13.9%), (27.8%) pencozeb (2.8%),
plantvax (2.8%), dithane (2.8%) Bean root rot Thiovit (57.5%), anvil (5.0%), pencozeb (5.0%), copper (2.5%), plantvax (2.5%),
dithane (2.5%) Key: ( )--- % farmers

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 121
Farmers were interviewed on how they make decision on when to spray the pesticides (Table 9 and Table 10). It was found that most farmers had routine spraying and that few did monitoring before spraying. Table 9: Farmers decision on when to spray
Decision to spray Percent farmers Scheduled treatments 75.8 Pest abundance 12.6 Pest damage 10.5
Table 10: farmers’ decision for next spray
Decision for next spray Percent farmers Routine schedule 92.6 Monitoring 6.3 Others 1.1
Farmers recorded the spray intervals preceding the first spray (Table 11) and how often they sprayed the French bean crop per season (Table 12). It was found that most farmers sprayed the crop weekly and had 8-10 sprays per one crop season. Table 11: Spraying interval for the pesticide
Spray interval (days) Percent farmers 2 3.2 3 3.8 5 1.1 7 80.8
10 1.1
Table 12: Frequency of insecticide application per season
Insecticide application per season Percent farmers 1-4 2.3 5-7 11.9
8-10 83.5 >10 2.3
Discussion The insect pests and diseases on French bean in Kenya are controlled mainly with pesticides as elsewhere in the world (Cardona and Corrales, 1992: Velasquez and Prada, 1992; Anyango et al. 1989). This is principally due to the demand by the export market for high quality produce (Nderitu et al.1997). Farmers interviewed expressed sufficient knowledge of insect pests and disease infesting French beans, which they encountered in the field. Of these pests and diseases, they were aware of the most serious and thus actively participated in the control of these pests in order to have maximum protection of the crop and ensure good yields while meeting the quality standards of unblemished pods. All the farmers used chemicals as the major control strategy for insect pests and diseases while small number of them supplemented the chemicals with cultural, physical and mechanical practices. Biological control was not a priority for use to control the pests by the farmers. Most of the pesticides were used against a wide range of pests and diseases on the French bean crop. The range of pesticides used in the area has reduced to 25 compared to survey done in 1993, which recorded 35 pesticides (Nderitu et al.1997). Most of these pesticides have been accepted by PCPB for use on French bean crop while others are not recommended but are used by farmers because they are cheap, effective and easily available in the area.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 122
Farmers used some few chemicals to target wide range of insect pests and diseases. Karate and Fastac were highly used to control the insect pests while Thiovit and Antracol were used to suppress most of the diseases. These pesticides have short pre-harvest interval and may not exceed the maximum residue levels. The cost of pesticides is always on increase and therefore farmers ought to reduce pesticide use. Farmers were observed to do prophylactic spraying and very few did pest monitoring. Monitoring of pest is an important factor as it enables the farmer to apply the chemical only when it is necessary. This can reduce the number of applications of the pesticides per season and therefore minimize the pesticide cost and pesticide residues. While chemical use may not be abandoned there is need to incorporate other control strategies which can help also reduce the pesticide application, minimize resistance build up and reduce dangers to the farmers during pesticide application in the field. Reliance on one chemical by the farmer to control the pests for the whole season could result in resistance build up by the pests and thus it is necessary to inform farmers of these dangers and how they can overcome them. It is necessary to make farmers aware of the actual losses caused by each pest and the effectiveness offered by the other control strategies. There is also need for researchers to collaborate with farmers and come up with control thresholds of each pest as this will enable the farmer to understand safe use and appropriate pesticide handling and application procedures.
References Anon (1999) Export market statistics figures for fresh fruits, vegetables, and cut flowers. 1997 -
1999. Horticultural Crops Development Authority, Kenya. Anyango, J..J., Ochiel, G.R.S., Alembi, D.K, Pete, S.O.K. and Kimutai, D.C. (1989) Survey,
identification and control of French bean pests with major emphasis on bean thrips in Kenya. NARL Annual report, KARl., Kenya, 1989. PP 109-113 Cardona, C, and Pastor-Corrales (1992) Strategies for management of pests and diseases of Snap beans in Latin America. In: Henry, G and Jansen, W (eds.) Snap beans in developing world: Proceedings of an international conference held in Cali. Colombia. 16-20 October 1989.CIAT, Cali, Colombia, 366pp.
Jaetzold, R. and Schmidt, H. (1983) Farm management handbook of Kenya. Crop development division, Ministry of Agriculture, Kenya. PP 179
Kibanga, C. (1996) Overview of Kenyan horticultural industry. 11orticulturalj. 4: 3-4 Nderitu, J.H, Anyango, .J.J and Ampofo, J.K.O (1997) A survey of insect pests and farmers.
control measures on Snap beans in Kirinyaga district, Kenya. CIAT , Occasional publications series. No.23 PP 16.
Velasquez, .J.G. and P. Prada (1992) Snap bean pests and diseases in Sumaz, Colombia: their present status and implications. In: Henry, G and Jansen, W (eds) Snap beans in developing world: Proceedings of an international conference held in Cali. Colombia. 16-20 October 1989.CIAT, Cali, Colombia, 366pp.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 123
APPLICATION OF REMOTE SENSING TECHNIQUES IN MANAGEMENT OF BIOTIC CROP STRESS Losenge, T.1 and Hau, B.2 1Department of Horticulture, JKUAT, 2Institute for Pant Diseases and Plant Protection, University of Hanover.
Abstract Remote sensing techniques is a method of acquiring and interpreting data from plants without any physical contact between the measuring devices and the plant. Various types of remote sensing instruments that measure electromagnetic radiation that is reflected or emitted from the plant canopies have been developed. In a series of field experiments, we investigated the application of remote sensing in disease identification and quantification. Reflected, transmitted and absorbed radiation were quantified using a hand held multispectral radiometer that is equipped with sensors that can estimated reflected and total incidence radiation at the range of 460 to 810 nanometers. A ceptometer was used to measure radiation transmitted through the canopy. From the results, the estimation of plant growth indicators such as the LAI and plant yield can be accurately achieved by utilising ratios such as the normalised difference vegetation indices (NDvi) and the Perpendicular vegetation indices (Pvi) that takes into account the variation in the zenith angle and the reflectance from the soil background beneath the canopy, respectively. Remote sensing especially by use of simple hand held instruments such as the multispectral radiometers and the ceptometers could allow non-destructive analyses of the healthy and stressed plant canopies. Current remote sensing techniques cannot identify the specific cause of stress in crops; however, we were able to detect and quantify the intensity of the stress early in the infection process before disease symptoms and signs become apparent.
USE OF NATURAL PESTICIDES FOR PEST AND DISEASE MANAGEMENT IN VEGATABLE PRODUCTION UNDER SMALL SCALE FARMS Moshi, C. and Nderitu, J. H. Siekuti Self Group, P. 0. Box 28, Tiliki / Crop protection Department, University of Nairobi P. 0. Box 30197, Nairobi
Abstract Preparations, uses and efficacy of the following natural pesticides on control of pests and diseases on vegetables will be discussed: garlic, onion, lantana, neem, spider weed. eucalyptus, pyrethrum, marigold, tomato, papaya, ash, chillies, tithonia, coriander, sour milk, cow urine, cowdug and tephrosia. Some of the pests killed or repelled are aphids, grasshoppers, caterpillars, ants, termites, mites, cutworms and diamondback moth. Some are also effective on control of nematodes and fungal diseases. There is need for scientists to confirm farmers findings.
SCREENING OF THREE STRIGA HERMONTHICA STIMULATING FLUORESCENS BACTERIA FOR PHYTOTOXICITY IN COWPEA Babalola, O.O.1; Sanni, A.I.1 and Odhiambo, G.D.2 lDepartment of Botany and Microbiology, University of Ibadan, Ibadan Nigeria 2Kenya Agricultural Research Institute, Kisumu, Kenya
Abstract The obligate root parasitic flowering plant Striga hermonthica (Del.) Benth devastates cereal crops in Africa. Constrains to maize production due to S. hermonthica, could be reduced by the use of trap crops, the stimulatory effect of which could be augmented by rhizobacteria. Virtually all studies on S. hermonthica control categorically state that no single method will be effective. In this study three cowpea varieties were used as trap crops on which the effect of three Pseudomonas strains introduced to the

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 124
rhizosphere of cowpea at planting was studied. Number of days to cowpea flowering was noted and at harvest, data were collected on pod characteristics and biomass. The data were subjected to ANOVA and means were compared using Tukey's Studentized Range Test. Results obtained revealed that the bacteria were significantly influenced by cowpea varieties. Significant varietal differences were found for pod weight, pod length, and pod number. Under the experimental conditions used, the Pseudomonas strains are not deleterious to cowpea plants. The bacteria introduced at planting affected the three tested cowpea varieties with better performance over the uninoculated control. Pseudomonas inoculation significantly increased (≥≥≥≥4.22%) cowpea fresh biomass. Average pod weight (g) with bacteria treatment was 37.97 for isolate 8MR5, 34.38 for isolate 1683 and 27.46 for isolate 883. Uninoculated control had an average weight of 27.98. Bacteria promoted a significant increase in pod weight (≥≥≥≥30.89%) and improved pod number (≥≥≥≥20.54%) and pod wall thickness (≥≥≥≥7.33%). Number of days to cowpea flowering and length of pod at harvest were less consistent. Interactions of variety x bacteria were not significant for any of the measured yield components.
Expression of resistance in. Brassica rapa sp. Infected with Plasmodiophora brassicae Wor Koech Joel Moi University, Chepkoilel Campus, Botany Department, P.O. Box 1125, Eldoret. E- mail: [email protected]
Abstract A study was undertaken to investigate the expression of resistance and the stage it operates in a resistant line of B. rapa infected with P. brassicae. A comparison was made using a susceptible cultivar in order to determine the stage in the life cycle when the resistance mechanism is expressed. The resistance mechanism in two lines with different inheritance of resistance was also compared. Light and electron microscopy was used to examine the expression of resistance in B. rapa. P. brassicae was able to invade the cortex, stele and vascular tissues of the resistant line as in susceptible lines up to approximately 15 days after infection. Resistance thus appears to be expressed at a late stage in infection. After 15 days, plasmodia within susceptible line developed rapidly and cell division and expansion was initiated, while in the resistant line, plasmodia could not be found. Resistance expression in the two resistant lines with different inheritance of resistance revealed a similarity in the infection process and in the early stages in the development of the pathogen. In both the resistant lines, the pathogen failed to develop beyond the earliest multinucleate stage.
Introduction Most previous work has concentrated on susceptible host cultivars infected with P. brassicae while little has been done on the pathogen in the resistant varieties. There are two phases in the susceptible cultivars, the primary phase in the root hairs and a secondary phase in the cortical region of the root system. Tommerup and Ingram (1971) and Ingram and Tommerup (1972) gave evidence that P. brassicae is haploid during most of its life cycle and that meiotic division takes place within the plasmodia of the secondary phase just before the formation of the resting spores. The severity of the club root phase is not closely related with the frequency of root-hair infection (Naiki, Tanahashi & Kageyama, 1984). There is little information about infection of the secondary phase. The possibility of root hair re-infection by the secondary zoospores of P. brassicae has been suggested (Karling, 1968; Macfarlane, 1952). Naiki, Kawaguchi and Ikegami (1984) showed that root hair infection occurred in healthy seedlings grown around the infected plant used as an infection source, in which the fungus occurred only in zoosporangial stage, indicating that re-infection might also occur. The secondary stage in the life cycle is associated with hyperplasia and hypertrophy in cortical cells, leading to clubroot formation. Little is known about the phase development following entry of secondary zoospores into the epidermis. Penetration of the cortical cells of the roots by zoosporangial zoospores has never been observed, but it is believed that these zoospores

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 125
migrate from the root hairs to the epidermal and cortical cells, where they cause re-infection (Colhoun, 1958). In a resistant variety of B. rapa, Dekhuijzen (1979) observed the presence of zoosporangia with secondary zoospores in the root hairs 9 days after inoculation; 2 to 4 days later, he noted a large number of dead host cells in the outer cortical layer whereas no apparent changes were found in the inner cortex. He suggested that a hypersensitive host reaction terminated growth of P. brassicae.
Objectives 1. To study the expression of resistance. 2. To determine the stage in the life cycle when the resistance mechanism is expressed. 3. To compare the resistance mechanism in two lines with different inheritance of resistance.
Materials and Methods Host Plants Two hosts lines, a susceptible B. rapa cv. Granaat and a resistant line 97 of B. rapa were selected. 80 seeds of each line were sown (one per pot) in a peat-sand mixture (1. 1 by volume). Inoculation A single spore isolate of the pathogen was used to inoculate 48 seedlings of each of the two lines. Inoculation was done by pipetting 1 ml spore suspension containing 9 x 106spores into the rhizosphere of each of the plants. Control plants comprising 48 seedlings of each line were left uninoculated. Sampling Sampling was done from the 10th day after inoculation and subsequently every two days till the 42nd day after inoculation. 4 inoculated and same number of non-inoculated plants was sampled. Segments were made approximately 1 cm in length, starting from the hypocotyl region. The sections were immediately fixed in gluteraldehyde in eppendorf tubes at room temperature overnight after which were embedded immediately. Root hair sections were made and stained with 1-% aceto-carmine. Tissue Embedding Freshly prepared root tissues were fixed in 2% gluteraldehyde/0.05M sodium cacodylate buffer and post fixed using 0.05M cacodylate buffer and 1% osmium tetroxide/0.05M sodium cacodylate buffer at pH 7. 0. After rinsing, they were immersed for 15 minutes in 25%, 50%,70% and 90% ethanol distilled water, and finally 100% ethanol before leaving overnight at room temperature. Tissues were then subjected to a sequence of resin changes involving acrylic resin 1 : 1 ethanol; acrylic resin 3 : 1 ethanol and 100% acrylic acid for two hours each. Tissues were further transferred to pure acrylic resin and rotated for 6 hours before being transferred into gelatin capsules (size 00) filled with pure acrylic resin and then allowed polymerizing for 12 hours at 60°C in the oven. Sectioning for Light and Electron Microscopy Semi-thin sections (1 urn in thickness) were cut with glass knives on an LKB ultramicrotome. The best sections were used to prepare permanent slides for light microscopy examination. Sections ranging from 70-90 nm in thickness were prepared

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 126
Staining for Light Microscopy The triple stain method described by Humprey & Pittman (1974) was used to stain resin embedded root tissues for light microscopy examination. The prepared sections, each measuring 1 urn in thickness were transferred to a drop of water on a slide and dried in an incubator at 70°C. A few drops of methylene blue azure II was then applied onto the dried sections and left for 40 minutes. The sections were then washed with sterile water and again dried before staining for 3 minutes in basic fuchsin. The slide was again washed with sterile water and dried before mounting with emmersion oil. Staining for Electron Microscopy The ultra-thin sections were mounted on grids, left for a couple of minutes to dry , then stained in 2% aqueous uranyl acetate (10 minutes). Excess uranyl acetate stain was washed off in sterile water and the second stain, lead citrate, was applied for 4 minutes.
Results Root Hair Infection Zoosporangia were observed within the root hairs in both the susceptible B. rapa cv Granaat and the resistant line 97 of B. rapa. They were visible from the first sampling 10 days after inoculation, but only a few root hairs contained the zoosporangia and many of them remained uninfected. There was variation in the number of zoosporangia observed in the infected root hairs, which ranged between I and 10. At this stage there was no obvious differences between the two lines. Primary plasmodia, as reported by Dekhuijzen (1979), were not observed. Root Infection From 10 days after inoculation, young plasmodia were observed mainly in epidermal cells in both the susceptible and resistant lines. At 12 days, the young plasmodia could be observed in epidermal as well as cortical cells in both cases. At 15 days, the first multinucleate plasmodia with abundant lipid droplets were visible in the susceptible line while the young plasmodia in the resistant line were still undeveloped and could be seen in vascular tissues. At 18 days, the multinucleate plasmodia had large number of nuclei and lipid droplets in the susceptible line while in the resistant line, they became dense and showed signs of degeneration. At 21 days, the plasmodia in the resistant line had completely disappeared and some dead cells could be seen while that in the susceptible line had plasmodial cytoplasm fissured around each nucleus and large lipid droplets were visible and the entire cells covered. Expression of resistance in two lines with different inheritance of resistance The results revealed a similarity in the infection process and in the early development of the pathogen. In both lines, infection and penetration resulted in the formation of zoosporangia in the epidermal cells. This was followed by the development of young plasmodia that penetrated into the cortex and the stelar region of the root. Some of the plasmodia were also observed in the vascular tissues in both lines. The pathogen in both lines resulted in a brief development for 15 days when the spherical plasmodia transformed into a small irregularly shaped plasmodium, which soon started to get dense and distorted. This stage was preceded by a gradual disintegration of the pathogen, host cell disruption and finally death. In both the resistant lines, the pathogen failed to develop beyond the earliest multinucleate stage.
Discussion The presence of the primary zoosporangia were observed within the root hairs shortly after inoculation. This is an indication that the first process of infection occurs within the root hairs. As mentioned by Naiki et al., (1984), the secondary zoospores released by the zoosporangia in

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 127
the root hairs may be able to re-infect the root hairs and repeat the zoosporangial stage within the root hairs or penetrate and infect the epidermal cells. Since the secondary zoospores were only observed in the epidermal cells, it is highly probable that the movement of the pathogen into the cortical cells, stelar and vascular tissues could have occurred in the young plasmodial stage. These appear to be the spore-like structures within the cortex described by Dekhuijzen (1976) and considered to represent secondary zoospores. It was possible to observe all the stages in the life cycle of the pathogen within the inoculated susceptible line, but not in the resistant line. The failure of the pathogen to develop beyond 15 days after inoculation may be an indication of a process that involve inhibition of the multiplication phase. While the plasmodium in susceptible line continued to divide, it seems to fail to do so in the resistant line beyond the 15th day after inoculation. Inhibition mechanism within the cell may prevent the growth of the pathogen after this time. A hypersensitive reaction of the host cell can be one cause of this mechanism. Disruption of the cell cytoplasm and its subsequent disintegration appear to trigger the same process in the pathogen. Unlike in the susceptible line, the infected cell of the resistant line tends to decrease in size and appear distorted. Plasmodia in the cortical cells were seen to undergo a process of disintegration starting from the outside, a portion in contact with the host and gradually disintegrating towards the inside of the plasmodium. This process could therefore be attributed to an inhibition mechanism within the host cytoplasm which as it occurs tends to inhibit the growth of the pathogen and cause its disruption starting from the outside. This observation has also been supported by the report of Dekhuijzen (l979) where he showed that resistance was manifested in the cortex of the main root 10 to 24 days after inoculation when necrotic host cells were found and attributed it to hypersensitive type of resistance occurring in the cortical cells that arrested the growth of the plasmodia. Although the inheritance of resistance differed in the two resistant lines of B. rapa, the expression of resistance was similar. This means that the expression of resistance to P. brassicae is similar irrespective of the number of genes involved.
References COLHOUN, J. 1958. Clubroot disease of crucifers caused by Plasmodiophora brassicae Wor.
Kew. Commonwealth Mycological Institute. Phytopath. Pap. 3, 1- 108 DEKHUIJZEN, H. M. 1976. Microscopical studies on the cortex and medullary rays of Brassica
campestris var. rapa infected with Plasmodiophora brassicae. Phytopath. Z 87, 171- 186.
DEKHUIJZEN, H. M. 1979. Electron microscopy studies on the root hairs and cortex of a susceptible and a resistant variety of Brassica campestris infected with Plasmodiophora brassicae. Neth. J PI. Path. 85, 1 - 17.
HUMPHREY, C. D. & PITTMAN, F. E. 1974. A simple methylene blue-azure II - basic fuchsin stain for epoxy - embedded tissue sections. Stain technology 49,9 - 14.
INGRAM, D. S. & TOMMERUP, I. C. 1972. The life history of Plasmodiophora brassicae Wor. Race 6. Eucarpia Cruiciferae Newsletter. 3,27.
KARLING, J. S. 1968. The Plasmodiophorales, 2nd ed. Hafner Publishing Company, New York.
MACFARLANE, I. 1952. Factors affecting the survival of Plasmodiophora brassicae Wor. in the soil and its assessment by host test. Annals of Applied Biology 39,239 - 256.
NAIKI, T. TANAHASHI, K. KAGEY AMA, K. 1984. Ibid 50, 211- 215. NAIKI, T.; KAWAGUCHI, C. & IKEGAMI, H. 1984. Root hair re-infection in Chinese cabbage
seedlings by the secondary zoospores of Plasmodiophora brassicae Wor. Annals of the Phytopathological Society of Japan. 50,216 - 220.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 128
TOMMERUP, I. C. & INGRAM, D. S. 1971. The life cycle of Plasmodiophora brassicae Wor. in Brassica tissue cultures and in intact roots. New Phytol. 70,327 - 332.
VEGETATIVE PROPAGATION OF MACADAMIA NUT (MACADAMIA INTEGRIFOLIA, (M. INTEGRIFOLIA * M. TETRAPHYLLA) HYBRIDS). Lucy Gitonga*1, W. Nyakundi2, S.T. Ruto1, B. Watiki1 and F. Balozi1, E. Takayama3
1Kenya Agricultural Research Institute (KARI); 2 Kenya Plant Health Inspectorate Service (KEPHIS); 3Japan International Cooperation Agency (JICA)
Abstract. Macadamia is highly heterozygous and therefore production of true-to-type macadamia seedlings would depend entirely on vegetative propagation. Experiments have been conducted for six years on the performance of macadamia seedlings grafted with scions from recommended varieties (KRG-15, EMB-1, MRG-20 and KMB-3) using four different grafting methods namely; top wedge (cleft), side wedge, splice and veneer. Seedlings ranging from 9-12 months were grafted with scion material that had been collected and stored at 4°°°°C for 1 wk prior to grafting. Grafted seedlings apart from those grafted with veneer method were tended in tunnels in greenhouse maintained at 25-28°°°°C, 80-90% relative humidity and 50% light intensity. Veneer grafted seedlings were tended in an open area under direct sunlight. All grafting methods were found to be successful but graft taking ratios varied slightly with varieties with each method. However, on average, top wedge grafting method gave the highest takes % (90-100) followed by splice (80-100) and side wedge (70-100) and finally veneer 40%. Top wedge was hence recommended for mass propagation of clonal macadamia seedlings augmented with splice and side wedge methods. Veneer grafting method, having not required greenhouse facilities was recommended for low-income nursery operators.
Introduction Macadamia nut (Macadamia integrifolia and M. tetraphylla) was introduced in Kenya in 1946 and later seedling progenies were distributed to farmers in various parts of Kenya. However, most of this material was low yielding (5-15 kg/tree/season), of low quality (<72% oil content) and had a long juvenile period (8-12 years before bearing). After conducting a national survey, high yielding trees with good quality nuts were selected. Due to the high heterozygosity of Macadamia (75% outcrossing) (Ondabu et al., 1997) clonal material of such selected varieties must be vegetatively produced and hence the need for an easy, fast and economical vegetative propagation method. Grafting, which involves the joining of two plant parts combining different desirable characteristics has been found to be successful in various fruit trees (Alexander, 1987; Hartmann and Kester, 1983; Hartmann et al., 1990; Nyakundi and Gitonga, 1993).
Project objective To evaluate four grafting methods namely top wedge, splice, side and veneer on their success in propagating clonal macadamia varieties.
Materials and methods Freshly harvested mature seeds were germinated on sand for 2 months. At the 4-6th hard leaf stage they were individually transplanted into polybags filled with about 2 kg of soil medium (soil: manure: compost: sand - 10:3:3:1) supplemented with 1.5% compound fertilizer (NPK). Seedlings were maintained under a shade (40-50% light reduction) for 2 months with regular watering and fortnightly sprays with Dithane M-45 to protect against fungal diseases. Regular

Wesonga, et al., 2002. Proceedings of ttropics October 3rd to 6th 2001. Jomo K
hand picking of weeds was done for successful establishment of seedlings in to the soil medium. They were then moved to an open area for further growth in preparation for grafting. Seedlings 9-12 months (5 months after establishment in the soil medium) were then grafted with scions obtained from selected varieties using four different methods namely top wedge (cleft), splice, side wedge and veneer. Scion material had been stored at 4-10°C for 1-2 wk prior to grafting to unify sprouting. Special care was taken to use scion material of the same age and size with the rootstock and to align the cambium layers of both the rootstock and the scion. Graft union was tightly fixed using a semi-permeable parafilm tape and the entire graft union sealed with a liquefied wax (paraffin : bees' wax - 9:1) to avoid desiccation and entry of water or pathogens. Top wedge grafted seedlings were tended in tunnels covered with polythene sheet in the green house maintained at 25-28°C, 80-100% relative humidity and 50 % light reduction. Splice and side wedge grafted seedlings were tended under the same conditions but not in tunnels while veneer grafted seedlings were maintained in an open area or under a shade (50% light reduction). After 3 wk in the greenhouse (open area/shade for veneer grafted seedlings), data was taken on successful graft takes, vigor of growth and time taken to first shooting, for a period of 3 months after which the seedlings were moved to a shade (50% light reduction) for hardening.
Results All the four grafting methods were successful for propagating macadamia. However, there were variations in graft takes % and time to first sprouting among the four methods with top wedge method having 90-100% takes and splice and side wedge methods with 80-100 and 70-100% respectively. The effect of variety was not significant when top wedge, splice and side wedge methods were used. However, when veneer method was used the effect of variety was significant with KMB-3 having significantly higher taking ratios (<80%) than MRG-20 (35-60%) but not KRG-15 and EMB-1 both with 60-80%. Shading veneer grafted seedlings did not improve graft taking frequencies. The following observations were made for the four methods; Top wedge method Graft takes % averaged at 90-100. This method was most successful with younger seedlings (5-9 months after establishment)
of pencil thickness and scion material of the same size was easily accessible It was easier and faster to prepare and also earlier in shooting (3 wk post-grafting) It formed a better and stronger graft union and scion shoots were more vigorous Seedlings were conveniently tended in tunnels that physically protected the seedlings from
mechanical damage and pests Seedlings were ready for field transplanting in 18-24 months from seeding through grafting
to hardening
Sproute
d top-wedge grafted seedlingshe Horticulture seminar on Sustainable Horticultural Production in the enyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 129

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 130
Splice method Graft takes % averaged at 80-100 It was successful on seedlings of all ages It was however more successful for pithy seedlings that were not flexible enough to allow a
tight fit of cambium layers if top wedge method was used eg. Seedlings that were raised during a dry spell probably due to lignification.
Side wedge Graft takes % averaged at 70-100 This method was more successful than top wedge method when older seedlings (9-12
months) were used (seedlings beyond pencil thickness) It demanded more skill to achieve maximum cambium alignment It took longer to sprout (4-5 months)
Veneer method Graft takes % averaged at 40-45 Time taken to first sprouting was similar to all other methods (3-5 wk) Seedlings were ready for field transplanting 3-5 months earlier than in other methods (15-
19 months from seeding)
Conclusions Top wedge method is fast and easy to make. It gives the highest number of successful takes and a nursery period of 18-24 months. It is therefore recommended for mass propagation of macadamia. Side wedge method is recommended for overgrown seedlings that cannot be successfully grafted by top wedge method, while splice method, if skillfully done, can compliment top wedge method since it is suitable for seedlings at all ages. Moreover, it is also suitable for seedlings that are not flexible enough for top wedge method. Such are seedlings that have been raised during a dry period. Veneer method is also simple to make, and since it does not require the conditions of a green house which unaffordable to most people, this method is recommended for small-scale nursery operators. Using this method, the total nursery period is also reduced by about 3-5 months thus saving on labor and material inputs.
Sprouted veneer grafted seedlings

Wesonga, et al., 2002. Proceedings of the Hortictropics October 3rd to 6th 2001. Jomo Kenyatta
Grafting of macadamia has achieved propagation and distribution to farmers clonal material of varieties with :- Improved in-shell nut yields (55-80 kg/tree/season)
Improved quality (≥72% kernel oil co Dwarfing effect and short juvenility (4
References Alexander, D.M. (1987) Practical Hints o
Commonwealth Science and IndHartmann, H.T. & Kester, D. (1983) Plan
Inc. Hartmann, H.T., Kester, D.E. & Davies, F
Practices. Prentice - Hall Inc. Ondabu, N., Nyaga A. and Tominaga T.
Proceedings of the 5th KARI conNyakundi, W. and Gitonga, L.N. (1993) M
THE EFFECT OF DIFFERENT SOROOTING OF HYPERICUM (HYPWatako, A.O. Department of Horticulture, Jomo Kenyatta Unive
Abstract The study, carried out in the greenhouse acompared the rooting of Hypericum androsaesand (control), sawdust and rice hulls in variwere tested for their suitability for the rootin2:3:1 (sand: sawdust: rice hull v/v) gave theand dry weight significantly increasing in 2:3
Improve
d grafted macadamia treeulture seminar on Sustainable Horticultural Production in the University of Agriculture and Technology, JKUAT, Juja, Kenya 131
ntent) yr)
f Budding and Grafting Fruit and Nut Trees. ustrial Research Organisation, Australia t Propagation: Principles and Practices. Prentice - Hall
.T. (1990) Plant Propagation: Principles and
(1997) Clonal selection of macadamia in Kenya. ference acadamia propagation handbook. JICA
ILESS SUBSTRATE MIXTURES ON THE ERICUM ANDROSAEMAM)
rsity of Agriculture and Technology
t Jomo Kenyatta University of Agriculture and Technology, mum in different substrate compositions. Three substrates,
ous compositions, and incubated for a period of six months, g of Hypericum cuttings. The results showed that the ratio
best performance. Root length percent rooting fresh weight :1 ratio over other mixture compositions.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 132
Incubation with sawdust and rice hulls decreased bulk density of sand. There was a slight decrease in the Cation Exchange capacity (C.E.C.) of the mixtures with incubation. The acidic properties of sawdust improved to near neutral pH with incubation.
Propagation of Lily Bulbs from Scales Muthoka N. M. National Horticultural Research Centre, Box 220, Thika
Abstract The lily cut flower is a relatively new introduction in the Kenyan flower industry. It is becoming a popular cut flower with a fairly steady demand in both the international and the local markets. The demand for the flower translates to demand for planting material. The demand for planting material is expected to be met partly by importation of the planting material of the varieties covered by breeder’s rights and multiplication of bulbs of the varieties not covered by breeders rights. One of the conventional methods of multiplication is by the use of scales. Scales are documented as able to produce up to 3-4 bulblet per scale (Rees 1992). An experiment was done on scaling using both the conventional and the tissue culture methods of multiplication with the additional use of auxins. In- vivo 5 parts sand: 1 part manure medium had relatively desirable results with 2 bulblets per scale on average with a size of between (1- 2) mm after four weeks. Rooting hormone-containing IBA (indole-butyric acid) was applied to induce bulblet formation at the base of the scales. There was no significant difference in bulblet number produced per scale but the bulblet size was significantly different between the treated scales and the untreated scales. In-vitro using Linsmaier and skoog media at least 3 bulblets were developed per scale section with an average size of 3 - 5 mm. The cultures were incubated for 4 weeks in the dark at a temperature of 26 ±±±± 2 oC. A follow-up study using LS media and different levels of NAA (Naphthalene acetic acid) showed that the number of bulblets formed at different levels of the auxin were not significantly different. They however, increased in size (10 - 15mm) following transfer to LS media supplemented with 0.1mgl -1 NAA and growing in a 16 hour light and 8 hour dark photoperiod at a temperature of 26 ±±±± 20C for 1 month.
Introduction The lily cut flower is a relatively new introduction in the Kenyan flower industry. It is becoming a popular cut flower with a fairly steady demand in both the international and the local markets. The demand for the flower translates to demand for planting material. The demand for planting material is expected to be met partly by importation of the planting material of the varieties covered by breeder’s rights and multiplication of bulbs of the varieties not covered by breeders rights. Lilies are becoming increasingly important cut flowers in the international and local markets. They compete very well with other cut flowers, ranking fourth in importance after roses, carnation and tulips. Lilies have a very great potential to contribute to small holder, resource poor farmers' incomes because they can be grown economically on small pieces of land without expensive grow-in structures and cultural practices. The flower has become an attractive cash crop to many small-scale farmers, resulting in a very high demand for planting material. Most of the popular varieties are covered by plant breeder's rights. Their cost is therefore out of reach of many small holder farmers. Some of the popular varieties, however, do not attract breeder's royalties and may therefore be multiplied for this category of farmers. Lilies produce one stem and one bulb under conventional methods. Bulblets have been produced in vivo from bulb-scales, but the system yields few propagules and takes a period of 2- 3 years to develop them to a size at which they are of commercial value. Efficient methods of bulb multiplication to enable these farmers to access more planting material of this flower are required.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 133
The genus Lilium is known to have a high capacity to generate bulblets through scaling (Niimi 1986). Bulblet production is highest from scales. The basal portion of a bulb scale regenerates into bulblets (Robb, 1957). These scales require a suitable media to be able to produce 3-4 bulbets per scale. Auxins are defined as chemicals that produce cell elongation; IBA (Indole-butryric acid) and NAA (Naphthalene acetic acid) are synthetic auxins. IBA is similar to IAA (indole-acetic acid) a naturally occurring hormone. Research since 1920’s has shown auxins to be involved with a multitude of plant responses (Mc conell et al 1981). Application of hormones enhance the naturally produced hormones in plants. Auxins may stimulate or prevent rooting owing to temperature changes, fertilizer levels, water practises and stages of growth.
General Objective To determine a suitable method for the multiplication of lily bulblets.
Specific objectives To compare the number and size of bulblets produced per scale using the conventional
and the tissue culture method To determine the effect of auxins on the number and size of bulblets produced.
Justification To meet the demand for the lily cutflower production of adequate planting material will be crucial. The method used should be effective and economical. Cut flowers developed from bulblets are of better quality in terms of stem length, bud number and bud length when compared with those produced from second generation bulbs. (Rees 1992) Bulblet production is highest from scales. This means that many bulbs can be generated within a certain period of time given the right management. Rooting hormone containing IBA( indole –butyric acid) can also be applied to induce bulblet formation at the base of the scales. Regeneration is increased when NAA (Naphthalene acetic acid) is incorporated into the medium (Hackett, 1969). IBA, NAA and IAA are auxins that enhance cell elongation. It is necessary to evaluate the effects of auxins on the number and size of bulblets.
Materials and Methods In-vivo culture The experiment was carried out at Thika- Jomo Kenyatta University of Agriculture and Technology ( Horticulture Department). Mature bulbs of cultivars 'Snow queen' were harvested from the field and stored at 2 0C for 6 weeks. The medium 5parts sand: 1part manure was replicated twice where by in one treatment the scales were treated with Indole-acteic acid (IBA) and the other without. 20 suitable scales were randomly selected and planted in the media with the bases on the
media The polyethene sheet was then covered over all the trays The media were kept moist all the time RCBD design was used
In-vitro culture The experiment was carried out at Thika- National Horticultural Research Centre
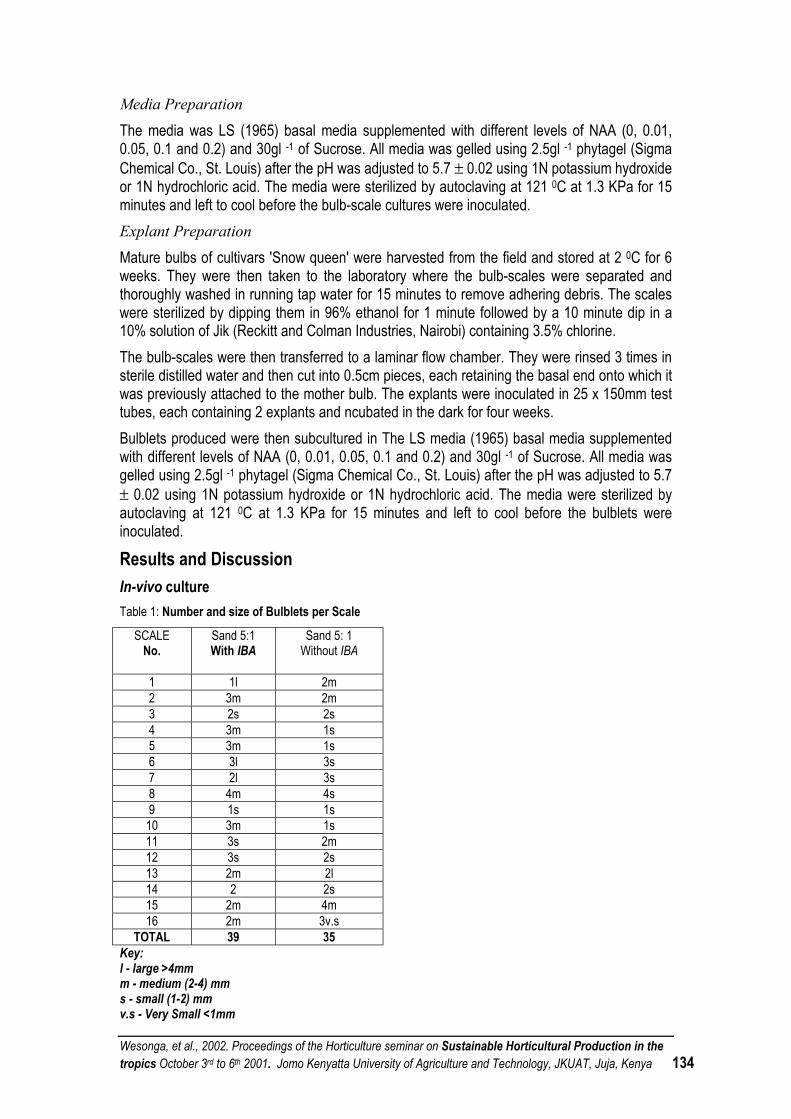
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 134
Media Preparation The media was LS (1965) basal media supplemented with different levels of NAA (0, 0.01, 0.05, 0.1 and 0.2) and 30gl -1 of Sucrose. All media was gelled using 2.5gl -1 phytagel (Sigma Chemical Co., St. Louis) after the pH was adjusted to 5.7 ± 0.02 using 1N potassium hydroxide or 1N hydrochloric acid. The media were sterilized by autoclaving at 121 0C at 1.3 KPa for 15 minutes and left to cool before the bulb-scale cultures were inoculated. Explant Preparation Mature bulbs of cultivars 'Snow queen' were harvested from the field and stored at 2 0C for 6 weeks. They were then taken to the laboratory where the bulb-scales were separated and thoroughly washed in running tap water for 15 minutes to remove adhering debris. The scales were sterilized by dipping them in 96% ethanol for 1 minute followed by a 10 minute dip in a 10% solution of Jik (Reckitt and Colman Industries, Nairobi) containing 3.5% chlorine. The bulb-scales were then transferred to a laminar flow chamber. They were rinsed 3 times in sterile distilled water and then cut into 0.5cm pieces, each retaining the basal end onto which it was previously attached to the mother bulb. The explants were inoculated in 25 x 150mm test tubes, each containing 2 explants and ncubated in the dark for four weeks. Bulblets produced were then subcultured in The LS media (1965) basal media supplemented with different levels of NAA (0, 0.01, 0.05, 0.1 and 0.2) and 30gl -1 of Sucrose. All media was gelled using 2.5gl -1 phytagel (Sigma Chemical Co., St. Louis) after the pH was adjusted to 5.7 ± 0.02 using 1N potassium hydroxide or 1N hydrochloric acid. The media were sterilized by autoclaving at 121 0C at 1.3 KPa for 15 minutes and left to cool before the bulblets were inoculated.
Results and Discussion In-vivo culture Table 1: Number and size of Bulblets per Scale
SCALE No.
Sand 5:1 With IBA
Sand 5: 1 Without IBA
1 1l 2m 2 3m 2m 3 2s 2s 4 3m 1s 5 3m 1s 6 3l 3s 7 2l 3s 8 4m 4s 9 1s 1s
10 3m 1s 11 3s 2m 12 3s 2s 13 2m 2l 14 2 2s 15 2m 4m 16 2m 3v.s
TOTAL 39 35 Key: l - large >4mm m - medium (2-4) mm s - small (1-2) mm v.s - Very Small <1mm
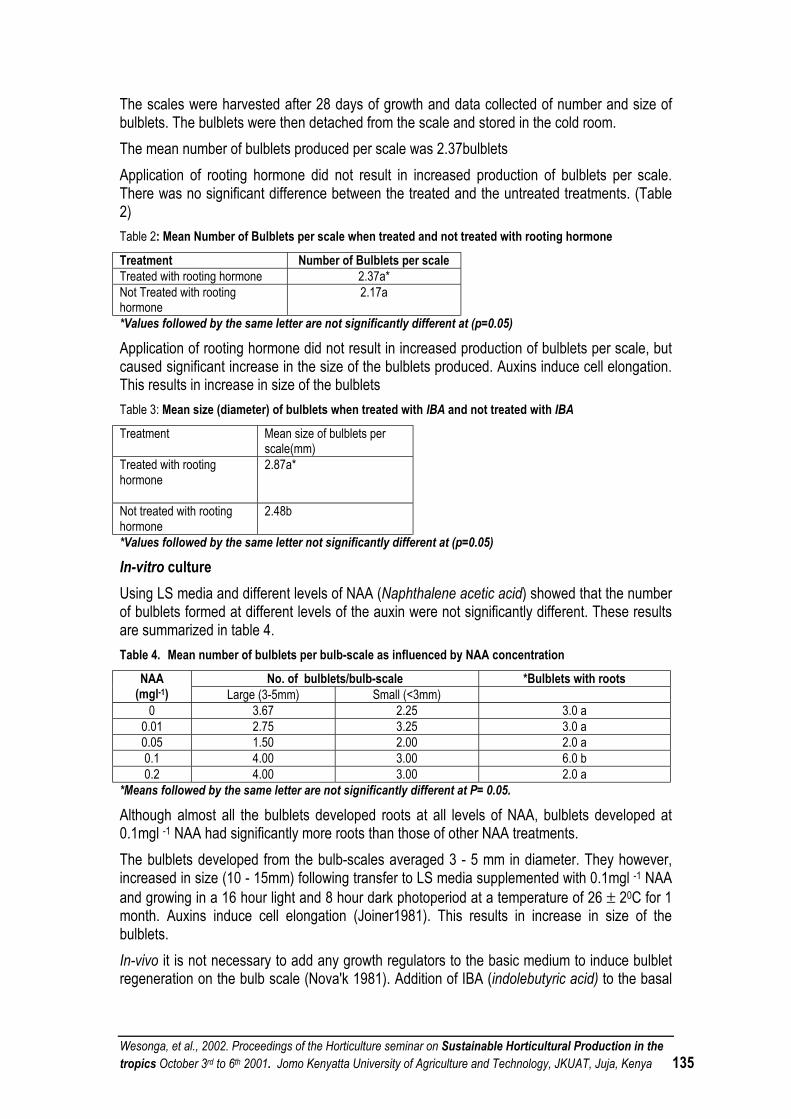
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 135
The scales were harvested after 28 days of growth and data collected of number and size of bulblets. The bulblets were then detached from the scale and stored in the cold room. The mean number of bulblets produced per scale was 2.37bulblets Application of rooting hormone did not result in increased production of bulblets per scale. There was no significant difference between the treated and the untreated treatments. (Table 2) Table 2: Mean Number of Bulblets per scale when treated and not treated with rooting hormone
Treatment Number of Bulblets per scale Treated with rooting hormone 2.37a* Not Treated with rooting hormone
2.17a
*Values followed by the same letter are not significantly different at (p=0.05)
Application of rooting hormone did not result in increased production of bulblets per scale, but caused significant increase in the size of the bulblets produced. Auxins induce cell elongation. This results in increase in size of the bulblets Table 3: Mean size (diameter) of bulblets when treated with IBA and not treated with IBA
Treatment Mean size of bulblets per scale(mm)
Treated with rooting hormone
2.87a*
Not treated with rooting hormone
2.48b
*Values followed by the same letter not significantly different at (p=0.05)
In-vitro culture Using LS media and different levels of NAA (Naphthalene acetic acid) showed that the number of bulblets formed at different levels of the auxin were not significantly different. These results are summarized in table 4. Table 4. Mean number of bulblets per bulb-scale as influenced by NAA concentration
No. of bulblets/bulb-scale *Bulblets with roots NAA (mgl-1) Large (3-5mm) Small (<3mm)
0 3.67 2.25 3.0 a 0.01 2.75 3.25 3.0 a 0.05 1.50 2.00 2.0 a 0.1 4.00 3.00 6.0 b 0.2 4.00 3.00 2.0 a
*Means followed by the same letter are not significantly different at P= 0.05.
Although almost all the bulblets developed roots at all levels of NAA, bulblets developed at 0.1mgl -1 NAA had significantly more roots than those of other NAA treatments. The bulblets developed from the bulb-scales averaged 3 - 5 mm in diameter. They however, increased in size (10 - 15mm) following transfer to LS media supplemented with 0.1mgl -1 NAA and growing in a 16 hour light and 8 hour dark photoperiod at a temperature of 26 ± 20C for 1 month. Auxins induce cell elongation (Joiner1981). This results in increase in size of the bulblets. In-vivo it is not necessary to add any growth regulators to the basic medium to induce bulblet regeneration on the bulb scale (Nova'k 1981). Addition of IBA (indolebutyric acid) to the basal

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 136
portions of scales showed no significant difference between the treated and the untreated scales (Table 2) in terms of bulblet number. NAA (Naphthalene acetic acid) addition to the medium has insignificant effect in increasing the number of bulblets per scale (Stimart and Asher 1978). Combination of auxins and cytokinin at certain ratios results in bulblet regeneration and growth that is used in tissue culture medium.
Conclusion and Areas for further research These studies indicate that it is possible to produce large numbers of lilies from bulb-scale sections within a short period using tissue culture technology as compared to the conventional method. More work, however, needs to be carried out to make the production protocol more efficient. The use of IBA is important but not necessary for in-vivo culture. The sizes achieved by the application of the same were significant, but with proper management of the bulblets after the initial harvest the increase in size can still be achieved. The use of NAA in in-vitro culture caused significant increase in size. However, further studies need to be done to look at the bulblet responses to combinations of auxins and cytokinin hormones.
Acknowledgements I would like to thank the following persons for their support and technical advice, which enable me to carry out this work Eston Njeru (Biotechnology section-NHRC), Wariara Kariuki, staff of Horticulture department- JKUAT. This paper has included some work that was funded by JICA Third Country Training Programme at JKUAT.
Literature Cited Boodley J. W., and R.S. Sheldrake 1977. Cornell peat – lite mixes for commercial plant
growing. Cornell plant science information Bulletin 43 Doerflinger F. (1982) Know your Bulbs. Manual No.1 “ Spring Flowering Bulbs” Horticultural
Trades Association Reading UK. Hackett W.P., 1969.Aseptic multiplication of lily bulblets from scales. Int plant Prop.soc.
proc,19:105-108 Joiner J.N (1981) Foliage Plant Production. Prentice-hall,inc.,Englewood cliffs pg.295,179 - 199 Niimi y.1986 Application of leaf segment culture to invitro bulblet production of six lilium
species, Acta Bot.neert. 35:189- 194 Nova’k, F.J and PetruE;1981. Tissue culture propagation of Lilium hybrids. Scientia hortic.14:
191- 199 Poole R.T 1969.Rooting response of four ornamental species propagates in various media
Proc.fla. Stte hort soc.82;393 – 397. Rees A.R. (1985) Ornamental bulbous plants. In: Halevy AH (ed), Handbook of Flowering Vol.
1. CRC press, Boca Raton . Florida pp259-308. Rees A.R. (1992) Ornamental Bulbs, Corms and tubers. CAB international Redwood Press Ltd.
Melksham Stimar and Asher, 1978 Tissue culture of bulb scale section for asexual propagation of lilium
longiflorum thumb. J. Amer.soc. Hort.Sc.103: 182 – 184. Yoshiji Niimi, 1995. Invitro propagation and post- in vitro Estblishment of bulblets of lilum
japonicum thunb ,J. Japan. Soc. Hort.sci.63 (4): 843 – 852. Water, W.E. W. Lewellyn and J. Nesmith 1970. The chemical physical and salinity
characteristic of 27 soil media Proc.Fla.state hort. Soc.482 – 488

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 137
GROWTH OF MANGO ROOTSTOCK SEEDLINGS UNDER DIFFERENT WATER REGIMES Ndung'u C. K. and Sugiura, H. Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P. O. Box 62000 Nairobi, Kenya.
Abtsract Mango trees in the tropics flower during the warm weather, after experiencing a period of drought. However, shoot growth and flowering are very erratic. In this study effect of water deficit stress on growth of young mango rootstocks was investigated. Plants were grown in the field, whereby one set of plants was allowed to grow without irrigation (rain-fed only) and another set was regularly irrigated twice or thrice per week. Irrigation treatments were continued for a period of nine months (April 1999 to January 2000). Soil water content ranged between 21% to 27%, and 11% to 17% for the irrigated and non-irrigated seedlings, respectively. Shoot length and shoot diameter were significantly reduced under water stress. Number of nodes also decreased under water stress. Effects of water stress on mango seedling growth under field conditions became evident after 6 months of drought. The study shows that drought significantly reduces early seedling growth of mango rootstocks.
PRELIMINARY STUDIES ON GRAFTING OF TOMA TO (LYCOPERSICON ESCULENTUM MILL.) ON SOLANUM INCANUM L. Ndung'u, C. K. Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P. 0. Box 62000 Nairobi, Kenya.
Abstract A preliminary study was conducted to test the graft compatibility of tomato (Lycopersicon esculentum L.) on Solanum incanum L. Seeds of S. incanum germinated 4 to 5 weeks after sowing, and were transplanted at the 6-leaf stage. Seeds of four varieties of tomato, i.e. Money Maker, Marglobe, Hofit, and Roma VF were sown 4 months after transplanting S. incanum. One month after sowing, tomato seedlings had attained the same size as S. incanum, at which stage wedge grafting of tomato on S. incanum was done. Graft union take occurred within 12 days after grafting. Percent success in grafting was 100%, 96%, 96%, and 80% for Money Maker, Marglobe, Hofit, and Roma VF , respectively. The study shows that S. incanum is compatible to tomato and can successfully be used as a rootstock.
TOMATO GRAFTING: OPTIMISATION OF THE TECHNIQUE AND ENVIRONMENTAL CONDITIONS FOR GRAFT SUCCESS Kusolwa, P. Sokoine University of Agriculture, Department of Crop Science and Production, P.O. Box 3005, Morogoro, Tanzania.
Abstract In order to exploit the potential of grafting technique in vegetable improvement, a field experiment was conducted to optimise the grafting conditions for successful grafting in tomato. Young seedlings of two varieties of tomato rootstocks known to possess vigour and resistance to nematode were used. Two varieties of tomato scions were also used for the study. The tested variables in this study were suitable age for both rootstock and scion seedlings, the techniques for supporting the graft point and the relative humidity where grafted plants were kept just after grafting. Results shows that a successful graft union formation is high when seedlings between 3 to 5 weeks old are used for both rootstock and scion. Water spraying on the grafted seedling results into excessive increase in relative humidity leading to fungal infestations on the graft union. Grafted plants kept under

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 138
ambient relative humidity showed high graft success. Tying strips provide an excellent support for successful graft union formation than the use of intravenous giving (I.V.) tubes. Significant difference in plant performance in terms of plant growth vigour; increase in leaf number and size was observed for each rootstock scion combination. The best rootstock -scion combination was observed when money maker was used as scion and when Brigeor HF 1 was used as rootstock.
EFFECTS OF FORCING METHODS AND ROOT PRUNING ON THE GROWTH OF BUDDED ‘ROUGH LEMON’ ROOTSTOCKS GEORGE OUMA Department of Horticulture Maseno University, P. 0. Box 333 Maseno, Kenya, Telephone: (035) 51622/51620
Abstract Experiments were conducted at Maseno University, Kenya to find the effect of three different bud forcing methods namely: bending, lopping and cutting-off and root pruning on the growth of 'rough lemon' (Citrus sinensis L.) rootstocks budded with Washington Navel' citrus. Forcing method significantly (P≤≤≤≤0.05) affected the growth of the 'rough lemon' rootstocks budded on to 'Washington Navel'. Cutting off method was significantly more superior to bending and lopping and it increased the dry weights of whole plants, stem, roots, scion leaves, shoot and the scion lengths. Forcing method did not however affect the root to shoot ratio. Between bending and lopping the former was superior to the other in terms of dry weights of whole plants, stem, roots, scion leaves, shoot etc. Bud break for the cutting off method was over 80% while that of bending and lopping was about 30%. Root pruning interacted with forcing method to reduce the dry weights of the above parameters. It is concluded that cutting off method in bud forcing 'rough lemon' rootstock is superior to bending and lopping methods in promoting growth of nursery stock and that root pruning interacts with it and other forcing methods applied in the present study to reduce the dry weights of the parameters determined. Finally it can be concluded that the manufacture of carbohydrates by leaves in bended or lopped rootstock shoots may not be the reason for the superior scion growth in some rootstock-scion combination as reported previously.
Introduction Cutting off, lopping or bending the rootstock shoot are common methods used in citrus nurseries to force scion bud growth (Rouse, 1988; Tucker and Youtsey, 1980; Williamson and Castle, 1989). Forcing methods such as lopping and bending which leave rootstock shoots attached usually result in greater nursery tree growth than cutting off rootstock shoots (Amih, 1980; Rouse, 1988; Williamson et al, 1992), most probably due to photosynthates supplied by the attached rootstock shoots. It has been reported that 14C-Iabelled photosynthates were found from rootstock shoots in scions and roots of Hamlin-carrizo trees following bud forcing by bending or lopping (Williamson et al. 1992). It has been consequently concluded that the benefit to nursery tree growth resulted from photsynthates produced in the rootstock shoot rather from stored carbohydrate reserves. (Williamson and Maust, 1995). The main disadvantage of bending or lopping is greater production costs which are associated with either of these methods compared to cutting off rootstock shoots. More labor, irrigation, fertilizer, pesticide and space are usually required for the production of plants forced by bending or lopping than for those forced by cutting off (Williamson, 1997). Root pruning has generally reduced tree size and shoot growth (Elfving et al. 1996; Ferree, 1992), but in some studies results were inconsistent (Baugher et al. 1995; Miller, 1995). Root pruning has reduced growth consistently in trees growing on the dwarfing Apple rootstocks (Ferree and Rhodus, 1993; Schupp et al, 1992) and mixed results were obtained from the semi dwarfing rootstocks of Apple such as M. T. (Baugher et al. 1995; Ferree and Rhodus, 1993; Miller, 1995) Well established root systems with high fibrous root content have improved transplant establishment of field grown citrus nursery trees (Grimm, 1956) Many other perennial crops are routinely root pruned in field nurseries to develop more compact, fibrous root terms and higher root shoot

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 139
ratios (Geisler and Ferree, 1984). In some cases (Mullin, 1966; Sutton, 1967) but not others (Janouch, 1972), this result has been correlated with higher transplant survival rates and better growth following transplanting. The present study attempts to find the effect of forcing method and root pruning on the growth of 'rough lemon' (C. Sinensis L) rootstocks budded to 'Washington Navel' scions. 'Rough lemons' are the most common rootstocks used in Kenya by the citrus nurseries and farmers and such studies have not been conducted on it before. Further, root pruning studies have not been conducted on citrus rootstocks at all, let alone on 'rough lemon' and with respect to the forcing methods.
Material and Methods: Location of Research Site: The study was conducted at the Maseno University's department of Horticulture nurseries, Kenya. The nurseries are situated at an altitude of 1515 metres above sea level and are at a longitude of 34°, 36° East and latitude of 00. The soils consist of a complex of somewhat excessively drained, shallow, stony and rocky soils of varying color, consistence and texture (dystric regosols with ferralic combisols, lithic phase and rock out crops). The area receives an annual rainfall of 1853mm which is fairly well distributed. Experimental Materials and Preparation: 'Rough lemon' (Citrus sinensis) fruits were obtained from a commercial farm near Kisumu town, Kenya after harvesting The fruits were then transported to Maseno University and stored in the refrigerator for 2 days at a temperature of 5C. The fruits were then washed and graded for uniformity of mass and freedom from blemishes. The fruits were subsequently cut in half and seeds extracted from them in warm water at a temperature of70oC. The seeds were then dried briefly for two days in trays in readiness for planting in the field. Land Preparation. Fertilization. Planting and Subsequent Care: Nursery beds measuring 20 metres by 10 meters were well prepared to a fine tilt using hoes and machetes and well mixed with 8 Kg of farm yard manure and 100Kg per hectare of Diammonium phosphate (18;46;0; N; P; K) fertilizer. The seeds were planted on the nursery beds on 10th May 1998 at spacing of 40cm by 15cm and a layer grass mulch put between the holes The beds were watered daily at 0800 hours and 1700 hours to saturation using a watering can For pest control Aldrin at 40% was applied at a rate of 5gm per Kg of seed for the control of cut worms and Dimethoate (dimethyl -5- (N-methvl carba-methyl) phosphorothiolo thionate) 40% EC at a rate of 1 litre in 500 litres of water per hectare sprayed at 2 weeks interval to control other insects. Diseases were controlled by Benomyl (Methyl N - (1-butylcarbomyl-2-benzimidazole) - carbamate) at a rate of 20gm per 20 litre of water. Transplanting of seedlings. The seedlings were transplanted into plastic pots measuring 4.5 litres. Before the seedlings were transplanted the nursery beds were well watered to facilitate up rooting and before that the seedlings under went a 'hardening off process when light was increased by reducing the shade, watering frequency was reduced to every other day and fertilization level reduced. The media in the pots comprised 1 part soil to 1 part sand and it was amended with 24g of farm yard manure and 12g Diammonium phosphate (18;46;0;N; P; K) fertilizer. After planting the seedlings they were watered every 2 days in the morning hours. The spacing between the pots was periodically adjusted to minimize shading between the plants.
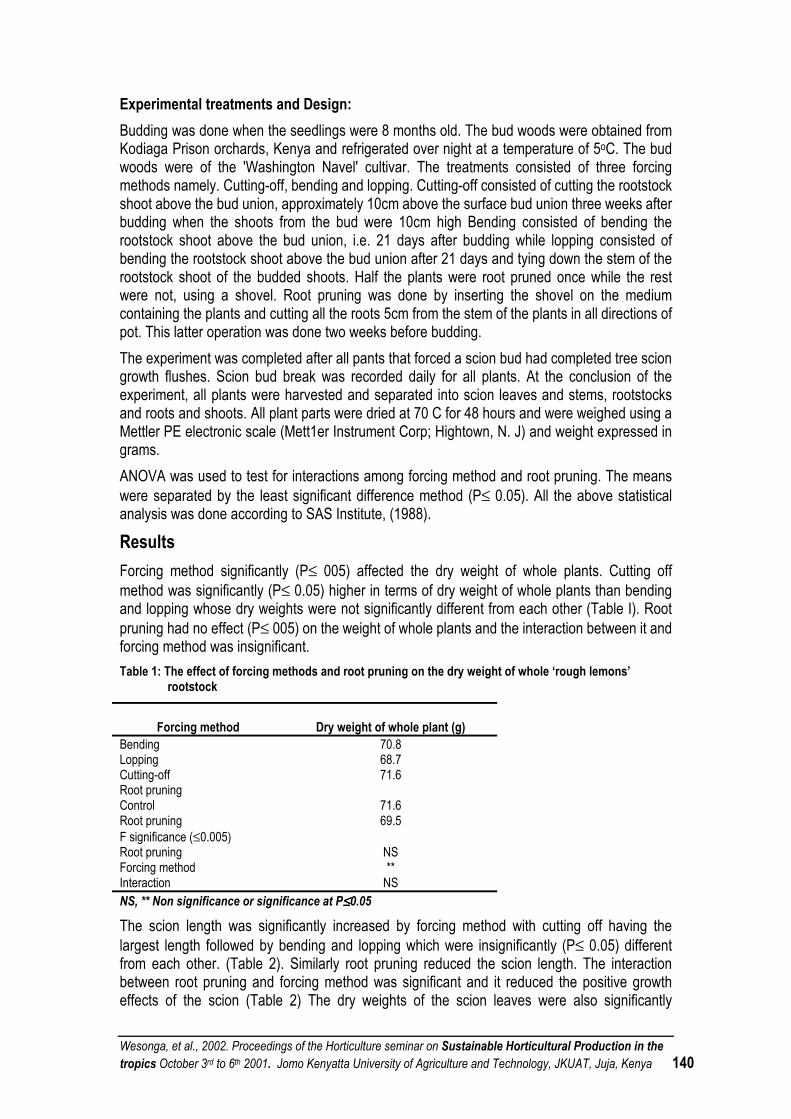
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 140
Experimental treatments and Design: Budding was done when the seedlings were 8 months old. The bud woods were obtained from Kodiaga Prison orchards, Kenya and refrigerated over night at a temperature of 5oC. The bud woods were of the 'Washington Navel' cultivar. The treatments consisted of three forcing methods namely. Cutting-off, bending and lopping. Cutting-off consisted of cutting the rootstock shoot above the bud union, approximately 10cm above the surface bud union three weeks after budding when the shoots from the bud were 10cm high Bending consisted of bending the rootstock shoot above the bud union, i.e. 21 days after budding while lopping consisted of bending the rootstock shoot above the bud union after 21 days and tying down the stem of the rootstock shoot of the budded shoots. Half the plants were root pruned once while the rest were not, using a shovel. Root pruning was done by inserting the shovel on the medium containing the plants and cutting all the roots 5cm from the stem of the plants in all directions of pot. This latter operation was done two weeks before budding. The experiment was completed after all pants that forced a scion bud had completed tree scion growth flushes. Scion bud break was recorded daily for all plants. At the conclusion of the experiment, all plants were harvested and separated into scion leaves and stems, rootstocks and roots and shoots. All plant parts were dried at 70 C for 48 hours and were weighed using a Mettler PE electronic scale (Mett1er Instrument Corp; Hightown, N. J) and weight expressed in grams. ANOVA was used to test for interactions among forcing method and root pruning. The means were separated by the least significant difference method (P≤ 0.05). All the above statistical analysis was done according to SAS Institute, (1988).
Results Forcing method significantly (P≤ 005) affected the dry weight of whole plants. Cutting off method was significantly (P≤ 0.05) higher in terms of dry weight of whole plants than bending and lopping whose dry weights were not significantly different from each other (Table I). Root pruning had no effect (P≤ 005) on the weight of whole plants and the interaction between it and forcing method was insignificant. Table 1: The effect of forcing methods and root pruning on the dry weight of whole ‘rough lemons’
rootstock
Forcing method
Dry weight of whole plant (g)
Bending 70.8 Lopping 68.7 Cutting-off 71.6 Root pruning Control 71.6 Root pruning 69.5 F significance (≤0.005) Root pruning NS Forcing method ** Interaction NS NS, ** Non significance or significance at P≤≤≤≤0.05
The scion length was significantly increased by forcing method with cutting off having the largest length followed by bending and lopping which were insignificantly (P≤ 0.05) different from each other. (Table 2). Similarly root pruning reduced the scion length. The interaction between root pruning and forcing method was significant and it reduced the positive growth effects of the scion (Table 2) The dry weights of the scion leaves were also significantly
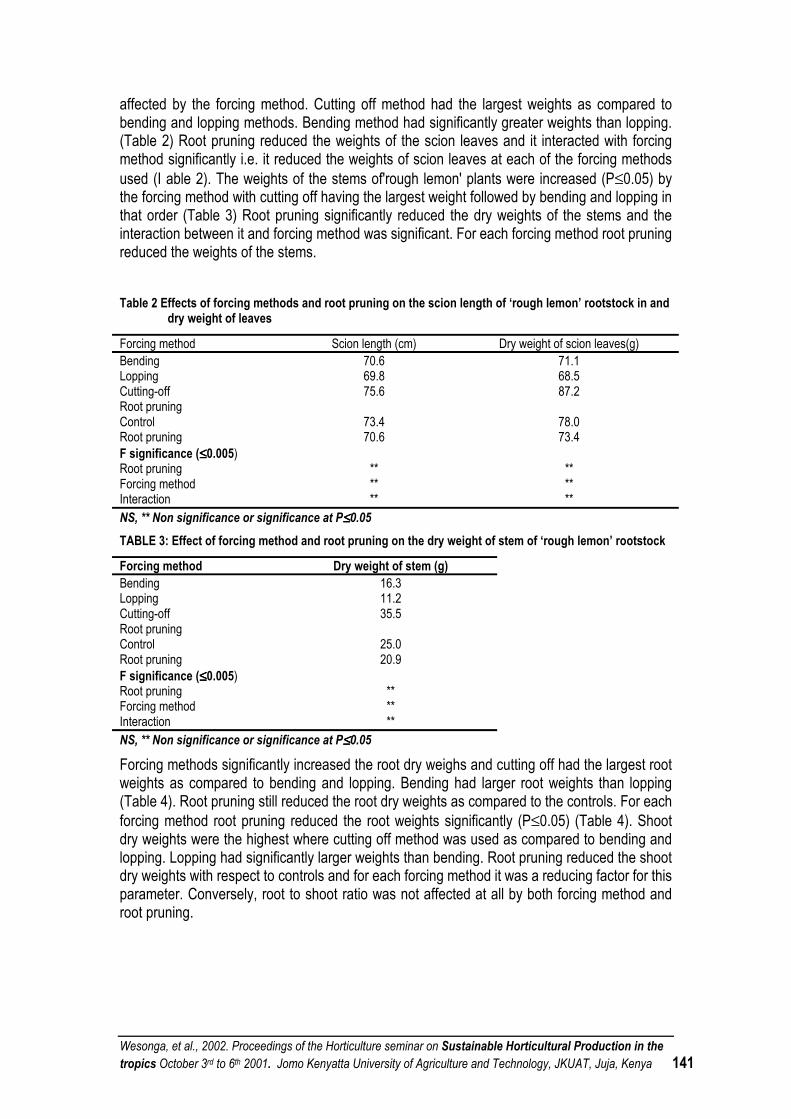
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 141
affected by the forcing method. Cutting off method had the largest weights as compared to bending and lopping methods. Bending method had significantly greater weights than lopping. (Table 2) Root pruning reduced the weights of the scion leaves and it interacted with forcing method significantly i.e. it reduced the weights of scion leaves at each of the forcing methods used (I able 2). The weights of the stems of'rough lemon' plants were increased (P≤0.05) by the forcing method with cutting off having the largest weight followed by bending and lopping in that order (Table 3) Root pruning significantly reduced the dry weights of the stems and the interaction between it and forcing method was significant. For each forcing method root pruning reduced the weights of the stems. Table 2 Effects of forcing methods and root pruning on the scion length of ‘rough lemon’ rootstock in and
dry weight of leaves
Forcing method Scion length (cm) Dry weight of scion leaves(g) Bending 70.6 71.1 Lopping 69.8 68.5 Cutting-off 75.6 87.2 Root pruning Control 73.4 78.0 Root pruning 70.6 73.4 F significance (≤≤≤≤0.005) Root pruning ** ** Forcing method ** ** Interaction ** ** NS, ** Non significance or significance at P≤≤≤≤0.05 TABLE 3: Effect of forcing method and root pruning on the dry weight of stem of ‘rough lemon’ rootstock
Forcing method Dry weight of stem (g) Bending 16.3 Lopping 11.2 Cutting-off 35.5 Root pruning Control 25.0 Root pruning 20.9 F significance (≤≤≤≤0.005) Root pruning ** Forcing method ** Interaction ** NS, ** Non significance or significance at P≤≤≤≤0.05
Forcing methods significantly increased the root dry weighs and cutting off had the largest root weights as compared to bending and lopping. Bending had larger root weights than lopping (Table 4). Root pruning still reduced the root dry weights as compared to the controls. For each forcing method root pruning reduced the root weights significantly (P≤0.05) (Table 4). Shoot dry weights were the highest where cutting off method was used as compared to bending and lopping. Lopping had significantly larger weights than bending. Root pruning reduced the shoot dry weights with respect to controls and for each forcing method it was a reducing factor for this parameter. Conversely, root to shoot ratio was not affected at all by both forcing method and root pruning.
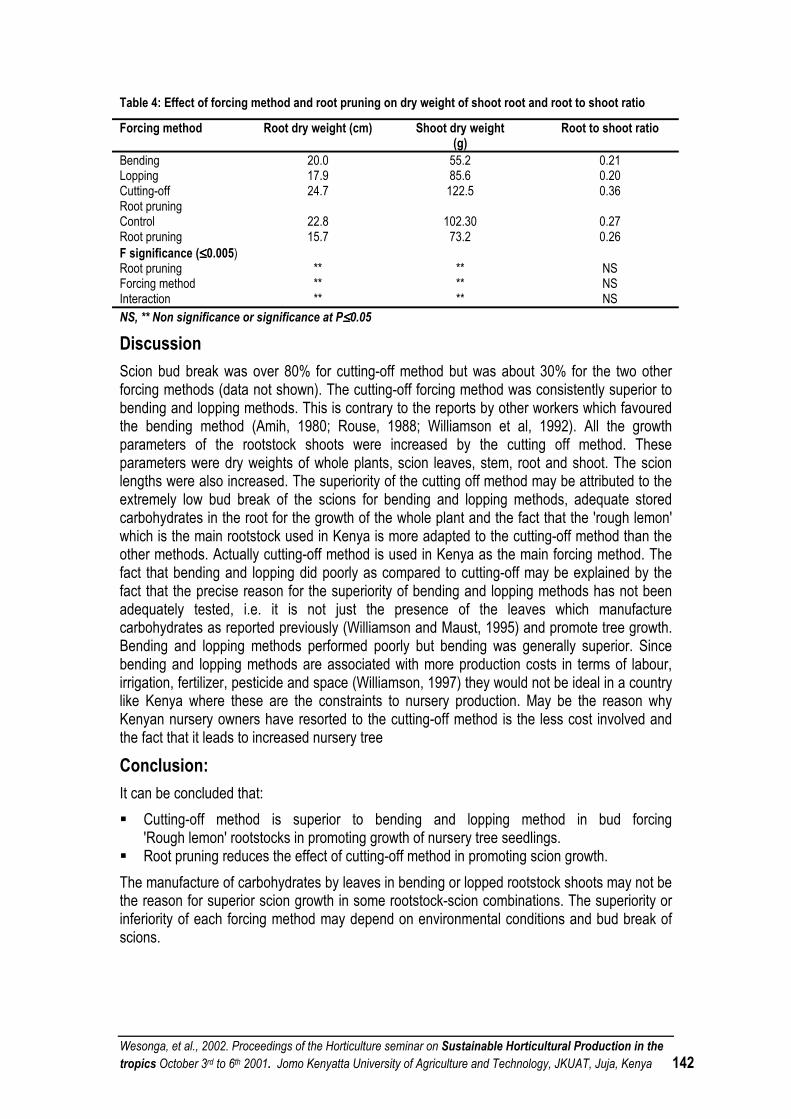
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 142
Table 4: Effect of forcing method and root pruning on dry weight of shoot root and root to shoot ratio
Forcing method Root dry weight (cm) Shoot dry weight (g)
Root to shoot ratio
Bending 20.0 55.2 0.21 Lopping 17.9 85.6 0.20 Cutting-off 24.7 122.5 0.36 Root pruning Control 22.8 102.30 0.27 Root pruning 15.7 73.2 0.26 F significance (≤≤≤≤0.005) Root pruning ** ** NS Forcing method ** ** NS Interaction ** ** NS NS, ** Non significance or significance at P≤≤≤≤0.05
Discussion Scion bud break was over 80% for cutting-off method but was about 30% for the two other forcing methods (data not shown). The cutting-off forcing method was consistently superior to bending and lopping methods. This is contrary to the reports by other workers which favoured the bending method (Amih, 1980; Rouse, 1988; Williamson et al, 1992). All the growth parameters of the rootstock shoots were increased by the cutting off method. These parameters were dry weights of whole plants, scion leaves, stem, root and shoot. The scion lengths were also increased. The superiority of the cutting off method may be attributed to the extremely low bud break of the scions for bending and lopping methods, adequate stored carbohydrates in the root for the growth of the whole plant and the fact that the 'rough lemon' which is the main rootstock used in Kenya is more adapted to the cutting-off method than the other methods. Actually cutting-off method is used in Kenya as the main forcing method. The fact that bending and lopping did poorly as compared to cutting-off may be explained by the fact that the precise reason for the superiority of bending and lopping methods has not been adequately tested, i.e. it is not just the presence of the leaves which manufacture carbohydrates as reported previously (Williamson and Maust, 1995) and promote tree growth. Bending and lopping methods performed poorly but bending was generally superior. Since bending and lopping methods are associated with more production costs in terms of labour, irrigation, fertilizer, pesticide and space (Williamson, 1997) they would not be ideal in a country like Kenya where these are the constraints to nursery production. May be the reason why Kenyan nursery owners have resorted to the cutting-off method is the less cost involved and the fact that it leads to increased nursery tree
Conclusion: It can be concluded that: Cutting-off method is superior to bending and lopping method in bud forcing
'Rough lemon' rootstocks in promoting growth of nursery tree seedlings. Root pruning reduces the effect of cutting-off method in promoting scion growth.
The manufacture of carbohydrates by leaves in bending or lopped rootstock shoots may not be the reason for superior scion growth in some rootstock-scion combinations. The superiority or inferiority of each forcing method may depend on environmental conditions and bud break of scions.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 143
Acknowledgement I am indebted to the Institute of Research and Post graduate Studies at Maseno University for the research funds, to Kibos prisons for research facilities and to Agatha for typing the manuscript.
Literature Cited Amih, C. A. (1980) The effects of time and method of forcing on bud survival and scion growth
of citrus. M.Sc. Thesis. Univ. of Florida, Gainesville. Baugher, T. A.; K. C. Elliot and D. M. Glenn (1995) Effect of Sod competition and root pruning
on 'Stayman' apple tree growth and fruit cracking. HortScience 30: 222-226. Elfving, D. C., I. Schechter and M. Born (1996) Effect of root pruning and Benzyladenine
application on tree growth and fruit size in 'Empire' and 'McIntosh' apple. J. Tree Fruit Prad. I: 1-13.
Ferree, D. C. (1992) Time of root pruning Influences Vegetative growth, fruit size, biennial bearing and yield of' Janathan' apple. J. Amer. Soc. Hort. Sci;. 117.198-202.
Ferree, D. C. and W. T. Rhodus (1993) Apple tree performance with mechanical hedging or root pruning in intensive orchards. J. Amer. Soc. Hort. Sci;. 118. 707-713.
Geisler, D. and D. C. Ferree (1984) Response of plants to root pruning. Hort. Rev. 6. 155-188. Grim, G. R. (1956) Preliminary Investigations on dieback of young transplanted citrus trees.
Proc. Fla. State. Hort. Soc. 69.31-34. Janouch, K. L. (1927) Effect of spacing and root pruning on the development of transplants. J.
Forest 25. 62-67. Miller, S. S. (1995) Root pruning and trunk scoring have limited effect on young bearing apple
trees. HortScience 30: 981-984. Mullin, R. D. (1966) Root pruning of nursery stock. For Chron. 42. 256-264 Rouse, R. E. (1988) Bud forcing method affects bud-break and Scion growth of citrus grown in
containers. J Rio Grande valley Hort. Soc. 41. 69- 73 Schupp, J. R., D. C. Ferree and I. J. Washington (1992) Interactions of root pruning and
deblossoming on growth development and yield of 'Golden Delicious' Apples J Hort. Sci. 67.465-480.
Sutton, R. F. (1967) Influence of root pruning on height increment and root development of out planted spruce Can. J Bot. 45. 1671-1682.
Tucker, D. P. H. and C. 0. Youtsey (1980) Citrus nursery practices. Fla. Coop. Ext. Serv., Inst. Food and Agr. Sci. University of Florida, Circ. 430.
Williamson, J G and W. S. Caste (1989) A survey of Florida Citrus nurseries. Proc. Fla. State Hort. Soc. 102: 78-82.
Williamson, J. G., W. s. Castle and K. E. Koch (1992) Growth and 14C-photosynthate allocation in citrus nursery trees subjected to one of three budforcing methods. J. Amer. Soc. Hort. Sci. 114. 180.
Williamson, J. G. and B. E. Maust (1995) Growth of budded, containerized citrusnursery plants when photosynthesis of rootstock shoots is limited. HortScience 30.1363-1365.
Williamson, J. G. (1997) Forcing method and time of rootstock shoot removal affect Growth of containerized citrus Nursery trees. HortTechnology 7 (3).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 144
GROWTH RESPONSES OF 'ROUGH LEMON'(Citrus Sinensis L.) ROOTSTOCKS TO DIFFERENT CONTAINER SIZES AND IRRIGATION FREQUENCY George Ouma Department of Horticul ture, Maseno university, P. 0. Box 333, Maseno, kenya., Telephone: (035) 51622 or 51620
Abstract Young 'rough lemon' (Citrus sinensis L.) seedlings were grown in container volumes of 1.7, 2.7 and 4.5 litres and subjected to three irrigation regimes namely: everyday irrigation, irrigation every other day and irrigation after three days, at Maseno University, Kenya. The media in the pots comprised 50 parts by volume of sand to 50 parts by volume of soil and it was amended with diammonium phosphate fertilizer (18, 46, 0, N, P, K) at a rate of 24gm per pot. Increasing container volumes significantly (P≤≤≤≤0.05) increased the dry weights of whole plants, roots and shoots, stem diameter, number of leaves, height of plants and canopy and vice versa for small container volumes. Conversely, increasing watering frequency only increased number of leaves, height of plants and height of canopy but had no effect (P≤≤≤≤0.05) on stem diameter, weight of whole plants, weight of roots and weight of shoots while the interaction between container volume and irrigation frequency was significant (P≤≤≤≤0.05) for stem diameter, weight of whole plants, number of leaves but not height of plants, height of canopy, weight of roots and weight of shoots.
Introduction: Commercial nursery produced are usually faced with two options, namely" to plant liners directly into market size containers or transplanting them into small containers and later transferring them into market size containers (upcanning). Upcanning is more labor intensive, it requires less space, provides more rapid shading of the container which cools the growth medium resulting in less crop failure (Beeson, 1991) In addition to the above, nursery owners are usually faced with the choice of various container sizes to use for their tree seedling production. Increased container size has been reported to cause increased canopy growth (Gillium et al.1984; Keever et al.1986; Keever and Cobb, 1987; Appleton and Whitcomb, 1983; Biran and Eliassaf, 1980), in pears (Pyrus caller:yana), Pecan (Carrya Illinoenis), Japanese euonymus (Eunymus Japonica Thumb) and other ornamental species respectively. Generally growing seedlings in small container sizes cause root restriction which causes reduced canopy growth (Hanson et al, 1987; Richards and Rowe, 1977; Ruff et al, 1987; Tschaplinski and Blake, 1985), reduced plant growth expressed as shoot length, fresh weight, dry weight accumulation and leaf area (Vizzotto, orietta and Costa, 1993), exhibited less expansion and caliper development of plants, reduced the number of primary shoots, total combined length of secondary shoots and total length of all shoots (Alvarez and Caula, 1993), reduced CO2 assimilation and leaf conductance (Riege and Marra, 1993), reduced leaf nutrient level s except N (Riege and Marra, 1993), reduced the dry weights of root, stem, leaves and fruit (Bar-Tal and Pressman, 1996). Root restriction reduces dry matter production but this has not been attributed to nutrient deficiency (Carmi and Heuer, 1981; Peterson et al.1991a; Peterson and Krizek, 1992; Reff et al., Heuer, 1981; Peterson et al, 1991a; Peterson and Krizek, 1992; Ruff et al, 1987) However, Bar- Tal et al .(1995) reported that root restriction reduced both dry matter production and K concentration in plant organs, indicating a possible K deficiency effect of restricting the roots. It has been argued however, that the reduction in plant growth under conditions of root restriction is not caused by nutrient deficiency but probably due to hormone synthesis and metabolism in the root system (Carmi and Heuer, 1981; Jackson, 1993' Peterson et al., 1991b; Richards and Rowe, 1977).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 145
Growth of containerized plants has also been reported to be slowed down by supraoptimal root zone temperatures (Graves, 1991; Martin and Ingram, 1991 a; Martin et al, 1989). Daily maximum temperatures greater than 50C have been recorded in rooting media adjacent to east and west containers walls and temperatures greater than 40C have been recorded for 6 to 10 hours daily during the summer at the centre of the medium profile (Martin and Ingram, 1988). The root and shoot growth responses of several plant species to supraoptimal root zone temperatures suggest that nursery operator should maintain root zone temperatures greater than 40C to attenuate injury and preserve satisfactory plant growth (Ingram et al, 1989; Martin et al, 1989; Wang et al, 1971; Yeager et al, 1991). Solar radiation striking container walls is the main source of thermal energy that causes supraoptimal rooting medium temperatures (Martin and Ingram, 1992). The most efficient cultural management strategies to correct high root zone temperatures therefore include those that reduce the influx as radiation energy into the rooting medium such as different container color, shape, close container spacing and more overhead shade (Ingram, 1981; Keever and Cobb, 1984; Laiche, 1985; Martin and Ingram, 1991 b). Different container dimensions also might alleviate supraoptimal rooting medium temperatures. Martin and Ingram (1993) reported that rooting medium temperatures were highest at the east and west container walls, half way down the container profile regardless of container height (20 to 50cm) or volume (10 to 70 litres). He also reported that the daily maximum medium temperatures at the centre was lower and occurred later in the day as container volume was increased due to increased distance to the container wall. The above literature review shows that no previous work has been conducted on 'rough lemon' rootstocks as far as container volume effect is concerned. Further, there is no previous work on the effect of container volume and irrigation frequency on the growth of 'rough lemon' citrus seedlings. The objectives of the present study were therefore, to investigate the effect of different container sizes and irrigation frequency on the growth of young citrus 'rough lemon'rootstocks. Materials and Methods Location of research site: The study was conducted at the Maseno University's department of Horticulture nurseries, Kenya. The nurseries are situated at an altitude of 1515 metres above sea level and are at a longitude of34°, 36° East and a latitude of 0° The soils comprise a complex of somewhat excessively drained, shallow, stony and rocky soils of varying color, consistence and texture (dystric regosols with ferralic combisols, lithic phase and rock outcrops). The area receives an annual rainfall of 1853mm. Experimental materials and their preparation: 'Rough lemon' (citrus sinensis L.) fruits were obtained from a farmer near Kisumu town, Kenya. The farmer had a history of high level management of his citrus grove, in terms of weeding, irrigation, fertilization, spraying against pests and diseases. The fruits were then transported to Maseno University, a distance of about 20 Km, and stored in the refrigerator for 2 days at a temperature of 5C. The fruits were subsequently washed and graded for uniformity of mass and freedom from blemishes. The fruits were then cut in half and seeds extracted from them in warm water at a temperature of 70C. The seeds were then dried briefly for two days in trays in readiness for planting in the nursery. Land Preparation. fertilization. planting and subsequent care: Nursery beds measuring 25 metres by 15 metres were well prepared to a fine tilth using hoes and machetes and well mixed with 8Kg of Farm Yard Manure and 100Kg per hectare of

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 146
Diammonium phosphate (18, 46, 0, N, P, K) fertilizer. The seeds were then planted on the nursery beds on loth May 1998 at spacing of 40cm by 15cm and a layer of grass mulch put between the holes. The beds were watered daily at 0800 hours and 1700 hours to saturation. Aldrin at 40% was applied at a rate of 5gm per Kg of seed for the control of cutworms. Insects were controlled by Dimethoate (dimethyl-5-(N-methyl carba-methyl) phosphorthiolothionate), 40% EC at a rate of 1 liter in 500 litres of water per hectare sprayed at 2 weeks interval. Diseases were controlled by Benomyl (methyl N- (l-butyl carbomoyl-2-benzimidazole) - carbamate) at a rate of 20 gm per 20 litres of water. Irrigation was done by watering cans using 16mm internal diameter hose and breaker nozzle. Transplanting of seedlings. treatments and experimental design: The treatments consisted of three container volumes namely. V 1 (1.7 litres), V2 (2.7 litres), V3 (4.5 litres) and three irrigation frequencies.growth1 (irrigating to saturation every day and twice a day), W2 (irrigating every other day and twice a day), W3 (irrigating every three days and twice a day). The design used was completely randomized design with three replications. After four months in the field the seedlings were transplanted into white plastic pots according to the treatments. The media put in the pots was sand and soil at a ratio of 1 to 1 (1 V 1 V) by volume. Diammonium phosphate (18, 46, 0, N, P, K) fertilizer was added to the pots at a rate of 24gm per pot. The pots were spaced 1 Metre from each other in a 30 percent shade structure made of timber and covered with grass. The size of the shade structure was approximately 10 metres in length, 40 metres in width and 10metres in height. Pests and diseases were controlled as described above for field seedlings while weeds were commenced one day after transplanting. Sampling statistical Analysis. Sampling was conducted destructively. The plants were uprooted 7 months after transplanting, from the containers after pouring some water on the media in the containers to soften it. The plants were washed free of soil, dried at 70C for 48 hours in an oven and before that they were weighed using a MettIer PE electronic scale (MettIer Instrument Corp; (Highstown, N. J.) and weighed expressed in kilograms. Whole plants were weighed. Roots and shoots were then separated and weighed. Other parts of the plant separated and weighed were leaves and stems. Heights of canopy and plants were measured using a tape measure. There were 4 plants per treatment per replication for each measurement. Statistical Analysis was done by the statistical analysis package (SAS Institute, Cary, N. C, 1988). Analysis of variance and separation of means by least significant difference method at five percent was carried out.
Results and Discussion: The number of leaves of the rootstock shoots was significantly (P≤0.05) increased by the volume of the container. (Table 1). The difference in number of leaves between the smallest and medium sized container was far much higher than between the medium and largest container. The interaction between watering frequency and container volume was significantly (P≤0.05) and the sma11er containers had plants with every few leaves particularly at very low watering frequencies as compared to the largest volumes. The large number of leaves at high container volumes may be attributed to increased dry matter production which cause increased plant growth rate (Carmi and Heuer, 1981 ; Jackson, 1993; Peterson and Krizeki, 1992; Ruff et at, 1987), reduced root restriction which increases soil volume and nutrient uptake, hormone synthesis and metabolism in the root system (Carmi and Heuer, 1981; Jackson, 1993; Peterson et al, 1991b; Richards and Rowe, 1977) and finally due to the fact that at greater container volumes the plants are shielded more from the highest medium temperatures than plants in small or narrow containers due to the greater distance to the container wall (Martin and Ingram, 1993) ensuring that water stays longer in the medium
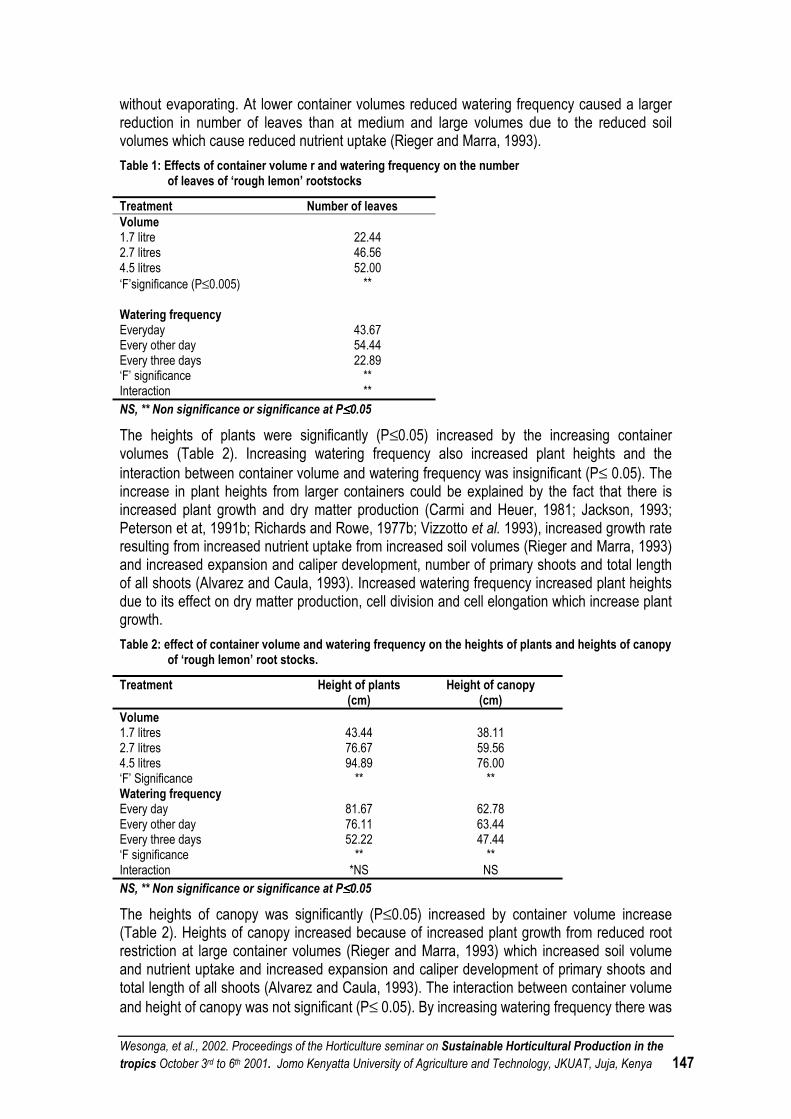
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 147
without evaporating. At lower container volumes reduced watering frequency caused a larger reduction in number of leaves than at medium and large volumes due to the reduced soil volumes which cause reduced nutrient uptake (Rieger and Marra, 1993). Table 1: Effects of container volume r and watering frequency on the number
of leaves of ‘rough lemon’ rootstocks
Treatment Number of leaves Volume 1.7 litre 22.44 2.7 litres 46.56 4.5 litres 52.00 ‘F’significance (P≤0.005) ** Watering frequency Everyday 43.67 Every other day 54.44 Every three days 22.89 ‘F’ significance ** Interaction ** NS, ** Non significance or significance at P≤≤≤≤0.05
The heights of plants were significantly (P≤0.05) increased by the increasing container volumes (Table 2). Increasing watering frequency also increased plant heights and the interaction between container volume and watering frequency was insignificant (P≤ 0.05). The increase in plant heights from larger containers could be explained by the fact that there is increased plant growth and dry matter production (Carmi and Heuer, 1981; Jackson, 1993; Peterson et at, 1991b; Richards and Rowe, 1977b; Vizzotto et al. 1993), increased growth rate resulting from increased nutrient uptake from increased soil volumes (Rieger and Marra, 1993) and increased expansion and caliper development, number of primary shoots and total length of all shoots (Alvarez and Caula, 1993). Increased watering frequency increased plant heights due to its effect on dry matter production, cell division and cell elongation which increase plant growth. Table 2: effect of container volume and watering frequency on the heights of plants and heights of canopy
of ‘rough lemon’ root stocks.
Treatment Height of plants (cm)
Height of canopy (cm)
Volume 1.7 litres 43.44 38.11 2.7 litres 76.67 59.56 4.5 litres 94.89 76.00 ‘F’ Significance ** ** Watering frequency Every day 81.67 62.78 Every other day 76.11 63.44 Every three days 52.22 47.44 ‘F significance ** ** Interaction *NS NS NS, ** Non significance or significance at P≤≤≤≤0.05
The heights of canopy was significantly (P≤0.05) increased by container volume increase (Table 2). Heights of canopy increased because of increased plant growth from reduced root restriction at large container volumes (Rieger and Marra, 1993) which increased soil volume and nutrient uptake and increased expansion and caliper development of primary shoots and total length of all shoots (Alvarez and Caula, 1993). The interaction between container volume and height of canopy was not significant (P≤ 0.05). By increasing watering frequency there was
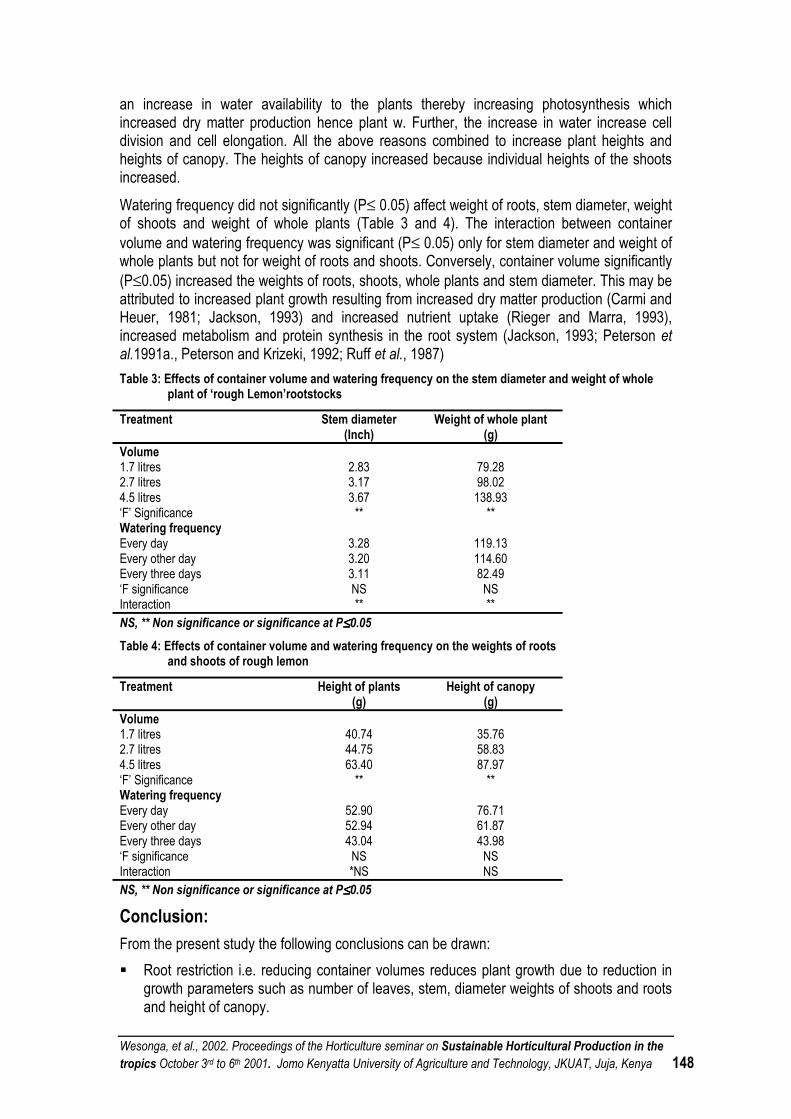
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 148
an increase in water availability to the plants thereby increasing photosynthesis which increased dry matter production hence plant w. Further, the increase in water increase cell division and cell elongation. All the above reasons combined to increase plant heights and heights of canopy. The heights of canopy increased because individual heights of the shoots increased. Watering frequency did not significantly (P≤ 0.05) affect weight of roots, stem diameter, weight of shoots and weight of whole plants (Table 3 and 4). The interaction between container volume and watering frequency was significant (P≤ 0.05) only for stem diameter and weight of whole plants but not for weight of roots and shoots. Conversely, container volume significantly (P≤0.05) increased the weights of roots, shoots, whole plants and stem diameter. This may be attributed to increased plant growth resulting from increased dry matter production (Carmi and Heuer, 1981; Jackson, 1993) and increased nutrient uptake (Rieger and Marra, 1993), increased metabolism and protein synthesis in the root system (Jackson, 1993; Peterson et al.1991a., Peterson and Krizeki, 1992; Ruff et al., 1987) Table 3: Effects of container volume and watering frequency on the stem diameter and weight of whole
plant of ‘rough Lemon’rootstocks
Treatment Stem diameter (Inch)
Weight of whole plant (g)
Volume 1.7 litres 2.83 79.28 2.7 litres 3.17 98.02 4.5 litres 3.67 138.93 ‘F’ Significance ** ** Watering frequency Every day 3.28 119.13 Every other day 3.20 114.60 Every three days 3.11 82.49 ‘F significance NS NS Interaction ** ** NS, ** Non significance or significance at P≤≤≤≤0.05 Table 4: Effects of container volume and watering frequency on the weights of roots
and shoots of rough lemon
Treatment Height of plants (g)
Height of canopy (g)
Volume 1.7 litres 40.74 35.76 2.7 litres 44.75 58.83 4.5 litres 63.40 87.97 ‘F’ Significance ** ** Watering frequency Every day 52.90 76.71 Every other day 52.94 61.87 Every three days 43.04 43.98 ‘F significance NS NS Interaction *NS NS NS, ** Non significance or significance at P≤≤≤≤0.05
Conclusion: From the present study the following conclusions can be drawn: Root restriction i.e. reducing container volumes reduces plant growth due to reduction in
growth parameters such as number of leaves, stem, diameter weights of shoots and roots and height of canopy.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 149
Reducing irrigation frequency reduces plant growth parameters such as number of leaves, height of plants, height of canopy whole plants but not stem diameter, weight of roots, shoots.
Root restriction under decreasing watering (irrigation) frequencies leads to more reduction in plant w. Applying water too frequently under increasing root restriction also leads to reduction of
plant w.
Acknowledgement: I am indebted to Maseno University for providing research facilities and funds and to Ms. Agatha for typing.
Liter ture Cited Alvarez, L. R. and Caula, B. (1993) Morphological effects of variable levels of rootrestriction on
'Loring' and 'Redhaven' Peaches in different environments. HortScicence28 (5). Appleton, B. and C. E. Whitcomb (1983) Effects of container size and transplant date onthe
growth of tree seedlings J. Environ. Hort. 1. 89-93. Bar- Tal, A. and E. Pressman (1996) Root restriction and potassium and calcium solution concentrations affect dry matter production, cation uptake and blossom end rot in green house Tomato. J. Amer. Soc. Hort. Sci. 121 (4).649-655
Bar-Tal, A., A. Feigin, S. Sheinfeld, R. Rosenberg, B. Sternbaum, T. Rylsk and E. Pressman (1995) Root restriction and N-NO3 solution concentration effects on tomato plant growth and fruit yield. Sci. Hort. 58. 91-103.
Beeson, R. C., Jr. (1991) Scheduling woody plants for production and harvest. HortTechnology 1. 30-36.
Beeson, R. C., Jr. (1993) Benefits of Progressively increasing container size during nursery production depend on Fertilizer Regime and Species. J. Amer. Soc Hort. Sci. 118 (6).752-756.
Bilderback, T. E. (1985)growth responses of Leyland Cypress to Media, N Application and container size after I and 2 growing seasons. J. Environ. Hort. 3. 132-135.
Biran, I. and I. Elliassaf (1980) The effect of container size and aeration conditions on growth of roots and canopy of woody plants. Scientia Hort. 12. 305-394.
Bish, E. B., D. J. Cantliffe; C. K. Chandler (1997) Transplant container shape and Strawberry Transplant Growth. HortScience 32. (3)
Chong, C.; G. P. Lumis; R. A. Cline and H J Reissmann (1987) Growth and Chemical composition of Populus deltoides X nigra grown in field grow fabric containers. J Environ. Hort. 5.45-48.
Chong, C.; G. P. Lumis and R. A. Cline (1989) Effect of fabric containers. Amer Nurseryman 170 (11).51-55.
Gillian, C. H.; G. S. Cobb and C. E. Evans (1984) Effects of nitrogen concentration and container size on growth of Pyrus calleyana 'Bradford' J. Environ. Hort. 2. 53-56.
Gilman, E. F. and R. C. Beeson, Jr.; G. W. Knox,; C. A. Neal and U. Yadav (1994) Micro-irrigation affects growth and root distribution of trees in fabric containers HortTechnology 4: 43-45.
Hanson, P. J.; R. K. Dixon and R. E. Dickson (1987) Effect of container size and shape on the growth of 'northern red' Oak seedling. HortScience 22: 1293-1295.
Ingram, D. L. (1981) Characterization of temperature fluctuations and woody plant growth in white poly bags and conventional black containers. HortScience 16. 762- 763.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 150
Ingram, D L.; U. Yadav and C. A. Neal (1987b) Production system comparisons for selected woody plants in Florida. HortScience 22 1285-1287.
Ingram, D. L.; C. A. Martin and J. M. Ruter (1989) Heat stress of container grown plants. IntI. Plant Prop. Soc. Proc. 39. 348-353.
Janet, C. R. Kjelgren and D. L. Hensler (1998) in ground fabric containers as an alternative Nursery Crop Production System. HortTechnology 8 (2).
Keever, G. S. and G. S. Cobb (1994) Container and production bed mulch effects on media temperatures and growth of 'Hershey' red azalea HortScience 19: 439-441.
Keever, G. J; G. S Cobb and R. MCDaniel (1986) Effects of container size, root pruning and fertilization on growth of seedlings of pecans. J Environ. Hort. 4. 11-13 Keever, G. J. and G S. Cobb (1987) Effects of container volume and fertility rate on growth of
two woody ornamentals. HortScience 22. 891-8943. Kjelgren, R.; C. Sphilman and B. R. Cleveland (1994) Effect of irrigation on crab apple growth
and water relations during field production with in ground fabric containers J. Environ. Hort. 12 108-111.
Laiche, A. J. (1985) Effect of time of spacing on the growth of container grown Ilex cornuta (DwarfBurford', Lindl. and Paxt; and Pittosprum tobira J. Environ Hort.3. 22-24.
Linder S.; M. L. Benson, B. J. Myers and R. J. Raison (1987) canopy dynamic and growth of Pinus radiata I. Effects of irrigation and fertilization during a drought. Con J. For. Res. 17. 1157-1165
Martin, C. A. and D. L. Ingram (1988) Temperature dynamics in black Poly containers. Southern Nurseryman Res. Confr. 33.71-74.
Lersel, M. V. (1998) Root restriction effects on growt hand development of (Salvia splendens) HortScience 32 (7). 1186.
Martin, C. A.; D. L. Ingram and T. A. NeIl (1989) Supraoptimal root zone temperature alters growth and photosynthesis of holly and elm. J. Arboricult. 15.272-276.
Martin, C. A. and D. L. Ingram (1991a) Root growth of Southern Magnolia following exposure to high root zone temperatures. HortScience 26.370-371.
Martin, C. A. and D. L. Ingram (1991b) Evaluation of thermal properties in container media and effect of irrigation on temperature J. Environ. Hort. 9 (1).24-28. Martin, C. A and D. L. Ingram (1992) Simulation modeling of temperatures in root container
media. J. Amer. Soc. Hort. Sci. 117: 571-577. Peterson, T. .I'\.; M. D Reinsel and D. T. Krizek (1991a) Tomato (Lycopersicon esculentum Mill.
cY. 'Bette Bush' Plant response to root restriction on I. alteration of plant morphology J. Expt. Bot. 42 1233-1240.
Peterson, T. A.; M. D. Reinsel and D. T. Krizek (1991b) Tomato (Lycopersicon esculentum Mill cy 'Better Bush') Plant response to root restriction I. Root respiration and ethylene generation. J. Expt. Bot. 42. 1241-1249.
Peterson, T. A. and D. T. Krizek (1992) A flow through hydropanic system for the study of root restriction J. Plant Nutri. 15.893-911.
G. W. Knox, C. A. Neal and U. Yadav (1994) Micro irrigation affects growth and root distribution of trees in fabric containers. HortTechnology 4. 43-45.
Graves, W. r. (1991) Growth and Iron content of three legume tree species at high root zone temperature J Arboriculture 17.313-317.
Martin, C A.; D. L Ingram (1993) container dimension Affects Rooting Medium Temperature Patterns. HortScience 28 (1): 18-19.
Rieger, M. and F. Marra (1993) Responses of young Peach trees to root confinement HortScience 28 (5).
Richards, D. and R. N. Rowe (1977) Effects of root restriction, root pruning and 6-benzyl amino purine on the growth of peach seedlings Ann. Bot. 41: 729- 740.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 151
Ruff, M. S.; D. L. Krizek; R. M. Mireckl and D. W. Inouye (1987) Restricted root zone development of Tomato. J. Amer. Soc. Hort. Sci. 112: 763- 769.
Ischaplinski, I. J. and I J. Blake (1995) Effects ofroot restriction on growth correlations, water relations and senescence of older seedlings. Physiol. Plant. 64 167- 176.
Vizzotto, G.; Orietta, L. and G. Costa (1993) Root restriction and photosynthetic response in a peach rootstock. HortScience 28 (5).
Yeager, I. H; R. H. Harrison and D. L. Ingram (1991). 'Rotundifolia' holly growth and N accumulation influenced by supraoptimal root zone temperatures. HortScience 26. 1387- 1388.
EFFECT OF CONTAINER VOLUME AND NITROGEN APPLICATION ON THE GROWTH OF YOUNG CITRUS SEEDLINGS George Ouma Department of Horticulture, Maseno University, P. 0. Box 333, MASENO, KENY A
Abstract Studies were conducted in a shade house at Maseno University, Kenya to determine the effect of Nitrogen fertilization and container volume on the growth of young citrus (Citrus sinensis) 'rough lemon' rootstocks. Nitrogen fertilizer was applied in the form of calcium ammonium nitrate to six months old seedlings at the rates of 0g/plant, 6g/plant, 12g/plant, 18g/plant and 24g/plant. The container volumes were 1.7 litres, 2.7litres and 4.5 litres. The media used was a mixture of soil and sand at a proportion of 50:50 (or half soil, half sand). The seedlings were watered every other day by hand to wetness. The results showed that nitrogen significantly increased the number of leaves, height of plants, height of canopy and weight of shoots. Volume of the containers significantly (P < 0.05) increased the number of leaves, height of plants, height of canopy but did not affect the weight of shoots and weight of roots as it increased. The interaction between volume and nitrogen was significant for number of leaves, height of plants, height of canopy, weight of whole plants, weight of roots and weight of shoots. The effects of N on the above parameters varied depending on the container size.
Introduction In commercial nursery container production there are two schools of thought namely planting of liners directly into market size containers or transplanting them into small containers and later transferring them into market size containers (upcanning). Upcanning is more labor intensive, it requires less space, provides more rapid shading of the container which cools the growth medium resulting in less crop failure (Beeson, 1991). In addition to the above, nursery owners are often faced with the choice of various container sizes to use for their tree seedling production. Generally, increased container size causes increased canopy growth (Gilliam et al, 1984; Keever et al, 1986; Keever and Cobb, 1987; Appleton and Whitcomb, 1983; Biran and Eliassaf, 1980). In pears (Pyrus calleryana), Pecan (Carrya illinoensis), Japanese euonymus (Eunymus JaQonica Thumb) and other ornamental species, respectively. Growing seedlings in small container sizes later cause root restriction which has often resulted in reduced canopy growth (Hanson et al, 1987; Richards and Rowe, 1977; Ruff et al, 1987; Tschoplinski and Blake, 1985), reduced plant growth expressed as shoot length, fresh weight and dry weight accumulation and leaf area (Vizzotto, Orietta and Costa, 1993 ), exhibited less expansion and caliper development of plants, reduced the number of primary shoots, total combined length of secondary shoots and total length of all shoots (Alvarez and Caula, 1993), reduced CO2 assimilation and leaf conductance (Riege and Marra, 1993 ), reduced leaf nutrient levels except N (Riege and Marra, 1993 ), reduced the dry weights of root, stem, leaves and fruit (Bar-Tal and Pressman, 1996). Although root restriction has been reported to reduce dry matter production it has been demonstrated that this reduction was not a result of nutrient deficiency.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 152
(Carmi, 1987) Conversely, Bar-Tal etal, (1995) reported that root restriction reduced both dry matter production and K concentration in plant organs, including the roots It has been reasoned that the production in plant growth under conditions of root restriction is not caused by nutrient deficiency but probably due to hormone synthesis and metabolism in the root system (Carmi and Heuer, 1981; Jackson, 1993; Peterson et al, 1991b; Richards and Rowe, 1977). Supraoptimal root zone temperatures also slow down the growth of containerized plants (Graves, 1991; Martin and Ingram, 1991a; Martin et al, 1989). Daily maximum temperatures greater than 50C have been reported in rooting media next to east and West container walls and temperatures greater than 40C have been recorded for 6 to 10 hour daily during the summer at the centre of the medium profile (Martin and Ingram, 1988). The root and shoot growth responses of several plant species to supraoptimal root zone temperature suggest that nursery operators should maintain root zone temperatures greater than 40C to attenuate root injury and preserve satisfactory plant growth (Ingram et al, 1989; Martin et al, 1989; Wang et al, 1971; Yeager et al, 1991). The main source of thermal energy that causes supraoptimal rooting medium temperatures is solar energy striking container walls (Martin and Ingram, 1992). The most efficient cultural management strategies to correct high root zone temperatures therefore consists of those that reduce the influx of radiation energy into the rooting medium such as different container color, shape, close container spacing and more overhead shade (Ingram, 1981; Keever and Cobb, 1984; Laiche, 1985; Martin and Ingram, 1991b). Supraoptimal rooting medium temperatures might also be alleviated by different container dimensions. It has been reported that rooting medium temperatures are highest at the East and West container walls, halfway down the container profile irrespective of container height (20 to 50cm) or volume (10 to 70 litres) (Martin and Ingram, 1993). Daily maximum medium temperatures at the centre is lower and occurs later in the day as container volume increased due to an increased distance to the container wall (Martin and Ingram, 1993). The effect of container volume on the growth of 'rough lemon' rootstocks, which is the main rootstock used by nursery owners in Kenya, has not been investigated. Further, no research, to the best of my knowledge has been conducted to study how varying container sizes interact with different N fertilizer levels to influence the growth of young citrus rootstock seedlings. The objectives of the present study was to investigate the effect of different container sizes and N fertilization on the growth of young citrus 'rough lemon' rootstocks.
Materials and Methods Location of research site. The study was conducted at the Maseno University's department of Horticulture nurseries, Kenya. The nurseries are located at an altitude of 1515 metres above sea level and are at a longitude of 34°, 36° East and a latitude of 00. The soils consist of a complex of somewhat excessively drained, shallow', stony and rocky soils of varying color, consistence and texture (dystric regosols with ferralic cambisols, lithic phase and rocky out crops). The area receives a fairly well distributed annual rainfall of 1853mm. Experimental Materials and Preparation. 'Rough lemon' (Citrus sinensis L.) fruits were obtained from a farm near Kisumu town, Kenya, after harvesting. Before harvesting the citrus trees had been well managed according to the recommended practices of weeding, irrigation, fertilization, crop protection to the etc. The fruits were subsequently transported to Maseno University's department of Horticulture laboratories where they were stored in the refrigerator for 2 days at a temperature of 5C. The fruits were then washed and graded for uniformity of mass and freedom from blemishes. The fruits were the cut in half and seeds extracted from them in warm water then dried briefly for two days in trays in readiness for planting in the field.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 153
Land Preparation, Fertilization, Planting and Subsequent Care Nursery beds measuring 25 metres by 15 metres were well prepared to a fine tilth using hoes and machetes and well mixed with 8Kg of Farm Yard Manure and 100 Kg per hectare of Diammonium phosphate (18, 46, 0, N, P, K) fertilizer. The seeds were then planted on the nursery beds on 15th May 1998 at spacing of 40cm by 15cm and a layer of grass mulch placed between the holes. The beds were watered daily at 0800 hours and 1700 hours to saturation F or pest control, Aldrin at 40% EC was applied at a rate of 5 gm per kilogram of seed for the control of cutworms while Dimethoate (dimethyl - 5 (N-Methyl carbomethyl) phosphorothiolothionate), 40% EC at a rate of 1 litre in 500 litres of water per hectare sprayed at 2 weeks interval w.as used to control other insect pests. Diseases were controlled by Benomyl (methyl N- (l-butyl carbomoyl-2-benzimidazole) -carbamate) at a rate of 20gm per 20 litres of water. Watering was done by watering cans using 16mm interval diameter hose and breaker nozzle. Transplanting of seedlings, Treatments and Experimental Design. After 4 months in the nursery the seedlings were transplanted into three sizes of containers according to treatments to be applied, which were V1 (1.7Iitres), V2 (2.7 litres), V3 (4 5 litres ). The containers comprised white plastic pots. The N treatment levels were 0,6, 12, 18 and 24g per pot of C. A. N. (Calcium Ammonium Nitrate, 26% N, Kenya Farmers Association). The experimental design was completely randomized design with three replications. The media placed in the pots were sand and soil at a ratio of 1 to 1 (1 V: 1 V) by volume. The pots were spaced 1 metre from each other in a 30 percent shade structure made of timber and covered with grass. The size of the shade structure was approximately 40cm in length 40m in width and 10m in height. Pests and diseases were controlled as described above for field seedlings while weeds were manually removed by hand. Watering w'as done by watering cans using 16mm internal diameter hose and breaker nozzle. Sampling and Statistical Analysis Sampling was conducted destructively. The plants were uprooted 7 months after transplanting from the containers after moistening the media in the containers to soften it. The plants were washed free of soil, dried at 700C for 48 hours in an oven and before that they were weighed using a Mettler PE electronic scale (MettIer Instrument Corp; Highstown, N. J.) and weight expressed in kilograms. Whole plants were weighed. Roots were then separated and weighed. Other weighings were for leaves and stems. Heights of canopy and plants were measured using a tape measure. There were 4 plants per treatment per replication for each measurement. Statistical Analysis was done by the statistical analysis package (SAS Institute, Cory, N. C., 1988). Analysis of variance was done and so was regression analysis and separation of means by least significant difference methods at five percent.
Results The number of leaves increased with increasing container volumes but Nitrogen fertilizer levels had no effect (P ≤ 0.05) The interaction between Nitrogen and container volume did not cause any significant (P ≤ 0.05) effect on the number of leaves (Table 1). Heights of plants were increased by increased container volumes up to the larger volume then decreased in contrast, increasing Nitrogen fertilizer levels consistently increased the height of plants (Table 1 ). The increases in plant heights were both linear and quadratic. The interaction between container volume and Nitrogen fertilizer was significant. Both heights of canopy and stem diameter were increased by increasing container volumes and like stem diameter, the farmer increased consistently up to the largest volume. In contrast,
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 154
only the height of canopy was increased by increased container volumes. Stem diameter was unaffected by the increasing Nitrogen fertilizer levels. Nitrogen fertilizer linearly and quadratically increased the heights of canopy and interacted significantly (P ≤ 0.05) with container volume (Table 2). Table 1: Effect of different container volumes and levels of Nitrogen on the height of plants and number of
leaves of rough lemon rootstocks
Treatment Number of leaves Height of Plants (cm) Volume 1.7 2.7 4.5 Significance (P ≤ 0.05) Nitrogen 0 6 12 18 24 Significance (P ≤ 0.05) Regression Linear Quadratic Interaction
233.69 264.11 334.78
**
165.67 154.89 178.00 184.00 187.00
NS NS NS
65.00 91.44 85.66
**
44.22 47.22 55.56 65.20 80.50
** ** **
NS, ** Not Significant or Significant at P ≤≤≤≤ 0.05 Table 2: Effect of different container volumes and levels of Nitrogen on the height of canopy and stem
diameter of rough lemon (Citrus sinensis L.) rootstocks
Treatment Height of canopy Stem diameter (cm) Volume 1.7 2.7 4.5 Significance (P ≤ 0.05) Nitrogen 0 6 12 18 24 Significance (P ≤ 0.05) Regression Linear Quadratic Interaction
64.33 72.11 51.67
**
44.22 47.22 55.56 59.02 63.55
**
** ** **
56.37 80.82 69.37
**
50.11 56.22 58.01 62.20 61.50
NS
NS NS NS
NS, ** Not Significant or Significant at P ≤≤≤≤ 0.05
The weight of whole plants increased quadratically as the container volumes increased and so was the weight of roots. The difference between the weights of whole plants in 1.7 litre and 2.7 litre one was far much bigger compared to between 2.7 litres and 4.5 litres (Table 3). The interactions between the two factors above were significant only for weight of whole plants but not weight of roots.
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 155
Table 3: Effect of different container volumes and levels of Nitrogen on the weight of whole plants and roots of ‘rough lemon’ rootstocks
Treatment Weight of whole plant (gm) Weight of roots (gm) Volume 1.7 2.7 4.5 Significance (P ≤ 0.05) Nitrogen 0 6 12 18 24 Significance (P ≤ 0.05) Regression Linear Quadratic Interaction
94.86 130.41 126.53 ** 47.20 80.64 94.59 117.24 100.11 ** NS ** **
42.15 48.54 54.57 ** 15.82 32.41 46.14 59.20 59.20 ** NS ** **
NS, ** Not Significant or Significant at P ≤≤≤≤ 0.05
The weights of shoots increased then decreased at the biggest container volume. Similarly Nitrogen fertilizer also increased the weights of shoots up to the second highest Nitrogen level then decreased. Container volume interacted significantly with Nitrogen fertilizer levels (Table 4). Table 4: Effect of different container volumes and levels of Nitrogen on the weight
of shoots of rough lemon rootstocks
Treatment Weight of shoots Volume 1.7 2.7 4.5 Significance (P ≤ 0.05) Nitrogen 0 6 12 18 24 Significance (P ≤ 0.05) Regression Linear Quadratic Interaction
56.37 80.82 69.37
**
31.28 48.50 59.10 73.14 68.05
**
** ** **
NS, ** Not Significant or Significant at P ≤≤≤≤ 0.05
Discussion Increasing container volumes increased the growth parameters measured or determined in the present study, with very few exceptions indeed. These were number of leaves, heights of plants, heights of canopy, stem diameter, weight of whole plants, weights of roots and shoots. These observations can be attributed to the following factors: The large container volumes had reduced root restriction, which led to increased dry matter
production, which increased plant growth rate (Carmi and Heuer, 1981; Jackson, 1993; Peterson et at, 1991b; Richards and Rowe, 1977; Peterson and Krizeki, 1992; Ruff et at, 1987; Vizzotto, et at, 1993).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 156
Reduced root restriction in large containers increases soil volume and nutrient up take, hormone synthesis and metabolism in the root system. (Carmi and Heuer, 1981; Jackson, 1993; Peterson et al, 1991b; Richards and Rowe, 1977; Rieger and Marra, 1993)
At larger volumes of container, there was increased expansions and caliper development, number of primary shoots and total length of all shoots (Alvarez and Caula, 1993), which caused increased heights of plants and canopies.
At greater volumes the plants are shielded more from the highest medium temperatures than plants in small or narrow containers due to the greater distance to the container wall (Martin and Ingram, 1993) ensuring that the water stays longer in the medium without evaporating.
Increasing Nitrogen fertilizer levels increased the heights of plants, heights of canopy, weights of whole plants, weights of roots and shoots but not stem diameter and number of leaves. This increase was more at the larger containers generally. It appears that the beneficial effects of reduced root restriction in larger containers are enhanced at higher N fertilizer levels. It would be interesting to find how different levels of different fertilizers such as P and K would behave under the present experimental circumstances. Studies are required to test these latter factors with respect to varying container volumes. High N fertilizer application in itself increases plant growth due to increased plant growth promoting processes such as cell division, cell enlargement, metabolic processes, etc Stem diameter and the number of leaves were however, not affected by N fertilizer levels. It may be due to the fact that they are not sensitive or responsive to increasing N levels. Therefore, at higher container volumes there are increased soil volumes which cause increased nutrient uptake and if soil nutrient levels are increased by N fertilizer applications at increased levels there will be more nutrients available for the plant growth processes. Most N fertilizer responses were quadratic. This implies that enough of fertilizer needed by the plants has been added and any more additions are not beneficial for plant growth.
Conclusions: From the results of the present study it can be concluded that When container volumes are increased there is usually increased in plant growth
parameters such as heights of plants, heights of canopy, stem diameter, number of leaves, weight of whole plants, weight of shoots and roots.
Increasing Nitrogen levels from 0gm to 24gm increases weights of shoots and roots, heights of canopy, weights of whole plants, heights of plants but not stem diameter and number of leaves.
Increasing container volumes lead to even higher weights of shoots and roots, heights of canopy and plants, etc when N fertilizer levels increases.
Acknowledgement: I am indebted to Maseno University for financial and equipment support for this project and to Ms Agatha for typing this manuscript. Literature Cited Alvarez, L. R. and Caula, B. (1993) Morphological effects of variable levels of root restriction on
'Loring' and 'red haven' Peaches in different environments. HortScience 28 (5). Appleton, B. and C. E. Whitcomb (1983) Effects of container size and transplant date on growth
of tree seedlings. J. Environ. Hort. 1. 89-93. Bar-Tal, A. and E. Pressman (1996) Root restriction and potassium and calcium solution
concentrations affect dry matter Production, Cation uptake and blossom end rot in Green house Tomato. J. Amer. Soc. Hort. Sci 121 (4): 649-655.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 157
Bar-Tal, A; A. Feigin; S. Sheinfeld; R. Rosenberg, B Sternbaum; T. Rylsk and E. Pressman (1995) Root restriction and N-NO3 solution concentration effects on tomato plant growth and fruit yield. Sci. Hort. 58.91-103.
Beeson, R. C. Jr. (1991) Scheduling woody plants for production and harvest. HortTechnology 1.30-366.
Beeson, R. C., Jr. (1993) Benefits of progressively increasing container size during nursery production depend on fertilizer regime and species. J Amer. Soc. Hort. Sci. 118 (6): 752-756. 7.
Bilderback, T. E. (1985) Growth responses ofLeyland Cypress to media, N application and container size after 1 and 2 growing seasons. J. Environ. Hort. 3.132-135.
Biran, I. and I. Elliassaf (1980) The effect of container size and aeration conditions on growth of roots and canopy of woody plants. Scientia. Hort 12.385-394.
Bish, E. B.; D. J. Cantiffe; C. K Chandler (1997) Transplant container shape and strawberry transplant growth. HortScience 32 (3).
Chong, C.; G. P. Lumis; R. A. Cline and H. J. Reissmann (1987) Growth and chemical composition of Populous deltoides X nigra grown in field grown fabric containers. J. Environ. Hort. 5. 45-48.
Chong, C.; G. P. Lumis and R. A. Cline (1989) Effect of fabric containers. Amer. NurseryMan. 170 (11).51-55.
Gilliam, C. H.; G. S. Cobb and C. E. Evans (1994) Effects of Nitrogen concentration and container size on growth of Pyrus calleryana 'Bradford' J. Environ. Hort. 2. 53-56
Gilman, E. F. and R. C. Beeson, Jr.; G W. Knox; C. A. Neal and U. Yadav ( 1994) Micro-irrigation affects growth and root distribution of trees in fabric containers. HortTechnology 4.43-45.
Hanson, P. J.; R. K. Dixon and R. E. Dickson (1987) Effect of container size and shape on the growth of northern red oak seedling HortScience 22. 1293-1295
Graves, W. R. (1991) Growth and Iron content of three legume tree species at high root zone temperature. J Arboriculture 17.313-317.
Ingram, D. L.; U. Yadav and C. A. Neal (1987) Production system comparisons for selected woody plants in Florida. HortScience 22. 1285-1287.
Ingram, D. L.; C. A Martin and J. M Ruter (1989) Heat stress of container grown plants. Intl. Plant Prop. Soc. Proc. 39. 348-353.
Janet, C. Cole; R. Kjelgren and D. L. Hensley (1998) In ground fabric containers as an alternative Nursery crop production system. HortTechnology 8 (2).
Keever, G. J. and G. S Cobb (1984) Container and Production bed mulch effects on media temperatures and growth of ‘Hershey’ red azalea. HortScience 19. 439-441.
Keever, G. J.; G. S. Cobb and R. McDaniel (1986) Effects of container size, root pruning and fertilizer on growth of seedlings of pecans. J. Environ. Hort. 41. 11-13.
Keever, G. J. and G. S. Cobb (1987) Effects of container volume and fertility rate on growth of two woody ornamentals. HortScience 22. 891-893.
Kjelgren, R.; C. Spihlman and B. R. Cleveland (1994) Effect of irrigation and 'crab' Apple growth and water relations during field production with in ground fabric containers. J. Environ. Hort. 12: 108-111.
Iaiche, A. J. ( 1985) Effect of time of spacing on the growth of container grown (Ilex cornutarum ('DwarfBurford'), Lindi and Paxt and Pittosporum tobira. J. Environ. Hort. 3: 22-24
Linder, S.; M. L. Benson, B. J. Myers and R. J. Raison (1987) Canopy dynamics and growth of Pinus radiata I. Effects of irrigation and fertilization during a drought. Can. J. for. Res. 17. 1157-1165.
Lersel, M. V. ( 1998) Root restriction effects on growth and development of (Salvia splendens) HortScience 32. (7): 1186.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 158
Martin, C. A. and D. L. Ingram (1988) Temperature dynamics in black poly containers. Southern Nurseryman Res. Confr. 33. 71- 74.
Martin, C. A.; D. L. Ingram and T. A NeIl (1989) Supra optimal root zone temperature alters growth and photosynthesis of holly and elm J. Arboricult. 15. 272-276
Martin, C. A. and D. L. Ingram (1991a) Root growth of Southern Magnolia following exposure to high root zone temperatures. HortScience 26. 370-371.
Martin, C. A. and D. L. Ingram (1991b) Evaluation of thermal properties in container media and effect of irrigation on temperature J Environ. Hart. 9 (1).24-28.
Martin, C. A. and D. L. Ingram (1992) Simulation modeling of temperatures in root container media. J. Amer. Soc. Hort. Sci. 117. 571-577.
Peterson, T. A.; M. D. Reinsel and D. T. Krizek (1991b) Tomato (Lycopersicon escutentum) Mill cv. 'Betterbush'. Plant response to root restriction 1 Root respiration and ethylene generation J. Expt. Bot. 42. 1241-1249.
Peterson, T. A. and D. T. Krizek (1992) A flow through hydroponic system for the study of root restriction. J Plant Nutri. 15. 893-911.
Rieger, M. and F. Marca (1993) Responses of young peach trees to root confinement. HortScience 28 (5).
Richards, D. and R. N. Rowe (1977) Effects of root restriction, root pruning and 6-benyl amino purine on the growth of peach seedlings Ann.Bot. 41: 729-740.
Ruff, M. S.; D. L. Krizek; R. M. Mireck and D. W. Inouye (1987) Restricted root zone volume; Influence on growth and development of Tomato. J. Amer. Soc. Hort. Sci. 112. 763- 769.
Tschaplinski, T. J. and T. J. Blake (1985) Effects ofroot restriction on growth correlations, water relations and senescence of older seedlings. Physiol plant 64: 167-176.
Vizzotto, G.; Orietta, L. and G. Costa (1993) Root restriction and photosynthetic responses in a peach rootstock. HortScience 28 (5).
Yeager, T. H.; R. H. Harrison and D. L. Ingram (1991) 'Rotundi foliar holly growth and N accumulation influenced by supra optimal root zone temperatures. HortScience 26. 1387-1388.
ACHIEVEMENTS IN ROOT AND TUBER CROPS IMPROVEMENT RESEARCH AND FUTURE PROSPECTS IN SOUTHWESTERN ETHIOPIA Amsalu, N.1 and Edossa, E.2 1Ethiopian Agricultural Research Organization, Jimma Research Center, P.O. Box 192, Jimma, Ethiopia 2Ethiopian Agricultural Research Organization, Nazareth Research Center, P.O. Box 431, Nazareth Ethiopia
Abstract Millions of people in Ethiopia depend on root and tuber crops as their major or supplementary food. Root and tuber crops give greater yield per unit area than the major grain crops used as basic food. These crops can support greater density of population than other food crops when produced on small plots of land. Research on root and tuber crops at Jimma was started nearly three decade ago as part of the coffee diversification program. The major areas of research given attention since the inception of the program were collection and/or introduction, maintenance, screening of adaptable and high yielding varieties identification of agronomic and compatible management practices. Taro, yam, cassava, aerial yam, sweet potato and Anchote were the major crops given attention. Over the past few years, although not as has been expected, some remarkable accomplishments have been made with regard to variety improvement and identification of appropriate agronomic and cultural practices. Therefore, three taros, three cassava, and three yam varieties were identified as high yielding, adaptable and quality materials in southwestern Ethiopia. These materials have been multiplied at Jimma

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 159
and distributed to local farmers, Agricultural Development offices and other NGOs involved in rural development programs. Improved cultural practices such as plant population/spacing, type of planting material, time of planting have been developed and recommended. Improved taro cultivars and their production recommendations were demonstrated in different agro ecologies where the crop is gown. Planting materials of taro, yam, and cassava have been also distributed to the users. In addition, Agricultural Development Agents and farmers have been trained on production and utilization technologies.
AGRONOMIC STUDIES FOR SUSTAINABLE PINEAPPLE (ANANAS COMOSUS) PRODUCTION IN SOUTHWESTERN ETHIOPIA Nebiyu, A.1 and Etissa, E.2 1Ethiopian Agricultural Research Organization, Jimma Research Center, P.O. Box 192, Jimma, Ethiopia 2Ethiopian Agricultural Research Organization, Nazareth Research Center, P.O. Box 431, Nazareth Ethiopia
Abstract The suboptimal agronomic and cultural techniques (type and size of planting material, optimum spacing, weed management and stand regulations) practiced by pinapple growers in Ethiopia are among the different factors that contributed to the low yield of the crop. In this paper, results of the major agronomic research activities conducted for sustainable pineapple production are discussed. To determine the type of planting material to be used for successful growth, development and sustainable yield of pineapple, different planting materials (slips, suckers and crowns) were compared. Results have shown that slips gave significantly higher marketable fruit yield (49.76 t/ha) than the suckers and crowns. On the other hand, in the study to determine the size of slips, different sized slips were compared for their effect on fruit yield. Hence, 40-50cm-sized slips were found best for an optimum yield. In addition, to determine the total plant population per hectare spacing trial was conducted. Results have shown that spacing combination of 90cm x 60cm x 30cm (1,444,444 plants/ha) was found optimum. These agronomic research results have been recommended to the subsistence farmers, private and state farms. On the other hand, trials on an integrated weed management practices and stand regulation are on progress.
EFFECT OF NITROGEN FOR SUSTAINABLE VEGETABLE PRODUCTION: RECENT DEVELOPMENTS Okeno, J. A.
Department of Horticulture, Moi University, P. 0. Box 1125, Eldoret, Kenya. E-mail: [email protected]
Abstract Sustainable food production or sustainable agriculture is a concept defined, among others, as 'a system that, over a long term, enhances environmental quality and resource base on which agriculture depends, provides for basic human food and fibre needs, is economically viable, socially acceptable, and enhances the quality of life for farmers and society as a whole'. A sustained food production, therefore, encompasses plant nutrient supply, a resource base of ecological aspect. Nitrogen (N) supply is the most universal deficient for optimum crop production considering its abundance in plants, central role in plant functions and reactivity in the biosphere. N is responsible for lush vegetative growth and a dark green color. N together with magnesium is one of the two soil-derived constituents of chlorophyll (C33H72O5N4Mg). Adequate N for plants promotes aerial vegetative growth, increases the top/root ratio, and is essential for fruit and seed formation. Being an essential constituent of amino acids, N is required in protein synthesis, making up 12 to 19% of various proteins and averaging about 16% by weight. In terms of mass, N in plant materials is often found in larger quantities than any other elements. In vegetable production, N supply affects both yield and quality characteristics. For example, in broccoli, N increased yield of head to an optimum level of 240 kg N ha-1 increased both nitrate and chlorophyll contents but decreased vitamin C content. In lettuce, N increased yield and nitrate content but did not affect vitamin C content.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 160
THE PARTITIONING OF ASSIMILATES IN FABA BEAN: A COMPARISON OF THE DETERMINATE AND INDETERMINATE GROWTH HABIT P.W. Mathenge Department of Crop Production and Seed Technology, Moi University, P.O. Box 1125, Eldoret
Abstract The manner in which crop plants partition and distribute photoassimilates at various stages of development, especially reproductive stage, is of immediate interest not only to plant scientists but also to farmers. Crop variants that partition more photoassimilates into yield components are to be preferred. The assimilate partitioning of an indeterminate and two determinate genotypes of field bean (Vica faba L.) was compared at the vegetative, flowering, early pod formation, mid pod filling and the late pod filling stages. At the vegetative stage, the bulk of 14C-photo assimilates was exported to the roots, the apical meristem and the stem. The distribution pattern at the flowering stage was only slightly different to that observed at the vegetative stage, which was indicative that at this stage the flowers were weak sinks for 14C-photoassimilates. However, less 14C-photoassimilate was exported to the vegetative organs after pod-set. Over 75% of 14C-photoassimilates translocated from the fed leaf was found in the pods at the pod-fi1ling stage. The pattern of 14C-photoassimilate distribution was similar at the vegetative stage in the two growth types studied but changed at the stage when the apical meristem of the determinate plant became reproductive. However, the physiological advantages of determinate type to produce more yield were cancelled in that though tillers evidently received photoassimilates, they were infertile. Key words:Vicia faba, assimilate partitioning, 14C, determinate genotype.
Introduction A number of mutants of the field bean with a determinate growth habit and which may have potential for the development of this crop are known and selective plant breeding of determinate varieties of field bean has been conducted in recent years. In one case the parent material, an X-ray induced mutant ti from the Swedish cultivar Primus (Sjodin, 1971) has bred true for the determinate habit enabling crosses to be made with both field and broad bean to produce a plant which has several distinct advantages over the indeterminate type (Chapman and peat, 1978). These advantages include. a shorter plant less prone to lodging, a shorter life cycle and an early pod set which results in aphid avoidance, ie there is an absence of a growing apex during the period of highest potential aphid infestation and a harvest index which is as high or higher than that of the indeterminate plant. Although Vicia faba has been the subject of a number assimilate partitioning studies (See Crompton et al., 1981) to date only a limited number of studies have been conducted on determinate material (Austin et al., 1981; Baker et al., 1983). These investigations showed that the inherent advantage of the terminal inflorescence may be lost when tillers and axillary branches, which develop asynchronously with the main shoot, remain infertile. Under such conditions these branches compete for assimilates as an alternative sink, in much the same manner as does the continued growth of the apex in the indeterminate habit. Furthermore, the determinate material is no more efficient at fixing CO2 than the indeterminate, but the individual leaves on the determinates are photosynthetically active for a longer period due to delayed senescence (Baker et al., 1983). In this paper the source: sink relationships of the two growth habits have been further characterised to provide physiological guidelines for future breeding strategies, in particular to remedy the effective source limitation of the determinate habit.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 161
Materials and Methods Two determinate (642;645) and one indeterminate (644) types of field bean were grown under glass in individual pots containing potting compost, using a randomised four block design and in the field at a density of 21 plants m-2. Plants were fed with 14CO2 at the vegetative stage, flowering, early pod set, mid-pod filling and late pod filling stages. Experimental plants were selected for uniformity in batches of three. 14CO2 feeding was done on a sunny day when photosynthetically active radiation (PAR) would not be limiting photosynthesis. Plants at the vegetative stage were fed with 14CO2 at the fifth leaf, plants at flowering and early pod formation at the leaf below the first flowering node and plants at mid and late pod filling at the leaf below the first podding node. Five ul of NaH14CO3 containing 10uCi was applied at the centre of a paper disc a few days before feeding and allowed to dry. This improved the efficiency of 14CO2 transfer to the fed leaf. At feeding, the paper disc containing NaH14CO3 was transferred to a transparent polythene bag, which was used as a 14CO2 chamber. The selected leaf was enclosed in the chamber and carefully secured to the leaf petiole with a wire and a piece of sponge to protect the petiole from damage. Three to five ml of 1% acetic acid were introduced into the chamber with a syringe and needle. The needle hole was carefully sealed with transparent cellotape to avoid leakage of 14CO2. Care was taken to release acetic acid onto the centre of the paper disc to ensure complete liberation of 14CO2 for 30 minutes before removing the polythene bag. Experimental plants were left to translocate 14C-photoassimilates for 6 hours and then harvested to provide samples of various organs (l.fed leaf, 2. Leaves above fed leaf, 3. Apical meristem, 4.flowers,5.flowers and young pods, 6.pods,7.stem above fed leaf, 8.leaves below fed leaf, 9.stem below fed leaf, 10.branches, 11.roots). These samples were dried to constant weight at 60°C, ground to powder and sub-samples of 0.2-0.5g weighed for combustion. Combustion was performed in an OX-400 Biological Material Oxidiser at 73Oc and 930°C in the catalyst and combustion zones respectively. 14CO2, trapped directly in a scintillation cocktail (27% methylated spirit, 27% phenylethylamine and 46% NE 233) was determined on an A211 Rackbeta liquid scintillation counter.
Results 14C- Photoassimilate Partitioning The pattern of 14C-photoassimilate partitioning and the related specific activities is presented in Tables 1 to 6. These results clearly indicate that the pattern of distribution of the 14C-photoassimilates varied with the plant growth habit and the stage of development. However, the general pattern of partitioning was basically similar in that the most actively growing organs not only imported more 14C-assimilates but also exhibited higher specific activities per unit of dry matter. At the vegetative stage (Tables 1 and 2), the bulk of 14C-photoassimilates was exported to the roots, stem and the shoot apex. For the data presented in Table 1, obtained from plants treated in the summer of 1988, it was observed that the two determinate growth types exhibited a similar pattern in distribution. The roots, stem and the shoot apex were the major sinks for assimilates. The branches in the determinate, 642, received more assimilates than those of type 645. On the other hand, the distribution in the indeterminate type was similar to that of the determinate types. In the data presented in Table 2, for the summer of 1989, again the pattern of assimilate distribution of the two growth habits was similar in that most of the assimilates were exported to the roots, stem and the shoot apex. The roots in the indeterminate type were observed to be particularly strong sinks at the time of feeding. There was an indication,

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 162
especially in Tables 1 and 2, that younger leaves attracted more 14C-phtoassimilates than older ones. Table 1: The % distribution of translocated 14C-photoassimilates and the activities min-1g-1 dry matter (in
DPM) for the two determinate growth habits 645(a) and 642(b) and the indeterminate type 644(C) at the vegetative stage in summer 1988. Values in parenthesis are the activities. Values are means of 3 replicates
Plant 645 642 644 Plant type a b C
Organ 1 - - - 2 10.8 (37684.7) 5.8 (19072.1) 15.8 (50515.1 3 22.7 (18244.4) 20.5 (263297.4) 27.4 (269818.5) 4 - - - 5 - - - 6 - - - 7 6.3 (24560.9) 6.0 (5431.1) 3.2 (55742.3) 8 4.4 (10529.1) 3.9 (491280.0) 4.1 (14713.6) 9 19.16 (22324.8) 24.8 (83442.8) 17.0 (60104.6)
10 2.58 (28099.9) 13.3 (63333.3) 7.5 (54288.9) 11 33.98 (25398.0) 25.59 (39437.6) 24.95 (30640.50)
Table 2: The % distribution of translocated 14C-photoassimilates and the activities in counts min-1g-1 dry matter (in parenthesis) for the two determinate growth habits 645(a), 642(b) and the indeterminate type 644(C) at the vegetative stage in summer 1989. Values are means of 3 replicates
Plant 645 642 644 Plant type a b C
Organ 1 - - - 2 6.8 (2325.0) 3.8 (1124.4) 1.87 (707.5) 3 22.9 (46585.8) 34.8 (11400.7) 8.05 (8602.3) 4 - - - 5 - - - 6 - - - 7 1.9 (5308.7) 2.4 (1461.1) 1.3 (4317.1) 8 4.5 (1898.6) 14.3 (1102.4) 7.2 (745.0) 9 23.3 (11794.7) 21.1 (67.8.1) 3.9 (6656.3)
10 4.3 (4144.2) 2.9 (1834.1) 3.5 (3910.3) 11 36.3 (12586.3) 22.18 (3010.9) 74.1 (89323.5)
Rapidly growing organs, i.e. shoot apex, roots and the stem had higher specific activities per unit of dry matter. The relatively higher activity that was observed in the stem than in several other organs may be an indication that some storage was taking place. Crompton et al. (1981) working with the indeterminate cultivar, ‘Maris Bead’, noted that the direction in which assimilates moved varied with the stage of development of the plants. It was observed that at pod set, i.e. at the vegetative and the flowering stages, most of 14C-assimilates were exported downwards from the expanded leaves to the lower stem and roots with little going to the growing shoot. These results are to some extent divergent from the results presented here, especially with reference to the amount going to the shoot apex which was found to not only import quite a considerable percentage of 14C-photoassimilates but also had, in most cases, higher specific activity The results presented in this study are consistent with those of Belikov (1973) on vegetative soybean plants in which lower leaves exported assimilates to the roots, midstem leaves to the roots and shoot apex while upper leaves capable of exporting assimilates were entirely under the influence of the shoot apex. Hume and Criswell (1973) also working with soybeans observed that at the vegetative stage

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 163
14Cphotoassimilates were incorporated into structural leaf and stem tissues. In young corn plants, 3 to 6 weeks old, the bulk of assimilates was exported mainly to the roots after 24 hours and young expanding leaves retained most of the label (Hofstra and Nelson, 1969). The relatively high percentages of 14C-assimilates observed in the stem, particularly the stem below the fed leaf, may reflect that some storage was taking place Ishag (1973) noted that the loss in dry matter of an indeterminate growth type was associated with increased dry weight in seed, and that the slow growth rate of the stem at pod filling indicated that dry matter was translocated from the stem to seeds or, as Allison and Watson (1966) observed in maize, more dry matter was lost from the stem by respiration than could not be replaced, because assimilates were preferentially diverted to the grain. That the stem might be the site of storage and remobilisation is indicated by the work of Peel and Weatherley (1962) who showed that with the change from light to dark, the source of sugar for phloem translocation in Salix species changed from the leaf to the stem tissue which indicates that the stem has an important role in the carbon economy of the plant. The pattern of 14C-photoassimilate partitioning at the flowering stage reveals that it was dependent on the growth habit (Table 3). This difference is due to the fact that the shoot apex of the determinate plants was now reproductive and hence most of the assimilates were exported to the roots, stem and branches, unlike in the indeterminate growth habit where the shoot apex remained vegetative. The low percentage of assimilates exported to the flowers is an indication that they are weak sinks which could be a significant implication on the pod set, and hence the often observed flower abscission in this crop (Kambal,1969; Jacquiery and Keller, 1980; Gates, Smith, White and Boulter, 1983). Tillers and axillary branches were now growing vigorously and received more assimilates, especially in the determinate plants. This observation concurs with that of Baker et al.(1983) in which they noted that in the determinate plant, the tillers and branches on the main stem behave in a similar fashion to the indeterminate apical growth in that they are both sources or sinks for assimilates to and from the main stem. On the other hand, most of the 14C-assimilates in the indeterminate plant were exported to the stem, roots and shoot apex. For the determinate plants, the stem below the fed leaf had the highest specific activity while the shoot apex had more activity in the determinate plant. Although flowers received less assimilates, their specific activity indicates that they were more active than some sinks, especially in the determinate growth habit. Table 3: The % distribution of translocated 14C-photoassimilates and the activities in counts min-1g-1 dry
matter (in parenthesis) for the two determinate growth habits 645(a), 642(b) and the indeterminate type 644(C) at the flowering stage. Values are means of 3 replicates
Plant 645 642 644 Plant type a b C
Organ 1 - - - 2 1.6 (3.15.5) 5.6 (258.3) 3.0 (213.4) 3 - - 19.0 (7212.6) 4 2.0 (1067.6) 9.1 (1404.8) 2.7 (681.1) 5 - - - 6 - - - 7 0.7 (325.8) 1.0 (186.9) 10.7 (3397.5) 8 3.2 (382.6) 10.0 (285.5) 1.9 (138.2) 9 28.8 (5263.1) 22.8 (2669.3) 26.4 (2100.9)
10 24.8 (4570.0) 24.4 (1024.7) 9.3 (2100.9) 11 38.4 (2173.9) 28.8 (589.5) 27.1 (833.9)

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 164
At flowering and early pod growth (Table 4a,b, and c) the reproductive organs (pods and flowers) were the main sinks for 14C-photoassimilates. Clearly the emphasis on assimilate demand had shifted to the pods and now less was exported to the roots, stem and shoot apex. In the determinate types, over 70 and 50% of the assimilates were exported to the reproductive organs in the determinate and the indeterminate types, respectively. More assimilates were found in the stem of the determinate types, while diminishing amounts were exported to the other organs. Specific activity counts show that the pods were the most active sinks in both growth habits, followed by the stem. The branches for determinate plants had more specific activity. Overall, there was a general decrease of assimilate export to other organs in favour of the growing pods. Table 4: The % distribution of translocated 14C-photoassimilates and the activities in counts min-1g-1 dry
matter (in parenthesis) for the two determinate growth habits 645(a), 642(b) and the indeterminate type 644(C) at the flowering stage and early pod stage in summer 1988. Values are means of 3 replicates
Plant 645 642 644 Plant type a b C
Organ 1 - - - 2 2.2 (69.9) 2.2 (695.9) 6.1 (480.3) 3 - - 1.6 (1657.3) 4 - - - 5 72.1 (12494.8) 71.0 (21461.2) 53.9 (6549.6) 6 - - - 7 1.0 (495.5) 0.3 (605.8) 5.2 (2919.4) 8 3.1 (464.8) 7.3 (2280.5) 1.6 (245.9) 9 16.1 (1737.6) 10.4 (3298.8) 21.61 (2705.9)
10 2.1 (113.6) 5.8 (1913) 1.5 (99.3) 11 3.6 (301.9) 2.9 (236.2) 8.3 (537.3)
For data presented in Table 5 for plants at the mid-pod filling stage, it was observed that pods were the major sinks. More assimilates were exported to the pods in the determinate plants than the indeterminate ones. This reflects the fact that determinate plants set pods earlier. Overall, the stem received more assimilates in the indeterminate plants, while roots were stronger sinks in the determinate growth habit. These observations agree with those of Crompton et al. (1981), who noted that at pod fill much of carbon from the lower and mid-stem leaves moved into the pods in the indeterminate type. At young pod development, Baker et al.(1983) observed that 62.6, 64.4 and 62.6% of translocated 14C-photoassimilates were exported to the pods when plants were fed at the 1st, 2nd and 3rd flowering nodes, while in the indeterminate plants the pods received 64.9,66.6 and 50.7% respectively. The pods and the shoot apex had more specific activity than the other organs in the determinate and the indeterminate plants respectively. The difference is most probably due to the fact that the determinate plants set pods earlier than the indeterminate ones.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 165
Table 5: The % distribution of translocated 14C-photoassimilates and the activities in counts min-1g-1 dry matter (in parenthesis) for the two determinate growth habits 645(a), 642(b) and the indeterminate type 644(C) at the mid pod filling stage in summer 1988. Values are means of 3 replicates
Plant 645 642 644 Plant type a B C
Organ 1 - - - 2 3.1 (131.9) 1.2 (243.5) 8.43 (133.2) 3 - - - 4 - - - 5 - - - 6 63.3 (1454.1) 24.2 (2699.4) 24.2 (979.1) 7 1.8 (218.7) 1.4 (1244.4) 21.1 (759.6) 8 5.0 (276.5) 4.7 (840.7) 8.8 (349.6) 9 4.2 (187.4) 5.8 (627.6) 2.99 (165.7)
10 9.1 (107.4) 3.0 (42.3) 20.7 9608.2) 11 13.5 (190.5) 34.0 (838.9) 8.82 (74.9)
At the late pod filling stage (Table 6), it was observed that over 75% of the 14C-assimilates was exported to the stem in both growth types. As expected, the pods had the highest specific activity, followed by the stem and leaves. The stem, apart from receiving an appreciable percentage of 14C-assimilates, also gave relatively higher activity than other organs. The results presented here therefore, clearly indicate that the distribution pattern of 14C-photoassimilates changed with the stage of development, type of growth habit and even the time of feeding, depending on the prevailing environmental conditions (Austin et al., 1981 ). The general pattern of distribution of assimilates for both growth types was similar at the vegetative stage but as the shoot apex of the determinate plants became reproductive at flowering, the subsequent pattern of assimilate distribution was different from that of the indeterminate plant. At the reproductive stages the pods were the major sinks for assimilates. In all stages of development considered in this study, none of the growth habits was observed to be unequivocally more efficient in the uptake and distribution of photoassimilates. The activities in the fed leaf (Table 7) after 6 h were higher than in the other organs. Crompton et al. (1981) observed that the amount of radioactivity moving out of a source leaf at a fruiting node increased over successive samplings up to 48h; the pattern of distribution, however, remained unchanged. Table 6: The % distribution of translocated 14C-photoassimilates and the activities in counts min-1g-1 dry
matter (in parenthesis) for the two determinate growth habits 645(a), 642(b) and the indeterminate type 644(C) at the late pod filling stage in summer 1988. Values are means of 3 replicates
Plant 645 642 644 Plant type a B C
Organ 1 - - - 2 1.2 (648.7) 1.1 (2296.8) 2.2 (281.7) 3 - - - 4 - - - 5 - - - 6 79.6 (3383.5) 75.5 (6902.3) 80.7 (7065.2) 7 0.6 (1143.2) 0.5 (742.9) 1.5 (501.9) 8 3.6 (947.9) 2.3 (1377.7) 4.3 (1221.4) 9 10.9 (2930.1) 14.3 (4932.5) 5.3 (1553.9)
10 3.4 (385.0) 1.7 (77.2) 1.1 9 (1.2) 11 0.65 (120.3) 4.8 (402.9) 3.2 9 (49.3)

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 166
Table 7: Activities min-1g-1 dry matter of the fed leaf of plant growth habits at various stages of growth as listed in Tables 1-6
Activities of fed leaf per g (DPM) Table a(645) b(642) c(644) 1 1222518.025 2604259.107 1808803.143 2 372938.845 360732.786 883823.990 3 167418.498 235609.745 221995.538 4 364447.425 381194.760 365526.536 5 205816.323 249636.992 227063.538 6 149140.245 23693.306 324182.471
Discussion This study was instigated with the objective of appraising the relative advantages and disadvantages of two Vicia faba growth habits, with reference to photosynthesis and photoassimilate partitioning. Chapman and Peat (1978) have enumerated the phenotypic advantages, which accrue to a determinate habit compared with an indeterminate one. The results presented here provide some physiological quantification of the relative performance and effectiveness of the two forms to utilise growth resources under similar conditions. As Baker et al. (1983) have pointed out, the development of a neotenous determinate field bean offers the potential for a protein yielding break crop, which could readily fit into a modern, cereal-dominated agricultural system. However, there is still ground to be covered, particularly when we consider that this new types is still under improvement programmes. A feature requiring urgent attention is the tendency of the determinate type to develop infertile branches on the main stem, which results in reduced plant yields. This is aggravated by the fact that the main stem has only 4 to 6 podding nodes (Sjodin, 1971). Gates et al. (1983) have pointed out that although the determinate type offers promising alternative for achieving yield stability, the concomitant loss of phtosynthetic leaf area has resulted in lower potential yield than in the indeterminate one. However, the patterns of leaf area development observed in this study for both types were similar, although that of the determinate plants were initially more rapid. Austin et al. (1981) observed the same trend, and also noted that the pattern of leaf area and dry matter increase was similar for both ground types. Possibly, the lower plant yields observed for the determinate growth habit are mainly due to the lower number of pods per plant as compared with the indeterminate type. Is it possible then than the determinate type is sink limited? The fact that the plant has fewer podding nodes compared with the presence of infertile tillers leads us to believe that the photosynthetic capacity of the leaf area (source) may not be fully utilised. Moreover, it was observed that leaf area duration was longer in the determine type and, with the limited sink capacity, this may well have encouraged the late growth of tillers. In fact the tillers acted in the same way as the indeterminate growth habit but without sinks (pods) to provide economic benefit from the imported assimilates. As Chapman and Peat (1978) observed, however, there were no significant differences in the average yields of the two phenotyes, although the normal indeterminate plant did have the highest average performance. Moreover, neither growth habit showed any major differences in its relative effectiveness in the uptake and assimilation of CO2. It is therefore, suggested that the future of the neotenous growth type for enhancing yields lies in the incorporation of more pod-forming nodes on the main stem and on the currently infertile tillers. Otherwise, this factor will continue to dilute the relative advantages of this type as compared with the conventional determinate ones. The results from the assimilate partitioning studies presented here have elucidated to some extent how the two growth habits behave in their pattern of assimilate distribution at different developmental stages. Before and at flowering

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 167
the pattern of assimilate distribution was similar in both growth habits. Interestingly, flowers were observed to be only weak sinks for the current assimilates, judging by the amount of 14C-assimilates transported to them. Heavy flower shedding was observed for both growth habits. It has been reported in a number of studies that fertilisation is not a major cause of flower shedding (Kambal et al., 1979; Gates et at. , 1981 ). In their study, Gates et al (1981) pointed out that the pattern of flower abscission may be directly related to sequential acropetal anthesis within racemes and to the architecture of the vascular system within raceme peduncles. The development of basal pods of racemes was found to induce cellular changes in the abscission layers of the apical flowers, resulting in shedding, even if fertilized. They also suggested that the physiological interaction between pods and flowers with the same raceme may be circumvented by selecting for synchrony of anthesis within racemes, or for independent vascular traces to each flower, so that direct interaction between flowers and young pods cannot occur. As synchrony of anthesis is strongly influenced by environmental factors such as temperature, the presence of an independent vascular supply to each flower within a raceme is the most reliable means of improving pod set, in which case any loss of yields would only be attributable to pod and ovule abortion. Charlesworth (1989) in answering the question "why do plants produce so many more ovules than seeds?" has suggested that abortion of ovules and fruits may result due to lack of enough pollen for fertilisation, competition over limited resources where excess flowers and fruits are produced and more importantly, due to high genetic loads, causing many fertilisation products to abort randomly early in development. Alternatively, Bawa et at. (1989) have proposed that both embryo and fruit abortion which are distinctly different contribute to the lowered maturation of ovules, which is caused by parent-offspring conflict over resource allocation and sibling rivalry. The production of extra ovules may be a means of selecting embryos in the face of diversity of pollen genotypes reaching the stigmas resulting in non-random abortions. As detailed studies have yet to be performed on the anatomy of the new determinate types, especially on the anatomy of flowering, the area of independent vascular supply to individual pods remains plausible and there is justification to believe that the massive flower and pod set observed in both growth types may have a bearing on the timeliness and pattern of vascular development in the racemes. Concerted and detailed studies in this area in future programmes may help in solving this puzzle and enhance the effectiveness of producing yields. It is the close coordination between the knowledge of anatomical development of photosynthetic tissue, vascular development, especially at the reproductive stage, and the pattern and availability of assimilates, that will help enhance the potential for yield of existing and new phenotypes in this crop. The relative advantages of the determinate growth habit over the indeterminate one are thus to a great extent cancelled by the fact that these types produce fewer pods on the main stem, coupled with the asynchronous production of infertile tillers. With this background, it is suggested that breeders should consider selecting an intermediate growth habit, which will combine the existing advantages of the determinate plants, while exhibiting a superior yield status. Perhaps lines with independent vascular traces to the individual pods on racemes will enhance the possibility of increased yields. Certainly, this type of plant will have to exploit the existing photosynthetic capacity in such a way that extended or current leaf area will be utilised more effectively than at present. It is assumed that the proposed intermediate determinate phenotype will have more pod-bearing nodes on the main stem and fewer tillers, with an improved capacity to set pods.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 168
It will also fit within a framework of optimum field agronomic practices of density, nutrients and other environmental factors. However, this is not a simple task and, for the interim, the indeterminate conventional types will continue to out yield the neotenous determinate ones.
References Allison, J.C. and Watson, D.J. (1966). The production and distribution of dry matter in maize
after flowering. Annals of Botany 30. 365-382. Ashley, D.A.(1972). 14C-labelled photosynthate translocation and utilisation in cotton plants.
Crop Science 12: 69- 74. Austin, R.B., Morgan, C.L. and Ford, M.A.(1981). A field study of the carbon economy of
normal and 'topless' field beans, Vicia faba L. In: World Crops Vol. 4. Production, utilisation and description. Vicia faba: Physiology and Breeding. Ed.R. Thompson. Martinus/Junk Publishers, The Hague.pp.66 - 77.
Baker, D.A., Chapman, G.P., Standish, M. and Bailey, M. (1983). Assimilate partitioning in a determinate variety of field bean. In: Temperate Legumes: Physiology, Genetics and Nodulation. Ed. D.G. Jones and D.R. Davies. Pitman Publishers, pp. 191-199.
Bawa, K.S., Hedge, S.G., Ganeshaiah, K.N. and Umashankev, R. (1989). Embryo and seed abortion in plants. Nature 342: 625 - 626.
Belikov, I.F. (1973). The basic laws of the transport and distribution of assimilates in agricultural plants. Proc.Inst.Biol.Pedol,(Vladivostok) 20: 154- 160.
Benedict, J. and Kohel, R.J. (1975). Export of 14C-assimilates in cotton leaves. Crop Science 15: 367- 372.
Chapman, G.P. and Peat, W.P. (1978). Procurement of yield in field and broad beans. Outlook on Agricutlure 9. 267- 272.
Chapman, G.P., Fagg, C.W. and Peat, W.E. (1978). Parthenocarpy and internal competition in Viciafaba L. Zeitschrift fur Pflanzenphysilogie 94 : 247- 255.
Charlesworth, D.(1989). Why do plants produce so many ovules than seeds? Nature 338.21-22.
Crompton, H.I., Llyoid-Jones, C.P. and Hill-Cottingham, D.G. (1981). Translocation of labelled assimilates following phtosynthesis of 14CO2 by the field bean Vicia faba L. Physiologia Plantarum 51: 189-194.
Eastin, J.A. (1970). 14C labelled photosynthate export from fully expanded corn (Zea mays L.) leaf blades. Crop Science 10:415-419
Evans, L.T. (1975). The physiological basis of crop yield. In: Crop Physiology. Ed.L.T Evans, Cambridge University Press. pp. 327- 355.
Gates, P.J., Yarwood, J.N., Harris, N., Smith, M.L. and Boulter, M.E. (1981). Cellular changes in the pedicel and peduncle during flower abscission in ~Ticia faba L. In rTicia faba, Physiology and Breeding, World Crops: production, utilization and description, Vol.4.Ed.R. Thompson. Martinus Nijhoff, The Hague. Pp. 299- 3212.
Gates, P.J., Smith, M.L., White, G. and Boulter, D.(1983). Reproductive physiology and yield stability in vriciafaba L. In: Temperate Legumes, Physiology, Genetics and Nodulation. Ed. D.G. Jones and D.R. Davies. Pitman Publishers, pp. 43- 54
Hall, A.J. and Milthorpe, F.L. (1978). Assimilate source sink relationships in Capsicum annum L. Australian Journal of Plant Physiology 5: 1-13.
Hartt, C.E. and Kortschak, H.P. (1967). Translocation of 14C in the sugarcane plant during the day and night. Plant Physiology 42: 89-94.
Ho, L.C. Grange, R.I. and Shaw, A.F.(1989). Source-sink regulation. In. Transport of Photoassimilates. Ed. D.A. Baker and J.A. Milburn. Monographs and Surveys in the Biosciences. Longman Scientific and Technical, New York. pp. 306- 322.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 169
Hofstra, G. and Nelson, C.D. (1969). The translocation of photosynthetically assimilated 14C in corn. Canadian Journal of Botany 47: 1435- 144.
Hume, D.J. and Criswell, J.G. (1973). The translocation of photosynthetically assimilated 14C in soybeans. Crop Science 13.519 - 524.
Ishag, H.M. (1973). Physiology of seed yield in field beans, r,Ticia faba L. Dry matter production. Journal of Agricultural Science 80: 191-199.
Jacquiery, R. and Keller, E.R. (1980). Influence of distribution ofassimilates on pod set in field beans, Vicia faba L. Part II. Angew.Bot. 54: 29- 39.
Kambal, A.E. (1969). Components of yield in faba beans (Vicia faba L.) Journal of Agricultural Science 72. 359- 364.
Kambal, A.E., Bond, D.A. and Toybee, G. (1976). A study on the pollination mechanism in field beans, Vicia faba L. Journal of Agricultural Science 87: 519-526.
Peel, A.J. and Weatherley, P.E. (1962). Studies on the sieve tube exudation through aphid mouth parts: the effects of light and girdling. Annals of Botany 26: 633- 646.
Sjodin, J. (1971). Induced morphological variation in Vicia faba L. Hereditas 67.155 180 Thorne, J.H. and Koller, H.R. (1974). Influence of assimilate demand on photosynthesis,
difusive resistances, translocation and carbohydrate levels of soybean leaves. Plant Physiology 54: 201 - 207.
Thrower, S.L (1962). Translocation of labelled assimilates in soybean. Australian Journal of Biological Sciences 15: 629- 649.
THE EFFECT OF SILICON AND MANAGANESE ON COWPEA (VIGNA UNGUICULATA) LEAF WATER LOSS Owino-Gerroh, C. Department of Horticulture, Maseno University P.O. Box 333, Maseno, Kenya
Abstract The effect of silicon (Si) and manganese (Mn) on cowpea leaf water loss was studied in nutrient solution in a growth chamber at the Institute of Plant Nutrition, University of Hannover, Germany. The plants were grown for 12 weeks after transplanting the seedlings. Both leaf transpiration and resistance were determined using a Locor porometer. The addition of Si significantly increased leaf transpiration and decreased leaf resistance. The opposite was true when Mn was added to the nutrient solution. The addition of Si to solution containing Mn significantly increased leaf transpiration and decreased resistance. The addition of Mn to solution containing Si had no effect on both leaf transpiration and resistance. These findings were attributed to better plant growth and suppression of the negative effect of Mn on addition of Si to the nutrient solution. Mn normally suppresses plant growth due to its toxic effect on plant growth.
ROLE OF MYCORRHIZAL FUNGI IN THE SUSTAINABLE HORTICULTURAL CROP PRODUCTION IN THE TROPICS Wamocho, L. S. Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P .O. Box 62000 Nairobi, Kenya
Abstract Mycorrhizal fungi are found in many soils around the world and the fungi are known to form an association with many horticultural plant species. Vesicular-arbuscular mycorrhizal fungi (VAM) colonization of horticultural plant roots can improve plant growth via increased uptake of phosphorus (P), Zn and other minerals and such association has shown tolerance to diseases. Moreover, root colonization by mycorrhiza has many advantages, including (a) enhancing rates of photosynthesis and transpiration of host plants under a high temperature stress condition (b) increasing transplant uniformity (c) reduction of transplant mortality and (d) injury to transplants. VAM fungi colonize the host plant roots by penetrating

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 170
the cortex of the host roots without harming them. Inside the host root, the fungi form characteristically specialized organs such as for food storage (vesicles), nutrient exchange between the fungi and the host plant (arbuscules), resting stage (spores) and the vegetative stage of the fungus (hyphae). The fungi also produce a network of hyphae in the soil. The network is the basis for the symbiotic association between the fungi and the roots of the host plant. It can lead to an enhancement of plant growth via the alleviation of P deficiency in a P-deficient soil. As a consequence of increasing plant nutrient uptake, infection of roots of the host plants by VAM fungi has often been found to enhance plant growth. P, even more than nitrogen (N) is the most critical limiting nutrient for plant growth in the tropics. It seems that under natural conditions, often a low effectiveness of VAM fungi occurs on plants. Occurrence of V AM fungi in soils and their subsequent infection in roots of host plants is often associated with fertility levels in soils.
ISOLATION AND TESTING OF LOCAL COMMON BEAN RHIZOBIUM FOR IMPROVED N SUPPLY THROUGH NITROGEN FIXATION. Joseph R. Chemwetich2, A. O. Onkware2* and Samuel Gudu
Abstract Legume-Rhizobium symbiosis can supply a substantial proportion of the nitrogen required by the legume for optimum growth and production, and reduce demand for N fertiliser. But, response of the common bean to rhizobium inoculation under local conditions has been variable, probably because of competition from indigenous rhizobia with low N-fixing efficiency, low P levels, or soil acidity and mineral toxicity. Use of locally adapted, effective rhizobia could overcome this problem. The objective of this study was to isolate rhizobia from different agroecological zones of Kenya and test them for nodulation and effectiveness, with the goal of getting isolates that would be most suitable for local soil conditions. Soil samples were collected from 12 different localities across the country. Rhizobium population in the soil samples was determined in Leonard jar assembly using a common bean cultivar GLP 24 (Canadian Wonder) as the trap plant. Resultant root nodules were aseptically extracted and plated for Rhizobium using yeast extract mannitol agar with BTB or Congo red for preliminary characterization2. Selected colonies were sub-cultured in broth and used to inoculate beans in Leonard jar assembly. Nine isolates that showed a high level of nodulation were selected and tested in a pilot field experiment using GLP 24 at the Chepkoilel Campus farm in Eldoret. Treatment with a commercial rhizobium strain (as a check) and recommended N fertilizer application as (standard) was also included. Data was recorded on the number of nodules, nodule dry weight, and shoot dry weight per plant at maximum flowering. The number of pods, seeds per pod and weight of seeds per plant were assessed and recorded at maturity. The data was subjected to statistical analysis and differences were taken as significant at p<a 0.05. Neutral or slightly acidic soils (pH 5.5 – 7) had significantly more rhizobia (>104 cell/g) than more acidic (pH <4.5; 1.6x102 cells/g) soils. All isolates were acid forming, fast growers that did not absorb Congo red in the dark. They successfully nodulated GLP 24 in Leonard jar assembly, in contrast to the control where there was no nodulation. Isolates E and Ki, and Ka produced significantly more nodules in Leonard jar assembly and field experiments respectively. Isolate Ro and E produced heavier nodules per plant than the other isolates or MIRCEN. None of the isolates significantly increased shoot height or dry weight in the Leonard jar assembly. In the field, only Ka had relatively more nodules per plant than either Mir or standard N application. In the field, plants inoculated with Ka and R had relatively higher shoot biomass than full N or Mir treatments. Isolate Kk induced significantly more branching than full N or Mir treatment at 43 DAS. The number of pods per plant was relatively higher in plants inoculated with Kk Ka and Crh than other treatments. None of the isloates had significant effect on number of seeds per pod, although Crh and R induced relatively more seeds per pod than Mir or standard N supply. Inoculation with Crh, Kk and Ku significantly increased seed weight per plant. It was concluded that some of the local rhizobium isolates improved common bean (GLP 24) growth and yield. Large-scale field experiments have been set up to test the responses further.
Introduction Nitrogen fixation by legume-Rhizobium association is a renewable, less expensive and more environment friendly source of plant nitrogen than mineral N-fertlisers. Rhizobium-common bean (Phaseolus vulgaris L) symbiosis can supply a substantial part of the nitrogen required by

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 171
the plant for optimum growth and production (FAO 1984, Giller et al 1998), thus reducing the need for mineral N fertiliser. Research reports show that inoculation with appropriate Rhizobium strains increases growth and production of common beans (FAO 1984, Amijee and Giller 1998). However, inoculation experiments with commercial Rhizobium inoculants have produced variable results under local, Kenyan conditions. Thus, Karanja et al (2000) reported that inoculation with elite Rhizobium strains improved general growth but not the grain. The Rhizobium that is introduced by inoculation must overcome several local soil factors in order to nodulate and form effective symbiosis with the common bean plants. Such factors include competition from native rhizobia (Anyango et al 1998), P deficiency (Lupwayi and Mkandawire 1996, Amijee and Giller 1998) or low soil pH and mineral toxicity (Franco and Munns 1982) in some cases. Soil acidity is often associated with both N and P deficiency, and mineral, especially aluminium and manganese, toxicity (Foy 1992). Consequently, Rhizobium isolates that may have shown superior nodulating and N-fixing ability under specific soil conditions may not be similarly effective under local conditions. Soils in many of the moist parts of Kenya are acidic, deficient in N and have high exchangeable aluminium. Rhizobium strains that can be used in such soils must be acid and aluminium tolerant. Attempts to improve bean production by inoculating with exotic acid resistant strains, such as CIAT 899, has produced inconclusive results (Karanja et al 2000). This problem could be overcome by use of effective local Rhizobium isolates that are both acid and aluminium tolerant. The objective of the current study was to isolate Rhizobium from different agroecological zones across Kenya and assess them for infectiveness and effectiveness, with the goal of finding strains, which would be most effective with local common bean cultivars and soil conditions.
Materials and Methods Composite soil samples were collected from twelve different locations in Kenya: Kavutiru (Embu), Kahuro (Muranga), Kuinet and Chepkoilel (Uasin Gishu), Ogembo and Nyatieko (Kisii), Kasaka and Mondoi (Baringo), Boito (Koibatek), Shariani (Kilifi), Mwangani (Malindi), and Mackinon Road (Taita Taveta). All samples were collected from farmers’ fields where no deliberate inoculation with common bean Rhizobium had been done before. A MIRCEN® commercial common bean inoculant was used as a check. The pH of soil samples in triplicate was determined both in distilled water and 0.01 M CaCl2 using 1: 2.5 (W/V) suspension and combined glass electrode. Rhizobium population was assessed using the plant nodulation technique (most probable number, MPN) in Leonard jar assembly inoculated with serial dilutions of soil suspension (4-1 to 4-9; Somasegaran et al 1982). Pregerminated seeds of Canadian Wonder (GLP 24), a local common bean cultivar, were planted in jars that were irrigated with N-free nutrient solution (Lupwayi and Haque 1994). 2 ml of the serially diluted soil suspension were separately added to different jars at 24 hours after planting. Four replicates were maintained for each dilution step, and the MIRCEN strain and full N were also included as check and standard respectively. Nodulation was assessed and recorded after 28 days, and the data on presence or absence at the various dilution levels was used to calculate the MPN (Somasegaran et al 1982). Nodules were surface-sterilized, extracted in sterile distilled water and plated on YEMA plates with either BTB or Congo red dyes for preliminary characterization. Single Rhizobium colonies on the YEMA plates were sub-cultured in YEM broth (Lupwayi and Haque 1994) at 29ºC for 48 hours. 1 ml of the broth culture, containing approximately 109 cell/ml, was used to inoculate GLP 24 seedlings in triplicate Leonard jar assembly that were
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 172
irrigated with N-free nutrient solution. A complete nutrient treatment was included as a standard. Nine of the isolates (Crh, E, Ka, Ku, Ki, R, Ro, KK, Mo - see foot note and Table 1 for isolate coding)3 that exhibited significantly higher levels of nodulation, or represented different AEZs, were re-cultured in YEM broth and used as inoculant in a pilot field experiment with GLP 24. Field plants were assessed for number nodules, shoot branches and pods per plant, number of seeds per pod and seed weight per plant at maturity. The data was analysed using a computer statistical program (SPSS Version 7.5.1, SPSS Inc.), and effects were taken as significant at p<0.05. Means were separated using Tukey’s HSD test, where applicable.
Results and Discussion Most soils from inland Kenya were acidic, pH < 6.0 (Table 1). The acidic inland soils, especially those with pH <5.5, are likely to adversely affect both growth and nitrogen fixing ability of most Rhizobium species. This is because acidity reduces Rhizobium activity (FAO 1984, Foy 1992, Graham and Swenson, Undated).
Table 1. Soil pH, Rhizobium denisty (cells/g) and nodulation properties for soils collected from different parts of Kenya
Site Code Soil pH Rhizobium density Nodule no. Nodule wt (mg)
Boito Bo 6.02 3.2*104 8bc 5.5bc Chepkoilel Crh 5.34 8.0*103 7bc 2.0c Kahuro Ka 5.55 1.8*104 1d 1.0c Kasaka Kk 5.55 2.3*104 1d 1.0c Kavutiri E 4.33 1.6*102 48a 10.5b Kuinet Ku 5.37 1.8*104 6bc 1.5c Mwangani M 7.15 6.5*104 2d 1.0c Mackinon Rd Ro 7.16 2.3*105 1d 1.0c Mondoi Mo 4.69 2.3*104 4c 1.2c Nyatieko Ro 4.79 8.5*104 28b 7.3bc Ogembo Ki 5.7 3.2*104 43ab 22.58a Shariani S 6.44 9.0*104 1d 1.0c Mircen Mir - 1*108 11bc 4.5bc
All the soils had a viable population of “bean’ rhizobium, but the population was significantly higher in the neutral or slightly acidic (pH > 6.0) soils (Table1). The rhizobium population in most of the soils would be considered adequate for bean nodulation (Amijee and Giller 1998), but effective symbiosis is also affected by other factors, in addition to rhizobium population (FAO 1984). All isolates were acid forming, fast growers, which did not absorb Congo red in the dark and successfully nodulated GLP 24 in Leonard jar assembly. This showed that they were bean Rhizobium species (Lupwayi and Haque 1994, Somasegaran et al 1982), although the species identity was not established. Only E and Ki produced significantly more nodules than the MIRCEN strain in the Leonard jar assembly (Table 1). In the field only isolate Ka had relatively more nodules per plant than the control or the Mir strain (Figure 1). The nodule weight per plant was closely correlated with
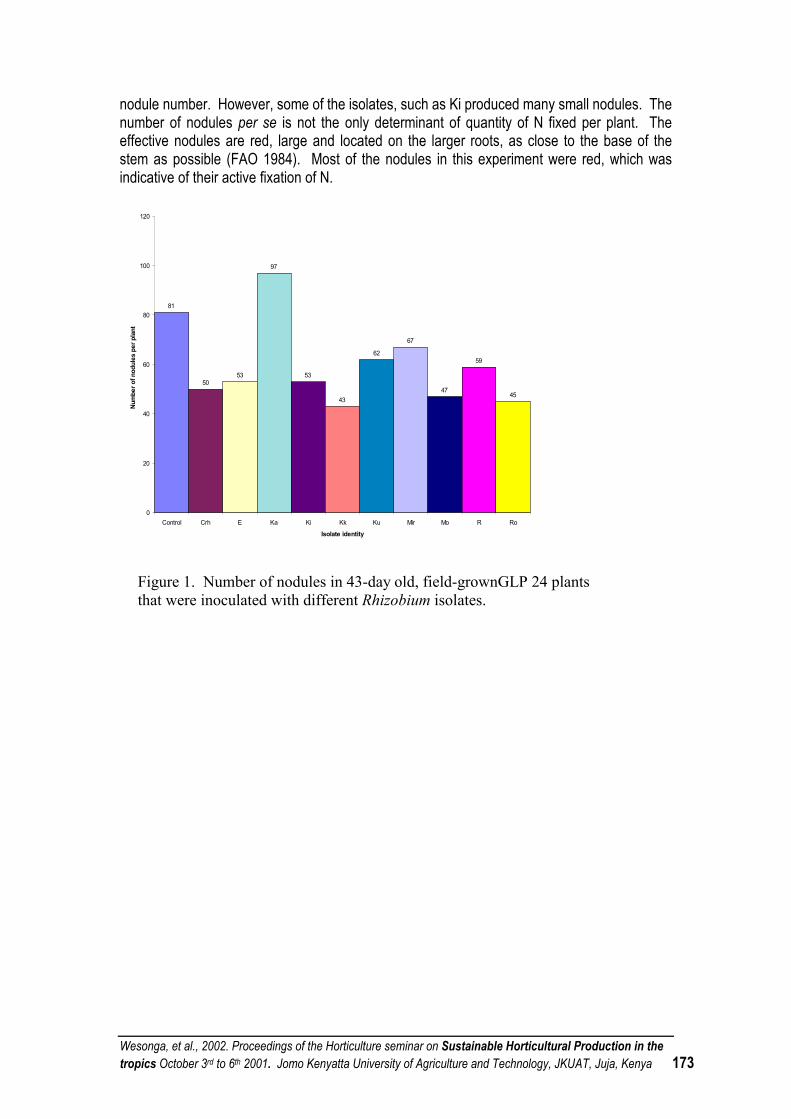
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 173
nodule number. However, some of the isolates, such as Ki produced many small nodules. The number of nodules per se is not the only determinant of quantity of N fixed per plant. The effective nodules are red, large and located on the larger roots, as close to the base of the stem as possible (FAO 1984). Most of the nodules in this experiment were red, which was indicative of their active fixation of N.
81
5053
97
53
43
62
67
47
59
45
0
20
40
60
80
100
120
Control Crh E Ka Ki Kk Ku Mir Mo R Ro
Isolate identity
Num
ber o
f nod
ules
per
pla
nt
Figure 1. Number of nodules in 43-day old, field-grownGLP 24 plants that were inoculated with different Rhizobium isolates.
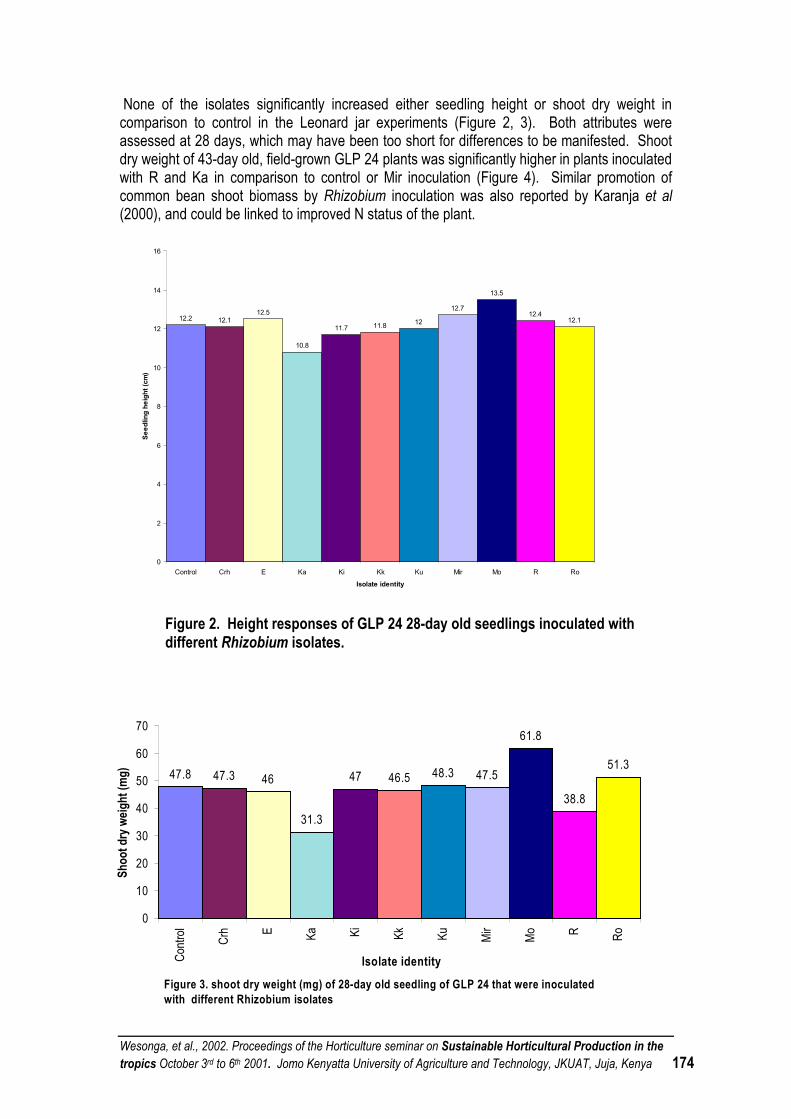
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 174
None of the isolates significantly increased either seedling height or shoot dry weight in comparison to control in the Leonard jar experiments (Figure 2, 3). Both attributes were assessed at 28 days, which may have been too short for differences to be manifested. Shoot dry weight of 43-day old, field-grown GLP 24 plants was significantly higher in plants inoculated with R and Ka in comparison to control or Mir inoculation (Figure 4). Similar promotion of common bean shoot biomass by Rhizobium inoculation was also reported by Karanja et al (2000), and could be linked to improved N status of the plant.
Figure 3. shoot dry weight (mg) of 28-day old seedling of GLP 24 that were inoculated with different Rhizobium isolates
47.8 47.3 46
31.3
47 46.5 48.3 47.5
61.8
38.8
51.3
0
10
20
30
40
50
60
70
Contr
ol Crh E Ka Ki Kk Ku Mir
Mo R Ro
Isolate identity
Shoo
t dry
weig
ht (m
g)
12.2 12.112.5
10.8
11.7 11.8 12
12.7
13.5
12.412.1
0
2
4
6
8
10
12
14
16
Control Crh E Ka Ki Kk Ku Mir Mo R Ro
Isolate identity
Seed
ling
heig
ht (c
m)
Figure 2. Height responses of GLP 24 28-day old seedlings inoculated with different Rhizobium isolates.
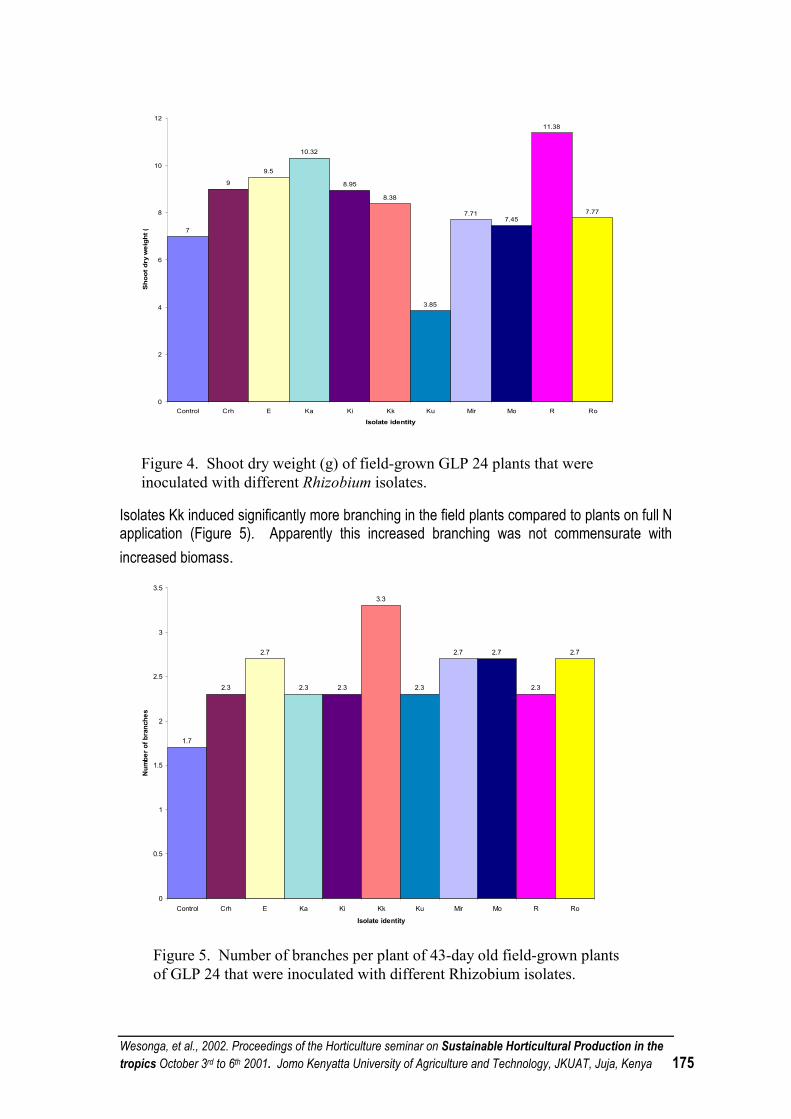
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 175
Isolates Kk induced significantly more branching in the field plants compared to plants on full N application (Figure 5). Apparently this increased branching was not commensurate with increased biomass.
1.7
2.3
2.7
2.3 2.3
3.3
2.3
2.7 2.7
2.3
2.7
0
0.5
1
1.5
2
2.5
3
3.5
Control Crh E Ka Ki Kk Ku Mir Mo R Ro
Isolate identity
Num
ber o
f bra
nche
s
Figure 5. Number of branches per plant of 43-day old field-grown plants of GLP 24 that were inoculated with different Rhizobium isolates.
7
9
9.5
10.32
8.95
8.38
3.85
7.717.45
11.38
7.77
0
2
4
6
8
10
12
Control Crh E Ka Ki Kk Ku Mir Mo R Ro
Isolate identity
Sho
ot d
ry w
eigh
t (
Figure 4. Shoot dry weight (g) of field-grown GLP 24 plants that were inoculated with different Rhizobium isolates.
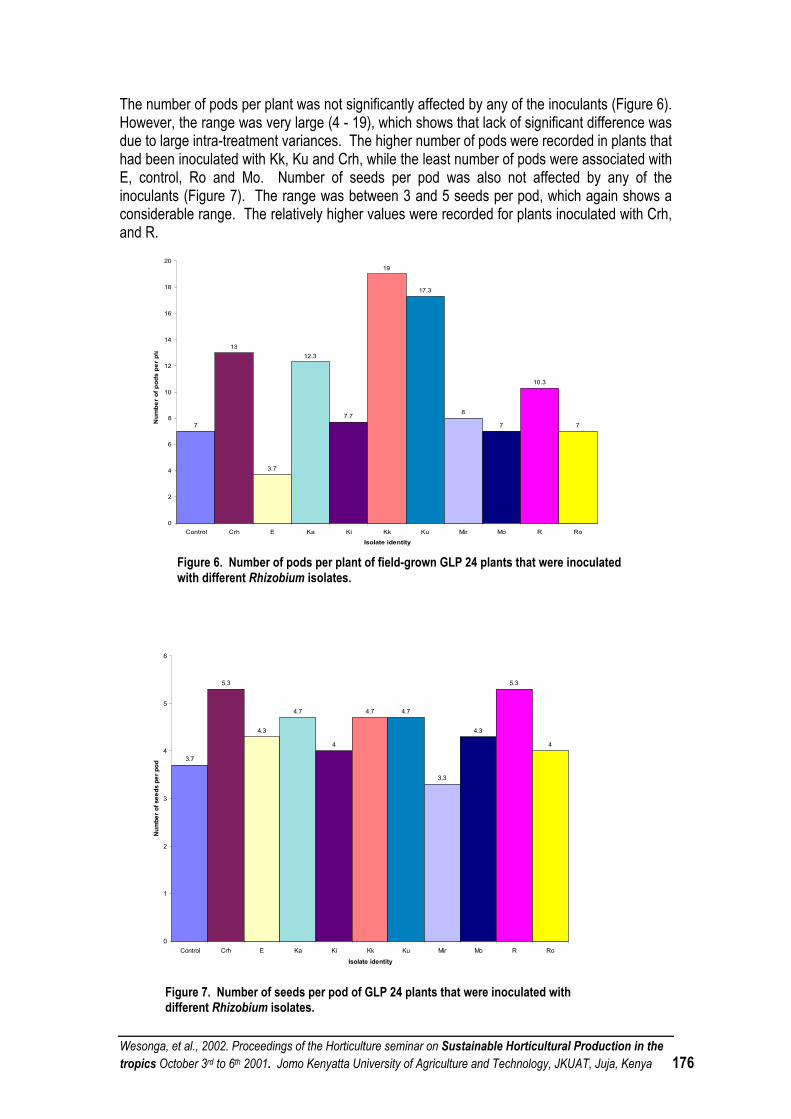
Wesongtropics
The number of pods per plant was not significantly affected by any of the inoculants (Figure 6). However, the range was very large (4 - 19), which shows that lack of significant difference was due to large intra-treatment variances. The higher number of pods were recorded in plants that had been inoculated with Kk, Ku and Crh, while the least number of pods were associated with E, control, Ro and Mo. Number of seeds per pod was also not affected by any of the inoculants (Figure 7). The range was between 3 and 5 seeds per pod, which again shows a considerable range. The relatively higher values were recorded for plants inoculated with Crh, and R.
3.7
5.3
4.3
4.7
4
4.7 4.7
3.3
4.3
5.3
4
0
1
2
3
4
5
6
Control Crh E Ka Ki Kk Ku Mir Mo R Ro
Isolate identity
Num
ber o
f see
ds p
er p
od
Figure 6. Number of pods per plant of field-grown GLP 24 plants that were inoculated with different Rhizobium isolates.
7
13
3.7
12.3
7.7
19
17.3
8
7
10.3
7
0
2
4
6
8
10
12
14
16
18
20
Control Crh E Ka Ki Kk Ku Mir Mo R Ro
Isolate identity
Num
ber
of p
ods
per
pla
Figure 7. Number of seeds per pod of GLP 24 plants that were inoculated with different Rhizobium isolates.
a, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 176
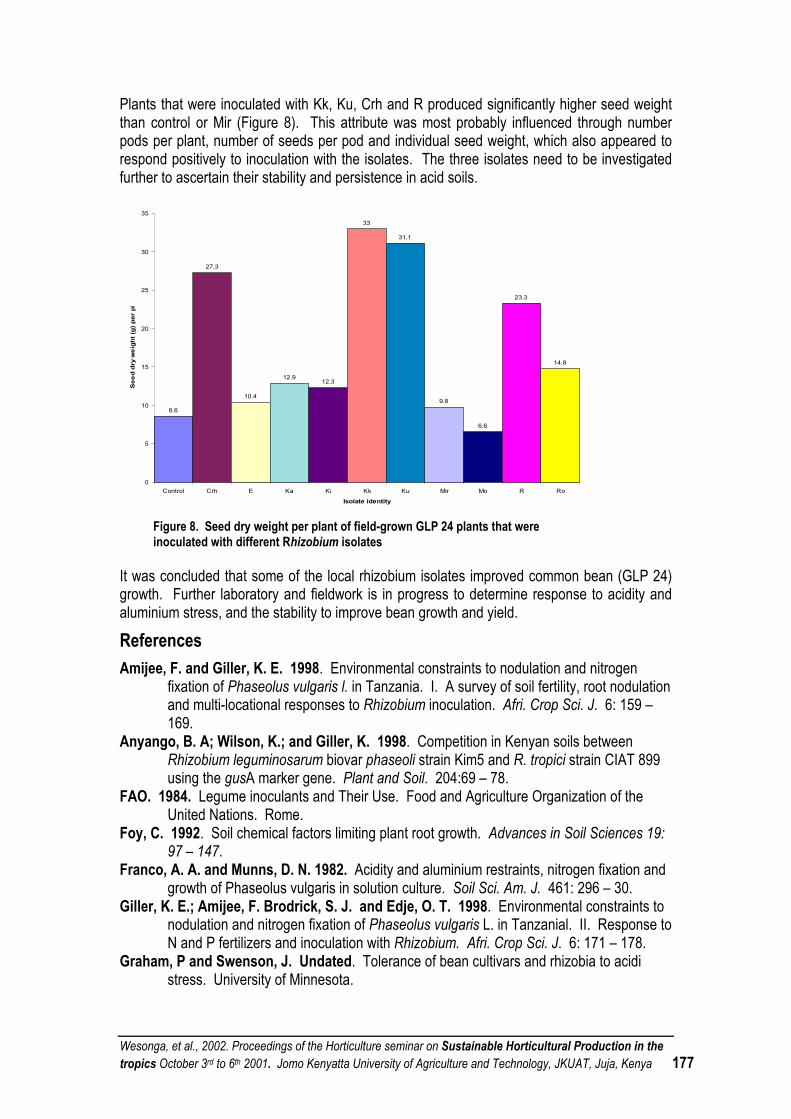
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 177
Plants that were inoculated with Kk, Ku, Crh and R produced significantly higher seed weight than control or Mir (Figure 8). This attribute was most probably influenced through number pods per plant, number of seeds per pod and individual seed weight, which also appeared to respond positively to inoculation with the isolates. The three isolates need to be investigated further to ascertain their stability and persistence in acid soils.
It was concluded that some of the local rhizobium isolates improved common bean (GLP 24) growth. Further laboratory and fieldwork is in progress to determine response to acidity and aluminium stress, and the stability to improve bean growth and yield.
References Amijee, F. and Giller, K. E. 1998. Environmental constraints to nodulation and nitrogen
fixation of Phaseolus vulgaris l. in Tanzania. I. A survey of soil fertility, root nodulation and multi-locational responses to Rhizobium inoculation. Afri. Crop Sci. J. 6: 159 – 169.
Anyango, B. A; Wilson, K.; and Giller, K. 1998. Competition in Kenyan soils between Rhizobium leguminosarum biovar phaseoli strain Kim5 and R. tropici strain CIAT 899 using the gusA marker gene. Plant and Soil. 204:69 – 78.
FAO. 1984. Legume inoculants and Their Use. Food and Agriculture Organization of the United Nations. Rome.
Foy, C. 1992. Soil chemical factors limiting plant root growth. Advances in Soil Sciences 19: 97 – 147.
Franco, A. A. and Munns, D. N. 1982. Acidity and aluminium restraints, nitrogen fixation and growth of Phaseolus vulgaris in solution culture. Soil Sci. Am. J. 461: 296 – 30.
Giller, K. E.; Amijee, F. Brodrick, S. J. and Edje, O. T. 1998. Environmental constraints to nodulation and nitrogen fixation of Phaseolus vulgaris L. in Tanzanial. II. Response to N and P fertilizers and inoculation with Rhizobium. Afri. Crop Sci. J. 6: 171 – 178.
Graham, P and Swenson, J. Undated. Tolerance of bean cultivars and rhizobia to acidi stress. University of Minnesota.
8.6
27.3
10.4
12.912.3
33
31.1
9.8
6.6
23.3
14.8
0
5
10
15
20
25
30
35
Control Crh E Ka Ki Kk Ku Mir Mo R Ro
Isolate identity
See
d dr
y w
eigh
t (g)
per
pl
Figure 8. Seed dry weight per plant of field-grown GLP 24 plants that were inoculated with different Rhizobium isolates

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 178
Karanja, N. K., Okalebo, J. R. and Obura, P. A. 2000. Assessing response of rhizobia inoculants bean (Phaseols vulgaris L) in low fertility soils: Preliminary findings. In BNF Research and Applications in Africa: Into the 21st Century. Book of Abstracts of the Ninth Congress of the African Association for Biological Nitrogen Fixation. Karanja, N., Kahindi, J. H. P, Anyango, B, Gitonga, n, M, Smithson, P. C, Woomer, P, L. and Odee, D. Nairobi, Kenya. Pp 34 - 36.
Lupwayi, N. Z and Haque, I. 1994. Working Document No. 29: Legume-Rhizobium Technology Manual. Environmental Sciences Division, ILCA, Addis Ababa. Pp 114.
Lupwayi, N. Z. and Mkandawire, A. B. C. 1996. Phosphorus, sulphur and molybdenum effects on response of Phaseolus beans to inoculation with Rhizobia in Malawi. Afri. Crop Sci. J. 4: 359 – 364.
Mugai, E. N. and Agong, S. G. 1997. The response of ‘Rosecoco” beans to aluminium treatment. Afri. Crop Sci. J. 5; 199 – 184.
Somasegaran, P.; Hoben, H and Halliday, J. 1982. The NifTAl Manual for Methods in Legume-Rhizobium Technology. University of Hawaii College of Tropical Agriculture and Human Resources.
VARIETY EVALUATION AND IMPROVED AGRONOMIC PRACTICES FOR GINGER (ZINGIBER OFFICINALE ROSC.) IN ETHIOPIA Nebiyu, A.1 and Etissa, E.2 1Ethiopian Agricultural Research Organization, Jimma Research Center, P.O. Box 192, Jimma, Ethiopia 2Ethiopian Agricultural Research Organization, Nazareth Research Center, P.O. Box 431, Nazareth Ethiopia
Abstract Ginger is known as a cultivated plant and its origin is probably from India or China. In Ethiopia, it has been known since ancient times and cultivated in wider environments especially in the hot and humid regions of the country. (Keffa, Illubabor, Gamogofa, Sidamo, Wellega, Gojam, Gonder) probably due to the ideality and the suitability of the region for ginger cultivation. In these parts of the country, ginger is cultivated as the major cash crop for the farmers. This spice crop is popular in the daily dish of every Ethiopian. It is used as sole or together with other spices to flavor a variety of foods and local drinks, for culinary purposes, for local medicines etc. It is well-known in Ethiopia and fresh or dry rhizomes are for sale on most markets. In addition, its derivative, the oleoresin, is being exported for additional foreign currency. During the past few years, research on variety improvement and agronomic practices and quality assessments have been successfully undertaken. Consequently, three ginger accession have been selected for their superiority in rhizome yield and essential oil content. In addition, the major agronomic and cultural practices like time of planting, land preparation, spacing, fertilizer requirement have been worked out. Hence, the improved varieties along with their agronomic packages have been recommended to users for sustainable ginger production in Ethiopia. In this paper, research results of variety improvement and agronomic aspects are discussed.
CHICKPEA ADAPTATION TO KENYA’S NORTHWESTERN HIGHLANDS Rheenen, H.A. van and Odindo, A.O. Department of Crop Production and Seed Technology, Moi University, P.O. Box 1125, Eldoret
Abstract Chickpea (Cicer arietinum) was introduced into Kenya probably during the late 19th century. It is mainly grown in relatively warm areas on vertisols and not in the cooler highlands. Preliminary adaptation studies suggest that the climatic conditions in the northwestern highlands are favourable for chickpea growth albeit that weather conditions during harvest may pose problems. A low pH of the soil may affect yields adversely. Large-, white-seeded kabuli types are probably better candidates for production in the
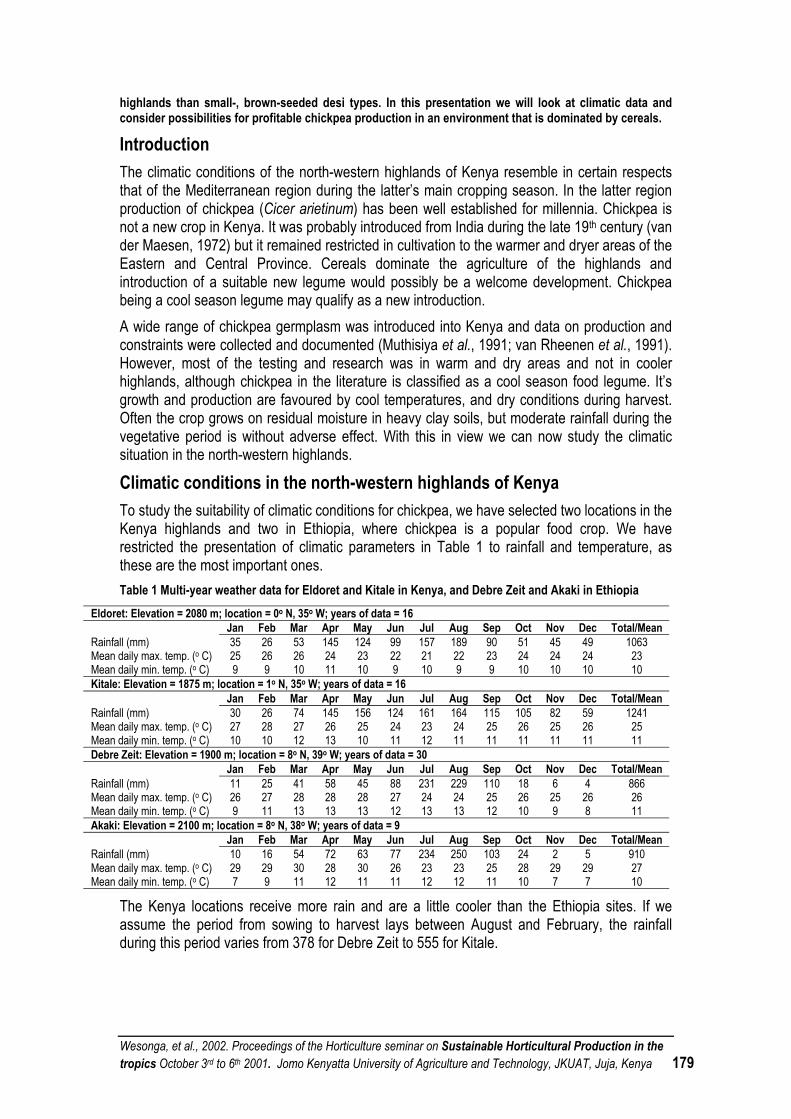
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 179
highlands than small-, brown-seeded desi types. In this presentation we will look at climatic data and consider possibilities for profitable chickpea production in an environment that is dominated by cereals.
Introduction The climatic conditions of the north-western highlands of Kenya resemble in certain respects that of the Mediterranean region during the latter’s main cropping season. In the latter region production of chickpea (Cicer arietinum) has been well established for millennia. Chickpea is not a new crop in Kenya. It was probably introduced from India during the late 19th century (van der Maesen, 1972) but it remained restricted in cultivation to the warmer and dryer areas of the Eastern and Central Province. Cereals dominate the agriculture of the highlands and introduction of a suitable new legume would possibly be a welcome development. Chickpea being a cool season legume may qualify as a new introduction. A wide range of chickpea germplasm was introduced into Kenya and data on production and constraints were collected and documented (Muthisiya et al., 1991; van Rheenen et al., 1991). However, most of the testing and research was in warm and dry areas and not in cooler highlands, although chickpea in the literature is classified as a cool season food legume. It’s growth and production are favoured by cool temperatures, and dry conditions during harvest. Often the crop grows on residual moisture in heavy clay soils, but moderate rainfall during the vegetative period is without adverse effect. With this in view we can now study the climatic situation in the north-western highlands.
Climatic conditions in the north-western highlands of Kenya To study the suitability of climatic conditions for chickpea, we have selected two locations in the Kenya highlands and two in Ethiopia, where chickpea is a popular food crop. We have restricted the presentation of climatic parameters in Table 1 to rainfall and temperature, as these are the most important ones. Table 1 Multi-year weather data for Eldoret and Kitale in Kenya, and Debre Zeit and Akaki in Ethiopia
Eldoret: Elevation = 2080 m; location = 0o N, 35o W; years of data = 16 Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Total/Mean Rainfall (mm) 35 26 53 145 124 99 157 189 90 51 45 49 1063 Mean daily max. temp. (o C) 25 26 26 24 23 22 21 22 23 24 24 24 23 Mean daily min. temp. (o C) 9 9 10 11 10 9 10 9 9 10 10 10 10 Kitale: Elevation = 1875 m; location = 1o N, 35o W; years of data = 16 Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Total/Mean Rainfall (mm) 30 26 74 145 156 124 161 164 115 105 82 59 1241 Mean daily max. temp. (o C) 27 28 27 26 25 24 23 24 25 26 25 26 25 Mean daily min. temp. (o C) 10 10 12 13 10 11 12 11 11 11 11 11 11 Debre Zeit: Elevation = 1900 m; location = 8o N, 39o W; years of data = 30 Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Total/Mean Rainfall (mm) 11 25 41 58 45 88 231 229 110 18 6 4 866 Mean daily max. temp. (o C) 26 27 28 28 28 27 24 24 25 26 25 26 26 Mean daily min. temp. (o C) 9 11 13 13 13 12 13 13 12 10 9 8 11 Akaki: Elevation = 2100 m; location = 8o N, 38o W; years of data = 9 Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Total/Mean Rainfall (mm) 10 16 54 72 63 77 234 250 103 24 2 5 910 Mean daily max. temp. (o C) 29 29 30 28 30 26 23 23 25 28 29 29 27 Mean daily min. temp. (o C) 7 9 11 12 11 11 12 12 11 10 7 7 10
The Kenya locations receive more rain and are a little cooler than the Ethiopia sites. If we assume the period from sowing to harvest lays between August and February, the rainfall during this period varies from 378 for Debre Zeit to 555 for Kitale.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 180
Discussion and conclusions It seems that the temperatures at the Kenya locations are good for chickpea crop growth. However, prerequisite for successful production is a reliable dry period at harvest. If we assume a growth period of 5 months, sowing in August – September will result in harvesting during January – February at Eldoret and Kitale. At Debre Zeit and Akaki, both good locations for chickpea production, sowing is usually during September and harvesting takes place in January. Yields of over two tons per ha have been obtained. We have observed excellent podding of chickpea at Eldoret, when the crop was sown during June. It seems that satisfactory chickpea production is feasible and that selection of adapted genotypes and the establishment of appropriate management practices will be key issues in introducing the crop in the north-western highlands of Kenya.
References Muthisiya, J., Omanga, P. and Rheenen, H.A. van 1990. A rapid survey of chickpea cultivation:
II. Machakos District, Kenya, 1989/90. International Chickpea Newsletter 23: 29-30. Van der Maesen, L.J.G. 1972. Cicer L., a monograph of the genus, with special reference to
the chickpea (Cicer arietinum L.), its ecology and cultivation. Mededelingen Landbouwhogeschool Wageningen, Nederland: 72-10. H. Veenman & Zonen N.V., Wageningen, The Netherlands.
Van Rheenen, H. A. , Abebe Tullu, Abdalla Ibrahim Sheikh M. and Omanga P. 1991. Growing chickpea in Eastern Africa. Patancheru, A.P. 502 324, India:International Crops Research Institute for the Semi-Arid Tropics.(Four languages; Kiswahili: Ukuzaji wa saina katika Afrika Mashariki).
MARKETING FOR SUSTAINABLE HORTICULTURAL PRODUCTlON IN KENYA Ong'eng'a, W.O.*,1; Odhiambo, M.O.1 and Okeno, J.2
1 Department of Agricultural Marketing and Co-operatives, Faculty of Agriculture, Moi University 2 Department of Horticulture, Faculty of Agriculture, Moi University * Address of Corresponding Author; Moi University, Box 1125 Eldoret. Tel: 0321-63160 E-mail: [email protected]
Abstract Sustainable horticultural production in Kenya, like any other economic activity, relies mainly on the ecological, political and economic conditions. Of these, economic conditions, particularly the marketing aspect is the most important to the highly perishable horticultural products. This paper analyses the domestic and export marketing channel constraints to sustainable horticultural production in the country. The major market challenges to sustainable horticultural production include: high physical and transaction costs, international trade stipulations, few and inefficient processing firms, restricted export product choices, inadequate market information, volatile domestic markets, and market policies. The low purchasing power occasioned by the declining economic growth not only reduces domestic demand for horticultural products but also hampers investment in the sector among market participants. All these influence the incentive structure to the farmers and other market participants. The consequence is forced home consumption of the edible produce by smallholder farmers, reduction in produce quality, heavy losses due to wastage and eventual loss of market share in the export market. Despite all these challenges, potentials exist for sustainability in the horticultural industry through enhancing efficiency in both the domestic and export markets. It is recommended that the achievement of efficiency in the marketing system for the country’s horticultural produce will

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 181
depend mainly on the reduction of both the transaction and physical production costs through improvements in the transport and communication networks and market integration for large-scale farms. Both government and private sector funding of research on the variety-specific marketing systems and quantification of the marketing margins received by all market participants is necessary for sustained growth and development of the subsector.
THE EFFECT OF MILD POSTHARVEST CHILLING ON THE EATING QUALITY OF BANANAS M. N. Muchui1* and A. Klieber 2
1 NHRC, Kenya Agricultural Research Institute. P. O. Kenya Agricultural. Box 220 Thika, Kenya Telephone: (0151) 21284 Email: karithi @kari.org, 2 University of Adelaide - Department of Horticulture , Viticulture Oenology-Australia
Abstract Bananas are highly chilling sensitive and suffer chilling injury when exposed to temperatures below 14oC that would occur accidentally during transport and storage. Mildlly chilled bananas attain a dull skin colour that is not attractive to the consumer. The main aim of this study was to find out through both sensory and chemical evaluation whether the eating quality of Cavendish bananas cv. Williams grown in Australia is affected by mild postharvest chilling. A secondary but important aim was to establish whether there is a correlation between sensory evaluation and chemical nose evaluation of banana aroma quality. Postharvest chilling was investigated by chilling summer grown bananas at 5oC for 24 hours and ripening at 16oC, 95% RH, and 300 ppm ethylene until ripeness colour stage 4. They were then transferred to 22 oC and ripened until ripeness colour stage 6. The fruits were then evaluated for aroma intensity, sweetness, soluble solids, pulp firmness and peel colour. This study showed that there was no perceptible difference in sweetness, aroma intensity and soluble solids between mildly chilled and control fruit. Fruit pulp pressure was less than 1 kg for both treatments. However, mildly chilled fruits had a dull yellow colour compared to control that had a bright yellow peel. Sensory evaluation was well corroborated by data from the chemical nose and this revealed that mild postharvest chilling did not have any effect specifically on aroma quality or the eating quality of bananas. The chemical nose is a viable tool for use in banana aroma quality studies as it performed similarly to the sensory panel.
Introduction Major banana varieties grown in Australia are Cavendish, Goldfinger and Ladyfinger. Cavendish is the main variety produced with Williams accounting for 95% of production. Most bananas are consumed locally at an average of 13kg/person/annum. Dessert bananas need to have certain quality characteristics in order to be acceptable to the consumers. These include, bright yellow peel, no bruises, a long shelf life, should also be sweet and have the desired flavour and aroma. Banana fruits are highly chilling-sensitive (Wills et al., 1998) and suffer chilling injury (CI) at chilling temperature below 14°C (Marriot, 1980). However, CI depends on temperature and duration of exposure (Hewage et al., 1996). CI lowers the visual quality of the fruits by giving them a sub-epidermal browning and blackening even on ripening (Olorunda et al., 1978). This Lowers banana marketability in Australia (Chatres, B. 2000, pers. comm.). Severe CI also lowers eating quality (Olorunda et al., 1978; Wang, 1990). Mild chilling temperatures may also affect the aroma quality of bananas (Mattei and Paillard, 1973).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 182
Objectives A study was carried out to determine the effect of accidental mild postharvest chilling during transportation and/or storage on eating quality of bananas. The study was also to find out whether there is a correlation between sensory and chemical sensor “chemical nose” evaluation of banana aroma quality.
Materials and Methods Cavendish bananas of cv. Williams grown in Australia during summer were used. They were harvested mature green and the third and fourth hands were used. The fruits were subjected to fungicidal treatments (Sportak, 0.55ml/L in water for 1 min.) and then subjected to two treatments. Mild chilling at 5°C for 24 hours. Control- left at 13°C.
The trial was replicated three times, of six fruits each, for three consecutive weeks. The fruits were ripened under standard Chiquita ripening conditions at 16°C-18°C, 95% RH, ethylene at 300 ppm (Johnson, T. 2000, pers. comm.) until colour stage 4 (Commonwealth Scientific and Industrial Research Organisation (CSIRO), 1972) Fig. 3.1. They were then stored in shelves at 22°C, ripened in ventilated polyethylene bags until colour stage 6. RCBD design was used and parameters analysed were volatiles using HP 4440 Chemical Sensor (Wirthenson, et al, 1999), soluble solids content using a refractometer (Bellinghan and Stanley Limited code 20-60, England) and data were subjected to analysis of variance at 5 % (ANOVA), fruit firmness using a penetrometer (FT 327 (3-37 Lbs), Italy) and data were subjected to ANOVA at 5 % and sensory evaluation by an untrained sensory panel using a simple difference test (Meilgaard et al., 1999).
Fig. 3.1: Standard colour index for banana ripening where 1 = green, 2 = green with a trace of yellow, 3 =
more green than yellow, 4 = more yellow than green, 5 = yellow with green tip, 6 = fully yellow, 7= yellow, lightly flecked with brown and 8 = yellow with increasing brown areas. (Source: CSIRO, 1972)

Wesonga, et altropics Octobe
Results Sensory analysis Effect on banana aroma Table 4.1:Number of responses for banana cv. Williams that were or were not chilled, and required
responses to show significant difference.
Number of responses for control fruit
Number of responses for
mildly chilled fruit
Minimum responses for one category to establish significant difference at P<0.05
(Meilgaard et al., 1999) Week 1 14 13 20 Week 2 16 13 21 Week 3 17 24 28
Effect on banana sweetness Table 4.2:Number of responses for banana cv. Williams that were or were not chilled, and required
responses to show significant difference.
Number of responses for control fruit
Number of responses for mildly chilled fruit
Minimum responses for one category to establish significant difference at P<0.05
(Meilgaard et al., 1999) Week 1 19 8 20 Week 2 16 13 21 Week 3 16 25 28
Effect on physical quality Effect on pulp firmness
Bananas in the three harvest weeks had a pulp firmness of less than 1 kg. Effect on peel colour
Mild postharvest chilling (MPHC) discoloured the banana peel giving it a dull yellow colour compared to the control that had a bright yellow peel.
Fig 4.11
: Peel colour of summer grown Cavendish bananas cv. Williams that were or were not chilled at 5°°°°C for 24 hours and then ripened at 16°°°°C, 95% RH and 300., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the r 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 183
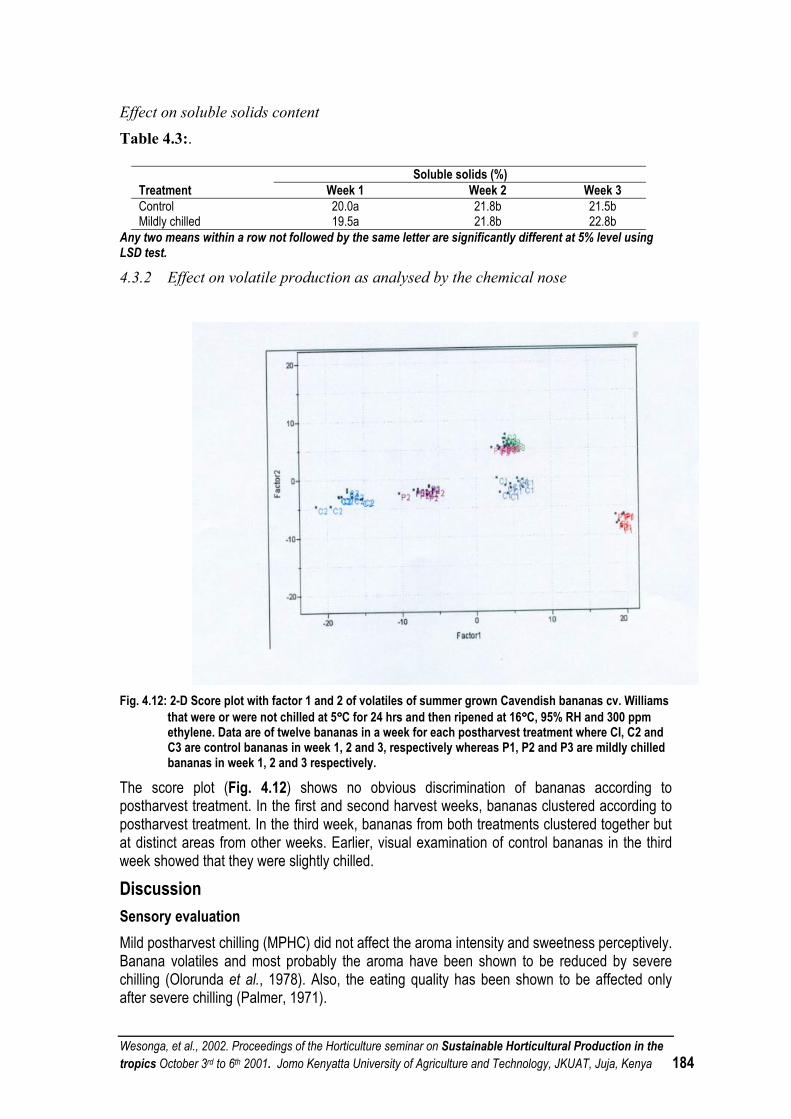
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 184
Effect on soluble solids content Table 4.3:.
Soluble solids (%) Treatment Week 1 Week 2 Week 3 Control 20.0a 21.8b 21.5b Mildly chilled 19.5a 21.8b 22.8b
Any two means within a row not followed by the same letter are significantly different at 5% level using LSD test.
4.3.2 Effect on volatile production as analysed by the chemical nose
Fig. 4.12: 2-D Score plot with factor 1 and 2 of volatiles of summer grown Cavendish bananas cv. Williams that were or were not chilled at 5°°°°C for 24 hrs and then ripened at 16°°°°C, 95% RH and 300 ppm ethylene. Data are of twelve bananas in a week for each postharvest treatment where CI, C2 and C3 are control bananas in week 1, 2 and 3, respectively whereas P1, P2 and P3 are mildly chilled bananas in week 1, 2 and 3 respectively.
The score plot (Fig. 4.12) shows no obvious discrimination of bananas according to postharvest treatment. In the first and second harvest weeks, bananas clustered according to postharvest treatment. In the third week, bananas from both treatments clustered together but at distinct areas from other weeks. Earlier, visual examination of control bananas in the third week showed that they were slightly chilled.
Discussion Sensory evaluation Mild postharvest chilling (MPHC) did not affect the aroma intensity and sweetness perceptively. Banana volatiles and most probably the aroma have been shown to be reduced by severe chilling (Olorunda et al., 1978). Also, the eating quality has been shown to be affected only after severe chilling (Palmer, 1971).

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 185
Pulp firmness (PF) was not affected by MPHC. This is desirable as PF determines texture and it positively affects eating quality (Israeli and Lahav, 1986). Poor texture leads to consumer criticism that results in reduced repeat buys, hence poor marketability. Peel colour was adversely affected by MPHC. Bright yellow peel positively influences purchase quality. Visual appearance is used to judge purchase quality (Kays, 1999). However, repeat purchase is influenced by flavour, aroma, texture and sweetness (Israeli and Lahav, 1986). Chemical nose The chemical nose was not able to discriminate bananas according to mildly chilled and control treatments in the three harvest weeks combined. Therefore, aroma intensity is also not expected to vary (Lavilla et al., 1999). This could be due to greater natural variation of bananas over weeks as opposed to variation due to mild postharvest chilling. Results from the chemical nose correlate very well with the sensory data.
Conclusion Mild chilling in storage and/or transport does not affect the eating quality of bananas perceptively. However, visual appearance was adversely affected. In the short term, the use of chemicals for removal of the discolouration is recommended. However, a long-term solution is to genetically engineer banana crops with low polyphenol oxidase activity in the peel (chilling resistant), but with good eating quality. Chemical nose can be used for the evaluation of banana aroma quality as it has several advantages as a quality assurance tool for the food industry.
References Commonwealth Scientific and Industrial Research Organisation, Australia Division of Research,
1972. Banana ripening guide, Circular 8. Melbourne, N.S.W. Hewage, K. S., Wainwright, H. and Wijeratnam, R. S. W. 1996. Quantitative assessment of
chilling injury in bananas using a calorimeter. J. Hort. Sci., 71(1), 135-139. Israeli, Y. and Lahav, E. 1983. Banana, In CRC Handbook of Fruit Set and Development. S. P.
Monselise (ed.), CRC Press, Boca Raton, pp. 45-73. Kays, S. J. 1999. Preharvest factors affecting appearance. J. Postharv. Biol. Technol., 15, 233-
247. Lavilla, T., Puy, J., Lopez, M. L., Recasens, I. and Vendrell, P. 1999. Relationships between
volatile production, fruit quality and sensory evaluation in Granny Smith apples stored in different controlled-atmosphere treatments by means of multivariate analysis. J. Agric. Food Chem., 47, 3791 - 3803.
Marriot, J. 1980. Bananas-Physiology and biochemistry of storage and ripening for optimum quality. CRC Critical reviews in Food Science and Nutrition, 13, 41-88.
Mattei, A. and Paillard, N. 1973. Effect of a variable storage period at the critical temperature of 12°C on the aroma of the banana (var. Poyo), at 20°C. Fruits, 33, 319.
Meilgaard, M., Civille, G.V and Carr, T. 1999. Sensory Evaluation Techniques. C. Anderson (ed), CRC Press, Boca Raton.
Olorunda, A.O., Meheriuk, M. and Looney, N.E. 1978. Some postharvest factors associated with the occurrence of chilling injury in banana. J. Sci. Food Agric., 29(3), 213-216.
Palmer, J.K. 1971. The banana, In Biochemistry of fruits and their products. A.C. Hulme (ed.), Academic Press, London. Vol. 2.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 186
Wang, C.Y. 1990. Alleviation of chilling injury of horticultural crops, In Chilling Injury of Horticultural Crops. C.Y. Wang (ed.), CRC Press, Boca Raton, pp. 281-302.
Wirthensohn, M.G, Francis, I.L., Gawel, R. and Jones, G.P. 1999. Sensory-Instrumental correlation of extra virgin olive oil aromas using a “Chemical Nose”, in Proceedings of the Australian International Symposium on Analytical Science, Melbourne, pp. 75-78.
Wills, R., McGlasson, B., Graham, D. and Joyce, D. 1998. An introduction to the Physiology & Handling of Fruit, Vegetable & Ornamentals. UNSW Press, Sydney.
POLLINATION CONSIDERATIONS IN SUSTAINABLE INTEGRATED HORTICULTURAL PRODUCTION SYSTEMS OF FRUIT AND SEED VEGETABLES IN THE TROPICS MRS GRACE NJERI NJOROGE JOMO KENYATTA UNIVERSITY, BOTANY DEPARTMENT, P.O. BOX 62000 NAIROBI E-MAIL: (I) [email protected] (ii) [email protected]
Abstract Pollination refers to the transfer of pollen from the male parts of the flower to the female parts. This is especially critical in plants where different sexes are found in different plants or flowers. Pollination is a resource that is vital to agricultural productivity. Insect pollinators for example are practically essential in fruit and vegetable crop production. This is especially because pollinators increase or enhance seed set, improve seed & fruit quality, as well as improve genotype progeny. Pollination may be a key to fruit and seed vegetable productivity when all the other conventional inputs of water, fertilizer and pest control are taken into consideration. The pollinators however are currently under threat arising from: Agricultural development, Habitat fragmentation, Agricultural chemicals (pesticides and herbicides), destruction of foraging and nesting sites, spread of pests and diseases. Bee poisonings for example from pesticides result in annual losses of $ 14.3 million in the USA alone. In Kenya and other parts of Africa pollination is viewed as a resource that will always be there. This implies that in cultivation of seed and fruit vegetables the pollination requirements of the crops are hardly considered. Because we still have the pollinators in the wild habitats they need documentation and conservation before they are decimated to unreasonable population levels. It is time we changed our view and seen pollination as a resource that can be depleted with very negative effects on our horticultural vegetable production. Many vegetables under cultivation in the tropics are directly dependent on pollination service for fruit and seed set. This paper explores the potential contribution of agricultural pollination service to vegetable production especially the cucurbits (Cucurbitaceae) by pollinators. Vegetable production management practices that favor pollination are discussed with the view that they need to be incorporated in sustainable production in future.
Introduction The Cucurbits (Cucurbitaceae) Cucurbitaceae Jussieu are a family of dicotyledonous plants comprising about 825 species in 118 genera (Zomlefer, 1994). This family is predominantly tropical, having 90% of the species in three main areas- Africa and Madagascar, Central and South America, and Southeast Asia and Malesia (Jeffrey, 1990). There are several cultivated species of the Cucurbitaceae that are thought to have their origin in Africa. These include Cucumis melo, Cucumis sativus, and Citrullus lanatus (Kirkbrid, 1993). The family stands out as the second important family in providing the world with fruit and vegetables, the first being the Solanaceae Juss. (Purseglove 1968). There are 15 species of Cucurbitaceae under cultivation, and many of these figure prominently in the export vegetable industry of Kenya.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 187
The fruits and vegetables obtained from this family are rich in vitamins, minerals and ß-carotene. Some of the important genera are: Benincasa Savi., Momordica L., Sechium P. Br., Cucumis L., Cucurbita L., Lagenaria Ser., Luffa Mill., Citrullus Eckl. & Zeyh., (syn. Colocynthis Mill. nom. rej) and Trichosanthes L., (Purseglove, 1968). Citrullus Cucurbita and Cucumis are the main genera cultivated in the tropics for their edible fruits (Cobley, 1965, Purseglove, 1968). In Kenya Cucurbitaceae, vegetables are gaining importance in the export market. Most of these are now being grown in arid and semi-arid regions of Kenya where Pastoralism is slowly giving way to horticulture. Pollination Service in Cucurbit Vegetable Production Cucurbits are largely cross-pollinated, and mostly this is accomplished by insects. Melons and Cucumbers for example produce large pollen grains but in low quantities; hence their dependence on effective pollinators (Free, 1993; Hodges & Baxendale, 1998) Pollinators also accelerate fruit set and early fruit development in cucumber. Pollen grains are sticky to the extent that even in hermaphroditic flowers which are self fertile no automatic self-pollination takes place because when the pollen is dehisced they adhere to the anthers (Free, 1993). Anthers in these species dehisce outwardly such that pollen is deposited at the based of the corolla and hence does not reach the stigmas unless bees and other insects carry them. Whiles these vegetables could be hand pollinated this is not only expensive but also produces fruit of uneven shape hence reducing vegetables quality especially those meant for export purposes. Cucurbits need abundant deposit of pollen to form viable fruits, and the more pollinators visit to a female flower, the larger and more vigorous is the fruit that results. The number of bee visitations to flowers of cucumber and watermelon can be a limiting factor in commercial production of these crops. In cucumber 8-10 bee visits are necessary for satisfactory fruit set while in water melons a minimum of 8 bee visits are necessary for normal fruit development. Pollination is such a concern in these vegetables production because of the added factor that majority of them are hybrids. Hybridisation reduces the amount of pollen available for pollination. Triploid watermelons for example have staminate flowers with much less pollen than the diploid cultivars. Because of their relative weights and stickiness, pollen grains in this family cannot be transported by wind. This therefore makes the cucurbits desperately dependent on animal pollen vectors for reproductive success. Cucurbits are particularly vulnerable to pollinator loss because their pollinators are thought to often be Oligolectic, that is, specializing on a few related plant species for pollen resources. Specialized pollinators may be less likely to adapt to changes in available plant resources, such as occurs with agricultural development. Of the 28 genera in East Africa nine of these have all their species as diocious (Jeffrey, 1967). The dioecious reproductive strategy is thought to have evolved under conditions of reliable pollinators (Darwin 1876, Renner and Feil, 1993); as ecosystems become disrupted, the cucurbit strategy may be an especially fragile one. Integration of Pollination Aspects in Horticultural Production Systems of Cucurbits In Kenya cucurbits are mainly grown for horticultural purposes in arid and semi arid lands (ASAL) usually under irrigation. The main pollinators of wild and cultivated species of the representative species of the genera Cucumis and Citrullus have been collected identified and
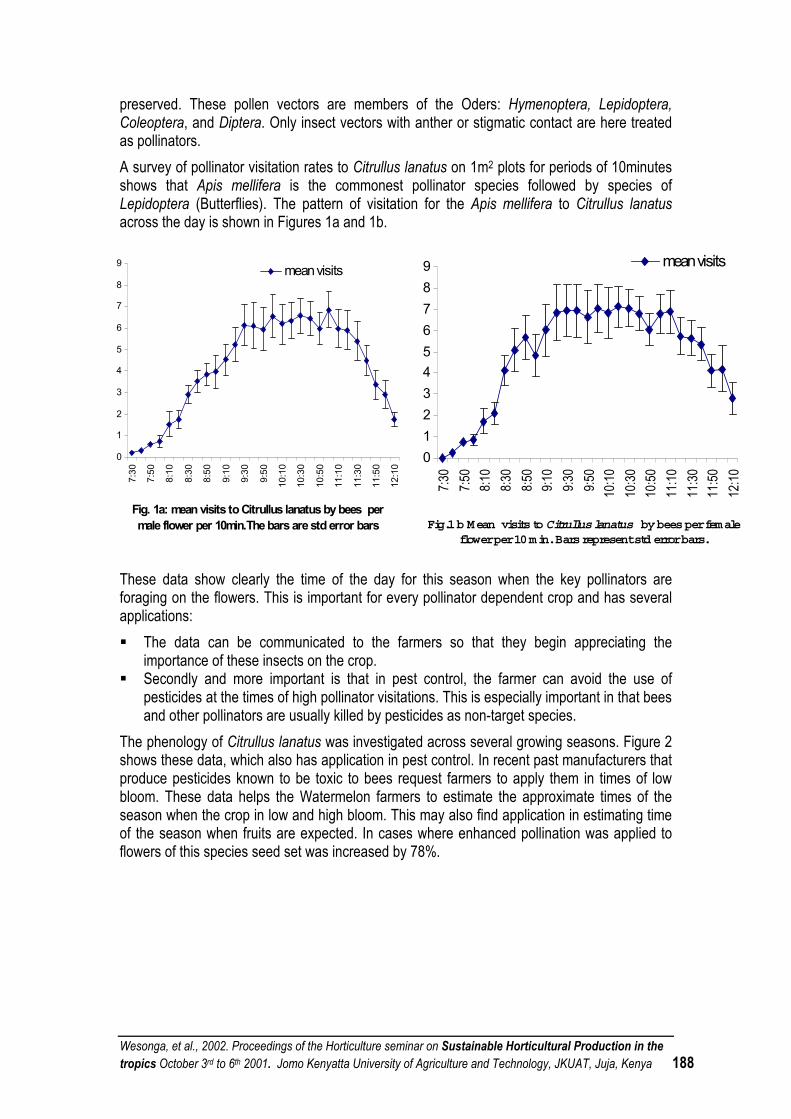
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 188
preserved. These pollen vectors are members of the Oders: Hymenoptera, Lepidoptera, Coleoptera, and Diptera. Only insect vectors with anther or stigmatic contact are here treated as pollinators. A survey of pollinator visitation rates to Citrullus lanatus on 1m2 plots for periods of 10minutes shows that Apis mellifera is the commonest pollinator species followed by species of Lepidoptera (Butterflies). The pattern of visitation for the Apis mellifera to Citrullus lanatus across the day is shown in Figures 1a and 1b.
These data show clearly the time of the day for this season when the key pollinators are foraging on the flowers. This is important for every pollinator dependent crop and has several applications: The data can be communicated to the farmers so that they begin appreciating the
importance of these insects on the crop. Secondly and more important is that in pest control, the farmer can avoid the use of
pesticides at the times of high pollinator visitations. This is especially important in that bees and other pollinators are usually killed by pesticides as non-target species.
The phenology of Citrullus lanatus was investigated across several growing seasons. Figure 2 shows these data, which also has application in pest control. In recent past manufacturers that produce pesticides known to be toxic to bees request farmers to apply them in times of low bloom. These data helps the Watermelon farmers to estimate the approximate times of the season when the crop in low and high bloom. This may also find application in estimating time of the season when fruits are expected. In cases where enhanced pollination was applied to flowers of this species seed set was increased by 78%.
Fig.1 b Mean visits to Citrullus lanatus by bees per female flower per 10 min. Bars represent std error bars.
0123456789
7:30
7:50
8:10
8:30
8:50
9:10
9:30
9:50
10:10
10:30
10:50
11:10
11:30
11:50
12:10
mean visits
Fig. 1a: mean visits to Citrullus lanatus by bees per male flower per 10min.The bars are std error bars
0
1
2
3
4
5
6
7
8
9
7:30
7:50
8:10
8:30
8:50
9:10
9:30
9:50
10:1
0
10:3
0
10:5
0
11:1
0
11:3
0
11:5
0
12:1
0
mean visits
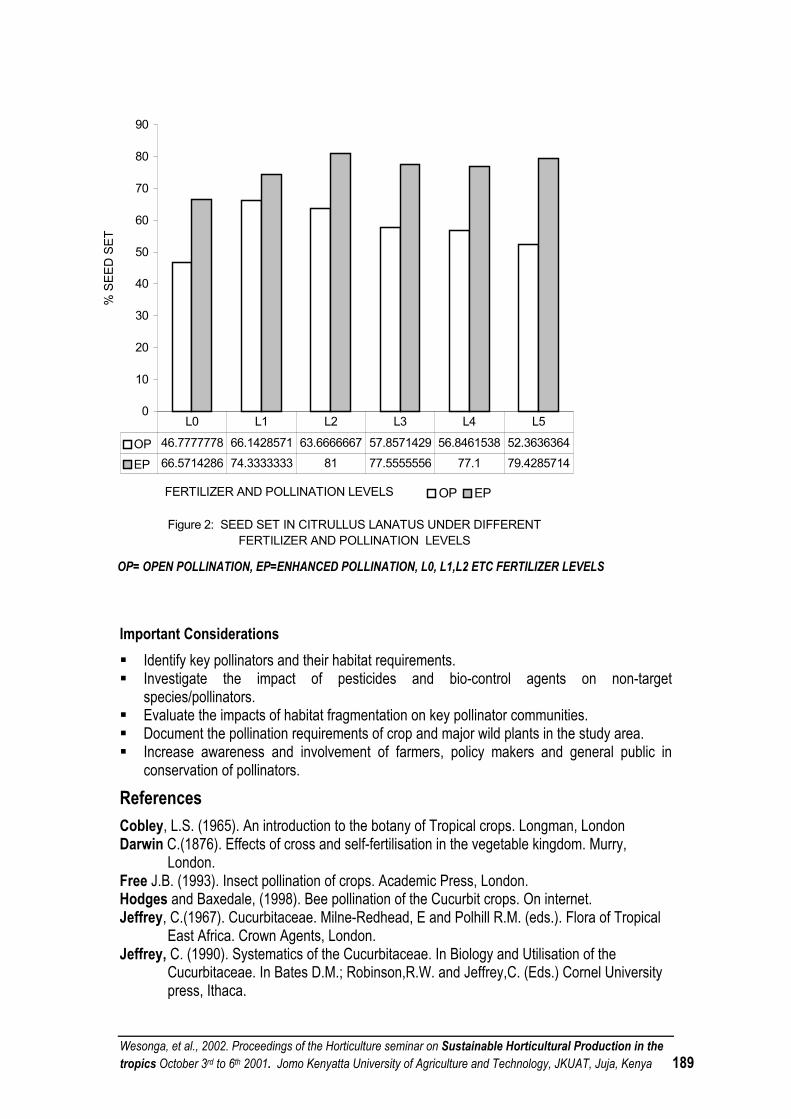
Figure 2: SEED SET IN CITRULLUS LANATUS UNDER DIFFERENT FERTILIZER AND POLLINATION LEVELS
0
10
20
30
40
50
60
70
80
90
FERTILIZER AND POLLINATION LEVELS
% S
EED
SET
OP EP
OP 46.7777778 66.1428571 63.6666667 57.8571429 56.8461538 52.3636364
EP 66.5714286 74.3333333 81 77.5555556 77.1 79.4285714
L0 L1 L2 L3 L4 L5
OP= OPEN POLLINATION, EP=ENHANCED POLLINATION, L0, L1,L2 ETC FERTILIZER LEVELS
Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 189
Important Considerations Identify key pollinators and their habitat requirements. Investigate the impact of pesticides and bio-control agents on non-target
species/pollinators. Evaluate the impacts of habitat fragmentation on key pollinator communities. Document the pollination requirements of crop and major wild plants in the study area. Increase awareness and involvement of farmers, policy makers and general public in
conservation of pollinators.
References Cobley, L.S. (1965). An introduction to the botany of Tropical crops. Longman, London Darwin C.(1876). Effects of cross and self-fertilisation in the vegetable kingdom. Murry,
London. Free J.B. (1993). Insect pollination of crops. Academic Press, London. Hodges and Baxedale, (1998). Bee pollination of the Cucurbit crops. On internet. Jeffrey, C.(1967). Cucurbitaceae. Milne-Redhead, E and Polhill R.M. (eds.). Flora of Tropical
East Africa. Crown Agents, London. Jeffrey, C. (1990). Systematics of the Cucurbitaceae. In Biology and Utilisation of the
Cucurbitaceae. In Bates D.M.; Robinson,R.W. and Jeffrey,C. (Eds.) Cornel University press, Ithaca.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 190
Kirkbried,J.H. (1993). Biosystematic monograph of the genus Cucumis (Cucurbitaceae) Botanical identification of cucumbers and melons. Parkway, Carolina.
Purseglove,J.W. (1968). Tropical crops. Longman group, Harlow. Renner,S.S. and Feil, J.P. (1993). Pollinators of Tropical dioecious Angiosperms. Amer. Journ.
Bot. 80(9): 1100-1107 Zomlefer, W.B. (1994). Guide to the flowering plant Families. Carolina press, Chapel Hill. Bernardello, L.; Galetto, L. & Rodriguel, I.G. (1994). Reproductive biology, variability of nectar
features and pollination of Combretum fruticosum (Combretaceae) in Argentina. Botanical Journal of the Linnean Society 114: 293-308.
Bowers, K. A. (1975). The pollination and Ecology of Solanum rostratum (Solanaceae). Amer. J. Bot. 62 (6): 633-638.
Buchmann,S. & Nabhan, G.P. (1996). The forgotten pollinators. Washington D.O. Island Press. Gemmill, B. 1998. Preliminary results of the pollination of Bitter gourd (Momordica charantia). Ingram, M.; Nabhan, GP.; Buchmann, S. (1996a).Ten essential reasons to protect the birds
and the bees. Arizona-Sonora Desert Museum, Tueson, AZ (www.desertmuseum.org/fp/ten reasons.html)
Kearns,C.A. and Inouye, D.W. (1997). Pollinators, Flowering Plants and Conservation Biology. Bioscience 47:297-307.
Kearns,C.A.; Inouye, D.W;Waser, N.M. (1998). Endangered mutualisms:The conservation of plnat-pollinator interactions. Annu. Rev. Ecol. Syst. 29:83-112
Newton L.E. (1994). Observations of flowering of Sansevieria robusta in Kenya. E.A.N.H.S Bulletin 24 (1): 8-10.
Roubik, D.W. (1995). Pollination of cultivated plants in the Tropics. FAO, Rome Spears, E.E. (1983). A direct measure of pollinator effectiveness. Oecologia 57:196-199 Wardelll, G.A.; Bernhardt, P;BitnerR,;Burquez,A.; Buchmann, S.(1998). The potential
consequences of pollinator declines on the conservation of Biodiversity and stability of food crops. Conservation biology 12(1): 8-17
Watanabe, M.E. (1994). Pollination worries rise as honey bees decline. Science 265:1170.
AGRO-METEOROLOGICAL INFLUENCES IN COFFEE PRODUCTION- IMPLICATIONS FOR IMPROVED COFFEE PRODUCTION IN KENYA Mutua, J. N. Coffee Research Foundation, P.O. Box 4 Ruiru. Email: [email protected]
Global weather forecasts are at the mercy of unpredictable variables and the most important is their impact on world agriculture. In the recent past, there has been a decline in coffee production in most coffee growing areas in Kenya. A combination of factors has been attributed to this negative trend but the most common of them all is the apparent increase in production costs and risks resulting from unfavourable weather conditions and insect pests and disease incidences. With the current climate change, global warming is likely to produce more erratic weather especially in lower altitudes. The effects of this has already been felt in most coffee growing areas in the country where rainfall has become unpredictable with the onset of rainfall in March and October tending to be delayed to late March and April for long rains and the short rains to November. Approximately £1 billion is spend each year on variable costs of growing arable crops mainly in fertilizer and chemical sprays, and much of this expenditure is weather dependent. This amount could be saved if agrometeorological information involving weather forecast is availed to farmers in good time. Timing, application technique and the quantity of nutrients to be applied is subject to weather conditions and any modification from the recommended period/amount is based on weather forecast. Disease control programmes, insect pest management, weed control and pruning are other major coffee management practices directly influenced by weather.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 191
This presentation is aimed at identifying coffee management practices that are weather dependant and how coffee farmers can identify and utilize agrometeorological parameters that influence these practices in making tactical and strategic farm management decisions and is based on monitoring survey carried out in 18 coffee growing districts. Critical parameters used on the basis of monitoring and forecasting changes in weather and the extent to which all these parameters affect production are discussed.
BIO-ALGEEN: ECONOMIC BENEFITS WITH ENVIRONMENTALLY FRIENDLY PRODUCTS AND PRODUCTION Freidel, J.W. Biological Environmental Systems and Products Ltd. (BESyP)
Growing awareness on environmental issues, especially in food production, demands production inputs that meet sustainable production quantity with required quality standards (e.g., 0-residue level). Plant extracts that are increasingly known and used in agriculture production are largely providing sustainable yields with the required quality standards. Most of these products, however, are merely replacing chemical pesticides as quick action solution with therapeutic results. But on the other hand there exist plant extracts that act in a prophylactic way, so to speak. They are known (and registered) as plant growth stimulants and soil conditioners. Hence the products are not fertilizers, nor pesticides - like the bio-algeen range of products. Bio-algeen products are plant extracts from marine algae (Ascophyllum nodosum) growing in the cold seas of the northern hemisphere. It is a renewable source being environmentally friendly right from the production, since the plants are only mown, then dried with geothemal heat and milled within 24 hours from harvest, with a cold extraction process. Those products are creating a healthy plant through stimulating increased root growth, balancing nutrient availability in the soil, improving soil fertility with a balanced soil life, stimulating microbiological activity with improved uptake of water and nutrients. A vigorous growth with healthy plants is the consequence, with increased yields (up to 35%) and less susceptibility towards health problems (less spray rounds); therefore they are even requested by the producers of commercial organic production. Through a so-called public private partnership program (PPP) between the manufacturer of bio-algeen and the German government, delegated to BESyP, we conduct training of farmers (out-growers and large scale) introducing environmentally friendly products and production.

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 192
OFFICIAL CLOSING SPEECH SPEECH BY MR. FREDRICK P. MUEMA, DEPUTY DIRECTOR OF AGRICULTURE IN-CHARGE OF HORTICULTURE DURING THE OFFICIAL CLOSING OF THE HORTICULTURE SEMINAR ON SUSTAINABLE HORTICULTURAL PRODUCTION IN THE TROPICS OCTOBER 4th, 2001
Regional Director, DAAD Dr. C. Etzold Dean Faculty of Agriculture, Directors of Institutes Distinguished Participates Ladies and Gentlemen. I feel greatly honoured to be with you this evening to perform the official closing ceremony of this workshop on sustainable horticultural production in the tropics. It is refreshing to note that you have for the last two days been following illuminating presentations from over 40 scientists drawn from Kenya, Tanzania, Ethiopia and the Federal Republic of Germany. The discussions, I am informed have revolved around the various horticultural themes such as crop physiology and harvest improvement and nursery management. The workshop also intensely examined at great length on the socio-economic impact of horticulture and on food harvesting technologies. Ladies and Gentlemen, the importance of the horticultural sector in the country cannot be overemphasised. It is currently the second largest foreign exchange earner in Kenya after tourism. The sector has grown steadily accounting for Kshs.3.6 and 10.6 billion in foreign exchange earnings in 1991 and 1995 respectively. Last year, foreign exchange earnings from horticulture stood at Kshs.14 billion. In addition the small-scale farmers who account for 70 percent of the sector dominate horticultural activities in the country. It is therefore the key income source to most of our people. Ladies and Gentlemen, the government would continue to encourage universities to hold workshops of this kind, because results from such sessions have a definite potential to stir up horticultural production. The consequences would clearly translate into improved incomes and reduced poverty to our farmers. As a ministry in charge of agriculture, we believe that a forum, as this one is crucial because it imparts useful knowledge to the participants. Such sessions also act as a good beginning for meaningful collaboration between the universities, national research institutions, the private sector and more importantly the farmers. As scientists, you must endeavour to provide solutions militating against the full exploitation of horticulture in the country. You should be able to quickly translate your research findings into appropriate applied technologies that would be available for use to our farmers. This way the objectives of research and development would be achieved. Africa, now plagued with endless calamities ranging from high poverty levels and soaring child mortality rates, raging famine and low agricultural earnings, is a great scandal to the academia of Africa. Your fellow countrymen are patiently but urgently waiting for solutions that would reduce their misery. Finally, I want to extend my gratitude to the German Academic Exchange resources Service; DAAD for making the financial that has culminated in the successful staging of this workshop. The joint effort between the Jomo Kenyatta University of Agriculture and Technology and the

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 193
University of Hannover, Germany represented in this workshop by several professors is commendable. It is now my pleasure to declare the workshop on sustainable horticulture production in the tropics officially closed. Thank you

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 194
SEMINAR PROGRAM
DAY 1: WEDNESDAY, October 3rd 2001
8:00 –10:00 AM REGISTRATION
10:30 AM POSTERS HAVE TO BE IN PLACE
11:00 - 12:30 PM OFFICIAL OPENING Welcome addresses
From the organizers:
Prof. Dr. S.G. Agong, JKUAT
Prof. Dr. H. Stuetzel, University of Hannover, Germany
From the hosts:
Dr. C.K. Ndung’u, Chairman, Department of Horticulture, JKUAT
Prof. Dr. F. Lenga, Dean, Faculty of Agriculture, JKUAT
Prof. Dr. K. Sinei, Deputy Vice-Chancellor (Research, Production & Extension), JKUAT
From the Sponsors:
Dr. C. Etzold, Regional Director, DAAD (German Academic Exchange Service)
Keynote address:
-Hon. Dr. Bonaya Godana, Minister of Agriculture
12:30 – 1:30 PM LUNCH
1:30 – 3:00 PM Session 1: Socio-Economic Issues
Chairperson: Prof. J.C. Onyango
Rapporteur: Mr. A.O. Watako
Assistant: Ms W. Kariuki
Mushroom Production in Sustainable Small-Scale Farming Systems: Opportunities and Constraints – A Survey of Uasin Gishu District Buigut, S.K.
Characteristics of French Bean (Phaseolus vulgaris) Production: A Case Study at Kibirigwi Irrigation Scheme in Central Province of Kenya Wamocho, L.S.; Ombwara, F.K.; Wesonga, J.M.; Sichangi, P.
Snow Pea and Sugar Snap Production in Mt. Kenya Region: Status. Challenges, and Opportunities Njuguna, L.; Kibaki, J.; Muchui, M.; Karnbo, C.M.; Wepukhulu, S.B.; Munene, S; Kinyua, G.K.
Post Harvest Handling of Cut Flowers Mutui, T.M.
3:00 – 3:30 PM: POSTER SESSION with Tea Break

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 195
4:00 - 6:00PM Session 2: Plant Protection
Chairperson: Prof. B. Hau
Co-Chairperson: Dr. S. Sithanntham
Rapporteur: Dr. P.G. Home
Assistant: Mr. T. Losenge
Introductory remarks: Interactions in Plant Disease Epidemics - Hau, B.
Diamondback Moth, Plutella xylostella (L.), on Peas in Kenya: Impact of the Host Shift on the Pest and Its Parasitoid Löhr, B.; Rossbach, A.
Phaeoramularia Leaf and Fruit Spot of Citrus: A Major Threat to Citrus Production in Tropical Africa - Past Experience and Future Perspectives. Mohammed, Y.
Raphanus raphanistrum L. as a Diversionary Host of Brevicoryne brassicae L. and Bagrada cruciferarum Kirk. Makwali, J.A; Wanjala, F.M.E.; Khaemba, B.M..
Towards Sustainable Crop Protection through Utilising Favourable Cultural Practices: Potential in Vegetable Crops Sithanantham, S.; Agong, S.; Matoka, C.; Raini, R.; Makatiani, J.
Potential for Enhancing the Impact of Augmentation Biocontrol of Two Key Caterpillar Pests on Important Vegetable Crops in Kenya Sithanantham, S.; Maniania, N.K.; Osir, E.O.; Baya, J.
Host Plant Resistance in Okra (Abelmoschus esculentus (L) Moench) Varieties Wanja, E.W.; Maranga, R.O.; Hallett, R.
6:30 - 8:30PM WELCOME PARTY

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 196
DAY 2: THURSDAY, October 4th 2001
8:30 – 10:00 AM Session 3: Tomato and Okra Production
Chairperson: Dr. A. Fricke
Rapporteur: Mr. G. Mwago
Assistant: Mr. P. Mulyungi
Introductory remarks: Some Aspects on Sustainable Tomato Production - Fricke, A.
Response of Tomato (Lycopersicon esculentum L.) Genotypes to Induced Salt Stress Agong, S.G.
Genetic Transformation of Tomato Lines for Resistance of Cucumber Mosaic Virus (CMV) Kusolwa, P.
Effects of Duration of Soaking and Concentration Levels of Salicylic Acid on Seed Germination of Cabbage (Brassica oleracea var. capitata L.), Cucumber (Cucumis sativus L.) and Tomato (Lycopersicon esculentum Mill. ) K’Opondo, F.O., Auma, E.O; Mathenge P.W.; Groot, S.P.C.
Effect of Irrigation Management on Yield, Water and Nitrogen Use of Okra (Abelmoschus esculentus L. ) Home, P.G.
10:00 – 11:00 AM POSTER SESSION with Tea Break
11:00-12:40 PM Session 4: Crop Improvement and Propagation
Chairperson: Prof. S.G. Agong
Co-Chairperson: Dr. J.A. Okeno
Rapporteur: Mr. P. Masinde
Assistant: Mr. H. Murage
Introductory Remarks: Genetic Improvement for Sustainable Vegetable Production - Agong, S.G.
Propagation of Kei Apple (Dovyalis caffra Warb.): TPRI Nursery Experience Bujulu, E.; Mkenda, J.E.
Selection of Macadamia Rootstocks Least Susceptible to Chlorosis and Necrosis Ngugi, S., Mbaka, J.; Gitonga, L.; Wepukhulu, J.B.; Wamocho, L.S.
Snap Bean Improvement in Kenya Odindo, A.O.; Van Rheenen, H.A.
Effect of Explant Type on GUS Gene Expression and Shoot Regeneration from Brussels Sprouts after Exposure to Agrobacterium tumefaciens - Gitonga, L.; Graham, C. The Effect of Preparation Methods on the Nutritional Quality of Some Vegetables Eaten in Kenya Makokha, A.O.; Kebenei, D.K.
12:40 - 1:30 PM LUNCH

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 197
1:30 - 3:30 PM Session 5: Plant Nutrition and Crop Physiology
Chairperson: Prof. H. Stützel
Co-Chairperson: Dr. C.K. Ndung’u
Rapporteur: Mr. F. Ombwara
Assistant: Dr. D. Shitanda
Introductory Remarks: Principles of Sustainable Nutrient Management in Vegetable Production - Stützel, H.
Rationalization of Nutritional Requirements for Sustainable Coffee Production in Kenya Bunyasi, S.W.; Mburu, J.N.
African Indigenous Vegetables: Opportunities and Constraints Onyango, M.O.A.
Influence of Water Level and Genotype on Transpiration of Spider Plant and Black Nightshade in Pot Experiments Masinde, P.W.
Laboratory Methods in Screening Crops for Aluminium Tolerance Mugai, E.N.; Agong, S.G.
Possible Role of Daylength on Grapevine Performance in the Tropics Ndung’u, C.K.
Storage and Processing Potentials for Horticultural Produce in Kenya Shitanda, D.
3:30 - 4:00 PM TEA BREAK
4:00 - 5:30 PM GROUP DISCUSSIONS ON RESEARCH NETWORKS
5:30 - 6:00 PM CLOSING CEREMONY
DAY 3: FRIDAY, October 5th 2001
8:00 AM - 6.00 PM EXCURSION: Kenya Wine Agencies Limited
Mwea Irrigation Scheme
DAY 4: SATURDAY, October 6th 2001
7.30 AM - 3:00 PM EXCURSION:
Sirimon/Ibis Farm, Timau
Kibirigwi Irrigation Scheme

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 198
LIST OF PARTICIPANTS & DISTINGUISHED GUESTS 1. Abiero-Gary
Department of Civil Engineering JKUAT P.O. Box 62000, NAIROBI - KENYA
2. Abukutsa Onyango, Mary Oyiela Department of Horticulture, Maseno University, P.O. Private Bag, Maseno, KENYA Tel: 254-35-51620/2, Fax: 254-35-51221/51153 E-mail: [email protected]
3. Agong StephenGaya Department of Horticulture, JKUAT, P .O. Box 62000 Nairobi, KENYA
4. Alice Nakhumicha Muriithi Kenya Methodist University P.O. Box 267 Meru, KENYA
5. Babalola, O. Olubukola., ICIPE P.O. Box 30772 NAIROBI - KENYA Email: [email protected]
6. Baya J. Mbanzu ICIPE P.O. Box 30772 NAIROBI - KENYA
7. Buigut, Stephen Department of Agricultural Economics, Egerton University, P.0. Box 536, Njoro, KENYA Email: [email protected]
8. Bujulu, Elvania Tropical Pesticides Research Institute (TPRI) P.O. Box 3024, Arusha, TANZANIA Email: [email protected]
9. Bunyasi Simon Wanyama. Coffee Research Foundation P.O. Box 4 Ruiru, KENYA
10. Catherine Ngamau African Biotechnology Stakeholders Forum (ABSF)/ISAAA P.O. Box 66069,Nairobi KENYA
11. Chemwetich, Joseph R. Department of Botany, Moi University, P.O. Box 1125 Eldoret, KENYA
12. E. N. Ireri Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
13. Elmada O. Auma Moi University P.O. Box 1125 Eldoret. KENYA Tel: 0321-63160
14. Esther Kabugi P.O. Box 35419, Nairobi, KENYA
15. Dr. C. Etzold, Regional Director, DAAD (German Academic Exchange Service) P.O. Box 14050, Nairobi, KENYA
16. F. K. lenga Biomechanical and Environmental Department (BEED)
JKUAT. P.O. Box 62000 Nairobi, KENYA
17. Freidel, J.W. Biological Environmental Systems and Products Ltd. (BESyP)
18. Gatahi Mwago Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
19. George Nyarega Omwoyo Department of Horticulture JKUAT. P.O. Box 62000 Nairobi
20. Gitonga, Lucy Njoki KARl, National Horticultural Research Centre, P.O. Box 220, Thika, KENYA, Email: [email protected]
21. Habib Mruttu Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
22. Hau, Bernhard Institute for Plant Diseases and Plant Protection, HerrenhauserStrasse 2 University of Hannover, D-30419 Hannover, GERMANY Email: [email protected]
23. Home, P.G. BEED, JKUAT, P.O. Box 62000, Nairobi, KENYA Email: [email protected]
24. Hunja Murage Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
25. J.C. Onyango Botany Department, Maseno University Private Bag Maseno, KENYA
26. John Wanjau Njoroge Kenya Insitute of Organic Farming (KIOF) P.O. Box 34972, Nairobi, KENYA
27. Joseph M. Kibaki National Horticultural Research Centre, P.O. Box 220, Thika, KENYA
28. Judith A. Makwali. Department of Zoology Moi University P.O. Box 1125, Eldoret-KENYA Email: [email protected]
29. K’Opondo, F.B.O. Department of Crop Production and Seed Technology, Moi University, P.O.Box 1125, Eldoret, KENYA. E-mail [email protected]
30. Kipsat Mary Jepkemboi Department of AMC Moi University P .0. Box 1125, Eldoret, KENYA E mail: [email protected]

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 199
31. Koech, Joel Moi University, P.O.Box 1125 Eldoret, KENYA Email: [email protected]
32. Kusolwa, Paul Sokoine University of Agriculture, Department of Crop Science and Production, P .0. Box 3005, Morogoro, TANZANIA.
33. Löhr, Bernhard Plant Health Division, International Centre of Insect Physiology and Ecology, P.O. Box 30772, Nairobi, KENYA, E-mail: [email protected]
34. Losenge Turoop Department of Horticulture JKUAT. P.O. Box 62000 Nairobi. Email: [email protected] or [email protected]
35. Macharia Ibrahim, Department of Horticulture JKUAT P.O. Box 62000, Nairobi, KENYA
36. Mahulo A.O Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
37. Makokha, Anselimo 0uma Department of Food Science and Post-harvest Technology, JKUAT, P.O. Box 62000, Nairobi, KENYA E-mail: [email protected]
38. Maranga, Rosebella Orangi Department of Zoology JKUAT P.O. Box 62000 Nairobi – KENYA Email: [email protected]
39. Masinde, Peter Wafula Department of Horticulture, JKUAT P.O. Box 62000 Nairobi. KENYA Email: [email protected]
40. Mathenge, P.W. Department of Crop Production and Seed Technology, Moi University P.O. Box 1125, Eldoret.
41. Matoka Chrales Mboya ICIPE P.O. Box 30772 NAIROBI – KENYA
42. Mohammed Yesuf Ethiopian Agricultural Research Organization (EARO) Melkassa Agricultural Research Center P.O. Box 436, Nazareth, Ethiopia. E-mail: [email protected]
43. Moshi, C. Siekuti Self Group, P. 0. Box 28, Tiliki/Crop protection Department, University of Nairobi P. 0. Box 30197, Nairob, KENYAi
44. Muchui, Margaret Nyatoro Kenya Agricultural Research Institute (K.A.R.I.) P.O. BOX 220 Thika, KENYA
45. Mugai, Elisha Njue Department of Horticulture, JKUAT P .O. Box 62000 Nairobi, KENYA
46. Muthoka, Nancy Munyiva National Horticultural Research Centre, P.O. Box 220, Thika, KENYA
47. Mutua, Joseph Ndunda. Coffee Research Foundation P.O. Box 4 Ruiru Email: [email protected]
48. Mutui Theophilus M. Department of Horticulture Moi University P.O. Box 1125, Eldoret, KENYA Email: [email protected]
49. Nderitu, John Huria Department of Crop Protection, University of Nairobi, P.O. Box 30197 Nairobi, KENYA. Email: [email protected]
50. Ndung’u, Cyrus K. Department of Horticulture JKUAT, P.O Box 62000 Nairobi, KENYA
51. Ng’ayu Beatrice Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
52. Ngamau Kamau Department of Horticulture JKUAT, P.O Box 62000 Nairobi, KENYA
53. Ngugi, Stephen National Horticultural Research Centre, P.O. Box 220, Thika, KENYA
54. Nicholas N. Kabare SCEM Farm Management Ltd. P.O. Box 260 Thika, KENYA
55. Njoroge, Grace Njeri Botany Department, JKUAT, P.O. Box 62000 Nairobi E-Mail: (i) [email protected] (ii) [email protected]
56. Njuguna, Lydia Wanjiru National Horticultural Research Centre, P.O. Box 220, Thika, KENYA
57. Odindo, A.O. Department of Crop Production and Seed Technology, Moi University, P.O. Box 1125, Eldoret, KENYA
58. Okeno, James Aketch. Department of Horticulture, Moi University, P. 0. Box 1125, Eldoret, KENYA. E-mail: [email protected]
59. Ombwara, FrancisKweyu., JKUAT, P.O. Box 62000, Nairobi, Kenya E-mail: [email protected]
60. Omunyin Michael Eleazar. Department of Horticulture, Moi University - Chepkoilel Campus, P.O. Box 1125, Eldoret, KENYA. E-mail: [email protected]

Wesonga, et al., 2002. Proceedings of the Horticulture seminar on Sustainable Horticultural Production in the tropics October 3rd to 6th 2001. Jomo Kenyatta University of Agriculture and Technology, JKUAT, Juja, Kenya 200
61. Ong'eng'a, W.O Department of Agricultural Marketing and Co-operatives, Moi University P.O. Box 1125 Eldoret. KENYA Tel: 0321-63160 E-mail: [email protected]
62. Onkware A. O. Moi University P.O. Box 1125 Eldoret. KENYA Tel: 0321-63160
63. Ouma, George Department of Horticulture, Maseno University P. 0. Box 333, Maseno, KENYA Telephone: (035) 51622 Or 51620. Email: [email protected]
64. Owino-Gerroh, C. Department of Horticulture Maseno University P.O.Box 333, Maseno, KENYA
65. Patrick Kavagi Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
66. Patrick M. Mulyungi Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
67. Samuel Gudu Moi University P.O. Box 1125 Eldoret. KENYA Tel: 0321-63160
68. Shitanda, Douglas Department of Biomechanical and Environmental Engineering JKUAT P.O. Box 62000 Nairobi, KENYA Tel: 0151-52029, E-mail: [email protected]
69. Sithanantham, S ICIPE P.O. Box 30772 NAIROBI - KENYA
70. Stuetzel, Hartmut Institute of Vegetables and Fruit Science HerrenhauserStrasse 2 University of Hannover, D-30419 Hannover, GERMANY Email: [email protected]
71. Waceke J. Wanjohi. Department of Botany, Kenyatta University, P.O. Box, 43844, Nairobi –KENYA.
72. Wamocho, L.S. JKUAT, P.O. Box 62000, Nairobi, Kenya E-mail: [email protected]
73. Wariara kariuki Department of Horticulture JKUAT. P.O. Box 62000 Nairobi, KENYA
74. Watako, Arnold Onyango Department of Horticulture JKUAT P.O. Box 62000 Nairobi - KENYA
75. Wesonga, J.M.. JKUAT P.O. Box 62000, Nairobi, Kenya E-mail: [email protected] or [email protected] or [email protected]