supply-side ecology, barnacle recruitment, and rocky intertidal community dynamics: do settlement...
TRANSCRIPT

Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
Contents lists available at ScienceDirect
Journal of Experimental Marine Biology and Ecology
j ourna l homepage: www.e lsev ie r.com/ locate / jembe
Supply-side ecology, barnacle recruitment, and rocky intertidal communitydynamics: Do settlement surface and limpet disturbance matter?
Bruce A. Menge ⁎, Melissa M. Foley 1, Jacque Pamplin, Gayle Murphy, Camryn PenningtonDepartment of Zoology, Oregon State University, Corvallis, OR 97331-2914, United States
⁎ Corresponding author.E-mail address: [email protected] (B.A. Men
1 Current address: Department of Ecology and EvoluCalifornia, Santa Cruz, CA 95064, United States.
0022-0981/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.jembe.2010.04.032
a b s t r a c t
a r t i c l e i n f oKeywords:Artificial collectorsBalanus glandulaBulldozingChthamalus dalliCypridsLimpetsOregonRecruitmentRocky intertidalSafety-walk platesSettlement
The supply of new recruits can be a critical determinant of community structure, but estimating recruitmentdensity can be challenging due to variation in larval supply, settlement, substratum, species interactions andphysical stresses. We evaluated the effect of surface type and limpet “bulldozing” on settlement andrecruitment of barnacles, using replicated field experiments sampled monthly at each of four sites along thecentral Oregon coast in 2001, 2003, and 2008. A particular goal was to evaluate the efficacy of Saf-T-Walk, awidely used textured surface, in quantifying actual patterns and levels of recruitment. In 2001 and 2003experiments, surfaces were bare rock (ROCK) and Saf-T-Walk (STW). In 2008 experiments, to furtherevaluate the influence of surface rugosity, we added two additional artificial substratum types — smooth PVCplates (no texture) and travertine plates (TRAV, low texture). Limpet activity was manipulated using barriersof anti-fouling paint. Results indicated the overwhelming importance of surface texture. Recruitment densitywas highest in 2001, lower in 2003, and lowest in 2008. Recruitment on STW, the most textured surface,ranged up to 87 times greater than on ROCK, and the rank order of surfaces for recruitment density wasSTWNROCKNTRAVNPVC which matches that for texture. Settlement differences were even greater, with upto 1180-fold higher settlement on STW than ROCK. Limpets usually had a negative effect on recruitment ofBalanus glandula on ROCK, likely due to bulldozing and direct consumption, but had a positive effect on STW.These impacts were reduced or negated when recruitment density was higher (e.g., 2001) or lower thanaverage (e.g., 2008). Limpets had no consistent effect on Chthamalus dalli. Abundances of settlers and recruitson the different surfaces were positively correlated (explaining up to 71% of the variance), and correlationsbetween STW and other surfaces generally were stronger in the absence of limpets. Although compared tobare rock, recruitment to STW overestimates absolute rates of recruitment, use of STW settlement platesprovides an effective and relatively efficient means of determining spatial and temporal patterns ofrecruitment at local to geographic scales.
ge).tionary Biology, University of
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
After decades of research focused at local scales, ecology hasundergone a dramatic shift in recent years towards investigation ofhow patterns and dynamics of species interactions vary acrossmultiple spatial and temporal scales (Levin, 1992; Brown, 1995;Holyoak et al., 2005). Investigations now routinely incorporatemultiple study sites often over vast spatial scales into their design(Jenkins et al., 2000; Sagarin and Gaines, 2002; Menge et al., 2004;O'Riordan et al., 2004, Navarrete et al., 2005; Coleman et al., 2006).Such studies are challenging because of variation among co-investigators, the biota, and physical conditions. For example,
incorporation of spatial scale requires standardization to ensure thatdifferences observed are independent of method.
In marine ecosystems, several recent large-scale studies exemplifythis approach and its challenges (Jenkins et al., 2000; Menge et al.,2003; O'Riordan et al., 2004; Coleman et al., 2006). These studies aretypically carried out by a consortium of institutions and a number ofinvestigators at each institution. For example, the PISCO (Partnershipfor Interdisciplinary Studies of Coastal Oceans) consortium is focusedon the linkages between coastal ocean environments and benthiccommunities, and how these vary through time and across space(Menge et al., 2004; Schoch et al., 2006).
A key potential determinant of community structure and animportant link between the coastal ocean and benthic habitats is thesupply of new recruits (Connell, 1985; Gaines and Roughgarden,1985; Connolly and Roughgarden, 1999). PISCO has focused mucheffort on the investigation of recruitment patterns of sessileinvertebrates to rocky intertidal regions and fish to kelp bed habitatsalong the coasts adjacent to the California current (Amman, 2004;

Fig. 1. Study sites along the Oregon coast. Cape Blanco and the Columbia River areshown for reference. Site codes are FC = Fogarty Creek, BB = Boiler Bay, YB = YachatsBeach, SH = Strawberry Hill, and TK = Tokatee Klootchman.
161B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
Broitman et al., 2008). In these large-scale investigations, animportant tool has been the adoption of standardized units toquantify recruitment that are inexpensive, uniform in size andcomposition, and readily deployed, retrieved and processed. Thesedevices include plastic mesh ovoids used to quantify musselrecruitment (Menge, 1992), and, our focus in this paper, settlementplates covered with a textured surface to quantify barnacle recruit-ment (Farrell et al., 1991).
Barnacles are ubiquitous in marine habitats, and in rocky intertidalsystems are usually dominant components of ecological communities(Lewis, 1964; Stephenson and Stephenson, 1972; Morton, 2004). Asexpressed in many influential studies, they are central to thedynamics of populations and communities around the world, servingas competitors, facilitators, and prey in a wide variety of settings (e.g.,Connell, 1961; Dayton, 1971; Menge, 1976 and many others).Barnacles are particularly prominent in studies of recruitmentbecause larvae, settlers and recruits are identifiable, abundant, andreadily studied in the field (e.g., Hawkins and Hartnoll, 1982;Wethey,1984; Connell, 1985; Caley et al., 1996).
In field investigations, two approaches have been taken to quantifysettlement and/or recruitment of barnacles. Some have investigatedrecruitment to natural rock substrata (e.g., Hawkins and Hartnoll,1982; Raimondi, 1988; Menge, 1991; Jenkins et al., 2000, 2008;Bertness et al., 2006) or to human-fabricated rock surfaces (Noda,2004), while others have used artificial settlement surfaces (e.g.,Pomerat and Weiss, 1946; Sutherland, 1974; Menge et al., 1999;Menge, 2000a). With artificial surfaces or collectors, a variety ofmaterials have been used including plexiglass plates with pits(Raimondi, 1990), unglazed ceramic plates (Broitman et al., 2005),and fiberglass, plexiglass, or PVC plates coated with a textured surface(Farrell et al., 1991; Pineda, 1994;Menge, 2000a) amongmany others.
The multiplicity of surfaces used in studies of recruitment raisesquestions of comparability among studies using different methodsand the extent to which artificial surfaces reflect patterns that occuron natural rock surface (Raimondi, 1990). Further, rock surfacevariation such as differences in rock type or texture may also causevariation in settlement on natural surfaces (Pomerat andWeiss, 1946;Barnes, 1956; Crisp, 1961; Raimondi, 1988). In addition, speciesinteractions can influence estimates of recruitment (Dayton, 1971,Hawkins, 1983). For example, Dayton (1971) showed that barnacleabundances were reduced by limpet “bulldozing,” defined asdislodgement of barnacle cyprids and recent metamorphs by theleading edge of the limpet shell as it grazes over the rock. Thus, ifbulldozing by limpets varies with their size, abundance, and speciescomposition, patterns of variation in recruitment of barnacles may beinfluenced by limpet abundance.
Here we address these issues using results from factorial experi-ments to evaluate the importance of substratum (natural rock orartificial settlement plates) and limpet bulldozing (present or absent)in determining barnacle recruitment density. Following earlierresearch (Connell, 1985; Raimondi, 1990), we define settlement asthe number of larvae attaching and surviving 24 h and recruitment asthe number of settlers surviving for up to one month. We addressedsix hypotheses:
H1. Recruitment of barnacles on natural rock surfaces will bepositively correlated with recruitment on artificial surfaces.
H2. With increasing texture on artificial surfaces, settlement andrecruitment densities will increase.
H3. Recruitment of barnacles will be lower and more spatiallyvariable on natural rock surfaces than on artificial surfaces becausenatural rock has lower and more spatially variable surface texture.
H4. Because of the negative effects of bulldozing, recruitment ofbarnacles will be lower in the presence of limpets than in their absence.
H5. Because of texturing that can provide refuges for new recruits,limpet bulldozing will have smaller effects on recruits on artificialsurfaces than on natural rock.
H6. recruitment of the coexisting acorn barnacle species Balanusglandula and Chthamalus dalli will be affected in similar ways bysubstratum and limpets.
2. Methods
2.1. Study sites
This studywas carried out atfive sites along the central Oregon coast(Fig. 1), with subsets of four sites used in each of 2001, 2003, and 2008.Replicate sites were located in regions of contrasting oceanographicconditions, which are associated with variation in the width of thecontinental shelf (Menge et al., 1997; Barth and Wheeler, 2005; Kosro,2005). In all years the two northern sites were located in the narrowcontinental shelf region (Fogarty Creek [both years] andBoiler Bay [bothyears], 44°51′ N and 44°50′ N, respectively) while the southern siteswere located in thewider continental shelf region (Yachats Beach [2003,2008], StrawberryHill [all years] and Tokatee Klootchman=Gull Haven[2001], southern sites; 44°17′N, 44°15′N, and 44°13′N, respectively). Atthe intertidal areas at Fogarty Creek and Boiler Bay (hereafter, FC andBB), abundance of macrophytes was relatively high, while abundancesof filter-feeders, predators, and macroherbivores were relatively low(Menge et al., 1994, 1997; Allison, 2004). In contrast, at Yachats Beach,Strawberry Hill, and Tokatee Klootchman (hereafter YB, SH, and TK) theabundance of macrophytes was relatively low and abundances of filter-feeders, predators, and macroherbivores were relatively high (Menge

162 B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
et al., 1994, 1997, 2004). Differences in community structure werereflected in differences in key dynamic processes. Rates of predation,mussel recruitment, and grazingwere all higher at southern sites thanatnorthern sites (Menge et al. 1994, 2004; Freidenburg et al., 2007).Barnacle recruitment, however, did not vary consistently among thefivesites (Barth et al., 2007). These community and process differences havepersisted since at least the early 1980s to present (Menge et al., 2004,Menge, B. A., pers. obs.).
2.2. Study design
To evaluate the simultaneous effects of limpet grazing andsettlement surface on the density of barnacle settlement (attachedcyprids) and recruitment (B. glandula and C. dalli), we establishedexperiments at wave-exposed locations at each site. All plots werepositioned at the lower edge of the mid zone, just below the musselbed. In 2001, five replicates of five treatments were set up in wave-exposed areas at FC, BB, SH, and TK (Table 1). In 2003, we substitutedYB for TK to make wave action more consistent across all sites. Sixtreatments were assigned randomly within each of five replicateblocks (Table 1). In 2008, we included two additional settlementsurfaces to the two used in the 2001 and 2003 designs to evaluate theinfluence of varying texture in addition to the effects of limpetgrazing. Eight treatments (four substrata×two grazing) in five blockswere included in the 2008 experiment (Table 1).
Although several barnacle species settled in these experiments(the above two plus Balanus crenatus, Balanus nubilis, and Pollicipespolymerus), recruitment of the latter three was sporadic and usuallyvery low except for occasional annual pulses of the gooseneckbarnacle Pollicipes. Thus we focused our analyses on B. glandula andChthamalus. Cyprids of these species are identifiable, but our methodof monitoring bare rock plots lacked sufficient resolution to separatethese species so we combined all cyprid counts for analysis.
Earlier sampling of daily plates indicated that cyprid metamor-phosis occurs within hours of settlement. We tested this assumption,and also evaluated the rate at which metamorphosis to juvenilebarnacles occurred by carrying out daily sampling for two four-dayperiods in July 2002 at the FC site. Three replicate settlement plateswere placed in the mid exposed zone on 09 July 2002 and collecteddaily through 14 July. This process was repeated 22–26 July. Based onthis study (see Results), we define cyprids attached to the rock assettlers and assume they reflect settlement over the past 24 h.
Table 1Design of experiments testing the effects of substratum and limpets on recruitment ofbarnacles.
Year Surface Treatment code Treatment description
2001 ROCK − limpets Paint barrier+limpets Marked plot
STW − limpets Painted plate− limpets Painted z-spar barrier+limpets Unpainted plate
2003 ROCK − limpets Paint barrier+limpets Marked plot+limpets Paint control
STW − limpets Painted plate+limpets Unpainted plate+limpets Paint control (partial paint barrier)
2003 STW − limpets Painted z-spar ramp (YB only)+limpets Unpainted z-spar ramp (YB only)
2008 ROCK − limpets Paint barrier+limpets Marked plot
STW − limpets Painted plate+limpets Unpainted plate
TRAV − limpets Painted plate+limpets unpainted plate
PVC − limpets Painted plate+limpets Unpainted plate
2.2.1. Substratum effectMany types of artificial surfaces have been tried (see Introduction),
and over the years we have tested unglazed ceramic tiles, a buildingmaterial called “Hardiflex,” plastic plates coated with latex paint withneoprenemicrospheresmixed in, Saf-T-Walk (see below), travertine andpolyvinyl chloride plate. Although each offers relative homogeneity insurface texture, all have shortcomings. For example, tile undersides weresmooth except for a square of grooves. Barnacles settled primarily alongthese grooves, leavingmost of the surface unoccupied (Gaines, S. D., pers.comm.; Blanchette, C. A., pers. comm.). Hardiflex and neoprenemicrosphere surfaces also proved similarly inadequate; we oftenobserved recruits on the rock nearby but almost none onHardiflex plates.
In this study,weused four different types of settlement surface. In the2001 and 2003 experiments, to evaluate the effect of settlement surface,we used 10×10 cm bare rock plots and 10×10 cm polyvinyl chloride(PVC) plates covered with Saf-T-Walk® tape (3M Company, St. Paul,Minnesota, USA)(Farrell et al., 1991; Menge, 2000a; Shkedy andRoughgarden, 1997). Saf-T-Walk tape (hereafter STW) is a rubberytextured surface that, since barnacles prefer to settle into pits andrugosities (e.g., Barnes, 1956, Crisp, 1961), provides barnacle cypridswithnearly ideal settlement surfaces andensures substratumuniformity(Farrell et al., 1991; see Fig. 2). The rock surface at all sites was basaltic, aconsiderably less rugose surface than STW. Bare rock (hereafter ROCK)plots were chosen to be representative of the texture of the rock at thesite (Fig. 2), but we avoided surfaces with crevices, pits, holes, grooves,and similar heterogeneities tomake these as comparable to the artificialcollectors as possible. This treatment allowed us to determine if platescovered by STW reflected the patterns of recruitment that occur onnatural substrata. We note that this design does not specifically test theeffects of rock texture or substratum type since neither of these wasvaried systematically in the design. Because of the differences in degreeof texturing onROCKvs. STW,weexpected toobserve lower recruitmentdensities on rock, but predicted that ROCK densities would be aconsistent fraction of the density on STW (Hypothesis H1).
In 2008, to evaluate the effects of surface texture, we added twoadditional surfaces to the design. The smooth surface of PVC plates withno STW (hereafter PVC) provided artificial surfaces with no texture andtravertine plates (hereafter TRAV) provided artificial surfaces with lowtomoderate texture (Fig. 2). Ranked by texture, the surface order is thusno texture (PVC), low texture (TRAV), moderate texture (ROCK) andhigh texture (STW)(Fig. 2). All artificial plates were soaked in seawaterfor at least a week to condition them before use in the field.
2.2.2. Limpet effectTo determine the effects of limpets on barnacle recruitment, we
established treatments with and without limpets (+limpet and− limpet, respectively). To exclude limpets from ROCK and STW plots,we surrounded one ROCK plot and one STW plate (2001 only) in eachreplicate with a 3 cm wide band of marine epoxy, z-spar™ (Kop-CoatInc., Los Angeles, California), which was then painted with a copper-based anti-fouling paint (hereafter termed z-spar+paint barriers).Copper paint barriers have been shown to impede limpets and chitons,in most cases with no artifacts due to the presence of the paint (Cubit,1984; Paine, 1992; Menge, 2000b; Freidenburg et al., 2007; Guerry,2008; Guerry et al., 2009; see Benedetti-Cecchi and Cinelli, 1997 forcritique). Since past experiments indicated that the z-spar+paintbarrier had to be repaired often, we also tested the effectiveness ofcopper paint in deterring grazers when coated on the sides and bottomof the artificial plates (all years). Because differences between thepainted plate and plateswith z-spar+paint barriers wereminimal (seeResults) we deployed only painted plates in the 2003 and 2008 studies.
2.2.3. Control experimentThe z-spar+paint barriers were relatively flat, so plate edges
protruded above the surrounding substratum by about 5 mm. Thisabrupt edge could possibly create differences in flow over plates vs.

Fig. 2. Close-up photographs of the artificial surfaces used in the experiments, and of the range of rugosity observed in the rock settlement plots. The holes in the plates are 8 mm indiameter. The area in the rock photos is scaled similarly to the area shown on the artificial plates. PVC= polyvinyl chloride plates, TRAV= travertine plates and STW= safety-walkplates. The differences in surface texture and relative homogeneity of the artificial plates is evident in the photos, as is the heterogeneity in natural rock surfaces.
163B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
ROCK plots, or even deter limpets from +limpet treatments, andcould also have affected barnacle recruitment. In 2003, we tested thispotential artifact at YB by deploying additional treatments thatincluded z-spar “ramps” around the plates to minimize eddies overthe plate edges. The two treatments were ramps with (− limpet) andwithout (+limpet) paint. All treatments are summarized in Table 1.
2.3. Monitoring procedures
The experiment was monitored monthly from June to September in2001 and 2003, and biweekly in July and August in 2008. Plates werereplaced at all sites during the same tide series, usually on consecutivedays. Limpets on plates (bottom and top) were counted in the field.Because small limpets are difficult to identify to species in the field, weidentified them to genus (Lottia) only. The species most likely to beincluded in this group are Lottia digitalis, L. pelta, L. strigatella, and anunidentified Lottia species. After collection, barnacle settlers (cyprids)and recruits (juveniles) on plateswere counted and identified to speciesin the laboratory under dissecting microscopes. ROCK plots were alsomonitored during the same tide series. All limpets in ROCK plots werecounted and in − limpet plots, removed. Barnacle recruits to the ROCKplots were counted and identified to species in the field using a fieldmicroscope (2001, 2003) or hand lens (2008). Field microscopes havegreater acuity than hand lenses but sampling the entire surface wasimpractical with microscopes so we sampled five haphazard fields ofview (0.1 cm in diameter) in each plot. Subsequent analysis suggestedthat subsampling introduced much variability, especially at lower
densities. Thus, in 2008 we switched to hand lenses and counted theentire plot. Recruit densitieswere standardized to anarea of 100 cm2 foranalysis. In the field, cyprids could not be resolved to species withmicroscopes, so in 2001 and 2003we lumped all taxa into a single countof “cyprids.”Hand lenseshave lower acuity thanmicroscopes, so in2008field counts of cyprids were unreliable and cyprid counts were onlyavailable from STW plates, which unlike other surfaces, were switchedout and processed in the laboratory.
ROCK plots were scrubbed clean using a stainless steel wire brushafter counts and rechecked to make sure all individuals were removed.On all surfaces filamentous algal blooms occurred at times, especially inthe absence of grazers. To help counting accuracy, we used bleach toremove the algae, either in the field (ROCK and TRAV) or in the lab(PVC). Counts in areas where recruits were visible prior to bleachingindicated that only algaewas removed, not settlers or recruits, allowingunobstructed counts of settlers and recruits prior to their removal. Priorto removal, percent cover of algae was recorded in all plots to allowtesting for potential interactions between ephemeral algae and recruits.
2.4. Data analysis
All analyses were done using JMP v. 8 (SAS, 2008). Establishment ofexperimental replicates takes 1–2 days, so it would be impractical toredo experimental setup eachmonth to vary the locationof each.Hence,ROCKplot locationswerefixed in space aswas the location of the plates,although the latter were replaced monthly. Because plot and replicatelocations were fixed, we used repeatedmeasures analysis of variance to
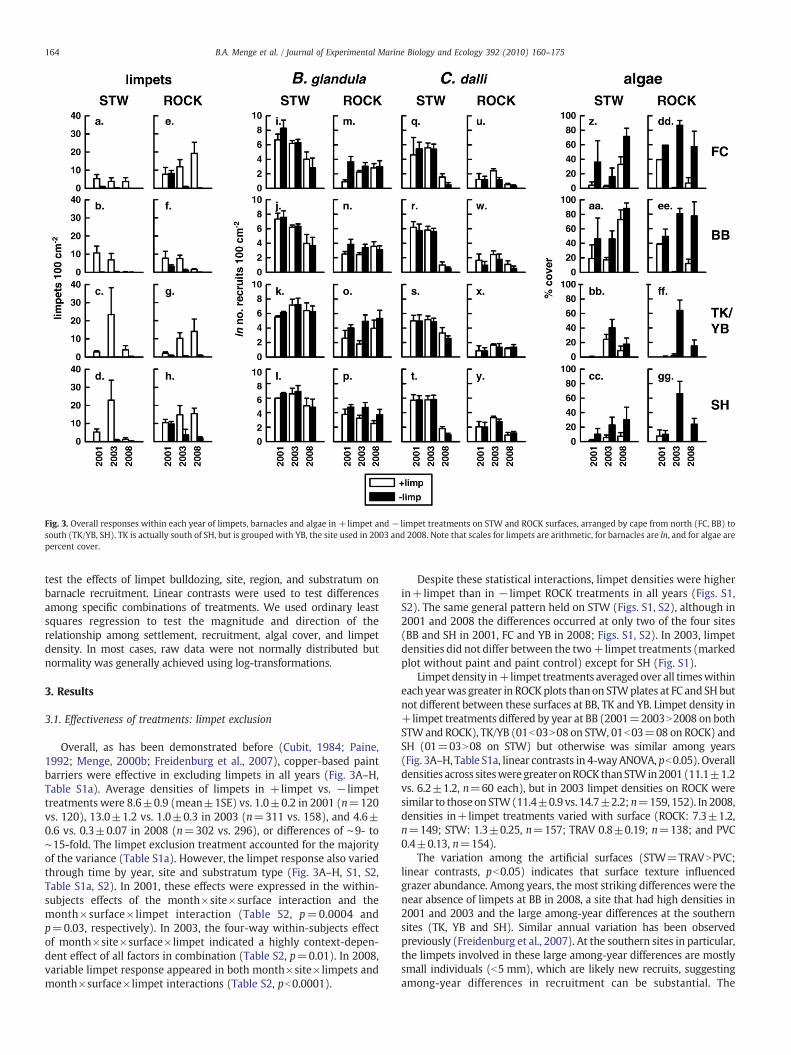
Fig. 3. Overall responses within each year of limpets, barnacles and algae in +limpet and − limpet treatments on STW and ROCK surfaces, arranged by cape from north (FC, BB) tosouth (TK/YB, SH). TK is actually south of SH, but is grouped with YB, the site used in 2003 and 2008. Note that scales for limpets are arithmetic, for barnacles are ln, and for algae arepercent cover.
164 B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
test the effects of limpet bulldozing, site, region, and substratum onbarnacle recruitment. Linear contrasts were used to test differencesamong specific combinations of treatments. We used ordinary leastsquares regression to test the magnitude and direction of therelationship among settlement, recruitment, algal cover, and limpetdensity. In most cases, raw data were not normally distributed butnormality was generally achieved using log-transformations.
3. Results
3.1. Effectiveness of treatments: limpet exclusion
Overall, as has been demonstrated before (Cubit, 1984; Paine,1992; Menge, 2000b; Freidenburg et al., 2007), copper-based paintbarriers were effective in excluding limpets in all years (Fig. 3A–H,Table S1a). Average densities of limpets in +limpet vs. − limpettreatments were 8.6±0.9 (mean±1SE) vs. 1.0±0.2 in 2001 (n=120vs. 120), 13.0±1.2 vs. 1.0±0.3 in 2003 (n=311 vs. 158), and 4.6±0.6 vs. 0.3±0.07 in 2008 (n=302 vs. 296), or differences of ∼9- to∼15-fold. The limpet exclusion treatment accounted for the majorityof the variance (Table S1a). However, the limpet response also variedthrough time by year, site and substratum type (Fig. 3A–H, S1, S2,Table S1a, S2). In 2001, these effects were expressed in the within-subjects effects of the month×site×surface interaction and themonth×surface× limpet interaction (Table S2, p=0.0004 andp=0.03, respectively). In 2003, the four-way within-subjects effectof month×site×surface×limpet indicated a highly context-depen-dent effect of all factors in combination (Table S2, p=0.01). In 2008,variable limpet response appeared in both month×site×limpets andmonth×surface×limpet interactions (Table S2, pb0.0001).
Despite these statistical interactions, limpet densities were higherin+limpet than in − limpet ROCK treatments in all years (Figs. S1,S2). The same general pattern held on STW (Figs. S1, S2), although in2001 and 2008 the differences occurred at only two of the four sites(BB and SH in 2001, FC and YB in 2008; Figs. S1, S2). In 2003, limpetdensities did not differ between the two+limpet treatments (markedplot without paint and paint control) except for SH (Fig. S1).
Limpet density in+limpet treatments averaged over all timeswithineach yearwas greater in ROCK plots thanon STWplates at FC and SH butnot different between these surfaces at BB, TK and YB. Limpet density in+limpet treatments differed by year at BB (2001=2003N2008 on bothSTWand ROCK), TK/YB (01b03N08 on STW, 01b03=08 on ROCK) andSH (01=03N08 on STW) but otherwise was similar among years(Fig. 3A–H, Table S1a, linear contrasts in 4-wayANOVA, pb0.05). Overalldensities across siteswere greater onROCK thanSTWin2001 (11.1±1.2vs. 6.2±1.2, n=60 each), but in 2003 limpet densities on ROCK weresimilar to thoseon STW(11.4±0.9 vs. 14.7±2.2; n=159, 152). In 2008,densities in+limpet treatments varied with surface (ROCK: 7.3±1.2,n=149; STW: 1.3±0.25, n=157; TRAV 0.8±0.19; n=138; and PVC0.4±0.13, n=154).
The variation among the artificial surfaces (STW=TRAVNPVC;linear contrasts, pb0.05) indicates that surface texture influencedgrazer abundance. Among years, the most striking differences were thenear absence of limpets at BB in 2008, a site that had high densities in2001 and 2003 and the large among-year differences at the southernsites (TK, YB and SH). Similar annual variation has been observedpreviously (Freidenburg et al., 2007). At the southern sites in particular,the limpets involved in these large among-year differences are mostlysmall individuals (b5 mm), which are likely new recruits, suggestingamong-year differences in recruitment can be substantial. The
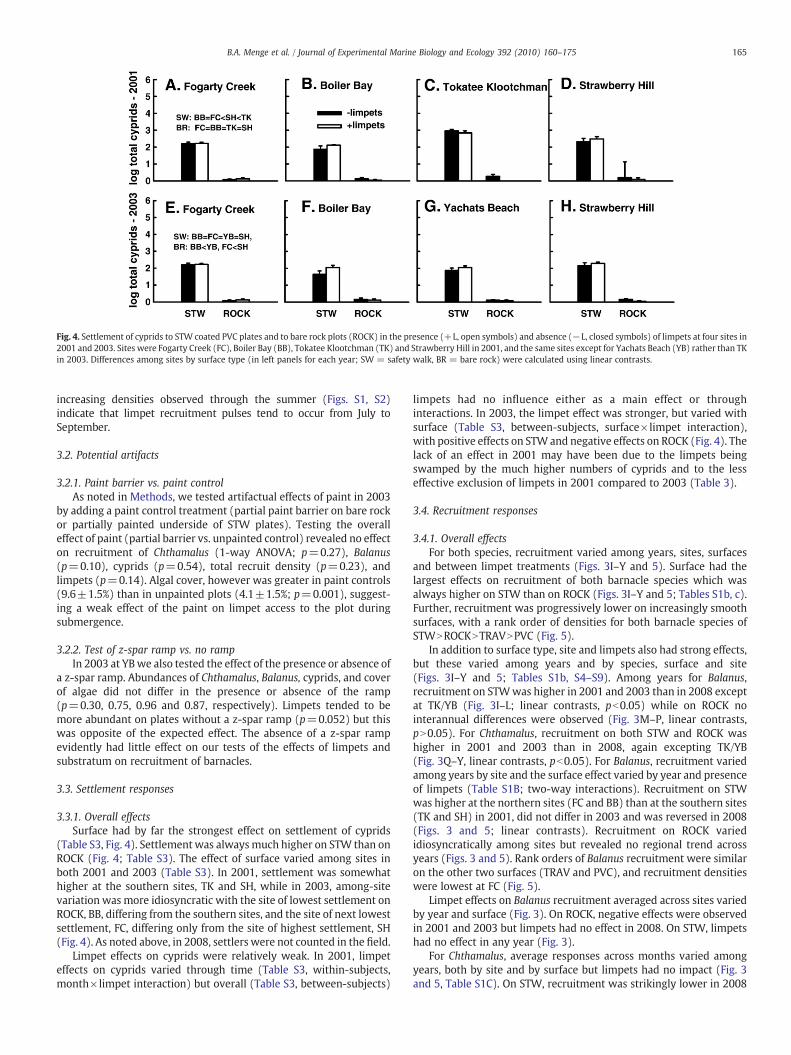
Fig. 4. Settlement of cyprids to STW coated PVC plates and to bare rock plots (ROCK) in the presence (+L, open symbols) and absence (−L, closed symbols) of limpets at four sites in2001 and 2003. Sites were Fogarty Creek (FC), Boiler Bay (BB), Tokatee Klootchman (TK) and Strawberry Hill in 2001, and the same sites except for Yachats Beach (YB) rather than TKin 2003. Differences among sites by surface type (in left panels for each year; SW = safety walk, BR = bare rock) were calculated using linear contrasts.
165B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
increasing densities observed through the summer (Figs. S1, S2)indicate that limpet recruitment pulses tend to occur from July toSeptember.
3.2. Potential artifacts
3.2.1. Paint barrier vs. paint controlAs noted in Methods, we tested artifactual effects of paint in 2003
by adding a paint control treatment (partial paint barrier on bare rockor partially painted underside of STW plates). Testing the overalleffect of paint (partial barrier vs. unpainted control) revealed no effecton recruitment of Chthamalus (1-way ANOVA; p=0.27), Balanus(p=0.10), cyprids (p=0.54), total recruit density (p=0.23), andlimpets (p=0.14). Algal cover, however was greater in paint controls(9.6±1.5%) than in unpainted plots (4.1±1.5%; p=0.001), suggest-ing a weak effect of the paint on limpet access to the plot duringsubmergence.
3.2.2. Test of z-spar ramp vs. no rampIn 2003 at YBwe also tested the effect of the presence or absence of
a z-spar ramp. Abundances of Chthamalus, Balanus, cyprids, and coverof algae did not differ in the presence or absence of the ramp(p=0.30, 0.75, 0.96 and 0.87, respectively). Limpets tended to bemore abundant on plates without a z-spar ramp (p=0.052) but thiswas opposite of the expected effect. The absence of a z-spar rampevidently had little effect on our tests of the effects of limpets andsubstratum on recruitment of barnacles.
3.3. Settlement responses
3.3.1. Overall effectsSurface had by far the strongest effect on settlement of cyprids
(Table S3, Fig. 4). Settlement was alwaysmuch higher on STW than onROCK (Fig. 4; Table S3). The effect of surface varied among sites inboth 2001 and 2003 (Table S3). In 2001, settlement was somewhathigher at the southern sites, TK and SH, while in 2003, among-sitevariation was more idiosyncratic with the site of lowest settlement onROCK, BB, differing from the southern sites, and the site of next lowestsettlement, FC, differing only from the site of highest settlement, SH(Fig. 4). As noted above, in 2008, settlers were not counted in the field.
Limpet effects on cyprids were relatively weak. In 2001, limpeteffects on cyprids varied through time (Table S3, within-subjects,month×limpet interaction) but overall (Table S3, between-subjects)
limpets had no influence either as a main effect or throughinteractions. In 2003, the limpet effect was stronger, but varied withsurface (Table S3, between-subjects, surface×limpet interaction),with positive effects on STW and negative effects on ROCK (Fig. 4). Thelack of an effect in 2001 may have been due to the limpets beingswamped by the much higher numbers of cyprids and to the lesseffective exclusion of limpets in 2001 compared to 2003 (Table 3).
3.4. Recruitment responses
3.4.1. Overall effectsFor both species, recruitment varied among years, sites, surfaces
and between limpet treatments (Figs. 3I–Y and 5). Surface had thelargest effects on recruitment of both barnacle species which wasalways higher on STW than on ROCK (Figs. 3I–Y and 5; Tables S1b, c).Further, recruitment was progressively lower on increasingly smoothsurfaces, with a rank order of densities for both barnacle species ofSTWNROCKNTRAVNPVC (Fig. 5).
In addition to surface type, site and limpets also had strong effects,but these varied among years and by species, surface and site(Figs. 3I–Y and 5; Tables S1b, S4–S9). Among years for Balanus,recruitment on STWwas higher in 2001 and 2003 than in 2008 exceptat TK/YB (Fig. 3I–L; linear contrasts, pb0.05) while on ROCK nointerannual differences were observed (Fig. 3M–P, linear contrasts,pN0.05). For Chthamalus, recruitment on both STW and ROCK washigher in 2001 and 2003 than in 2008, again excepting TK/YB(Fig. 3Q–Y, linear contrasts, pb0.05). For Balanus, recruitment variedamong years by site and the surface effect varied by year and presenceof limpets (Table S1B; two-way interactions). Recruitment on STWwas higher at the northern sites (FC and BB) than at the southern sites(TK and SH) in 2001, did not differ in 2003 and was reversed in 2008(Figs. 3 and 5; linear contrasts). Recruitment on ROCK variedidiosyncratically among sites but revealed no regional trend acrossyears (Figs. 3 and 5). Rank orders of Balanus recruitment were similaron the other two surfaces (TRAV and PVC), and recruitment densitieswere lowest at FC (Fig. 5).
Limpet effects on Balanus recruitment averaged across sites variedby year and surface (Fig. 3). On ROCK, negative effects were observedin 2001 and 2003 but limpets had no effect in 2008. On STW, limpetshad no effect in any year (Fig. 3).
For Chthamalus, average responses across months varied amongyears, both by site and by surface but limpets had no impact (Fig. 3and 5, Table S1C). On STW, recruitment was strikingly lower in 2008
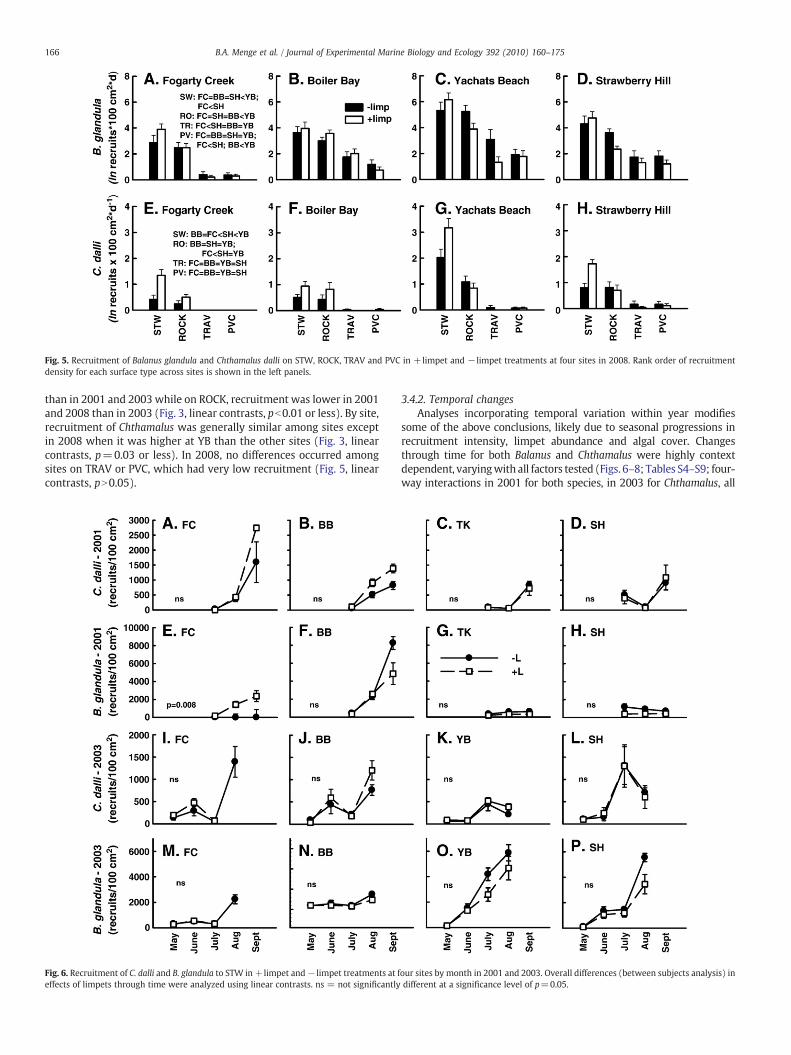
Fig. 5. Recruitment of Balanus glandula and Chthamalus dalli on STW, ROCK, TRAV and PVC in +limpet and − limpet treatments at four sites in 2008. Rank order of recruitmentdensity for each surface type across sites is shown in the left panels.
166 B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
than in 2001 and 2003 while on ROCK, recruitment was lower in 2001and 2008 than in 2003 (Fig. 3, linear contrasts, pb0.01 or less). By site,recruitment of Chthamalus was generally similar among sites exceptin 2008 when it was higher at YB than the other sites (Fig. 3, linearcontrasts, p=0.03 or less). In 2008, no differences occurred amongsites on TRAV or PVC, which had very low recruitment (Fig. 5, linearcontrasts, pN0.05).
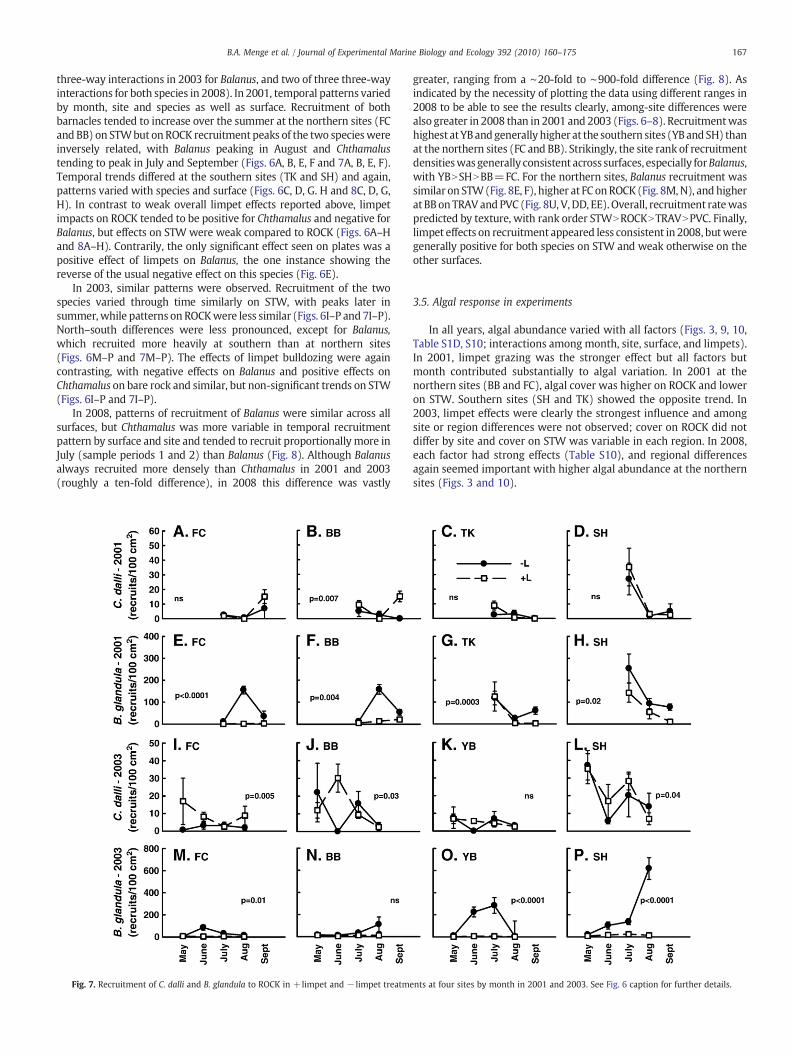
Fig. 6. Recruitment of C. dalli and B. glandula to STW in +limpet and− limpet treatments at feffects of limpets through time were analyzed using linear contrasts. ns = not significantly
3.4.2. Temporal changesAnalyses incorporating temporal variation within year modifies
some of the above conclusions, likely due to seasonal progressions inrecruitment intensity, limpet abundance and algal cover. Changesthrough time for both Balanus and Chthamalus were highly contextdependent, varyingwith all factors tested (Figs. 6–8; Tables S4–S9; four-way interactions in 2001 for both species, in 2003 for Chthamalus, all
our sites by month in 2001 and 2003. Overall differences (between subjects analysis) indifferent at a significance level of p=0.05.
167B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
three-way interactions in 2003 for Balanus, and two of three three-wayinteractions for both species in 2008). In 2001, temporal patterns variedby month, site and species as well as surface. Recruitment of bothbarnacles tended to increase over the summer at the northern sites (FCand BB) on STWbut on ROCK recruitment peaks of the two specieswereinversely related, with Balanus peaking in August and Chthamalustending to peak in July and September (Figs. 6A, B, E, F and 7A, B, E, F).Temporal trends differed at the southern sites (TK and SH) and again,patterns varied with species and surface (Figs. 6C, D, G. H and 8C, D, G,H). In contrast to weak overall limpet effects reported above, limpetimpacts on ROCK tended to be positive for Chthamalus and negative forBalanus, but effects on STW were weak compared to ROCK (Figs. 6A–Hand 8A–H). Contrarily, the only significant effect seen on plates was apositive effect of limpets on Balanus, the one instance showing thereverse of the usual negative effect on this species (Fig. 6E).
In 2003, similar patterns were observed. Recruitment of the twospecies varied through time similarly on STW, with peaks later insummer,while patterns on ROCKwere less similar (Figs. 6I–P and 7I–P).North–south differences were less pronounced, except for Balanus,which recruited more heavily at southern than at northern sites(Figs. 6M–P and 7M–P). The effects of limpet bulldozing were againcontrasting, with negative effects on Balanus and positive effects onChthamalus on bare rock and similar, but non-significant trends on STW(Figs. 6I–P and 7I–P).
In 2008, patterns of recruitment of Balanus were similar across allsurfaces, but Chthamalus was more variable in temporal recruitmentpattern by surface and site and tended to recruit proportionally more inJuly (sample periods 1 and 2) than Balanus (Fig. 8). Although Balanusalways recruited more densely than Chthamalus in 2001 and 2003(roughly a ten-fold difference), in 2008 this difference was vastly
Fig. 7. Recruitment of C. dalli and B. glandula to ROCK in +limpet and − limpet treatme
greater, ranging from a ∼20-fold to ∼900-fold difference (Fig. 8). Asindicated by the necessity of plotting the data using different ranges in2008 to be able to see the results clearly, among-site differences werealso greater in 2008 than in 2001 and 2003 (Figs. 6–8). Recruitmentwashighest atYBandgenerally higher at the southern sites (YBandSH) thanat the northern sites (FC and BB). Strikingly, the site rank of recruitmentdensitieswas generally consistent across surfaces, especially forBalanus,with YBNSHNBB=FC. For the northern sites, Balanus recruitment wassimilar on STW(Fig. 8E, F), higher at FC onROCK (Fig. 8M,N), and higherat BBon TRAV and PVC (Fig. 8U, V,DD, EE).Overall, recruitment ratewaspredicted by texture, with rank order STWNROCKNTRAVNPVC. Finally,limpet effects on recruitment appeared less consistent in 2008, butweregenerally positive for both species on STW and weak otherwise on theother surfaces.
3.5. Algal response in experiments
In all years, algal abundance varied with all factors (Figs. 3, 9, 10,Table S1D, S10; interactions among month, site, surface, and limpets).In 2001, limpet grazing was the stronger effect but all factors butmonth contributed substantially to algal variation. In 2001 at thenorthern sites (BB and FC), algal cover was higher on ROCK and loweron STW. Southern sites (SH and TK) showed the opposite trend. In2003, limpet effects were clearly the strongest influence and amongsite or region differences were not observed; cover on ROCK did notdiffer by site and cover on STW was variable in each region. In 2008,each factor had strong effects (Table S10), and regional differencesagain seemed important with higher algal abundance at the northernsites (Figs. 3 and 10).
nts at four sites by month in 2001 and 2003. See Fig. 6 caption for further details.
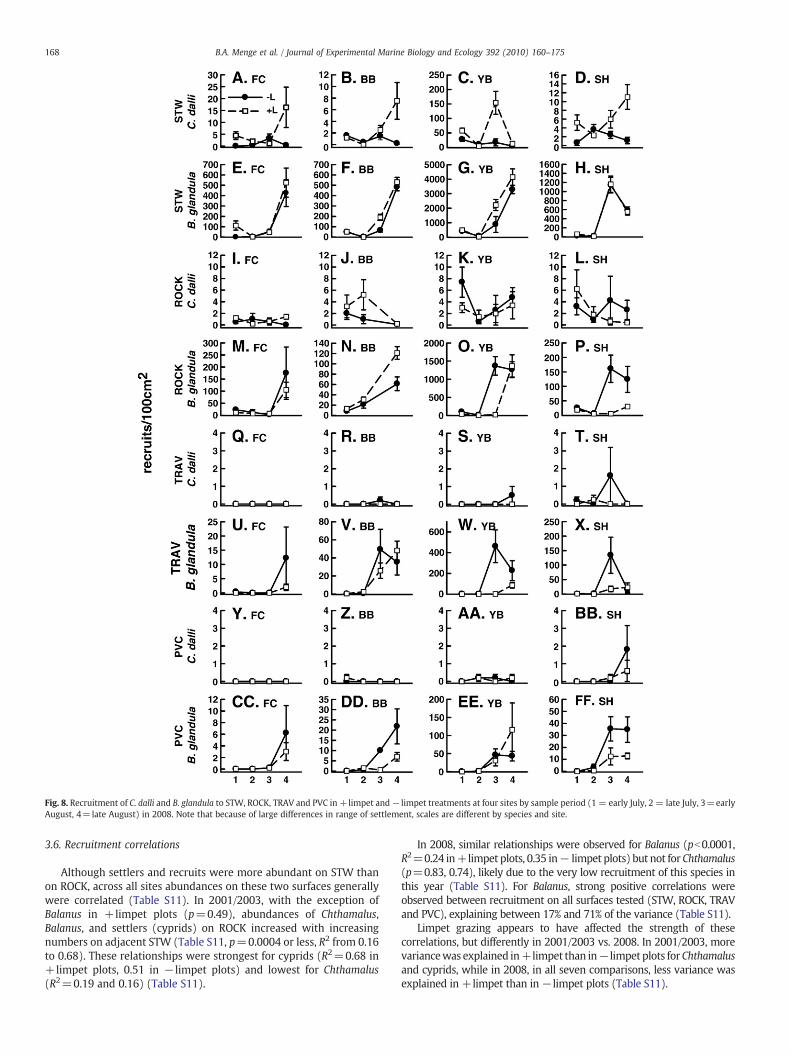
Fig. 8. Recruitment of C. dalli and B. glandula to STW, ROCK, TRAV and PVC in +limpet and− limpet treatments at four sites by sample period (1 = early July, 2 = late July, 3=earlyAugust, 4=late August) in 2008. Note that because of large differences in range of settlement, scales are different by species and site.
168 B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
3.6. Recruitment correlations
Although settlers and recruits were more abundant on STW thanon ROCK, across all sites abundances on these two surfaces generallywere correlated (Table S11). In 2001/2003, with the exception ofBalanus in +limpet plots (p=0.49), abundances of Chthamalus,Balanus, and settlers (cyprids) on ROCK increased with increasingnumbers on adjacent STW (Table S11, p=0.0004 or less, R2 from 0.16to 0.68). These relationships were strongest for cyprids (R2=0.68 in+limpet plots, 0.51 in − limpet plots) and lowest for Chthamalus(R2=0.19 and 0.16) (Table S11).
In 2008, similar relationships were observed for Balanus (pb0.0001,R2=0.24 in+limpet plots, 0.35 in− limpet plots) but not for Chthamalus(p=0.83, 0.74), likely due to the very low recruitment of this species inthis year (Table S11). For Balanus, strong positive correlations wereobserved between recruitment on all surfaces tested (STW, ROCK, TRAVand PVC), explaining between 17% and 71% of the variance (Table S11).
Limpet grazing appears to have affected the strength of thesecorrelations, but differently in 2001/2003 vs. 2008. In 2001/2003, morevariancewas explained in+limpet than in−limpet plots forChthamalusand cyprids, while in 2008, in all seven comparisons, less variance wasexplained in +limpet than in−limpet plots (Table S11).
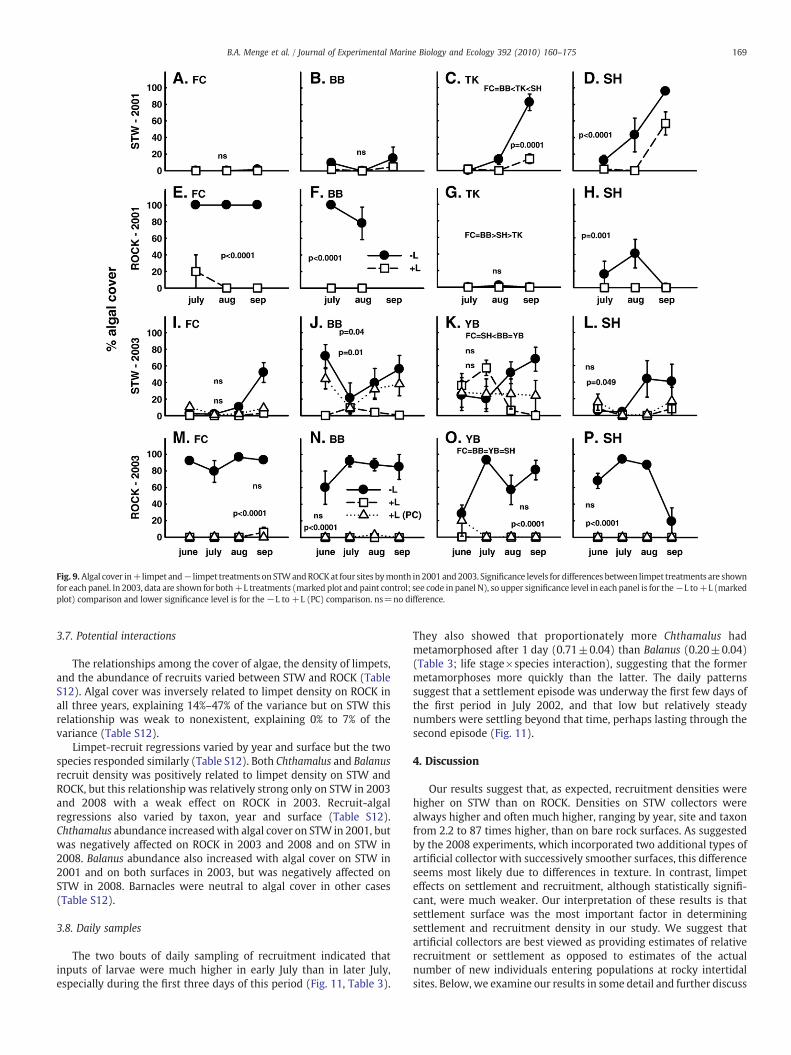
Fig. 9. Algal cover in+limpet and− limpet treatments on STWandROCKat four sites bymonth in 2001 and2003. Significance levels for differences between limpet treatments are shownfor each panel. In 2003, data are shown for both+L treatments (marked plot and paint control; see code in panel N), so upper significance level in each panel is for the−L to+L (markedplot) comparison and lower significance level is for the−L to +L (PC) comparison. ns=no difference.
169B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
3.7. Potential interactions
The relationships among the cover of algae, the density of limpets,and the abundance of recruits varied between STW and ROCK (TableS12). Algal cover was inversely related to limpet density on ROCK inall three years, explaining 14%–47% of the variance but on STW thisrelationship was weak to nonexistent, explaining 0% to 7% of thevariance (Table S12).
Limpet-recruit regressions varied by year and surface but the twospecies responded similarly (Table S12). Both Chthamalus and Balanusrecruit density was positively related to limpet density on STW andROCK, but this relationship was relatively strong only on STW in 2003and 2008 with a weak effect on ROCK in 2003. Recruit-algalregressions also varied by taxon, year and surface (Table S12).Chthamalus abundance increasedwith algal cover on STW in 2001, butwas negatively affected on ROCK in 2003 and 2008 and on STW in2008. Balanus abundance also increased with algal cover on STW in2001 and on both surfaces in 2003, but was negatively affected onSTW in 2008. Barnacles were neutral to algal cover in other cases(Table S12).
3.8. Daily samples
The two bouts of daily sampling of recruitment indicated thatinputs of larvae were much higher in early July than in later July,especially during the first three days of this period (Fig. 11, Table 3).
They also showed that proportionately more Chthamalus hadmetamorphosed after 1 day (0.71±0.04) than Balanus (0.20±0.04)(Table 3; life stage×species interaction), suggesting that the formermetamorphoses more quickly than the latter. The daily patternssuggest that a settlement episode was underway the first few days ofthe first period in July 2002, and that low but relatively steadynumbers were settling beyond that time, perhaps lasting through thesecond episode (Fig. 11).
4. Discussion
Our results suggest that, as expected, recruitment densities werehigher on STW than on ROCK. Densities on STW collectors werealways higher and often much higher, ranging by year, site and taxonfrom 2.2 to 87 times higher, than on bare rock surfaces. As suggestedby the 2008 experiments, which incorporated two additional types ofartificial collector with successively smoother surfaces, this differenceseems most likely due to differences in texture. In contrast, limpeteffects on settlement and recruitment, although statistically signifi-cant, were much weaker. Our interpretation of these results is thatsettlement surface was the most important factor in determiningsettlement and recruitment density in our study. We suggest thatartificial collectors are best viewed as providing estimates of relativerecruitment or settlement as opposed to estimates of the actualnumber of new individuals entering populations at rocky intertidalsites. Below, we examine our results in some detail and further discuss
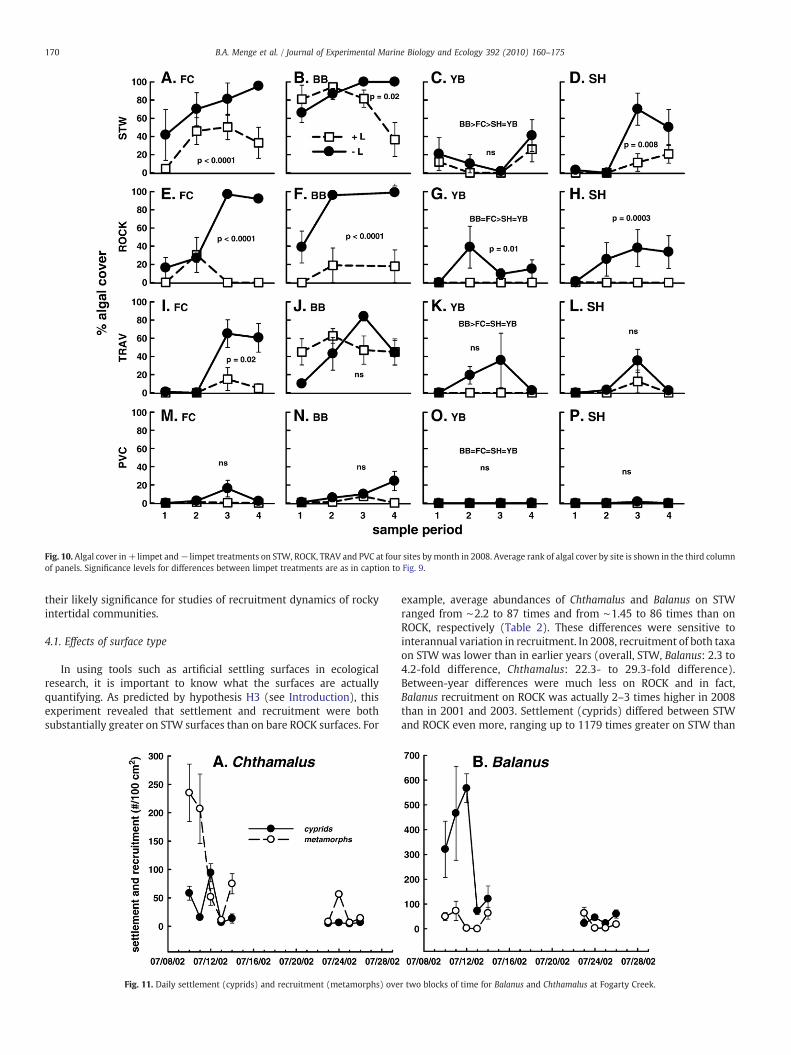
Fig. 10. Algal cover in+limpet and− limpet treatments on STW, ROCK, TRAV and PVC at four sites bymonth in 2008. Average rank of algal cover by site is shown in the third columnof panels. Significance levels for differences between limpet treatments are as in caption to Fig. 9.
170 B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
their likely significance for studies of recruitment dynamics of rockyintertidal communities.
4.1. Effects of surface type
In using tools such as artificial settling surfaces in ecologicalresearch, it is important to know what the surfaces are actuallyquantifying. As predicted by hypothesis H3 (see Introduction), thisexperiment revealed that settlement and recruitment were bothsubstantially greater on STW surfaces than on bare ROCK surfaces. For
Fig. 11. Daily settlement (cyprids) and recruitment (metamorphs) ove
example, average abundances of Chthamalus and Balanus on STWranged from ∼2.2 to 87 times and from ∼1.45 to 86 times than onROCK, respectively (Table 2). These differences were sensitive tointerannual variation in recruitment. In 2008, recruitment of both taxaon STW was lower than in earlier years (overall, STW, Balanus: 2.3 to4.2-fold difference, Chthamalus: 22.3- to 29.3-fold difference).Between-year differences were much less on ROCK and in fact,Balanus recruitment on ROCK was actually 2–3 times higher in 2008than in 2001 and 2003. Settlement (cyprids) differed between STWand ROCK even more, ranging up to 1179 times greater on STW than
r two blocks of time for Balanus and Chthamalus at Fogarty Creek.
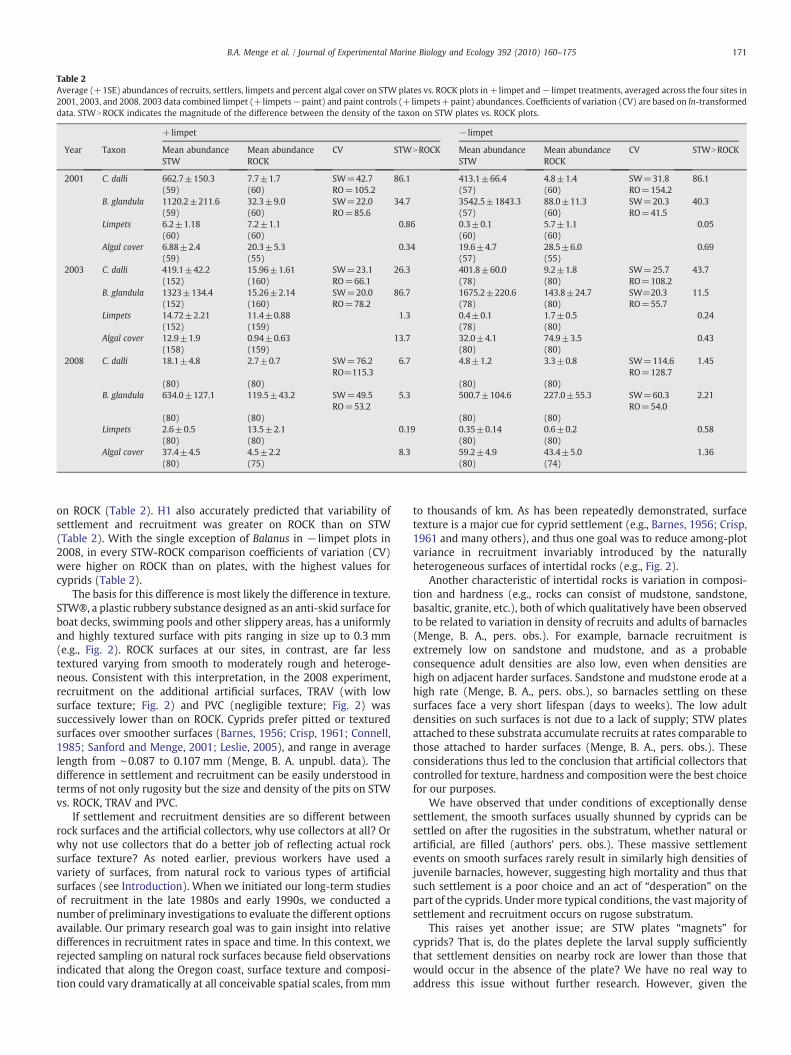
Table 2Average (+1SE) abundances of recruits, settlers, limpets and percent algal cover on STW plates vs. ROCK plots in +limpet and− limpet treatments, averaged across the four sites in2001, 2003, and 2008. 2003 data combined limpet (+limpets−paint) and paint controls (+limpets+paint) abundances. Coefficients of variation (CV) are based on ln-transformeddata. STWNROCK indicates the magnitude of the difference between the density of the taxon on STW plates vs. ROCK plots.
+limpet − limpet
Year Taxon Mean abundanceSTW
Mean abundanceROCK
CV STWNROCK Mean abundanceSTW
Mean abundanceROCK
CV STWNROCK
2001 C. dalli 662.7±150.3(59)
7.7±1.7(60)
SW=42.7RO=105.2
86.1 413.1±66.4(57)
4.8±1.4(60)
SW=31.8RO=154.2
86.1
B. glandula 1120.2±211.6(59)
32.3±9.0(60)
SW=22.0RO=85.6
34.7 3542.5±1843.3(57)
88.0±11.3(60)
SW=20.3RO=41.5
40.3
Limpets 6.2±1.18(60)
7.2±1.1(60)
0.86 0.3±0.1(60)
5.7±1.1(60)
0.05
Algal cover 6.88±2.4(59)
20.3±5.3(55)
0.34 19.6±4.7(57)
28.5±6.0(55)
0.69
2003 C. dalli 419.1±42.2(152)
15.96±1.61(160)
SW=23.1RO=66.1
26.3 401.8±60.0(78)
9.2±1.8(80)
SW=25.7RO=108.2
43.7
B. glandula 1323±134.4(152)
15.26±2.14(160)
SW=20.0RO=78.2
86.7 1675.2±220.6(78)
143.8±24.7(80)
SW=20.3RO=55.7
11.5
Limpets 14.72±2.21(152)
11.4±0.88(159)
1.3 0.4±0.1(78)
1.7±0.5(80)
0.24
Algal cover 12.9±1.9(158)
0.94±0.63(159)
13.7 32.0±4.1(80)
74.9±3.5(80)
0.43
2008 C. dalli 18.1±4.8 2.7±0.7 SW=76.2RO=115.3
6.7 4.8±1.2 3.3±0.8 SW=114.6RO=128.7
1.45
(80) (80) (80) (80)B. glandula 634.0±127.1 119.5±43.2 SW=49.5
RO=53.25.3 500.7±104.6 227.0±55.3 SW=60.3
RO=54.02.21
(80) (80) (80) (80)Limpets 2.6±0.5 13.5±2.1 0.19 0.35±0.14 0.6±0.2 0.58
(80) (80) (80) (80)Algal cover 37.4±4.5 4.5±2.2 8.3 59.2±4.9 43.4±5.0 1.36
(80) (75) (80) (74)
171B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
on ROCK (Table 2). H1 also accurately predicted that variability ofsettlement and recruitment was greater on ROCK than on STW(Table 2). With the single exception of Balanus in − limpet plots in2008, in every STW-ROCK comparison coefficients of variation (CV)were higher on ROCK than on plates, with the highest values forcyprids (Table 2).
The basis for this difference is most likely the difference in texture.STW®, a plastic rubbery substance designed as an anti-skid surface forboat decks, swimming pools and other slippery areas, has a uniformlyand highly textured surface with pits ranging in size up to 0.3 mm(e.g., Fig. 2). ROCK surfaces at our sites, in contrast, are far lesstextured varying from smooth to moderately rough and heteroge-neous. Consistent with this interpretation, in the 2008 experiment,recruitment on the additional artificial surfaces, TRAV (with lowsurface texture; Fig. 2) and PVC (negligible texture; Fig. 2) wassuccessively lower than on ROCK. Cyprids prefer pitted or texturedsurfaces over smoother surfaces (Barnes, 1956; Crisp, 1961; Connell,1985; Sanford and Menge, 2001; Leslie, 2005), and range in averagelength from ∼0.087 to 0.107 mm (Menge, B. A. unpubl. data). Thedifference in settlement and recruitment can be easily understood interms of not only rugosity but the size and density of the pits on STWvs. ROCK, TRAV and PVC.
If settlement and recruitment densities are so different betweenrock surfaces and the artificial collectors, why use collectors at all? Orwhy not use collectors that do a better job of reflecting actual rocksurface texture? As noted earlier, previous workers have used avariety of surfaces, from natural rock to various types of artificialsurfaces (see Introduction). When we initiated our long-term studiesof recruitment in the late 1980s and early 1990s, we conducted anumber of preliminary investigations to evaluate the different optionsavailable. Our primary research goal was to gain insight into relativedifferences in recruitment rates in space and time. In this context, werejected sampling on natural rock surfaces because field observationsindicated that along the Oregon coast, surface texture and composi-tion could vary dramatically at all conceivable spatial scales, frommm
to thousands of km. As has been repeatedly demonstrated, surfacetexture is a major cue for cyprid settlement (e.g., Barnes, 1956; Crisp,1961 and many others), and thus one goal was to reduce among-plotvariance in recruitment invariably introduced by the naturallyheterogeneous surfaces of intertidal rocks (e.g., Fig. 2).
Another characteristic of intertidal rocks is variation in composi-tion and hardness (e.g., rocks can consist of mudstone, sandstone,basaltic, granite, etc.), both of which qualitatively have been observedto be related to variation in density of recruits and adults of barnacles(Menge, B. A., pers. obs.). For example, barnacle recruitment isextremely low on sandstone and mudstone, and as a probableconsequence adult densities are also low, even when densities arehigh on adjacent harder surfaces. Sandstone and mudstone erode at ahigh rate (Menge, B. A., pers. obs.), so barnacles settling on thesesurfaces face a very short lifespan (days to weeks). The low adultdensities on such surfaces is not due to a lack of supply; STW platesattached to these substrata accumulate recruits at rates comparable tothose attached to harder surfaces (Menge, B. A., pers. obs.). Theseconsiderations thus led to the conclusion that artificial collectors thatcontrolled for texture, hardness and composition were the best choicefor our purposes.
We have observed that under conditions of exceptionally densesettlement, the smooth surfaces usually shunned by cyprids can besettled on after the rugosities in the substratum, whether natural orartificial, are filled (authors' pers. obs.). These massive settlementevents on smooth surfaces rarely result in similarly high densities ofjuvenile barnacles, however, suggesting high mortality and thus thatsuch settlement is a poor choice and an act of “desperation” on thepart of the cyprids. Undermore typical conditions, the vast majority ofsettlement and recruitment occurs on rugose substratum.
This raises yet another issue; are STW plates “magnets” forcyprids? That is, do the plates deplete the larval supply sufficientlythat settlement densities on nearby rock are lower than those thatwould occur in the absence of the plate? We have no real way toaddress this issue without further research. However, given the

172 B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
turbulence of intertidal habitats and our observation that cyprids willsettle on rock within cm of the collector plates, it seems unlikely thatthis possible behavior is the primary cause of the higher densities onSTW plates and lower densities on rock.
Shanks (2009) recently criticized the use of STW plates, citingresults of his research at a site on the southern Oregon that suggestedthat STW plates heated rapidly in the sun, killing settled cypridswithin hours. In his studies, he found much higher recruitment onceramic floor tiles than on STW plates. His studies were carried out in2007 from late May to the end of July.
Our results are inconsistent with Shanks' (2009) results. Forexample, we find much lower recruitment on artificial tiles (bothTRAV and PVC) than on STW. One difference that could account forour different results is that Shanks' studies were carried out in Maythrough July while ours were carried out from June/July throughAugust/September. The significance of this difference is that on theOregon coast, timing of low tides gradually falls earlier in the day ineach successive tide series through the summer. Thus, exposure ofsettlers and recruits to warmer mid-day temperatures graduallylessens as the summer progresses, presumably meaning lowerexposure to thermal stress. This could explain why numbers ofrecruits on STW earlier in summer were generally lower in ourexperiments than they were later in summer (e.g., Figs. 6–8).However, numbers of recruits on other surfaces (TRAV, PVC andROCK) were usually also lower earlier in summer (Figs. 6–8). Ifmortality explains the lower numbers on STW, and STW (and PVC)plates get hotter than ROCK or TRAV, then it seems likely thatconsistently higher numbers would be seen on the latter than on theformer surfaces. Further, we have also measured plate vs. rocktemperatures (B. Menge et al., unpublished data), and like Shanksfound that STW plates often reached temperatures in the 40 °C range.
It is possible that the differences between Shanks' (2009) resultsand ours may be influenced by other differences in spatial or temporalcoverage. Shanks' study was done in one summer at one site whileours spanned three separate years at four sites per summer. Further,examination of our long-term data set spanning six sites along thecentral and southern Oregon coast (including Cape Arago, which is afew km south of Shanks' Bastendorff Beach site) and 20 years (1989–2008) suggests other time/space differences that could be important(B. Menge et al., unpublished data). Average recruitment of Balanuswas relatively high from March through November, with some siteshaving peaks of recruitment in August or September and othershaving relatively uniform recruitment through this 9-month period.Importantly, variance among successive months could be very large,especially at Cape Arago. Thus, it is possible that a repeat of Shanks'(2009) study would have a different outcome. Without furtherinvestigation, it is impossible to determine the causes for the differentresults obtained by Shanks (2009) and the present paper.
What other alternatives to STW might we consider? Our sense isthat the best form of standard collector would be one that mimicked
Table 3Three way analysis of variance of variation in numbers of settlers by period (early vs.late July 2002), life history stage (cyprid vs. metamorph) and species (Chthamalus vs.Balanus). Analysis was on ln-transformed data. R2=0.48. Boldface values are significantat p=0.05.
Analysis of variance
Source SS df MS F p
Period 40.70672 1 40.70672 29.43461 b0.0001Life stage 5.51549 1 5.51549 3.98819 0.049Species 5.91069 1 5.91069 4.27396 0.04Period⁎ life stage 3.94577 1 3.94577 2.85314 0.09Period⁎species 1.08498 1 1.08498 0.78453 0.38Life stage⁎species 58.15874 1 58.15874 42.05399 b0.0001Period⁎ life stage⁎species 5.70853 1 5.70853 4.12778 0.045Error 138.29542 100 1.38295
natural rock surface texture of moderate rugosity. After our initialtrials, we explored the possibility of having unglazed ceramic tileswith appropriate surface texture manufactured for us, but high costmade this option unfeasible. We felt such tiles should be used onlyonce since the porous nature of the tile could lead to accumulation ofbiofilm residues that might not be removable and might influencesettlement in later uses (see Bourget, 1988; Keough and Raimondi,1995; Thompson et al., 1998). (In contrast, STW plates are reusable;the STW can be removed and a new layer applied to the non-porousPVC plastic plate.) Similar restrictions applied to other surfacesconsidered such as travertine, slate, granite, and other types of naturalrock. Nonetheless, given the extensive use of STW plates around theglobe, the apparent success (contra Shanks, 2009) of these collectorsin providing relative measures of recruitment in space and time, andthe lengthy time series that have been accumulated in PISCO andother programs (Broitman et al., 2008; Navarrete et al., 2008), we arehesitant to recommend switching to new surfaces at this point. If thegoals are to estimate absolute rates of input of recruits or the study isshort-term, more local-scale, and not aimed at making comparisonswith other investigations, however, then using natural rock surfacesor molds of natural rock surfaces would be a good choice.
4.2. Effects of limpets
The results of the limpet manipulations suggest that their effectsare more complicated than the simple, uniformly negative effectspredicted by H4 (see Table 2). Overall, limpet activity had strongeffects on settlement and recruitment of Balanus (2-way ANOVA withlimpets and surface as factors, p=0.008) but no effects on Chthamalus(2-way ANOVA, p=0.25). Further, the effect on Balanus varied amongyears and surface type. Balanus recruitment densities were lower in+limpet than in − limpet plots in 2001 and 2003 (suggesting anegative effect of limpets), but in 2008 recruit density differencessuggested a positive effect on STW and a negative effect on ROCK(Tables 2, S1B; surface×limpet interaction).
Our results suggest that limpet effects were contingent on densityof settlers or recruits. For example, on ROCK limpets negativelyaffected settlement of cyprids strongly in 2003 (p=0.005) andweakly in 2001 (p=0.049; linear contrasts in 3-way ANOVA),possibly due to swamping by much higher cyprid densities in 2001(Table 2). On STW, limpets had no effect on cyprids in 2001, again,possibly due to swamping, but in 2003 had a positive effect (Table 3).
As predicted by H5, this positive effect may be a consequence ofthe influence of rugosity on limpet crawling and grazing. Limpets,which range in length from about 2 to 15 mm, with most in the 5–10 mm range, are too large to fit in the pits and thus skim over the pitson STW as they crawl. While the radula (tongue-like feedingapparatus) of a 5 to 10 mm limpet is likely to be ∼0.12 to 0.25 mmin width (Kitting, 1980; assuming proportional scaling using a limpet24.2 mmwith a radula 0.6 mmwide), and thus could fit into the largerpits, it seems unlikely to be able to affect most cyprids that settle intopits. In contrast, due to growth and change in morphology,metamorphs extend further out of the pits in STW and may bemore vulnerable to bulldozing than are cyprids, at least during thefirst several days after metamorphosis. A direct effect of limpets onmicroalgae may also influence survival of cyprids by removing apotential barrier to settlement. Finally, a weaker effect of bulldozingon cyprids than on metamorphs may also reflect a shorter period ofvulnerability of cyprids (≤24 h) than metamorphs (several days)coupled with spatial and temporal variability in limpet grazing. Thus,the positive effect of limpets on STW may result from the removal ofmicroalgae from the ridges of the pits and a reduction in the ability ofthe limpets to bulldoze cyprids in pits. Conversely, the negative effecton ROCK seems likely due to the greater vulnerability of cypridsbecause of the much smoother texture of the rock surface and thusrelative lack of settlement micro-refuges. Further investigation of the

173B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
interaction between grazing activity, substratum and life history stageof barnacles seems warranted.
Effects of limpets similar to thoseon cypridswere also observedwithrecruits, but counter to H6, which predicts similar effects of substratumand bulldozing on both species, the taxonomic resolution reveals yetanother difference in how species responded to limpet bulldozing.Withthe exception of STW in 2008,when grazing had a positive effect (Fig. 3,3-way ANOVA on site, surface, and limpets, surface×limpets interac-tion, pb0.0001), limpets had either no or weakly positive effects onChthamalus andnegative effects onBalanus (Figs. 3 and5; Tables S4–S9).These effects varied in space (among sites) and time, likely as aconsequence of spatio-temporal variation in recruitment density,abundance of limpets and algal cover (Figs. 3, 6–10).
Themechanism underlying the relative resistance of Chthamalus tobulldozing compared to Balanus is unclear. Chthamalus recruits aresimilar in size to those of Balanus (0.932±0.01 vs. 0.95±0.05 mm,means of means of 9 measurements per species across 8 sample datesin 2002), suggesting resistance is not based on size. HoweverChthamalus do appear to metamorphose faster than Balanus. Twosets of daily samples taken in July 2002 indicate that after one day, themajority of Chthamalus were metamorphs while the majority ofBalanus were still cyprids (Fig. 11). Thus a possible mechanismunderlying resistance is that Chthamalus can quickly metamorphoseand thus attachmore firmly faster than can Balanus.We conclude thatthe effects of limpet bulldozing on settlement and recruitment arecontext-dependent, varying with the density of settlers and recruits(as indicated by the differences between 2001 and 2003 for cypridsand 2001, 2003 and 2008 for metamorphs), surface rugosity, anddifferences in the abilities of settlers or recruits to resist limpetdisturbance. Hence, the prediction of H5, that barnacle recruitmentwill be similarly affected by surface and limpet grazing is notsupported. Bulldozing reduced the density of Balanus but had littleeffect on the density of Chthamalus (excepting a positive effect in 2008on STW), most likely due to more rapid metamorphosis and thusfirmer attachment of the latter.
4.3. Relationship between densities on different surfaces
Hypothesis H1 predicts that even though magnitudes ofsettlement to different surfaces may differ, settlement andrecruitment on STW will be correlated to that on ROCK. Ouranalyses support this prediction. For both settlers (cyprids) andrecruits (abundances of Chthamalus and Balanus), numbers on STWwere usually positively related to numbers on ROCK, the excep-tions being Balanus in +limpet plots in 2001/2003 and Chthamalusin 2008 (Table S11). These positive relationships explainedbetween 16% and 68% of the variance. In 2008, correlationsamong the four different surfaces were also relatively strong(explaining 17% to 71% of the variance; Table S11).
These relationships seem to be relatively insensitive to countingmethod or location. In 2001 and 2003, counts on ROCK were made infive fields of view of known area using field microscopes and thenadjusting numbers to 100 cm2. In 2008, we used hand lenses to countrecruits in the whole area of ROCK, TRAV and PVC plates, and countedrecruits on STW in the lab using both a hand lens and a dissectingmicroscope (the method also used in 2001/2003). Despite thisvariation in method, the amount of variance explained between2001/2003 and 2008 on STW vs. ROCK tended to be similar for Balanus(again excepting the 2001/2003 +limpet plots)(Table S11). Further,STW counts using hand lenses and lab microscopes were stronglycorrelated (62% to 86% of the variance explained for Balanus;estimates for Chthamaluswere compromised by very low recruitmentof this species in 2008). On the other hand, hand lenses did notprovide the resolution needed to detect and identify cyprids, which iswhy these were not counted in 2008.
4.4. Spatial variation
Most analyses revealed strong differences among sites in recruit-ment, limpet densities, and algal cover (e.g., Fig. 3). The mostconsistent spatial trend, however, was the regional differencebetween the two northern sites on Cape Foulweather and the threesouthern sites on Cape Perpetua (see also Menge et al., 1997, 2004;Leslie et al., 2005; Barth et al., 2007; Freidenburg et al., 2007). Patternsat FC and BB were consistent for barnacle recruitment, algalabundance and limpet density. Similarly, patterns at SH tended tobe the same as those at TK and YB despite the switch from TK to YBbetween 2001 and 2003, and, overall, different from FC and BB, at leastin the three years covered by this study. For barnacles, however,longer-term data suggest that the regional differences observedduring these experiments is not consistent. Examination of longerterm (1989–2008) data indicated that in some years one cape hadhigher recruitment, in others, the other cape had higher recruitmentand in still other years, the capes did not differ (e.g., Leslie et al., 2005;Barth et al., 2007; Broitman et al., 2008).
5. Conclusions
We conclude that surface texture has the greatest effect onsettlement and recruitment of these two barnacles, an observationthat is consistent with classic studies in the United Kingdom andelsewhere (e.g., Barnes, 1956; Crisp, 1961; Chabot and Bourget, 1988).In addition, limpet grazing activity can influence settlement andrecruitment, but this effect is variable through time, with the densityof settlement and recruitment, with the specific barnacle taxa involved,and with surface rugosity. Important variation can also occur in space,both at the local site scale, and at the regional (cape) scale.
A primary goal in this study was to evaluate the extent to whichsettlement on artificial surfaces provided relatively comparableestimates across space and time that were independent of variationin surface texture, and reflected actual recruitment rates to rocksurfaces in the intertidal. Our conclusions are that recruitment onSTW plates does provide a method for allowing comparisons ofpotential recruitment across multiple spatial and temporal scales, butthat, not surprisingly, recruitment densities on STW substantiallyoverestimate actual recruitment to rock. The tradeoff between two ofthe goals in a recruitment sampling program remains: STW platesprovide the ability to sample many sites across large spatial scalesover long time periods but overestimate recruitment densities, whilebare rock counts provide more direct estimates of actual recruitdensities but, due to much longer “handling times” in the field, aremore limiting in spatio-temporal coverage. It is possible that with acorrection factor, recruitment on STW potentially could provide anapproximation of actual recruitment to bare rock surfaces and therebyallow for both precision in estimates and comparisons at multiplespatial and temporal scales. However, the fractions of recruits onROCK compared to STW varied by species and year (e.g., for 2001,2003, and 2008, respectively: Chthamalus, 1.1, 2.9, and 11.0% andBalanus, 2.3, 5.4, and 26.8%), suggesting the need to maintain parallelsampling schemes of both types of surface, at least at a few locations.
Acknowledgements
We thank the legions of undergraduate interns for their assistancein fabricating collectors, and deploying, replacing, and processing themany samples analyzed here. A. Iles, J. Tyburczy, S. Dudas, F. Chan, andG. Rilov and anonymous reviewers provided constructive criticism onthe manuscript. The research was funded by an endowment from theWayne and Gladys Valley Foundation, and grants from the A. W.Mellon Foundation, the David and Lucile Packard Foundation, and theGordon and Betty Moore Foundation. This is publication # 291 fromPISCO, the Partnership for Interdisciplinary Studies of the Coastal

174 B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
Ocean, a Large-Scale and Long-Term Ecological Consortium, fundedprincipally by the David and Lucile Packard and Gordon and BettyMoore Foundations. [SS]
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.jembe.2010.04.032.
References
Allison, G.W., 2004. The influence of species diversity and stress intensity oncommunity resistance and resilience. Ecological Monographs 74, 117–134.
Amman, A.J., 2004. SMURFs: standard monitoring units for the recruitment of temperatereef fishes. Journal of Experimental Marine Biology and Ecology 99, 135–154.
Barnes, H., 1956. Balanus balanoides L. in the Firth of Clyde: the development andannuary variation in the larval population and the causative factors. Journal ofAnimal Ecology 25, 72–84.
Barth, J.A., Menge, B.A., Lubchenco, J., Chan, F., Bane, J.M., Kirincich, A.R., Mcmanus, M.A.,Nielsen, K.J., Pierce, S.D., Washburn, L., 2007. Delayed upwelling alters nearshorecoastal ocean ecosystems in the northern California current. Proceedings of theNational Academy of Sciences of the United States of America 104, 3719–3724.
Barth, J.A., Wheeler, P.A., 2005. Introduction to special section: coastal advances in shelftransport. Journal of Geophysical Research 110, C10S01. doi10.1029/1205JC003124002005.
Benedetti-Cecchi, L., Cinelli, F., 1997. Confounding in field experiments: direct andindirect effects of artifacts due to the manipulation of limpets and macroalgae.Journal of Experimental Marine Biology and Ecology 209, 171–184.
Bertness, M.D., Crain, C.M., Silliman, B.R., Bazterrica, M.C., Reyna, M.V., Hidalgo, F.,Farina, J.K., 2006. The community structure of western Atlantic Patagonian rockyshores. Ecological Monographs 76, 439–460.
Bourget, E., 1988. Barnacle larval settlement: the perception of cues at different spatialscales. In: Chelazzi, G., Vannini, M. (Eds.), Behavioural Adaptations to Intertidal Life.NATO ASI Series. Plenum Press, pp. 155–173.
Broitman, B.R., Blanchette, C.A., Gaines, S.D., 2005. Recruitment of intertidalinvertebrates and oceanographic variability at Santa Cruz Island, California, USA.Limnology and Oceanography 50, 1473–1479.
Broitman, B.R., Blanchette, C.A., Menge, B.A., Lubchenco, J., Raimondi, P.A., Krenz, C., Foley,M.M., Lohse, D., Gaines, S.D., 2008. Spatial and temporal patterns of recruitment ofintertidal invertebrates along theU. S.West coast. EcologicalMonographs 78, 403–421.
Brown, J.H., 1995. Macroecology. University of Chicago Press.Caley, M.J., Carr, M.H., Hixon, M.A., Hughes, T.P., Jones, G.P., Menge, B.A., 1996.
Recruitment and the local dynamics of open marine populations. Annual Review ofEcology and Systematics 27, 477–500.
Chabot, R., Bourget, E., 1988. Influence of substratum heterogeneity and settledbarnacle density on the settlement of cypris larvae. Marine Biology 97, 45–56.
Coleman, R.A., Underwood, A.J., Benedetti-Cecchi, L., Aberg, P., Arenas, F., Arrontes, J.,Castro, J., Hartnoll, R.G., Jenkins, S.R., Paula, J., Santina, P.D., Hawkins, S.J., 2006. Acontinental scale evaluation of the role of limpet grazing on rocky shores. Oecologia147, 556–564.
Connell, J.H., 1961. Effects of competition, predation by Thais lapillus, and other factorson natural populations of the barnacle Balanus balanoides. Ecological Monographs31, 61–104.
Connell, J.H., 1985. The consequences of variation in initial settlement vs. postsettlement mortality in rocky intertidal communities. Journal of ExperimentalMarine Biology and Ecology 93, 11–45.
Connolly, S.R., Roughgarden, J., 1999. Theory of marine communities: competition,predation, and recruitment-dependent interaction strength. Ecological Monographs69, 277–296.
Crisp, D.J., 1961. Territorial behavior in barnacle settlement. Journal of ExperimentalBiology 38, 429–446.
Cubit, J., 1984. Herbivory and the seasonal abundance of algae on a high intertidal rockyshore. Ecology 65, 1904–1917.
Dayton, P.K., 1971. Competition, disturbance, and community organization: theprovision and subsequent utilization of space in a rocky intertidal community.Ecological Monographs 41, 351–389.
Farrell, T.M., Bracher, D., Roughgarden, J., 1991. Cross-shelf transport causesrecruitment to intertidal populations in central California. Limnology andOceanography 36, 279–288.
Freidenburg, T.L., Menge, B.A., Halpin, P.M., Webster, M.S., Sutton-Grier, A., 2007. Cross-scale variation in top-down and bottom-up control of algal abundance. Journal ofExperimental Marine Biology and Ecology 347, 8–29.
Gaines, S.D., Roughgarden, J., 1985. Larval settlement rate: a leading determinant ofstructure in an ecological community of the marine intertidal zone. Proceedings ofthe National Academy of Sciences of the United States of America 82, 3707–3711.
Guerry, A.D., 2008. Interactive effects of grazing and enrichment on diversity:conceptual implications of a rocky intertidal experiment. Oikos 117, 1185–1196.
Guerry, A.D., Menge, B.A., Dunmore, R., 2009. Effects of consumers and enrichment onabundance and diversity of benthic algae in a rocky intertidal community. Journalof Experimental Marine Biology and Ecology 369, 155–164.
Hawkins, S.J., 1983. Interactions of Patella and macroalgae with settling Semibalanusbalanoides (L.). Journal of Experimental Marine Biology and Ecology 71, 55–72.
Hawkins, S.J., Hartnoll, R.G., 1982. Settlement patterns of Semibalanus balanoides (L.) inthe Isle of Man (1977–1981). Journal of Experimental Marine Biology and Ecology62, 271–283.
Holyoak, M., Leibold, M.A., Holt, R.D., 2005. Metacommunities: Spatial Dynamics andEcological Communities. University of Chicago Press.
Jenkins, S.R., Aberg, P., Cervin, G., Coleman, R.A., Delany, J., Della Santina, P., Hawkins, S.J.,LaCroix, E.,Myers, A.A., Lindegarth,M., Power, A.-M., Roberts,M.F., Hartnoll, R.G., 2000.Spatial and temporal variation in settlement and recruitmentof the intertidal barnacleSemibalanus balanoides (L.)(Crustacea: Cirripedia) over a European scale. Journal ofExperimental Marine Biology and Ecology 243, 209–225.
Jenkins, S.R., Murua, J., Borrows, M.T., 2008. Temporal changes in the strength ofdensity-dependent mortality and growth in intertidal barnacles. Journal of AnimalEcology 77, 573–584.
Keough, M.J., Raimondi, P.T., 1995. Responses of settling invertebrate larvae to bio-organic films: effects of different types of films. Journal of Experimental MarineBiology and Ecology 185, 235–253.
Kitting, C.L., 1980. Herbivore–plant interactions of individual limpets maintaining amixed diet of intertidal marine algae. Ecological Monographs 50, 527–550.
Kosro, P.M., 2005. On the spatial structure of coastal circulation off Newport, Oregon,during spring and summer 2001 in a region of varying shelf width. Journal ofGeophysical Research 110, C10S06. doi10.1029/2004JC002769.
Leslie, H.M., 2005. Positive interspecific effects trump negative effects in high-densitybarnacle aggregations. Ecology 86, 2716–2725.
Leslie, H.M., Breck, E.N., Chan, F., Lubchenco, J., Menge, B.A., 2005. Hotspots of barnaclereproduction associated with nearshore ocean conditions. Proceedings of theNational Academy of Sciences of the United States of America 102, 10534–10539.
Levin, S.A., 1992. The problem of pattern and scale in ecology. Ecology 73, 1943–1967.Lewis, J.R., 1964. The Ecology of Rocky Shores, First ed. The English Universities Press
Ltd.Menge, B.A., 1976. Organization of the New England rocky intertidal community: role of
predation, competition and environmental heterogeneity. Ecological Monographs46, 355–393.
Menge, B.A., 1991. Relative importance of recruitment and other causes of variation onrocky intertidal community structure. Journal of Experimental Marine Biology andEcology 146, 69–100.
Menge, B.A., 1992. Community regulation: under what conditions are bottom-upfactors important on rocky shores? Ecology 73, 755–765.
Menge, B.A., 2000a. Recruitment vs. post-recruitment processes as determinants ofbarnacle population abundance on Oregon rocky shores. Ecological Monographs70, 265–288.
Menge, B.A., 2000b. Top-down and bottom-up community regulation in marine rockyintertidal habitats. Journal of ExperimentalMarine Biology and Ecology 250, 257–289.
Menge, B.A., Berlow, E.L., Blanchette, C.A., Navarrete, S.A., Yamada, S.B., 1994. Thekeystone species concept: variation in interaction strength in a rocky intertidalhabitat. Ecological Monographs 64, 249–286.
Menge, B.A., Daley, B.A., Wheeler, P.A., Dahlhoff, E., Sanford, E., Strub, P.T., 1997.Benthic–pelagic links and rocky intertidal communities: bottom-up effects on top-down control? Proceedings of the National Academy of Sciences of the UnitedStates of America 94, 14530–14535.
Menge, B.A., Daley, B.A., Lubchenco, J., Sanford, E., Dahlhoff, E., Halpin, P.M., Hudson, G.,Burnaford, J.L., 1999. Top-down and bottom-up regulation of New Zealand rockyintertidal communities. Ecological Monographs 69, 297–330.
Menge, B.A., Lubchenco, J., Bracken, M.E.S., Chan, F., Foley, M.M., Freidenburg, T.L.,Gaines, S.D., Hudson, G., Krenz, C., Leslie, H., Menge, D.N.L., Russell, R., Webster, M.S.,2003. Coastal oceanography sets the pace of rocky intertidal community dynamics.Proceedings of the National Academy of Sciences of the United States of America 100,12229–12234.
Menge, B.A., Blanchette, C.A., Raimondi, P., Freidenburg, T.L., Gaines, S.D., Lubchenco, J.,Lohse, D., Hudson, G., Foley, M.M., Pamplin, J., 2004. Species interaction strength:testing model predictions along an upwelling gradient. Ecological Monographs 74,663–684.
Morton, J.E., 2004. Seashore Ecology of New Zealand and the Pacific. David Bateman, Ltd.Navarrete, S.A., Broitman, B.R., Wieters, E.A., Castilla, J.C., 2005. Scales of benthic–
pelagic coupling and the intensity of species interactions: from recruitmentlimitation to top down control. Proceedings of the National Academy of Sciences ofthe United States of America 102, 18046–18051.
Navarrete, S.A., Broitman, B.R., Menge, B.A., 2008. Interhemispheric comparison ofrecruitment to intertidal communities: pattern persistence and scales of variation.Ecology 89, 1308–1322.
Noda, T., 2004. Large-scale variability in recruitment of the barnacle Semibalanuscariosus: it cause and effects on the population density and predator. MarineEcology Progress Series 278, 241–252.
O'Riordan, R.M., Arenas, F., Arrontes, J., Castro, J., Cruz, J.J., Delany, J., Martinez, B.,Fernandez, C., Hawkins, S.J., McGrath, D., Myers, A.A., Oliveros, J., Pannacciulli, F.G.,Power, A.-M., Relini, G., Rico, J.M., Silva, T., 2004. Spatial variation in the recruitmentof the intertidal barnacles Chthamalusmontagui Southward and Chthamalus stellatus(Poli) (Crustacea: Cirripedia) over a European scale. Journal of ExperimentalMarineBiology and Ecology 304, 243–264.
Paine, R.T., 1992. Food-web analysis through field measurement of per capitainteraction strength. Nature 355, 73–75.
Pineda, J., 1994. Spatial and temporal patterns in barnacle settlement rate along asouthern California rocky shore. Marine Ecology Progress Series 107, 125–138.
Pomerat, C.M.,Weiss, C.M., 1946. The influence of texture and composition of surface onthe attachment of sedentary marine organisms. Biological Bulletin 91, 57–65.
Raimondi, P.T., 1988. Settlement cues and determination of the vertical limit of anintertidal barnacle. Ecology 69, 400–407.

175B.A. Menge et al. / Journal of Experimental Marine Biology and Ecology 392 (2010) 160–175
Raimondi, P.T., 1990. Patterns, mechanisms, and consequences of variability insettlement and recruitment of an intertidal barnacle. Ecological Monographs 60,283–309.
Sagarin, R., Gaines, S.D., 2002. Geographical abundance distributions of coastalinvertebrates: using one-dimensional ranges to test biogeographic hypotheses.Journal of Biogeography 29, 985–997.
Sanford, E., Menge, B.A., 2001. Spatial and temporal variation in barnacle growth in acoastal upwelling system. Marine Ecology Progress Series 209, 143–157.
SAS Institute Inc, 2008. JMP 8 Statistics and Graphics Guide. SAS Institute Inc., Cary, NC.Schoch, G.C., Menge, B.A., Allison, G.W., Kavanaugh, M., Wood, S., 2006. Fifteen degrees
of separation: latitudinal gradients of rocky intertidal biota along the CaliforniaCurrent. Limnology and Oceanography 51, 2564–2585.
Shanks, A.L., 1974. Barnacle settlement versus recruitment as indicators of larval delivery.I. Effects of post-settlement mortality and recruit density. Marine Ecology ProgressSeries 385, 205–216.
Shkedy, Y., Roughgarden, J., 1997. Barnacle recruitment and population dynamicspredicted from coastal upwelling. Oikos 80, 487–498.
Stephenson, T.A., Stephenson, A., 1972. Life Between Tidemarks on Rocky Shores. W. H.Freeman and Co.
Sutherland, J.P., 1974. Multiple stable points in natural communities. American Naturalist108, 859–873.
Thompson, R.C., Norton, T.A., Hawkins, S.J., 1998. The influence of epilithic microbialfilms on the settlement of Semibalanus balanoides cyprids— a comparison betweenlaboratory and field experiments. Hydrobiologia 375 (376), 203–216.
Wethey, D.S., 1984. Spatial pattern in barnacle settlement: day to day changes duringthe settlement season. Journal of the Marine Biological Association (UnitedKingdom) 64, 687–698.