subdivisions and connectional networks of the lateral prefrontal cortex in the macaque monkey
TRANSCRIPT

Subdivisions and Connectional Networks of theLateral Prefrontal Cortex in the Macaque Monkey
Kadharbatcha S. Saleem,1,2* Brad Miller,1 and Joseph L. Price1
1Department of Anatomy and Neurobiology, Washington University School of Medicine, St. Louis, Missouri 631102Laboratory of Neuropsychology, National Institute of Mental Health, National Institute of Health, Bethesda, Maryland 20892
ABSTRACTNeuroanatomical studies have long indicated that corti-
cocortical connections are organized in networks that
relate distinct sets of areas. Such networks have been
emphasized by development of functional imaging
methods for correlating activity across the cortex. Previ-
ously, two networks were recognized in the orbitome-
dial prefrontal cortex, the “orbital” and “medial”
networks (OPFC and MPFC, respectively). In this study,
three additional networks are proposed for the lateral
prefrontal cortex: 1) a ventrolateral network (VLPFC) in
and ventral to the principal sulcus; 2) a dorsal network
(DPFC) in and dorsal to the principal sulcus and in the
frontal pole; 3) a caudolateral network (CLPFC) in and
rostral to the arcuate sulcus and the caudal principal
sulcus. The connections of the first two networks are
described here. Areas in each network are connected
primarily with other areas in the same network, with
overlaps around the principal sulcus. The VLPFC and
DPFC are also connected with the OPFC and MPFC,
respectively. Outside the prefrontal cortex, the VLPFC
connects with specific areas related to somatic/visceral
sensation and vision, in the frontoparietal operculum,
insula, ventral bank/fundus of the superior temporal
sulcus, inferior temporal gyrus, and inferior parietal cor-
tex. In contrast, the DPFC connects with the rostral
superior temporal gyrus, dorsal bank of the superior
temporal sulcus, parahippocampal cortex, and posterior
cingulate and retrosplenial cortex. Area 45a, in caudal
VLPFC, is unique, having connections with all the net-
works. Its extrinsic connections resemble those of the
DPFC. In addition, it has connections with both auditory
belt/parabelt areas, and visual related areas. J. Comp.
Neurol. 522:1641–1690, 2014.
VC 2013 Wiley Periodicals, Inc.
INDEXING TERMS: LPFC; VLPFC; DPFC; CLPFC; OMPFC; OPFC; MPFC; superior temporal gyrus; parietal cortex; infe-
rior temporal cortex; superior temporal sulcus; insula; connections
Although the prefrontal cortex (PFC) in primates,
defined as the region rostral to the “frontal” motor
areas, does not have obvious sensory or motor roles, it
has been associated with many functions. Perhaps the
most common is “executive function,” but this vague
and ultimately unsatisfactory designation subsumes a
number of subordinate functions. Attention, spatial and
nonspatial working memory, assessment of food stimuli,
reward-guided behavior, anticipation of reward or aver-
sion, emotional behavior, and mood have all been
attributed to the PFC (Mishkin and Manning, 1978;
Levy and Goldman-Rakic, 2000; Miller, 2000; €Ong€ur
and Price, 2000; Rolls, 2000; Rushworth et al., 2005;
Petrides, 2005; Meyer et al., 2011; Noonan et al.,
2011; Rudebeck and Murray, 2011b; Price and Drevets,
2012; Passingham and Wise, 2012). Because the PFC
as a whole is large and heterogeneous, it is likely that
these and other functions are not represented evenly
across the many architectonic areas that make it up.
Instead, it may be presumed that the PFC is divided
into several networks that have distinct roles.
The development of methods that are purported to
demonstrate “functional connections” with MRI (fcMRI)
has led to the description of several corticocortical net-
works. These link widespread cortical areas in several
parts of the cortex and are presumed to underlie
Grant sponsor: National Institutes of Health; Grant number:MH70941; Grant sponsor: McDonnell Center for Higher Brain Function;Grant sponsor: National Institute of Mental Health Intramural ResearchProgram.
*CORRESPONDENCE TO: Kadharbatcha Saleem, PhD, Laboratory ofNeuropsychology, National Institute of Mental Health, 49 Convent Drive,Bldg. 49, 1B80 MSc. 4415, Bethesda, MD 20892.E-mail: [email protected]
Received June 12, 2013; Revised October 31, 2013;Accepted October 31, 2013.DOI 10.1002/cne.23498Published online November 9, 2013 in Wiley Online Library(wileyonlinelibrary.com)VC 2013 Wiley Periodicals, Inc.
The Journal of Comparative Neurology | Research in Systems Neuroscience 522:1641–1690 (2014) 1641
RESEARCH ARTICLE

Abbreviations
1–2 somatosensory areas 1 and 23a/b somatosensory areas 3a and 3b3v third ventricle4 primary motor cortex (or F1)5 superior parietal lobule area (or PE / PEa / PEc)7a caudal inferior parietal lobule area (or Opt / PG)7b rostral inferior parietal lobule area (or PFG / PF)7m medial parietal area (or PGm)7op parietal opercular area8Ad periarcuate area (or frontal eye field), dorsal subdivison8Av periarcuate area (or frontal eye field), ventral subdivison8Bd/m dorsal prefrontal area9d/m dorsal prefrontal area10mr frontal pole area10mc medial prefrontal area10o frontal pole area11l orbital prefrontal area11m orbital prefrontal area12l ventrolateral prefrontal area12m orbital prefrontal area12o orbital prefrontal area12r ventrolateral prefrontal area13a,b orbital prefrontal area13l/m orbital prefrontal area14r/c orbitomedial prefrontal area (gyrus rectus)23 posterior cingulate cortexv23 or 23v subregion of posterior cingulate cortex24a/b/c subregions of anterior cingulate cortex24a0/b0/c0 subregions of anterior cingulate cortex25 medial prefrontal area29/30 retrosplenial cortex31 posterior cingulate area32 medial prefrontal area35 area 35 of the perirhinal cortex36c area 36 of the perirhinal cortex, caudal subregion36p area 36 of the perirhinal cortex, temporal-polar subregion36r area 36 of the perirhinal cortex, rostral subregion44 ventrolateral prefrontal area45a/b ventrolateral prefrontal area46d dorsal prefrontal area46v ventrolateral prefrontal area46f ventrolateral prefrontal area46d/v caudal caudolateral prefrontal areaAAA anterior amygdaloid areaAB accessory basal nucleus of amygdalaac anterior commissureAI auditory area I, core region of the auditory cortexAIP anterior intraparietal areaAL anterior lateral, belt region of the auditory cortexamts anterior middle temporal sulcusamy amygdalaArh arcuate hypothalamic nucleusasl arcuate sulcus lower limbasu arcuate sulcus upper limbB basal nucleus of amygdalacas calcarine sulcusCC corpus callosumcd caudate nucleuscis cingulate sulcusCL caudal lateral, belt region of the auditory cortexcla claustrumclc central latocellular nucleusCLPFC caudolateral prefrontal (network) cortexCM caudomedial, belt region of the auditory cortexCOa anterior cortical nucleusCP cerebral peduncleCPB caudal parabelt region of the auditory cortexcs central sulcusDP dorsal prelunate areaDPFC dorsal prefrontal (network) cortexF1 agranular frontal area F1 (or 4)F2 agranular frontal area F2F3 agranular frontal area F3 (or SMA)F4 agranular frontal area F4F5 agranular frontal area F5F6 agranular frontal area F6 (or preSMA)F7 agranular frontal area F7FST floor of superior temporal areaG gustatory cortexGP globus pallidusHC hippocampusIa agranular insula
Iac caudal agranular insular areaIai intermediate agranular insula areaIal lateral agranular insula areaIam medial agranular insula areaIapl posterolateral agranular insula areaIapm posteromedial agranular insula areaId dysgranular insula,Ig granular insulaios inferior occipital sulcusipd infraprincipal dimpleips intraparietal sulcusL lateral nucleus of amygdalaLd lateral nucleus of amygdala, dorsal subdivision,LGN lateral geniculate nucleusLIP lateral intraparietal areaLIPd lateral intraparietal area, dorsal subdivisionLIPv lateral intraparietal area, ventral subdivisionLP lateral posterior nucleusls lateral sulcuslus lunate sulcusLv lateral nucleus of amygdala, ventral subdivision,lv lateral ventricleMD medial dorsal nucleusMDpc mediodorsal nucleus, parvicellular divisionML middle lateral, belt region of the auditory cortexMPFC medial prefrontal (network) cortexMST medial superior temporal areaMT middle temporal area,NA nucleus accumbensNBM nucleus basalis of MeynertOPf frontal opercular areaOPFC orbital prefrontal (network) cortexOpt caudal inferior parietal lobule area (or 7a)ot optic tractots occipitotemporal sulcusP pulvinarPa paraventricular nucleusPAC periamygdaloid cortexPcn paracentral nucleusPE superior parietal lobule area (or 5)PEa superior parietal lobule area (or 5)PEc superior parietal lobule area (or 5)PF rostral inferior parietal lobule area (or 7b)Pf parafascicular nucleusPFG rostral inferior parietal lobule area (or 7b)PG caudal inferior parietal lobule area (or 7a)PGm medial parietal area (or 7m)PI inferior pulvinarPi parainsular areaPL lateral pulvinarPM medial pulvinarpmts posterior middle temporal sulcusPO parieto-occipital area (or V6/V6Av)pos parieto-occipital sulcusPrCO precentral opercular areaPreSMA presupplementary motor areaps principal sulcuspu putamenPV parietal ventral areaR rostral, core region of the auditory cortexRe reunions nucleusRM rostromedial, belt region of the auditory cortexRPB rostral parabelt region of the auditory cortexrs rhinal sulcusRT rostrotemporal, core region of the auditory cortexRTL lateral rostrotemporal, belt region of the auditory cortexRTM medial rostrotemporal, belt region of the auditory cortexRTp rostrotemporal (“p” refers to polar)sas spur of the arcuate sulcusSII secondary somatosensory areaSMA supplementary motor areaSTGr rostral superior temporal gyrussts superior temporal sulcusSTSd dorsal bank of the superior temporal sulcusSTSf fundus of the superior temporal sulcusSTSv ventral bank of the superior temporal sulcusTEad dorsal subregion of anterior TETEav ventral subregion of anterior TETEOd area TEO, dorsal partTEOv area TEO, ventral partTEpd dorsal subregion of posterior TETEpv ventral subregion of posterior TE
K.S. Saleem et al.
1642 The Journal of Comparative Neurology |Research in Systems Neuroscience

specific functions (see, e.g., Power et al., 2011).
Although the relationship between these functional net-
works and anatomical connections is still uncertain,
such networks, and previous neuroanatomical analyses
of corticocortical connections (see, e.g., Pandya and
Kuypers, 1969; Jones and Powell, 1970), imply that the
cortex is a mosaic of interwoven circuits that connect
restricted cortical areas. Any region as large as the PFC
can therefore be expected to be heterogeneous in
function.
For the ventromedial or orbital and medial parts of
the PFC (OMPFC), previous neuroanatomical studies
have defined two systems or “networks.” These were
based initially on distinctions in intrinsic connections
within the OMPFC (Carmichael and Price, 1996), but
the two systems also have distinct connections with
other brain areas. The so-called orbital (OPFC) network
is connected to several sensory-related cortical areas,
such as the olfactory and taste cortices, visual-related
areas in the inferior temporal gyrus, and somatic
sensory-related areas in the insula and frontoparietal
operculum (Carmichael and Price, 1995; €Ong€ur and
Price, 2000; Saleem et al., 2008b). This network
appears to integrate multimodal sensory information,
especially related to food, and to be involved in assess-
ment of stimuli in terms of reward and/or aversion and
their relative “value” in relation to other stimuli (€Ong€ur
and Price, 2000; Padoa-Schioppa and Assad, 2006;
Rudebeck and Murray, 2011a,b). The “medial” (MPFC)
network has few sensory inputs but has substantial out-
puts to visceral control areas in the hypothalamus and
brainstem (An et al., 1998; €Ong€ur et al., 1998). It is
also interconnected with a well-defined cortical circuit
involving the anterior and posterior cingulate cortex, an
area in the rostral superior temporal gyrus and dorsal
bank of the superior temporal sulcus, and the posterior
parahippocampal cortex (Saleem et al., 2008b) as well
as limbic structures, including the amygdala, hippocam-
pus, and entorhinal cortex. This network is related to
modulation of visceral function in relation to emotion
and is critically involved in mood disorders (Drevets
et al., 1997, 1998; Mayberg et al., 1999, 2005; Price
and Drevets, 2012).
In the lateral part of the PFC (LPFC), there are also
indications that relatively restricted regions have distinct
functions. Thus, in monkeys, the “ventrolateral prefrontal
cortex” (VLPFC), below the principal sulcus, has been
related to the assessment and convergence of sensory
information, including the processing of faces, visual
association, and integration of species-specific face and
vocal stimuli, and has also been thought to be involved in
object working memory and memory retrieval (Wilson
et al., 1993; O Scalaidhe et al., 1997; Asaad et al., 1998;
Levy and Goldman-Rakic, 2000; Passingham et al., 2000;
Cadoret and Petrides, 2007; Tsao et al., 2008b; Roman-
ski, 2012). The “dorsal prefrontal cortex” (DPFC) above
the principal sulcus has been thought to be involved in
self-centered functions, including behaviors that depend
on the previous actions or feelings of the individual (Pet-
rides, 2005). Finally, the “caudolateral prefrontal cortex”
(CLPFC), the region around the arcuate sulcus, and parts
of the caudal principal sulcus have been related to eye
movements (frontal eye field), attention, shape selectivity,
and working memory for spatial position (Bruce and Gold-
berg, 1985; Bruce et al., 1985; Gamlin and Yoon, 2000;
Tehovnik et al., 2000; Schall 2004; Peng et al., 2008;
Amiez and Petrides, 2009; Monosov and Thompson,
2009; Zhou and Desimone, 2011).
Several anatomical studies have documented the
connections of the LPFC with other cortical areas
(Kuypers et al., 1965; Pandya and Kuypers, 1969; Jones
and Powell, 1970; Chavis and Pandya, 1976; Barbas
and Mesulam, 1981, 1985; Kawamura and Naito, 1984;
Shiwa, 1987; Barbas, 1988; Barbas and Pandya, 1989;
Seltzer and Pandya, 1989; Ungerleider et al., 1989;
Webster et al., 1994; Romanski et al. 1999a,b). Pet-
rides and Pandya (1999, 2002, 2006, 2007), in particu-
lar, have approached this topic with several studies of
the efferent (or afferent) connections of the dorsome-
dial, ventrolateral, caudal, and rostral parts of the
LPFC. Based on these studies, Pandya and his col-
leagues have proposed that the prefrontal cortex can
be divided into two architectonic “trends,” separated
along the principal sulcus (e.g., Yeterian et al., 2012).
The dorsal trend includes the medial and dorsomedial
prefrontal surface, whereas the ventral trend includes
TF area TF of the parahippocampal cortexTFO area TFO of the parahippocampal cortexTGa agranular part of the temporal poleTGdd dysgranular part of the dorsal temporal poleTGdg granular part of the dorsal temporal poleTGsts sts part of the temporal poleTGvd ventral dysgranular part of the temporal poleTGvg ventral granular part of the temporal poleTH area TH of the parahippocampal cortexTpt temporoparietal areaV1 visual area 1 (primary visual cortex)V2 visual area 2V3v visual area 3, ventral partV4d visual area 4, dorsal part
V4t V4 transitional areaV4v visual area 4, ventral partV6 visual area 6 (or PO)V6Ad visual area 6A, dorsal divisionV6Av visual area 6A, ventral division (or PO)VA ventral anterior nucleusVAmc ventral anterior nucleus, magnocellular divisionVIP ventral intraparietal areaVLc ventral lateral caudal nucleusVLPFC ventrolateral prefrontal (network) cortexVMH ventromedial hypothalamic nucleusVPLo ventral posterior lateral oral nucleusVPM ventral posterior medial nucleus
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1643
the orbital and ventrolateral surfaces. Areas within each
trend are preferentially interconnected with each other
and with similar areas in other parts of the cortex,
although substantial individual variation is reported
between areas, especially for the caudal areas of each
trend (Yeterian et al., 2012).
In this study, the intrinsic and extrinsic cortical con-
nections of the LPFC were analyzed with anterograde
and retrograde tracers in an attempt to define distinct
connectional and functional networks. Tracers were
also injected into different parts of the superior and
inferior temporal cortex that are connected with the
LPFC. Initial results on intrinsic, intra-PFC connections
suggest that there are at least three relatively distinct
networks within the dorsal, ventrolateral, and caudolat-
eral parts of the LPFC (DPFC, VLPFC, CLPFC, respec-
tively). Further analysis of the extrinsic, or extra-PFC,
corticocortical connections of the networks indicated
that each of three networks has a unique pattern of
corticocortical connections. These constitute three
functionally different cortical circuits. In addition, there
is a small ventrolateral region, just rostral to the lower
limb of the arcuate sulcus (area 45a), that has connec-
tions with all of three networks in the LPFC. Some of
the present results have been reported in abstract form
(Saleem et al., 2008a).
MATERIALS AND METHODS
Retrograde tracers were injected into subregions of
the LPFC, including the frontal pole, in adult cynomol-
gous monkeys (Macaca fascicularis). In addition, some
cases with anterograde and retrograde tracer injections
in the inferior and superior temporal cortex, which had
been prepared and used in previous studies (Saleem
et al., 2008b), were reexamined and reanalyzed in rela-
tion to the connections with the LPFC (see Table 1 for
all tracer injections). All animal protocols were reviewed
and approved by the Animal Studies Committee of
Washington University and were in compliance with the
guidelines of the NIH for the care and use of laboratory
animals.
MRI, surgery, and tracer injectionThe details of the MR imaging, anesthesia, aseptic
surgery, and tracer injections have been described by
Saleem et al. (2008b). In brief, prior to surgery, each
monkey was anesthetized (see below) and placed in an
MRI-compatible stereotaxic frame. An MRI scan was
then obtained using a 1.5-T scanner, with a receive-only
or volume coil placed over the top of the head of the
animal. Stereotaxic coordinates for each desired injec-
tion site in the PFC that were specific for each individ-
ual animal were derived without correction from the MR
images. These individual-specific coordinates were com-
pared with coordinates from the cynomolgous monkey
brain atlas of Szabo and Cowan (1984).
For the surgery and MRI scans, anesthesia was
induced by intramuscular injection of ketamine (10
mg/kg) and xylazine (0.67 mg/kg). The animals were
then intubated, and surgical anesthesia was initiated
with a gaseous mixture of oxygen, nitrous oxide, and
halothane or isofluorane. Once anesthesia had been
established, the animals were placed in a stereotaxic
apparatus, and the scalp was incised. Craniotomies
were made in the skull at the sites indicated by the ste-
reotaxic analysis. The midline was defined from the
midsagittal sinus. After surgery, a longlasting analgesic,
buprenorphine (0.1 mg/kg, i.m.), was given as the ani-
mal was brought out of anesthesia.
For most of the LPFC injections, the expected locus
of the injection was determined from the coordinates
derived from the MRI scan, an appropriate craniotomy
was made, and the principal sulcus and the arcuate sul-
cus were exposed. Similarly, for most of the inferior
and superior temporal cortical injections, the superior
TABLE 1.
Tracer Injections in the Lateral Prefrontal and Temporal
Cortex
Case No.
Area(s)
injected Tracer Figure No.
Retrograde tracer injections in VLPFC (12r, 12l, and 46v cases)1 (OM84) 12r DY 2A, 3A, 42 (OM80) 12r DY 2B3 (OM9) 12l DY 2C4 (OM87) 12l FB 2D, 3B, 55 (OM77) 46v middle FB 2E, 3C, 66 (OM76) 46v middle FB 2F7 (OM82) 46v middle CTB 2G, 3D, 78 (OM76) 46v caudal DY 2HRetrograde tracer injections in VLPFC (45a and 45b cases)9 (OM86) 45a rostral FB 3E, 8A, 910 (OM84) 45a caudal FB 3F, 8B, 1011 (OM77) 45a/b LY 3G, 8C, 11Retrograde tracer injections in DPFC (9, 10, 46d, and 8B cases)12 (OM19) 10o FB 13A13 (OM81) 46d rostral DY 13B, 14D, 1714 (OM69) 10mr FR 13C, 14E, 1815 (OM64) 10mr FB 13D, 14A, 1516 (OM64) 9d rostral DY 13E, 14B, 1517 (OM77) 9 d/m FR 13F, 14F, 1618 (OM81) 9d caudal FR 13G19 (OM85) 46d caudal/8B? FB 13H20 (OM80) 46d middle FB 13I, 14C, 1921 (OM86) 8B LY 13J, 20Anterograde and retrograde tracer injections in the temporal
cortex22 (OM59) STSd/STGr and STSv LY and BDA 12A23 (OM55) STGr DY 21A24 (OM68) STGr and belt/
parabeltDY, FB, and CTB 21B, C, D
K.S. Saleem et al.
1644 The Journal of Comparative Neurology |Research in Systems Neuroscience

temporal sulcus and lateral sulcus were exposed after
appropriate craniotomy. The injection sites were then
determined with reference to these sulci, together with
the stereotaxic coordinates derived from the MRI scan.
Aqueous solutions of three retrograde tracers (fast
blue [FB; Sigma, St. Louis, MO; Dr. Illing; 2%], diamidino
yellow [DY; Sigma; 4%], and cholera toxin subunit B
[CTb; List Biological, Campbell, OR; 1–2%]) and an
anterograde tracer (biotinylated dextran amine [BDA;
Molecular Probes, Eugene, OR; 10%]) and two bidirec-
tional tracers (fluoro-ruby [FR; Molecular Probes; 5% or
10%], and Lucifer yellow [LY; Molecular Probes; 5% or
10%]) were injected in each animal. The injections were
made through micropipettes with an air pressure sys-
tem. The volume of tracers injected varied between 0.1
and 1.2 ll, depending on the sensitivity of tracers. To
avoid spread of tracer into areas along the pipette
track, the micropipette was left in place for 10–20
minutes after the injection was finished. With this pro-
cedure, there was little spread of tracer into the overly-
ing cortex or white matter.
Perfusion and histological processingAfter a survival period of 2 weeks, the animals were
anesthetized with ketamine (10 mg/kg, i.m.), followed
by sodium pentobarbital (25–30 mg/kg i.v.), and per-
fused with a pH shift fixation method as described by
Carmichael and Price (1994), with slight modifications
(Saleem et al., 2008b). In this method, the animals
were first perfused transcardially with warm heparinized
saline, followed by a sequence of cold 4% paraformalde-
hyde in 0.1 M sodium acetate buffer (pH 6.5), then 4%
paraformaldehyde in 0.1 M borate buffer (pH 9.5), and
finally 4% paraformaldehyde and 10% sucrose in borate
buffer. The brain was blocked stereotaxically, removed,
photographed, and postfixed for 6 hours in the final fix-
ative/sucrose solution and then transferred through 20
and 30% sucrose in 0.1 M phosphate buffer (pH 7.2–
7.4) at 4�C.
After 3–4 days, the brain blocks were frozen in dry
ice and isopentane, and cut coronally at 50 lm thick-
ness on a sliding microtome. An alternating series of
sections was processed for each tracer, usually one
section out of 10 in each series, with 500-lm intervals
between adjacent sections. The fluorescent tracers FB
and DY were analyzed from unstained sections. BDA
was processed directly with the avidin-biotin-peroxidase
method. The other tracers, CTb, FR, and LY, were proc-
essed immunohistochemically with an avidin-biotin-
horseradish peroxidase method (Carmichael et al.,
1994; Haber et al., 2000). For these tracers, the sec-
tions were first processed to block the biotin from the
injected BDA (Avidin/Biotin Blocking Kit; Vector, Burlin-
game, CA) and then incubated for 3–3.5 days in the pri-
mary antibody (anti-CTb: List Biological; No. 703;
1:10,000–20,000; antitetramethylrhodamine [for FR]
and anti-LY: Molecular Probes; Nos. A-6397 and
A-5750; 1:1,000). The sections were then processed
with the appropriate biotinylated secondary antibody
and avidin/biotin staining kit (Vector) with diaminoben-
zidene as the chromogen (for other details see Saleem
et al., 2008b). For BDA, LY, and FR, the immunostaining
was enhanced with a silver/gold intensification method,
which made the labeled axons and cells visible with
darkfield illumination (Carmichael and Price, 1994).
Additional series of sections were processed with Nissl,
acetylcholinesterase (AChE), and myelin (Gallyas) stains,
or immunohistochemically with antibodies against par-
valbumin (Sigma.; Nos. P3171 and P3088) and a non-
phosphorylated epitope of the neurofilament protein
(recognized by the SMI-32 antibody; Sternberger Mono-
clonals, Baltimore, MD). The specificity of antibodies
against parvalbumin and SMI-32 has been described in
our previous article (Saleem et al., 2007). In brief, the
parvalbumin antibody was raised against parvalbumin
from carp muscle. It was determined to be specific by
immunoblotting (Western blot), staining specifically the
12,000-molecular-weight band that was identified as
parvalbumin by Ca binding (Sigma data sheet). The par-
valbumin has been shown to stain subpopulations of
nonpyramidal neurons (GABAergic) in the neocortex and
to label different types of neurons in subcortical struc-
tures (Jones and Hendry, 1989; Saleem and Logothetis,
2012). The SMI-32 antibody recognizes a nonphos-
phorylated epitope of neurofilament H. It was shown to
be specific by immunoblot, in which it recognizes a
double band at MW 200,000 and 180,000; this double
band merges into a single neurofilament H line on two-
dimensional blots (Sternberger and Sternberger, 1983;
Goldstein et al., 1987). The antibody has been shown
to stain a subpopulation of pyramidal cells in the neo-
cortex (see, e.g., Campbell and Morrison, 1989; Hof
and Morrison, 1995); the pattern of staining seen in
this study corresponds to well-established patterns
from many previous studies.
Data analysis and presentationof illustrations
The location and the extent of each injection site and
the spatial distribution of labeled cell bodies and axonal
varicosities were plotted from the histological sections
with a microscope digitizer system that has encoders
attached to the microscope stage and is interfaced
with a personal computer (AccuStage, Shoreview, MN).
Cortical boundaries and other landmarks were added to
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1645

these plots by camera lucida drawings of adjacent
Nissl-, parvalbumin-, AChE-, and myelin-stained sec-
tions. Each labeled cell was plotted as a single point.
The files prepared with the microscope digitizer were
transferred to Adobe Illustrator with a utility supplied
by AccuStage and then into the drawing program Can-
vas (Deneba; ACD Systems), to add the section out-
lines, boundaries between gray and white matter,
architectonic areas, and labels. To ensure that all cells
were plotted, and were not double plotted, in many
cases the sections were scanned in rows, resulting in
the appearance of rows of cells on the plots. Beyond
this, the distribution of the labeled cells was not
altered, and they retained their exact position as plot-
ted at the microscope. The maps of individual sections
were then combined to make the figures. Photomicro-
graphs to document the tracer injection sites and pat-
terns of labeling were taken with a Nikon DXM1200
CCD camera attached to the microscope, using Nikon
ACT-1 image acquisition software. The images were
transferred into Adobe Photoshop for adjustment of the
brightness and contrast to show the injection sites and
labeled axons optimally and to size the images for
inclusion into a plate.
RESULTS
Subdivisions of the LPFCBased on the observation of intrinsic and extrinsic
connections in the present study, three relatively dis-
tinct networks were recognized in the LPFC (Fig. 1A).
1) The ventrolateral prefrontal network (VLPFC) includes
the cortex ventral to the principal sulcus on the ventro-
lateral convexity of the PFC. This network consists of
areas 12r, 12l, and 46v (except for its most caudal
part, area 45a). Area 45 (45a/b) is part of the ventro-
lateral convexity but has distinct and different connec-
tions and will be considered separately. 2) The dorsal
prefrontal network (DPFC) includes cortex dorsal to the
principal sulcus, extending around the dorsomedial con-
vexity. It consists of areas 8B, 9, and 46d (except for
its most caudal part), and the frontal polar parts of
areas 10m and 10o. 3) The caudolateral prefrontal net-
work (CLPFC) that includes cortex in and rostral to the
arcuate sulcus, extending forward within the caudal por-
tion of the principal sulcus (areas 8Ad/8Av and 46d/v).
The connections that constitute CLPFC network will be
described in detail in a later publication; in this article,
the CLPFC will be described only for comparison with
the other two networks.
Note that we have chosen not to use the term dorso-
lateral prefrontal cortex or DLPFC in this study in order
to avoid confusion with the common usage of this term
to denote either the entire LPFC or specifically the
region both dorsal and ventral to the principal sulcus
(e.g., Levy and Goldman-Rakic, 2000; Wallis and Miller,
2003; Fox et al., 2012).
The architectonic areas of the LPFC (Fig. 1A) are
delineated based on the descriptions by Walker (1940),
Preuss and Goldman-Rakic (1991), and Petrides and
Pandya (1999). It is useful to consider different areas in
relation to the principal and arcuate sulci. Caudal to
the principal sulcus, area 8A (frontal eye fields) occu-
pies the rostral bank of the arcuate sulcus and the
gyrus immediately rostral to the sulcus; it is usually
divided into dorsal and ventral parts (areas 8Ad and
8Av, respectively; Petrides and Pandya, 2002). Dorsal
to the principal sulcus, areas 8B, 9, and 10 are found
from caudal to rostral on the dorsomedial convexity
(Walker, 1940). Area 9 can be further divided into 9m
on the medial wall and 9d on the dorsal surface (Preuss
and Goldman-Rakic, 1991).
The most rostral part of the PFC is occupied by area
10, which has been previously divided into a medial
subdivision (area 10m) and a rostral orbital subdivision
(area 10o; Carmichael and Price, 1994). Area 10o is in
the most rostral part of the orbital cortex and extends
around the ventral aspect of the frontal pole. Area 10m
is located on the medial wall, between areas 32 and
14, and extends rostrally around the dorsal part of the
frontal pole. The area increases in granularity rostrally,
and it can be divided into a rostral area 10mr and a
caudal area 10mc (Saleem et al., 2008b; Fig. 1B).
These areas have previously been described as part of
the MPFC; areas 10mr and 10o will also be discussed
here because these areas extend around the frontal
pole into the LPFC.
Ventral to the principal sulcus, area 12 occupies the
rostral part of the ventrolateral convexity. Carmichael
and Price (1994) divided this region into areas 12r and
12l on the lateral side and areas 12m and 12o on the
orbital surface (Fig. 1A,B). Area 45 is found in the cau-
dal aspect of the convexity, back to the inferior limb of
the arcuate sulcus (Walker, 1940), and is characterized
by having large pyramidal cells in layer III (Walker,
1940; Petrides and Pandya, 2002; see also Carmichael
and Price, 1994, for the architectonic distinction
between areas 12l and 45). Based on the comparative
cytoarchitectonic analysis in both human and nonhu-
man primates, Petrides and Pandya (1999, 2002) have
subdivided this area into 45a and 45b, on the convexity
rostral to the arcuate sulcus and in the rostroventral
bank of the sulcus, respectively (see also Gerbella
et al., 2007, for the architectonic analysis of these
areas). We adapted this subdivision in the present
study (Fig. 1). For discussion related to area 45 and
K.S. Saleem et al.
1646 The Journal of Comparative Neurology |Research in Systems Neuroscience
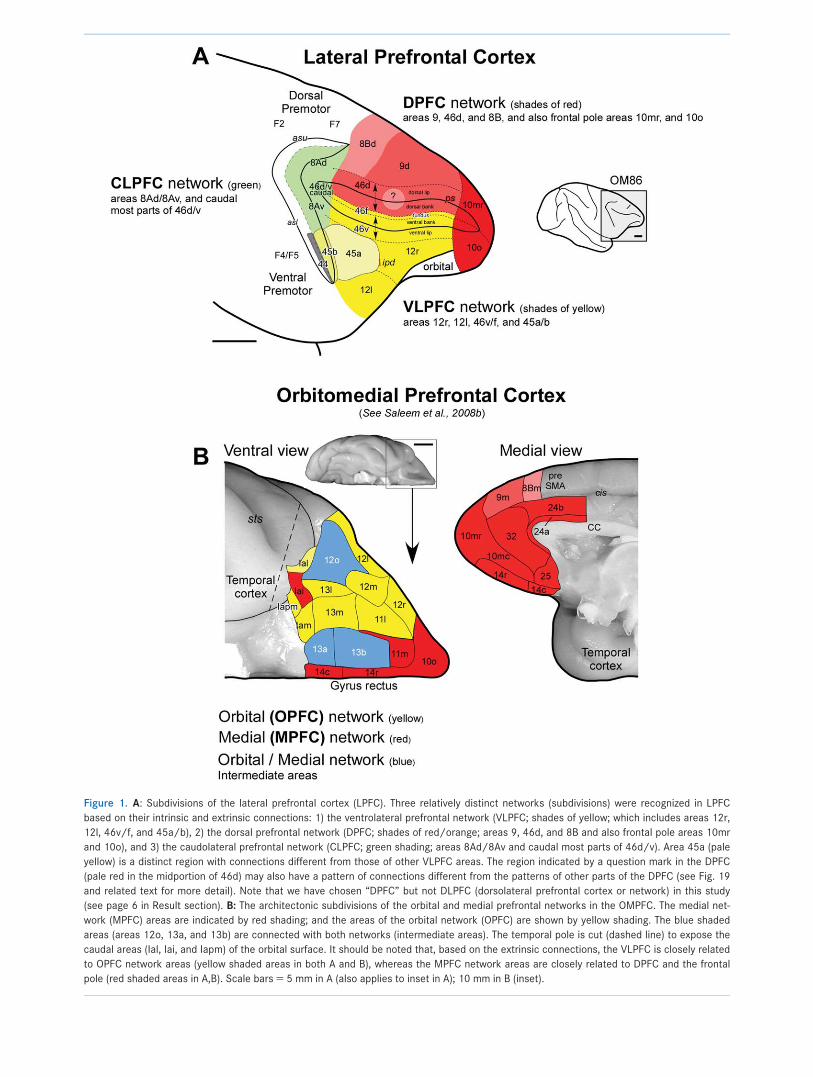
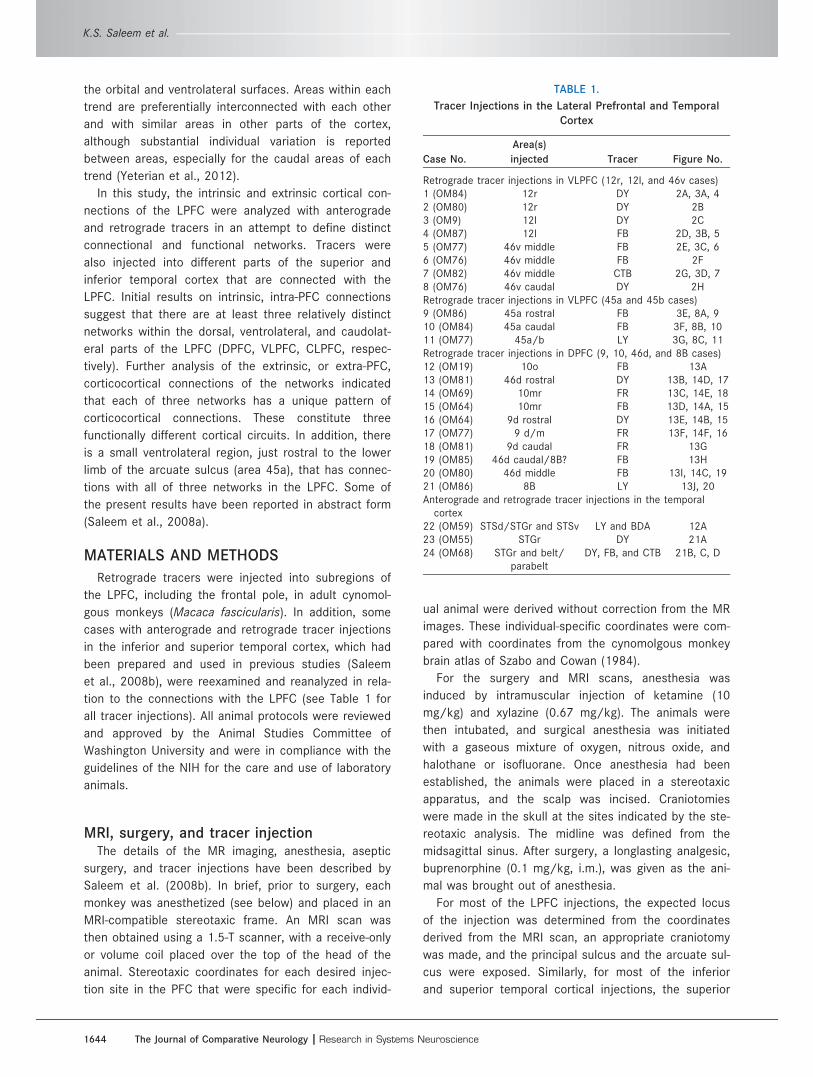
Figure 1. A: Subdivisions of the lateral prefrontal cortex (LPFC). Three relatively distinct networks (subdivisions) were recognized in LPFC
based on their intrinsic and extrinsic connections: 1) the ventrolateral prefrontal network (VLPFC; shades of yellow; which includes areas 12r,
12l, 46v/f, and 45a/b), 2) the dorsal prefrontal network (DPFC; shades of red/orange; areas 9, 46d, and 8B and also frontal pole areas 10mr
and 10o), and 3) the caudolateral prefrontal network (CLPFC; green shading; areas 8Ad/8Av and caudal most parts of 46d/v). Area 45a (pale
yellow) is a distinct region with connections different from those of other VLPFC areas. The region indicated by a question mark in the DPFC
(pale red in the midportion of 46d) may also have a pattern of connections different from the patterns of other parts of the DPFC (see Fig. 19
and related text for more detail). Note that we have chosen “DPFC” but not DLPFC (dorsolateral prefrontal cortex or network) in this study
(see page 6 in Result section). B: The architectonic subdivisions of the orbital and medial prefrontal networks in the OMPFC. The medial net-
work (MPFC) areas are indicated by red shading; and the areas of the orbital network (OPFC) are shown by yellow shading. The blue shaded
areas (areas 12o, 13a, and 13b) are connected with both networks (intermediate areas). The temporal pole is cut (dashed line) to expose the
caudal areas (Ial, Iai, and Iapm) of the orbital surface. It should be noted that, based on the extrinsic connections, the VLPFC is closely related
to OPFC network areas (yellow shaded areas in both A and B), whereas the MPFC network areas are closely related to DPFC and the frontal
pole (red shaded areas in A,B). Scale bars 5 5 mm in A (also applies to inset in A); 10 mm in B (inset).

neighboring areas in human and nonhuman primate
macaque see Petrides et al. (2012).
Area 46 is located in the both dorsal bank and ven-
tral bank of the principal sulcus and is clearly heteroge-
neous. Differences in the architectonic structure of
both banks of the principal sulcus suggest that further
subdivisions are possible in the dorsoventral axis (see
Preuss and Goldman-Rakic, 1991). We have consis-
tently noted differences in structure and connections
between the dorsal bank/lip and ventral bank/lip of
the principal sulcus and will refer to these as areas 46d
and 46v, respectively (see Fig. 1A; Saleem et al.,
2008a). Other distinctions between the fundus and dor-
sal and ventral banks of the principal sulcus may be
possible, but differences between these structures are
less consistent. As a working definition, we have
marked the fundus of the principal sulcus as 46f,
between 46d and 46v (Fig. 1A and other illustrations).
In the tables, 46f will be combined with 46v and
labeled 46v/f (e.g. see VLPFC section in Table 2).
The rostral and caudal parts of the principal sulcus
were distinguished by Petrides and Pandya (1999) as
areas 46 and 9/46d,v, respectively. Although we have
also found differences in the rostrocaudal axis, these
are less definitive, and these subdivisions will not be
explicitly delineated.
The subdivisions of areas in the orbitomedial prefron-
tal cortex (OMPFC; Fig. 1B), inferior and superior tem-
poral cortex, and other cortical areas are described and
illustrated in our previous work (Carmichael and Price,
1994; Saleem et al., 2007, 2008b; see also Saleem
and Logothetis, 2012).
Seltzer and Pandya (1978) and others (e.g., Baylis
et al., 1987) have differentiated several cytoarchitec-
tonic areas from rostral to middle parts of STS in the
rhesus monkey (Macaca mulatta): areas TAa, TPO, and
PGa in the lateral (including lip), central, and medial
part of the dorsal bank, respectively; IPa in the fundus
and very medial portion of the ventral bank; and TEa
and TEm in the medial and lateral part (including the
lip) of the ventral bank of the STS, respectively.
Although we found it useful to describe some of our
results in terms of these subdivisions, we found it diffi-
cult to define these areas precisely, especially their ros-
trocaudal borders. We therefore refer to the dorsal
bank, fundus, and ventral bank of the STS (d, f, and v,
respectively) in most descriptions.
Overview of connectionsFrom injections of retrograde tracers in all parts of
the lateral PFC, three patterns of corticocortical con-
nections within the PFC were apparent, related to the
three networks described above (VLPFC, DPFC, and
CLPFC). With a substantial degree of consistency, the
pattern of axonal label in each case corresponded to
one of these patterns. The majority of corticocortical
connections from each network are distinct from those
of the other two networks, both within the PFC and
with other parts of the cortex. The intrinsic connections
of each network (within the PFC) are generally with
other parts of the same network (see Tables 2 and 6).
Area 45a is an exception; it is connected to all three
networks (Table 4). The VLPFC is also connected to the
OPFC and the ventral premotor cortex, and the DPFC is
connected to the MPFC and the dorsal premotor
cortex.
Extrinsic connections outside the PFC link each net-
work into larger, connectionally distinct cortical cir-
cuits. The VLPFC is connected mainly to the
dysgranular insula, frontal operculum, somatic sensory-
related areas in the parietal operculum and inferior
parietal cortex, visual-related areas in the inferior tem-
poral cortex, and anterior cingulate areas (Table 3).
The DPFC is connected to the rostral part of the
superior temporal gyrus and adjacent dorsal bank of
the superior temporal sulcus (STGr/STSd), parahippo-
campal cortex, anterior and posterior cingulate, and
retrosplenial cortex (Table 6). For comparison, the
CLPFC is connected primarily with the posterior parie-
tal cortex, anterior and posterior cingulate cortex, and
caudal STS.
Ventrolateral prefrontal network (VLPFC)Area 12r, 12l, and 46v injectionsData from eight tracer injections in the VLPFC were
analyzed, four in area 12r or 12l in the ventrolateral
convexity and four in area 46v in the ventral bank and
lip of the principal sulcus (Fig. 2, Tables 2 and 3). An
additional three injections in area 45a/b were also
studied; these produced a substantially different pattern
of labeling and will be considered separately (see
below).
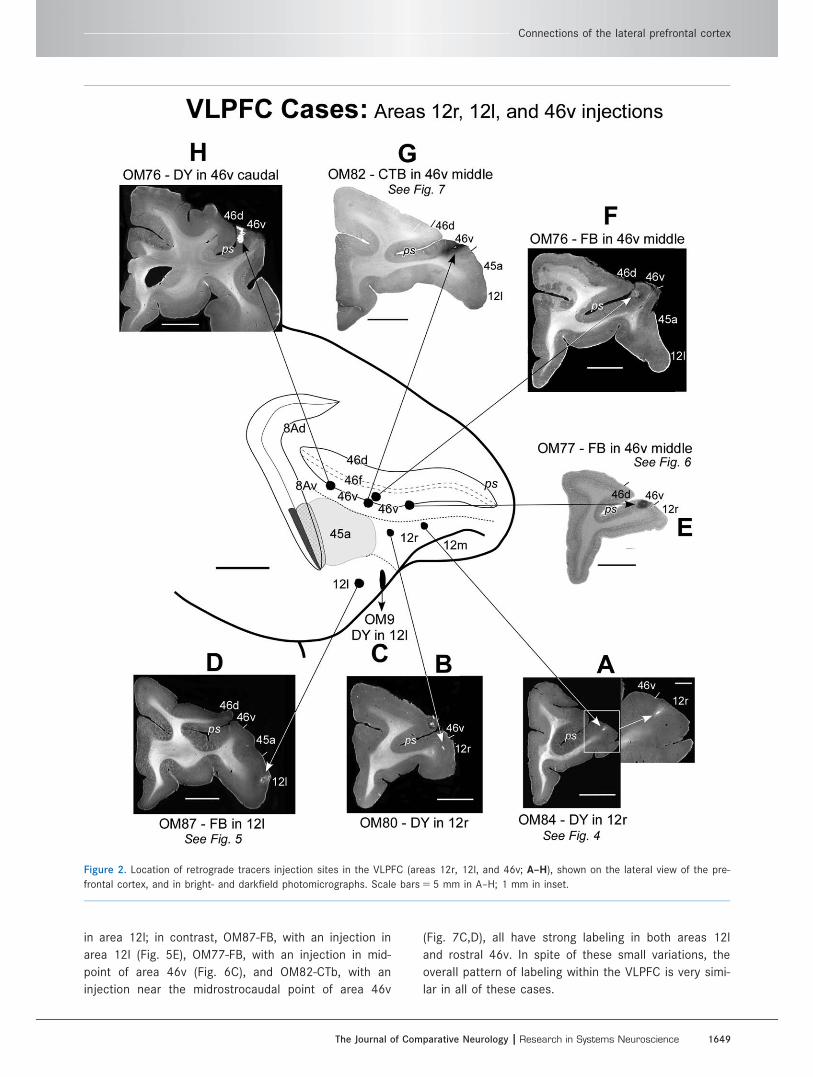
Intrinsic connections of the VLPFC within thePFC (Figs. 3–7, Table 2)Tracer injections in the VLPFC (except in area 45a; see
below) produced retrograde neuronal label mainly in the
VLPFC itself, in the ventral bank and lip of the principal
sulcus, and on the ventrolateral convexity (Fig. 3A–D).
In OM84-DY (Figs. 3A, 4A–E), in which an injection was
made into area 12r, labeled cells were found in ventro-
lateral areas 46v/f, 45a, 12r, 12m, and 12l. Similar pat-
terns of retrograde labeling were present in other cases
with injections in the VLPFC, but there are some differ-
ences in the detailed distribution (Table 2). For exam-
ple, in OM84-DY there are relatively fewer labeled cells
K.S. Saleem et al.
1648 The Journal of Comparative Neurology |Research in Systems Neuroscience
in area 12l; in contrast, OM87-FB, with an injection in
area 12l (Fig. 5E), OM77-FB, with an injection in mid-
point of area 46v (Fig. 6C), and OM82-CTb, with an
injection near the midrostrocaudal point of area 46v
(Fig. 7C,D), all have strong labeling in both areas 12l
and rostral 46v. In spite of these small variations, the
overall pattern of labeling within the VLPFC is very simi-
lar in all of these cases.
Figure 2. Location of retrograde tracers injection sites in the VLPFC (areas 12r, 12l, and 46v; A–H), shown on the lateral view of the pre-
frontal cortex, and in bright- and darkfield photomicrographs. Scale bars 5 5 mm in A–H; 1 mm in inset.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1649
There were few labeled cells in the dorsal or caudo-
lateral subdivisions of the LPFC (DPFC and CLPFC) fol-
lowing injections in areas 12r or 12l. With injections in
area 46v, in the ventral lip of the principal sulcus, how-
ever, cells are labeled in the dorsal lip of the principal
sulcus, in area 46d (e.g., Figs. 6C, 7B). The density and
position of this label varies along the rostrocaudal
extent of 46d from case to case, but it is present in all
four available cases with injections in area 46v (Table
2). Furthermore, the label in area 46d is not found only
at the level of the injection site and therefore cannot
be attributed to diffusion of the tracer from the injec-
tion site. The labeling therefore indicates that there are
at least some connections between areas 46v and 46d
and, to that extent, some connections between the
VLPFC and DPFC.
Figure 3. Extent of intrinsic connections within the LPFC labeled by injections into different parts of the VLPFC. A–D: Injections in areas
12r, 12l, and 46v. Note that the label following these injections is confined to the VLPFC. E–G: Injections in area 45a. In these cases,
labeled cells are found in all three prefrontal networks (DPFC, CLPFC, and VLPFC). Scale bars 5 5 mm.
K.S. Saleem et al.
1650 The Journal of Comparative Neurology |Research in Systems Neuroscience

Figure 4. A–J: Distribution of cells in the frontal and other cortical areas that were retrogradely labeled from an injection of DY into VLPFC
area 12r (case OM84). The inset at top left shows the location of the injection site and the extent of intrinsic labeling mapped onto the
lateral view of the brain (gray-shaded area; same as Fig. 3A). The actual plottings of retrogradely labeled cells in the frontal and other cort-
ical areas are illustrated in the coronal sections A–J. Each dot represents one labeled cell; because the section was plotted in rows, the
dots appear to be aligned into lines across the cortex. The thin radial lines through the cortex indicate architectonic areal boundaries, and
the dashed line indicates layer IV. Note that cells projecting to 12r (and other VLPFC areas; see below) are located in the sensory associa-
tion areas related to vision (STSv/f and TE) and somatic sensation (OPf, Id, SII, and 7b [PFG/PF]). Also note that no auditory-related areas
are labeled after this and other VLPFC injections, except those in area 45a (see Fig. 10). For abbreviations see list. Scale bars 5 5 mm.

The injection in OM76-DY is at the caudal edge of
the VLPFC, very near the CLPFC (Fig. 2H), and this
case serves to mark the junction between these two
networks. Within the PFC, most of the neuronal label
in the case is found in other parts of the VLPFC, as
with other injections in the VLPFC. In addition, how-
ever, there are labeled cells in areas of the CLPFC,
including areas 8Ad, 8Av, and the caudal part of 46d,v
(Table 2, column H).
Elsewhere in the PFC, the experiments with injections
in the VLPFC labeled neurons in the areas of the orbital
prefrontal network, especially in rostral areas 11l, 12m,
13l, and 13m. Label in more caudal areas in the orbital
network, especially those on the rostral part of the
Figure 5. A–I: Distribution of retrogradely labeled cells in the cerebral cortex after an FB injection into VLPFC area 12l (case OM87). The
other details are the same as in Figure 4. Note that there is no labeling in the auditory core/belt/parabelt areas. Scale bars 5 5 mm.
K.S. Saleem et al.
1652 The Journal of Comparative Neurology |Research in Systems Neuroscience
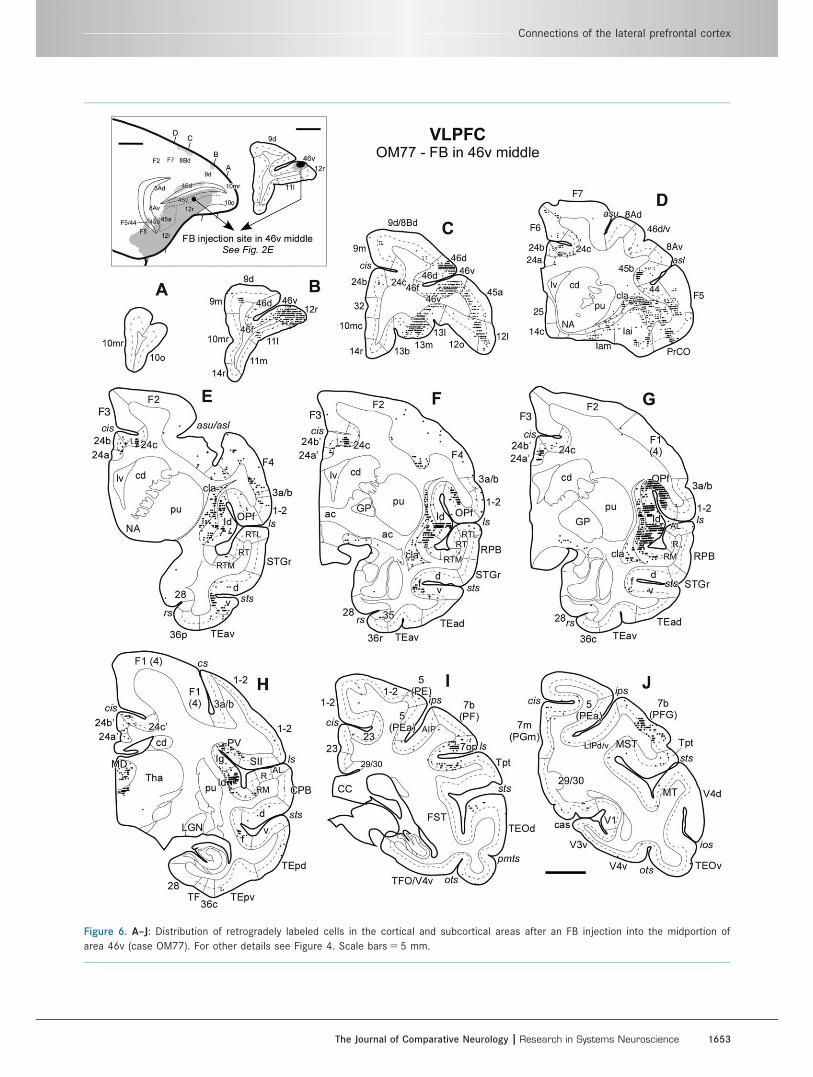
Figure 6. A–J: Distribution of retrogradely labeled cells in the cortical and subcortical areas after an FB injection into the midportion of
area 46v (case OM77). For other details see Figure 4. Scale bars 5 5 mm.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1653
insula (Iam, Iapm, and Ial), is more variable but is sub-
stantial in cases with strong overall labeling (see orbital
network section in Table 2).
There are almost no labeled cells in the areas of the
medial prefrontal network (areas 10m, 11m, 14c, 14r,
25, 32 or Iai; compare medial and orbital network sec-
tions in Table 2). In the caudal part of the frontal
cortex labeled neurons are found in area 44 (in the
fundus/caudal bank of the ventral limb of the
arcuate sulcus), in area PrCO on the frontal opercu-
lum, and in the ventral premotor area F5 (Figs. 3–7,
Table 2).
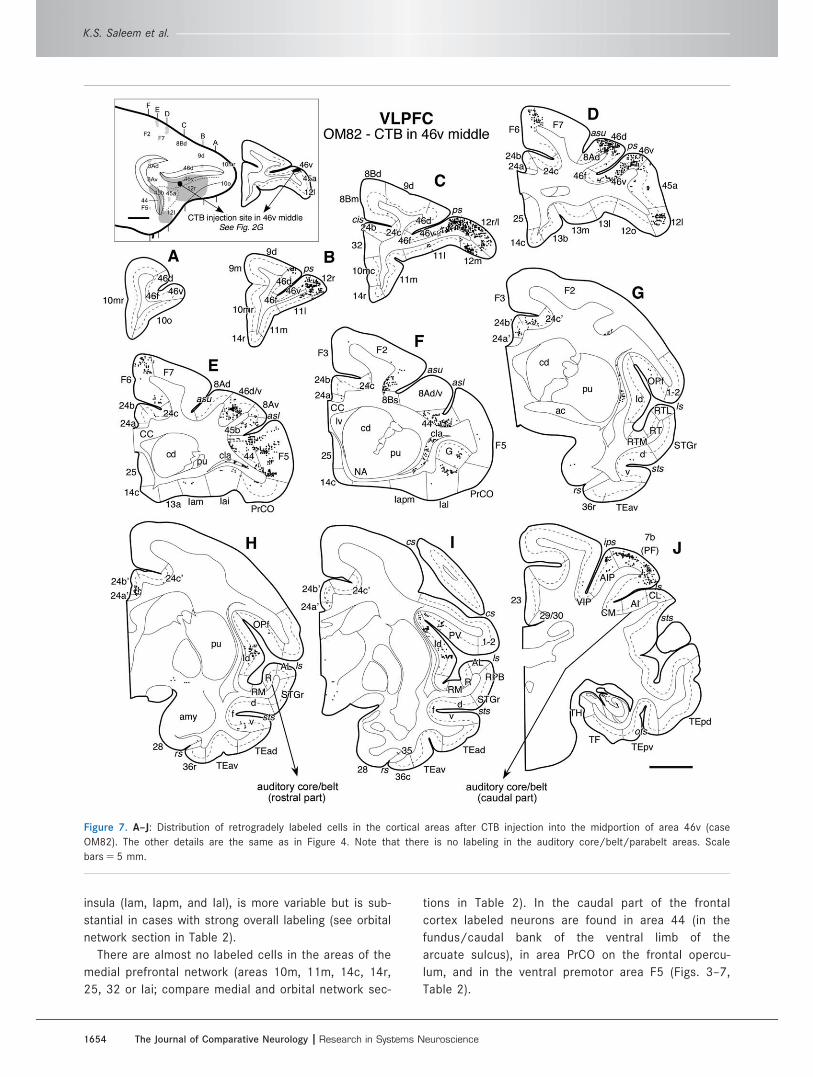
Figure 7. A–J: Distribution of retrogradely labeled cells in the cortical areas after CTB injection into the midportion of area 46v (case
OM82). The other details are the same as in Figure 4. Note that there is no labeling in the auditory core/belt/parabelt areas. Scale
bars 5 5 mm.
K.S. Saleem et al.
1654 The Journal of Comparative Neurology |Research in Systems Neuroscience

TABLE 2.
Distribution of Labeled Neurons Within the Prefrontal Cortex (Intrinsic Connections) After Injections in VLPFC Areas 12r,
12l, and 46v.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1655
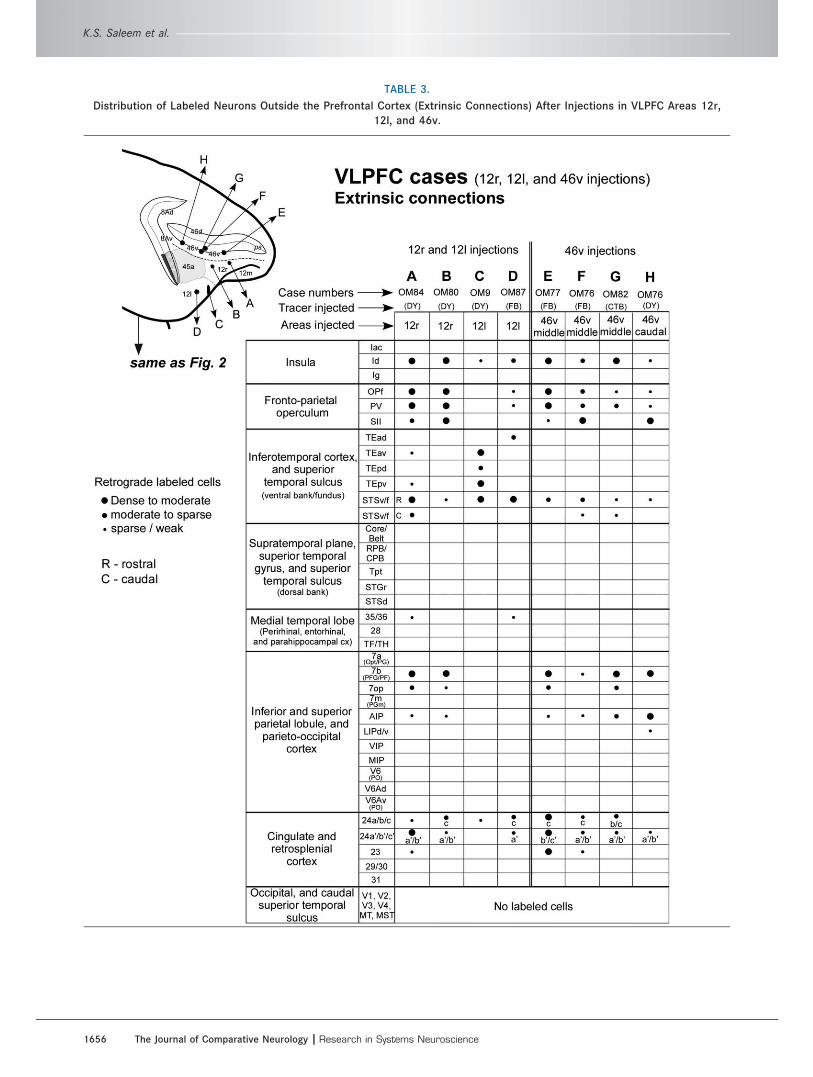
TABLE 3.
Distribution of Labeled Neurons Outside the Prefrontal Cortex (Extrinsic Connections) After Injections in VLPFC Areas 12r,
12l, and 46v.
K.S. Saleem et al.
1656 The Journal of Comparative Neurology |Research in Systems Neuroscience

Extrinsic connections of the VLPFC with othercortical areas (Figs. 4–7, Table 3)The experiments with injections in the VLPFC also show
a consistent pattern of extrinsic connections with corti-
cal areas outside the PFC. In general, these connec-
tions are with sensory association areas related to
somatic sensation and/or vision. As with the intrinsic
connections of the VLPFC, a major exception to this
pattern is seen only with injections in area 45a (see
below).
The constellation of connections of the VLPFC is shown
well in case OM77-FB, in which a retrograde tracer was
injected into area 46v, about midway along the ventral lip
of the principal sulcus (Figs. 2E, 6). Caudal to the frontal
cortex, most of the retrogradely labeled neurons are
found in the insula and somatic sensory related areas
(Fig. 6E–H). In particular, the dysgranular insula (Id) has
dense patches of labeled neurons through most of its
extent. There are also substantial numbers of labeled cells
in the frontal operculum (OPf), within the dorsal bank of
the lateral sulcus. The cells in OPf decrease in number
caudally, but they extend into the somatic sensory area
PV. There are very few labeled cells in area SII in this
case (Fig. 6H), but in other cases with injections in the
VLPFC area SII has substantial numbers of labeled cells
(Fig. 4H, Table 3). In the parietal cortex of OM77-FB,
labeled neurons are also found in the somatic sensory-
related area 7b (including its subdivisions PFG/PF), and in
nearby areas 7op and AIP (Fig. 6I,J, Table 3).
In the vision-related areas of the inferior temporal cortex
in OM77-FB, labeled cells are found in the ventral bank
and fundus of the superior temporal sulcus (STSv/f),
including areas IPa, PGa, and TEa/m of Seltzer and Pandya
(1978), but labeled cells are mostly absent in area TE in
the inferior temporal gyrus (Fig. 6E–H, Table 3). This is simi-
lar to other cases with injections in areas 46v and 12r. It
differs, however, from the cases with injections in area 12l
(OM9-DY and OM87-FB), in which there are fewer labeled
cells in Id and OPf and none in the parietal areas 7b and
7op (Fig. 5, Table 3). On the other hand, these cases have
more labeled neurons in the inferior temporal cortex, both
in STSv/f and in several subdivisions of area TE (TEad,
TEav, TEpd, and TEpv; Table 3).
In several cases, the labeled cells in STSf extend
from the fundus onto the medial part of STSd, in the
region identified as PGa/TPO by Seltzer and Pandya
(1978; e.g., Figs. 4E,H, 5G, 6E). These cells are not
found in the more lateral part, including the lip of the
STSd (areas TAa and TS of Seltzer and Pandya, 1978).
In the cingulate cortex, labeled cells are most consis-
tently found in the middle part of this region, in area
24a0/b0. Some cells are found in the more rostral cingu-
late region, but most of these are confined to area 24c,
within the ventral bank and fundus of the cingulate sul-
cus (Fig. 6D–H, Table 3).
In OM76-DY, in which the injection was at the bound-
ary between the VLPFC and CLPFC, a few cells were
also found in area LIP in the lateral bank of the intrapar-
ietal sulcus (Fig. 2H, Table 3). Neuronal label is not seen
in adjacent and related area 7a in this case, and no
other injection in the VLPFC produced label in these pos-
terior parietal areas. Both of these areas were consis-
tently labeled in experiments with injections in CLPFC.
Area 45a injectionsThree other, more caudal tracer injections in the VLPFC
were centered in area 45a or involved this area along
with area 45b (Fig. 8A–C). These experiments demon-
strated a pattern of intrinsic and extrinsic connections
very different from that in the experiments with more
rostral injections. They show that area 45a has exten-
sive connections with the DPFC and CLPFC as well as
the same intrinsic connections with other parts of the
VLPFC that are seen in other VLPFC injections
(although they are generally weaker). Area 45a is also
connected with cortical areas outside the PFC that are
related to the DPFC instead of the VLPFC.
An injection that was restricted to area 45b was not
available in this study. A previous study from another
group (Gerbella et al., 2010) indicates, however, that
the connections of area 45b are similar to those from
more rostral parts of the VLPFC (see discussion).
Intrinsic connections of area 45a within thePFC (Figs. 3, 9–11, Table 4)The distribution of transported label is well shown in
OM86-FB and OM84-FB, in which the tracer injections
are centered in rostral and caudal parts of area 45a,
respectively (Figs. 3E,F, 8A,B, 9, 10, Table 4). These
cases had labeled neurons in the VLPFC and in the
OPFC, although there were fewer labeled cells than in
other experiments with injections in other areas of the
VLPFC (compare orbital [OPFC] network section in
Tables 2, 4). Some labeled neurons were found in the
frontal polar part of area 10; in areas 8B, 9, and 46d of
the DPFC; and also in area 8Ad (frontal eye field) of the
CLPFC. There are almost no labeled neurons in the
medial PFC, but there are labeled cells in medial
network-associated areas 12o and Iai (Table 4). These
experiments suggest that area 45a may provide a link
for interaction between the VLPFC and other parts of
the lateral PFC.
The cases with injections in area 45a also have
labeled cells in the dorsal premotor areas F2 and F7
(e.g., Fig. 10C–E). Such connections are not seen in
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1657
most cases with injections in the VLPFC, although two
cases (OM9-DY and OM82-CTB) have labeled cells at
the border of F6 and F7 (e.g., Fig. 7D, Table 2).
Extrinsic connections of area 45a with othercortical areas (Figs. 9–11, Table 5)Caudal to the prefrontal cortex, the experiments with
injections in area 45a have relatively few or no retro-
gradely labeled cells in the somatic sensory or visually
related areas (in the Id, OPf, STSv/f, and TE) that are
connected with more rostral parts of the VLPFC (com-
pare Tables 3 and 5). Instead, there are many labeled
neurons in the superior temporal cortex, in areas that
are also connected to the DPFC (see below). OM86-FB
(Fig. 9) and OM77-LY (Fig. 11) had labeled cells in the
dorsal bank of the STS (STSd) and a few cells in the ros-
tral superior temporal gyrus (STGr). OM84-FB, with an
injection centered in caudal area 45a (Fig. 10, Table 5),
also had many cells in STSd and STGr, but in this case
many labeled cells extended into the auditory core,
medial and lateral belt, and parabelt areas caudal and
dorsal to STSd and STGr. This case had substantially
more neuronal labeling in the auditory areas than any
other case with a retrograde tracer injection in the pre-
frontal cortex. In all of the cases, the labeled cells are
found in both lateral and medial parts of STSd and
extend into STSf, including rostral parts of areas TS,
TAa, TPO, PGa, and IPa of Seltzer and Pandya (1978;
see Fig. 10F,G).
In addition to the experiments with injections in the
PFC, other cases are available from a previous study
(Saleem et al., 2008b) with injections in the parts of
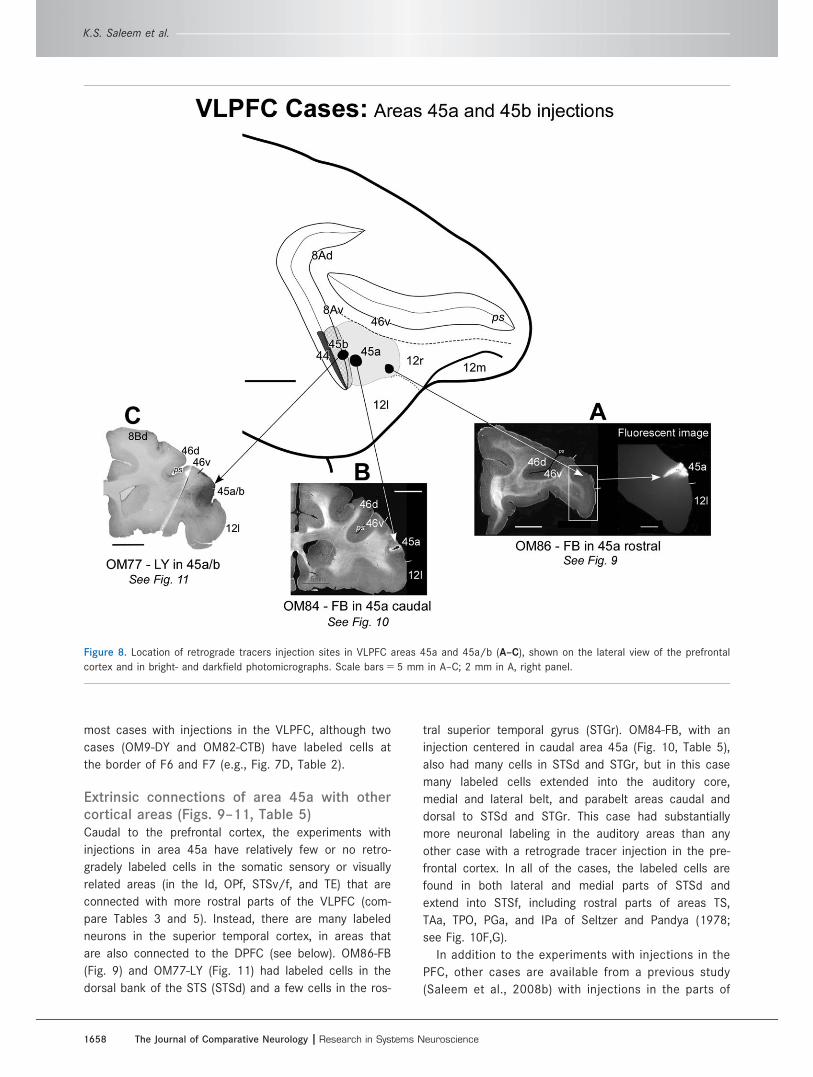
Figure 8. Location of retrograde tracers injection sites in VLPFC areas 45a and 45a/b (A–C), shown on the lateral view of the prefrontal
cortex and in bright- and darkfield photomicrographs. Scale bars 5 5 mm in A–C; 2 mm in A, right panel.
K.S. Saleem et al.
1658 The Journal of Comparative Neurology |Research in Systems Neuroscience
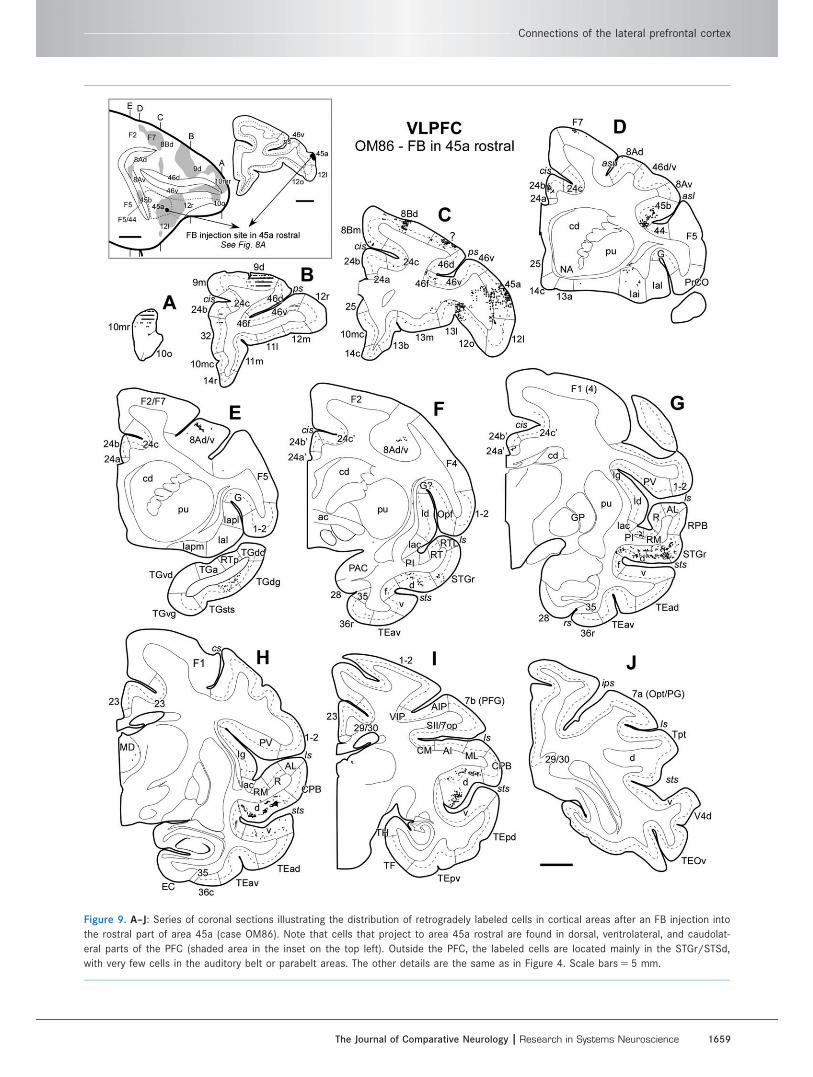
Figure 9. A–J: Series of coronal sections illustrating the distribution of retrogradely labeled cells in cortical areas after an FB injection into
the rostral part of area 45a (case OM86). Note that cells that project to area 45a rostral are found in dorsal, ventrolateral, and caudolat-
eral parts of the PFC (shaded area in the inset on the top left). Outside the PFC, the labeled cells are located mainly in the STGr/STSd,
with very few cells in the auditory belt or parabelt areas. The other details are the same as in Figure 4. Scale bars 5 5 mm.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1659
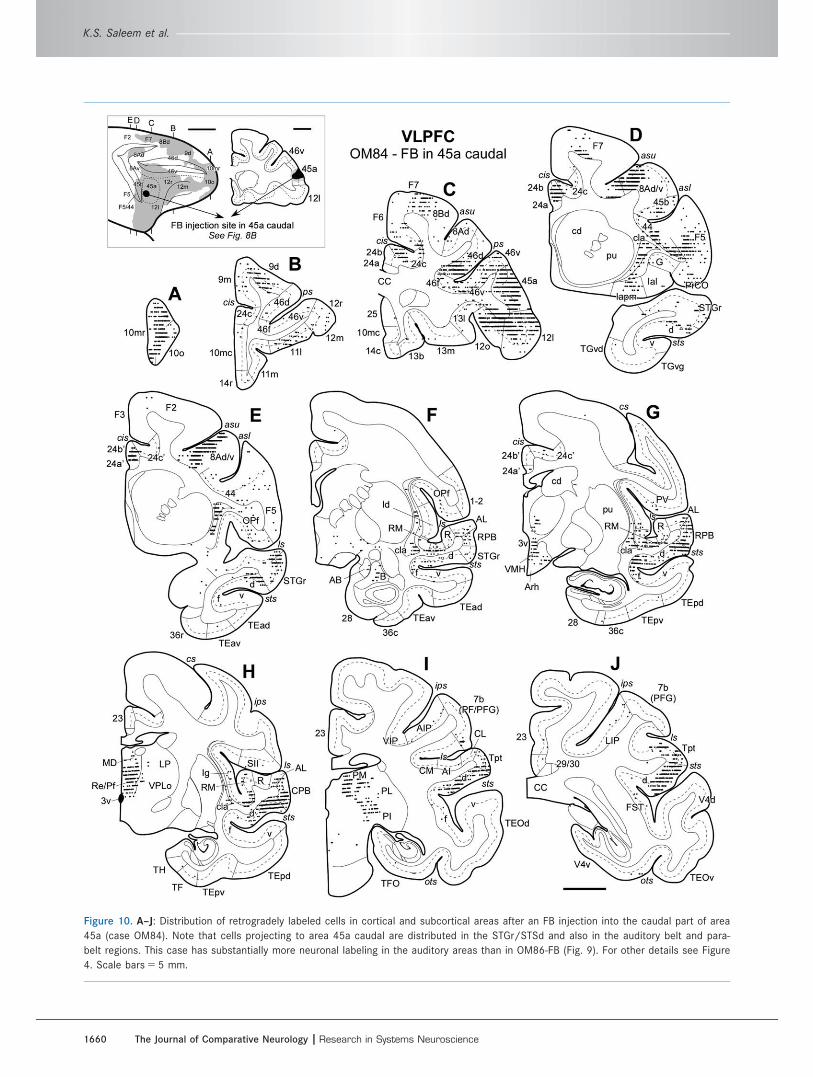
Figure 10. A–J: Distribution of retrogradely labeled cells in cortical and subcortical areas after an FB injection into the caudal part of area
45a (case OM84). Note that cells projecting to area 45a caudal are distributed in the STGr/STSd and also in the auditory belt and para-
belt regions. This case has substantially more neuronal labeling in the auditory areas than in OM86-FB (Fig. 9). For other details see Figure
4. Scale bars 5 5 mm.
K.S. Saleem et al.
1660 The Journal of Comparative Neurology |Research in Systems Neuroscience
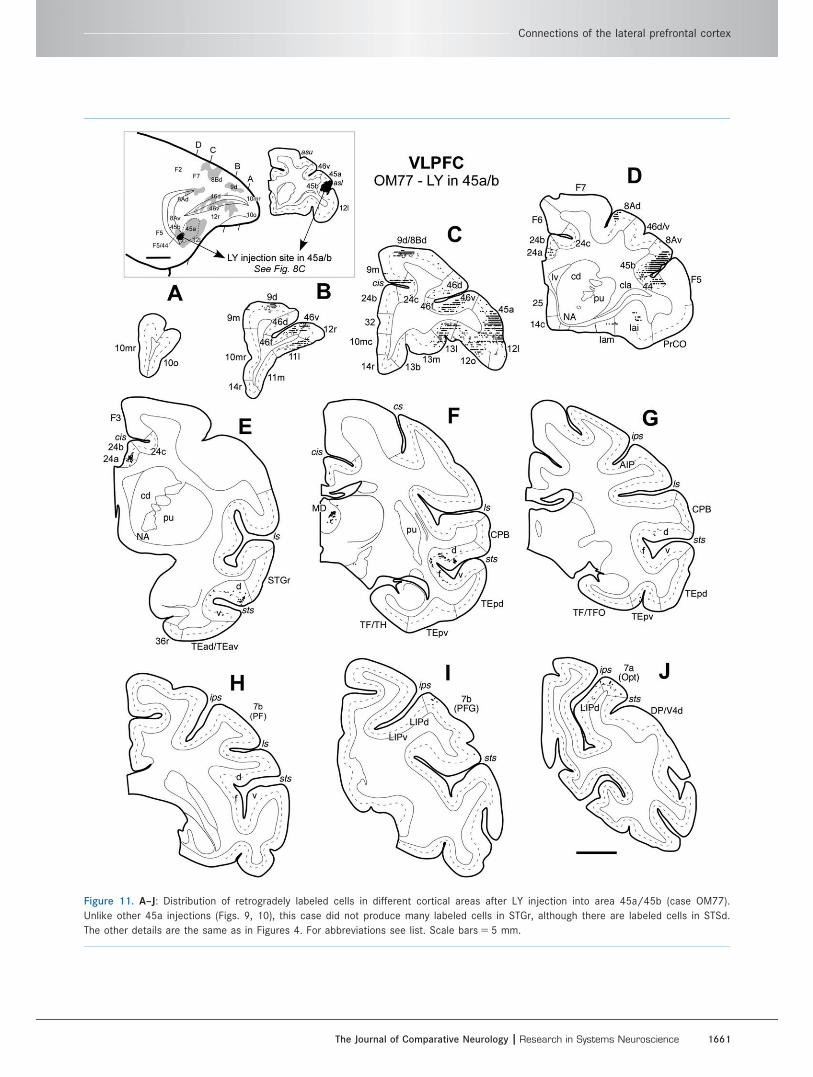
Figure 11. A–J: Distribution of retrogradely labeled cells in different cortical areas after LY injection into area 45a/45b (case OM77).
Unlike other 45a injections (Figs. 9, 10), this case did not produce many labeled cells in STGr, although there are labeled cells in STSd.
The other details are the same as in Figures 4. For abbreviations see list. Scale bars 5 5 mm.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1661
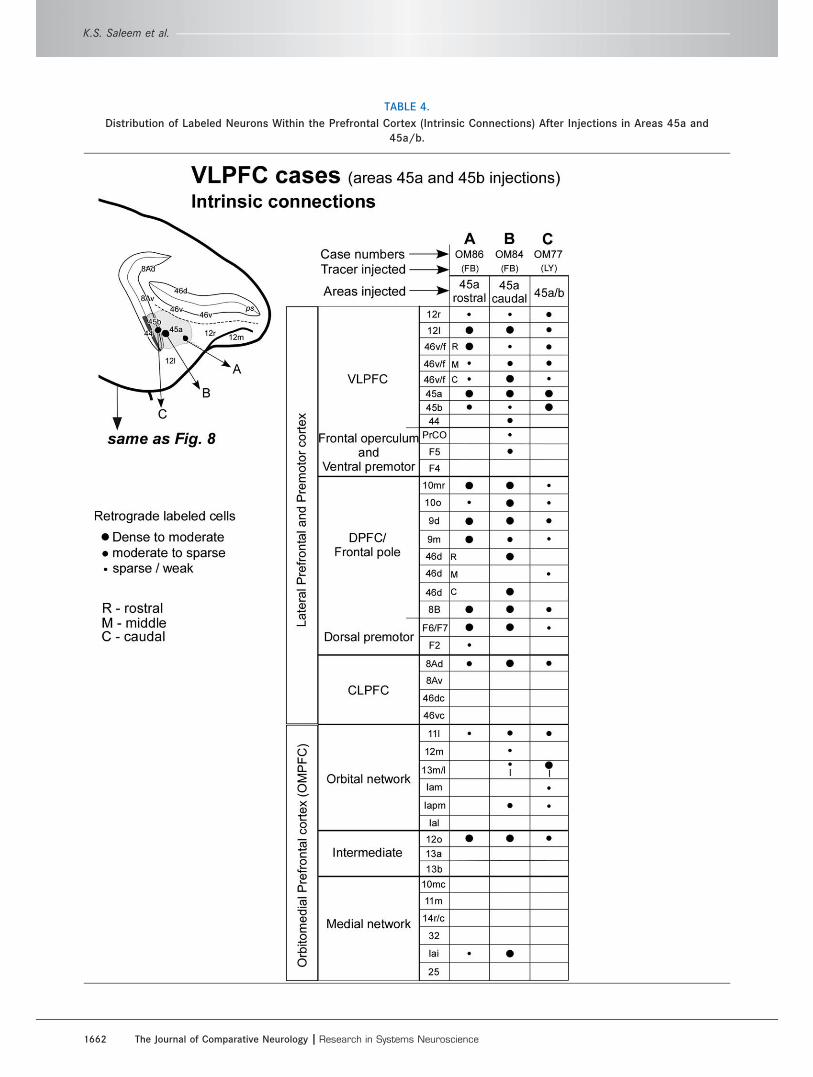
TABLE 4.
Distribution of Labeled Neurons Within the Prefrontal Cortex (Intrinsic Connections) After Injections in Areas 45a and
45a/b.
K.S. Saleem et al.
1662 The Journal of Comparative Neurology |Research in Systems Neuroscience
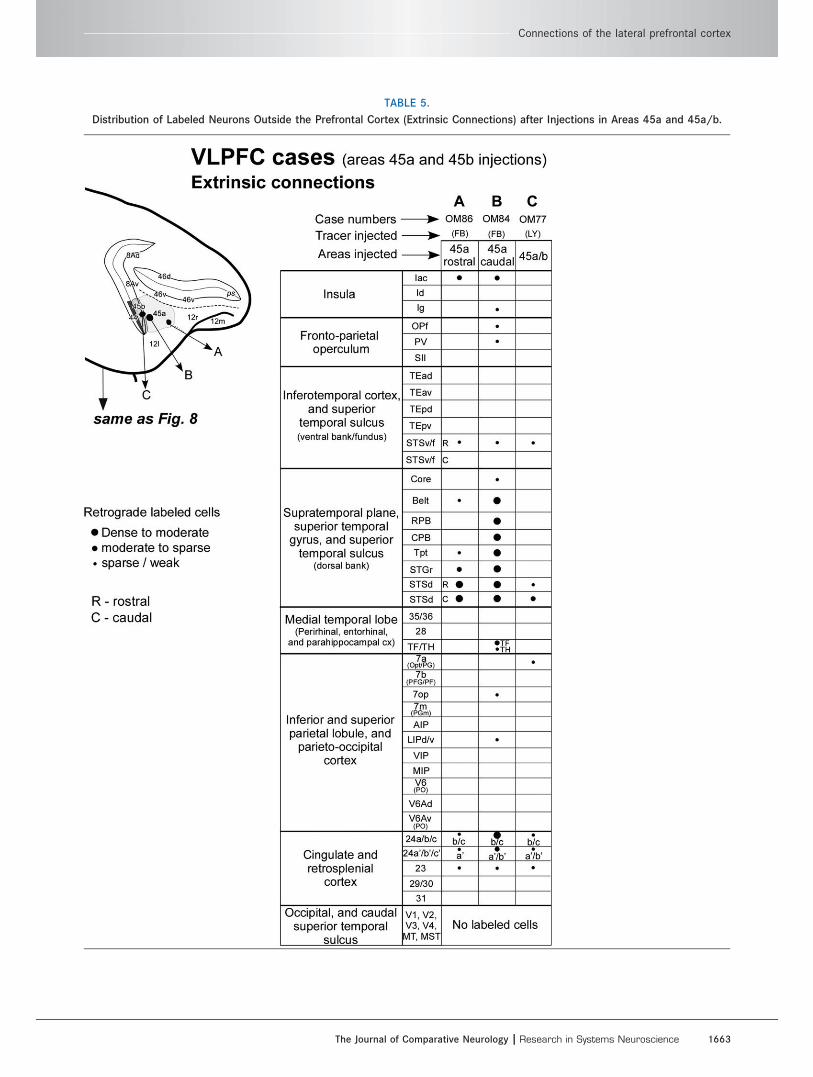
TABLE 5.
Distribution of Labeled Neurons Outside the Prefrontal Cortex (Extrinsic Connections) after Injections in Areas 45a and 45a/b.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1663
the superior and inferior temporal cortex areas that
have labeled cells following tracer injections in the
PFC. In particular, in case OM59, there are two
anterograde tracer injections in the STSd/STGr (LY)
and in STSv (BDA; Fig. 12). The injection in STSd/
STGr resulted in substantial axonal label in area 45a,
whereas the injection in STSv produced axonal label
in overlapping parts of area 45a, together with label
in areas 12l, 45b, and 46v. These confirm the previ-
ously described connections from STSd/STGr to area
45a (Figs. 9–11) as well as the connections from
STSv to area 12l (Fig. 5) and other parts of the VLPFC
(Table 3).
Dorsal prefrontal network (DPFC; includingfrontal pole)Area 9d/m, 46d, and 10mr/10o injectionsTen experiments are available in which injections of ret-
rograde axonal tracers were made into the DPFC. Four
of the injections are at the frontal pole (Fig. 13A–D;
OM19-FB in area 10o ventrally, OM81-DY in rostral
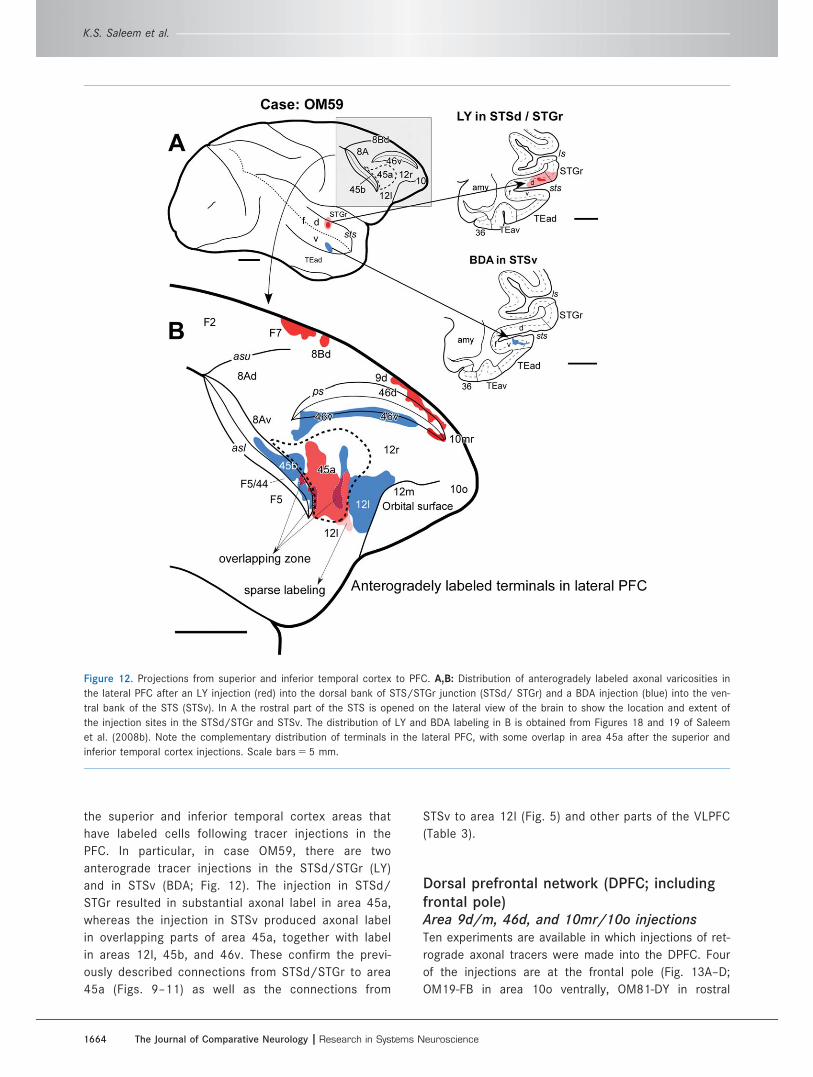
Figure 12. Projections from superior and inferior temporal cortex to PFC. A,B: Distribution of anterogradely labeled axonal varicosities in
the lateral PFC after an LY injection (red) into the dorsal bank of STS/STGr junction (STSd/ STGr) and a BDA injection (blue) into the ven-
tral bank of the STS (STSv). In A the rostral part of the STS is opened on the lateral view of the brain to show the location and extent of
the injection sites in the STSd/STGr and STSv. The distribution of LY and BDA labeling in B is obtained from Figures 18 and 19 of Saleem
et al. (2008b). Note the complementary distribution of terminals in the lateral PFC, with some overlap in area 45a after the superior and
inferior temporal cortex injections. Scale bars 5 5 mm.
K.S. Saleem et al.
1664 The Journal of Comparative Neurology |Research in Systems Neuroscience
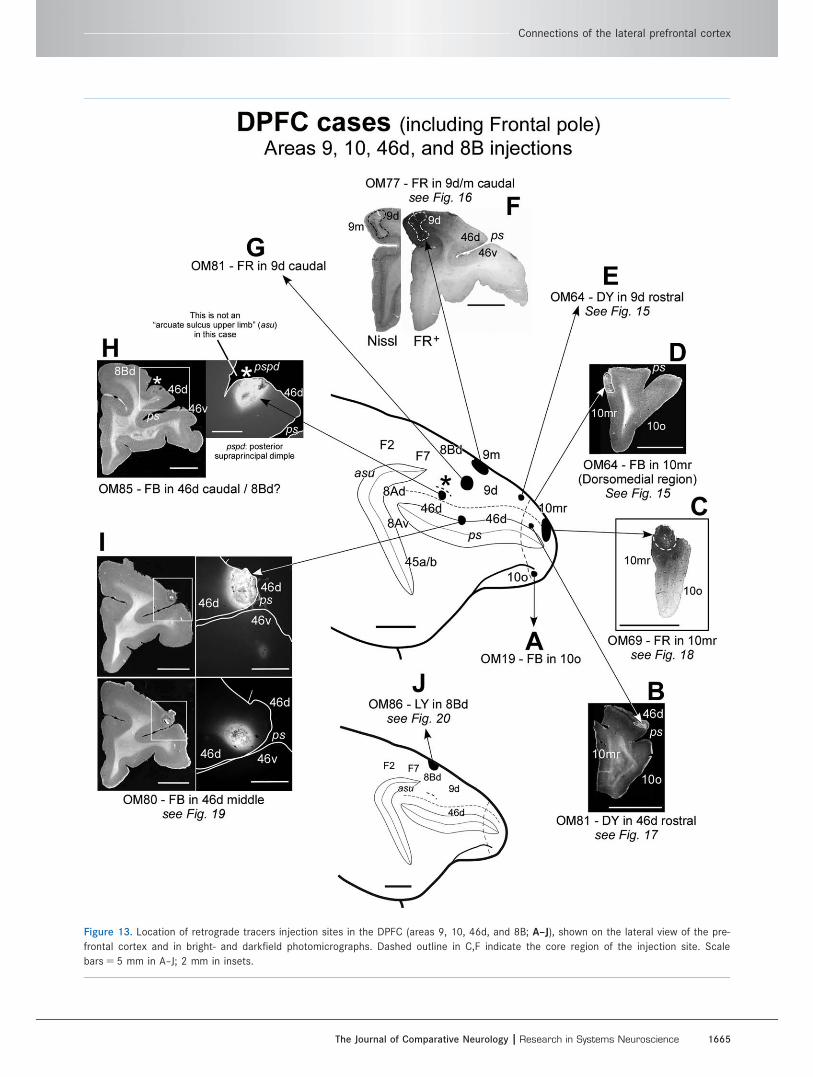
Figure 13. Location of retrograde tracers injection sites in the DPFC (areas 9, 10, 46d, and 8B; A–J), shown on the lateral view of the pre-
frontal cortex and in bright- and darkfield photomicrographs. Dashed outline in C,F indicate the core region of the injection site. Scale
bars 5 5 mm in A–J; 2 mm in insets.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1665
area 46d, OM69-FR and OM64-FB in area 10mr dor-
sally). Three injections are in area 9 (Fig. 13E–G;
OM64-DY and OM81-FR in 9d and OM77-FR in both 9d
and 9m). Two injections are in area 46d (Fig. 13H,I;
OM85-FB more caudally, overlapping the border
between areas 46d and 8B; OM80-FB in mid-46d at the
dorsal lip of the principal sulcus). Finally, one injection
was placed in area 8B (Fig. 13J; OM86-LY).
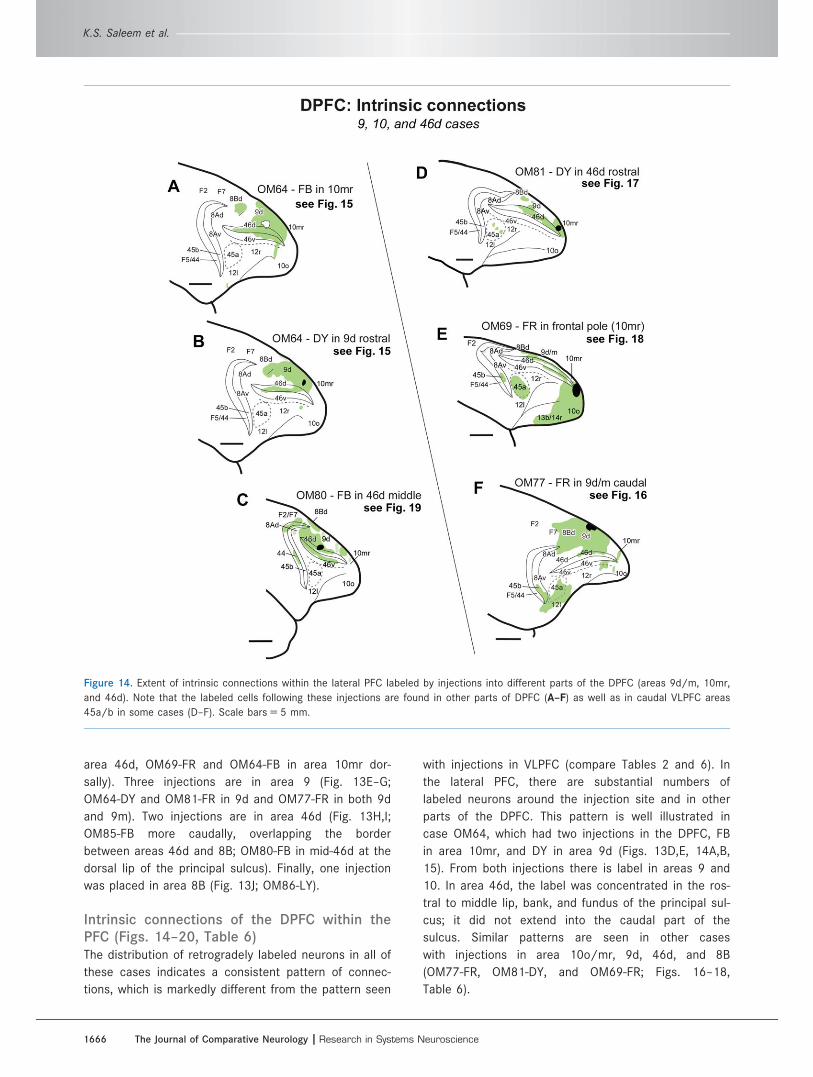
Intrinsic connections of the DPFC within thePFC (Figs. 14–20, Table 6)The distribution of retrogradely labeled neurons in all of
these cases indicates a consistent pattern of connec-
tions, which is markedly different from the pattern seen
with injections in VLPFC (compare Tables 2 and 6). In
the lateral PFC, there are substantial numbers of
labeled neurons around the injection site and in other
parts of the DPFC. This pattern is well illustrated in
case OM64, which had two injections in the DPFC, FB
in area 10mr, and DY in area 9d (Figs. 13D,E, 14A,B,
15). From both injections there is label in areas 9 and
10. In area 46d, the label was concentrated in the ros-
tral to middle lip, bank, and fundus of the principal sul-
cus; it did not extend into the caudal part of the
sulcus. Similar patterns are seen in other cases
with injections in area 10o/mr, 9d, 46d, and 8B
(OM77-FR, OM81-DY, and OM69-FR; Figs. 16–18,
Table 6).
Figure 14. Extent of intrinsic connections within the lateral PFC labeled by injections into different parts of the DPFC (areas 9d/m, 10mr,
and 46d). Note that the labeled cells following these injections are found in other parts of DPFC (A–F) as well as in caudal VLPFC areas
45a/b in some cases (D–F). Scale bars 5 5 mm.
K.S. Saleem et al.
1666 The Journal of Comparative Neurology |Research in Systems Neuroscience
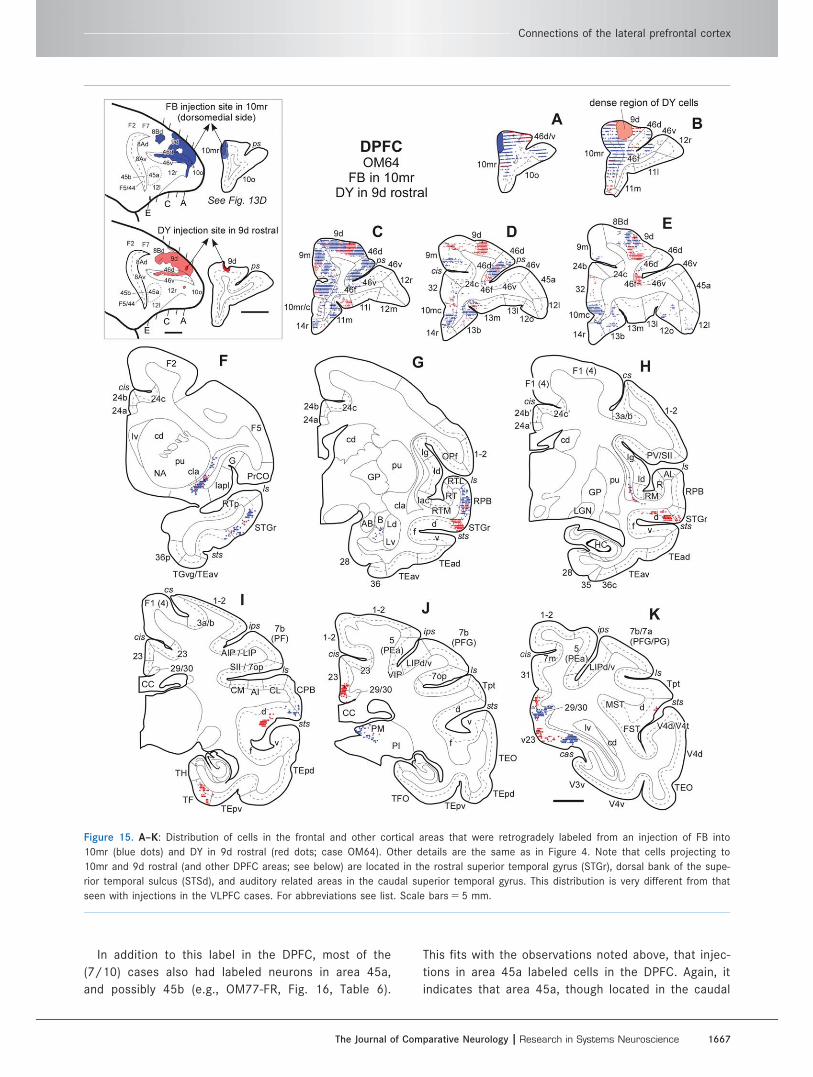
In addition to this label in the DPFC, most of the
(7/10) cases also had labeled neurons in area 45a,
and possibly 45b (e.g., OM77-FR, Fig. 16, Table 6).
This fits with the observations noted above, that injec-
tions in area 45a labeled cells in the DPFC. Again, it
indicates that area 45a, though located in the caudal
Figure 15. A–K: Distribution of cells in the frontal and other cortical areas that were retrogradely labeled from an injection of FB into
10mr (blue dots) and DY in 9d rostral (red dots; case OM64). Other details are the same as in Figure 4. Note that cells projecting to
10mr and 9d rostral (and other DPFC areas; see below) are located in the rostral superior temporal gyrus (STGr), dorsal bank of the supe-
rior temporal sulcus (STSd), and auditory related areas in the caudal superior temporal gyrus. This distribution is very different from that
seen with injections in the VLPFC cases. For abbreviations see list. Scale bars 5 5 mm.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1667

part of the VLPFC, is specifically connected with the
DPFC.
Most other parts of the VLPFC contained few if
any labeled cells from injections in the DPFC. In
many cases, however, labeled neurons were found in
the ventral bank of the principal sulcus (e.g., OM80-
FB; Fig. 19C,D), confirming the connections across
the principal sulcus described above. With injections
in the caudal part of DPFC, there were also a vari-
able number of labeled cells in area 8Ad and
Figure 16. A–J: Distribution of retrogradely labeled cells in the cerebral cortex after an FR injection into dorsomedial portion of caudal
area 9 (9d/m; case OM77). The other details are the same as in Figure 4. Scale bars 5 5 mm.
K.S. Saleem et al.
1668 The Journal of Comparative Neurology |Research in Systems Neuroscience
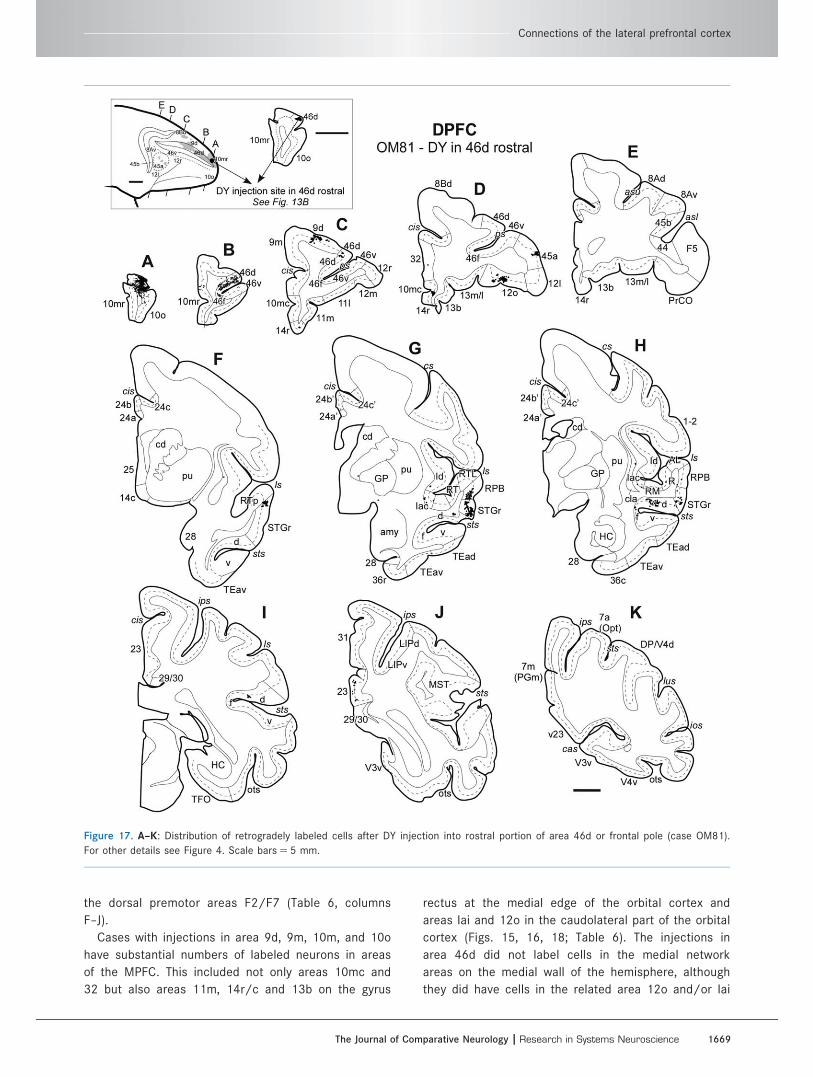
the dorsal premotor areas F2/F7 (Table 6, columns
F–J).
Cases with injections in area 9d, 9m, 10m, and 10o
have substantial numbers of labeled neurons in areas
of the MPFC. This included not only areas 10mc and
32 but also areas 11m, 14r/c and 13b on the gyrus
rectus at the medial edge of the orbital cortex and
areas Iai and 12o in the caudolateral part of the orbital
cortex (Figs. 15, 16, 18; Table 6). The injections in
area 46d did not label cells in the medial network
areas on the medial wall of the hemisphere, although
they did have cells in the related area 12o and/or Iai
Figure 17. A–K: Distribution of retrogradely labeled cells after DY injection into rostral portion of area 46d or frontal pole (case OM81).
For other details see Figure 4. Scale bars 5 5 mm.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1669
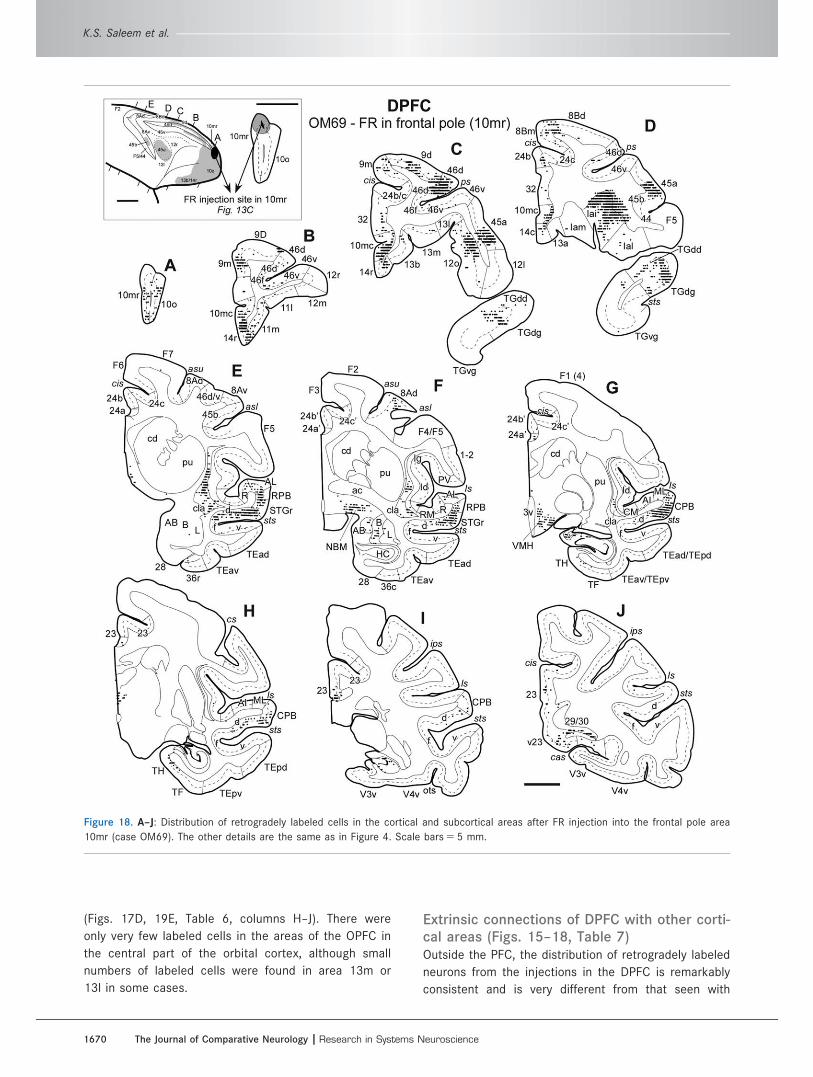
(Figs. 17D, 19E, Table 6, columns H–J). There were
only very few labeled cells in the areas of the OPFC in
the central part of the orbital cortex, although small
numbers of labeled cells were found in area 13m or
13l in some cases.
Extrinsic connections of DPFC with other corti-cal areas (Figs. 15–18, Table 7)Outside the PFC, the distribution of retrogradely labeled
neurons from the injections in the DPFC is remarkably
consistent and is very different from that seen with
Figure 18. A–J: Distribution of retrogradely labeled cells in the cortical and subcortical areas after FR injection into the frontal pole area
10mr (case OM69). The other details are the same as in Figure 4. Scale bars 5 5 mm.
K.S. Saleem et al.
1670 The Journal of Comparative Neurology |Research in Systems Neuroscience
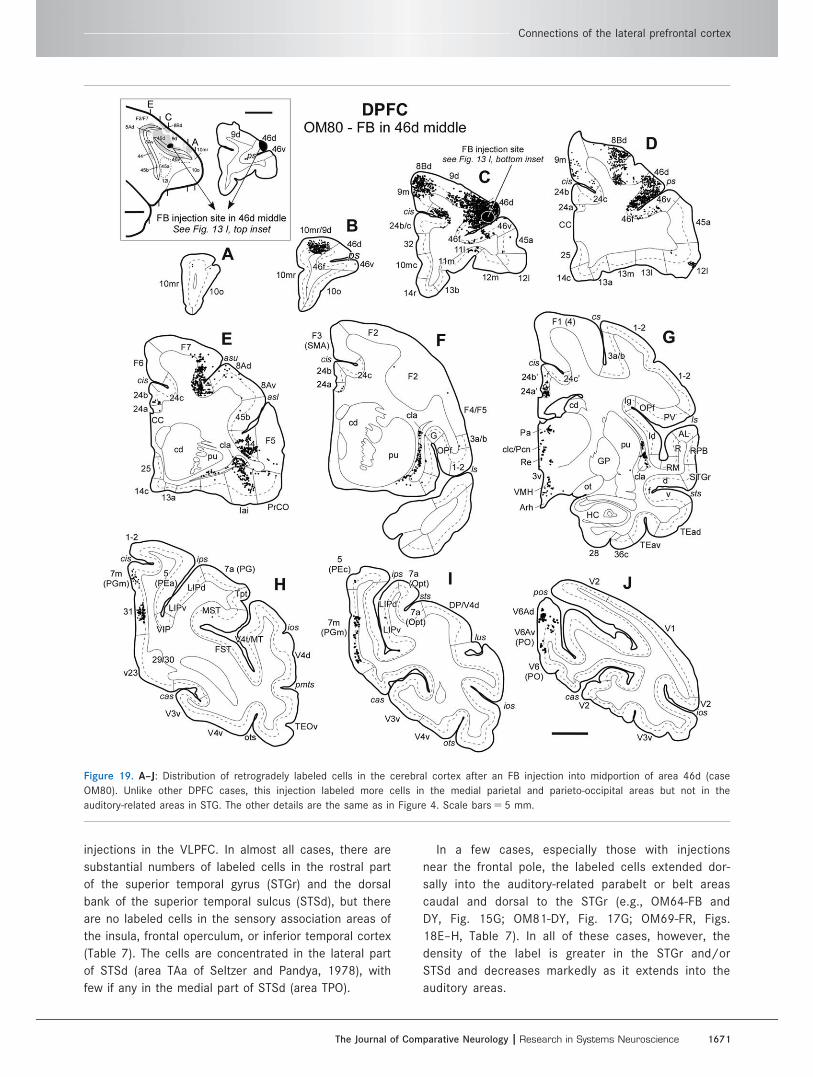
injections in the VLPFC. In almost all cases, there are
substantial numbers of labeled cells in the rostral part
of the superior temporal gyrus (STGr) and the dorsal
bank of the superior temporal sulcus (STSd), but there
are no labeled cells in the sensory association areas of
the insula, frontal operculum, or inferior temporal cortex
(Table 7). The cells are concentrated in the lateral part
of STSd (area TAa of Seltzer and Pandya, 1978), with
few if any in the medial part of STSd (area TPO).
In a few cases, especially those with injections
near the frontal pole, the labeled cells extended dor-
sally into the auditory-related parabelt or belt areas
caudal and dorsal to the STGr (e.g., OM64-FB and
DY, Fig. 15G; OM81-DY, Fig. 17G; OM69-FR, Figs.
18E–H, Table 7). In all of these cases, however, the
density of the label is greater in the STGr and/or
STSd and decreases markedly as it extends into the
auditory areas.
Figure 19. A–J: Distribution of retrogradely labeled cells in the cerebral cortex after an FB injection into midportion of area 46d (case
OM80). Unlike other DPFC cases, this injection labeled more cells in the medial parietal and parieto-occipital areas but not in the
auditory-related areas in STG. The other details are the same as in Figure 4. Scale bars 5 5 mm.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1671
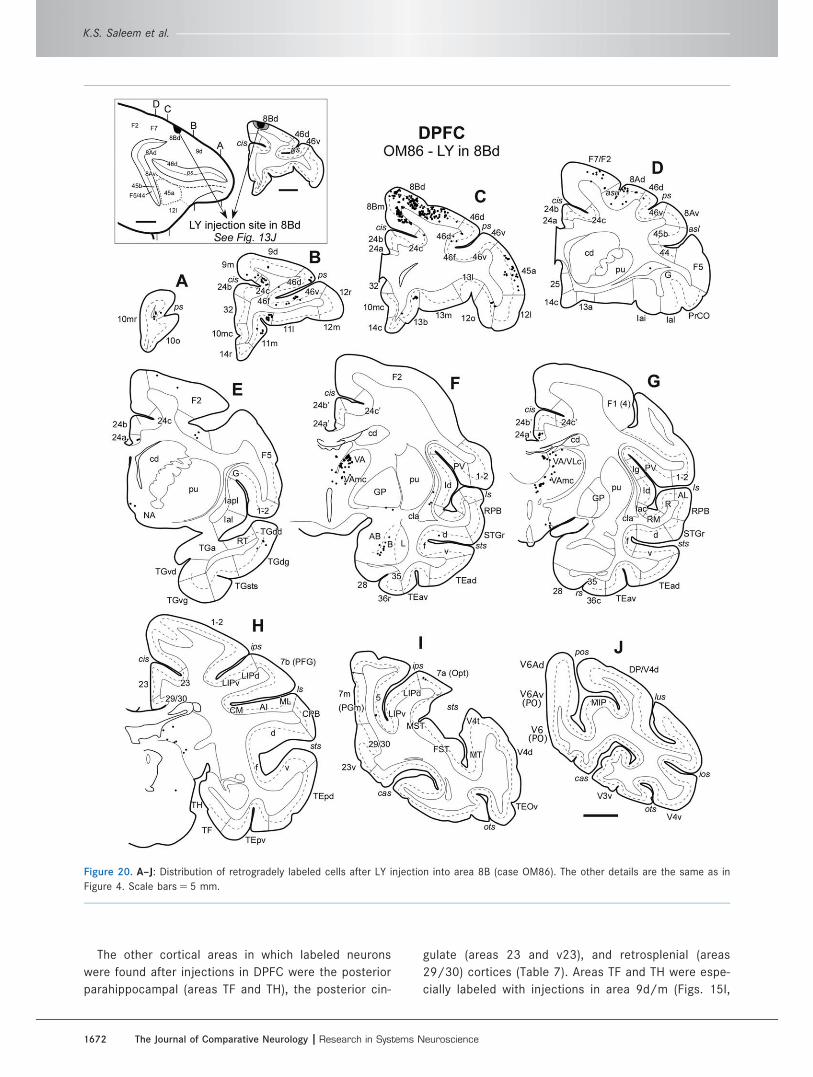
The other cortical areas in which labeled neurons
were found after injections in DPFC were the posterior
parahippocampal (areas TF and TH), the posterior cin-
gulate (areas 23 and v23), and retrosplenial (areas
29/30) cortices (Table 7). Areas TF and TH were espe-
cially labeled with injections in area 9d/m (Figs. 15I,
Figure 20. A–J: Distribution of retrogradely labeled cells after LY injection into area 8B (case OM86). The other details are the same as in
Figure 4. Scale bars 5 5 mm.
K.S. Saleem et al.
1672 The Journal of Comparative Neurology |Research in Systems Neuroscience

16G). The label in the posterior cingulate cortex was
found in virtually every case, but the distribution dif-
fered slightly among the subdivisions. In most DPFC
cases, there was also label in the rostral cingulate
area 24, but there were few if any cells in the middle
cingulate area 240.
Exceptional cases in DPFC (Figs. 19 and 20,Table 7)Although most of the cases with injections in the DPFC
have remarkably consistent patterns of retrograde neu-
ronal labeling, two cases produced variant patterns of
label. OM80-FB (Figs. 13I, 19), with an injection in the
TABLE 6.
Distribution of Labeled Neurons Within the Prefrontal Cortex (Intrinsic Connections) After Injections in DPFC and Frontal
Pole Areas 8B, 9, 10, and 46d.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1673

midportion of area 46d, differed from other DPFC cases
in that labeled cells were not seen in the STGr/STSd
but were found in the medial parietal area 7m (or area
PGm), area 31, and the parieto-occipital areas V6Ad
and V6Av (or area PO; Fig. 19G–J). This distribution of
labeling was not seen in any of the other DPFC cases
in this study. A possible explanation is that the midpor-
tion of the dorsal bank of the principal sulcus (indicated
by a question mark in Fig. 1A) is a distinct area with
connections different from those of the cortex rostral,
dorsal, and caudal to it. This is supported by the pat-
tern of label in the LPFC in available cases that have
TABLE 7.
Distribution of Labeled Neurons Outside the Prefrontal Cortex (Extrinsic Connections) After Injections in DPFC and Frontal
Pole Areas 8B, 9, 10, and 46d.
K.S. Saleem et al.
1674 The Journal of Comparative Neurology |Research in Systems Neuroscience

tracer injections in STGr (Fig. 12; OM59-LY; see also
Figs. 6F,G, 15A,B in Saleem et al., 2008b). None of
these cases has transported label in the midportion of
area 46d, in the dorsal lip of the principal sulcus.
OM86-LY, with an injection in area 8B (Fig. 20), also
had only a few if any labeled cells in STGr/STSd or in
the posterior parietal area 7a, which is usually labeled
only from the CLPFC. Because LY is a bidirectional
tracer, however, in this case it was possible to see that
there were anterogradely labeled axons in STGr/STSd
and areas 7m, retrosplenial cortex (29/30), and parie-
tooccipital area V6Ad/v. There is relatively little label
outside the PFC in this case, and the paucity of retro-
grade cellular label in STGr/STSd may be due to a low
efficacy of the tracer injection. Petrides and Pandya
(1999) reported a comparable experiment with a some-
what larger tracer injection in area 8B, which had a pat-
tern of labeling similar to that seen in OM86-LY but
with more label in STSd and area 7a (their areas TPO
and Opt, respectively).
Connections within the STG (Fig. 21)The results described above indicate that the STGr/
STSd region provides a major input to the DPFC net-
work, and it has previously been shown to provide to
the MPFC network (Saleem et al., 2008b). Because the
STG as a whole is usually presumed to have an auditory
function, an important question is whether there are
connections between the auditory-defined belt/parabelt
areas and STGr/STSd. Experiments were therefore per-
formed and analyzed that have tracer injections into
either the belt/parabelt areas or the STGr (OM55 DY;
OM68 DY, FB, and CTB; Fig. 21A–D). The results indi-
cate that neurons labeled from an injection in STGr are
distributed mostly throughout the STGr/STSd but also
extend caudally and dorsally to overlap the edge of the
rostral and caudal parabelt (RPB and CPB, respectively;
see Fig. 21A,B). Conversely, after injections in the belt/
parabelt, retrogradely labeled neurons are most densely
distributed in the core/belt/parabelt but also extend
rostrally and ventrally to overlap the STGr/STSd (see
Fig. 21C,D).
Laminar distribution of labeled cellsAlthough detailed cell counts were not made, inspection
of the plots of retrogradely labeled neurons indicates
that labeled cells were generally distributed in both
superficial and deep layers of the cortex (see discus-
sion below). Little indication of any more specific lami-
nar organization was found, except the connections
from medial temporal lobe areas 28 and TF/TH. In
these areas, the labeled neurons were distributed
mostly in the deep layers after injections in DPFC area
9d/m (Fig. 16F,G) and frontal pole area 10mr (Fig.
18G,H; see also Kondo et al., 2005, their Fig. 14A for
the projections from area 28 to medial prefrontal
network).
In some cases, the labeled neurons in some architec-
tonic areas within and outside the prefrontal cortex
(orbitofrontal and STS areas) appeared to be in superfi-
cial layers (2–3) and gradually moved to the deep
layers (5–6) in the adjacent sections, or vice versa, pos-
sibly because of the tangential plane of sections
through cortical gyri (e.g., see labeled neurons in cen-
tral orbital area 13m in Fig. 5D,E). This pattern was
also reported previously for the distribution of antero-
gradely labeled terminals in the temporal cortex (see
the connections from area TEav to STS in Saleem et al.,
2000, their Fig. 10, top right).
DISCUSSION
The results presented here, based on analysis of 24
axonal tracer injections in 14 monkeys, indicate that
the areas on the lateral surface of the PFC can be
divided into three distinct networks, in the dorsal, ven-
trolateral, and caudal parts of this large cortical region.
Areas in each of these networks are preferentially con-
nected to other areas within the same part of the PFC
and to a distinctive set of areas elsewhere in the cere-
bral cortex. In addition, area 45a in the ventrolateral
part of the PFC is a special region that is connected
across all three PFC networks and has a unique set of
extrinsic connections outside the PFC. A second unique
region may exist in the midrostrocaudal part of area
46d (in the dorsal bank/lip of the principal sulcus), but
there is not currently sufficient evidence for a definitive
definition of this area.
These results expand previous observations on the
OMPFC that recognized two networks, which were
termed the “orbital” and “medial” prefrontal networks
(although neither corresponds solely to the orbital or
medial cortical surface; Carmichael and Price, 1996;
Saleem et al., 2008b). The areas within each network
are preferentially interconnected by intrinsic connec-
tions (Carmichael and Price, 1996) and have distinct
and selective connections to other cortical and subcort-
ical structures (€Ong€ur et al., 1998; An et al., 1998;
Ferry et al., 2000; Kondo et al., 2003, 2005; Hsu and
Price 2007; Saleem et al., 2008b). Perhaps the most
striking observation in the current study is that VLPFC
and DPFC networks are very similar in their connections
to the orbital and medial networks (OPFC and MPFC),
respectively. The combined MPFC/DPFC and OPFC/
VLPFC networks are similar to previously proposed dor-
sal and ventral architectonic “trends” in the prefrontal
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1675
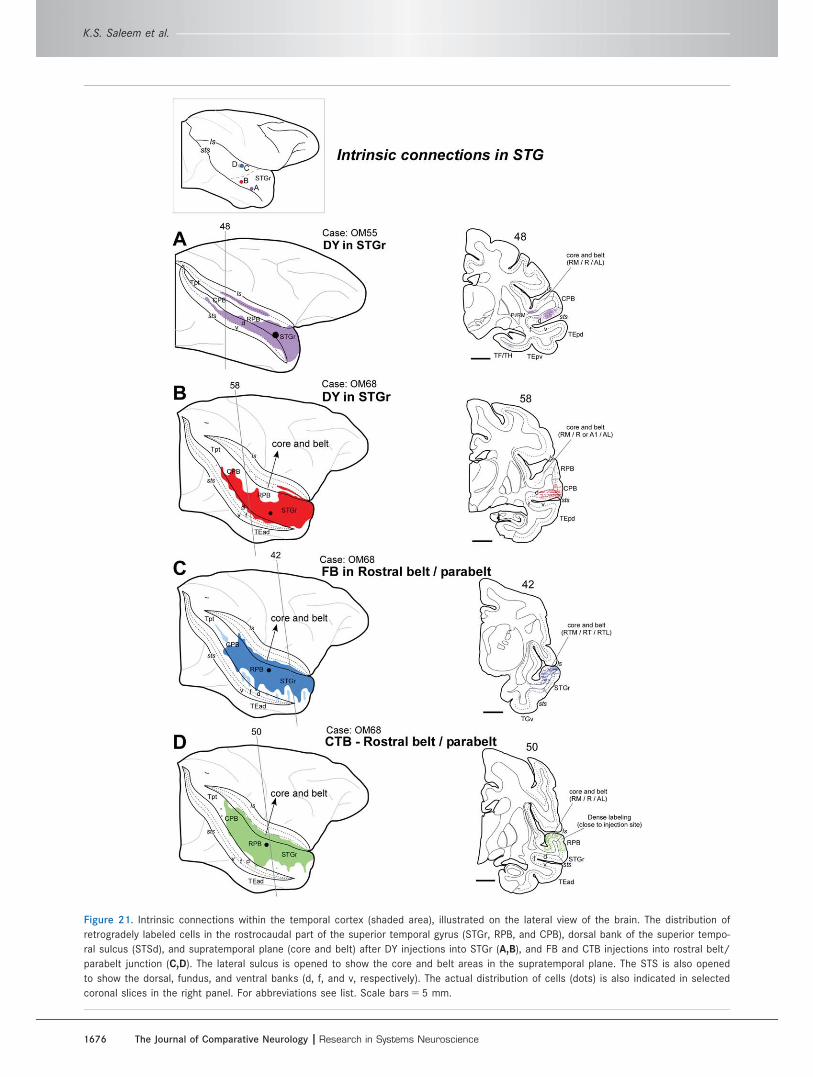
Figure 21. Intrinsic connections within the temporal cortex (shaded area), illustrated on the lateral view of the brain. The distribution of
retrogradely labeled cells in the rostrocaudal part of the superior temporal gyrus (STGr, RPB, and CPB), dorsal bank of the superior tempo-
ral sulcus (STSd), and supratemporal plane (core and belt) after DY injections into STGr (A,B), and FB and CTB injections into rostral belt/
parabelt junction (C,D). The lateral sulcus is opened to show the core and belt areas in the supratemporal plane. The STS is also opened
to show the dorsal, fundus, and ventral banks (d, f, and v, respectively). The actual distribution of cells (dots) is also indicated in selected
coronal slices in the right panel. For abbreviations see list. Scale bars 5 5 mm.
K.S. Saleem et al.
1676 The Journal of Comparative Neurology |Research in Systems Neuroscience

cortex (Yeterian et al., 2012). The interpretation pro-
posed here differs in that area 45a is considered a
unique area and that the caudal part of the PFC has
been separated out as the CLPFC network, which is
especially connected with the posterior parietal cortex
(Saleem et al., 2008a).
It should be emphasized that each of the networks is
composed of a number of architectonic areas, each of
which has its own distinctive characteristics, especially
as shown by architectonic differences. Presumably
there are also functional differences between areas
within each network, although in most cases these
have not yet been defined. In spite of these differences,
there is remarkable consistency in the connections of
areas within a network, particularly in relation to con-
nections with regions outside the PFC. Such consis-
tency appears to indicate that each network as a whole
is involved in the same general function (e.g., sensory-
motor integration vs. self-referential functions), although
individual areas may function in relation to different
aspects of that general function (see below for further
discussion).
The absence of a clear laminar organization in the
pattern of retrogradely labeled cells is initially surpris-
ing, because of previous reports of differential laminar
patterns in projections between prefrontal areas, espe-
cially between areas postulated to be at different hier-
archical levels (Barbas and Rempel-Clower, 1997;
Rempel-Clower and Barbas, 2000). Several factors may
explain this discrepancy, however. The most striking
patterns previously reported were in the lamination of
axonal terminations. Almost all of the experiments
reported here used retrograde tracers, so we cannot
make any statement about laminar patterns of axonal
termination. In addition, we did not make counts of
cells in different layers, so it is possible that more
subtle quantitative differences in lamination were over-
looked. Finally, most of the connections reported in this
study were between areas that would have been con-
sidered to be at higher levels, so differences may not
be as clearcut.
VLPFC networkThe VLPFC, which extends from the ventral bank of
the principal sulcus onto the ventrolateral convexity,
has extrinsic connections that are similar to those of
the orbital network areas. These connections are gener-
ally with cortical regions that are involved in sensory
processing or integration (Fig. 22). Thus, there are
strong connections with the dysgranular insula (Id) and
the frontal opercular area (OPf), which in some cases
extend caudally into areas PV and SII in the lateral sul-
cus. These areas are related to tactile, somatic sensory,
and visceral afferent processing in primates (Robinson
and Burton, 1980; Cusick et al., 1989; Krubitzer et al.,
1995; Burton et al., 1995, 2008; Disbrow et al., 2000;
Kaas and Collins, 2003; Fitzgerald et al., 2004; Eickhoff
et al., 2010). Other connections are with the ventral
bank/fundus of the superior temporal sulcus (STSv/f),
and the inferior temporal area TE that are involved in
the visual discrimination and recognition of objects and
faces (Tanaka, 1996; Janssen et al., 2000; Tsao et al.,
2003, 2008a; Op de Beeck et al., 2008; Ungerleider
and Bell, 2011; Kravitz et al., 2013). On the motor side,
there are connections to the ventral premotor area F5
and to a lesser extent with the dorsal premotor areas
F6/F7 (area F7 corresponds to the supplementary eye
field) and with parietal areas 7b (PFG/PF) and AIP (see
also Borra et al., 2011; Gerbella et al., 2013). These
areas are themselves interconnected and appear to
function in somatosensory, visual, and motor interac-
tions for goal-directed motor acts and sensory-motor
transformations for grasping of objects (Jennerod et al.,
1995; Sakata et al., 1995; Murata et al., 2000; Rozzi
et al., 2008; Kravitz et al., 2011). Connections were not
found between the VLPFC and posterior parietal areas
LIP and 7a, although relatively strong connections were
found between these areas and the CLPFC (Saleem
et al., 2008a).
Connections of areas 12r, 12l, and 46vAlthough the visually related inputs to the VLPFC (12r,
12l, and 46v) originate from the inferotemporal cortex
(IT), the present data indicate that these projections do
not arise equally from all parts of IT or terminate evenly
throughout the VLPFC. Projections to areas 12r, 12l, and
46v all arise in the ventral bank and fundus of the rostral
STS (STSv/f). In contrast, the rostral or caudal subre-
gions of TE (areas TEad, TEav, TEpd, and TEpv; see
Table 3) project primarily to area 12l. This difference pre-
sumably reflects functional distinctions. For example,
Janssen et al. (2000) found that, whereas neurons in
both STSv and TE (TEad) respond to and are selective
for two-dimensional (2D) shapes, many more of the neu-
rons in STSv are selective for 3D shape than in area
TEad.
Somatic sensory inputs terminate throughout the
VLPFC but have a relatively diverse origin. The most
obviously somatic sensory projections are from areas
PV and SII, areas that have been clearly established as
responding to somatic sensory stimuli, with connections
to primary somatic sensory cortex or thalamus (see,
e.g., Robinson and Burton, 1980; Krubitzer et al.,
1995). The strongest connection to the VLPFC, how-
ever, is with areas Id and OPf, rostral to areas SII and
PV. The nature of somatic sensory information
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1677
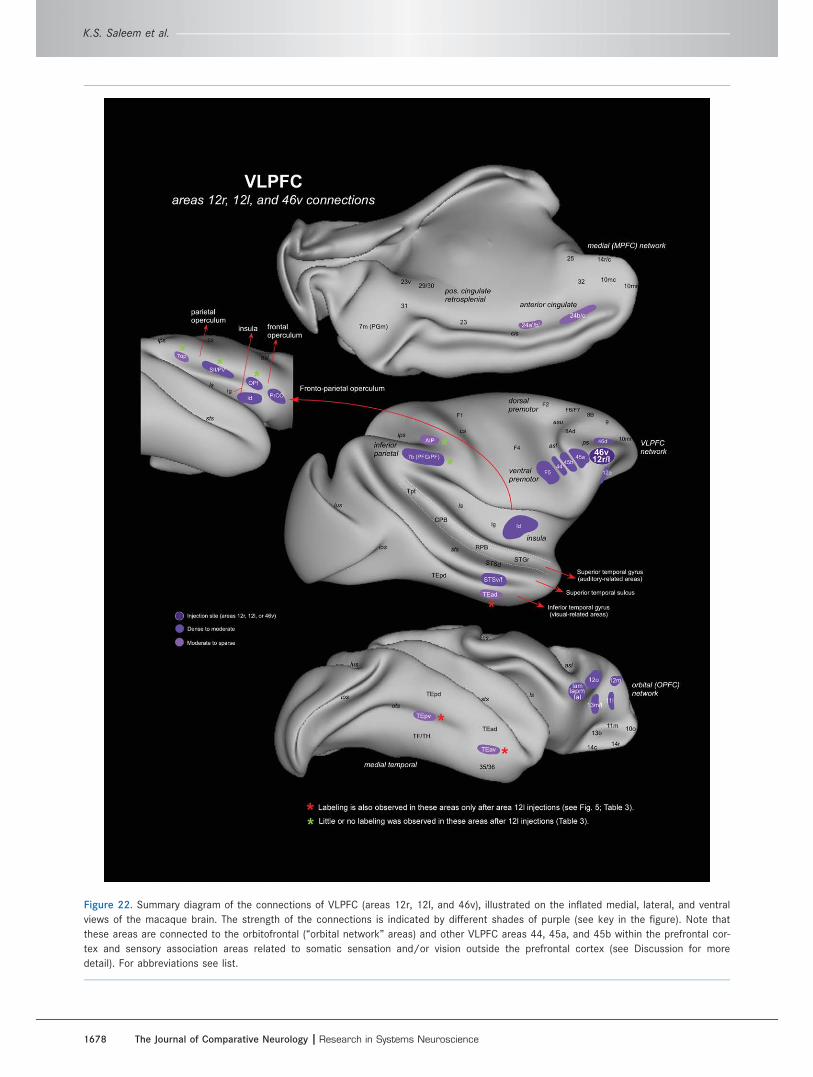
Figure 22. Summary diagram of the connections of VLPFC (areas 12r, 12l, and 46v), illustrated on the inflated medial, lateral, and ventral
views of the macaque brain. The strength of the connections is indicated by different shades of purple (see key in the figure). Note that
these areas are connected to the orbitofrontal (“orbital network” areas) and other VLPFC areas 44, 45a, and 45b within the prefrontal cor-
tex and sensory association areas related to somatic sensation and/or vision outside the prefrontal cortex (see Discussion for more
detail). For abbreviations see list.
K.S. Saleem et al.
1678 The Journal of Comparative Neurology |Research in Systems Neuroscience

represented is unclear because there have been few
reported physiological recordings in these areas. Many
of the studies that explored the opercular areas have
used anesthetized animals, and it is possible that neu-
rons in the OPf respond to relatively complex stimuli
and are not responsive in anesthetized animals. Various
physiological and anatomical data suggest that area Id
may process visceral afferent activity related to the
state of the body (Craig, 2009). This will be discussed
below in a section on the functional significance of the
VLPFC network.
It is notable that none of the VLPFC areas is con-
nected with auditory association cortex in the superior
temporal gyrus and supratemporal plane, except for the
special case of area 45a (see below). Even when cells
were labeled in STSd following VLPFC injections, they
were in the very medial part of the sulcus (area PGa
and TPO of Seltzer and Pandya, 1978), where visual
responses but not auditory responses have been
reported (Baylis et al., 1987).
The present findings of VLPFC connections with tem-
poral, parietal, and insular cortical areas are consistent
with previous descriptions (Barbas, 1988; Seltzer and
Pandya, 1989; Petrides and Pandya, 2002; Borra et al.,
2011; Gerbella et al., 2010, 2013; see also Cavada and
Goldman-Rakic, 1989; Rozzi et al., 2006; Borra et al.,
2008), although many of these studies used large injec-
tions that obscured the connectional patterns and
made the interpretation more difficult. As a group,
these studies have been interpreted to show a varied
set of different connections, without an attempt to
identify cortical networks.
Petrides and Pandya (2002) described the connec-
tions of the VLPFC, including areas 45, 12l (their 47/
12), and 46v (their 9/46v), as part of a series of stud-
ies on all parts of the PFC. For all of the VLPFC areas,
they reported complex multimodal inputs, which varied
with the target prefrontal area. Area 12l (47/12) was
shown to be primarily connected to STSv and anterior
area TE, whereas area 46v (9/46v) was related to areas
Id, the frontal operculum, SII, and the inferior part of
the intraparietal sulcus, in the region of areas 7b and
AIP. Both of these areas were also strongly intercon-
nected with other VLPFC areas. Area 45 was reported
to have the most complex connections, interacting with
extensive parts of the DPFC as well as the VLPFC; out-
side the PFC, its major connection was with the STGr/
STSd and auditory association areas (their Fig. 10). All
of these results are in excellent agreement with the
observations presented here (see below for a discus-
sion of area 45a).
In two recent studies, Luppino and his colleagues
examined the connections of areas 12r and 46v, with
the use of many small, well placed tracer injections
(Borra et al., 2011; Gerbella et al., 2013). Their delinea-
tion of area 12r was somewhat larger than that used in
this study, so their most caudal injections in this area
impinged on our area 12l (Borra et al., 2011). Conse-
quently, they reported that the middle portion of area
12r was related to somatic sensory/visceral afferent
areas in Id, the frontal operculum, and area SII as well
as the STSv. Conversely, the more caudal region (our
12l) was connected primarily to the visually related
regions STSv and anterior TE, without connections to
the insular or opercular areas. Interestingly, they found
that the most rostral part of area 12r has a third pat-
tern of connections, primarily with the fundus of the
STS.
Similar patterns emerged from their study of area
46v, with variation along the principal sulcus from ros-
tral to caudal (Gerbella et al., 2013). The mid- to rostral
part of 46v was shown to have its main extrinsic con-
nection (outside the PFC) with the STSv/f. In the mid-
to caudal part of 46v, this pattern of connections
shifted to the insular and opercular areas. Finally, the
most caudal part of area 46v showed a very different
pattern, with connections to the posterior STS and the
posterior parietal area LIP, areas that are related to
hand–eye interaction and attention. This pattern corre-
sponds to our results for the caudal part of area 46
that is part of the CLPFC network.
Connections of area 45aArea 45a stands out from the other parts of the VLPFC
(12r, 12l, and 46v; compare Figs. 23 and 22), in that it
has substantial intrinsic connections with areas in the
other subdivisions of the PFC (DPFC and CLPFC) as
well as with those of the VLPFC. Furthermore, the
extrinsic connections of area 45a with cortical areas
outside the PFC resemble the connections of the DPFC
network (see below). In particular, there are strong con-
nections with areas STGr/STSd, which are seen with
dorsal prefrontal areas but not with other ventrolateral
prefrontal areas. These connections extend more cau-
dally in the temporal cortex than the connections of
most of the DPFC areas (with the exception of the fron-
tal pole), to involve the auditory association areas in
the belt and parabelt region. This study has confirmed
the previous reports of connections of area 45a, both
within and outside the PFC (Petrides and Pandya 2002;
Saleem et al., 2008b; Gerbella et al., 2010). Another
study by Romanski et al. (1999a, their Fig. 9) also
agrees with the present results, although the architec-
tonic delineation of area 45 in their maps is slightly dif-
ferent from ours. In their study, a tracer injection in
area 12vl, which is anterior to area 45 in their map (but
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1679
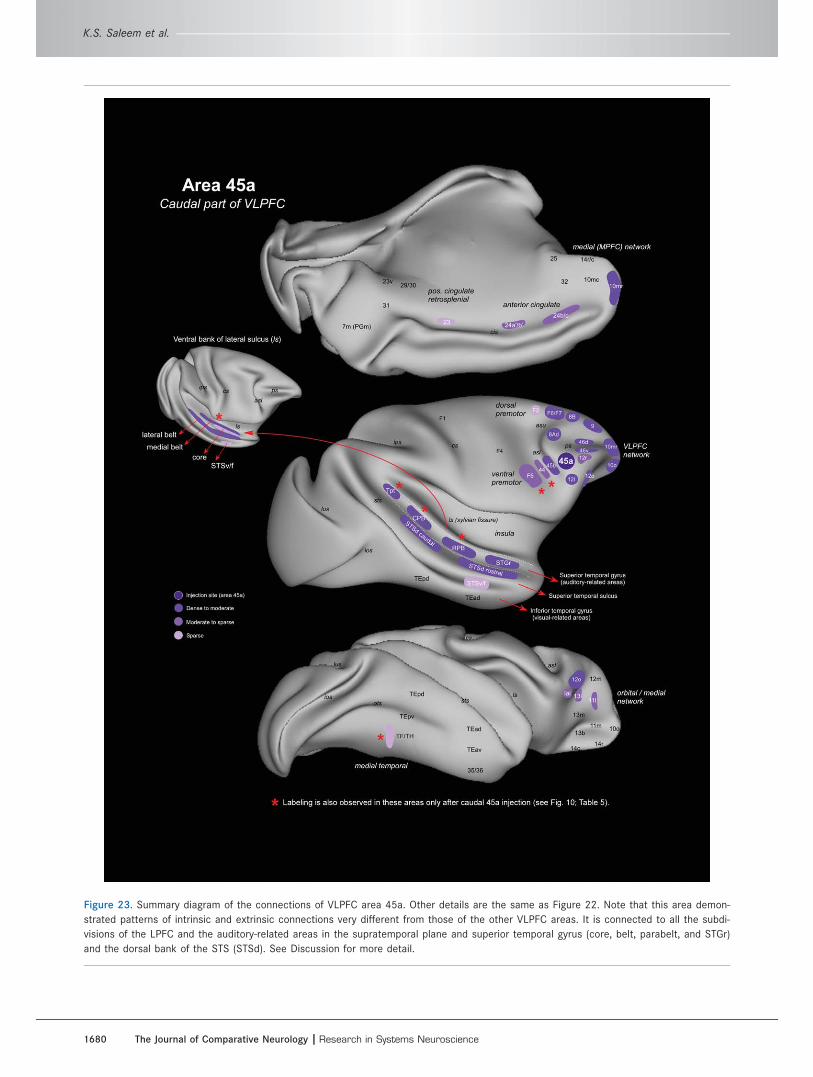
Figure 23. Summary diagram of the connections of VLPFC area 45a. Other details are the same as Figure 22. Note that this area demon-
strated patterns of intrinsic and extrinsic connections very different from those of the other VLPFC areas. It is connected to all the subdi-
visions of the LPFC and the auditory-related areas in the supratemporal plane and superior temporal gyrus (core, belt, parabelt, and STGr)
and the dorsal bank of the STS (STSd). See Discussion for more detail.
K.S. Saleem et al.
1680 The Journal of Comparative Neurology |Research in Systems Neuroscience

partially overlaps with areas 45a, 12l, and 12o on the
orbital cortex as defined in the present study) labeled
both terminals and neurons in the auditory belt and
parabelt areas as well as the upper bank of the STS.
Complementary experiments with injections in the ros-
tral belt areas of the auditory cortex labeled axon termi-
nals in areas 12vl, 45, and medial part of 12o
(Romanski et al., 1999b, see their Fig. 3e). It is impor-
tant to note that injections into area 12l (ventral to
area 45a; Fig. 5) in the present study did not label any
cells in the auditory association areas but only cells in
the inferior temporal cortex (area TE and STSv). Further-
more, an anterograde tracer injection that involved area
STGr/STSd in our previous study produced dense ter-
minal labeling in the medial part of 12o and in 45a but
very sparse labeling in 12l (Fig. 12; see also Fig. 18D,E
in Saleem et al., 2008b).
There may be further organization in the input from
superior temporal cortex to area 45a. We found that
the rostral part of area 45a receives projections mainly
from STSd, whereas the caudal part (just anterior to
the ventral limb of the arcuate sulcus) receives input
from the auditory association areas (belt and parabelt)
in STG as well as STSd. These results suggest func-
tional subdivisions within area 45a.
At least a portion of area 45a also receives inputs
from visually related areas in STSv (Fig. 12; see also
Petrides and Pandya, 2002; Saleem et al., 2008b; Ger-
bella et al., 2010). The cells in STSd that project to
area 45a also extend substantially into the medial part
(areas PGa and TPO of Seltzer and Pandya, 1978),
where visual responses have been reported (Baylis
et al., 1987). Such observations, especially the conver-
gence of visual and auditory inputs, indicate that area
45a is a functionally distinct region. Recordings that
seem to correspond to 45a have found convergent
responses to visual and auditory stimuli in individual
neurons; the auditory responses were particularly to
species-specific calls (Sugihara et al., 2006; Romanski,
2012), and visual responses to face stimuli have also
been found (Wilson et al., 1993; O’Scalaidhe et al.,
1997; Tsao et al., 2008b). It is worth noting that the
area identified by Romanski and colleagues (1999a,b;
2012) in rhesus macaques is reported to be just rostral
to area 45a. Regardless of terminology, it seems proba-
ble that these areas coincide, but this remains to be
conclusively determined.
In any case, area 45a and the region around it
appear to be involved in vocal and facial communica-
tion between monkeys. It is fascinating to speculate
that area 45a may be a precursor for the language-
related areas found in the caudal ventrolateral PFC in
humans (Kelly et al., 2010; Petrides et al., 2012).
Area 45b may be more similar to other parts of
VLPFC than area 45a. Although we do not have an
injection restricted to area 45b, experiments with injec-
tions in the temporal cortex showed that the visual
area STSv projects to 45b (Fig. 12; see also Fig. 19E of
Saleem et al., 2008b). In addition, Gerbella et al.
(2010) showed that tracer injections restricted to area
45b labeled cells in the STSv and in several parts of TE,
but not in the STGr/STSd or the auditory belt and para-
belt areas. However, based on reports of direct connec-
tions of 45b with 8A and LIP (Gerbella et al., 2010), it
is possible that further work will indicate that area 45b
is also related to CLPFC.
Functional significance of the VLPFC networkThe areas of the VLPFC network (except 45a) are inter-
connected with the orbital network (OPFC), and both
networks have a very similar pattern of extrinsic con-
nections with several sensory processing areas, includ-
ing the insular and opercular areas for somatic
sensation and inferior temporal areas for vision; in the
case of the OPFC, there are also multisensory inputs
from the perirhinal cortex (Carmichael and Price, 1995;
Kondo et al., 2005; Saleem et al., 2008b). The two net-
works are also connected with ventral premotor areas
(Carmichael and Price, 1995; Saleem et al., 2008b; cur-
rent study). It is reasonable to suggest that both net-
works have a similar function, which is related in some
way to integration of multisensory stimuli and possibly
the promotion of appropriate motor responses. The
major difference between the OPFC and the VLPFC is
that the OPFC has strong connections with the primary
olfactory cortex and the gustatory cortex (Carmichael
et al., 1994; Carmichael and Price, 1995), which are
not found with the VLPFC. Although the OPFC network
therefore appears to be involved in processing food-
related stimuli, it is more likely that the VLPFC is
related to nonfood stimuli.
In the case of the orbital network, there is now sub-
stantial data indicating that it serves to assess the rela-
tive value of sensory objects, as well as playing a
central role in reward processing (Hikosaka and Wata-
nabe, 2000; Rolls, 2000; Schultz et al., 2000; Padoa-
Schioppa and Assad, 2006; Noonan et al., 2011; Rude-
beck and Murray, 2011a). Presumably, the VLPFC plays
a similar role in object assessment. Such a role implies
that the networks receive information both about the
sensory nature of the stimuli and about their affective
quality that could serve as the basis of a judgment that
the stimuli are rewarding or aversive.
It is readily apparent that areas such as PV and SII
may provide somatic sensory information, whereas
areas STSv and TE presumably provide visual
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1681

information about objects that are to be assessed, just
as the primary olfactory and gustatory cortex provide
the OPFC with information about flavor. The source of
positive or negative affective information is less appa-
rent. Among the connections that have been identified,
the most likely source of such information is the dys-
granular cortex in the rostral and central part of the
insula. This cortical region appears to receive taste/vis-
ceral afferents that could provide information about the
state of the body (Cechetto and Saper, 1987; Saper
et al., 2002; Scott, 2011), and it has substantial limbic
connections with the amygdala (Amaral and Price,
1984; Price and Drevets, 2010). It has been suggested
that activity in the insula underlies “body sense,” or
how we “feel about ourselves” (Craig, 2009, 2011).
Such sensibility could well provide the affective signals
that are needed for analysis of value. The most basic
element of reward in a phylogenetic sense is the avail-
ability of adequate food and warmth, and to be in safe
surroundings, factors that are signaled by visceral
afferents.
There has also been a considerable amount of atten-
tion to the anterior insula as a “control” area for behav-
iors (Nelson et al., 2010; Power and Petersen, 2013). In
humans, the anterior insula is among the most fre-
quently activated regions across a wide variety of fMRI
studies, involving many task situations (Nelson, et al.,
2010). It is not clear exactly what this human area cor-
responds to in monkeys, but it is likely to include the
posterior agranular and the anterior dysgranular insular
regions. It is possible that in phylogenetically earlier
animals this region was an area in which visceral affer-
ent information was evaluated and that in primates it
has evolved into a structure for more complex repre-
sentation of bodily sensibility and wellbeing. From the
current study, it is notable that the anterior insula has
such substantial and specific connections with the
orbital and VLPFC networks.
DPFC networkThe DPFC network includes cortex on the dorsal part
of the medial hemispheric surface above the cingulate
sulcus and cortex on the dorsolateral surface, dorsal to
and including the dorsal bank of the principal sulcus. It
also includes the frontal pole. It does not include the
cortex around the caudal end of the principal sulcus or
in the rostral bank of the arcuate sulcus, however, which
is part of the CLPFC network and has axonal connec-
tions distinctly different from the DPFC network. There is
some terminological confusion between these regions, at
least in part because the region around the caudal end
of the principal sulcus has often been referred to as the
“dorsolateral” PFC or DLPFC (e.g., Levy and Goldman-
Rakic, 2000; Wallis and Miller, 2003; Fox et al., 2012);
because of this confusion, we have not used the term
“dorsolateral” in referring to the DPFC.
The axonal connections of the DPFC network are simi-
lar to those of the “medial PFC network,” including inter-
connections with the STGr/STSd in the rostral temporal
cortex, the posterior cingulate/retrosplenial cortex, and
the posterior parahippocampal cortex (Fig. 24; see also
Saleem et al., 2008b; their Fig. 22B). Furthermore, just
as different areas within the DPFC network are intercon-
nected with each other, they are also connected to
areas within the MPFC network. The major difference in
the connections of the MPFC and DPFC networks is that
the MPFC network has stronger connections with limbic
structures such as the amygdala, entorhinal cortex, and
hippocampal formation. The MPFC network also has
stronger outputs to the hypothalamus and periaqueduc-
tal gray than most parts of the DPFC, although area 9
has relatively substantial projections to these visceral
control regions (€Ong€ur et al., 1998; An et al., 1998).
Our results agree well with previous studies of the
connections of the DFPC, although the interpretations
have differed. The earliest studies with axonal tracer
methods (Barbas and Mesulam, 1985; Barbas and Pan-
dya, 1989) used relatively large injections and inter-
preted the results in terms of phylogenetic “streams”
from supposedly less well differentiated orbital and
medial “paralimbic” cortex to supposedly more devel-
oped cortex in the region of the arcuate sulcus. None-
theless, these studies identified the major targets of
the DPFC (e.g., in the rostral STG and STS region; Bar-
bas and Mesulam, 1985).
Three other studies have provided more detailed
descriptions. Petrides and Pandya (1999) described five
experiments with retrograde tracer injections in areas
8b, 9d, and 46d (including their area 9/46d) plus three
more caudal injections. The distribution of labeled cells
is very similar to that reported here. In all of the cases
with injections in the DPFC (as defined here), most of
the labeled connections within the PFC were with other
parts of the DPFC, the MPFC, and area 45a. The con-
nections outside the PFC were mainly with the posterior
cingulate cortex, the STG/STSd region, and the poste-
rior parahippocampal cortex (areas TF and TH). A sec-
ond article by Petrides and Pandya (2007) described
seven cases with large tracer injections in the rostral
PFC, the results of which again correspond closely to
the results of the present study. The major connections
of the frontal pole (area 10) and area 9 were reported
to be with the posterior cingulate cortex and with the
rostral part of the superior temporal gyrus and sulcus.
Interestingly, the only one of their cases in which the
labeling clearly extended into the auditory belt/parabelt
K.S. Saleem et al.
1682 The Journal of Comparative Neurology |Research in Systems Neuroscience
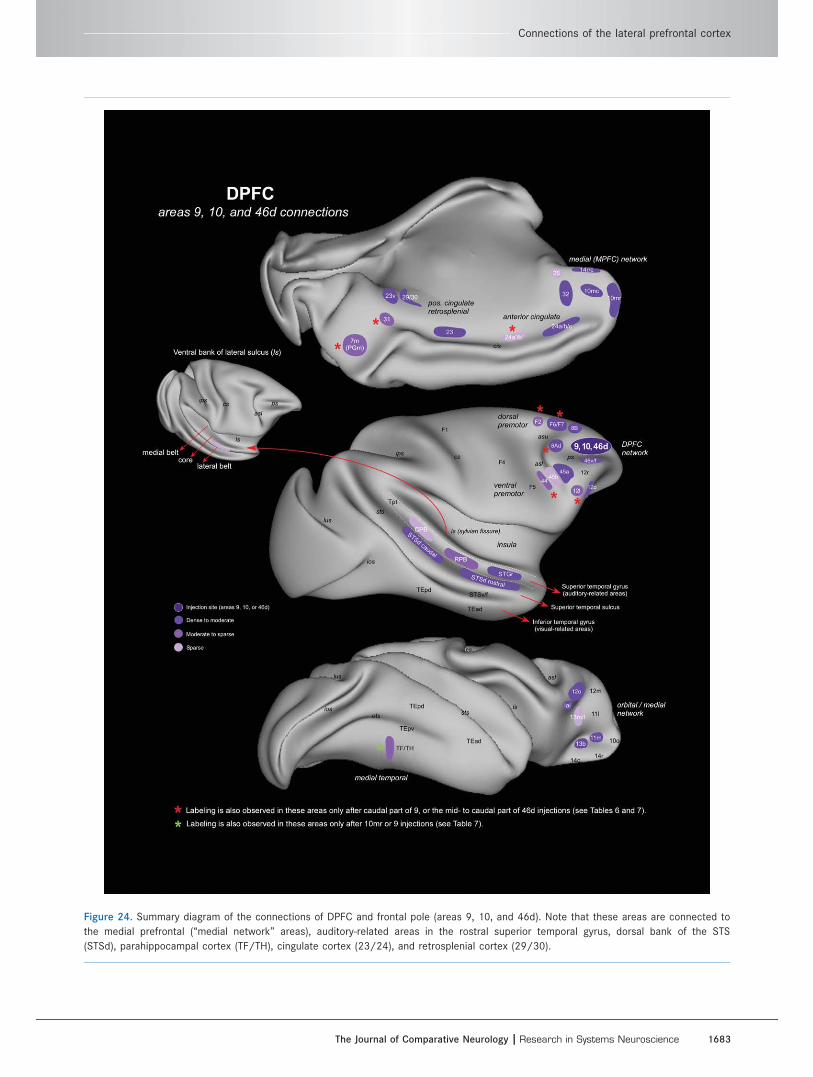
Figure 24. Summary diagram of the connections of DPFC and frontal pole (areas 9, 10, and 46d). Note that these areas are connected to
the medial prefrontal (“medial network” areas), auditory-related areas in the rostral superior temporal gyrus, dorsal bank of the STS
(STSd), parahippocampal cortex (TF/TH), cingulate cortex (23/24), and retrosplenial cortex (29/30).
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1683

(their areas TS2 and TS3) was a large injection directly
in the frontal pole (their case 1).
In a third study, Romanski et al. (1999a) analyzed
possible auditory projections to the PFC from the STG,
based on 16 experiments with tracer injections in sev-
eral parts of the PFC. The tracer used was horseradish
peroxidase (WGA-HRP, or fluorescent tracers), and many
of the injections were large, invaded the white matter,
and crossed areal boundaries, making them difficult to
interpret. Connections from the STG were identified in
11 of the cases, especially those with injections that
involved areas of the DPFC and MPFC. In eight cases,
the pattern of labeling was reported to include the para-
belt. Among these, six injections were in or near the
frontal pole (area 10o/10mr) or area 45a, areas that
were identified in the present study as connected to the
parabelt; another was in area 8A. Therefore, although it
is difficult to make a precise comparison, there is rela-
tively good agreement in the data between the Roman-
ski et al. study and the present study.
Auditory inputs to the PFCIt is generally thought that there are auditory inputs to
the PFC (see, e.g., Kaas and Hackett, 2000), although
many of the details on the precise origin and areas of
termination of such connections have been somewhat
vague. Many of these can now be filled in, but ques-
tions remain, and the functional nature of the input(s)
is still uncertain. The key points are that there are con-
nections with both defined parabelt areas and the
“auditory-related area” in the rostral superior temporal
cortex (STGr and possibly STSd) and that these possibly
different projections are distributed in distinct patterns
to different parts of the PFC.
As discussed above, there is good agreement that
the rostral parabelt projects both to the frontal pole
(area 10mr/10o) and to a region in the caudal part of
the VLPFC; in our observations and those of Gerbella
et al. (2010), this latter region corresponds to area
45a. As discussed above, area 45a is an intriguing can-
didate for a monkey communication area that might be
related to the prefrontal language areas in humans. The
role of the auditory input to the frontal pole is not
known. From perhaps the only study with awake mon-
keys that recorded specifically from the frontal pole,
Wise and his colleagues reported that neurons in the
frontal pole “encode chosen goals at feedback time
and nothing else” (Tsujimoto et al., 2010). If this is so,
auditory input would be needed if the chosen goal
includes evocation of a call or other vocalization.
Areas STGr/STSd, rostral and ventral to the auditory
belt/parabelt areas, project to area 45a and the frontal
pole but also project much more widely to the DPFC
and MPFC. The evidence linking STGr/STSd to auditory
function is much less strong and direct than for the
auditory parabelt areas. This question would seem to
depend on two additional, related questions. 1) Are
there corticocortical connections from the core, belt, or
parabelt to STGr/STSd? 2) Can reliable auditory
responses or activation be found outside the auditory
core/belt/parabelt regions? The answer to both ques-
tions appears to be a qualified yes, although the evi-
dence is not as definitive as one would like.
A study by Hackett et al. (1998) placed tracer injec-
tions in several parts of the auditory parabelt. Although
the focus of the study was connections within the
defined auditory areas, they reported “isolated patches
of labeled cells in the STS and rostral STG” following
the most rostral injections in the parabelt. In the pres-
ent study, we found that tracer injections in STGr or
the rostral parabelt produced label mainly in the
injected area but that there was overlap between the
zone labeled from the STGr and that labeled from the
rostral parabelt.
Most of the neurophysiological studies that have
recorded responses to auditory stimuli have been con-
centrated in the supratemporal plane and in belt/para-
belt areas in the middle to caudal STG. For example, a
recent study (Kikuchi et al., 2010) on responses to
complex auditory stimuli, including monkey calls, was
restricted to areas within the lateral sulcus. In part this
may be due to the difficulty of getting an electrode into
more lateral areas in unanesthetized monkeys, but it
leaves a gap in the published data on STGr/STSd.
Neurophysiological studies of STSd have mostly
stressed the polysensory nature of the region and have
referred to it as the superior temporal polysensory area
(STP). Cells in the STP respond differentially to several
modalities and are also responsive to motion-related
stimuli (Bruce et al., 1981; Heitanen and Perrett, 1993;
Oram et al., 1993; Oram and Parrett, 1996). The exact
boundaries of the STP are not well defined, but it is
said to correspond to areas TPO and PGa of Seltzer
and Pandya (1978; see also Heitanen and Perrett,
1993), which occupy the very medial part of the STSd.
Most studies on STP have also reported units in the
medial part of the STSd (see, e.g., Heitanen and Per-
rett, 1993; Oram and Perrett, 1996). The area that we
have referred as STGr/STSd occupies the more lateral
part and lip of STSd and the adjacent surface of the
STG. It is likely, therefore, that the cortical area that is
connected to MPFC and DPFC is separate from the
STP. From one of the few comprehensive studies of
this region, Baylis et al. (1987) reported responsive
neurons in many areas within the STS; almost all of the
areas responded mainly to visual stimuli, including all
K.S. Saleem et al.
1684 The Journal of Comparative Neurology |Research in Systems Neuroscience

parts of STSv/f and area TPO in the medial part of
STSd. Only the lateral part of STSd (their area TAa) and
STGr (their area TS) had substantial responses to both
visual and auditory stimuli. A particularly useful study
was reported by Poremba and Mishkin (2007), who
used the 2-deoxyglucose method of labeling metabol-
ically (and synaptically) active brain areas to compare
areas activated by auditory stimuli on one side of the
cortex with areas in the contralateral hemisphere that
had been deafferented by cutting the auditory pathways
and commissures. Their study indicates that the entire
STG is activated in the “hearing” hemisphere and deac-
tivated in the “deaf” hemisphere. A close examination
of their illustrated data, however, shows that the pri-
mary auditory area and the belt and parabelt regions
are strongly affected, whereas the STGr/STSd region is
affected to a substantially lesser degree (see their Figs.
3, 7). The activation seen in STGr/STSd was in isolated
columns; that in the core/belt/parabelt was more
continuous.
Taken together, all of these observations suggest
that STGr/STSd may not be exclusively or primarily
related to audition but may be more fully involved in a
related function, such as communication or vocalization.
In this respect, it is notable that an earlier study from
this laboratory that defined cortical projections to the
midbrain periaqueductal gray (PAG) showed that these
arise from STGr/STSd as well as from most of the
areas of the MPFC and DPFC (An et al., 1998). The
PAG and the MPFC have long been recognized as sites
where stimulation can evoke vocalization in monkeys
(for review see Jurgens, 2009). The PAG appears to
function through coordinating control of laryngeal
motorneurons and other vocalization-related nuclei in
the caudal brainstem. The PAG also coordinates behav-
ioral and autonomic responses to stress, especially pain
(Keay and Bandler, 2001), and it is likely that the role
of the PAG in vocalization is related to its general role
in relation to stress. The MPFC/DPFC appears to pro-
vide cortical modulation of this stress response (see
below), and it may be suggested that STGr/STSd con-
tributes to this function.
Functional significance of DPFC networkRelation to default mode networkTaken together, the DPFC, MPFC, and their connected
cortical areas (STGr/STSd, posterior cingulate and para-
hippocampal cortices), as defined in monkeys, closely
resemble the network identified in humans as the
“default mode network” (DMN). As analyzed with func-
tional imaging methods, the DMN is active under rest-
ing conditions and decreases its activity under most
active test conditions (Raichle et al., 2001; Raichle and
Snyder, 2007). The core regions of the DMN in humans
are the posterior cingulate cortex and adjacent precu-
neus and the medial and dorsal prefrontal cortex, which
match the combined MPFC and DPFC in monkeys, and
the posterior cingulate region to which they are con-
nected. Other cortical regions that are usually included
in the DMN are the parietal/temporal/occipital junction
region, the rostral part of the middle temporal gyrus,
and the entorhinal and parahippocampal cortices. The
only one of these regions that does not appear to cor-
respond to a part of the MPFC/DPFC network and con-
nected cortical areas in monkeys is the parietal/
temporal/occipital junction region. Although the middle
temporal gyrus region in humans does not immediately
correspond to the STGr/STSd region in monkeys, this
may be because of expansion of the temporal cortex in
humans, which produces a middle temporal gyrus that
is not present in monkeys. The visual association areas
in the inferior temporal cortex in monkeys have moved
onto the ventral surface of the temporal lobe in humans
(Denys et al., 2004; Orban et al., 2004; Tsao et al.,
2008a). It may be suggested that the middle temporal
gyrus has developed from an expansion of the cortex in
the STS, such that the STGr/STSd in monkeys is repre-
sented in the middle temporal gyrus in humans.
Although the function of the DMN is still unclear, it has
been implicated in self-referential functions, among
others (Gusnard et al., 2001; Gusnard, 2005). Mood is
a prime example of such functions, and areas within
the DMN have been frequently implicated in mood dis-
orders, especially areas in the MPFC, rostral and ventral
to the genu of the corpus callosum (Sheline et al.,
2009; Price and Drevets, 2010, 2012; Holtzheimer
et al., 2012). Petrides has presented evidence that that
the DPFC is involved in a different type of self-
referential function, namely, monitoring previous behav-
ior as a guide to subsequent choices (Petrides et al.,
2002; Petrides, 2005).
The function of the middle temporal gyrus in humans
has been unclear. Recently, however, Kanwisher and
her colleagues have reported a new method of analyz-
ing fMRI data in individual subjects that allows the
demonstration of language-related activation in an
extended strip along the middle temporal gyrus (Feder-
enko et al., 2010). This language-related strip includes
the rostral part of the gyrus that is part of the DMN
and may correspond to the STGr/STSd in monkeys.
Convergent region around principal sulcusThe three networks of the lateral PFC (DPFC, VLPFC,
and CLPFC) all extend into the principal sulcus, with
the DPFC largely in the dorsal bank, the VLPFC in the
ventral bank, and the CLPFC in the caudal part of the
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1685

sulcus. The extrinsic connections of these parts of the
sulcus are distinct, but intrinsic corticocortical connec-
tions suggest that there is considerable interaction
within the principal sulcus. Most of the local connec-
tions of PFC networks are restricted to other areas of
the same network, but there are substantial connec-
tions between the dorsal and the ventral banks of the
principal sulcus, linking the DPFC and VLPFC networks.
Similarly, experiments with injections in areas of the
CLPFC show that there are connections with more ros-
tral parts of the principal sulcus.
A recent study of neuronal responses in the dorsal
and ventral banks of the principal sulcus suggests how
this might work (Cai and Padoa-Schippo, 2014). As
mentioned above, previous work had shown that neu-
rons in the OPFC network function in assessment of
stimuli by encoding the relative value of offered
rewards (Padoa-Schioppa and Assad, 2006). Given the
similarities and connections between the OPFC and
VLPFC networks, it would be expected that neurons in
the VLPFC might show similar responses. Indeed, neu-
rons were found that respond to the relative value of
stimuli in the ventral bank of the principal sulcus.
Remarkably, the same types of responses were also
found in the dorsal bank, although at a longer average
latency (see also Kennerly et al., 2009), suggesting that
the activity was processed in the VLPFC network and
then relayed to the DPFC networks. Presumably, the
two networks use the activity for different purposes,
but this remains to be determined.
Correlation with other speciesFor rats, there is evidence for the existence of orbital
and medial PFC networks similar to those defined for
monkeys, although they are substantially less complex
(Floyd et al., 2000, 2001). The medial PFC of monkeys
has obvious correlation with medial agranular frontal
areas in rats, and several small areas in the dorsal
bank of the rostral rhinal sulcus of rats appear to corre-
spond to areas in the orbital cortex of monkeys. On the
other hand, there are considerable questions about
whether the granular areas in the lateral PFC of prima-
tes exist in rodents (Preuss, 1995, 2000). Functionally,
it has been suggested that the agranular medial PFC in
rodents has a function similar to that of much of the
granular cortex of the primate lateral PFC, but anatomi-
cally the two regions are very different. There is virtu-
ally no granular frontal cortex in rodents. The current
finding that the DPFC and VLPFC networks have similar
connections to the MPFC and OPFC networks, respec-
tively, suggests a possible solution to this question.
That is, the dorsal and ventral parts of the primate lat-
eral PFC might have developed phylogenetically by elab-
oration and expansion of the medial and orbital
networks in some precursor species. The functions rep-
resented in two networks in rodents would then be
spread over larger regions, which would develop novel
structural and functional characteristics as they become
further subdivided. In that sense, the medial PFC in
rodents would be expected to have some of the func-
tions of the lateral PFC in primates, but these would be
much less well developed and would be combined with
functions that are represented primarily by the MPFC
network in primates.
The suggestion that the DPFC has developed out of
the MPFC and the VLPFC out of the OPFC is very simi-
lar to a longstanding proposal by Pandya and his col-
leagues for dorsal and ventral streams or trends of
phylogenetic development of the PFC (see Yeterian
et al., 2012). Their “dorsal architectonic trend” corre-
sponds largely to the combined MPFC and DPFC net-
works, and their “ventral architectonic trend” is similar
to the combined OPFC and VLPFC networks. There are
several differences in the detailed definition of these
systems (for example, the MPFC network includes
some areas along the medial and lateral edges of the
orbital cortical surface), but the major difference is that
the CLPFC is included in the dorsal and ventral trends,
whereas we have separated it out as a distinct system.
A corollary of this argument is that the PFC in
humans is essentially an expansion and elaboration of
the same networks found in monkeys. This cannot be
determined at present, but it is notable that the same
architectonic areas found in the orbital and medial pre-
frontal cortices of monkeys can be identified in humans
(€Ong€ur et al., 2003), and these can be used to outline
the MPFC and OPFC networks. Petrides and colleagues
(2012) have also proposed correlated architectonic
maps for the LPFC for monkeys and humans, although
these maps are not sufficiently detailed to account for
the subdivisions defined in this study. The presence of
the DMN in the dorsal as well as ventral parts of the
MPFC in humans (see above) suggests that the MPFC
and DPFC networks are present in humans and that
they are located mostly on the medial wall and dorsal
convexity of the hemisphere. It is reasonable to expect
that the VLPFC network is located on the ventrolateral
convexity, but it is difficult to delineate it precisely. If
area 45a is indeed part of a communication system, it
is likely to have expanded substantially with the devel-
opment of language areas in the LPFC.
ACKNOWLEDGMENTSWe thank Hieu Van Luu, Abhisek C. Kandai, and Sze-
chuan Suen for their excellent technical assistance and
Colin Reveley for providing inflated brain images. We also
K.S. Saleem et al.
1686 The Journal of Comparative Neurology |Research in Systems Neuroscience

thank two anonymous reviewers for useful comments on
the manuscript.
CONFLICT OF INTEREST STATEMENT
The authors have no conflicts of interest.
ROLE OF AUTHORS
All authors were involved in this study. Study concept
and design: KSS, JLP. Acquisition of data: KSS, BM, JLP.
Analysis and interpretation of data: KSS, BM, JLP. Draft-
ing of the article: KSS, JLP.
LITERATURE CITEDAmaral DG, Price JL. 1984. Amygdalo-cortical projections in
the monkey (Macaca fascicularis). J Comp Neurol 230:465–496.
Amiez C, Petrides M. 2009. Anatomical organization of theeye fields in the human and non-human primate frontalcortex. Prog Neurobiol 89:220–230.
An X, Bandler R, €Ong€ur D, Price JL. 1998. Prefrontal cortical pro-jections to longitudinal columns in the midbrain periaqueduc-tal gray in macaque monkeys. J Comp Neurol 401:455–479.
Asaad WF, Rainer G, Miller EK. 1998. Neural activity in theprimate prefrontal cortex during associative learning.Neuron 21:1399–1407.
Barbas H. 1988. Anatomic organization of basoventral andmediodorsal visual recipient prefrontal regions in the rhe-sus monkey. J Comp Neurol 276:313–342.
Barbas H, Mesulam MM. 1981. Organization of afferent inputto subdivisions of area 8 in the rhesus monkey. J CompNeurol 200:407–431.
Barbas H, Mesulam MM. 1985. Cortical afferent input to theprincipalis region of the rhesus monkey. Neuroscience15:619–637.
Barbas H, Pandya DN. 1989. Architecture and intrinsic con-nections of the prefrontal cortex in the rhesus monkey. JComp Neurol 286:353–375.
Barbas H, Rempel-Clower N. 1997. Cortical structure predictsthe pattern of corticocortical connections. Cereb Cortex7:635–646.
Baylis CG, Rolls ET, Leonard CM. 1987. Functional subdivisionsof the temporal lobe neocortex. J Neurosci 7:330–342.
Borra E, Belmalih A, Calzavara R, Gerbella M, Murata A, RozziS, Luppino G. 2008. Cortical connections of the maca-que anterior intraparietal (AIP) area. Cereb Cortex 18:1094–1111.
Borra E, Gerbella M, Rozzi S, Luppino G. 2011. Anatomicalevidence for the involvement of the macaque ventrolat-eral prefrontal area 12r in controlling goal-directedactions. J Neurosci 31:12351–12363.
Bruce, CJ, Goldberg ME. 1985. Primate frontal eye fields. I.Single neurons discharging before saccades. J Neurophy-siol 53:603–635.
Bruce C, Desimone R, Gross CG. 1981. Visual properties ofneurons in a polysensory area in superior temporal sul-cus of the macaque. J Neurophysiol 46:369–384.
Bruce CJ, Goldberg ME, Bushnell MC, Stanton GB. 1985. Pri-mate frontal eye fields. II. Physiological and anatomicalcorrelates of electrically evoked eye movements. J Neu-rophysiol 54:714–734.
Burton H, Fabri M, Alloway K. 1995. Cortical areas within thelateral sulcus connected to cutaneous representations inareas 3b and 1: a revised interpretation of the second
somatosensory area in macaque monkeys. J Comp Neu-rol 355:539–562.
Burton H, Sinclair RJ, Wingert JR, Dierker DL. 2008. Multipleparietal operculum subdivisions in humans: tactile activa-tion maps. Somatosens Mot Res 25:149–162.
Cadoret G, Petrides M. 2007. Ventrolateral prefrontal neuro-nal activity related to active controlled memory retrievalin nonhuman primates. Cereb Cortex 17:i27–i40.
Cai X, Padoa-Schioppa C. 2014. Contributions of orbitofrontaland lateral prefrontal cortices to economic choice andthe good-to-action transformation. Neuron, In press.
Campbell MJ, Morrison JH. 1989. Monoclonal antibody to neu-rofilament protein (SMI-32) labels a subpopulation ofpyramidal neurons in the human and monkey neocortex.J Comp Neurol 282:191–205.
Carmichael ST, Price JL. 1994. Architectonic subdivision ofthe orbital and medial prefrontal cortex in the macaquemonkey. J Comp Neurol 346:366–402.
Carmichael ST, Price JL. 1995. Sensory and premotor connec-tions of the orbital and medial prefrontal cortex of maca-que monkeys. J Comp Neurol 363:642–664.
Carmichael ST, Price JL. 1996. Connectional networks withinthe orbital and medial prefrontal cortex of macaquemonkeys. J Comp Neurol 371:179–207.
Carmichael ST, Clugnet M-C, Price JL. 1994. Central olfactoryconnections in the macaque monkey. J Comp Neurol346:403–434.
Cavada C, Goldman-Rakic PS. 1989. Posterior parietal cortexin rhesus monkey: II. Evidence for segregated cortico-cortical networks linking sensory and limbic areas withthe frontal lobe. J Comp Neurol 287:422–445.
Cechetto DF, Saper CB. 1987. Evidence for a viscerotopicsensory representation in the cortex and thalamus in therat. J Comp Neurol 262:27–45.
Chavis DA, Pandya DN. 1976. Further observations on cortico-frontal connections in the rhesus monkey. Brain Res117:369–386.
Craig AD. 2009. How do you feel—now? The anterior insulaand human awareness. Nat Rev Neurosci 10:59–70.
Craig AD. 2011. Significance of the insula for the evolution ofhuman awareness of feelings from the body. Ann N YAcad Sci 1225:72–82.
Cusick CG, Wall JT, Felleman DJ, Kaas JH. 1989. Somatotopicorganization of the lateral sulcus of owl monkeys: area3b, S-II, and a ventral somatosensory area. J Comp Neu-rol 282:169–190.
Denys K, Vanduffel W, Fize D, Nelissen K, Peuskens H, VanEssen D, Orban GA. 2004. The processing of visualshape in the cerebral cortex of human and nonhumanprimates: a functional magnetic resonance imagingstudy. J Neurosci 24:2551–2565.
Disbrow E, Roberts T, Krubitzer L. 2000. Somatotopic organi-zation of cortical fields in the lateral sulcus of Homosapiens: evidence for SII and PV. J Comp Neurol 418:1–21.
Drevets WC, Price JL, Simpson JR, Todd RD, Reich T, VannierM, Raichle ME. 1997. Subgenual prefrontal cortex abnor-malities in mood disorders. Nature 386:824–827.
Drevets WC, €Ong€ur D, Price JL. 1998. Neuroimaging abnormal-ities in the subgenual prefrontal cortex: implications forthe pathophysiology of familial mood disorders. Mol Psy-chiatry 3:220–226.
Eickhoff SB, Jbabdi S, Caspers S, Laird AR, Fox PT, Zilles K,Behrens TE. 2010. Anatomical and functional connectiv-ity of cytoarchitectonic areas within the human parietaloperculum. J Neurosci 30:6409–6421.
Fedorenko E, Hsieh PJ, Nieto-Casta~n�on A, Whitfield-Gabrieli S,Kanwisher N. 2010. New method for fMRI investigations
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1687

of language: defining ROIs functionally in individual sub-jects. J Neurophysiol 104:1177–1194.
Ferry AT, €Ong€ur D, An X, Price JL. 2000. Prefrontal corticalprojections to the striatum in macaque monkeys: evi-dence for an organization related to prefrontal networks.J Comp Neurol 425:447–470.
Fitzgerald PJ, Lane JW, Thakur PH, Hsiao SS. 2004. Receptivefield properties of the macaque second somatosensorycortex: evidence for multiple functional representations. JNeurosci 24:11193–11204.
Floyd NS, Price JL, Ferry AT, Keay KA, Bandler R. 2000. Orbi-tomedial prefrontal cortical projections to distinct longi-tudinal columns of the periaqueductal gray in the rat. JComp Neurol 422:556–578.
Floyd NS, Price JL, Ferry AT, Keay KA, Bandler R. 2001. Orbi-tomedial prefrontal cortical projections to hypothalamusin the rat. J Comp Neurol 432:307–328.
Fox MD, Liu H, Pascual-Leone A. 2012. Identification of repro-ducible individualized targets for treatment of depressionwith TMS based on intrinsic connectivity. Neuroimage66C:151–160.
Gamlin PD, Yoon K. 2000. An area for vergence eye move-ment in primate frontal cortex. Nature 407:1003–1007.
Gerbella M, Belmalih A, Borra E, Rozzi S, Luppino G. 2007.Multimodal architectonic subdivision of the caudal ven-trolateral prefrontal cortex of the macaque monkey.Brain Struct Funct 212:269–301.
Gerbella M, Belmalih A, Borra E, Rozzi S, Luppino G. 2010.Cortical connections of the macaque caudal ventrolateralprefrontal areas 45A and 45B. Cereb Cortex 20:141–168.
Gerbella M, Borra E, Tonelli S, Rozzi S, Luppino G. 2013. Con-nectional heterogeneity of the ventral part of the maca-que area 46. Cereb Cortex 23:967–987.
Goldstein ME, Sternberger LA, Sternberger NH. 1987. Varyingdegrees of phosphorylation determine microheterogene-ity of the heavy neurofilament polypeptide (Nf-H). J Neu-roimmunol 14:135–148.
Gusnard DA. 2005. Being a self: considerations from func-tional imaging. Conscious Cogn 14:679–697.
Gusnard DA, Akbudak E, Shulman GL, Raichle ME. 2001.Medial prefrontal cortex and self-referential mental activ-ity: relation to a default mode of brain function. ProcNatl Acad Sci U S A 98:4259–4264.
Haber SN, Fudge JL, McFarland NR. 2000. Striatonigrostriatalpathways in primates form an ascending spiral from theshell to the dorsolateral striatum. J Neurosci 20:2369–2382.
Hackett TA, Stepniewska I, Kaas JH. 1998. Subdivisions ofauditory cortex and ipsilateral cortical connections of theparabelt auditory cortex in macaque monkeys. J CompNeurol 394:475–495.
Hietanen JK, Perrett DI. 1993. Motion sensitive cells in themacaque superior temporal polysensory area I. Lack ofresponse to the sight of the animal’s own limb move-ment. Exp Brain Res 93:117–128.
Hikosaka K, Watanabe M. 2000. Delay activity of orbital andlateral prefrontal neurons of the monkey varying with dif-ferent rewards. Cereb Cortex 10:263–271.
Hof PR, Morrison JH. 1995. Neurofilament protein definesregional patterns of cortical organization in the macaquemonkey visual system: a quantitative immunohistochemi-cal analysis. J Comp Neurol 352:161–186.
Holtzheimer PE, Kelley ME, Gross RE, Filkowski MM, GarlowSJ, Barrocas A, Wint D, Craighead MC, Kozarsky J,Chismar R, Moreines JL, Mewes K, Posse PR, GutmanDA, Mayberg HS. 2012. Subcallosal cingulate deep brainstimulation for treatment-resistant unipolar and bipolardepression. Arch Gen Psychiatry 69:150–158.
Hsu DT, Price JL. 2007. Midline and intralaminar thalamic con-nections with the orbital and medial prefrontal networksin macaque monkeys. J Comp Neurol 504:89–111.
Janssen P, Vogels R, Orban GA. 2000. Selectivity for 3Dshape that reveals distinct areas within macaque inferiortemporal cortex. Science 288:2054–2056.
Jeannerod M, Arbib MA, Rizzolatti G, Sakata H. 1995. Grasp-ing objects: the cortical mechanisms of visuomotortransformation. Trends Neurosci 18:314–320.
Jones EG, Hendry SH. 1989. Differential calcium binding pro-tein immunoreactivity distinguishes classes of relay neu-rons in monkey thalamic nuclei. Eur J Neurosci 1:222–246.
Jones EG, Powell TP. 1970. An anatomical study of convergingsensory pathways within the cerebral cortex of the mon-key. Brain 93:793–820.
J€urgens U. 2009. The neural control of vocalization in mam-mals: a review. J Voice 23:1–10.
Kaas JH, Collins CE. 2003. The organization of somatosensorycortex in anthropoid primates. Adv Neurol 93:57–67.
Kaas JH, Hackett TA. 2000. Subdivisions of auditory cortexand processing streams in primates. Proc Natl Acad SciU S A 97:11793–11799.
Kawamura K, Naito J. 1984. Corticocortical projections to theprefrontal cortex in the rhesus monkey investigated withhorseradish peroxidase techniques. Neurosci Res 1:89–103.
Keay KA, Bandler R. 2001. Parallel circuits mediating distinctemotional coping reactions to different types of stress.Neurosci Biobehav Rev 25:669–678.
Kelly C, Uddin LQ, Shehzad Z, Margulies DS, Castellanos FX,Milham MP, Petrides M. 2010. Broca’s region: linkinghuman brain functional connectivity data and non-humanprimate tracing anatomy studies. Eur J Neurosci 32:383–398.
Kennerley SW, Dahmubed AF, Lara AH, Wallis JD. 2009. Neu-rons in the frontal lobe encode the value of multipledecision variables. J Cogn Neurosci 21:1162–1178.
Kikuchi Y, Horwitz B, Mishkin M. 2010. Hierarchical auditoryprocessing directed rostrally along the monkey’s supra-temporal plane. J Neurosci 30:13021–13030.
Kondo H, Saleem KS, Price JL. 2003. Differential connectionsof the temporal pole with the orbital and medial prefron-tal networks in macaque monkeys. J Comp Neurol 465:499–523.
Kondo H, Saleem KS, Price JL. 2005. Differential connectionsof the perirhinal and parahippocampal cortex with theorbital and medial prefrontal networks in macaque mon-keys. J Comp Neurol 493:479–509.
Kravitz DJ, Saleem KS, Baker CI, Mishkin M. 2011. A new neu-ral framework for visuospatial processing. Nat Rev Neu-rosci 12:217–230.
Kravitz DJ, Saleem KS, Baker CI, Ungerleider LG, Mishkin M.2013. The ventral visual pathway: an expanded neuralframework for the processing of object quality. TrendsCogn Sci 17:26–49.
Krubitzer L, Clarey J, Tweedale R, Elston G, Calford M. 1995.A redefinition of somatosensory areas in the lateral sul-cus of macaque monkeys. J Neurosci 15:3821–3839.
Kuypers HG, Szwarcbart MK, Mishkin M, Rosvold HE. 1965.Occipitotemporal corticocortical connections in the rhe-sus monkey. Exp Neurol 11:245–262.
Levy R, Goldman-Rakic PS. 2000. Segregation of workingmemory functions within the dorsolateral prefrontal cor-tex. Exp Brain Res 133:23–32.
Mayberg HS, Liotti M, Brannan SK, McGinnis S, Mahurin RK,Jerabek PA, Silva JA, Tekell JL, Martin CC, Lancaster JL,Fox PT. 1999. Reciprocal limbic–cortical function and
K.S. Saleem et al.
1688 The Journal of Comparative Neurology |Research in Systems Neuroscience

negative mood: converging PET findings in depressionand normal sadness. Am J Psychiatry 156:675–682.
Mayberg HS, Lozano AM, Voon V, McNeely HE, SeminowiczD, Hamani C, Schwalb JM, Kennedy SH. 2005. Deepbrain stimulation for treatment-resistant depression. Neu-ron 45:651–660.
Meyer T, Qi X, Stanford TR, Constantinidis C. 2011. Stimulusselectivity in dorsal and ventral prefrontal cortex aftertraining in working memory tasks. J Neurosci 31:6266–6276.
Miller EK. 2000. The prefrontal cortex and cognitive control.Nat Rev Neurosci 1:59–65.
Mishkin M, Manning FJ. 1978. Non-spatial memory after selec-tive prefrontal lesions in monkeys. Brain Res 143:313–323.
Monosov IE, Thompson KG. 2009. Frontal eye field activityenhances object identification during covert visualsearch. J Neurophysiol 102:3656–3672.
Murata A, Gallese V, Luppino G, Kaseda M, Sakata H. 2000.Selectivity for the shape, size, and orientation of objectsfor grasping in neurons of monkey parietal area AIP. JNeurophysiol 83:2580–2601.
Nelson SM, Dosenbach NU, Cohen AL, Wheeler ME,Schlaggar BL, Petersen SE. 2010. Role of the anteriorinsula in task-level control and focal attention. BrainStruct Funct 214:669–680.
Noonan MP, Mars RB, Rushworth MFS. 2011. Distinct roles ofthree frontal cortical areas in reward-guided behavior. JNeurosci 31:14399–14412.
€Ong€ur D, Price JL. 2000. The organization of networks withinthe orbital and medial prefrontal cortex of rats, monkeysand humans. Cereb Cortex 10:206–219.
€Ong€ur D, An X, Price JL. 1998. Prefrontal cortical projectionsto the hypothalamus in macaque monkeys. J Comp Neu-rol 401:480–505.
€Ong€ur D, Ferry AT, Price JL. 2003. Architectonic subdivision ofthe human orbital and medial prefrontal cortex. J CompNeurol 460:425–449.
Op de Beeck HP, Dicarlo JJ, Goense JB, Grill-Spector K,Papanastassiou A, Tanifuji M, Tsao DY. 2008. Fine-scalespatial organization of face and object selectivity in thetemporal lobe: do functional magnetic resonance imag-ing, optical imaging, and electrophysiology agree? J Neu-rosci 28:11796–11801.
Oram MW, Perrett DI. 1996. Integration of form and motionin the anterior superior temporal polysensory area(STPa) of the macaque monkey J Neurophysiol 76:109–129.
Oram MW, Perrett DI, Hietanen JK. 1993. Directional tuning ofmotion-sensitive cells in the anterior superior temporalpolysensory area of the macaque. Exp Brain Res 97:274–294.
Orban GA, Van Essen D, Vanduffel W. 2004. Comparativemapping of higher visual areas in monkeys and humans.Trends Cogn Sci 8:315–324.
O’Scalaidhe SP, Wilson FA, Goldman-Rakic PS. 1997. Arealsegregation of face-processing neurons in prefrontal cor-tex. Science 278:1135–1138.
Padoa-Schioppa C, Assad JA. 2006. Neurons in the orbitofron-tal cortex encode economic value. Nature 441:223–226.
Pandya DN, Kuypers HG. 1969. Cortico-cortical connectionsin the rhesus monkey. Brain Res 13:13–36.
Passingham RE, Wise SP. 2012. The neurobiology of the pre-frontal cortex: anatomy, evolution, and the origin ofinsight. London: Oxford University Press.
Passingham RE, Toni I, Rushworth MF. 2000. Specializationwithin the prefrontal cortex: the ventral prefrontal cortexand associative learning. Exp Brain Res 133:103–113.
Peng X, Sereno ME, Silva AK, Lehky SR, Sereno AB. 2008.Shape selectivity in primate frontal eye field. J Neurophy-siol 100:796–814.
Petrides M. 2005. Lateral prefrontal cortex: architectonic andfunctional organization. Philos Trans R Soc Lond B BiolSci 360:781–795.
Petrides M, Pandya DN. 1999. Dorsolateral prefrontal cortex:comparative cytoarchitectonic analysis in the human andthe macaque brain and corticocortical connection pat-terns. Eur J Neurosci 11:1011–1036.
Petrides M, Pandya DN. 2002. Comparative cytoarchitectonicanalysis of the human and the macaque ventrolateralprefrontal cortex and corticocortical connection patternsin the monkey. Eur J Neurosci 16:291–310.
Petrides M, Pandya DN. 2006. Efferent association pathwaysoriginating in the caudal prefrontal cortex in the maca-que monkey. J Comp Neurol 498:227–251.
Petrides M, Pandya DN. 2007. Efferent association pathwaysfrom the rostral prefrontal cortex in the macaque mon-key. J Neurosci 27:11573–11586.
Petrides M, Alivisatos B, Frey S. 2002. Differential activationof the human orbital, mid-ventrolateral, and mid-dorsolateral prefrontal cortex during the processingof visual stimuli. Proc Natl Acad Sci U S A 99:5649–5654.
Petrides M, Tomaiuolo F, Yeterian EH, Pandya DN. 2012. Theprefrontal cortex: comparative architectonic organizationin the human and the macaque monkey brains. Cortex48:46–57.
Poremba A, Mishkin M. 2007. Exploring the extent and func-tion of higher-order auditory cortex in rhesus monkeys.Hear Res 229:14–23.
Power JD, Petersen SE. 2013. Control-related systems in thehuman brain. Curr Opin Neurobiol 23:223–228.
Power JD, Cohen AL, Nelson SM, Wig GS, Barnes KA, ChurchJA, Vogel AC, Laumann TO, Miezin FM, Schlaggar BL,Petersen SE. 2011. Functional network organization ofthe human brain. Neuron 72:665–678.
Preuss TM. 1995. Do rats have prefrontal cortex? The Rose-Woolsey-Akert program reconsidered. J Cogn Neurosci 7:1–24.
Preuss TM. 2000. Taking the measure of diversity: compara-tive alternatives to the model-animal paradigm in corticalneuroscience. Brain Behav Evol 55:287–299.
Preuss TM, Goldman-Rakic PS. 1991. Myelo- and cytoarchitec-ture of the granular frontal cortex and surrounding regionsin the strepsirhine primate Galago and the anthropoid pri-mate Macaca. J Comp Neurol 310:429–474.
Price JL, Drevets WC. 2010. Neurocircuitry of mood disorders.Neuropsychopharmacology 35:192–216.
Price JL, Drevets WC. 2012. Neural circuits underlying thepathophysiology of mood disorders. Trends Cogn Sci 16:61–71.
Raichle ME, Snyder AZ. 2007. A default mode of brain func-tion: a brief history of an evolving idea. Neuroimage 37:1083–1090.
Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, GusnardDA, Shulman GL. 2001. A default mode of brain function.Proc Natl Acad Sci U S A 98:676–682.
Rempel-Clower NL, Barbas H. 2000. The laminar pattern ofconnections between prefrontal and anterior temporalcortices in the rhesus monkey is related to corticalstructure and function. Cereb Cortex 10:851–865.
Robinson CJ, Burton H. 1980. Somatotopographic organizationin the second somatosensory area of M. fascicularis. JComp Neurol 192:43–67.
Rolls ET. 2000. The orbitofrontal cortex and reward. CerebCortex 10:284–294.
Connections of the lateral prefrontal cortex
The Journal of Comparative Neurology | Research in Systems Neuroscience 1689

Romanski LM. 2012. Integration of faces and vocalizations inventral prefrontal cortex: implications for the evolution ofaudiovisual speech. Proc Natl Acad Sci U S A 109:10717–10724.
Romanski LM, Bates JF, Goldman-Rakic PS. 1999a. Auditorybelt and parabelt projections to the prefrontal cortex inthe rhesus monkey. J Comp Neurol 403:141–157.
Romanski LM, Tian B, Fritz J, Mishkin M, Goldman-Rakic PS,Rauschecker JP. 1999b. Dual streams of auditory affer-ents target multiple domains in the primate prefrontalcortex. Nat Neurosci 2:1131–1136.
Rozzi S, Calzavara R, Belmalih A, Borra E, Gregoriou GG,Matelli M, Luppino G. 2006. Cortical connections of theinferior parietal cortical convexity of the macaque mon-key. Cereb Cortex 16:1389–1417.
Rozzi S, Ferrari PF, Bonini L, Rizzolatti G, Fogassi L. 2008.Functional organization of inferior parietal lobule convex-ity in the macaque monkey: electrophysiological charac-terization of motor, sensory and mirror responses andtheir correlation with cytoarchitectonic areas. Eur J Neu-rosci 28:1569–1588.
Rudebeck PH, Murray EA. 2011a. Balkanizing the primateorbitofrontal cortex: distinct subregions for comparingand contrasting values. Ann N Y Acad Sci 1239:1–13.
Rudebeck PH, Murray EA. 2011b. Dissociable effects of sub-total lesions within the macaque orbital prefrontal cortexon reward-guided behavior. J Neurosci 31:10569–10578.
Rushworth MFG, Buckley MJ, Gough PM, Alexander IH,Kyriazis D, McDonald KR, Passingham RE. 2005. Atten-tional selection and action selection in the ventral andorbital prefrontal cortex. J Neurosci 25:11628–11636.
Sakata H, Taira M, Murata A, Mine S. 1995. Neural mecha-nisms of visual guidance of hand action in the parietalcortex of the monkey. Cereb Cortex 5:429–438.
Saleem KS, Logothetis NK. 2012. A combined MRI and histol-ogy atlas of the rhesus monkey brain in stereotaxic coor-dinates. 2nd edition with horizontal, coronal, and sagittalseries. San Diego: Academic Press/Elsevier.
Saleem KS, Suzuki W, Tanaka K, Hashikawa T. 2000. Connec-tions between anterior inferotemporal cortex and supe-rior temporal sulcus regions in the macaque monkey. JNeurosci 20:5083–5101.
Saleem KS, Price JL, Hashikawa T. 2007. Cytoarchitectonicand chemoarchitectonic subdivisions of the perirhinaland parahippocampal cortices in macaque monkeys. JComp Neurol 500:973–1006.
Saleem KS, Miller B, Kandai AC, Suen SC, Price JL. 2008a.Distinct connectional systems in the lateral prefrontalcortex of macaque monkeys. Soc Neurosci 465.12.
Saleem KS, Kondo H, Price JL. 2008b. Complementary circuitsconnecting the orbital and medial prefrontal networkswith the temporal, insular, and opercular cortex in themacaque monkey. J Comp Neurol 506:659–693.
Saper CB, Chou TC, Elmquist JK. 2002. The need to feed:homeostatic and hedonic control of eating. Neuron 36:199–211.
Schall JD. 2004. On the role of frontal eye field in guidingattention and saccades. Vis Res 44:1453–1467.
Schultz W, Tremblay L, Hollerman JR. 2000. Reward process-ing in primate orbitofrontal cortex and basal ganglia.Cereb Cortex 10:272–284.
Scott TR. 2011. Taste as a basis for body wisdom. PhysiolBehav 104:57–63.
Seltzer B, Pandya DN. 1978. Afferent cortical connections andarchitectonics of the superior temporal sulcus and surround-ing cortex in the rhesus monkey. Brain Res 149:1–24.
Seltzer B, Pandya DN. 1989. Frontal lobe connections of thesuperior temporal sulcus in the rhesus monkey. J CompNeurol 281:97–113.
Sheline YI, Barch DM, Price JL, Rundle MM, Vaishnavi SN,Snyder AZ, Mintun MA, Wang S, Coalson RS, Raichle ME.2009. The default mode network and self-referentialprocesses in depression. Proc Natl Acad Sci U S A 106:1942–1947.
Shiwa T. 1987. Corticocortical projections to the monkey tem-poral lobe with particular reference to the visual process-ing pathways. Arch Ital Biol 125:139–154.
Sternberger LA, Sternberger NH. 1983. Monoclonal antibodiesdistinguish phosphorylated and nonphosphorylated formsof neurofilaments in situ. Proc Natl Acad Sci U S A 80:6126–6130.
Sugihara T, Diltz MD, Averbeck BB, Romanski LM. 2006. Inte-gration of auditory and visual communication informationin the primate ventrolateral prefrontal cortex. J Neurosci26:11138–11147.
Szabo J, Cowan WM. 1984. A stereotaxic atlas of the brain ofthe cynomolgus monkey (Macaca fascicularis). J CompNeurol 222:265–300.
Tanaka K. 1996. Inferotemporal cortex and object vision.Annu Rev Neurosci 19:109–139.
Tehovnik EJ, Sommer MA, Chou IH, Slocum WM, Schiller PH.2000. Eye fields in the frontal lobes of primates. BrainRes Rev 32:413–448.
Tsao DY, Freiwald WA, Knutsen TA, Mandeville JB, Tootell RB.2003. Faces and objects in macaque cerebral cortex.Nat Neurosci 6:989–995.
Tsao DY, Moeller S, Freiwald WA. 2008a. Comparing facepatch systems in macaques and humans. Proc Natl AcadSci U S A 105:19514–19519.
Tsao DY, Schweers N, Moeller S, Freiwald WA. 2008b.Patches of face-selective cortex in the macaque frontallobe. Nat Neurosci 11:877–879.
Tsujimoto S, Genovesio A, Wise SP. 2010. Evaluating self-generated decisions in frontal pole cortex of monkeys.Nat Neurosci 13:120–126.
Ungerleider LG, Bell AH. 2011. Uncovering the visual“alphabet”: advances in our understanding of object per-ception. Vis Res 51:782–799.
Ungerleider LG, Gaffan D, Pelak VS. 1989. Projections frominferior temporal cortex to prefrontal cortex via the unci-nate fascicle in rhesus monkeys. Exp Brain Res 76:473–484.
Walker AE. 1940. A cytoarchitectural study of theprefrontal area of the macaque monkey. J. Comp Neurol73:59–86.
Wallis JD, Miller EK. 2003. Neuronal activity in primate dorso-lateral and orbital prefrontal cortex during performanceof a reward preference task. Eur J Neurosci 18:2069–2081.
Webster MJ, Bachevalier J, Ungerleider LG. 1994. Connectionsof inferior temporal areas TEO and TE with parietal andfrontal cortex in macaque monkeys. Cereb Cortex 4:470–483.
Wilson FA, O’Scalaidhe SP, Goldman-Rakic PS. 1993. Dissoci-ation of object and spatial processing domains in pri-mate prefrontal cortex. Science 260:1955–1958.
Yeterian EH, Pandya DN, Tomaiuolo F, Petrides M. 2012. Thecortical connectivity of the prefrontal cortex in the mon-key brain. Cortex 48:58–81.
Zhou H, Desimone R. 2011. Feature-based attention in thefrontal eye field and area V4 during visual search. Neu-ron 70:1205–1217.
1690 The Journal of Comparative Neurology |Research in Systems Neuroscience
K.S. Saleem et al.