study of the selective uptake progress of aptamer-modified plga particles by liver cells
TRANSCRIPT

Full Paper
Study of the Selective Uptake Progress ofAptamer-Modified PLGA Particles byLiver Cellsa
Dahai Yu, Yuying Zhang, Zhengwei Mao,* Changyou Gao*
It is of paramount importance to study the cellular uptake processes of particles with definedsurface property, especially the uptake pathways and intracellular transportation. In thisstudy, aptamer AS1411 molecules, which are known to specifically bind the over-expressed
nucleolin on cancer cell membrane, were conjugatedonto bovine serum albumin-decorated poly(D,L-lactide-co-glycolide; PLGA, F400nm) particles with a density of1–1.7 molecule/10 nm2. The aptamer-modified PLGAparticles were preferably ingested by liver cancer cellswith higher amount and faster rate. The clathrin-mediated endocytosis and macropinocytosis pathwaysplayed a more important role in uptake of the aptamermodified particles.D. Yu, Y. Zhang, Z. Mao, C. GaoMOE Key Laboratory of Macromolecular Synthesis andFunctionalization, Department of Polymer Science andEngineering, Zhejiang University, Hangzhou 310027, ChinaE-mail: [email protected]. GaoState Key Laboratory of Diagnosis and Treatment for InfectiousDiseases, First Affiliated Hospital, College of Medicine, ZhejiangUniversity, Hangzhou 310003, China,E-mail: [email protected]
aSupporting Information is available from theWiley Online Library orfrom the author.
� 2013 WILEY-VCH Verlag GmbH & Co. KGaA, WeinheimMacromol. Biosci. 2013, 13, 1413–1421
wileyonlinelibrary.com
[8–11] [12–15]
1. IntroductionIn order to decrease the dosage and subsequent toxicity,
several active and passive strategies have been developed
toenhance thedrugconcentration indesiredcells or tissues,
amongwhich decoration of the particle surfacewith ligand
molecules for specific cells or tissues has attracted
tremendous attention in recent years.[1–4] The folate,[5–7]
Arg-Gly-Asp (RGD), transferrin, and monoclonal
antibody[16–19] are the typical ligands used to target
particles to many kinds of carcinomas.
Due to their particular small size and/or other function-
alities, the particles are possibly internalized into cells
when they come into contact with the cells. In the
endocytosis process the particles are firstly attached to
the cell membrane through specific or nonspecific inter-
actions, and then are wrapped by cell membrane to form
membrane-bounded vesicles. The particles can enter cells
by several different endocytic pathways, such as phagocy-
tosis, clathrin-mediated endocytosis, caveolae-mediated
endocytosis, macropinocytosis, and clathrin- and caveolin-
independent endocytosis.[20] The internalization pathways
not only influence the uptake efficiency but also the
intracellular fate of the particles.[21] Moreover, the release
of the drug into the enzymatic environment of the
lysosomes or directly in the cell cytoplasm will, indeed,
has different pharmacological activities.[22] Therefore, the
uptake pathways and intracellular transportation of
particles should be clarified with great care.
It is known that the cellular uptake process and
intracellular fate of the particles are dominated by their
DOI: 10.1002/mabi.201300165 1413

www.mbs-journal.de
D. Yu, Y. Zhang, Z. Mao, C. Gao
1414
surface chemistry especially the existence of ligand
molecules.[22] Generally, the surface-tethered ligand mol-
eculesare feasible foruptakeof theparticles.However, their
influence on theparticle uptakepathways and intracellular
location is still quite controversial. For example, Ferris et al.
observed that transferrin and cyclic-RGD peptides-modifi-
cation enhanced the adherence of mesoporous silica
particles to the cancer cell membrane, suggesting the
membrane receptor-mediated endocytosis.[23] Tung et al.
found that human lung carcinoma A549 cells endocyto-
sized more silica particles decorated with monoclonal
antibody against epidermal growth factor receptor (EGFR,
which is over-expressed on A549 cells) compared with
the unmodified ones. Both particles were internalized
by macropinocytosis and clathrin-mediated pathways,
and resided in lysosomes.[24] Fujiwara et al. found that
octaarginine-modified liposomes were internalized by
Madin-Darby canine kidney cells (MDCK cells) and located
in endosomes and lysosomes. By contrast, ligand-free
liposomes were always located in lysosomes.[25]
Mao et al.[26] demonstrated that Tat peptide decora-
tion affected the subcellular distribution of silica
particles, resulting in localization of the particles in the
cell nuclei.
Recently, aptamers,[27] which are oligonucleic acids or
peptides that bind to a type of specific target molecules,
have shown potential applications in diagnostics, purifica-
tion processes, drug discovery, and even therapeutic
approaches.[28,29] Previous studies have demonstrated
that functional particles with surface-tethered aptamers
have potential applications in tumor diagnosis and
therapy.[30–35] AS1411 is the first oligodeoxynucleotide
aptamer to reach phase I and II clinical trials for the
potential treatment of cancers, including acute myeloge-
nous leukemia.[36,37] It shows high binding affinity to
nucleolin, which is overexpressed on the plasma
membrane of tumor cells.[38–41] Therefore, covalent conju-
gation or physical immobilization of AS1411 onto
particles will largely enhance their targeting efficiency to
tumor cells. Nevertheless, so far most attention has been
paid to realizing either high selective targeting and anti-
tumor efficacy,[42–44] or bioimaging and tracking cancer
cells.[45–47] Little attention is focused on the uptake
processes. It is of high demand to clarify the cellular uptake
processes of the AS1411-modified particles, especially
the uptake pathways and intracellular transportation
because targeting therapeutics to individual intracellular
compartments has resulted in benefits to therapies
associated with these unique organelles.
Our previous study have shown that the surface
modification of PLGA particles with anti-foulingmolecules
bovine serum albumin (BSA) could significantly increase
their stability in physiological fluids and reduce the
adsorption of serum proteins and thereby the uptake by
Macromol. Biosci. 201
� 2013 WILEY-VCH Verlag GmbH
endothelial cells.[48,49] Taking this advantage, in this work
the PLGA–BSA particles were further covalently immobi-
lizedwith the aptamer AS1411molecules, and their uptake
by liver cancer cells (QGY-7703 cells) with over-expressed
nucleolin and normal liver cells (Hepli cells) was compared
in terms of uptake kinetics, pathways and subcellular
distributions. Herein, we investigated the cell–particle
interactions at molecular and subcellular levels, and
provided insight in design of functional particles for
selective intracellular delivery.
2. Experimental Section
2.1. Materials
Poly(D,L-lactide-co-glycolide (LA: GA¼75: 25,Mw ¼ 130 kD), BSA, 1-
ethyl-3-(3-dimethyl aminopropyl) carbodiimide hydrochloride
(EDC), N-hydroxysuccinimide (NHS), amiloride–HCl, amanta-
dine–HCl, sodium azide, genistein, 40,6-diamidino-2-phenylindole
(DAPI), cytochalasin D (CytD), and 3-(4,5-dimethylthiazol-2-yl)-2,5-
diphe-nyltetrazolium bromide (MTT) were purchased from Sigma-
Aldrich. Sodiumdodecyl sulfate (SDS)was purchased fromHaotian
Co., Ltd. (China). Anti-EEA-1 (early endosome marker) and
LysoTrackerGreenwereobtained fromInvitrogenCo., Ltd.Aptamer
AS1411-NH2 and Cy3—AS1411—NH2 were purchased from
Sangon Biotech Co., Ltd. (Shanghai, China). All other chemicals
were analytical grade andused as received.Milli-Qwaterwas used
throughout the experiments.
2.2. Particle Preparation
The PLGA–BSA (PB) particles were prepared by an O/W emulsion–
solvent evaporation method as described previously.[48] Briefly,
after 1mL 2w/v% PLGA dichloromethane solution (organic phase)
was added into 4mL 3% BSA solution (water phase), the mixture
solution was emulsified with an ultrasonicator (MISONIX Ultra-
sonic liquid Processors, 5W) for 20 s. The obtained emulsion was
poured into 150mL water, and stirred for 3 h at room temperature
with a magnetic stirrer until the organic solvent was completely
evaporated. The PB particles were collected by centrifugation at 12
500 rpm for 10min, and washed with water five times to remove
free BSA in the water phase and freeze-dried for storage. The PB
particles containing Nile red (NR) were similarly prepared by
adding 0.2mgmL�1 NR into the PLGA solution before mixing with
the BSA solution, respectively.
2.3. Surface Conjugation of Aptamer AS1411
The sequence of AS1411 is 50-(GGTGGTGGTGGTTGTGGTGGT-GGTGG). The AS1411 aptamer capped with an amino end group
(aptamer-NH2)was covalently conjugated to the PB particles using
carbodiimide chemistry. In brief, the 0.5mL suspension of 1mg
mL�1 PB particles was mixed with 1mg EDC and 0.5mg NHS to
activate the –COOH groups. 20min later, 0.5mL aptamer–NH2 (or
Cy3–aptamer–NH2) solution was added and the reaction was
maintained for 40min under shaking at room temperature. The
3, 13, 1413–1421
& Co. KGaA, Weinheim www.MaterialsViews.com

Study of the Selective Uptake Progress of Aptamer-Modified PLGA Particles . . .
www.mbs-journal.de
PLGA–BSA–aptamer (PB-Apt) particles were collected by centrifu-
gation at 12 500 rpm for 10min, andwashedwithwater five times
under ultrasonication to remove physically adsorbed aptamer
molecules.
The fluorescence intensity of the aptamer solution before and
after reaction was compared to assess the grafting amount of the
aptamer by fluorescence spectroscopy (LS55, PerkinElmer, USA)
using Cy3–aptamer–NH2.
2.4. Particle Characterization
2.4.1. Morphology
Themorphology and the size of the PB and PB-Apt particles in a dry
state were analyzed by transmission electron microscopy (TEM,
Philips TECNAL-10). A drop of the particles suspension was added
onto a copper grid with a carbon membrane and dried at room
temperature. 100 particles were analyzed to provide statistical
result of the particle size.
2.4.2. Size and Surface Charge
The size and surface charge of the PB and PB-Apt particles in water
(pH 6.6) and DMEM/10% fetal bovine serum (pH 7.4) were
determined by dynamic light scattering with Beckman Delsa
Nano (Beckman Coulter), respectively.
2.5. Cell Uptake
2.5.1. Cell Culture
The immortalized human normal liver cells (Hepli cells) were
kindly donated by Dr. Jun Li from the First Affiliated Hospital,
College of Medicine, Zhejiang University. The human liver cancer
cells (QGY-7703 cells) were obtained from the Cell Bank of Typical
Culture Collection of Chinese Academy of Sciences (Shanghai,
China). The Hepli and QGY cells were maintained at 37 8C in a 5%
CO2 humidified environment in low glucose DMEM and high
glucose RPMI 1640 (Gibco, USA), respectively. Both culture
mediumswere supplementedwith 10%FBS, 100UmL�1 penicillin,
and 100mgmL�1 streptomycin.
2.5.2. Cellular Uptake of the PLGA Particles
The cellswere seeded on a 24-well plate at a density of 1� 105 cells
per well and allowed to attach for 16 h. Then they were incubated
with 50mgmL�1 of NR-labeled PLGA particles (PB and PB-Apt) for
different time, or with various concentrations of NR-labeled PLGA
particles for 12h. After washed five times with PBS to remove
the free particles, the cells were detached by trypsin. The uptake
amount of the NR-labeled PLGA particles was determined by flow
cytometry (FACS Calibur, BD).
2.5.3. Uptake Pathway
In order to clarify the role of ligand–receptor interactions in the
processof endocytosis, theQGY-7703cellswere incubatedwithNR-
Macromol. Biosci. 20
� 2013 WILEY-VCH Verlag Gmwww.MaterialsViews.com
labeled PLGA particles for 2 h in the presence and absences of 3mL
100mM free AS1411 at 4 8C, respectively. The cellular uptake of thePLGA particles was then determined by flow cytometry. To
determine the uptake pathways, the cells were firstly incubated
at 4 8C andwith 100mM sodium azide in the presence of particles,
respectively. Different pharmacological inhibitors, including2mM
amiloride–HCl, 1�10�3M amantadine–HCl, 100�10�3
M genis-
tein, and10mgmL�1 cytochalasinDwerealsoused to treat the cells
for 1 h before incubation with the PLGA particles, respectively.
2.5.4. Intracellular Distribution
Fluorescent stainingof early endosomes, lysosomes, and cell nuclei
was performed to display the intracellular distribution of the PLGA
particles by confocal laser scanning microscopy (CLSM, TCS SP5,
Leica). Briefly, after incubatedwith 50mgmL�1 of NR-labeled PLGA
particles for desired time, the cells were carefullywashedwith PBS
three times, and continually culturedwith anti-EEA-1, LysoTracker
Green, and DAPI at 37 8C for another 30min, respectively.
2.6. Statistical Analysis
All values are expressed as mean� standard deviation (SD).
Statistically significant value is set as p <0.05 based on the
Student’s t-test.
3. Results and Discussion
3.1. Characterization of PLGA–BSA–Aptamer Particles
The PLGA–BSA particles were prepared via an O/W
emulsion-solvent evaporationmethodwith BSAmolecules
as stabilizers in the water phase. As a result, the obtained
PLGA particles had a shallow shell (several tens of nm) of
BSA molecules (Figure 1C), contributing about 6% of the
particle weight. These BSA molecules were very stable and
cannot be removed even under strong ultrasonication.[48]
This feature enabled the covalent tether of amino end-
capped AS1411 by the carboxyl groups of BSAmolecules in
the presence of EDC/NHS. To quantify the grafting amount,
Cy3 labeled aptamer molecules were used. Of the 2.0mg
feeding amount, 1.9mg aptamer molecules were conjugat-
ed onto 1mg PB particles (PB-Apt2), suggesting the very
high conjugation efficiency. Taking into account the
average particle size (Table 1), this value equaled to
3.8� 104 molecules per particle or 1 molecule/10 nm2.
When the feeding amount of aptamer was raised to 5mg
per mg PB particles, the grafting amount reached to 3.3mg
(PB-Apt3),which equaled to 6.3� 104molecules per particle
or 1.7 molecules/10 nm2.
As representatively shown in Figure 1 and Supporting
Information, Figure S1, both the PB and PB-Apt3 particles
possessed a size of 170nm in a dry state and a typical core–
shell structure regardlessof theconjugationofaptamer.The
size of PB-Apt3 particles (�430nm due to the hydration of
13, 13, 1413–1421
bH & Co. KGaA, Weinheim 1415

Figure 1. A,B) SEM and C,D) TEM images of A,C) PLGA–BSA (PB)particles and B,D) PB-Apt3 (3.3mg aptamer per mg PB particle)particles, respectively.
www.mbs-journal.de
D. Yu, Y. Zhang, Z. Mao, C. Gao
1416
surface protein layer and possible aggregation) kept
unchanged in comparison with that of the PB particles in
both water and serum-containing cell culture medium,
suggesting thegoodcolloidal stabilityof theparticlesdue to
the surface coating of BSA molecules. The PB particles
possessed negative charge on their surface due to the BSA
protecting layer, which was about �30mV in water and
�10mV in serum-containing medium.[48] The surface
charge property was not significantly altered after the
EDC/NHS treatment and conjugation of the AS1411
aptamer molecules (Table 1). Therefore, the potential
influence of physical parameters, such as size and surface,
charge on the cellular uptake of the aptamer-tethered PLGA
particles could be safely ruled out.
3.2. Cellular Uptake
The aptamer-tethered PLGA particles are expected to be
selectively delivered to the tumor cells, which could then
improve the therapeutic effects of the encapsulated
substances.[50] In this regard, the cellular internalization
Table 1. The size and surface charge properties of PB, PB-Apt2, and
Sample
Size [nm]
Water 1 640/1
PB particles 435� 13 438
PB-Apt2 particles 434� 26 441
PB-Apt3 particles 435� 27 441
Macromol. Biosci. 201
� 2013 WILEY-VCH Verlag GmbH
of the nanoparticles is very important. In this study two
types of liver-derived cellswereused.QGY7703 cell is a type
of hepatic carcinoma cell that over expresses nucleolin (the
receptor of aptamerAS1411) on cellmembrane. Hepli cell is
a type of immortalized normal liver cells, which express a
normal level of nucleolin on cell membrane. By the
comparison, the differences in delivery and uptake of the
aptamer-modified particles can be distinguished. To make
the PLGA particles detectable via fluorescence microscopy
andflowcytometry, a hydrophobic dye, NR,was pre-loaded
during the particle fabrication. Due to its very poor
solubility in culture medium, the release of NR from the
particleswasneglectable, asdemonstratedbythe transwell
experiments.[48] In the FCMmeasurement, the logarithmic
fluorescence intensity of untreated cells was set between
100 and 101, and those cells with the intensity larger than
101 were considered as the positive ones (the cells ingested
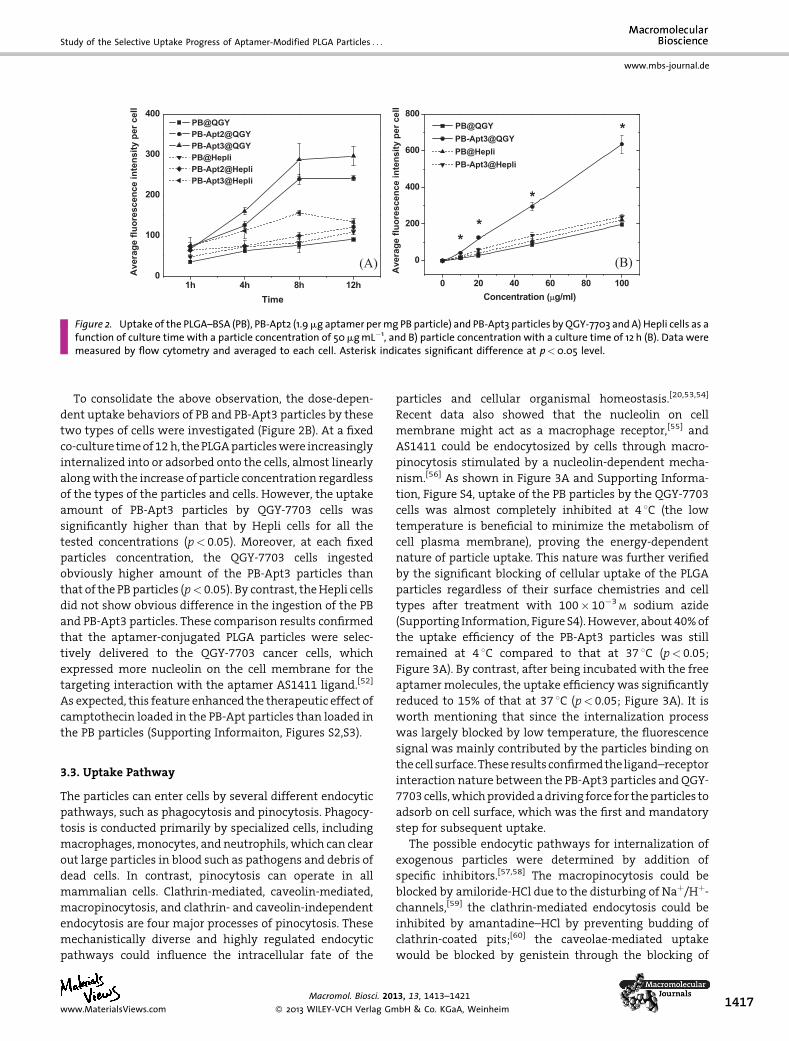
the PLGA particles).[51] As shown in Figure 2A, the average
fluorescence intensity per cell increased along with the
prolongation of culture time for all types of the PLGA
particles andboth types of cells,with faster rates during the
first 8 h. In general, the normal cells (Hepli) internalized a
similar amount of PB and PB-Apt2 particles during the
whole experimental period (p< 0.05). However, the fluo-
rescence intensity of the Hepli cells incubated with the PB-
Apt3 particles was higher compared to that of the cells
incubatedwith the PBandPB-Apt2particles during thefirst
8 h, suggestingmorePB-Apt3particleswere takenupby the
Hepli cells. This difference disappeared at 12h. This might
be attributed to the fact that someamount of nucleolin also
existed on the surface of Hepli cells. Thus higher density of
aptamer AS1411 would lead to the enhanced recognition
and internalization of PB-Apt particles by Hepli cells to
some extent.
TheQGY-7703 cells took up a significantly larger amount
of PB-Apt2 particles than PB particles (p< 0.05) during the
whole experimental period, whichwas about four times at
the maximum at 8h. They internalized evenmore PB-Apt3
particles except at the first hour. During all the time points
(p< 0.05) except the first hour, the QGY-7703 cells also took
up significantly larger amounts of the PB-Apt2 and PB-Apt3
particles, which were 2 and 2.2 times to the Hepli cells at
12h, respectively.
PB-Apt3 particles.
Zeta potential [mV]
0% FBS Water 1 640/10% FBS
� 30 �34� 0.7 �17� 1.4
� 20 �35� 4.2 �16� 1.5
� 24 �36� 5.6 �15� 0.7
3, 13, 1413–1421
& Co. KGaA, Weinheim www.MaterialsViews.com
1h 4h 8h 12h0
100
200
300
400
Ave
rage
fluo
resc
ence
inte
nsity
per
cel
l
Time
PB@QGY PB-Apt2@QGY PB-Apt3@QGY PB@Hepli PB-Apt2@Hepli PB-Apt3@Hepli
0 20 40 60 80 100
0
200
400
600
800
*
*
**
Ave
rage
fluo
resc
ence
inte
nsity
per
cel
l
Concentration (µg/ml)
PB@QGY PB-Apt3@QGY PB@Hepli PB-Apt3@Hepli
(B)(A)
Figure 2. Uptake of the PLGA–BSA (PB), PB-Apt2 (1.9mg aptamer permg PB particle) and PB-Apt3 particles by QGY-7703 and A) Hepli cells as afunction of culture time with a particle concentration of 50mgmL�1, and B) particle concentration with a culture time of 12 h (B). Data weremeasured by flow cytometry and averaged to each cell. Asterisk indicates significant difference at p<0.05 level.
Study of the Selective Uptake Progress of Aptamer-Modified PLGA Particles . . .
www.mbs-journal.de
To consolidate the above observation, the dose-depen-
dent uptake behaviors of PB and PB-Apt3 particles by these
two types of cells were investigated (Figure 2B). At a fixed
co-culture timeof12h, thePLGAparticleswere increasingly
internalized into or adsorbed onto the cells, almost linearly
alongwith the increase of particle concentration regardless
of the types of the particles and cells. However, the uptake
amount of PB-Apt3 particles by QGY-7703 cells was
significantly higher than that by Hepli cells for all the
tested concentrations (p< 0.05). Moreover, at each fixed
particles concentration, the QGY-7703 cells ingested
obviously higher amount of the PB-Apt3 particles than
that of the PB particles (p< 0.05). By contrast, theHepli cells
did not show obvious difference in the ingestion of the PB
and PB-Apt3 particles. These comparison results confirmed
that the aptamer-conjugated PLGA particles were selec-
tively delivered to the QGY-7703 cancer cells, which
expressed more nucleolin on the cell membrane for the
targeting interaction with the aptamer AS1411 ligand.[52]
As expected, this feature enhanced the therapeutic effect of
camptothecin loaded in the PB-Apt particles than loaded in
the PB particles (Supporting Informaiton, Figures S2,S3).
3.3. Uptake Pathway
The particles can enter cells by several different endocytic
pathways, such as phagocytosis and pinocytosis. Phagocy-
tosis is conducted primarily by specialized cells, including
macrophages,monocytes, and neutrophils, which can clear
out large particles in blood such as pathogens and debris of
dead cells. In contrast, pinocytosis can operate in all
mammalian cells. Clathrin-mediated, caveolin-mediated,
macropinocytosis, and clathrin- and caveolin-independent
endocytosis are four major processes of pinocytosis. These
mechanistically diverse and highly regulated endocytic
pathways could influence the intracellular fate of the
Macromol. Biosci. 20
� 2013 WILEY-VCH Verlag Gmwww.MaterialsViews.com
particles and cellular organismal homeostasis.[20,53,54]
Recent data also showed that the nucleolin on cell
membrane might act as a macrophage receptor,[55] and
AS1411 could be endocytosized by cells through macro-
pinocytosis stimulated by a nucleolin-dependent mecha-
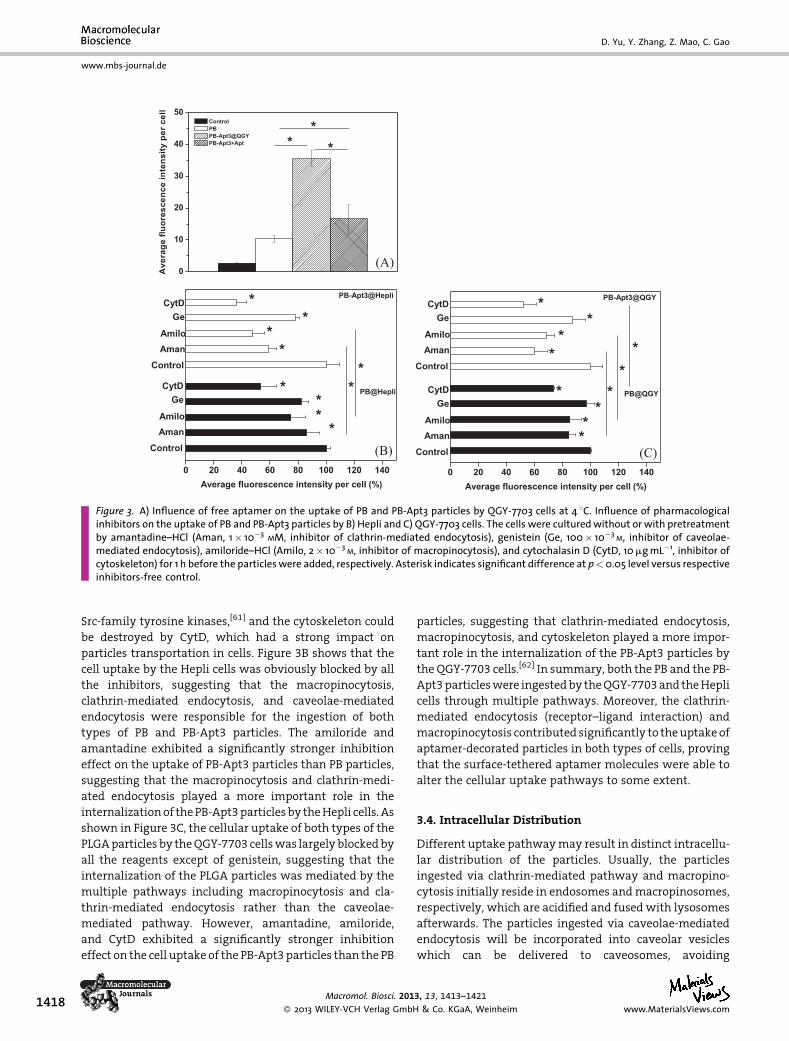
nism.[56] As shown in Figure 3A and Supporting Informa-
tion, Figure S4, uptake of the PB particles by the QGY-7703
cells was almost completely inhibited at 4 8C (the low
temperature is beneficial to minimize the metabolism of
cell plasma membrane), proving the energy-dependent
nature of particle uptake. This nature was further verified
by the significant blocking of cellular uptake of the PLGA
particles regardless of their surface chemistries and cell
types after treatment with 100� 10�3M sodium azide
(Supporting Information, Figure S4). However, about 40%of
the uptake efficiency of the PB-Apt3 particles was still
remained at 4 8C compared to that at 37 8C (p< 0.05;
Figure 3A). By contrast, after being incubated with the free
aptamermolecules, the uptake efficiencywas significantly
reduced to 15% of that at 37 8C (p< 0.05; Figure 3A). It is
worth mentioning that since the internalization process
was largely blocked by low temperature, the fluorescence
signal was mainly contributed by the particles binding on
thecell surface.These results confirmedthe ligand–receptor
interaction nature between the PB-Apt3 particles andQGY-
7703cells,whichprovidedadriving force for theparticles to
adsorb on cell surface, which was the first and mandatory
step for subsequent uptake.
The possible endocytic pathways for internalization of
exogenous particles were determined by addition of
specific inhibitors.[57,58] The macropinocytosis could be
blocked by amiloride-HCl due to the disturbing of Naþ/Hþ-channels,[59] the clathrin-mediated endocytosis could be
inhibited by amantadine–HCl by preventing budding of
clathrin-coated pits;[60] the caveolae-mediated uptake
would be blocked by genistein through the blocking of
13, 13, 1413–1421
bH & Co. KGaA, Weinheim 1417
0
10
20
30
40
50llec rep ytisnetni e cne cs eroulf egarevA
Control PB PB-Apt3@QGY PB-Apt3+Apt
***
(A)
0 20 40 60 80 100 120 140
PB-Apt3@Hepli
**
Average fluorescence intensity per cell (%)
PB@Hepli***
*
**
**
Control
AmanAmilo
GeCytD
Control
AmanAmilo
GeCytD
0 20 40 60 80 100 120 140
*
PB-Apt3@QGY
**
Average fluorescence intensity per cell (%)
PB@QGY**
**
**
**
Control
AmanAmilo
GeCytD
Control
AmanAmilo
GeCytD
(C)(B)
Figure 3. A) Influence of free aptamer on the uptake of PB and PB-Apt3 particles by QGY-7703 cells at 4 8C. Influence of pharmacologicalinhibitors on the uptake of PB and PB-Apt3 particles by B) Hepli and C) QGY-7703 cells. The cells were culturedwithout or with pretreatmentby amantadine–HCl (Aman, 1� 10�3 MM, inhibitor of clathrin-mediated endocytosis), genistein (Ge, 100� 10�3 M, inhibitor of caveolae-mediated endocytosis), amiloride–HCl (Amilo, 2� 10�3 M, inhibitor of macropinocytosis), and cytochalasin D (CytD, 10mgmL�1, inhibitor ofcytoskeleton) for 1 h before the particleswere added, respectively. Asterisk indicates significant difference at p<0.05 level versus respectiveinhibitors-free control.
www.mbs-journal.de
D. Yu, Y. Zhang, Z. Mao, C. Gao
1418
Src-family tyrosine kinases,[61] and the cytoskeleton could
be destroyed by CytD, which had a strong impact on
particles transportation in cells. Figure 3B shows that the
cell uptake by the Hepli cells was obviously blocked by all
the inhibitors, suggesting that the macropinocytosis,
clathrin-mediated endocytosis, and caveolae-mediated
endocytosis were responsible for the ingestion of both
types of PB and PB-Apt3 particles. The amiloride and
amantadine exhibited a significantly stronger inhibition
effect on the uptake of PB-Apt3 particles than PB particles,
suggesting that the macropinocytosis and clathrin-medi-
ated endocytosis played a more important role in the
internalizationof thePB-Apt3particlesby theHepli cells. As
shown in Figure 3C, the cellular uptake of both types of the
PLGAparticles by theQGY-7703 cellswas largelyblockedby
all the reagents except of genistein, suggesting that the
internalization of the PLGA particles was mediated by the
multiple pathways including macropinocytosis and cla-
thrin-mediated endocytosis rather than the caveolae-
mediated pathway. However, amantadine, amiloride,
and CytD exhibited a significantly stronger inhibition
effect on the cell uptake of the PB-Apt3particles than the PB
Macromol. Biosci. 201
� 2013 WILEY-VCH Verlag GmbH
particles, suggesting that clathrin-mediated endocytosis,
macropinocytosis, and cytoskeleton played a more impor-
tant role in the internalization of the PB-Apt3 particles by
the QGY-7703 cells.[62] In summary, both the PB and the PB-
Apt3particleswere ingestedby theQGY-7703and theHepli
cells through multiple pathways. Moreover, the clathrin-
mediated endocytosis (receptor–ligand interaction) and
macropinocytosis contributed significantly to theuptakeof
aptamer-decorated particles in both types of cells, proving
that the surface-tethered aptamer molecules were able to
alter the cellular uptake pathways to some extent.
3.4. Intracellular Distribution
Different uptake pathwaymay result in distinct intracellu-
lar distribution of the particles. Usually, the particles
ingested via clathrin-mediated pathway and macropino-
cytosis initially reside in endosomes andmacropinosomes,
respectively, which are acidified and fused with lysosomes
afterwards. The particles ingested via caveolae-mediated
endocytosis will be incorporated into caveolar vesicles
which can be delivered to caveosomes, avoiding
3, 13, 1413–1421
& Co. KGaA, Weinheim www.MaterialsViews.com
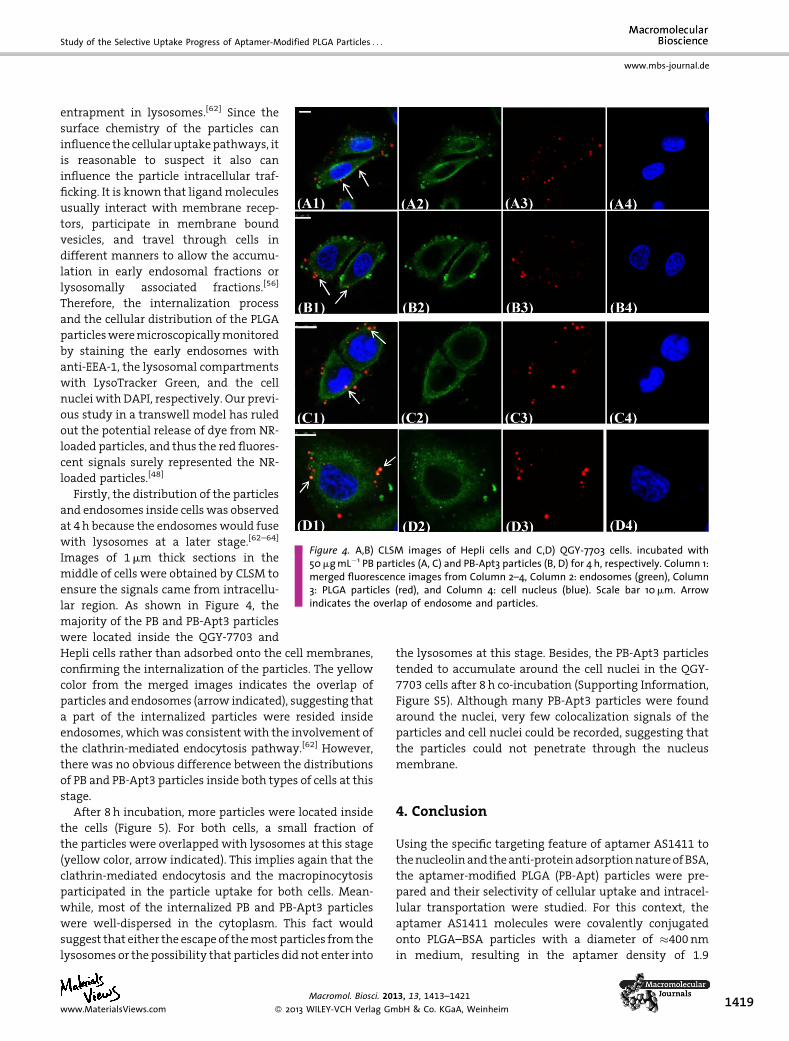
Figure 4. A,B) CLSM images of Hepli cells and C,D) QGY-7703 cells. incubated with50mgmL�1 PB particles (A, C) and PB-Apt3 particles (B, D) for 4 h, respectively. Column 1:merged fluorescence images from Column 2–4, Column 2: endosomes (green), Column3: PLGA particles (red), and Column 4: cell nucleus (blue). Scale bar 10mm. Arrowindicates the overlap of endosome and particles.
Study of the Selective Uptake Progress of Aptamer-Modified PLGA Particles . . .
www.mbs-journal.de
entrapment in lysosomes.[62] Since the
surface chemistry of the particles can
influence the cellular uptakepathways, it
is reasonable to suspect it also can
influence the particle intracellular traf-
ficking. It is known that ligandmolecules
usually interact with membrane recep-
tors, participate in membrane bound
vesicles, and travel through cells in
different manners to allow the accumu-
lation in early endosomal fractions or
lysosomally associated fractions.[56]
Therefore, the internalization process
and the cellular distribution of the PLGA
particlesweremicroscopicallymonitored
by staining the early endosomes with
anti-EEA-1, the lysosomal compartments
with LysoTracker Green, and the cell
nuclei with DAPI, respectively. Our previ-
ous study in a transwell model has ruled
out the potential release of dye from NR-
loaded particles, and thus the red fluores-
cent signals surely represented the NR-
loaded particles.[48]
Firstly, the distribution of the particles
and endosomes inside cells was observed
at 4h because the endosomeswould fuse
with lysosomes at a later stage.[62–64]
Images of 1mm thick sections in the
middle of cells were obtained by CLSM to
ensure the signals came from intracellu-
lar region. As shown in Figure 4, the
majority of the PB and PB-Apt3 particles
were located inside the QGY-7703 and
Hepli cells rather than adsorbed onto the cell membranes,
confirming the internalization of the particles. The yellow
color from the merged images indicates the overlap of
particles and endosomes (arrow indicated), suggesting that
a part of the internalized particles were resided inside
endosomes, whichwas consistentwith the involvement of
the clathrin-mediated endocytosis pathway.[62] However,
there was no obvious difference between the distributions
of PB and PB-Apt3 particles inside both types of cells at this
stage.
After 8 h incubation, more particles were located inside
the cells (Figure 5). For both cells, a small fraction of
the particles were overlapped with lysosomes at this stage
(yellow color, arrow indicated). This implies again that the
clathrin-mediated endocytosis and the macropinocytosis
participated in the particle uptake for both cells. Mean-
while, most of the internalized PB and PB-Apt3 particles
were well-dispersed in the cytoplasm. This fact would
suggest that either the escapeof themost particles fromthe
lysosomes or the possibility that particles did not enter into
Macromol. Biosci. 20
� 2013 WILEY-VCH Verlag Gmwww.MaterialsViews.com
the lysosomes at this stage. Besides, the PB-Apt3 particles
tended to accumulate around the cell nuclei in the QGY-
7703 cells after 8 h co-incubation (Supporting Information,
Figure S5). Although many PB-Apt3 particles were found
around the nuclei, very few colocalization signals of the
particles and cell nuclei could be recorded, suggesting that
the particles could not penetrate through the nucleus
membrane.
4. Conclusion
Using the specific targeting feature of aptamer AS1411 to
thenucleolinand theanti-proteinadsorptionnatureofBSA,
the aptamer-modified PLGA (PB-Apt) particles were pre-
pared and their selectivity of cellular uptake and intracel-
lular transportation were studied. For this context, the
aptamer AS1411 molecules were covalently conjugated
onto PLGA–BSA particles with a diameter of �400nm
in medium, resulting in the aptamer density of 1.9
13, 13, 1413–1421
bH & Co. KGaA, Weinheim 1419
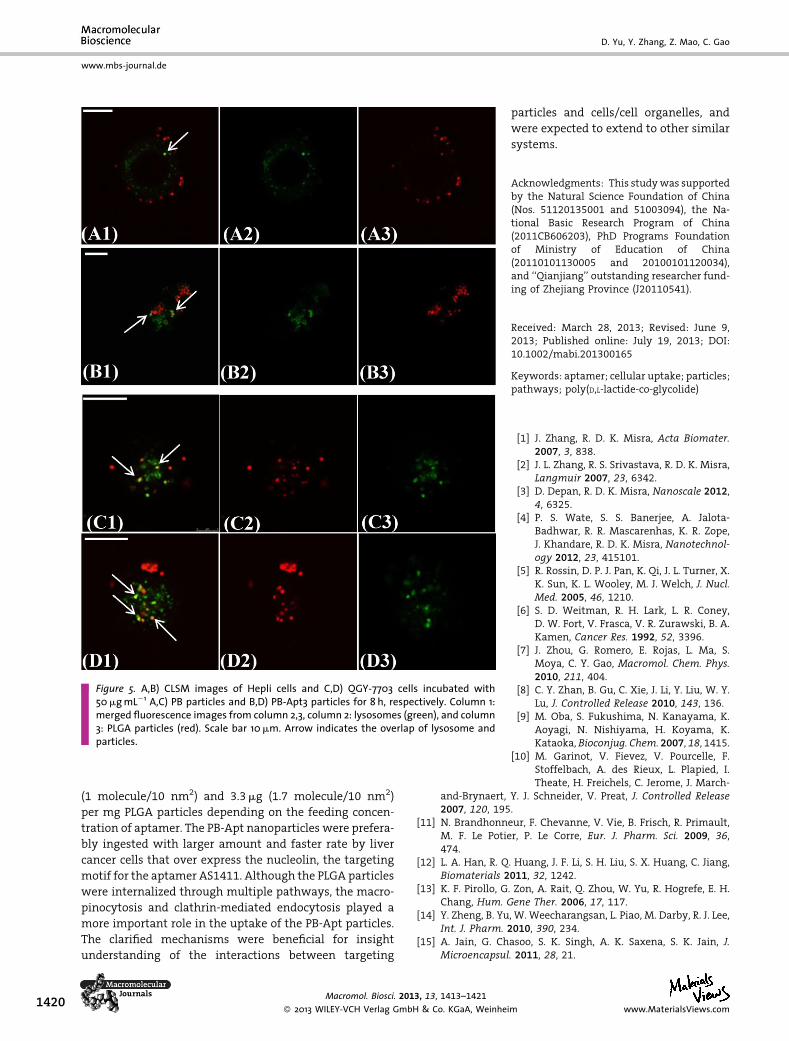
Figure 5. A,B) CLSM images of Hepli cells and C,D) QGY-7703 cells incubated with50mgmL�1 A,C) PB particles and B,D) PB-Apt3 particles for 8 h, respectively. Column 1:merged fluorescence images from column 2,3, column 2: lysosomes (green), and column3: PLGA particles (red). Scale bar 10mm. Arrow indicates the overlap of lysosome andparticles.
www.mbs-journal.de
D. Yu, Y. Zhang, Z. Mao, C. Gao
1420
(1 molecule/10 nm2) and 3.3mg (1.7 molecule/10 nm2)
per mg PLGA particles depending on the feeding concen-
tration of aptamer. The PB-Apt nanoparticles were prefera-
bly ingested with larger amount and faster rate by liver
cancer cells that over express the nucleolin, the targeting
motif for the aptamer AS1411. Although the PLGA particles
were internalized through multiple pathways, the macro-
pinocytosis and clathrin-mediated endocytosis played a
more important role in the uptake of the PB-Apt particles.
The clarified mechanisms were beneficial for insight
understanding of the interactions between targeting
Macromol. Biosci. 2013, 13, 1413–1421
� 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinhe
particles and cells/cell organelles, and
were expected to extend to other similar
systems.
Acknowledgments: This studywas supportedby the Natural Science Foundation of China(Nos. 51120135001 and 51003094), the Na-tional Basic Research Program of China(2011CB606203), PhD Programs Foundationof Ministry of Education of China(20110101130005 and 20100101120034),and ‘‘Qianjiang’’ outstanding researcher fund-ing of Zhejiang Province (J20110541).
Received: March 28, 2013; Revised: June 9,2013; Published online: July 19, 2013; DOI:10.1002/mabi.201300165
Keywords: aptamer; cellular uptake; particles;pathways; poly(D,L-lactide-co-glycolide)
[1] J. Zhang, R. D. K. Misra, Acta Biomater.2007, 3, 838.
[2] J. L. Zhang, R. S. Srivastava, R. D. K. Misra,Langmuir 2007, 23, 6342.
[3] D. Depan, R. D. K. Misra, Nanoscale 2012,4, 6325.
[4] P. S. Wate, S. S. Banerjee, A. Jalota-Badhwar, R. R. Mascarenhas, K. R. Zope,J. Khandare, R. D. K. Misra, Nanotechnol-ogy 2012, 23, 415101.
[5] R. Rossin, D. P. J. Pan, K. Qi, J. L. Turner, X.K. Sun, K. L. Wooley, M. J. Welch, J. Nucl.Med. 2005, 46, 1210.
[6] S. D. Weitman, R. H. Lark, L. R. Coney,D. W. Fort, V. Frasca, V. R. Zurawski, B. A.Kamen, Cancer Res. 1992, 52, 3396.
[7] J. Zhou, G. Romero, E. Rojas, L. Ma, S.Moya, C. Y. Gao, Macromol. Chem. Phys.2010, 211, 404.
[8] C. Y. Zhan, B. Gu, C. Xie, J. Li, Y. Liu, W. Y.Lu, J. Controlled Release 2010, 143, 136.
[9] M. Oba, S. Fukushima, N. Kanayama, K.Aoyagi, N. Nishiyama, H. Koyama, K.Kataoka, Bioconjug. Chem. 2007, 18, 1415.
[10] M. Garinot, V. Fievez, V. Pourcelle, F.Stoffelbach, A. des Rieux, L. Plapied, I.Theate, H. Freichels, C. Jerome, J. March-
and-Brynaert, Y. J. Schneider, V. Preat, J. Controlled Release2007, 120, 195.
[11] N. Brandhonneur, F. Chevanne, V. Vie, B. Frisch, R. Primault,M. F. Le Potier, P. Le Corre, Eur. J. Pharm. Sci. 2009, 36,474.
[12] L. A. Han, R. Q. Huang, J. F. Li, S. H. Liu, S. X. Huang, C. Jiang,Biomaterials 2011, 32, 1242.
[13] K. F. Pirollo, G. Zon, A. Rait, Q. Zhou, W. Yu, R. Hogrefe, E. H.Chang, Hum. Gene Ther. 2006, 17, 117.
[14] Y. Zheng, B. Yu, W. Weecharangsan, L. Piao, M. Darby, R. J. Lee,Int. J. Pharm. 2010, 390, 234.
[15] A. Jain, G. Chasoo, S. K. Singh, A. K. Saxena, S. K. Jain, J.Microencapsul. 2011, 28, 21.
im www.MaterialsViews.com

Study of the Selective Uptake Progress of Aptamer-Modified PLGA Particles . . .
www.mbs-journal.de
[16] H. J. Lee, B. Engelhardt, J. Lesley, U. Bickel, W. M. Pardridge, J.Pharmacol. Exp. Ther. 2000, 292, 1048.
[17] J. M. Luk, K. F. Wong, Clin. Exp. Pharmacol. Physiol. 2006, 33,482.
[18] B. Sun, B. Ranganathan, S. S. Feng, Biomaterial 2008, 29, 475.[19] R. T. Carroll, D. Bhatia, W. Geldenhuys, R. Bhatia, N. Miladore,
V. Sutariya, J. Drug Target. 2010, 18, 665.[20] S. D. Conner, S. L. Schmid, Nature 2003, 422, 37.[21] S. Biswas, N. S. Dodwadkar, P. P. Deshpande, V. P. Torchilin, J.
Controlled Release 2012, 159, 393.[22] Z. W. Mao, X. Y. Zhou, C. Y. Gao, Biomater. Sci., in press.[23] D. P. Ferris, J. Lu, C. Gothard, R. Yanes, C. R. Thomas, J. C. Olsen,
J. F. Stoddart, F. Tamanoi, J. I. Zink, Small 2011, 7, 1816.[24] W. L. Tung, S. H. Hu, D. M. Liu, Acta Biomater. 2011, 7, 2873.[25] T. Fujiwara, H. Akita, H. Harashima, Int. J. Pharm. 2010, 386,
122.[26] Z. W. Mao, L. Wan, L. Hu, L. Ma, C. Y. Gao, Colloid Surf. B 2010,
75, 432.[27] S. M. Nimjee, C. P. Rusconi, B. A. Sullenger, Annu. Rev. Med.
2005, 56, 555.[28] D. Proske, M. Blank, R. Buhmann, A. Resch, Appl. Microbiol.
Biotechnol. 2005, 69, 367.[29] S. Tombelli, A.Minunni, A.Mascini, Biosens. Bioelectron. 2005,
20, 2424.[30] O. C. Farokhzad, S. Y. Jon, A. Khademhosseini, T. N. T. Tran,
D. A. LaVan, R. Langer, Cancer Res. 2004, 64, 7668.[31] O. C. Farokhzad, J. J. Cheng, B. A. Teply, I. Sherifi, S. Jon, P. W.
Kantoff, J. P. Richie, R. Langer, Proc. Natl. Acad. Sci. USA 2006,103, 6315.
[32] S. Dhar, F. X. Gu, R. Langer, O. C. Farokhzad, S. J. Lippard, Proc.Natl. Acad. Sci. U. S. A. 2008, 105, 17356.
[33] V. Bagalkot, L. Zhang, E. Levy-Nissenbaum, S. Jon, R. Langer,O. C. Farokhzad, Nano Lett. 2007, 7, 3065.
[34] L. L. Xie, W. J. Tong, J. X. Xu, C. Y. Gao, Chin. J. Polym. Sci. 2012,30, 719.
[35] R. Tong, L. D. Yala, T. M. Fan, J. J. Cheng, Biomaterials 2010, 31,3043.
[36] F. Mongelard, P. Bouvet, Curr. Opin. Mol. Ther. 2010, 12, 107.[37] K. Shah, H. Djeha, C. Richie, G. McGeever, C. Green, D. M.
Miller, Blood 2006, 108, 564A.[38] P. J. Bates, D. A. Laber, D. M. Miller, S. D. Thomas, J. O. Trent,
Exp. Mol. Pathol. 2009, 86, 151.[39] H. Iwasaki, K. Nabeshima, J. Nishio, S. Jimi, M. Aoki, K. Koga,
M. Hamasaki, H. Hayashi, A. Mogi, Pathol. Int. 2009, 59, 501.[40] T. Watanabe, K. Hirano, A. Takahashi, K. Yamaguchi, M.
Beppu, H. Fujiki, M. Suganuma, Biol. Pharm. Bull. 2010, 33,796.
Macromol. Biosci. 20
� 2013 WILEY-VCH Verlag Gmwww.MaterialsViews.com
[41] C. R. Ireson, L. R. Kelland, Mol. Cancer. Ther. 2006, 5, 2957.[42] Y. A. Shieh, S. J. Yang, M. F. Wei, M. J. Shieh, ACS Nano 2010, 4,
1433.[43] J. W. Guo, X. L. Gao, L. N. Su, H. M. Xia, G. Z. Gu, Z. Q. Pang, X. G.
Jiang, L. Yao, J. Chen, H. Z. Chen, Biomaterials 2011, 32, 8010.[44] A. Aravind, P. Jeyamohan, R. Nair, S. Veeranarayanan, Y.
Nagaoka, Y. Yoshida, T. Maekawa, D. S. Kumar, Biotechnol.Bioeng. 2012, 109, 2920.
[45] D. W. Hwang, H. Y. Ko, J. H. Lee, H. Kang, S. H. Ryu, S. Kim, J.Nucl. Med. 2010, 51, 98.
[46] W. J. Kang, M. H. Ko, D. S. Lee, S. Kim, Proteomics Clin. Appl.2009, 3, 1383.
[47] M. H. Ko, S. Kim, W. J. Kang, J. H. Lee, H. Kang, D. S. Lee, Small2009, 5, 1207.
[48] D. H. Yu, Y. Y. Zhang, X. Y. Zhou, Z. W. Mao, C. Y. Gao,Biomacromolecules 2012, 13, 3272.
[49] B. Wang, G. Q. Chen, Z. W. Mao, Y. Y. Zhang, D. H. Yu, C. Y. Gao,Chin. Sci. Bull. 2012, 57, 3985.
[50] A. Lamprecht, N. Ubrich, H. Yamamoto, U. Schafer, H.Takeuchi, P.Maincent, Y. Kawashima, C.M. Lehr, J. Pharmacol.Exp. Ther. 2001, 299, 775.
[51] T. H. Chung, S. H. Wu, M. Yao, C. W. Lu, Y. S. Lin, Y. Hung, C. Y.Mou, Y. C. Chen, D. M. Huang, Biomaterials 2007, 28, 2959.
[52] L. L. Xie, W. J. Tong, D. H. Yu, J. Q. Xu, J. Li, C. Y. Gao, J. Mater.Chem. 2012, 22, 6053.
[53] M. D. Chavanpatil, A. Khdair, J. Panyam, J. Nanosci. Nano-technol. 2006, 6, 2651.
[54] G. Sahay, D. Y. Alakhova, A. V. Kabanov, J. Controlled Release2010, 145, 182.
[55] K. Hirano, Y. Miki, Y. Hirai, R. Sato, T. Itoh, A. Hayashi, M.Yamanaka, S. Eda, M. J. Beppu, Biol. Chem. 2005, 280, 39284.
[56] E. M. Reyes-Reyes, Y. Teng, P. J. Bates, Cancer Res. 2010, 70,8617.
[57] H. Y. Nam, S. M. Kwon, H. S. Chung, Y. Lee, S. H. Kwon, H. Jeon,Y. Kim, J. H. Park, J. Kim, S. Her, Y. K. Oh, I. C. Kwon, K. Kim, S. Y.Jeong, J. Control. Release 2009, 135, 259.
[58] L. Hu, Y. Y. Zhang, C. Y. Gao, Prog. Chem. 2009, 21, 1254.[59] L. J. Hewlett, A. R. Prescott, C. Watts, J. Cell Biol. 1994, 124, 689.[60] D. G. Perry, G. L. Daugherty, W. J. Martin, J. Immunol. 1999,
162, 380.[61] R. G. Parton, B. Joggerst, K. Simons, J. Cell Biol. 1994, 127,
1199.[62] H. Hillaireau, P. Couvreur, Cell. Mol. Life. Sci. 2009, 66, 2873.[63] S. Mukherjee, R. N. Ghosh, F. R. Maxfield, Endocytosis Physiol.
Rev. 1997, 77, 759.[64] L. M. Bareford, P. W. Swaan, Adv. Drug Delivery Rev. 2007, 59,
748.
13, 13, 1413–1421
bH & Co. KGaA, Weinheim 1421