stable isotope profiles in whale shark (rhincodon typus) suggest segregation and dissimilarities in...
TRANSCRIPT

Stable isotope profiles in whale shark (Rhincodon typus)suggest segregation and dissimilarities in the diet dependingon sex and size
Asunción Borrell & Alex Aguilar & Manel Gazo &
R. P. Kumarran & Luis Cardona
Received: 13 March 2010 /Accepted: 27 June 2011 /Published online: 7 July 2011# Springer Science+Business Media B.V. 2011
Abstract We investigated the sex- and size-relateddifferences in the diet of whale sharks from theArabian Sea (north-western Indian Ocean) usingcarbon and nitrogen stable-isotope analyses in whitemuscle. The samples were collected during thecommercial fishing season between April and May of2001 in Veraval (Gujarat, India). The overall isotopesignature was similar to that of the pelagic-neriticzooplanktivore Ilisha melastoma, which suggests thatboth species are feeding on similar prey. In whalesharks, a positive relationship was found between δ15Nand δ13C. This, together with a significant enrichmentof both heavy stable isotopes with total length indicatesthat the contribution to the diet of small fish and/orlarger zooplankton of higher trophic level increaseswith the movement from offshore areas to coastal areasas they grow. Gender differences in the isotopic ratioswere not statistically significant, but small sample sizecannot rule out completely the existence of somedegree of spatial or dietary segregation between sexes.
Keywords Habitat selection . Arabian Sea . IndianOcean .Whale shark . δ13C . δ15N
Introduction
The whale shark, Rhincodon typus, is the largest fishon Earth, with a maximum length of 20°m reported inTaiwan (Chen et al. 2002). It is found in all oceansfrom tropical to temperate latitudes (generally be-tween 30°N and 35°S), where it occupies bothoceanic and coastal waters, including the lagoons ofcoral atolls and reefs (Colman 1997). It migrates overthousands of kilometres (Eckert and Stewart 2001;Sleeman et al. 2010), and its movements are related tolocal productivity (Wilson et al. 2001; Stevens 2007;but see Sleeman et al. 2010). Its occurrence is oftenassociated with schools of pelagic fish that may feedon similar prey items (Compagno 1984).
In the 1990s, whale sharks from the Arabian Sea(north-western Indian Ocean) were caught annuallyfrom March to June in the Gujarat coasts. The numberof whale sharks caught during the 1988–1998 periodwas at least of 1823 units only in Verabal (Pravin2000). However, the catches declined in 2000,probably because of the excessive fishing pressure(Hanfee 2001). In 2001, the Government of Indiagranted protection from fishing to the whale sharkthrough the Wildlife (Protection) Act (Dutta 2001)and, in 2002, the Convention on Migratory Speciesincluded the species in Appendix II (IUCN 2003) thus
Environ Biol Fish (2011) 92:559–567DOI 10.1007/s10641-011-9879-y
A. Borrell (*) :A. Aguilar :M. Gazo : L. CardonaIrBio; Institut de Recerca de la Biodiversitat andDepartment of Animal Biology, Faculty of Biology,University of Barcelona,Avinguda Diagonal 645,08028 Barcelona, Spaine-mail: [email protected]
R. P. KumarranMullai Nagar, Anna Nagar,Chennai 600040, India

implementing restrictions on its international com-merce. However, poaching activities and illegal tradecontinued on small scale in the region (Riley et al.2009).
Despite the short but intense period in which thewhale shark was commercially exploited, the mainbiological characteristics, which could be investigatedfrom individuals captured by the fishery, remainunknown, particularly habitat use and diet composi-tion. The sparse available information indicates thatwhale sharks are probably born in either open oceanor deep waters and that juveniles move closer to theshore to form coastal aggregations that may be sex-biased, often in favour of males (Eckert and Stewart2001; Graham and Roberts 2007; Rowat et al. 2007;Rowat et al. 2008; Hobbs et al. 2009).
Whale sharks are generally considered suction-filter feeders that forage on dense aggregations ofzooplankton. However, other organisms such as algae,squids, tunas and other nektonic species of medium orlarge size have been identified as potential prey(Compagno 1984; Taylor 1994; Norman 1999; Wilsonand Newbound 2001; Duffy 2002; Stevens 2007).This information comes from the examination of thestomach contents of opportunistically studied strand-ed individuals (Compagno 1984; Last and Stevens1994; Colman 1997) and from observations ofindividuals from coastal aggregations while feeding(Clark and Nelson 1997; Heyman et al. 2001; Duffy2002; Nelson and Eckert 2007; Rowat et al. 2007).The sample size of stomach content studies isinvariably limited to a single individual. Moreover,although these analyses offer a high degree oftaxonomic precision, they provide only a snapshot intime of the consumers’ diet, and they are, therefore,limited in scope (Cortés 1999). In addition, stomachcontents may contain incidentally ingested items suchas plant material or mangrove seedpods (Beckley etal. 1997). On the other hand, the prey consumedwhile engaged at the coastal feeding aggregations isof unclear relevance because whale sharks are highlymobile and spend only a short time at any particularlocation (Eckert and Stewart 2001; Eckert et al. 2002;Wilson et al. 2006; Graham and Roberts 2007; Rowatet al. 2007; Sleeman et al. 2010).
The analysis of stable isotopes offers an alternativemethod for investigating diet compositions and hasproved reliable for establishing trophic levels in manyspecies including sharks (Rau et al. 1983; Fisk et al.
2002; Estrada et al. 2003; Domi et al. 2005; MacNeilet al. 2005; Estrada et al. 2006). This technique doesnot produce species-specific dietary information butprovides a measure of the assimilated prey over aperiod of time, depending on the tissue analysed, andcomplete isotope turnover is muscle shark may takemore than 2 years (Logan and Lutcavage 2010).
Animal tissues are typically more enriched in theheavy isotopes of both nitrogen (15N) and carbon(13C) than their food source; as a result, the values ofthese isotopes increase with trophic level (DeNiro andEpstein 1978; DeNiro and Epstein 1981; Minagawaand Wada 1984; Peterson and Fry 1987; Cabana andRasmussen 1996). This effect is more pronounced innitrogen (N) than in carbon (C), and, in the muscle oflarge sharks, it leads to increases of ~2.3‰ in δ15Nand of ~0.9‰ in δ13C above their prey items (Husseyet al. 2010).
Moreover, the primary producers of marine eco-systems show strong spatial gradients in isotopiccarbon ratios (Fry and Wainright 1991; Hemmingaand Mateo 1996; Rau et al. 2001) with valuestypically increasing from offshore to near-shoreecosystems and peaking in macrophyte dominatedecosystems (kelp and seagrass beds). This pattern ofvariation makes C isotopic ratios useful for investi-gating habitat use by predatory species (Burton andKoch 1999).
In this paper, we determined the stable isotopeprofiles of C and N in the muscle of 19 whale sharkscaught in Gujarat, India, to investigate potentialintrapopulation segregation in feeding habitats anddifferences in migratory patterns associated to the sizeand sex of the sharks.
Material and methods
Sampling
The samples used in this study were obtained fromthe whale shark fishing operations off the coast ofVeraval in Gujarat, India, (Fig. 1). During April andMay of 2001, we examined 19 whale sharks that hadbeen caught with hooks and lines and subsequentlytowed to the shore. We do not know the exactlocations where these sharks were captured but whalesharks captured in previous years were caught in areasshallower than 70 m (Pravin 2000). We determined
560 Environ Biol Fish (2011) 92:559–567

the sex and the standard length (SL) before they werecut into pieces while afloat in the water. Once thebody parts were brought to shore, we collected forisotopic determination a sample of white muscle nextto the dorsal fin and below the collagen-like materialand the red muscle (Table 1). The total length (TL)was calculated using the equation TL=1.125*SL+0.0203, as described in Wintner (2000).
We also collected muscle from two teleost fish of wellknown feeding habits, captured from the same area. Theywere 1) the longarmmullet, Valamugil cunnesius (n=10;TL range 10.6–12.5 cm), which is a benthic, coastalfish that mainly feeds on decaying organic matter(Sommer et al. 1996), and 2) the Indian ilisha, Ilishamelastoma, (n=10; TL range 20–28 cm), a pelagiczooplanktivorous fish (Whitehead 1985; Carpenter et al.
Date of capture Standard length (m) Total length (m) Sex δ 15 N δ 13C C:N
11-apr-01 8.1 10.2 F 15.3 −15.1 3.2
11-apr-01 9.0 11.3 F 13.3 −16.4 4.1
12-apr-01 6.0 7.5 F 13.8 −16.2 3.1
12-apr-01 6.0 7.5 F 12.0 −16.3 3.0
17-apr-01 3.2 4.0 M 11.2 −17.9 2.8
30-apr-01 6.8 8.5 ? 14.7 −14.7 3.3
30-apr-01 4.9 6.2 ? 13.6 −16.2 3.7
01-may-01 6.3 7.9 M 15.2 −15.4 3.7
04-may-01 4.2 5.3 ? 15.2 −16.1 4.5
07-may-01 7.0 8.8 F 15.3 −15.7 3.8
08-may-01 15.0 18.8 F 15.1 −15.7 3.9
08-may-01 4.9 6.2 F 15.2 −16.8 3.3
08-may-01 5.9 7.4 M 15.5 −15.5 3.9
08-may-01 9.0 11.3 M 14.6 −15.1 3.6
08-may-01 ? ? M 15.5 −15.5 3.6
09-may-01 4.0 5.0 F 13.2 −16.1 3.3
09-may-01 5.4 6.8 F 14.6 −15.9 4.1
10-may-01 2.4 3.0 F 11.0 −17.0 3.1
11-may-01 6.3 7.9 F 12.0 −17.4 3.4
Table 1 Biological charac-teristics of the sampledwhale sharks and results ofthe stable isotope analysesconducted in their muscle
Fig. 1 Sampling location
Environ Biol Fish (2011) 92:559–567 561

1997). As a consequence of their diet, these teleostswere expected to differ both in the δ15N (higher for theilisha than for the mullet) and in the δ13C (higher for themullet than for the ilisha) and were therefore consideredto provide a suitable baseline for interpreting the stableisotope values of whale sharks.
The collected samples were initially preserved incooking salt, a method considered suitable forpreserving fish muscle tissue (Arrington and Wine-miller 2002). Once in the laboratory, the sampleswere soaked for 30 h in distilled water and thenrinsed to remove the salt; subsequently, they werelong-term preserved at −25°C.
Stable isotope analysis
Prior to analysis, the muscle tissue (1 g) was dried for3 days at 70°C. After being ground, its lipids wereremoved by rinsing the ground tissue several timeswith a 2:1 chloroform: methanol mixture followingthe Folch method (Folch et al. 1957). Approximately1 mg of the powdered sample was weighed in tincapsules, automatically-loaded and combusted at1000°C to be analysed in a continuous flow isotoperatio mass spectrometer (Flash 1112 IRMS Delta CSeries EA Thermo Finnigan).
The results were presented according to the delta (δ)notation, where the relative variation of stable isotoperatios are expressed in parts-per-thousand from prede-fined standards. This variation is calculated as:
d ¼ RS RR=ð Þ � 1½ �»1000where RS is the ratio of the heavy isotope to the lightisotope of the sample, and RR is the ratio of the heavyisotope to the light isotope in the reference.
The Rstandard for13C and 15N are the Vienna Peedee
Belemnite (V-PDB) standard and atmospheric nitrogen(air), respectively. The international isotope secondarystandards of known 13C/12C ratios in relation toV-PDB, namely, polyethylene (IAEA CH7; δ13
C=−31.8‰), graphite (USGS24; δ13C=−16.1‰) andsucrose (IAEA-CH6; δ13C=−10.4‰), were used forthe calibration of δ13C at a precision of 0.2‰. Fornitrogen, the international isotope secondary standardsof known 15N/14N ratios in relation to air, namely,ammonium sulphate (IAEA N1; δ15N=+0.4‰ andIAEA N2; δ15N=+20.3‰) and potassium nitrate(IAEA NO3; δ15N=+4.7‰), were used for thecalibration of δ15N to a precision of 0.3‰. Atropine
(70.56%C, 4.84%N) was used as a standard forelemental composition of C and N. The experimentalprecision based on the standard deviation of replicatesof an atropine standard was 0.3‰ for both carbon andnitrogen.
The reference materials used for the analysis aredistributed by the International Atomic Energy Agency(IAEA). The analyses were carried out in the laborato-ries of the University of Barcelona.
Statistics
Prior to any statistical analysis, the data were testedfor normality with a Kolmogorov-Smirnov test ofgoodness of fit and for homogeneity of variances withLevene’s test. As all data sets were normallydistributed, the differences between groups wereinvestigated using the Student’s t-test. To investigatesex-related variation, we only used the subsample ofwhale sharks that measured more than 4.0 m (i.e.,three with known lengths of 7.4, 7.9, and 11.3 m andone with unknown length) and six females of similarsize (i.e., 7.5, 7.5, 7.9, 8.8, 10.2 and 11.3 m). A simpleleast squares linear regression was used to determine therelationships between δ15N and δ13C and betweenisotopic signatures and the total shark length afterlogarithmic transformation. For the latter analyses thelength values were converted to loge of length becausethe relationship of length with delta values fitted betteran exponential than a linear relationship.
All the statistical calculations were carried outusing the SPSS-15 statistical package.
Results
The main biological characteristics, the δ15N and δ13Cvalues and the C:N ratio of the muscle tissue of thesharks analysed are summarised in Table 1. The meanvalue of the C:N ratio was 3.5 (SD ±0.43), whichindicates that the lipid extraction process was effectivein the tissues analysed. The stable isotope ratios in themuscle tissue of whale shark ranged from −17.9to −14.7‰ for δ13C and from 11 to 15.5‰ for δ 15N(Table 1, Fig. 2); this implied a difference between theextreme values of 4.5‰ in δ15N and of 3.2‰ in δ13C.A statistically significant and positive correlation wasfound between the δ15N and δ13C values (R2=0.6; p<0.001; Fig. 2).
562 Environ Biol Fish (2011) 92:559–567
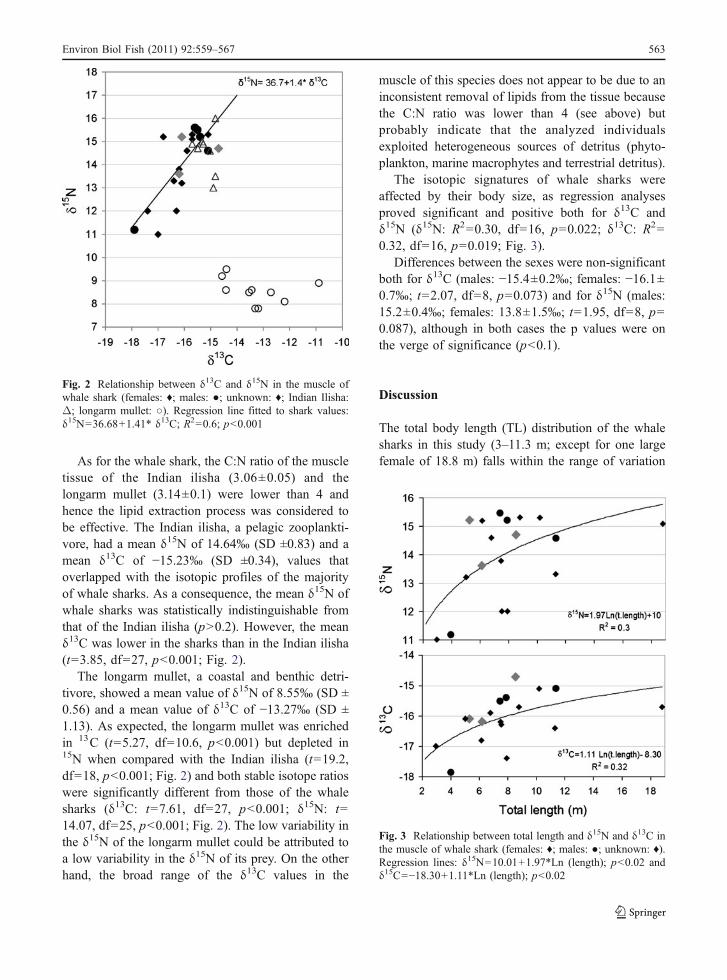
As for the whale shark, the C:N ratio of the muscletissue of the Indian ilisha (3.06±0.05) and thelongarm mullet (3.14±0.1) were lower than 4 andhence the lipid extraction process was considered tobe effective. The Indian ilisha, a pelagic zooplankti-vore, had a mean δ15N of 14.64‰ (SD ±0.83) and amean δ13C of −15.23‰ (SD ±0.34), values thatoverlapped with the isotopic profiles of the majorityof whale sharks. As a consequence, the mean δ15N ofwhale sharks was statistically indistinguishable fromthat of the Indian ilisha (p>0.2). However, the meanδ13C was lower in the sharks than in the Indian ilisha(t=3.85, df=27, p<0.001; Fig. 2).
The longarm mullet, a coastal and benthic detri-tivore, showed a mean value of δ15N of 8.55‰ (SD ±0.56) and a mean value of δ13C of −13.27‰ (SD ±1.13). As expected, the longarm mullet was enrichedin 13C (t=5.27, df=10.6, p<0.001) but depleted in15N when compared with the Indian ilisha (t=19.2,df=18, p<0.001; Fig. 2) and both stable isotope ratioswere significantly different from those of the whalesharks (δ13C: t=7.61, df=27, p<0.001; δ15N: t=14.07, df=25, p<0.001; Fig. 2). The low variability inthe δ15N of the longarm mullet could be attributed toa low variability in the δ15N of its prey. On the otherhand, the broad range of the δ13C values in the
muscle of this species does not appear to be due to aninconsistent removal of lipids from the tissue becausethe C:N ratio was lower than 4 (see above) butprobably indicate that the analyzed individualsexploited heterogeneous sources of detritus (phyto-plankton, marine macrophytes and terrestrial detritus).
The isotopic signatures of whale sharks wereaffected by their body size, as regression analysesproved significant and positive both for δ13C andδ15N (δ15N: R2=0.30, df=16, p=0.022; δ13C: R2=0.32, df=16, p=0.019; Fig. 3).
Differences between the sexes were non-significantboth for δ13C (males: −15.4±0.2‰; females: −16.1±0.7‰; t=2.07, df=8, p=0.073) and for δ15N (males:15.2±0.4‰; females: 13.8±1.5‰; t=1.95, df=8, p=0.087), although in both cases the p values were onthe verge of significance (p<0.1).
Discussion
The total body length (TL) distribution of the whalesharks in this study (3–11.3 m; except for one largefemale of 18.8 m) falls within the range of variation
Fig. 3 Relationship between total length and δ15N and δ13C inthe muscle of whale shark (females: ♦; males: ●; unknown: ♦).Regression lines: δ15N=10.01+1.97*Ln (length); p<0.02 andδ15C=−18.30+1.11*Ln (length); p<0.02
Fig. 2 Relationship between δ13C and δ15N in the muscle ofwhale shark (females: ♦; males: ●; unknown: ♦; Indian Ilisha:Δ; longarm mullet: ○). Regression line fitted to shark values:δ15N=36.68+1.41* δ13C; R2=0.6; p<0.001
Environ Biol Fish (2011) 92:559–567 563

previously observed both in India (n:164; range:3.15–14.5 m; modal length: 5.1–6.0 m) and world-wide (most specimens reported to measure 4–12 m)(Compagno 1984; Colman 1997; Pravin 2000; Duffy2002; Graham and Roberts 2007; Norman andStevens 2007). However, several authors havereported a maximum length of 20 m for this species(Compagno 2001; Chen et al. 2002). Sexual maturityis reached at a TL of about 9 m (Colman 1997;Norman and Stevens 2007), which means that themajority of the individuals in this study were stillimmature. This is consistent with the fact that inVeraval the fishery was conducted with relativelysmall boats which towed the catch to shore, andtherefore could not operate far away from the coast,and that immature whale sharks prevail in coastallocations worldwide (Eckert and Stewart 2001; Eckertet al. 2002; Meekan et al. 2006; Wilson et al. 2006;Graham and Roberts 2007; Rowat et al. 2007; Hobbset al. 2009). This indicates that this fishery exploitedan on-shore fishing ground, as previously suggestedby Hanfee (2001), although the exact locationremained unknown for the authors. However, in otherlocations such aggregations of immatures are mostlycomposed of males shorter than 9 m of TL, while oursample was dominated by females. This may indicatea female-biased sex ratio in the area, a featureinfrequently described in other localities (Eckert andStewart 2001; Hobbs et al. 2009).
The analysis of stable isotopes indicated that theoverall δ15N values for all the species in the currentstudy was high, possibly resulting from the influenceof the high levels of organic pollution that occur inthe coastal ecosystem of the Gujarat coast. Pollutionis particularly intense in the waters around Veraval,due to extensive raw urban and industrial sewage aswell as by runoff of agricultural products (Zingde2005). Thus, previous studies in similarly organicpolluted areas in the Adriatic Sea have shown ahigher than expected enrichment of the δ15N signal incoastal invertebrates such as Anemonia sulcata(Dolenec et al. 2005).
When isotopic values of whale sharks caught inVeraval are compared with those of other componentsof the food web, the results corroborate that, at leastin the Arabian Sea, the species has a primarilyzooplanktivorous diet. Thus, despite the wide spec-trum of prey that it is known to consume (Compagno1984; Taylor 1994; Colman 1997; Norman 1999;
Wilson and Newbound 2001; Duffy 2002) its stableisotope profile is close to that of the Indian ilisha.However, the positive relationship between δ15N andδ13C (explaining 60% of the variance) and therelationship between each isotope ratio and the totallength of sharks (explaining 30% of the variance) allsuggest that, as the size of the sharks increases, thecontribution to the diet of small fish and/or ofzooplankton of larger size and higher trophic levelincreases. The lack of highly developed filteringstructures on the gill-rakers in neonatal whale sharkscompared to adults (Garrick 1964) could explain thelow ability of small individuals to forage on largerprey. Indeed, studies on the intrapopulation variabilityof isotope signatures in fish frequently show apositive relationship between δ15N values and size, arelationship that is commonly attributed to ontogenet-ic or size-based changes in diet (Overman and Parrish2001).
Nevertheless, a reduction with age of the 15Ntissue-diet fractionation factor, as described in otherspecies (Focken 2001; Gaye-Siessegger et al. 2003;Trueman et al. 2005), may also contribute to theobserved increase of δ15N with total length. Indeed,this effect would explain why the slope of increasewith size of δ15N, and to a lesser degree of δ13C, wasmore abrupt in the group of smaller, growing fasterindividuals than in those of larger size and compar-atively lower growth rates. Whale shark neonatallength is approximately 50 cm±10 (Joung et al. 1996)and the growth rate is extremely high during the fewyears after birth (more than 100 cm per year) but,after reaching 3–4 m in length, it slows down byabout 75% (Chang et al. 1997; Hsu et al. 2000;Uchida et al. 2000; Nishida 2001). In this growingscenario, the sharp increase of muscle δ15N values insharks 3–4 m or longer may be partially caused bythis effect.
On the other hand, the increase of δ13C with totalsize is likely attributable to intraspecific differences inmovement patterns and heterogeneities in the locationwhere individuals forage. Thus, whale sharks smallerthan ca 4 m showed a δ13C of about −17‰, while inlarger individuals it increased to about −15‰, achange that indicates a transition from a pelagicoffshore life to a relatively more coastal habitat.
Despite the lack of knowledge of where and whenwhale sharks give birth or about the way how earlylife develops, the fact that all neonatal captures have
564 Environ Biol Fish (2011) 92:559–567

occurred in, or very close to, deep waters seems toindicate that nursery areas are located offshore (Wolfson1983; Kukuyev 1995; Rowat et al. 2008). This appearsto be confirmed by the scarcity of records of whalesharks of <4 m in length, especially in the IndianOcean (Rowat et al. 2008), which would suggest thatneonates and very small whale sharks frequent deeperwaters than their larger conspecifics. Such preferencemay be due to natal philopatry, a behaviour describedin other species of sharks (Hueter et al. 2005) or,alternatively, to a protective behaviour of very youngwhale sharks, which would remain in deep waters toavoid predators (Rowat et al. 2008).
When comparing sharks of similar total length,differences between sexes in the stable isotope signalwas found to be marginally non-significant (p<0.1), aresult likely to be due to the low statistical power ofthe small-sized sample than to an actual homogeneityin signatures (Hoem, 2008). Thus, males appeared tohave higher values of δ13C and δ15N than females.Virtually nothing is known about abundance, segre-gation and migration patterns of the whale shark andwhether there are sex-related differences in these traits(Taylor 1996; Colman 1997; Heyman et al. 2001;Wilson and Newbound 2001; Duffi 2002; Stewart andWilson 2005; Rowat 2007), but coastal aggregationsare often observed to be sex-biased (Eckert andStewart 2001; Wilson et al. 2006; Graham andRoberts 2007; Rowat et al. 2007). In our case,regardless of isotope signal signatures, the higherproportion of females in the catch in a coastal fishery,as discussed above, would support the hypothesis ofspatial segregation between sexes.
Sex and size segregation have been observed inother shark species (Bres 1993; Sims 2005). Thus,tagged individuals and DNA studies have establishedthat male white sharks (Carcharodon carcharias) mayundertake transoceanic movements, whereas femalestend to remain in the coastal waters of the continentwhere they had been born (Pardini et al. 2001;Boustany et al. 2002); likewise, intermediate-sizefemales of scalloped hammerhead sharks (Sphyrnalewini) have been observed to form schools thatsegregate from males and feed more frequently onpelagic prey (Klimley 1987). Like other sharks, whalesharks may perform either short distance (Gunn et al.1999) or large-scale migrations (Eckert et al. 2002;Schmidt et al. 2009). These latter migrations may takeyears to complete, and their range seems to be
governed by the timing and location of productionpulses as well as by breeding behaviour, which differsbetween sexes (Colman 1997; Eckert and Stewart2001). Further research, potentially conductedthrough skin biopsies obtained from free-rangingindividuals, should be made to clarify whether thesedifferential patterns of movement result in actualsegregation between sexes in foraging habitat.
Acknowledgments We acknowledge the Fundació pelDesenvolupament Sostenible (FDS) for providing funds forthe sampling fieldwork, to the Serveis Cientifico Tècnics (SCT)of the University of Barcelona for analytical assistance, and tothe Vicerectorate of International Relations of the University ofBarcelona for travel funding during the elaboration andcorrection of the manuscript.
References
Arrington DA, Winemiller KO (2002) Preservation effects onstable isotope analysis of fish muscle. Trans Am Fish Soc131:337–342
Beckley LE, Cliff G, Smale MJ, Compagno LJV (1997) Recentstrandings and sightings of whale sharks in South Africa.Environ Biol Fish 50:343–348
Boustany AM, Davis SF, Pyle P, Anderson SD, Le Boeuf BJ,Block BA (2002) Expanded niche for white sharks. Nature415:35–36
Bres M (1993) The behaviour of sharks. Rev Fish Biol Fish3:133–159
Burton RK, Koch PL (1999) Isotopic tracking of foraging andlong distance migration in northeastern Pacific pinnipeds.Oecologia 119:578–585
Cabana G, Rasmussen JB (1996) Comparison of aquatic foodchains using nitrogen isotopes. Proc Natl Acad Sci USA93:10844–10847
Carpenter K, Krupp F, Jones DA, Zajonz W (1997) Livingmarine resources of Kuwait, eastern Saudi Arabia,Bahrain, Qatar, and the United Arab Emirates. FAO, Rome
Chang WB, Leu M, Fang LS (1997) Embryos of the whaleshark, Rhincodon typus: early growth and size distribution.Copeia 97:444–446
Chen CT, Liu KM, Joung SJ (2002) Preliminary report onTaiwan’s whale shark fishery. In: Fowler SL, Reed TM,Dipper FA (eds) Elasmobranch biodiversity, conservationand management: proceedings of the international seminarand workshop, Sabah, Malaysia, July 1997 IUCN SSC.Shark Specialist Group, IUCN, Gland, Switzerland, pp162–167
Clark E, Nelson DR (1997) Young whale sharks, Rhincodontypus, feeding on a copepod bloom near La Paz, Mexico.Environ Biol Fish 50:63–73
Colman JG (1997) A review of the biology and ecology of theWhale Shark. J Fish Biol 51:1219–1234
Compagno LJV (1984) Sharks of the world. An annotated andillustrated catalogue of shark species known to date.
Environ Biol Fish (2011) 92:559–567 565

Carcharhiniformes. FAO species catalogue for fisherypurposes 1, vol. 4, 269 FAO, Rome pp 251–655
Compagno LJV (2001) Sharks of the world. An annotated andillustrated catalogue of shark species known to date.Bullhead, mackerel and carpet sharks (Heterodontiformes,Lamniformes and Orectolobiformes). FAO species cata-logue for fishery purposes 1, vol. 2, 269 pp FAO, Rome
Cortés E (1999) Standardized diet compositions and trophiclevels of sharks. ICES J Mar Sci 56:707–717
DeNiro MJ, Epstein S (1978) Influence of diet on thedistribution of carbon isotopes in animals. GeochimCosmochim Acta 42:495–506
DeNiro MJ, Epstein S (1981) Influence of diet on thedistribution of nitrogen isotopes in animals. GeochimCosmochim Acta 45:341–351
Dolenec T, Vokal B, Dolenec M (2005) Nitrogen—15 signals ofanthropogenic nutrient loading in Anemonia sulcata as apossible indicator of human sewage impacts on marinecoastal ecosystems: a case study of Pirovac Bay and theMurter Sea (Central Adriatic). Croat Chem Acta 78(4):593–600
Domi N, Bouquegneau J, Das K (2005) Feeding ecology of fivecommercial shark species of the Celtic Sea through stableisotope and trace metal analysis. Mar Environ Res60:551–569
Duffy CAJ (2002) Distribution, seasonality, lengths, andfeeding behaviour of whale sharks (Rhincodon typus)observed in New Zealand waters. NZ J Mar Fresh Res36:565–570
Dutta R (2001) Whale Sharks. The gentle giants of the Sea.Traffic Dispatches 17:10
Eckert SA, Stewart BS (2001) Telemetry and satellite trackingof whale sharks, Rhincodon typus, in the Sea of Cortez,Mexico, and the north Pacific Ocean. Environ Biol Fish60:299–308
Eckert SA, Dolar LL, Kooyman GL, Perrin W, Ridzwan RA(2002) Movements of whale sharks (Rhincodon typus) inSoutheast Asia waters as determined by satellite telemetry.J Zool (Lond) 257:111–115
Estrada JA, Rice AN, Lutcavage ME, Skomal GB (2003)Predicting trophic position in sharks of the north-westAtlantic Ocean using stable isotope analysis. J Mar BiolAss UK 83:1347–1350
Estrada JA, Rice AN, Natanson LJ, Skomale GB (2006) Use ofisotopic analysis of vertebrae in reconstructing ontogeneticfeeding ecology in white sharks. Ecology 87:829–834
Fisk AT, Tittlemier ShA, Pranschke JL, Norstrom RJ (2002)Using anthropogenic contaminants and stable isotopes toassess the feeding ecology of Greenland sharks (Somnio-sus microcephalus). Ecology 83:2162–2172
Focken U (2001) Stable isotopes in animal ecology: the effectof ration size on the trophic shift of C and N isotopesbetween feed and carcass. Isot Environ Health Stud37:199–211
Folch J, Lees M, Sloane Stanley GH (1957) A simple methodfor the isolation and purification of total lipides fromanimal tissues. J Biol Chem 226:497–509
Fry B, Wainright SC (1991) Diatom sources of 13C-rich carbonin marine food webs. Mar Ecol Progr Ser 76:149–158
Garrick JAF (1964) Additional information on the morphologyof an embryo whale shark. Proc US Nat Mus 115:10
Gaye-Siessegger J, Focken U, Abel HJ, Becker K (2003)Feeding level and diet quality influence trophic shift of Cand N isotopes in Nile tilapia (Oreochromis niloticus L.).Isot Environ Health Stud 39:125–134
Graham RT, Roberts CM (2007) Assessing the size, growth rateand structure of a seasonal population of whale sharks(Rhincodon typus Smith 1828) using conventional taggingand photo identification. Fish Res 84:71–80
Gunn JS, Stevens JD, Davis TLO, Norman BM (1999)Observations on the short-term movements and behaviourof whale sharks (Rhincodon typus) at Ningaloo Reef,Western Australia. Mar Biol 135:553–559
Hanfee F (2001) Trade in Whale shark and its products in thecoastal state of Gujarat, India. Report to the RuffordFoundation from TRAFFIC India
Hemminga MA, Mateo MA (1996) Stable carbon isotopes inseagrasses: variability in ratios and use in ecologicalstudies. Mar Ecol Prog Ser 140:285–298
Heyman WD, Graham RT, Kjerfve B, Johannes RE (2001)Whale sharks Rhincodon typus aggregate to feed on fishspawn in Belice. Mar Ecol Prog Ser 215:275–282
Hobbs JPA, Frisch AJ, Hamanaka T, McDonald CA, GilliganJJ, Neilson J (2009) Seasonal aggregation of juvenilewhale sharks (Rhincodon typus) at Christmas Island,Indian Ocean. Coral Reefs 28:577–577
Hoem J (2008) The reporting of statistical significance inscientific journals. Demographic Res 18:437–442
Hsu CK, Chang TY, Zu HC, Huang KF, Tsai WS (2000)Preliminary attempt on Whale Shark ranching (in Chi-nese). TFRI Newsletter 143:6–11
Hueter RE, Heupel MR, Heist EJ, Keeney DB (2005)Evidence of philopatry in sharks and implications forthe management of shark fisheries. J Northw Atl FishSci 35:239–247
Hussey NE, Brush J, McCarthy ID, Fisk AT (2010) δ15N andδ13C diet–tissue discrimination factors for large sharksunder semi-controlled conditions. Comp Biochem Physiol155:445–453
IUCN (2003) Inclusion of whale shark Rhincodon typus inAppendix II. Proponent: India and Philippines. TheIUCN/TRAFFIC Analyses of Proposals to Amend theCITES Appendices Prepared for the 12th meeting of theConference of the Parties (CoP12) Santiago, Chile, 3–15
Joung SJ, Chen CT, Clark E, Uchida S, Huang WYP (1996)The whale shark, Rhincodon typus, is a livebearer: 300embryos found in one ‘megamamma' supreme. EnvironBiol Fish 46:219–223
Klimley AP (1987) The determinants of sexual segregation inthe scalloped hammerhead shark, Sphyrna lewini. EnvironBiol Fish 18:27–40
Kukuyev EI (1995) The new finds in recently born individualsof the whale shark Rhiniodon typus (Rhiniodontidae) inthe Atlantic Ocean. J Ichthyol 36:203–205
Last PR, Stevens JD (1994) Sharks and Rays of Australia.CSIRO, Australia
Logan JM, Lutcavage ME (2010) Stable isotope dynamics inelasmobranch fishes. Hydrobiol 644:231–244
MacNeil MA, Skomal GB, Fisk AT (2005) Stable isotopes frommultiple tissues reveal diet switching in sharks. Mar EcolProg Ser 302:199–206
566 Environ Biol Fish (2011) 92:559–567

Meekan MG, Bradshaw CJA, Press M, McLean C, Richards A,Quasnichka S, Taylor JA (2006) Population size andstructure of whale sharks (Rhincodon typus) at NingalooReef, Western Australia. Mar Ecol Prog Ser 319:275–285
Minagawa M, Wada E (1984) Stepwise enrichment of 15Nalong food chains: further evidence and the relationbetween δ15N and animal age. Geochim CosmochimicActa 48:1135–1140
Nelson J, Eckert S (2007) Foraging ecology by whale sharks(Rhincodon typus) within Bahia de los Angeles, BajaCalifornia Norte, Mexico. Fish Res 84:47–64
Nishida K (2001) Whale shark—the world's largest fish. In:Kakabo T, Machida Y, Yamaoka K, Nishida K (eds) Fishesof the Kuroshio Current, Japan. Osaka Aquarium KAIYU-KAN, Minato-ku, Osaka, pp 20–26
Norman BM (1999) Aspects of the biology and ecotourismindustry of the Whale Shark Rhincodon typus in North-western Australia. MPhil. Thesis. Murdoch University,Western Australia
Norman BM, Stevens JD (2007) Size and maturity status of thewhale shark (Rhincodon typus) at Ningaloo Reef inWestern Australia. Fish Res 84:81–86
Overman NC, Parrish DL (2001) Stable isotope composition ofwalleye: 15N accumulation with age and area-specificdifferences in 13C. Can J Fish Aquatic Sci 58:1253–1260
Pardini AT, Jones CS, Noble LR, Kreiser B, Malcolm H, BruceBD, Stevens JD, Cliff G, Scholl MC, Francis M, DuffyCAJ, Martin AP (2001) Sex-biased dispersal of greatwhite sharks. Nature 412:139–140
Peterson BJ, Fry B (1987) Stable isotopes in ecosystem studies.Annu Rev Ecol Syst 18:293–320
Pravin P (2000) Whale shark in the Indian coast—Need forconservation. Curr Sci 79:310–315
Rau GH, Mearns AJ, Young DR, Olson RJ, Schafer HA,Kaplan IR (1983) Animal 13C/12C correlates with trophiclevel in pelagic food webs. Ecology 64:1314–1318
Rau GH, Chavez FP, Andfriederich GE (2001) Plankton13C/12C variations in Monterey Bay, CA: evidence ofnon-diffusive inorganic carbon uptake by phytoplankton inan upwelling environment. Deep Sea Res 48:79–94
Riley MJ, Harman A, Rees RG (2009) Evidence of continuedhunting of whale sharks Rhincodon typus in the Maldives.Environ Biol Fish 86:371–374
Rowat D (2007) Occurrence of whale shark (Rhincodon typus)in the Indian Ocean: a case for regional conservation. FishRes 84:96–101
Rowat D, Meekan MG, Engelhardt U, Pardigon B, Vely M(2007) Aggregation of juvenile whale shark (Rhincodontypus) in the Gulf of Tadjoura, off Djibouti. Environ BiolFish 80:465–472
Rowat D, Gore MA, Baloch BB, Islam Z, Ahmed E, Ali QM,Culloch R, Hameed S, Hasnain SA, Hussain B, Kiani S,Siddiqui J, Ormond RF (2008) New records of neonatalwhale shark (Rhincodon typus) from the Northern IndianOcean. Environ Biol Fish 82:215–219
Schmidt JV, Schmidt CL, Ozer F, Ernst RE, Feldheim KA,Ashley MV, Levine M (2009) Low genetic differentiationacross three major ocean populations of the Whale Shark,Rhincodon typus. PLoS ONE; 4(4):e4988. doi:10.1371/journal.pone.0004988
Sims DW (2005) Differences in habitat selection and repro-ductive strategies of male and female sharks. In: RuckstuhlKE, Neuhaus P (eds) Sexual segregation in vertebrates:ecology of the two sexes. Cambridge University Press,Cambridge, pp 127–14
Sleeman JC, Meekan MG, Wilson SG, Polovina JJ, Stevens JD,Boggs GS, Bradshaw CJA (2010) To go or not to go withthe flow: environmental influences on whale sharkmovement patterns. J Exp Mar Ecol Biol 390:84–98
Sommer CW, Schneider J, Poutiers M (1996) FAO speciesidentification field guide for fishery purposes. The livingmarine resources of Somalia. FAO, Rome, 376
Stevens JD (2007) Whale shark (Rhincodon typus) biologyand ecology: a review of the primary literature. Fish Res84:4–9
Stewart B, Wilson S (2005) Threatened Fishes of the World:Rhincodon typus (Smith 1828) (Rhincodontidae). EnvironBiol Fish 74:184–185
Taylor G (1994) Whale Sharks. Angus & Robertson Publishers,Sydney, Australia
Taylor JG (1996) Seasonal occurrence, distribution andmovements of the whale shark, Rhincodon typus, atNingaloo reef, Western Australia. Mar Freshwater Res47:637–642
Trueman CN, McGill R, Guyard PH (2005) The effect ofgrowth rate on tissue-diet isotopic spacing in rapidlygrowing animals. An experimental study with Atlanticsalmon (Salmo salar). Rapid Comm Mass Spec 19:3239–3247
Uchida S, Toda M, Kamei Y, Teruya H (2000) The husbandry of16 whale sharks Rhincodon typus, from 1980 to 1998 atthe Okinawa expo aquarium. American ElasmobranchSociety 16th Annual Meeting June 14–20, 2000 La Paz,B.C.S., México, U.S.A. Okinawa Expo Aquarium, Oki-nawa, Japan
Whitehead PJP (1985) FAO species catalogue. Vol. 7. Clupeoidfishes of the world (suborder Clupeioidei). An annotated andillustrated catalogue of the herrings, sardines, pilchards,sprats, shads, anchovies and wolf-herrings.. Part 1—Chirocentridae, Clupeidae and Pristigasteridae. FAO Fish.Synop. 125(7/1):1–303
Wilson SG, Newbound DR (2001) Two whale shark faecal.samples from Ningaloo Reef, Western Australia. Bull MarSci 68:361–362
Wilson SG, Taylor JG, Pearce AF (2001) The seasonalaggregation of whale sharks at Ningaloo Reef, WesternAustralia: currents, migrations and the El Niño/SouthernOscillation. Environ Biol Fish 61:1–11
Wilson SG, Polovina JJ, Stewart BS, Meekan MG (2006)Movements of Whale Sharks (Rhincodon typus) tagged atNingaloo Reef, Western Australia. Mar Biol 148:1157–1166
Wintner SP (2000) Preliminary study of vertebral growth ringsin the whale shark, Rhincodon typus, from the east coast ofSouth Africa. Environ Biol Fish 59:441–451
Wolfson FH (1983) Records of seven juveniles of the whaleshark Rhiniodon typus. J Fish Biol 22:647–655
Zingde MD (2005) Inputs into the oceans from land/rivers andpollution. In: Gupta HK (ed) Oceanology. UniversitiesPress, India, pp 92–117
Environ Biol Fish (2011) 92:559–567 567