space partitioning and interactions in an intertidal sand-burrowing amphipod guild
TRANSCRIPT

Marine Biology 61, 79-88 (1980) MARINE BIOLOGY �9 by Springer-Verlag 1980
Space Partitioning and Interactions in an Intertidal Sand-Burrowing Amphipod Guild
R. A. Croker and E. B. Hatfield
Department of Zoology and Jackson Estuarine Laboratory, University of New Hampshire; Durham, NH 03824, USA
Abstract Introduction
A nine-year study at a moderately exposed, fine sand intertidal beach at Long Sands, Maine, USA identified marked seasonal changes in the foci of sand-burrowing amphipod activities. These included seasonal population movements, degree of interspecific horizontal overlap, life stage segregation within and between species, and sand depth stratification. Behavioral interactions related to competition for available space are considered impor- tant in affecting the distribution and abundance of amphipod species, and community structure. Significant negative correlations were shown between the abundance and upper distributional limits of the smaller, lower shore Acanthohaustorius millsi and the abundance of the larger, higher shore species Haustorius canadensis. Tolerance limits to a variety of prevailing physical fac- tors alone, were not considered a serious obstacle to occupation of higher intertidal level sands by A. millsi. Summer brooding females of both species occupied similar 5 to 10 cm sand depths, but were strongly segregated horizontally. Laboratory coexistence ex- periments during the reproductive period showed in- creased mortality of adults and negligible reproductive output of A. millsi when in combination with H. cana- densis compared to controls where the former species was alone. A third species, Amphiporeia virginiana, oc- cupied primarily very shallow sand (0 to 2.5 cm), and performed seasonal movements opposite in direction to A. millsi and H. canadensis. The presence, amount, and refinement of biological interactions across the wave exposure gradient requires further study. Until then, we consider as inappropriate the blanket designation of all types of intertidal sand communities as "physically controlled".
A wealth of information has appeared concerning the ecology of amphipod-dominated intertidal sand dom- munities in the USA (Croker, 1967; Dexter, 1967, 1969; Sameoto, 1969a; Croker et al., 1975; Holland and Polgar, 1976; Holland and Dean, 1977; Bosworth, 1977), in the British Isles (Colman and Segrove, 1955; Mclntyre, 1970; Fincham, 1971), and Europe (DAN, 1952; Salvat, 1967). These studies have yielded tantaliz- ing hints of interspecific competition and ecological release, mention of seasonal population movements, and the suggested masking of biological interactions by physical factors in what is considered a relatively harsh envrionment. We believe that long-term field studies at well-characterized habitats are an important ingredient for thoroughly exploring these aspects of the species network (sensu Elton and Miller, 1954). Such studies have been lacking so far. In addition, with the exception of some preliminary information (Watkin, 1942; Croker, 1967; Nicolaisen and Kanneworff, 1969; Sameoto, 1969a), the partitioning of vertical space (sand depth) has not been adequately examined.
Long-term ecological field studies began during 1971 at Long Sands, York, Maine, USA, a moderately exposed sandy shore where sedimentary parameters and beach morphology are substantially less variable than at very exposed locations (Hager and Croker, 1979). We doc. umented relatively modest fluctuations in amphi- pod abundance and biomass at Long Sands during early years (Croker et al., 1975; Croker, 1977) suggesting that amphipod species populations were at, or near carrying capacities. We also obtained some preliminary evidence for species interactions, and subsequently hypothesized that behavioral interactions related to available space (downshore, and depth in sand) were important in affecting the distribution and abundance of amphipod species. Community structure would presumably be affected in turn, since the 3 dominant amphipod species, Acanthohaustorius millsi, Amphiporeia virginiana, and
0025-3162/80/0061/00791502.00

80 R. A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild
Haustorius canadensis make up more than 90% of both abundance and biomass in a community generally total- ling 8 to 10 species (Croker et al., 1975; Croker, 1977).
We report here on evidence from field studies during 1971-1979, and laboratory experiments during 1978 related to the patterns of space partitioning and inter- actions among amphipod species. We conclude by discussing the implications of our findings for views on intertidal sand community structure.
Materials and Methods
Quantitative sampling to determine horizontal distribu- tion and abundance was accomplished by taking triplicate 0.04 m 2 samples of sand 12.5 cm deep. Samples were taken at 5 or 6 levels on a transect form the highest level of occurrence of amphipods to approximately Mean Low Water (MLW) level at Long Sands, Maine during March and July from 1971 through 1979. Individ- uals were separated from sand by washing through a 0.5 mm sieve. Coefficients of variation of species popula- tion estimates averaged about 12%.
Cores to determine vertical distribution and abundance were taken with a split plastic corer (83 cm 2 area) secured with an alligator clamp. The corer was forced 20 cm into the sand during low tide. Sand cores were cut into 2.5 cm sections to a depth of 10 cm, with an additional core section to 12 or 15 cm depth. Six to eight replicate cores were taken at each of 3 to 5 beach levels seasonally from June 1977 to August 1978, for a total of 150 cores. This sampling design provided a 95% probability of estimating amphipod numbers in cores within 10% of their true value.
Experiments were performed to simulate the co- existence of Acanthohuastorius millsi and Haustorius canadensis in the lower intertidal zone, or in higher beach pools and runoffs. In addition to being abundant between the tides, H. canadensis is also known from sandy marsh creeks, and shallow intertidal sand bars inundated by sea water on all but the lowest of low tides (Sameoto, 1969a; Bousfield, 1973). A. millsi occurs subtidally up to 50 m (Bousfield, 1973). Survival and reproductive output of these species alone and together were compared using freshly-collected amphipods (with- in 24 h). Individuals were sexed live (by the presence of reproductive structures visible through the dorsal exoskeleton), and placed in presieved sand (8 cm depth) held in 8.5 x 8.5 x 11.5 cm plastic containers. Numbers of amphipods were similar to field abundances and bio- mass. The ranges and approximate average lengths of A. millsi and/-/, canadensis were 2.8 to 5.3 mm (4.0 mm), and 5.1 to 9.0 mm (6.6 ram), respectively. Fine nylon mesh windows in the containers allowed passage of flowing water through the sand while the containers were held on a sea water table. Experiments were run for 3 wk during July 11 to August 1, 1978, with 3 replicates for each single species control, and for the species together. Temperature and salinity during
experiments averaged 20.7 ~ -+ 1.2 C ~ and 29.1 + 1.8 ~ S, respectively.
Water content of sand at 5 cm depths was measured by inserting pre-weighed, 20 ml glass vials horizontally into a quickly-opened sand trench at half-hour intervals during a half tidal cycle. Vials were then capped, wrapped with electrical tape, and brought to the laboratory. The vials were wiped free of excess moisture, uncapped, and immediately reweighed. Vials were then dried for 48 h at 70 ~ and reweighed again. Water content as percent weight of sand was obtained by difference. The depth of the water table in the beach was measured during ebbing, low, and flooding tides (6 h) with a meter stick placed in dug holes.
Temperature measurements of sand were made at half-hour intervals over a similar (above) 6 h tidal period with a Yellow Springs Telethermometer and the appropriate thermistors. Thermistors were calibrated against a laboratory mercury thermometer, while both were placed in melting ice. Salinity of surf and inter- stitial water was measured with a refractometer. Inter- stitial water was collected with a microsyringe. Oxygen content of interstitial water was measured with a Hach Chemical Kit (Ames, Iowa, USA), after carefully filling a small vial with seawater slowly draining into dug holes. Relative humidity of sand during low tide emersion was measured with paper humidity indicators (Andrews Tech. Co., Evanston, Ill., USA).
Results
Physical Environment
The study habitat is a gently sloping (1:48 to 1:68), sandy shore with well-sorted (o I = 0.37 to 0.43), fine sand with median diameters 2.07 to 2A7 6, and with generally < 1% of coarse sand present. Mean tide range is about 2.6 m, and monthly sea water temperatures average 0 ~ to 17 ~ during the year. Salinity of inter- stitial water in beach sands generally averaged within - 3 ~ S of surf water salinity (annual average range 28.5 to 33.0 %o S) with the exception of lower salinities during spring thaw (as low as 22.5 %o S), and lower salinities deeper than 10 cm in beach sands between 1.7 to 2.0 m levels apparently influenced by ground water (as low as 22 ~ S). The habitat exhibited a relatively stable beach profile, with the exception of moderate to severe erosion of the upper beach (> 2.0 m) during periodic winter storms. Additional habitat characteristics were presented in Croker et aL (1975).
The temperatures reached by intertidal sands during low tide emersion on a very warm summer day, with a 0.0 low tide at 1226 hrs, are given in Table 1. As ex- pected, wider temperature ranges occurred at the higher intertidal level and at shallower sand depths, while temperatures were lower both deeper in the sand and at the lower intertidal level (0.8 m). Similar temperature profiles were common for warm summer days from late June through early August. During one of the hottest
R. A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild
Table 1. Temperature of intertidal sands during low tide emersion, Long Sands, Maine, USA. June 28, 1978 a
81
Level(m) Emersion(h) Sanddepth(cm) 3.8 6.3 8.8
1.4 6.2 25.2• b 22.8~4.5 21.0• 0.8 4.3 24.3• 21.7• 19.2•
11.0
19.5• 17.9•
aAir temperature 0.5 m over beach 29.5 ~ • 1.0 C ~ ; surf temperature at time of low tide 13.3 ~ bMean • I Standard deviation (DS)
Table 2. Some environmental factors related to low tide emersion, Long Sands, Maine, USA. June 28 and 30, 1978
% Water content at 5 cm depth
Just after % Water Low tide Nos./0.04 m 2c Level (m) Emersion (h) emersion Low tide loss water table
(cm) A. millsi H. canadensis
1.7 7.6 20.1• b 10.7• 46.8 >35 2.5• 6.9 1.4 6.2 21.2• 9.4• 55.7 >35 44.7• 0.8 4.3 21.7• 11.3• 47.9 >35 26.3•
42.3 -+ 22.7
0.6
aJune 30, low tide 1424 hrs (0.6) bMean • [ SD, based on 3 to 5 samples; water content > 21% weight of sand - oversaturation CMean -+ I SD for 8 core samples to 10 cm per beach level
�9 EHWS
3 _MHWS
MHW
~IHWN .SHWN
MTL
ELWN MLWN
.MLW
~LWS
~'ELWS
(218)
koe 2524 Jut},
A. ~giniaua
(~5) (27)
Mac duty
H. canadensis
(51}
Mac duty A. mittsi
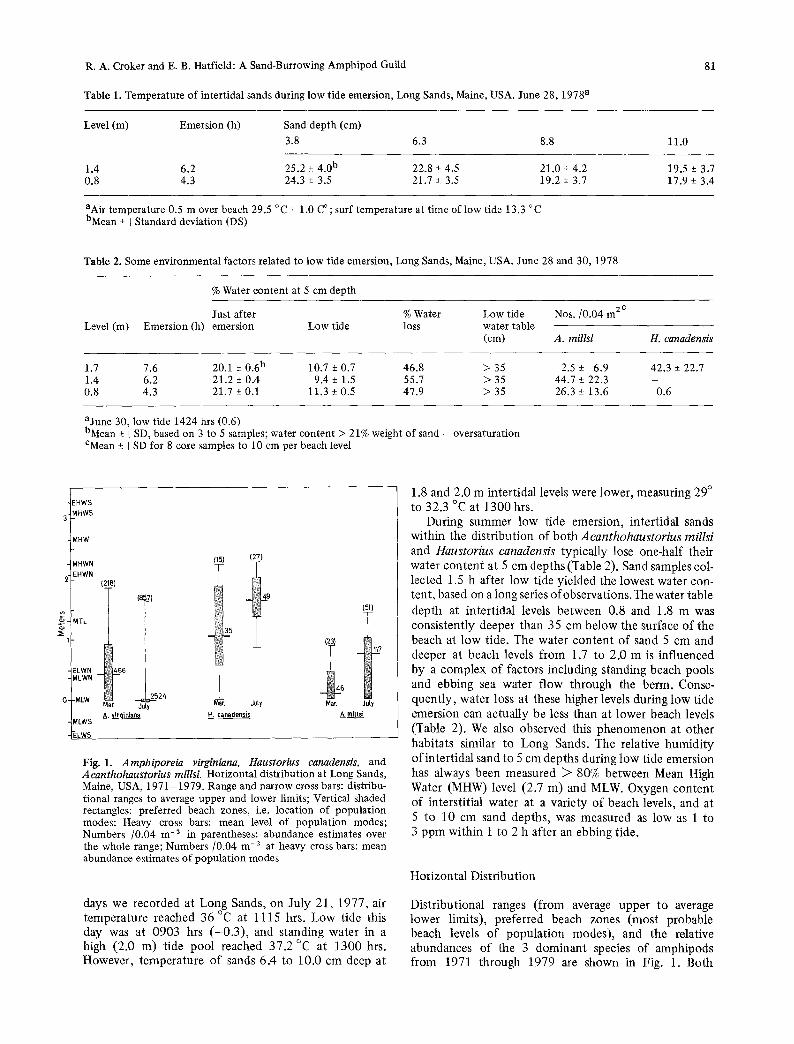
Fig. 1. Amphiporeia virginiana, Haustorius canadensis, and Acanthohaustorius millsi. Horizontal distribution at Long Sands, Maine, USA, 1971 1979. Range and narrow cross bars: distribu- tional ranges to average upper and lower limits; Vertical shaded rectangles: preferred beach zones, i.e. location of population modes; Heavy cross bars: mean level of population modes; Numbers /0.04 m -~ in parentheses: abundance estimates over the whole range; Numbers/0.04 m -2 at heavy cross bars: mean abundance estimates of population modes
days we recorded at Long Sands, on July 21, 1977, air temperature reached 36 ~ at 1115 hrs. Low tide this day was at 0903 hrs ( -0 .3 ) , and standing water in a high (2.0 m) tide pool reached 37.2 ~ at 1300 hrs. However, temperature of sands 6.4 to 10.0 cm deep at
1.8 and 2.0 m intertidal levels were lower, measuring 29 ~ to 32.3 ~ at 1300 hrs.
During summer low tide emersion, intertidal sands within the distribution of bothAcanthohaustorius rnillsi and Haustorius canadensis typically lose one-half their water content at 5 cm depths (Table 2), Sand samples col- lected 1.5 h after low tide yielded the lowest water con- tent, based on a long series of observations. The water table depth at intertidal levels between 0.8 and 1.8 m was consistently deeper than 35 cm below the surface of the beach at low tide. The water content of sand 5 cm and deeper at beach levels from 1.7 to 2.0 m is influenced by a complex of factors including standing beach pools and ebbing sea water flow through the berm. Conse- quently, water loss at these higher levels during low tide emersion can actually be less than at lower beach levels (Table 2). We also observed this phenomenon at other habitats similar to Long Sands. The relative humidity of intertidal sand to 5 cm depths during low tide emersion has always been measured > 80% between Mean High Water (MHW) level (2.7 m) and MLW. Oxygen content of interstitial water at a variety of beach levels, and at 5 to 10 cm sand depths, was measured as low as 1 to 3 ppm within 1 to 2 h after an ebbing tide.
Horizontal Distribution
Distributional ranges (from average upper to average lower limits), preferred beach zones (most probable beach levels of population modes), and the relative abundances of the 3 dominant species of amphipods from 1971 through 1979 are shown in Fig. 1. Both
82 R.A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild
10C
6C q 0
0 March
�9 Jury
I " 0 y = 77.2- 20.07 tn x r 077 p<O001
0 0 0 �9
0 �9 0
' 2 '0 ' 20 ' H= canadensis abundance/O.04 m 2
6b
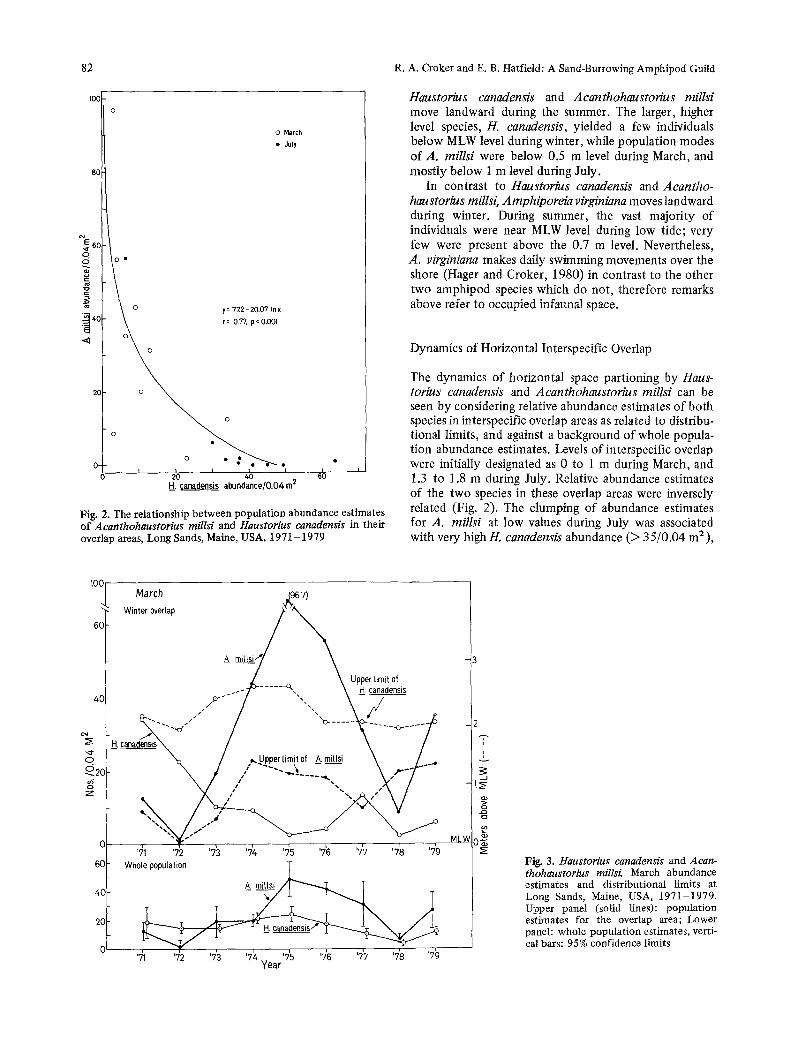
Fig. 2. The relationship between population abundance estimates of Acanthohaustorius millsi and Haustorius canadensis in their overlap areas, Long Sands, Maine, USA, 1971-1979
Haustorius canadensis and Acanthohaustorius millsi move landward during the summer. The larger, higher level species, 11. canadensis, yielded a few individuals below MLW level during winter, while population modes of A. rnillsi were below 0.5 m level during March, and mostly below 1 m level during July.
In contrast to Hctustorius canadensis and Acantho- haustorius millsi, Amphiporeia virginiana moves landward during winter. During summer, the vast majority of individuals were near MLW level during low tide; very few were present above the 0.7 m level. Nevertheless, A. virginiana makes daily swimming movements over the shore (Hager and Croker, 1980) in contrast to the other two amphipod species which do not, therefore remarks above refer to occupied infaunal space.
Dynamics of Horizontal Interspecific Overlap
The dynamics of horizontal space partioning by Haus- torius canadensis and Acanthohaustorius millsi can be seen by considering relative abundance estimates of both species in interspecific overlap areas as related to distribu- tional limits, and against a background of whole popula- tion abundance estimates. Levels of interspecific overlap were initially designated as 0 to 1 m during March, and 1.3 to 1.8 m during July. Relative abundance estimates of the two species in these overlap areas were inversely related (Fig. 2). The clumping of abundance estimates for A. millsi at low values during July was associated with very high//, canadensis abundance (2> 35/0.04 m2),
I00
60
40
-4" o. ~ g Z
0
60
4C
20
March ,t,~.7) Winter overlap / ' % ~
J ~ Upper [irnit of . . . . ~ - . . . . -o,\ \ H canadensis
" % / ,. Upper limit of A. millsi \ /
i i I "=~%%% i I
'71 72 73 '74 '75 76 '77 '78 79
Whole population L
, T , , ~ , , ., , -; /rs '71 '72 '73 '74 '75 '76 '77 '78 '79
Year
ML~
2
T i
o
O~
Fig. 3. Haustorius canadensis and Acan- thohaustorius millsL March abundance estimates and distributional limits at Long Sands, Maine, USA, 1971-1979. Upper panel (solid lines): population estimates for the overlap area; Lower panel: whole population estimates, verti- cal bars: 95% confidence limits
R. A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild 83
3
JuLy * March o . . . . .
i y= -O.02x + 2.17
"~0,~ �9 . ~ r=-O.67, p<O.05
" �9 y:-O.03x 124 �9 - . r= -06g, p<O05
r I 2 ~ U o I "1~ I I 40 6tO 810 H. canadensis abundance/0.04 m 2
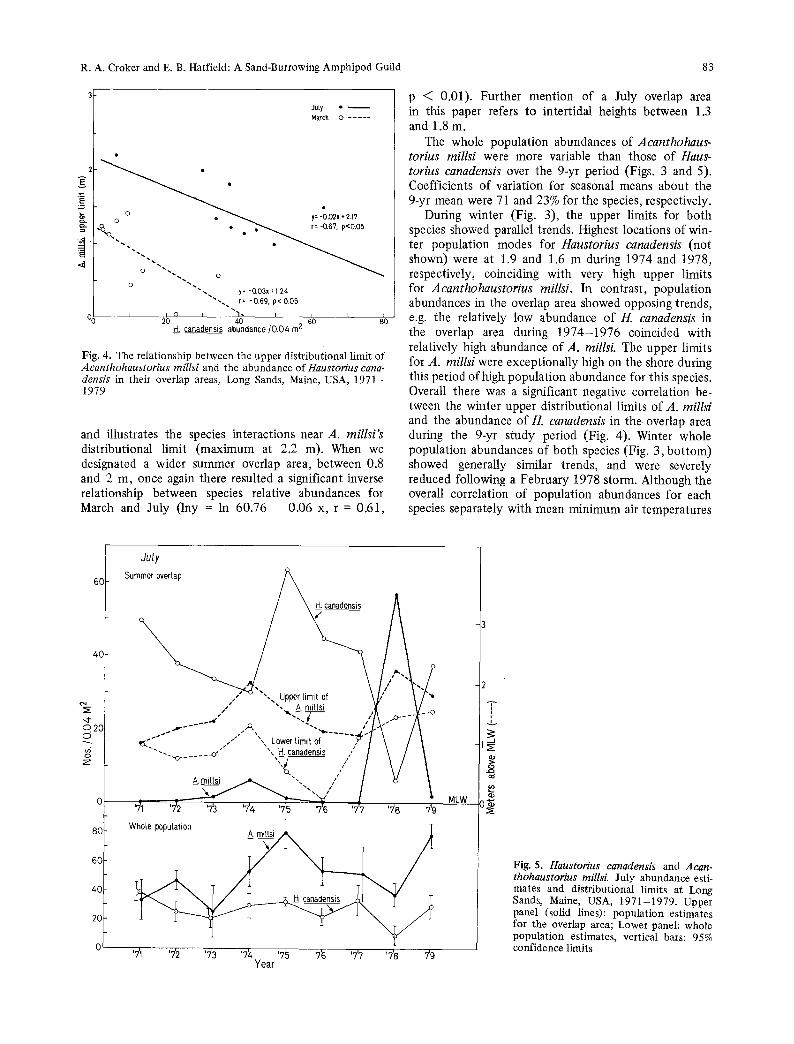
Fig. 4. The relationship between the upper distributional limit of Aeanthohaustorius millsi and the abundance of Haustorius cana- densis in their overlap areas, Long Sands, Maine, USA, 1971- 1979
and illustrates the species interactions near A. millsi's distributional limit (maximum at 2.2 m). When we designated a wider summer overlap area, between 0.8 and 2 m, once again there resulted a significant inverse relationship between species relative abundances for March and July (lny = in 60.76 - 0.06 x, r = 0.61,
p < 0.01). Further mention of a July overlap area in this paper refers to intertidal heights between 1.3 and 1.8 m.
The whole population abundances of Acanthohaus- torius millsi were more variable than those of Haus- torius canadensis over the 9-yr period (Figs. 3 and 5). Coefficients of variation for seasonal means about the 9-yr mean were 71 and 23% for the species, respectively.
During winter (Fig. 3), the upper limits for both species showed parallel trends. Highest locations of win- ter population modes for Haustorius canadensis (not shown) were at 1.9 and 1.6 m during 1974 and 1978, respectively, coinciding with very high upper limits for Aeanthohaustorius millsi. In contrast, population abundances in the overlap area showed opposing trends, e.g. the relatively low abundance of H. canadensis in the overlap area during 1974-1976 coincided with relatively high abundance of A. millsi. The upper limits for A. millsi were exceptionally high on the shore during this period of high population abundance for this species. Overall there was a significant negative correlation be- tween the winter upper distributional limits of A. millsi and the abundance of H. canadensis in the. overlap area during the 9-yr study period (Fig. 4). Winter whole population abundances of both species (Fig. 3, bo t tom) showed generally similar trends, and were severely reduced following a February 1978 storm. Although the overall correlation of population abundances for each species separately with mean minimum air temperatures
60
40
% -..1" Q 2C o Z
0 80 60 40 20
July Summer overlap ~ k
X / \ H canadensis ] ~
s SS �9 �9 7
,,'/L.Y:g
." / \ / / -.o. . . . . . ,.< / / V /
, . 1 '7'1 '?'2 ?~ '7'4 '7~ '7'6 '7T7 '7'8 7'9
WhoLe population A mittsi 2
'7'1 '7'2 '7'3 '72+ '7~ '7'6 '7~7 '7'8 7'9 Year
2
T 14
MLW 0
Fig. 5. Haustorius canadensis and Acan- thohaustorius millsi. July abundance esti- mates and distributional limits at Long Sands, Maine, USA, 1971-1979. Upper panel (solid lines): population estimates for the overlap area; Lower panel: whole population estimates, vertical bars: 95% confidence limits
84 R.A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild
5.0
75
10.0
> 1 0
0
2,5
Preferred beach zones (Summer) H~ canadensir~ P A. millsi
Jan.-early May
z:v
~E!;!
z; ..........
llsi \: :;':!~ es)
H. canadensis (??cores)
' ~ k ,!o o'.s ' MLWN 1.5 Beach level (N)
2.5
f ol
2~
MLW
Preferred beach zones A.v., H, canadensis ~ A~miUsi ~
T
Adults Summer
A.mitlsi (77 cores)
I
?.5 .k l.'O o'.5'.Lw. ~w Beach level (M)
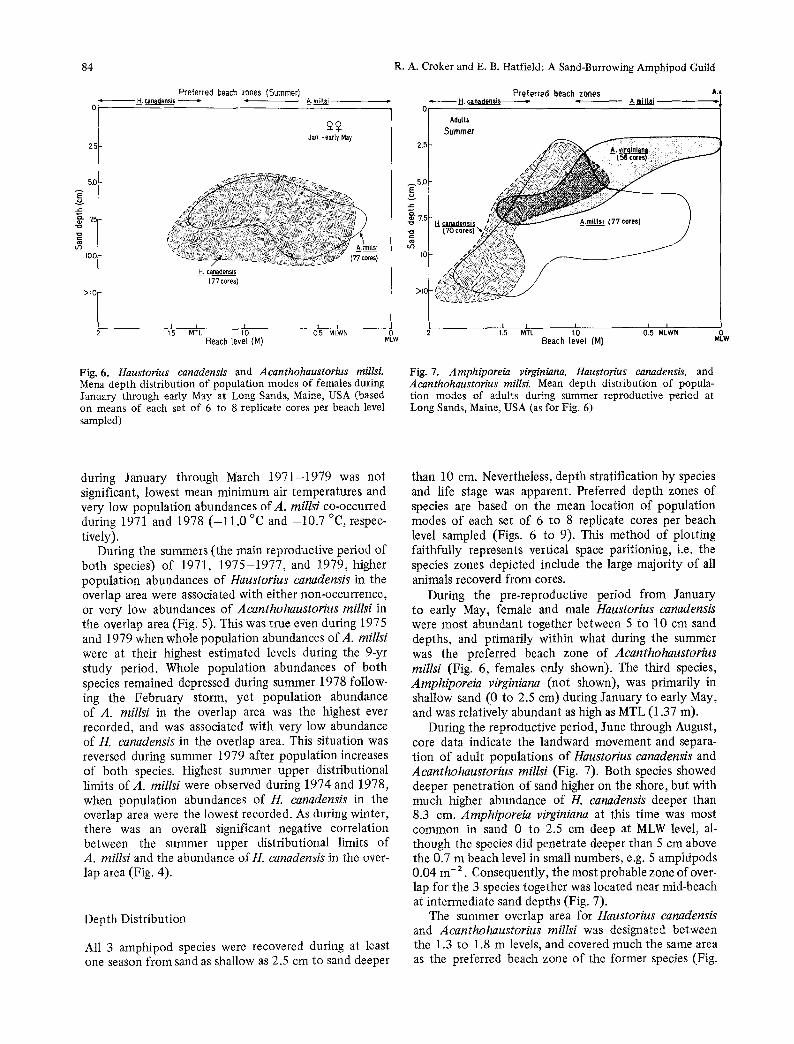
Fig. 6. Haustorius canadensis and Acanthohaustorius millsL Mena depth distribution of population modes of females during January through early May at Long Sands, Maine, USA (based on means of each set of 6 to 8 replicate cores per beach level sampled)
Fig. 7. Amphiporeia virginiana, Haustorius canadensis, and Acanthohaustorius millsi. Mean depth distribution of popula- tion modes of adults during summer reproductive period at Long Sands, Maine, USA (as for Fig. 6)
during January through March 1971-1979 was not significant, lowest mean minimum air temperatures and very low population abundances ofA. millsi co-occurred during 1971 and 1978 ( -11.0 ~ and -10.7 ~ respec- tively).
During the summers (the main reproductive period of both species) of 1971, 1975-1977, and 1979, higher population abundances of Haustorius canadensis in the overlap area were associated with either non-occurrence, or very low abundances of Acanthohaustorius millsi in the overlap area (Fig. 5). This was true even during 1975 and 1979 when whole population abundances ofA. millsi were at their highest estimated levels during the 9-yr study period. Whole population abundances of both species remained depressed during summer 1978 follow- ing the February storm, yet population abundance of A. milIsi in the overlap area was the highest ever recorded, and was associated with very low abundance of H. canadensis in the overlap area. This situation was reversed during summer 1979 after population increases of both species. Highest summer upper distributional limits of A. millsi were observed during 1974 and 1978, when population abundances of H. canadensis in the overlap area were the lowest recorded. As during winter, there was an overall significant negative correlation between the summer upper distributional limits of A. millsi and the abundance ofH. canadensis in the over- lap area (Fig. 4).
Depth Distribution
All 3 amphipod species were recovered during at least one season from sand as shallow as 2.5 cm to sand deeper
than 10 cm. Nevertheless, depth stratification by species and life stage was apparent. Preferred depth zones of species are based on the mean location of population modes of each set of 6 to 8 replicate cores per beach level sampled (Figs. 6 to 9). This method of plotting faithfully represents vertical space paritioning, i.e. the species zones depicted include the large majority of all animals recoverd from cores.
During the pre-reproductive period from January to early May, female and male Haustorius canaclensis were most abundant together between 5 to 10 cm sand depths, and primarily within what during the summer was the preferred beach zone of Acanthohaustorius millsi (Fig. 6, females only shown). The third species, Amphiporeia virginiana (not shown), was primarily in shallow sand (0 to 2.5 cm) during January to early May, and was relatively abundant as high as MTL (1.37 m).
During the reproductive period, June through August, core data indicate the landward movement and separa- tion of adult populations of Haustorius canadensis and Acanthohaustorius millsi (Fig. 7). Both species showed deeper penetration of sand higher on the shore, but with much higher abundance of H. canadensis deeper than 8.3 cm. Amphiporeia virginiana at this time was most common in sand 0 to 2.5 cm deep at MLW level, al- though the species did penetrate deeper than 5 cm above the 0.7 m beach level in small numbers, e.g. 5 amphipods 0.04 m- 2. Consequently, the most probable zone of over- lap for the 3 species together was located near mid-beach at intermediate sand depths (Fig. 7).
The summer overlap area for Haustorius canadensis and Acanthohaustorius millsi was designated between the 1.3 to 1.8 m levels, and covered much the same area as the preferred beach zone of the former species (Fig.
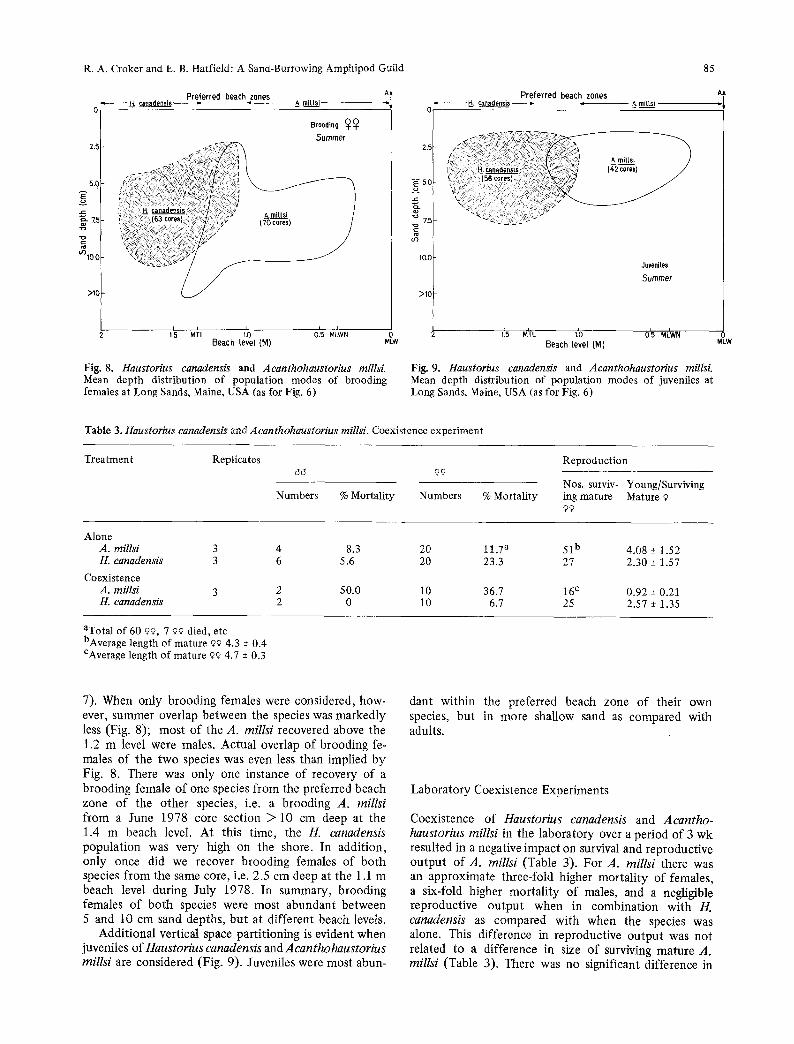
R. A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild 85
2.5
5.(
~10.(
>1(
A.v. Preferred beach zones ! H~ r �9 ~ A. mittsi .~
Brooding ~
Summer
;,'~ Mk ,'.o o; ' MLWN Beach level (M) MLW
Fig. 8. Haustorius canadensis and Acanthohaustorius millsL Mean depth distribution of population modes of brooding females at Long Sands, Maine, USA (as for Fig. 6)
2.5
5.0
7.5
0.0
>10
Preferred beach zones 147 %anadens s - - A. mi|tsi
Juveniles
Summer
2 l!s M~L ;I0 015 ' MLWN Beach [ever (M)
Fig. 9. Haustorius canadensis and Acanthohaustorius millsi. Mean depth distribution of population modes of juveniles at Long Sands, Maine, USA (as for Fig. 6)
A~
-1
MLW
Table 3. Haustorius canadensis and Acanthohaustorius millsi. Coexistence experiment
Treatment Replicates d d 9 9
Numbers % Mortality Numbers % Mortality
Reproduction
Nos. surviv- Young/Surviving ing mature Mature 9 97
Alone A. millsi 3 4 8.3 1t. canadensis 3 6 5.6
Coexistence A. millsi 3 2 50.0 H. canadensis 2 0
20 11.7 a 51 b 4.08 _+ 1.52 20 23.3 27 2.30 _+ 1.57
10 36.7 16 c 0.92 +_ 0.21 10 6.7 25 2.57 -+ 1.35
aTotal of 60 99, 7 99 died, etc bAverage length of mature 99 4.3 -+ 0.4 eAverage length of mature 99 4.7 -+ 0.3
7). When only brooding females were considered, how- ever, summer overlap between the species was markedly less (Fig. 8); most of the A. millsi recovered above the 1.2 m level were males. Actual overlap of brooding fe- males of the two species was even less than implied by Fig. 8. There was only one instance of recovery of a brooding female of one species from the preferred beach zone of the other species, i.e. a brooding A. millsi from a June 1978 core section > 10 cm deep at the 1.4 m beach level. At this time, the H. canadensis populat ion was very high on the shore. In addition, only once did we recover brooding females of both species from the same core, i.e. 2.5 cm deep at the 1.1 m beach level during July 1978. In summary, brooding females of both species were most abundant between 5 and 10 cm sand depths, but at different beach levels.
Addit ional vertical space parti t ioning is evident when juveniles of Haustorius canadensis and Acanthohaustorius millsi are considered (Fig. 9). Juveniles were most abun-
dant within the preferred beach zone of their own species, but in more shallow sand as compared with adults.
Laboratory Coexistence Experiments
Coexistence of Haustorius canadensis and Aeantho- haustorius millsi in the laboratory over a period of 3 wk resulted in a negative impact on survival and reproductive output of A. millsi (Table 3). For A. millsi there was an approximate three-fold higher mortal i ty of females, a six-fold higher mortal i ty of males, and a negligible reproductive output when in combination with H. canadensis as compared with when the species was alone. This difference in reproductive output was not related to a difference in size of surviving mature A. millsi (Table 3). There was no significant difference in

86 R. A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild
reproductive output of experimental and control H. canadensis.
Mortality ofHaustorius canadensis females was higher when the species was alone. This may have been due to a density effect in the single species control. A similar effect was not seen for Acanthohaustorius millsi; adults of this species are on the average only 20% as heavy as adult H. canadensis.
Discussion and Conclusions
Long-term studies have shown marked seasonal changes in the loci of amphipod activities within a moderately exposed, fine sand habitat. These include seasonal population movements, degree of interspecific horizon- tal overlap, increased segregation of a species pair during reproductive period, and a dynamic pattern of changing distributional limits and population abundances of that species pair. Combined with details of life stage segrega- tion and vertical stratification, these population charac- teristics affect community structure in intertidal sand beach space.
Acanthohaustorius millsi has been characterized as a common, essentially subtidal species from Maine to central Florida (Bousfield, 1973). It is also a prominent member of US Atlantic intertidal sand communities (Dexter, 1969; Croker et al., 1975; Croker, 1977; Holland and Polgar, 1976). More variable seasonal abundances characterize this species compared with co-occurring closely-related species, both in northern New England, and in southeastern US and habitats (Holland and Polgar, 1976). We view the intertidal distribution and abundance of this species in northern New England as associated partly with temperature, e.g. very low abundance correlated with extremely low mean minimum winter air temperatures, but also with the distribution and abundance of the confamilial Haustorius canadensis. By migrating landward in warmer months, A. millsi has liberal access to food and space at inter- mediate sand depths below 1 m beach level, when the larger H. canadensis moves landward as well. Additional species occurring here during summer include Amphi- poreia virginiana, Micrura sp., Nephtys bucera, Scole- lepis squamata, and Paraonis fulgens. These species all exhibited abundances < 3 individuals (and mostly < 1) 0.04 m -2 at intermediate sand depths; none are consid- ered as a serious competitor with A. millsi.
The horizontal overlap ofAcanthohaustorius millsi and Haustorius canadensis was maximal during winter and early spring, and minimal for brooding females and juveniles of both species during the summer reproductive period. On the other hand, with the exception of in- creased numbers of H. canadensis (males and maturing females) deeper in the sand at high intertidal levels during warmer months, the overall occupation of vertical space was similar for these two species. Other within- species differences were noted, however, e.g. deeper penetration of higher intertidal sands by male A. millsi
during summer, and the more shallow occurrence of juveniles of both species.
There were strong negative correlations for the upper distributional limits and population abundance of Acan- thohaustorius millsi with the population abundance of Haustorius canadensis, implying competition for habitat space. The evidence shown for ecological release of A. millsi could be based on a volume relationship as Fig. 2 suggests. It appears that H. canadensis abundances < about 20 individuals 0.04 m -2 are correlated with a display of higher abundances ofA. millsi. The results of laboratory coexistence experiments when space was obviously limited further indicate the negative impact ofH. canadensis with abundances ~20 individuals 0.04 m -2 on survival and reproductive success of the smaller species, and underlines the advantage of the segregation of ovigerous A. millsi to the lower shore and away from areas of high density H. canadensis.
The consistently higher penetration of the intertidal zone by Acanthohaustorius millsi as Haustorius canadensis abundance decreased was observed by us early on, and not only at this habitat. This represents a departure from the more common situation where the upper limits of a lower shore species are set primarily by tolereances to physical factors (Connell, 1961). An alternate ex- planation is that both species are responding to similar (or different) physical factors which we have not yet identified (see below).
In summary, vertical distribution and reproductive periods are similar for Acanthohaustorius millsi and Haustorius canadensis while the amount of horizontal overlap changes with season. Despite differences in mouthpart morphology of these filter feeders (Bous- field, 1970), there have been no comparative studies on food utilization. We view the dynamic relationship between the species in fine sand intertidal habitats as an example of the ability of haustoriid (sensu Bousfield, 1978) species to inhabit successively more landward environments (Bousfield, 1970). Accordingly, H. cana- densis plays a prominent role in affecting the realized niche of A. millsi. No, or very little overlap in the horizontal distribution of these species is apparent at habitats where lower intertidal sediments are coarse, and where A. millsi is found primarily in subtidal sands (Sameoto, 1969a).
Despite the somewhat higher overall tolerance of Haustorius canadensis to higher temperature, desicca- tion, and low salinities as compared to Acanthohaus- torius millsi (Sameoto, 1969b), North American Atlantic coast haustoriid species are generally tolerant to more extreme environmental conditions than they experience in nature (Croker, 1967; Sameoto, 1969b). Furthermore, significant correlations between population abundances of most New England sand-burrowing amphipod species and such physical factors as organic carbon, nitrogela, and water content of sand, sand grain size, sand tem- perature, and oxygen content of pore water have not appreared (Sameoto, 1969a; Croker et al., 1975).
We consider the suite of physical variables as back- ground conditioning factors for amphipod spatial and

R. A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild 87
temporal dynamics in fine sand beaches. Variables such as sediment temperature, water content, salinity, and sand grain size are similar enough within-season over the intertidal zone of fine sand habitats, so that except for occasional random, large stresses, e.g. very hot days coinciding with spring tides, or major storm erosion, we do not consider them as primarily controlling the within-habitat distribution and abundance of most sand-burrowing amphipod species. On the other hand, seasonal temperature variation probably modulates population movements of amphipod species (Fincham, 1970, 1971), and significant correlations between the population abundance of Amphiporeia virginiana and sedimentary cycles on very exposed shores were reported (Hager and Croker, 1979).
Concerning the role of predation on these amphipods, excepting for the obvious predation we have seen on swimming and shallow-burrowing Amphiporeia vir- giniana by migrating shorebirds and fish, we are not aware of any predation pressure that could potentially regulate populations of Haustorius canadensis and Acanthohaustorius millsi in intertidal habitats. Neither species appears in the surf plankton (R. A. Croker, unpublished data), nor do they swim in beach runoff as A. virginiana does. The occupation of deeper sand by these species may serve as a refuge (sensu Woodin, 1978) from potential predators. From what is known of fish predation on sandy shores when covered by the tides, the majority of predators are Young flatfish that feed primarily by sight, and mostly during the day. Frequent prey include polychaetes (whole and palps), bivalves (young and siphons), and tidally active peracarid crustaceans (i.e. like A. virginiana), e.g. species of Bathyporeia and Eurydice (Edwards and Steele, 1968; Gibson, 1969, 1973; Tyler, t971; Thijssen etal., 1974). The impact of predation by migrating shore birds on burrowing crustaceans would depend on the length of the bird's bill, and efficiency of capture of various life stages of prey (Puttick, 1977, 1978). The whole subject of predation requires study in North American Atlantic sandy habitats.
We do not agree that all sandy intertidal communities should be designated per se as "physically controlled" (Holland and Polgar, 1976). Although the number and kinds of species are probably largely determined by the suite of physical factors, we agree that the designation "physically controlled" is proper only if in addition, distribution and abundance patterns are directly and primarily caused by environmental perturbations (Menge and Sutherland, 1976)
For the different types of intertidal sand habitats, e.g. protected, moderately exposed, and exposed, we envision a change in the relative degree of control of abundance and distribution of constituent species by physical factors and biological interactions across the gradient of physiological stress (Sanders, 1968, 1969). Simply put, we suggest a spread in the location of inter- tidal sand communities across the gradient. Two tasks at hand are to determine how refined biological organiza- tion is across the wave exposure gradient, and to what
degree environmental variability modulates potential competition for space.
Acknowledgements. We are indebted to many people for assistance in the laboratory and in the field, particu- larly T. Donn, P. Groden, C. McBane, C. Scarano, and K. Stapelfeldt. Discussions with W. Bosworth and J. Levinton were helpful. This work was supported by the Hubbard Fund of the University of New Hampshire, and partially by NSF (Oceanography Section) Grant GA 33743 to R. A. Croker. It is Contribution No. 99 of the Jackson Estuarine Laboratory, University of New Hampshire.
Literature Cited
Bosworth, W. S., Jr.: Biology of the genus Eohaustorius (Amphi- poda: Haustoriidae) on the Oregon coast. 194 pp. Ph.D. Thesis, Oregon St. Univ., Corvallis, 1977
Bousfield, E. L.: Adaptive radiation in sand-burrowing amphi- pod crustaceans. Chesapeake Sci. 11,143-154 (1970)
Bousfield, E. L.: Shallow-water gammaridean Amphipoda of New England. 312 pp. Cornell Univ. Press., Ithaca, N. Y. 1973
Bousfield, E. L.: A revised classification and phylogeny of amphipod crustaceans. Trans. R. Soc. Can. 16, 343-390 (1978)
Colman, J. S. and F. Segrove: The fauna living in Stoupe Beck Sands, Robin Hood's Bay (Yorkshire, North Riding). J. Anita. Ecol. 24, 426-444 (1955)
Connell, J. H.: The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalus stellatus. Ecology 42, 710-723 (1961)
Croker, R. A.: Niche diversity in five species of intertidal amphi- pods (Crustacea: Haustoriidae). Ecol. Monogr. 37, 173-200 (1967)
Croker, R. A.: Macro-infauna of northern New England marine sand: Long-term intertidal community structure. In: Ecology of marine benthos, Vol. 6, pp 439-450, Ed. by B. C. Coull. Belle W. Baruch Library in Marine Science, Columbia, S. C.: Univ. of South Carolina Press 1977
Croker, R. A., R. P. Hager and K. J. Scott: Macroinfauna of northern New England marine sand. II. Amphipod-dominated intertidal communities. Can. J. Zool. 53, 42-51 (1975)
Dahl, E.: Some aspects of the ecology and zonation of the fauna on sandy beaches. Oikos 4, 1-27 (1952)
Dexter, D. M.: Population dynamics of the sandy-beach amphipod Neohaustorius schmitzi Bousfield. 105 pp. Ph.D. Thesis, Univ. of North Carolina, Chapel Hill, 1967
Dexter, D. M.: Structure of an intertidal sandy-beach community in North Carolina. Chesapeake Sci. 10, 93-98 (1969)
Edwards, R. and J. H. Steele: The ecology of O-group plaice and common dabs at Loch Ewe I. Population and food. J. exp. mar. Biol. Ecol. 2, 215-238 (1968)
Elton, C. S. and R. S. Miller: The ecological survey of animal communities: With a practical system of classifying habitats by structural characters. J. Ecol. 42,460-496 (1954)
Fincham, F. A.: Amphipods in the surf plankton. J. mar. biol. Ass. U.K. 50, 177-198 (1970)
Fincham, A. A.: Ecology and population studies of some inter- tidal and sublittoral sand-dwelling amphipods. J. mar. biol. Ass. U.K. 51,471-488 (1971)
Gibson, R. N.: The biology and behaviour of littoral fish. Oce- anogr. Mar. Biol. Ann. Rev. 7, 367-410 (1969)
Gibson, R. N.: The intertidal movements and distribution of young fish on a sandy beach with special reference to the plaice (Pleuronectes platessa L.) J. exp. mar. Biol. Ecol. 12, 79-102 (1973)

88 R. A. Croker and E. B. Hatfield: A Sand-Burrowing Amphipod Guild
Hager, R. P. and R. A. Croker: Macroinfauna of northern New England marine sand. IV. In: faunal ecology of Amphiporeia virginiana Shoemaker, 1933 (Crustacea: Amphipoda). Can. J. Zool. 57, 1511-1519 (1979)
Hager, R. P. and R. A. Croker: The sand-burrowing amphipod, Amphiporeia virginiana Shoemaker 1933 in the tidal plankton. Can. J. Zool. 58, 860-864 (1980)
Holland, A. F. and J. M. Dean: The community biology of intertidal macrofauna inhabiting sandbars in the North Inlet area of South Carolina, U.S.A. In: Ecology of marine benthos, Vol. 6, pp. 423-438, Ed. by B. C. Coull. Belle W. Baruch Library in Marine Science, Columbia, S.C.: Univ. of South Carolina Press 1977
Holland, A. F. andT. T. Polgar: Seasonal changes in the structure of an intertidal community. Mar. Biol. 37, 341-348 (1976)
McIntyre, A. D.: The range of biomass in intertidal sand, with special reference to the bivalve, Tellina tenuis. J. mar. biol. Ass. U.K. 50, 561-575 (1970)
Menge, B. A. and J. P. Sutherland: Species diversity gradients: Synthesis of the roles of predation, competition, and temporal heterogeneity. Am. Nat. 110, 351-369 (1976)
Nicolaisen, W. and E. Kanneworff: On the burrowing and feeding habits of the amphipods Bathyporeia piIosa Lind- strom and Bathyporeia sarsi Watkin. Ophelia 6, 231-250 (1969)
Puttick, G. M.: Spatial and temporal variations in the inter- tidal animal distribution at Langebaan Lagoon, South Africa. Trans. R. Soc. S. Aft. 42, 403-440 (1977)
Puttick, G. M.: The diet of the curlew sandpiper at Langebaan Lagoon, South Africa. Ostrich 49, 158-167 (1978)
Salvat, B.: La macrofaune carcinologique endogge des sediments meubles intertidaux (Tanaidac~s, Isopodes et Amphipodes),
gthologie, bionomie et cycle biologique. M~m. Mus. natn. Hist. nat. (Paris) Ser. A, Zool. 45, 1-275 (1967)
Sameoto, D. D.: Comparative ecology, life histories and be- havior of intertidal sand burrowing amphipods (Crustacea: Haustoriidae) at Cape Cod. J. Fish. Res. Bd Can. 26, 3 6 1 - 388 (196%)
Sameto, D. D.: Physiological tolerances and behaviottr responses in five species of Haustoriidae (Amphipoda: Crustacea) to five environmental factors. J. Fish. Res. Bd Can. 26, 2283- 2295 (1969b)
Sanders, H. L.: Marine benthic diversity: A comparative study. Am. Nat. 102, 243-282 (1968)
Sanders, H. L.: Bentic marine diversity and the stability-time hypothesis. Brookhaven Syrup. Biol. 22, 71-81 (1969)
Thijssen, R., A. J. Lever, and J. Lever: Food composition and feeding periodicity of O-group plaice (Pleuronecte~ platessa) in the tidal area of a sandy beach. Neth. J. Sea Res. 8, 3 6 9 - 374 (1974)
Tyler, A. V.: Surges of the winter flounder, Pseudopleuronectes americana, into the intertidal zone. J. Fish. Res. Bd Can. 28, 1727-1732 (1971)
Watkin, E. E.: The macrofauna of the intertidal sand of Kames Bay, Millport, Buteshire. Trans. R. Soc. Edinb. 60, 5 4 3 - 561 (1942)
Woodin, S. A.: Refuges, disturbance, and community structure: a marine soft-bottom example. Ecology 59,274-284 (1978)
Date of final manuscript acceptance: September 23, 1980. Communicated by I. Morris, West Bothbay Harbor