sound localisation in a habitat: an analytical approach to quantifying the degradation of...
TRANSCRIPT

This article was downloaded by: [University of Connecticut]On: 13 October 2014, At: 06:00Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number:1072954 Registered office: Mortimer House, 37-41 Mortimer Street,London W1T 3JH, UK
Bioacoustics: TheInternational Journalof Animal Sound and itsRecordingPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/tbio20
SOUND LOCALISATIONIN A HABITAT: ANANALYTICAL APPROACHTO QUANTIFYING THEDEGRADATION OFDIRECTIONAL CUESAXEL MICHELSEN a & KRISTIN ROHRSEITZ aa Centre for Sound Communication , Instituteof Biology, Odense University , DK-5230 ,Odense M , DenmarkPublished online: 13 Apr 2012.
To cite this article: AXEL MICHELSEN & KRISTIN ROHRSEITZ (1997)SOUND LOCALISATION IN A HABITAT: AN ANALYTICAL APPROACH TOQUANTIFYING THE DEGRADATION OF DIRECTIONAL CUES, Bioacoustics: TheInternational Journal of Animal Sound and its Recording, 7:4, 291-313, DOI:10.1080/09524622.1997.9753341
To link to this article: http://dx.doi.org/10.1080/09524622.1997.9753341
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of allthe information (the “Content”) contained in the publications on ourplatform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy,

completeness, or suitability for any purpose of the Content. Anyopinions and views expressed in this publication are the opinionsand views of the authors, and are not the views of or endorsed byTaylor & Francis. The accuracy of the Content should not be reliedupon and should be independently verified with primary sources ofinformation. Taylor and Francis shall not be liable for any losses,actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directlyor indirectly in connection with, in relation to or arising out of the useof the Content.
This article may be used for research, teaching, and private studypurposes. Any substantial or systematic reproduction, redistribution,reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of accessand use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

Bioacoustics The International Journal of Animal Sound and its Recording, 1997, Vol. 7, pp. 291-313 0952-4622/97 $10 © 1997 AB Academic Publishers
SOUND LOCALISATION IN A HABITAT: AN ANALYTICAL APPROACH TO QUANTIFYING THE DEGRADATION OF DIRECTIONAL CUES
AXEL MICHELSEN AND KRISTIN ROHRSEITZ*
Centre for Sound Communication, Institute of Biology, Odense University, DK-5230 Odense M, Denmark
ABSTRACT
Although much research has been done to describe the degradation of sound signals propagating in natural habitats, the directional cues of sound have so far been neglected. This paper describes a first approach to quantifying the degradation of directional cues in sound propagating parallel to the ground in a grassland habitat of orthopteran insects. A matched pair of probe microphones measured the sound amplitude and phase close to the ears of grasshopper carcasses for 12 evenly spaced directions of sound incidence. The degradation was found to increase with frequency and distance from the sound source and to decrease with distance from the ground. The acoustical data were used to predict how well animals with different auditory systems can determine the direction of the sender. At one position in the habitat, the predictions were compared with the pattern of phonotactic responses of live grasshoppers. Amplitude cues appear to degrade much faster with distance than phase cues. Animals exploiting phase cues may therefore maintain a reasonable directional hearing when the amplitude cues no longer make sense. The pressure-difference-receiver type of ears responds to phase differences, and these ears may be particularly suited to overcoming the degradation of directional cues. This suggests that the possession of such ears may be an adaptation not only to small body size (relative to wavelength), but also to the acoustic properties of the habitat.
Key words: sound, localisation, degradation, directional hearing, grasshopper, pressure difference receiver
INTRODUCTION
Since the mid-1970s, several biologists have been interested in the acoustic properties of the habitats of animals that communicate by sound. The pioneering studies were concerned with how sounds can be adapted to penetrate a habitat over as long distances as possible. More
*Present address: Zoology II, Biozentrum, Am Ruhland, D-97074 Wurzburg, Germany.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

292
recent research has focused on how to obtain reliable communication over the required distances, not necessarily the longest distances. The focus of the research has therefore changed from sound attenuation to wider aspects of sound degradation (e.g. Wiley and Richards 1978, 1982, Michelsen 1978, Michelsen and Larsen 1983, Dabelsteen et al. 1993). The processes leading to sound degradation are known (ground effects; multiple reflections from vegetation and other objects; effects of wind and gradients of temperature or humidity), but not understood in all details. So far the degradation of the cues for directional hearing has been neglected.
For decades, the directional properties of hearing in both vertebrates and insects have remained a popular subject for study, but almost entirely in the laboratory. Next to nothing is known about how well the biophysical and neuronal mechanisms of directional hearing work in natural habitats where sound fields are supposed to vary between the extremes of free field and diffuse field (that is, no redirected sound components and sound arriving evenly from all directions, respectively). A major step forward was the demonstration, by means of a movable neurophysiological preparation, that sounds received by a bushcricket (Orthoptera, Tettigoniidae) in dense vegetation appear to have lost their directional properties although other aspects (like rhythm) may still be available for coding by auditory neurons (Rheinlaender and Romer 1986, Romer and Lewald 1992).
Directional cues and mechanisms of directional hearing
Human ears respond to the sound pressure at the external surface of the ear drum. Above 200 Hz, the diameter of the head approaches the wavelength of sound, and diffraction causes the amplitude of sound at the ear drum to vary with the direction of sound incidence (Shaw 1974). The interaural phase difference, caused partly by diffraction but mainly by different path lengths, also varies with the direction of sound incidence. The timing of the neuronal activity may encode the phase of the sound up to a few kHz. Directional hearing may thus be based on both amplitude and phase cues within the range of sound frequencies used for speech.
Although the sound pressures acting on the ears of insects may differ much with respect to phase at the kHz-frequencies used for communication, the receptor cells show phase locking only at much lower frequencies (Michelsen 1966). Furthermore, insects are often too small relative to the wavelength of sound for diffraction to cause much difference in sound amplitude at the ears. Nevertheless, in most insect ears the amplitude of vibration of the ear drum varies much with the direction of sound incidence. The reason for this apparent paradox is
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

293
that sound reaches both the outer surface of the ear drum and the inner surface. The ear drum thus operates as a pressure difference receiver, in which the force causing the drum to vibrate depends on the relative phase of the sounds at its surfaces (Autrum 1940; for a recent review see Michelsen 1994). A similar mechanism operates in some vertebrates (frogs: J0rgensen et al. 1991; birds: Calford 1988).
A method exists for measuring the changes in amplitude and phase during the propagation of sound to the internal surface of the ear drum (Michelsen et al. 1994a), and in some insects the physics of directional hearing is now understood (crickets: Michelsen et al. 1994b, Michelsen and Li::ihe 1995; grasshoppers: Michelsen and Rohrseitz 1995). In short, the remarkable directionality found in these ears is based on sound propagating from one side of the body, through the tracheal system, to the inner surface of the ear drum at the other side of the body. The relative time of arrival of sound at the two sides of the body is very sensitive to the direction of sound incidence. Sound direction therefore affects the phase relationship between the sounds acting on the two surfaces of the ear drum, and this may lead to substantial variation in vibration amplitude.
One may say, therefore, that interaural phase differences are important in the directional hearing of most hearing animals. Some vertebrates (most mammals and birds) measure this phase difference directly by means of phase-sensitive neurons (Kuwada and Yin 1987), while most insects and some vertebrates transform differences in phase into differences in vibration amplitude at the ear drums (Michelsen 1994).
The present approach
The work described here consists mainly of acoustical measurements of the directional cues available at various positions and distances in a grassland habitat of orthopteran insects. We use the results to predict how well animals with pressure or pressure difference receiving ears can determine the approximate direction of the sender. In one case, the predictions are compared with the actual performance of grasshoppers, which indicate the perceived direction of singing conspecifics through phonotactic responses.
METHODS
Study site
The experiments were carried out in July-August 1994 near Komen, Slovenia Gust north of Triest). A wide variety of biotopes and
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

294
orthopteran insects are found in this region. A grassland biotope typical for many bushcrickets (Tettigoniidae) and short-horned grasshoppers (Acrididae) was selected for the studies. The ground at the site was flat and was covered mainly with grass with an average height of approximately 50 em. The "level of sight" (the distance from the ground at which one can see distant objects if the head is moved from side to side) was approximately 25 em. Several kinds of thinstemmed plants with narrow leaves were found among the grass plants, but no objects had widths of more than 1-2 em.
Acoustical measurements
A fast-Fourier-transform signal analyser (Hewlett-Packard 3562A) generated "chirps", which were amplified (Xelex DDS) and emitted by a loudspeaker of 43 mm diameter (VIFA D26NC). The sound was measured with two probe microphones (Bruel & Kjrer 4170, used with the dual microphone amplifier WB1057). The recorded signals returned to the analyser for processing and analysis.
The "chirps" (brief sine-sweeps similar to the FM-cries of hunting bats) were chosen for this study because they have an almost flat spectrum from 0 to 100 kHz and are so brief that the loudspeaker transmitted the part between 1 and 20 kHz in less than 2 ms. The window functions and the delay-compensating facilities of the signal analyser made it possible to restrict the analysis to a few ms after the arrival of the sound at the position of the microphone(s). This reduced the effect of extraneous noise (e.g. cicadas), the possible effects of which were evaluated by computation of the coherence function. This is a statistical measure of whether the power in the microphone response is coherent with, and thus may be caused by, the power in the chirp generated by the analyser. The coherence goes from 1 (all the output power at that frequency is coherent with the input) to 0 (none of the output power is coherent with the input). At a value of 0.8 for 25 averages, the 90% confidence limits for amplitude and phase are approximately ±15% and ±10°, respectively. We rejected all data with coherence values below 0.8.
The sound pressure amplitude and relative phase angle were measured close to the ears of female Calliptamus italicus (L.). The length of the 8 grasshoppers used varied between 26 and 30 mm. All were c. 6 mm broad at the region of the ears. A freshly killed grasshopper was glued in a natural position to the tips (1 mm diameter) of two matched probe microphones (Figure 1). 10 mm above the tips, the probe tubes were shielded by steel tubes hidden inside an external cylinder of 12 mm diameter. The shielding was necessary, because the probe tubes are somewhat transparent to sound. The direction of sound incidence was perpendicular to the long axis of the
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

295
Figure 1. The experimental set-up for measuring the degradation ofthe directional cues. The tips of two probe microphones measured the sound close to the ears of a grasshopper. Rotation of the vertical rod carrying the microphones and the animal caused the sound from the loudspeaker to arrive at various angles of sound incidence relative to the length axis of the grasshopper. The rods carrying the frame are 2 m long and 1 or 2 m apart. Further details are explained in the text. (Drawn by Per Erik Hedman).
external cylinder. With this geometry, diffraction by the cylinder caused only a small change in the sound pressure at the position of the probe tips.
The probe microphones were attached to a vertical rod, which was fastened c. 1 m above the tips and the grasshopper. The rod could be rotated and moved in the horizontal plane (Figure 1). A pointer and a protractor indicated the angle between the body of the grasshopper and the direction of sound incidence. Four rods 2 m long and 1 or 2 m apart supported the frame carrying the entire setup. All parts of the set-up were at least one meter from the grasshopper, and any possible
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

296
echoes therefore arrived too late to be included in the brief time window of the FFT -analyser.
The phase angle between the sounds at the two ears as well as the sound amplitude at each ear were determined at 12 directions of sound incidence. The loudspeaker and the microphones were at identical distances from the ground (in most experiments 2-5 or 20 em). The distance between the loudspeaker and the microphones varied between 0.6 and 4 m. In total, 1,138 sound spectra were recorded during 28 experiments. The exact positions in the habitat of the loudspeaker and microphones were not constant, even in experiments with similar distances.
Preliminary measurements showed that the interaural phase difference had to be obtained by simultaneous measurement of the sounds at the two ears, because the time spent on propagation from the loudspeaker to the animal often varied (probably due to fluctuating temperature and winds). The choice of procedure (simultaneous versus consecutive measurements) was not so critical for the amplitude component of sound.
The values were normalised: at the beginning of a series of measurements, the sound amplitude at one of the ears was designated the value 1 (0 dB) and the phase the value 0° when sound arrived from the frontal direction. All other values were expressed relative to this reference. The sound spectra were computed from time averages of 25 presentations (see Discussion).
The temperature and wind velocity were measured with a Thermo- & Anemometer (Testo 452 with the miniature probe 0635-1049 mounted at the end of a thin rod). Both acoustic and behavioural experiments took place in the morning and late afternoon (it was too hot in the middle of the day, both for the grasshoppers and for us).
Behavioural experiments
For the behavioural experiments we took advantage of the phonotactic behaviour of males of the acridid grasshopper Chorthippus biguttulus (L.): To achieve pair formation, the male sings its calling song, to which a receptive female may answer with her own song. Most males then make an abrupt turn (followed by some walking) towards the direction of the female. This orientation is based on a simple left-right decision: A difference of only 1-2 dB in the sound levels at the ears is needed for eliciting a turn towards the louder side (Helversen and Rheinlaender 1988). Other males respond by jumping towards the sound source (Gilbert 1995), probably because the difference in sound level is below 1-2 dB. The male may start a new sequence by singing again. Using this strategy, the male eventually reaches the female (Helversen 1972, Helversen and Helversen 1983).
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

297
The males were caught close to the study site and tested for responsiveness to female song. Three responsive males were selected for the study. The highly motivated male was placed at the centre of a circular platform (radius 2.5 em) 5 em above the ground in dense vegetation very similar to that at the site for the acoustic measurements. The 800 ms sequence of female song used in our experiments had earlier been selected by Dagmar von Helversen as an efficient releaser of the phonotactic response in a wide variety of males. It was played back to the males from a tape recorder (Uher 4200 Report Monitor, frequency range 0.8-18 kHz) through a loudspeaker (Vifa D26NC) 5 em from the ground and pointing at the platform carrying the male 4 m away. The sound level at the platform was kept constant at approximately 50 dB SPL (the optimum value for release of the phonotactic response).
The phonotactic turning response is very simple: The males turn their bodies about 90°, and the magnitude of the turn is not scaled to the target angle (the angle between the body axis and the direction of sound incidence). The absence of scaling, which occurs in, for example, crickets and bushcrickets, and the stereotyped nature of the response made it easy for us to score the responses into three classes: turns to the correct side, erroneous turns, and jumps. The approximate target angle was estimated by means of a protractor on the platform. The males were free to move on the platform between the tests, and they therefore never occupied exactly the same position at the beginning of the tests. Each of the three males contributed approximately one third of the 126 responses recorded. The response patterns of the three males were fairly similar, and the data for Figure 4A were therefore pooled.
RESULTS
Acoustical measurements
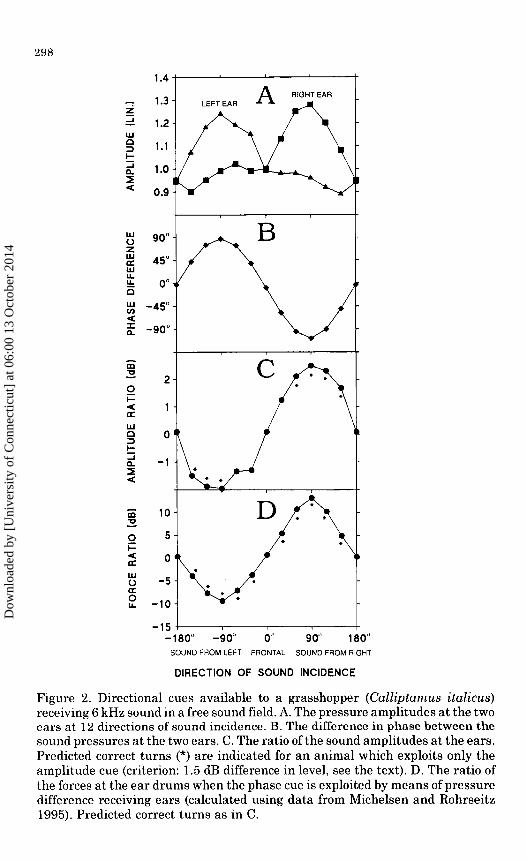
The females of the grasshopper Calliptamus italicus (L.) used for the acoustical part of the study were 26-30 mm long and c. 6 mm broad at the region of the ears. Figure 2A shows an example of the pressure amplitude at 6 kHz, measured at each ear for 12 directions of sound incidence in a free sound field (the loudspeaker and animal were 40 em apart and 40 em from the ground). Very similar amplitudes were recorded at the two ears when the sound arrived from the forward and backward directions (0° and 180°, respectively). Diffraction of sound due to the presence of the grasshopper caused a surplus pressure at an ear when the sound source was ipsilateral to that ear (90° for the right ear, -90° for the left ear). Only a slight "shadow" is seen at the other ear. As expected, the pattern observed around -90° at the left ear was
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4
298
:Z :::::;
w c :::J 1-:::::; a. ~ <
w u z w a: w u.. u.. 0 w (/)
< ::c a.
0 j::: < a: w
1.4
1.3
1.2
1.1
1.0
0.9
90"
45"
0"
-45"
-90"
2
g 0 1-:::::; a. -1 ~ <
iii 10 ~
0 5 j::: < 0 a: w -5 u a: 0 -10 u..
-15
RIGHT EAR
c
D
-180" -90" 0" 90" 180" SOUND FROM LEFT FRONTAL SOUND FROM RIGHT
DIRECTION OF SOUND INCIDENCE
Figure 2. Directional cues available to a grasshopper (Calliptamus italicus) receiving 6kHz sound in a free sound field. A. The pressure amplitudes at the two cars at 12 directions of sound incidence. B. The difference in phase between the sound pressures at the two ears. C. The ratio of the sound amplitudes at the ears. Predicted correct turns (*) are indicated for an animal which exploits only the amplitude cue (criterion: 1.5 dB difference in level, see the text). D. The ratio of the forces at the ear drums when the phase cue is exploited by means of pressure difference receiving ears (calculated using data from Michelsen and Rohrseitz 1995). Predicted correct turns as in C.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

299
almost a mirror image of that seen around 90° at the right ear. The central nervous system needs the driving forces at the ears
to differ by 1-2 dB in order to decide whether the sound source is to the right or the left (Helversen and Rheinlaender 1988). The ratio of the sound amplitudes at the two ears is shown in Figure 2C, which also indicates the expected decisions of a hypothetical animal of the size of C. italicus, equipped with pressure receiving ears, and supposed to be incapable of exploiting phase or other time cues. A 1.5 dB difference was chosen as the criterion for the left-right decision. A decision to turn to the correct side is indicated by "*". As expected, turns are absent at the directions 0° and 180°, but they are also absent at -60°, -30°, and 30°. At these three directions, the 1.5 dB criterion is not met, although a turn is expected.
The message of Figure 2C is that an animal of the size of C. italicus, which relies only on measuring the pressure amplitudes at the ears, would make mainly correct decisions when listening to a 6 kHz tone in a free sound field (no turn at oo and 180°; a turn towards the most stimulated ear at most other directions of sound incidence). At three directions, however, the animal would not be able to decide on a correct turn. The data in Figure 2C and other similar observations show that the hypothetical animal would often have troubles at the 30° and -30° directions.
So far, our hypothetical animal has made use only of amplitude cues. In addition, a phase cue (the interaural phase difference) is available (Figure 2B). Grasshoppers and many other small animals can exploit this phase cue in their directional hearing by allowing sound to propagate from one side of the body, through some internal pathway (in grasshoppers tracheal air sacs separating the ears), to the internal surface of the ear drum (see Introduction). This mechanism allows the animal to exploit the phase cue, and it causes the ratio of the forces acting on the two ear drums to vary much more with the direction of sound incidence than the ratio of the sound amplitudes. In other words, the exploitation of the phase cue improves the pattern of directionality. Calculations show that a real grasshopper in a free sound field would make correct decisions at all the 12 directions of sound incidence (Figure 2D). [The fairly complicated calculations are described by Michelsen and Rohrseitz (1995). We assumed the gain of the sound path to be 0.5 and -60°, typical values for 6 kHz in a grasshopper of this size. These figures mean that sound arriving at the internal surface of an ear drum has been attenuated by a factor of 2 and delayed by 60° by propagating through the other ear and through the tracheal air sacs].
The free sound field used for obtaining the data in Figure 2 is the most favourable acoustical situation that one can imagine for directional hearing. In a total of 28 similar experiments, the preparation was placed at various distances from the loudspeaker in a natural habitat (see Methods: Study site). The distance from the ground also
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

:300
2.0
A RIGHT EAR
~ 1.5 ....1
w 0 1.0 ::::l 1-....1 a.. ::::!: <
0.5
w 180" B (.) z w 90" a: w u.. u.. 0" 0 w -90" (J)
< it -180"
iii c ~ 10 0 1- 5 E < a: E
w 0 0 ::::l -5 E E 1-....1 a.. -10 ::::!: <
iii 10 D ~
0 5 . . 1-< 0 a: w -5 (.) a: 0 -10 u..
E
-15 -180" -90" 0" 90" 180"
SOUND FROM LEFT FRONTAL SOUND FROM RIGHT
DIRECTION OF SOUND INCIDENCE
Figure 3. Degraded directional cues available to a grasshopper receiving 10kHz sound. 2 em from ground and 4 m from the loudspeaker. A. The pressure amplitudes at the two ears at 12 directions of sound incidence. B. The phase cue (cf. Figure 2B). C. The ratio of the sound amplitudes at the ears. Predicted lateralisation responses as in Figure 2C (E: erroneous turns). D. The ratio of the forces at the ear drums in a pressure difference receiving grasshopper (cf. Figure 2D).
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

301
varied (it was always the same for the loudspeaker and the preparation). In each experiment, the expected decisions of a hypothetical animal responding only to pressure amplitude were calculated from the pressures at the two ears.
Several examples were found of positions in the habitat at which directional hearing should be very difficult. One of these examples (2 em from ground, 4 m from a 10kHz sound source) is shown in Figure 3. The amplitude cue (Figure 3A) is obviously much degraded (compare with Figure 2A), and the amplitude ratio (Figure 3C) is far from the pattern of 2C. Our hypothetical amplitude-receiving animal makes only 5 correct turns ("*") plus the correct decision not to turn at the 180° direction. However, it makes 4 errors (E): a turn at 0° and a turn to the wrong side at the directions -150°, 30°, and 150°. Furthermore, it fails to make a turn at the directions -90° and 60°. This is only a slightly better score than a fifty-fifty situation, in which sound localisation is impossible. The phase cue (Figure 3B) is much less degraded than the amplitude cue, however, and animals that are able to exploit the phase cue may still be able to determine the sound direction with only few errors.
The calculated ratio of the forces acting on the ear drums is shown in Figure 3D (the assumed gain was 0.5 and -80°, which are typical values for 10 kHz, see Michelsen and Rohrseitz 1995). The animal now makes 9 correct turns ("*") and fails to turn at the oo direction. However, it fails to turn at 150° and makes a (wrong) turn at 180°. A close look at the data reveals that the turn at 180° was caused by a large degradation of the phase cue at that direction.
Although the degradation of the phase cue may thus be substantial, in general the phase cue is less affected by degradation than the amplitude cue. The robustness of the phase cue probably reflects the fact that both the time of arrival of sound at the ears and the sound diffraction contribute to the phase cue, whereas only the latter process is responsible for the amplitude cue.
Monitoring the effect of degradation with a live grasshopper
Although the protective cylinder around the microphone probes had little effect on the accuracy of the acoustical measurements, it forced us to use a fairly large species of grasshopper for the acoustical measurements. Unfortunately, the large species is lacking the phonotactic lateralisation response to conspecific sound found in males of the smaller Chorthippus biguttztlzts (see the Introduction). The dimensions of female Calliptamus italicus are approximately twice those of male C. biguttulus, but the scaling of diffraction around grasshoppers is not simple (Michelsen and Rohrseitz 1995). We find that the amplitude and phase data obtained in a free sound field at 6 kHz (Figure 2) are
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

302
very similar to those obtained under similar conditions at 10kHz in C. biguttulus (Michelsen and Rohrseitz 1995).
Despite these complications, we decided to use the phonotactic behaviour of male C. biguttulus for monitoring the accuracy of directional hearing in an environment with degraded directional cues. On hearing the sound from a female, most of the motivated males made an abrupt turn towards the direction of the most stimulated ear (see Methods). However, some males did not respond to the female song by turning, but by jumping. This reaction was especially common when the direction to the "female" (here the loudspeaker) was in the forward (0°) direction. Such jumps are generally directed at the sound source (Gilbert 1995), and we assume that jumping is released when the two ears are almost equally stimulated.
The results of the acoustical measurements mentioned above indicated that a sound path 5 em above ground in dense grass and a distance of 4 m provided a degradation of the directional cues that was sufficient to affect the accuracy of the directional hearing, but not so large that the animals were facing an impossible task. In each experiment, the angle between the body axis of the male and the direction of sound incidence was noted at the time of sound stimulation. 90° means that the loudspeaker was ipsilateral to one of the ears. The female song was not presented when the males faced away from the loudspeaker.
The pooled results of the observation of 126 reactions from 3 animals are shown in Figure 4A (note the different numbers of experiments in each direction). Obviously, the animals performed very well, despite the large distance to the loudspeaker. At 45-90°, the animals always turned to the correct side. In the more frontal directions, several turns away from the loudspeaker (errors, indicated with solid fill) and jumps (cross-hatched) occurred. In the frontal direction (0°), a turn to either side is an error.
Acoustical measurements were performed 2-5 em above ground and at a distance of 3 or 4 m between microphones and loudspeaker. The predictions (Figure 4B) were made by the pooling of data from 6, 8, and 10 kHz, because this frequency range is necessary for eliciting phonotaxis in C. biguttulus (D. von Helversen, personal communication). A comparison of Figures 4A and 4B shows much similarity. Obviously, both the live grasshoppers and the hypothetical amplitudereceiving animal have troubles in the forward direction. At other directions, the grasshoppers tend to make fewer errors than predicted, but because of the small number of behavioural observations, the scores in 4A and 4B are not significantly different at the individual directions of sound incidence.
We should like to stress that we used C. biguttulus only as a monitor for estimating the effect of the degradation on hearing. These experiments are not relevant for the phonotactic behaviour of this animal in its own habitat (see the Discussion).
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

303
DIRECTION OF SOUND INCIDENCE
Figure 4. Observed (A) and predicted (B) phonotactic responses. A. Reactions of three male Chorthippzts biguttulu.s presented with degraded female response song at various directions of sound incidence. *:turns to the correct side. Crosshatched: jumps. Solid filled: erroneous turns. The number of experiments at each direction are indicated above the bars. B. Predicted responses of a hypothetical animal responding only to the amplitude component of sound pressure (cf. Figures 2C and 3C). Symbols as in A, but cross-hatched means that the amplitudes differed by less than 1.5 dB. The figure is based on the amplitudes measured at the ears of female Calliptamus italicus at 6, 8, and 10 kHz. The number of data at each direction are indicated above the bars.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

304
Scatter as a measure of degradation
It is possible, then, to get some insight into the magnitude of the degradation of the directional cues by determining the number of errors in the directional hearing of both hypothetical animals with various kinds of auditory periphery and live animals. Further analysis using the former approach revealed that the degradation tended to increase with the distance to the sound source and to decrease with the distance from the ground.
The number of errors also tended to increase with sound frequency. The trend was weak, however, except at the 0° and 180° directions (where the expected number of errors at 6 kHz was significantly lower than at 8 and 10 kHz). The reason for the weak trend at other directions is that the data reflect two opposite trends. The scatter of sound amplitude increases with frequency, but so does the diffraction of sound by the body of the grasshopper. The method is thus biased by the size of the animal used during the collection of the primary acoustical data.
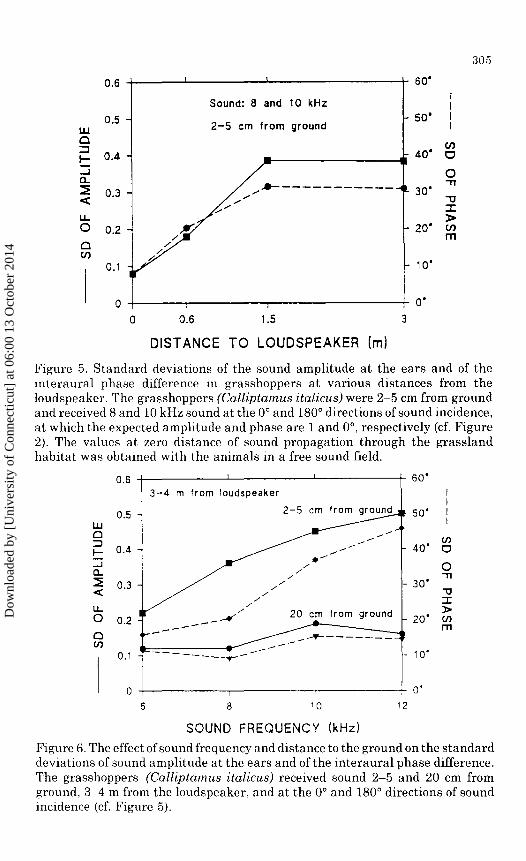
Diffraction around a cylindrical body like that of a grasshopper causes some change in the sound pressure at the surface at all directions of sound incidence, but the change is a minimum at the oo and 180° directions. A minimum of diffraction means that one may here have a better chance to observe variations in the scatter of sound amplitude. We took advantage of this minimum by calculating the amount of scatter in the pressure amplitudes and interaural phase differences recorded at these directions. This procedure resulted in very significant trends (Figures 5 and 6).
Figure 5 shows how the standard deviations of sound amplitude and interaural phase increase as the sound is propagating 2-5 em from ground (data from 8 and 10 kHz were pooled). The value at "distance zero" is the standard deviation calculated from measurements in a free sound field. The differences between the amplitude values at 0, 0.6, and 1.5 m are significant at the p < 0.01 level (F test for variances; Armitage and Berry 1987).
The standard deviation of both the amplitude and phase cues was much larger 2-5 em from the ground than 20 em from ground (Figure 6). A clear dependence on sound frequency was also found, but only at a distance of 2-5 em from ground. Both of these findings were also highly significant.
DISCUSSION
The present approach is quite pragmatic. We do not attempt to analyse the physics of sound degradation, but merely describe how the sounds at the ears change as a result of the degradation. We have tried to
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4
305
0.6 so•
Sound: 8 and 10 kHz
0.5 so· w 2-5 em from ground a (J) :::::J
0.4 40' 0 I-..J 0 a.. _._ ___________ .., ~ 0.3 /
/ 30' <( / -u
'/ / :I:
u.. )> 0 0.2 ~ 20' (J)
/ m a /
(J) /. h
0.1 10'
0 o· 0 0.6 1.5 3
DISTANCE TO LOUDSPEAKER (m)
Figure 5. Standard deviations of the sound amplitude at the ears and of the interaural phase difference in grasshoppers at various distances from the loudspeaker. The grasshoppers (Calliptamu.s italicu.s) were 2-5 em from ground and received 8 and 10kHz sound at the 0° and 180° directions of sound incidence, at which the expected amplitude and phase are 1 and 0°, respectively (cf. Figure 2). The values at zero distance of sound propagation through the grassland habitat was obtained with the animals in a free sound field.
0.6 60'
3-4 m from loudspeaker
0.5 2-5 em 50' UJ Cl (Jl :::::J -0.4 -- 40' 0 I- .---l / 0 a. /
:::E /
.., 0.3 / 30'
<( / -u / / I
u.. / )> / 20 em from ground 0 0.2
_ _.-20' (Jl ---- m
Cl (Jl
0.1 ---~- 10'
0 a· 6 8 10 12
SOUND FREQUENCY (kHz)
Figure 6. The effect of sound frequency and distance to the ground on the standard deviations of sound amplitude at the ears and of the interaural phase difference. The grasshoppers (Calliptamus italicus) received sound 2-5 and 20 em from ground, 3-4 m from the loudspeaker, and at the 0° and 180° directions of sound incidence (cf. Figure 5).
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

306
predict the possibilities for directional hearing in animals that exploit only sound amplitudes and in animals that also exploit interaural phase differences. In one case, we have compared the predictions with the phonotactic behaviour of a live grasshopper. The observed and predicted performances at various angles of sound incidence look similar (Figures 4A and 4B), but the interpretation is not simple (see below).
Directional hearing in degraded sound fields
Directional hearing requires that the sound field has some directional characteristics. The optimum situation for directional hearing is some distance away from the sound source in a homogeneous space, in which sound propagates outwards from the source, and no redirected components are present. In most laboratory studies of directional hearing, the investigators aim at establishing such a uni-directional free sound field, but free sound fields are rare in natural habitats.
The other extreme is the diffuse field. Some acoustic laboratories contain rooms with a complex geometry, hard walls, and a minimum of absorption. Under such conditions, the energy density may be almost the same at all points in space, and there may be an equal probability that sound arrives from any direction. So, directional hearing has become impossible (unless the animal is able to exploit a precedence effect).
At some positions in the habitat studied here, the acoustics showed clear signs of degradation. The amplitude cue (Figures 3A and 3C) may be so degraded that directional hearing is difficult or even impossible in animals which only measure sound amplitudes. This seems to be the case in many bushcrickets. In these animals, the ear drums also receive sounds both at their external and internal surfaces, but the sound path to the internal surface (a horn-shaped tracheal tube, which opens at the lateral surface of the body) has a considerable gain in most species. The sound at the internal surface may dominate the ear, which therefore is a pressure receiver with its input at the body surface (Lewis 1983, Michelsen et al. 1994a).
As already mentioned, Rheinlaender and Romer (1986) used a movable neurophysiological preparation for demonstrating the degradation of the directional cues. A bushcricket (Tettigonia viridissima) received 20 kHz sound pulses from a loudspeaker which was 1.5 m from ground and 10 m from the animal. Figure 7 shows the responses of two interneurons when the preparation was at three different distances from the ground. At 1.5 m (the height of the dense bushes) the directional hearing was as perfect as in Figure 2. At 0. 75 m, two errors occurred. Finally, directional hearing was abolished when the preparation was standing on the ground (cf. Figure 3A), although the
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

l.Sm above ground ~ 8~----~------~----~-----r----~----~
~ ·- ----· - o_ -• E61--o ___ b l right :;; left 1' t interneuron ~ interneuron ', -4 ' ~ '\
~ T ~'- T - • ·---- -o ~ 2 ] _____ 1 1 ..... _ ~ - -o---o :0 I I I I I ' 0 ..... ~ 0.75m above ground
E 4 ./"·o • .............- 1' ........
T/ ' 0 ---o 2·~·--r ] o~l--~~~--~~~-
• 60
sound from left
30 •
frontal
on ground
sound from right
307
Figure 7. Responses recorded from a pair of direction-sensitive auditory interneurones (T -fibres) in a bushcricket, at various directions of sound incidence in dense bushland. Further information in the text. Reproduced from Rheinlaender and Romer (1986).
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

308
neurons still coded the rhythmical structure of the song. (Note that in this experiment the sound level was constant at the loudspeaker, not at the receiving animal, so the sound was also attenuated near the ground).
A similar technique was used by Romer and Lewald (1992), but they used the responses of the non-directional omega neuron to characterise the acoustical properties of a plantation of shrubs. The amount of scatter in the neural responses to sound led them to expect "a more or less diffuse sound field around the receiver. As a consequence of these effects, the localisation, but not the detection, of sound may be impossible at unfavourable positions in the habitat". In contrast, we find that degraded sound fields with useless amplitude cues may still be very different from a diffuse field, since a phase cue is available. The phase cue has also degraded, but it is sufficient for allowing a very useful directional hearing, for example in animals responding to pressure differences across the ear drum (Figure 3D).
A comparison of Figures 7 and 3D may serve to illustrate the improvement in directionality made possible by the mechanical exploitation of the phase cue in a pressure difference receiver. The finding that ears working as pressure difference receivers are particularly well suited for overcoming the degradation of the directional cues suggests that the possession of such ears may be an adaptation not only to small body size, but also to the kind of habitat. This may perhaps be the reason why some bushcrickets have acoustic tracheae with a much reduced gain, thus providing the ears with pressuredifference properties (Stumpner and Heller 1992). It would be interesting to compare the directional hearing made possible by pressuredifference-receiving ears with that based on exploiting a neural phase processing in the brain. Which mechanism is superior, and why?
The problem with forward directions
The percentage of correct lateralisation responses in a degraded sound field depends on the angle between the body axis and the direction of sound incidence (Figure 4). It is more difficult to make correct decisions when the sound arrives from frontal directions than when sound is ipsilateral to one of the ears. This appears to be true for animals that exploit only amplitude cues (Figure 4B) as well as for animals that are equipped with pressure-difference-receiving ears capable of exploiting also the phase cue (Figure 4A). This result is not surprising, considering the respective mechanisms involved. Both the amplitude cue and the phase cue are zero when sound arrives from the frontal direction, and they are a maximum when the sound source is ipsilateral to one of the ears. Apparently, the scatter in degraded sound fields may be larger than these directional cues at the frontal
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

309
directions of sound incidence, whereas the ratio between cues and scatter is more favourable in more lateral directions. The dominant role of lateralisations with an absence of scaling (see Methods) in the phonotaxis of grasshoppers like C. biguttulus may be a strategy for a voiding the critical forward direction.
It is tempting, but too ambitious, to compare the percentages at each direction in Figures 4A and 4B. Several assumptions were made in the measurements and calculations leading to Figure 4B, and only little is known about the neural processing in the lateralisation behaviour of Figure 4A. The animals of Figure 4A were listening to songs carried by a broad range of frequencies, and it is not known how much different frequency bands contributed to the lateralisation response. In contrast, the predictions in Figure 4B were made by pooling data from three frequency bands centred at 6, 8, and 10 kHz, respectively (because of the short duration of the sound pulses, the effective bandwidth around each frequency was approximately 500 Hz). In bushcrickets, Romer and Lewald (1992) have shown that the variability of the neuronal responses to sounds decreases with the bandwidth of the stimulus, and recent experiments similar to that in Figure 7 have indicated a smaller number of errors with broad band sounds instead of pure tones (Heiner Romer, personal communication).
The temporal integration of directional information in the listening grasshopper is also unknown. This is true both for the duration of each syllable in the song and for the much longer lasting repetition of syllables. The chirps used in the acoustical measurements were much briefer than the syllables of C. biguttulus (approximately 2 and 80 ms, respectively). We chose to average 25 chirps in each acoustical measurement, but it remains to be learned whether the temporal integration in the grasshopper causes a similar reduction of scatter.
It should be noted that C. biguttulus was chosen for the experiments because of its remarkable phonotactic response behaviour, and that the experimental conditions differed much from those prevailing during the normal social communication in this species. The songs of C. biguttulus are not very intense, and the range of communication in a grassland habitat is· generally much shorter than the 4 m chosen here. Furthermore, this species prefers habitats with much shorter grass (Gilbert 1995). We simply adjusted the output level from the loudspeaker to provide the optimum sound level for the release of the phonotactic response at the position of the listening males, since we were only interested in the ability of the animals to deal with the degradation of the directional cues.
Some insects have similarly loud calls. Male field crickets calling on the ground may be heard by females on the ground at a distance of 6 m; the distance increases to 15-22 m if the animals move away from ground (Paul and Walker 1979). The results of Figure 6 suggest that
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

310
the species communicating on the ground should prefer rather low frequencies (European field crickets do indeed use 4-5 kHz), whereas species communication at some distance from ground may have a wider choice. Of course, other aspects than directional hearing may affect the optimum frequency for communication.
Mechanisms leading to directional degradation
One mysterious finding in this study was the lack of right-left symmetry in the amplitudes recorded in degraded sound fields (compare Figures 3A and 2A). We do not have any convincing explanation for this. Several physical mechanisms are likely contributors to the degradation of the directional cues, but more sophisticated investigations will be needed if we are to understand the nature of the processes involved. The interactions of sound waves with the ground are complicated and not well understood. Most studies of outdoor acoustics have concentrated on frequencies up to a few kHz, and sound reflections from grass-covered ground change dramatically at higher frequencies (Michelsen and Larsen 1983). The interactions between sound waves and vegetation are also likely to change much with frequency, because diffractional effects depend on object size relative to wavelength.
Sound propagation outdoors is further complicated by being affected by processes that vary with time. Temperature gradients are not stable, and air turbulences are among the processes known to affect sound propagation. Wind can create large turbulences, and in addition it causes the vegetation to move, thus giving rise to fluctuations in the spatial distribution of sound pressure in the vegetation. The experiments reported here were performed when both wind speeds and temperature gradients were small. The degradation of the directional cues at normal weather conditions may thus turn out to be more prominent than that reported here.
The relative magnitude of the effects of the various factors appears to depend on the distances and habitats studied. Wiley and Richards (1978) measured amplitude fluctuations below 12 kHz, at distances up to 80 m, and 2 m from ground, in a mixed deciduous forest. They found the fluctuations to increase with frequency and distance between emitter and receiver, but even a slight wind caused this dependence to disappear. In contrast, in several studies of sound propagation over shorter ranges in vegetation this dependence has been reported to persist (Michelsen and Larsen 1983, Romer and Lewald 1992, this study). Similarly, the frequency spectrum of the amplitude fluctuations was found to be independent of carrier frequency by Wiley and Richards (1978), whereas Michelsen and Larsen (1983) found the fluctuation frequency to increase with carrier
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

311
frequency. The physical interpretation of the latter finding might be that plants with thin stems and leaves (which are only affecting high frequency sounds) may vibrate with higher frequencies than larger plants. It seems safe to conclude that different physical processes are leading to degradation of the directional cues in different environments.
Where do we go from here?
Obviously, the present study has resulted in more questions than answers. One approach in future work would be to use the methods described here to study a number of different environments. So far, estimates of the optimum frequency for communication in a certain environment have been based mainly on measurements of excess attenuation, but the degradation of the directional cues may also depend on frequency (Figure 6). In the present study, the sound emitter and sound receiver were always at the same distance from ground. In several animals, this is not so.
It remains to be learned, whether some communication strategies are adapted to minimise the degradation of the directional cues. It would be interesting to compare the directional hearing in natural habitats in animals with pressure receiver ears and animals with pressure difference receiver ears. Are some animals with pressure difference receivers adapted to special environments rather than just compensating for being small? The clear and reliable phonotactic behaviour makes males of the grasshopper Chorthippus biguttulus ideally suited for studies of sound localization. However, the complicated acoustics of its auditory system and unknown features of the neuronal processing makes it difficult to interpret the results. So, for further studies the relative contributions of different frequency bands to eliciting the phonotactic behaviour should be known. Also, it remains to be learned whether grasshoppers exploit temporal integration in order to reduce scatter. It would be advantageous for future work, if a similar reliable behaviour could be demonstrated in other animals. Especially, it would be interesting to study the behaviour of animals that allowed a clear separation of the use of amplitude and phase cues.
ACKNOWLEDGMENTS
The Centre for Sound Communication is financed by the Danish National Research Foundation. We are most grateful to Matija Gogala for hospitality and help during the field work, to the company Vifa A/S for donating the loudspeakers, and to Dagmar von Helversen for
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

312
placing a recording of female song at our disposal. We thank Jakob Christensen-Dalsgaard, Norbert Elsner, Morten Buhl J0rgensen, Ole Nmsbye Larsen, Bertel M0hl, Simon Boel Pedersen, and two anonymous referees for their comments on this paper.
REFERENCES
Armitage, P. and Berry, G. (1987). Statistical Methods in Medical Research. Oxford; Blackwell.
Autrum, H. (1940). Uber Lautii.usserungen und Schallwahrnehmung bei Arthropoden. II. Das Richtungshi:iren von Locusta und Versuch einer Hi:irtheorie fiir Tympanalorgane vom Locustidentyp. z. vergl. Physiol, 28, 326-352.
Calford, M. B. (1988). Constraints on the coding of sound frequency imposed by the avian interaural canal. J.comp.Physiol.A, 162, 491-502.
Dabelsteen, T., Larsen, O.N. and Pedersen, S.B. (1993). Habitat-induced degradation of sound signals: Quantifying the effects of communication sounds and bird location on blur ratio, excess attenuation, and signal-to-noise ratio in blackbird song. J. Acoust. Soc. Am., 93, 2206-2220.
Gilbert, F. (1995). Partnerfindung und akustische Orientierung bei Ch. biguttulus im nati.irlichen Lebensraum. Thesis, University of Gi:ittingen.
Helversen, D. von. (1972). Gesang des Mannchens und Lautschema des Weibchens bei der Feldheuschrecke Chorthippus biguttulus (Orthoptera, Acrididae). J.comp. Physiol., 81, 381-422.
Helversen, D. von and Helversen, 0. von. (1983). Species recognition and acoustic localization in Acridid grasshoppers: a behavioral approach. In Neuroethology and behavioral physiology (F. Huber and H. Mark!, eds.). Springer-Verlag; Berlin. Heidelberg, pp. 95-107.
Helversen, D. von and Rheinlaender, J. (1988). Interaural intensity and time discrimination in an unrestraint grasshopper: a tentative behavioural approach. J. comp.Physiol. A, 162, 333-340.
Jorgensen, M. B., Schmitz, B. and Christensen-Dalsgaard, J. (1991). Biophysics of directional hearing in the frog Eleutherodactylus coqui. J.comp.Physiol. A, 168, 223-232.
Kuwada, S. and Yin T.C.T. (1987). Physiological studies of directional hearing. In Directional hearing (W.A Yost and G. Gourevitch, eds.). Springer-Verlag; New York, pp. 146-176.
Lewis, B. (1983). Directional cues for auditory localization. In Bioacoustics, a comparative approach (B. Lewis, ed.). Academic Press; London, pp. 233-257.
Michelsen, A (1966). Pitch discrimination in the locust ear: Observations on single sense cells. J. Insect Physiology, 12, 1119-1131.
Michelsen, A (1978). Sound reception in different environments. In Sensory ecology (M.A. Ali, ed.). Plenum; New York, pp. 345-373.
Michelsen, A (1994). Directional hearing in crickets and other small animals. In Neural basis of behavioural adaptations (K. Schildberger and N. Elsner, eds.). Fortschr. Zoo!. 39, Gustav Fischer; Stuttgart, pp. 195-207.
Michelsen, A and Larsen, O.N. (1983). Strategies for acoustic communication in complex environments. In Neuroethology and behavioral physiology (F. Huber and H. Markl, eds.). Springer-Verlag; Berlin, pp. 321-331.
Michelsen, A, Heller, K.-G., Stumpner, A and Rohrseitz, K. (1994a). A new biophysical method to determine the gain of the acoustic trachea in bushcrickets. J.comp. Physiol. A. 175. 145-151.
Michelsen, A, Popov, A.V. and Lewis, B. (1994b). Physics of directional hearing in the cricket Gryllus bimaculatus. J.comp.Physiol. A., 174, 153-164.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4

313
Michelsen, A. and LOhe, G. (1995). Tuned directionality in cricket ears. Nature, 375, 639. Michelsen, A. and Rohrseitz, K. (1995). Directional sound processing and interaural
sound transmission in a small and a large grasshopper. J. exp. Biol., 198,1817-1827. Paul, R.C. and Walker, T.J. (1979). Arboreal singing in a burrowing cricket,
Anurogryllus arboreus. J.comp.Physiol., 132, 217-224. Rheinlaender, J. and Romer, H. (1986). Insect hearing in the field. I. The use of
identified nerve cells as "biological microphones". J. comp. Physiol. A, 158, 647-651. Romer, H. and Lewald, J. (1992). High-frequency sound transmission in natural habitats:
implications for the evolution of insect acoustic communication. Behav. Ecol. Sociobiol., 29, 437-444.
Shaw, E. A. G. (1974). The external ear. In Handbook of sensory physiology Vol. V/1 (W. D. Keidel and W. D. Neff, eds.). Springer-Verlag; Berlin, Heidelberg, New York, pp. 455-490
Stumpner, A. and Heller, K.-G. (1992). Morphological and physiological differences of the auditory system in three related bushcrickets (Orthoptem: Phaneropteridae, Poecilimon). Physiol. Entomol., 17, 73-80.
Wiley, R.H. and Richards, D.G. (1978). Physical constraints on acoustic communication in the atmosphere: implications for the evolution of animal vocalisations. Behav. Ecol. Sociobiol., 3, 69-94.
Wiley, R.H. and Richards, D.G. (1982). Adaptations for acoustic communication in birds: transmission and signal detection. In Acoustic communication in birds. Vol. I (D.E. Kroodsma and E.H. Miller, eds.). Academic Press; New York, pp. 131-181.
Received 12 August 1996, revised 2 December 1996 and accepted 6 December 1996.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
onne
ctic
ut]
at 0
6:00
13
Oct
ober
201
4