some aspects of the morphology of the ovule and seed ofcostus malortieanus (zingiberaceae)
TRANSCRIPT

Proc. Indian Acad. Sci., Vol. 86 B, No. 3, September 1977, pp. 175-179, ~) Printed in India.
Some aspects of the morphology of the ovule and seed of Costus malortieanus (Zingiberaceae)
JOSE K MANGALY and K SWORRUPANANDAN Department of Botany, University of Calicut, Calicut 673 635
MS received 7 February 1977; revised 18 July 1977
Abstract. The ovules of Costus malortieanus are typically anatropous and are arranged along two lines on each septum, of the tricarpellary syncarpous inferior ovary. The ovular traces develop from the peripheral bundles of the axial vasculature. The ovule has a number of linear, translucent, multicellular appendages arising from the base and they are ontogenetically similar to ovular appendages in other Zingiberaceae. These appendages are derived partly from the funicle and partly from the free end of the outer integument; Jn C. malortieanus they are developed before anthesis and fertiliza- tion. The ovular appendages elongate considerably in the seed and entwine it irregu- larly. Seeds are more or less ovoid, compressed dorsiventrally and with .the ventral surface highly wrinkled. It is concluded that the dual origin of the ovular appendages is a result of the anatropous nature of the ovule and may be of phylogenetic significance.
Keywords. Costus malortieanus; Zingibexaeeae; Ovules and seed morphology.
1. Introduction
Ovules of Zingiberaceae are typically anatropus, bitegmic and bearing characteristic ovular apppendages (Panchaksharappa 1959, 1961, 1962a, 1962b, 1962c, 1966, 1970; Davis 1966; Sachar and Arora 1963). In most cases the appendages are lacerate as in Zingiber, but may be connate and veil-like as in Elettaria (Panehaksharappa 1962b, 1966). In the genus Costus, which recently is separated by some taxonomists to Costaeeae, the ovules differ from other Zingiberaceae in having a soft parenehymatous pad of tissue at the micropylar end instead of ovular appendages (Panchaksharappa 1961, 1962a).
Ovular morphology, particularly the number of integuments and presence of addi- tional structures like aril, caruncle, etc. are usually characteristic of natural groups (Maheshwari 1950), and therefore, are of taxonomic significance. The characteristic difference in ovular morphology of Costus (C. malortieanus H. Wendl.), thus gains significance in the light of the recent suggestion (Nakai 1941 ; Tomlinson 1956, 1962), that the genus must be removed from Zingiberaeeae to a new family Costaceae. During the course of detailed taxonomic study of South Indian Zingiberaeeae, we observed that the characteristic peculiarity of the ovule of C. malortieanus is not universal in the genus.
175
e.Ce)--3

176 Jose K Mangaly and K Sworrupanandan
2. Materials and Methods
Materials for the present study were collected from plants cultivated in the Calicut University Botanical Garden. Fresh material was used in most cases. For detailed observation microtome sections were prepared following the NBA-paraffm method; preceded by fixing in FAA; crystal violet and erythrosin were used for staining. In addition, ovules were cleared by KOH at 60°C and stained with saffranin.
3. Observation
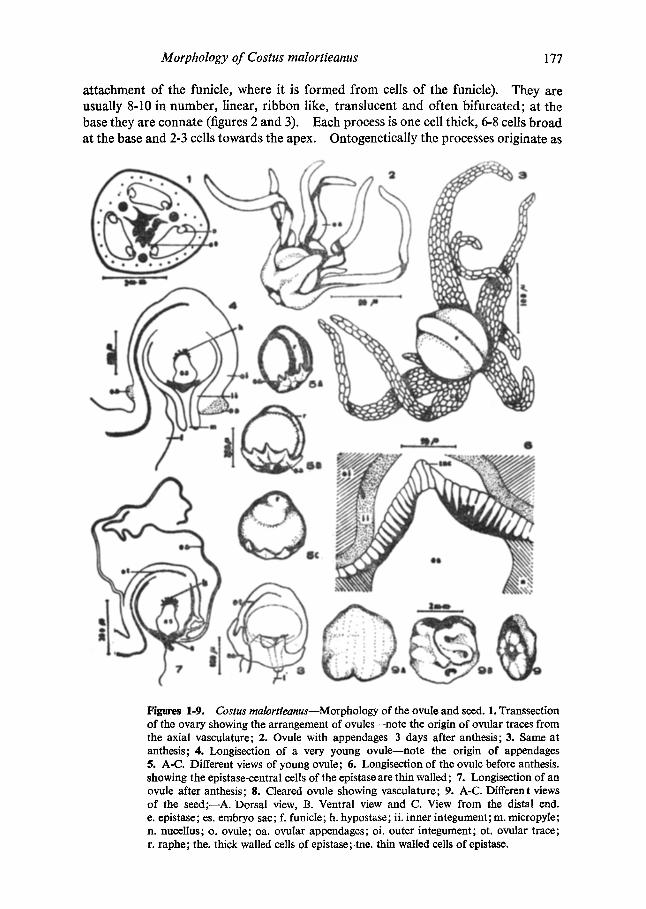
The ovary of Costus malortieanus is three chambered, with each chamber containing few ovules in two longitudinal rows on the septum (figure 1), slightly away from the central axis of the ovary. Placentation in Costus (and other Zingiberaceae having 3- chambered ovary) is commonly reported as axile (Lawrence 1951; Hooker 1890; Tomlinson 1956; Panchaksharappa 1962, 1963, 1967). But in the present study no instance of strictly axile placentation could be observed (our unpublished observations on several other South Indian taxa reveal that the ovules are attached to the central axis in Curcuma, Hedychium and Zingiber species, but in Alpinia spp. the ovules are attached to the septum and distinctly away from the central axis and in A. sandarae the ovules are attached near the outer edge of the septum close to the ovary wall). The central axis is progressively swollen towards the distal end so that the ovary chambers are constricted upwards and the ovules are restricted to the basal, more spacious part of the chambers. The ovules are bitegmic and typically anatropous with the raphe forming a longitudinal lateral rib extending upto the chalazal end of the ovule (figures 3, 4 and 5A-C). They are ovoid and are oriented obliquely on the placentum with the raphe on the axial side so that the micropyle is directed downwards and away from the axis (facing the outer wall of the ovary). The outer integument is four cells thick and shorter while the inner is two cells thick and protrudes out from the micropyle. The outer integument, towards the micropylar region is thicker and often five or six cells thick towards the anterior end. Both integuments are free.
There is a well developed epistase consisting of a single layer of vertically elongated cells, the inner tangential walls of which are lignified and thickened (figures 4 and 6). In the embryo sac stage of the ovule the epistase has a few central thin wailed cells opposite the micropyle so that the thickened region appears annular (figure 6). At a later stage (probably after fertilization) the central cells also become lignified and by then the epistase becomes two or three layered by the cells undergoing periclinal divisions. There is also, a well developed hypostase (figures 4 and 7) formed by the lignification of the cells at the chalazal end of the embryo sac.
The vascular connection to the ovule (figures 7 and 8) is a slender vascular strand consisting of 1-3 rows of tracheids and arises from the peripheral lateral bundle of the central axis. It is given off, a little below the level of the funicle. In the post fertiliz- ation stages there is a depression in the testa, opposite to where the vascular trace terminates (figure 9b); this depression persists in the mature seed.
An interesting feature of the ovule of C. malortieanus is the presence of characteri- stic lacerate ovular appendages which surround the ovule. The appendages arise from an annular ridge encircling the ovule at the micropylar end (figures 5A-C), formed by proliferation of cells of the outer integument (except at the region of
Morphology of Costus malortieanus 177
a t t achment o f the funicle, where i t is f o rmed f rom cells o f the funicle). They are usual ly 8-10 in number , l inear, r i bbon like, t rans lucent and of ten b i furca ted ; a t the base they are conna te (figures 2 and 3). Each process is one cell th ick, 6-8 cells b r o a d at the base and 2-3 cells t owards the apex. Ontogenet ica l ly the processes or iginate as
2 3
Figures 1-9. Costus malortieanus--Morphology of the ovule and seed. 1. Transsection of the ovary showing the arrangement of ovules--note the origin of ovular traces from the axial vasculature; 2. Ovule with appendages 3 days after anthesis; 3. Same at anthesis; 4. Longisection of a very young ovule--note the origin of appendages 5. A-C. Different views of young ovule; 6. Longisection of the ovule before anthesis. showing the epistase-central cells of the epistase are thin walled; 7. Longisection of an ovule after anthesis; 8. Cleared ovule showing vasculature; 9. A-C. Different views of the seed;--A. Dorsal view, B. Ventral view and C. View from the distal end. e. epistase; es. embryo sac; L funicle; h. hypostase; ii. inner integument; m. micropyle; n. nucellus; o. ovule; oa. ovular appendages; oi. outer integument; ot. ovular trace; r. raphe; the. thick walled cells of epistase; tne. thin walled cells of epistase.

178 Jose K Mangaly and K Sworrupanandan
proliferations partially from the funicular tissue and partially from the integument tissue. The proliferations appear at the ovular base (figure 4) even before the flower opens and grows rapidly to become longer than the ovule at the time of anthesis (figure 3). They continue to grow and as the seed matures, become considerably elongated and irregularly entwine the seed.
Seed setting is rare in C. malortieanus. The seed is ovoid, more or less dorsivent- rally compressed and ca. 3.00 mm across. It is dull brown in colour with an almost angular outline and a dorsal notch at the distal end. The dorsal surface (figure 9A), is smooth and fiat whereas the ventral surface (figure 9B) has wrinkled seed-coat. There is a depression at the point of attachment of the funicle which is slightly towards the ventral surface. In a polar view (figure 9C), from the distal end, the seed is elliptic in outline with an orbicular depression which is apparently formed by the overarching of the collar from the seed coat. In a polar view from the proximal end, the seed has a more or less rectangular outline with the depression of attachment at the centre.
4. Discussion
Structural variations in the angiosperm ovule are considered to be of phylogenetic significance by many (Maheshwari 1950, Van Tieghem 1901, Philipson 1974). A major deviation in the general structure of the ovule is the presence of ovular appendages in several taxa. Similar appendage-like arils have been reported in a number of families of dicots, not necessarily related and in Scitamineae (Arillatae of Engler and Prantle 1892), of monocotyledons, and their phylogenetic significance seems rather doubtful (Mauritson 1936), unless what are termed arils generally differ morpho- logically or ontogenetically. Proliferations developed from the funicle of the ovule are termed arils whereas those developed from the tissue of the integument at the micropylar end are termed as caruncles. In both cases the proliferations are developed after fertilization of the ovule.
All Zingiberaceae are described as arillate, signifying that the ovular outgrowths in the family are developed from the funicular tissue. However, the ovular outgrowths of C. malortieanus differ from arils as commonly understood, in developing very early during the ontogeny of the ovule (being often as long or even longer than the ovule at the time of anthesis), and in originating from the tissue of the funicle as well as the integument. The taxonomic significance of these proliferations are yet to be ascer- tained. However, the presence of such proliferations gains significance since some taxonomists (Nakai 1941 ; Tomlinson 1956, 1962), separate Costus from Zingiberaceae to constitute a separate family Costaceae. In the related species, C. speciosus, though the seed is 'axillate' no trace of such proliferations are formed in the pre-fertilization stages (Panchaksharappa 1962a).
The nature and significance of ovular appendages are to be considered, in the light of the nature of the ovule itself. In many cases, the ontogeny of these structures can be directly governed by the shape and structure of the ovule itself. In the case of an anatropous ovule, the outer integument is not developed on the side towards funieulus p a s the curvature of the ovule increases before the outer integument is complete (Coulter and Chamberlain 1965). Thus, in Zingiberaceae in which the ovules are anatropous the ovular appendages are formed mostly by the proliferation and folding

Morphology o f Costus malortieanus 179
back of the outer integument, but on the funicular side, it develops from the funiele itself, as the outer integument is absent there and its place is occupied by the funicle, as seen in C. malortieanus. On the other hand, in the orthotropous ovules occasional- ly met with in Hitchenia eaulina (Panchaksharappa 1966), the outer integument is in tact all around and proliferations of the funicle is seen all around, below the outer integument. This indicates that the dual origin of the ovular appendages as seen in C. malortieanus in particular and members of Zingiberaceae in general, results more by the anatropous nature of the ovule and consequent fusion of the funicle with the outer integument. However, the characteristic origin of the ovular appendages (viz. partly from the funicular tissue and partly from the integument) may be of phylogenetic significance. The significance of the appearance of these appendages before anthesis and fertilization in C. malortieanus can be assessed only when more data are available on the ovular appendages in tile family.
References
Coulter J M and Chamberlain C J 1965 Morphology of Angiosperms (Morphology o f Spermatophytes, Part II) (Central Book Depot, Allahabad) p. 348
Davis G L 1966 Systematic Embryology of Angiosperms (London: John Wiley and Sons Inc.) p. 528 Engler A and Prantl H 1892 Die Naturlichen Pflanzenfamilien (Leipzig) Hooker J D 1890 Flora of British India, Part XVII (L Reeve and Co., Ltd. Brook, Ashford, Kent) Lawrence G H M 1951 Taxonomy of Vascular Plants (New York: The Macmillan Co.) p. 823 Maheshwari P 1950 An Introduction to the Embryology of Angiosperms (New York: McGraw Hill
Book Co. Inc.) p. 453 Mauritzon J 1936e Smenbau und Embryologie Einiger Scitamineen. Acta Univ. London 31 1-31 Nakai T 1941 Notulae and Plantas Asiae Orientalis (XVI) J. Jpn. Bot. I7 189-210 Panchaksharappa M G 1959 Embryological Studies in some members of Zingiberaceae. Ph.D.
Thesis, Delhi University Panchaksharappa M G 1961 Embryological Studies in the family Zingiberaceae: Curcuma aromatica
Salisb; J. Karnatak Univ. 6 49-56 Panchaksharappa M G 1962a Embryological Studies in the Family Zingiberaceae I. Costus speciosus
Smith; Phytomorphology 12 418-430 Panchaksharappa M G 1962b Embryological Studies in Some members of Zingiberaceae; Plant
Embryology. A Symposium, (CSIR India) 224-238 Panchaksharappa M G 1962c Taxonomic Evaluation of Zingiberaceae; Bull. Bot. Surv. India 4
129-135 Panchaksharappa M G 1966 Embryological Studies in Some Members of Zingiberaceae II. Elettaria
cardamomum, Hitchenia caulina and Zingiber macrostachyum ; Phytomorphology 16 412-417 Panchaksharappa 1970 Zingiberaceae--Comparative embryology of Angiosperms; Bull. Indian Nat.
Sci. Acad. 41 380-385 Philipson W R 1974 Ovular morphology and the major classification of the Dicotyledons; J. Linn
Soc. (Bot.). 68 89-108 Sachar R C and Arora U 1963 Some Embryological aspects ofAmomum dealbratum and H.edychium
acuminatum; Bot. Gaz. 124 356-360 Tomlinson P B 1956 Studies in the Systematic Anatomy of Zingiberaceae; J. Linn. Soc. (Bot). 55
547-592 Tomlinson P B 1962 Phylogeny of the Scitamineae-Morphological and Anatomical considerations;
Evolution 16 192-213 Van Tieghem P 1901 L'oeuf des plantes considere Comme base de leur Classification; Ann. Sci. Nat.
(Bot.) Set. 8 114 213-390