somatostatin and brain-derived neurotrophic factor mrna expression in the primate brain: decreased...
TRANSCRIPT

Ž .Brain Research 749 1997 283–289
Research report
Somatostatin and brain-derived neurotrophic factor mRNA expression in theprimate brain: decreased levels of mRNAs during aging
Motoharu Hayashi a,) , Akiko Yamashita b, Keiko Shimizu a
a Department of Cellular and Molecular biology, Primate Research Institute, Kyoto UniÕersity, Inuyama, Aichi 484, Japanb Department of Anatomy, Nihon UniÕersity School of Medicine, Tokyo , Japan
Accepted 22 October 1996
Abstract
Ž . Ž .The expression of the genes for somatostatin SRIF and brain-derived neurotrophic factor BDNF was investigated in the centralŽ . Ž .nervous system CNS of the macaque monkey Macaca fuscata fuscata . Using Northern blot analysis, one SRIF mRNA transcript, 0.65
Žkb, and two BDNF mRNA transcripts, 1.6 and 4.0 kb in length, were detected in the monkey brain tissues. During the aging process 2. Ž .years, 10 years, and )30 years , the ratio of SRIF mRNArglyceraldehyde-3 phosphate dehydrogenase G3PDH mRNA significantly
Ž .decreased 60–70% in the hippocampus and in several cerebral subdivisions such as frontal cortex, temporal cortex, motor cortex,somatosensory cortex and visual cortex. BDNF mRNA was expressed in the various cerebral subdivisions and in the hippocampus.
ŽDuring the aging process, the gene expression of BDNF declined 20–50% for the 4.0 kb transcript, and 40–70% for the 1.6 kb.transcript in the various cerebral subdivisions. In the hippocampus, the level of the 1.6 kb mRNA at )30 years old declined to 60% of
the level at 2 years old, while the 4.0 kb mRNA did not change significantly during the aging process. Recent studies have shown thatŽ Ž .BDNF enhances the expression of SRIF mRNA in the rodent cerebral cortex Nawa, H. et al., J. Neurochem., 60 1993 772–775; Nawa,
Ž . .H. et al., J. Neurosci., 14 1994 3751–3765 . These studies and our present results suggest that the decrease in gene expression for aneurotrophic molecule, such as BDNF, might cause the levels of SRIF mRNA to decline in the primate brain during the aging process.
Keywords: Somatostatin; Brain-derived neurotrophic factor; Gene expression; Primate; Central nervous system; Aging
1. Introduction
Ž .Somatostatin SRIF is one of the neuroactive peptidesŽ . w xin the mammalian central nervous system CNS 4,14,42 .
The most well known role of this peptide in the CNS is toregulate release of growth hormone from the pituitarygland. Moreover, SRIF has also been detected in variousbrain tissues such as the cerebral cortex, hippocampus, andamygdala. The physiological function of SRIF in the mam-malian CNS is still unclear. SRIF has been reported to
w xboth excite and inhibit neuronal firing rates 11,41 . Man-w xcillas et al. 30 reported that SRIF enhances
acetylcholine-induced excitations in hippocampal and cor-tical neurons in the rat. Together, these findings suggestthat the peptide may have a role as a neurotransmitter or aneuromodulator in the CNS. Furthermore, treatment withcysteamine, which depletes SRIF in the mammalian CNS,
w ximpairs memory in rats 3,16,20 , suggesting that the pep-
) Ž .Corresponding author. Fax: q81 568 630085; e-mail:[email protected]
tide might participate in higher brain functions such ascognition and memory.
In previous studies, we observed that the gene expres-sion of SRIF, the content of SRIF, and the numbers ofSRIF-containing cells in the monkey cerebral and cerebel-lar cortices were transiently enhanced during the embry-
w xonic period 21–23,25,50,51 . The developmental stage atwhich these phenomena were observed coincided with theperiod of fiber elongation in the cerebral and cerebellarcortices. These results indicate that the peptide may beinvolved in the formation of the connection of fibers in thedeveloping monkey CNS. In fact, SRIF has been reportedto induce neurite outgrowth in the molluscan neuronsw x w x5,19 , PC12 cells 15 , and rat cerebellar granule cellsw x47 .
In brain tissue of patients with Alzheimer’s disease, thew xmarked decrease in gene expression of SRIF 13,49 and in
w xthe levels of SRIF 12,44 , have been reported. Moreover,SRIF immunoreactivity has been detected in senile plaques
w xof patients with Alzheimer’s disease 1,34,43 . Degenerat-ing SRIF containing neurons have also been observed in
0006-8993r97r$17.00 Copyright q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 96 01317-0

( )M. Hayashi et al.rBrain Research 749 1997 283–289284
w xthe brain of the aged macaque monkey 45 . Furthermore,w xthe gene expression for SRIF 17 and the numbers of
w xSRIF-containing neurons 33 are reduced in the cerebralcortex of aged rat.
The molecular mechanism of the gene expression ofSRIF in the mammalian CNS has not yet been clarified.Nawa et al. recently reported, however, that one of theneurotrophic factors, brain-derived neurotrophic factorŽ .BDNF , enhanced the expression of SRIF, neuropeptide Y
Ž .and cholecystokinin CCK in rat cerebral neurons both inw x w xvivo 37 and in vitro 36 . Thus, BDNF seems to be one of
the factors that regulate the expression of SRIF genes inthe mammalian CNS.
Ž .Neurotrophic factors neurotrophins are known to beinvolved in the survival and differentiation of selectiveneurons during embryonic stages, and in the maintenance
w xof those neurons in adulthood 29,48 . A recent studyindicates that neurotrophins are also involved in neuronal
w xplasticity 48 . For example, the expression of BDNFmRNA in the CNS was reported to be regulated by
w x w xphysiological stimuli such as light 7 and exercise 38 .Furthermore the gene expression of BDNF has been de-scribed to decrease in the hippocampus of patients with
w xAlzheimer’s disease 35,40 , whose neuronal plasticityseems to be severely affected. In the present study, weinvestigated whether the gene expression of SRIF andBDNF might change during the aging process of theprimate CNS. Preliminary results have been reported else-
w xwhere 24 .
2. Materials and methods
2.1. Preparation of samples
Ž .Japanese monkeys Macaca fuscata fuscata , four at 2Žyears, three at 10 years, and three at )30 years 30 years
.male, 30 years female, 33 years female were pretreatedŽ .with ketamine hydrochloride 10 mgrkg, i.m. , deeply
Žanesthetized with pentobarbital sodium Nembutal; 25.mgrkg, i.p. , and killed by bloodletting from the carotid
artery. Dissection of the various brain regions was per-w xformed on crushed ice, as described previously 23 . The
dissected brain tissues were stored at y808C until use. Thecare and use of the living animals in the present studywere approved by the Animal Research Committee of thePrimate Research Institute, Kyoto University.
2.2. Nucleic acid probes for SRIF, G3PDH, and BDNF
Ž . ŽThe plasmid pMC4G8 -containing monkey Macaca.fascicularis SRIF cDNA, which was kindly supplied by
Dr. J.G. Sutcliffe, was propagated in Escherichia coliŽ .HB101 . The 0.65 kb insertion was cut from the plasmidby digestion with EcoRI and HindIII. The probe was
w 32 x Žlabeled with a- P dCTP 3000 Cirmmol; 1 Cis37
.GBq; ICN, Costa Mesa, USA using a Random PrimedŽDNA Labeling Kit Boehringer Mannheim, Mannheim,
.Germany . Human glyceraldehyde-3-dehydrogenaseŽ .G3PDH cDNA probe was obtained from Clontech labo-
Ž .ratories Palo Alto, USA .The mouse BDNF plasmid, which was kindly supplied
Ž .by Dr. H. Thoenen, was propagated in E. coli HB101 .The plasmid was linearized with HincII and a single-stranded RNA probe was transcribed using a riboprobe
Ž .transcription system Promega, Madison, USA with T7w 32 xpolymerase and a- P UTP.
2.3. Preparation of RNA
Total RNA was purified by the method of Chomczynskiw xand Sacchi 9 with slight modification. The brain tissue
Ž .0.5–1.0 g was homogenized in 10 vols. of 4 M guani-dinium thiocyanate, 0.1 M 2-mercaptoethanol, 25 mM
Ž .sodium citrate pH 7.0 , and 0.5% sodium N-lauroyl sar-cosinate. To the homogenate, 0.1 vol. of 2 M sodiumacetate buffer, pH 4.0, 1 vol. of phenol saturated withwater, and 0.2 vol. of a mixture of chloroform and isoamy-
Ž .lalcohol 49:1, vrv was added. The solution was incu-bated on ice for 15 min and centrifuged at 10 000=g for30 min. The supernatant phase was mixed with 1 vol. ofisopropyl alcohol, and then incubated at y208C for 1 h.The precipitate was dissolved in the homogenization solu-tion and two volumes of ethanol were added. The mixturewas incubated at y208C overnight. After centrifugation,the resultant pellet was resuspended in water and stored aty808C until use.
2.4. Northern blot analysis of SRIF and G3PDH
Total RNA was denatured in 50% formamide, 6.5%formaldehyde at 608C for 15 min and subjected to elec-
Ž .trophoresis on a horizontal agarose gel 1.0% that con-Ž . Ž .tained formaldehyde 6.5% . Total RNA 15 mg was
loaded per lane. RNA was transferred by capillary blottingŽonto nylon membrane Hybond-N, Amersham, Little Chal-
.font, UK and prehybridized at 428C for 5 h in 5=standardŽ .saline citrate SSC , 50% formamide, and 50 mM sodium
phosphate buffer, pH 7.0, containing 0.1% Ficoll, 0.1%BSA, 0.1% polyvinylpyrrolidone, and sheared denatured
Ž .salmon sperm DNA 200 mgrml . Hybridization was car-ried out overnight at 428C in prehybridization solutioncontaining 5=106 cpmrml of SRIF cDNA. The mem-brane was washed four times for 5 min at room tempera-ture in 2=SSC and 0.1% SDS and twice for 30 min at558C in 0.1=SSC and 0.1% SDS. The membrane was
Ž .then exposed to X-ray film Hyperfilm MP; Amersham aty808C for 24–48 h. The membrane was boiled in 0.1=SSC and 0.4% SDS for 30 min and rehybridized with theG3PDH cDNA probe. The autoradiographic signals forSRIF mRNA and G3PDH mRNA were quantified with a
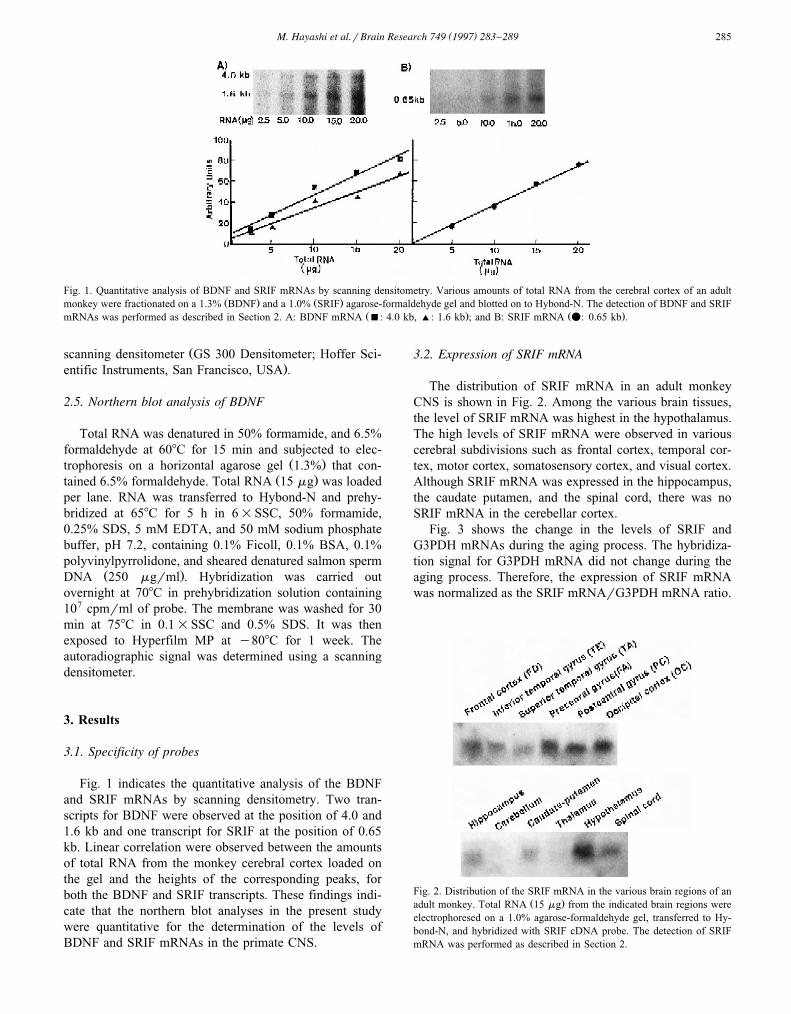
( )M. Hayashi et al.rBrain Research 749 1997 283–289 285
Fig. 1. Quantitative analysis of BDNF and SRIF mRNAs by scanning densitometry. Various amounts of total RNA from the cerebral cortex of an adultŽ . Ž .monkey were fractionated on a 1.3% BDNF and a 1.0% SRIF agarose-formaldehyde gel and blotted on to Hybond-N. The detection of BDNF and SRIF
Ž . Ž .mRNAs was performed as described in Section 2. A: BDNF mRNA B: 4.0 kb, ': 1.6 kb ; and B: SRIF mRNA v: 0.65 kb .
Žscanning densitometer GS 300 Densitometer; Hoffer Sci-.entific Instruments, San Francisco, USA .
2.5. Northern blot analysis of BDNF
Total RNA was denatured in 50% formamide, and 6.5%formaldehyde at 608C for 15 min and subjected to elec-
Ž .trophoresis on a horizontal agarose gel 1.3% that con-Ž .tained 6.5% formaldehyde. Total RNA 15 mg was loaded
per lane. RNA was transferred to Hybond-N and prehy-bridized at 658C for 5 h in 6=SSC, 50% formamide,0.25% SDS, 5 mM EDTA, and 50 mM sodium phosphatebuffer, pH 7.2, containing 0.1% Ficoll, 0.1% BSA, 0.1%polyvinylpyrrolidone, and sheared denatured salmon sperm
Ž .DNA 250 mgrml . Hybridization was carried outovernight at 708C in prehybridization solution containing107 cpmrml of probe. The membrane was washed for 30min at 758C in 0.1=SSC and 0.5% SDS. It was thenexposed to Hyperfilm MP at y808C for 1 week. Theautoradiographic signal was determined using a scanningdensitometer.
3. Results
3.1. Specificity of probes
Fig. 1 indicates the quantitative analysis of the BDNFand SRIF mRNAs by scanning densitometry. Two tran-scripts for BDNF were observed at the position of 4.0 and1.6 kb and one transcript for SRIF at the position of 0.65kb. Linear correlation were observed between the amountsof total RNA from the monkey cerebral cortex loaded onthe gel and the heights of the corresponding peaks, forboth the BDNF and SRIF transcripts. These findings indi-cate that the northern blot analyses in the present studywere quantitative for the determination of the levels ofBDNF and SRIF mRNAs in the primate CNS.
3.2. Expression of SRIF mRNA
The distribution of SRIF mRNA in an adult monkeyCNS is shown in Fig. 2. Among the various brain tissues,the level of SRIF mRNA was highest in the hypothalamus.The high levels of SRIF mRNA were observed in variouscerebral subdivisions such as frontal cortex, temporal cor-tex, motor cortex, somatosensory cortex, and visual cortex.Although SRIF mRNA was expressed in the hippocampus,the caudate putamen, and the spinal cord, there was noSRIF mRNA in the cerebellar cortex.
Fig. 3 shows the change in the levels of SRIF andG3PDH mRNAs during the aging process. The hybridiza-tion signal for G3PDH mRNA did not change during theaging process. Therefore, the expression of SRIF mRNAwas normalized as the SRIF mRNArG3PDH mRNA ratio.
Fig. 2. Distribution of the SRIF mRNA in the various brain regions of anŽ .adult monkey. Total RNA 15 mg from the indicated brain regions were
electrophoresed on a 1.0% agarose-formaldehyde gel, transferred to Hy-bond-N, and hybridized with SRIF cDNA probe. The detection of SRIFmRNA was performed as described in Section 2.
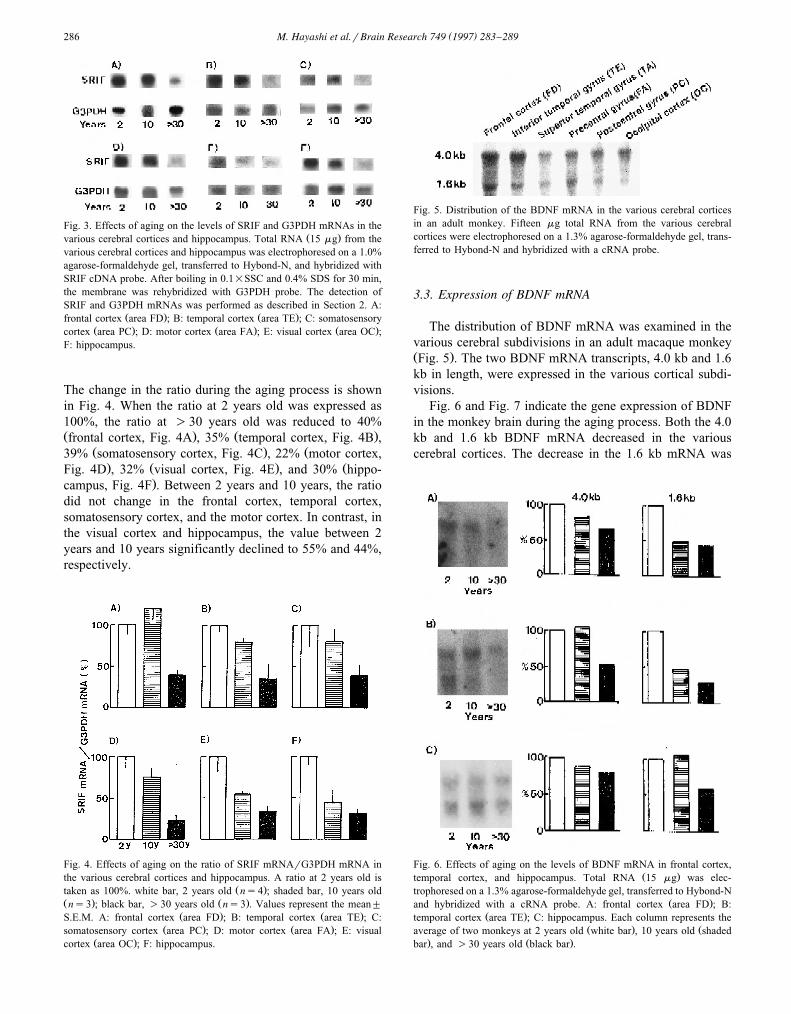
( )M. Hayashi et al.rBrain Research 749 1997 283–289286
Fig. 3. Effects of aging on the levels of SRIF and G3PDH mRNAs in theŽ .various cerebral cortices and hippocampus. Total RNA 15 mg from the
various cerebral cortices and hippocampus was electrophoresed on a 1.0%agarose-formaldehyde gel, transferred to Hybond-N, and hybridized withSRIF cDNA probe. After boiling in 0.1=SSC and 0.4% SDS for 30 min,the membrane was rehybridized with G3PDH probe. The detection ofSRIF and G3PDH mRNAs was performed as described in Section 2. A:
Ž . Ž .frontal cortex area FD ; B: temporal cortex area TE ; C: somatosensoryŽ . Ž . Ž .cortex area PC ; D: motor cortex area FA ; E: visual cortex area OC ;
F: hippocampus.
The change in the ratio during the aging process is shownin Fig. 4. When the ratio at 2 years old was expressed as100%, the ratio at )30 years old was reduced to 40%Ž . Ž .frontal cortex, Fig. 4A , 35% temporal cortex, Fig. 4B ,
Ž . Ž39% somatosensory cortex, Fig. 4C , 22% motor cortex,. Ž . ŽFig. 4D , 32% visual cortex, Fig. 4E , and 30% hippo-
.campus, Fig. 4F . Between 2 years and 10 years, the ratiodid not change in the frontal cortex, temporal cortex,somatosensory cortex, and the motor cortex. In contrast, inthe visual cortex and hippocampus, the value between 2years and 10 years significantly declined to 55% and 44%,respectively.
Fig. 4. Effects of aging on the ratio of SRIF mRNArG3PDH mRNA inthe various cerebral cortices and hippocampus. A ratio at 2 years old is
Ž .taken as 100%. white bar, 2 years old ns4 ; shaded bar, 10 years oldŽ . Ž .ns3 ; black bar, )30 years old ns3 . Values represent the mean"
Ž . Ž .S.E.M. A: frontal cortex area FD ; B: temporal cortex area TE ; C:Ž . Ž .somatosensory cortex area PC ; D: motor cortex area FA ; E: visual
Ž .cortex area OC ; F: hippocampus.
Fig. 5. Distribution of the BDNF mRNA in the various cerebral corticesin an adult monkey. Fifteen mg total RNA from the various cerebralcortices were electrophoresed on a 1.3% agarose-formaldehyde gel, trans-ferred to Hybond-N and hybridized with a cRNA probe.
3.3. Expression of BDNF mRNA
The distribution of BDNF mRNA was examined in thevarious cerebral subdivisions in an adult macaque monkeyŽ .Fig. 5 . The two BDNF mRNA transcripts, 4.0 kb and 1.6kb in length, were expressed in the various cortical subdi-visions.
Fig. 6 and Fig. 7 indicate the gene expression of BDNFin the monkey brain during the aging process. Both the 4.0kb and 1.6 kb BDNF mRNA decreased in the variouscerebral cortices. The decrease in the 1.6 kb mRNA was
Fig. 6. Effects of aging on the levels of BDNF mRNA in frontal cortex,Ž .temporal cortex, and hippocampus. Total RNA 15 mg was elec-
trophoresed on a 1.3% agarose-formaldehyde gel, transferred to Hybond-NŽ .and hybridized with a cRNA probe. A: frontal cortex area FD ; B:
Ž .temporal cortex area TE ; C: hippocampus. Each column represents theŽ . Žaverage of two monkeys at 2 years old white bar , 10 years old shaded
. Ž .bar , and )30 years old black bar .

( )M. Hayashi et al.rBrain Research 749 1997 283–289 287
Fig. 7. Effects of aging on the levels of BDNF mRNA in mortor cortexŽ .and somatosensory cortex. Total RNA 15 mg was electrophoresed on a
1.3% agarose-formaldehyde gel, transferred to Hybond-N and hybridizedŽ .with a cRNA probe. A: motor cortex area FA ; B: somatosensory cortex
Ž .area PC . Each column represents the average of two monkeys at 2 yearsŽ . Ž . Ž .old white bar , 10 years old shaded bar and )30 years old black bar .
larger than that of the 4.0 kb mRNA. The expression of the1.6 kb mRNA at )30 years old was 29–40% of the valueat 2 years old. In contrast, the expression of 4.0 kb mRNAat )30 years old was 53–65% of the level at 2 years old.On the other hand, in the hippocampus the 1.6 kb mRNAdecreased to 60% of the value at 2 years old, but the 4.0kb mRNA did not decline during the aging process.
4. Discussion
4.1. Expression of SRIF mRNA in the monkey CNS
In the present study, we observed heterogeneous ex-pression of the gene for SRIF in the primate CNS. Highexpression of the gene was found in the hypothalamus andthe various cerebral cortices. A relatively high expressionwas also observed in the hippocampus, the caudate-puta-men, and in the spinal cord. In contrast, no SRIF mRNAwas found in the cerebellum. These results are fairlyconsistent with the levels of SRIF immunoreactive peptide
w xin the CNS of the human 10 and macaque monkeyw x22,23 . High levels of SRIF mRNA were expressed in theprimate cerebral cortices. In a previous study, we foundthat SRIF is one of the most abundant neuropeptides in theprimate cerebral cortex, particularly in the association
w xcortices 21 . Thus, these findings suggest that SRIF is anevolutionarily important neuropeptide for primate corticalfunctions.
With respect to the function of the peptide in themammalian CNS, SRIF is reported to enhance the acetyl-choline-induced excitation in rat hippocampal and cortical
w x Ž .neurons 30 . Enhancement of long-term potentiation LTP
by SRIF has also been reported in the mossy fiber-CA3w xsystem in the guinea-pig hippocampal slice 31 . Further-
more, cysteamine, a depletor of SRIF, impairs memory inw xthe rat 3,16,20 . Thus, SRIF is likely to participate in the
neuronal transmission that may be involved in the higherbrain functions such as learning and memory.
4.2. Expression of SRIF mRNA during aging
In the present study, we found that the expression of theSRIF gene was reduced in various cerebral cortices and thehippocampus in the aged monkey. Recently, a similarreduction of SRIF gene expression during the aging pro-
w xcess was reported in the cerebral cortex of the rat 17 .Moreover, in the brain tissues of patients with Alzheimer’sdisease, marked decreases in both levels of SRIF proteinw x w x12,44 and SRIF mRNA 13,49 have been demonstrated.Furthermore, among the various neuropeptides, SRIF de-creased more than other peptides in the CNS of these
w xpatients 2 . The reduction of the density of SRIF mRNA-containing neurons has been reported in various cerebralcortices, the parahippocampal cortex, and the hippocampus
w xof patients with Alzheimer’s disease 13 . Therefore, SRIFis apparently one of the neuropeptides which is vulnerableduring the normal aging process and dementia.
The decrease in the level of SRIF is reported to becorrelated to the loss of cognitive ability in patients with
w xAlzheimer’s disease 46 . This finding is consistent withthe assumption that the peptide may be involved in higherbrain functions.
4.3. Expression of BDNF mRNA in the monkey CNS
BDNF, one of the neurotrophins, is thought to be asurvival factor for various neurons, such as cholinergic
w xneurons in the basal forebrain 27 , the mesencephalicw xdopaminergic neurons 26,27 , and the cerebral cortical
w xneurons 18 . Furthermore, BDNF has been shown toparticipate in the formation of ocular dominance columns
w x w xin the cat visual cortex 6 . Recently, McAllister et al. 32reported that BDNF enhanced dendritic growth in thedeveloping visual cortex of the rat. These findings indicatethat BDNF may participate in cortical development duringthe embryonic stages.
In the present study, we observed that the gene forBDNF was expressed in the various cortical subdivisionsand in the hippocampus, suggesting that BDNF may be animportant factor for the maintenance of higher brain func-tions at the adult stage. The synthesis of mRNA of theneurotrophins, in particular BDNF and NGF, was de-scribed to be regulated by neuronal activity in adulthoodw x8,52 . Furthermore, the expression of the genes for BDNF
w xand NGF is increased by LTP in the hippocampus 7,39 .More recently, hippocampal LTP was shown to be im-paired in mice with a deletion in the coding sequence of
w xthe BDNF gene 28 . Together, these findings suggest that

( )M. Hayashi et al.rBrain Research 749 1997 283–289288
BDNF may be essential for neuronal plasticity at adult-hood.
4.4. Similarity of the expression of BDNF and SRIF mR-NAs during the aging process
In the present study, we found that SRIF and BDNFmRNAs decreased similarly during the aging process inthe monkey cerebral cortices and in the hippocampus. Thedecrease in both SRIF and BDNF mRNAs has also beenreported in the hippocampus of patients with Alzheimer’s
w xdisease 13,35,40 . A recent study showed that BDNFincreases the expression of the gene for SRIF in the rat
w x w xcerebral cortex in vivo 37 and in vitro 36 , suggestingthat the expression of SRIF mRNA might be regulated byneurotrophins such as BDNF. Thus, the decreases in thelevels of SRIF mRNA in the aged monkey brain might bedue to the decrease in the gene expression of BDNF.
Furthermore, various stimuli such as retinal activationw x w x7 and physical exercise 38 , increased the expression ofBDNF mRNA in the rat cerebral cortices. Therefore, thedecrease in the levels of SRIF mRNA might be due to thereduction of BDNF mRNA expression, which may becaused by a low level of neuronal stimulation in the agedmonkey CNS. The changes in the numbers of the cellswhich express BDNF mRNA and BDNF protein duringthe aging process remain to be clarified in future experi-ments.
Acknowledgements
We wish to express our thanks to Drs. H. Thoenen andD. Lindholm for their supply of mouse BDNF plasmid,and to Dr. J.G. Sutcliffe for his supply of monkey SRIFplasmid. This work was supported by Grants no. 05261205
Ž .and no. 08838012 M.H. from the Ministry of Education,Science and Culture, Japan.
References
w x1 Armstrong, D.M., Benzing, W.C., Evans, J., Terry, R.D., Shields, D.and Hansen, L.A., Substance P and somatostatin coexist withinneuritic plaques: implications for the pathogenesis of Alzheimer’s
Ž .disease, Neuroscience, 31 1989 663–671.w x2 Auchus, A.P., Green, R.C. and Nemeroff, C.B., Cortical and subcor-
tical neuropeptides in Alzheimer’s disease, Neurobiol. Aging, 15Ž .1994 589–595.
w x3 Bakhit, C. and Swerdlow, N., Behavioral changes following centralŽ .injection of cysteamine in rats, Brain Res., 365 1986 159–163.
w x4 Beal, M.F., Uhl, G., Mazurek, M.F., Kowall N. and Martin, J.B.,Somatostatin: alterations in the central nervous system in neurologi-
Ž .cal diseases. In J.B. Martin and J.D. Barchas Eds. , Neuropeptidesin Neurologic and Psychiatric Disease, Raven Press, New York,1986, pp. 215–257.
w x5 Bulloch, A.G.M., Somatostatin enhances neurite outgrowth and elec-trical coupling of regenerating neurons in Helisoma, Brain Res., 412Ž .1987 6–17.
w x6 Cabelli, R.J., Hohn, A. and Shatz, C.J., Inhibition of ocular domi-nance column formation by infusion of NT4r5 or BDNF, Science,
Ž .267 1995 1662–1666.w x7 Castren, E., Pitkanen, M., Sirvio, J., Parsadanian, A., Lindholm, D.,´ ¨ ¨
Thoenen, H. and Riekkinen, P.J., The induction of LTP increasesBDNF and NGF mRNA but decreases NT-3 mRNA in the dentate
Ž .gyrus, NeuroReport, 4 1993 895–898.w x8 Castren, E., Zafra, F., Thoenen, H. and Lindholm, D., Light regu-´
lates expression of brain-derived neurotrophic factor mRNA in ratŽ .visual cortex, Proc. Natl. Acad. Sci. USA, 89 1992 9444–9448.
w x9 Chomczynski, P. and Sacchi, N., Single-step method of RNA isola-tion by acid guanidinium thiocyanate-phenol-chloroform extraction,
Ž .Anal. Biochem., 162 1987 156–159.w x10 Cooper, P.E., Fernstrom, M.H., Rorstad, O.P., Leeman, S.E. and
Martin, J.B., The regional distribution of somatostatin, substance PŽ .and neurotensin in human brain, Brain Res., 218 1981 219–232.
w x11 Dodd, J. and Kelly, J.S., Is somatostatin an excitatory transmitter inŽ .the hippocampus?, Nature, 273 1978 674–675.
w x12 Davies, P., Katzman, R. and Terry, R.D., Reduced somatostatin-likeimmunoreactivity in cerebral cortex from cases of Alzheimer disease
Ž .and Alzheimer senile dementia, Nature, 288 1980 279–280.w x13 Dournaud, P., Cervera-Pierot, P., Hirsch, E., Javoy-Agid, F., Kor-
don, C.L., Agid, Y. and Epelbaum, J., Somatostatin messengerRNA-containing neurons in Alzheimer’s disease: an in situ hy-bridization study in hippocampus, parahippocampal cortex and
Ž .frontal cortex, Neuroscience, 61 1994 755–764.w x14 Epelbaum, J., Somatostatin in the central nervous system: physiol-
Ž .ogy and pathological modifications, Prog. Neurobiol., 27 198663–100.
w x15 Ferriero, D.M., Sheldon, R.A. and Messing, R.O., Somatostatinenhances nerve growth factor-induced neurite outgrowth in PC12
Ž .cells, DeÕ. Brain Res., 80 1994 13–18.w x16 Fitzgerald, L.W. and Dokla, C.P.J., Morris water task impairment
and hypoactivity following cysteamine-induced reductions of so-Ž .matostatin-like immunoreactivity, Brain Res., 505 1989 246–250.
w x17 Florio, T., Ventra, C., Postiglione, A. and Schettini, G., Age-relatedalterations of somatostatin gene expression in different rat brain
Ž .areas, Brain Res., 557 1991 64–68.w x18 Ghosh, A., Carnahan, J. and Greenberg, M.E., Requirement for
BDNF in activity-dependent survival of cortical neurons, Science,Ž .263 1994 1618–1623.
w x19 Grimm-Jørgensen, Y., Somatostatin and calcitonin stimulate neuriteŽ .regeneration of molluscan neurons in vitro, Brain Res., 403 1987
121–126.w x20 Haroutunian, V., Mantin, R., Campbell, G.A., Tsuboyama, G.K. and
Davis, K.L., Cysteamine-induced depletion of central somatostatin-like immunoreactivity: effects on behavior, learning, memory and
Ž .brain neurochemistry, Brain Res. 403 1987 234–242.w x21 Hayashi, M., Ontogeny of some neuropeptides in the primate brain,
Ž .Prog. Neurobiol., 38 1992 231–260.w x22 Hayashi, M., Ontogeny of glutamic acid decarboxylase, tyrosine
hydroxylase, choline acetyltransferase, somatostatin and substance PŽ .in monkey cerebellum, DeÕ. Brain Res., 32 1987 181–186.
w x23 Hayashi, M. and Oshima, K., Neuropeptides in cerebral cortex ofŽ .macaque monkey Macaca fuscata fuscata : regional distribution
Ž .and ontogeny, Brain Res., 364 1986 360–368.w x24 Hayashi, M. and Shimizu, K., BDNF and somatostatin gene expres-
sion in the primate brain: decreased levels of mRNA during aging,Ž .Soc. Neurosci. Abstr. 21 1995 2030.
w x25 Hayashi, M., Yamashita, A., Shimizu, K., Sogawa, K. and Fujii, Y.,Somatostatin gene expression in the developing monkey frontal and
Ž .cerebellar cortices, DeÕ. Brain Res. 57 1990 37–41.w x26 Hyman, C., Hofer, M., Brade, Y.-A., Juhasz, M., Yancopoulos,
G.D., Squinto, S.P. and Lindsay, R.M., BDNF is a neurotrophicfactor for dopaminergic neurons of the substantia nigra, Nature, 350Ž .1991 230–232.
w x27 Knusel, B., Winslow, J.W., Rosenthal, A., Burton, L.E., Seid, D.P.,¨

( )M. Hayashi et al.rBrain Research 749 1997 283–289 289
Nikolics, K. and Hefti, F., Promotion of central cholinergic anddopaminergic neuron differentiation by brain-derived neurotrophic
Ž .factor but not neurotrophin 3, Proc. Natl. Acad. Sci. USA, 88 1991961–965.
w x28 Korte, M., Carroll, P., Wolf, E., Brem, G., Thoenen, H. andBonhoeffer, T., Hippocampal long-term potentiation is impaired inmice lacking brain-derived neurotrophic factor, Proc. Natl. Acad.
Ž .Sci. USA, 92 1995 8856–8860.w x29 Lewin, G.R. and Barde, Y.-A., Physiology of the neurotrophins,
Ž .Annu. ReÕ. Neurosci., 19 1996 289–317.w x30 Mancillas, J.R., Siggins, G.R. and Bloom, F.E., Somatostatin selec-
tively enhances acetylcholine-induced excitations in rat hippocam-Ž .pus and cortex, Proc. Natl. Acad. Sci. USA, 83 1986 7518–7521.
w x31 Matsuoka, N., Kaneko, S. and Satoh, M., Somatostatin augmentslong-term potentiation of the mossy fiber-CA3 system in guinea-pig
Ž .hippocampal slices, Brain Res., 553 1991 188–194.w x32 McAllister, A.K., Lo, D.C. and Katz, L.C., Neurotrophins regulate
Ž .dendritic growth in developing visual cortex, Neuron, 15 1995791–803.
w x33 Miettinen, R., Sirvio, J., Riekkinen Sr., P., Laakso, M.P., Riekkinen,¨M. and Riekkinen Jr., P., Neocortical, hippocampal and septalparvalbumin- and somatostatin-containing neurons in young andaged rats: correlation with passive avoidance and water maze perfor-
Ž .mance, Neuroscience, 53 1993 367–378.w x34 Morrison, J.H., Rogers, J., Scherr, S., Benoit, R. and Bloom, F.E.,
Somatostatin immunoreactivity in neuritic plaques of Alzheimer’sŽ .patients, Nature, 314 1995 90–92.
w x35 Murray, K.D., Gall, C.M., Jones, E.G. and Isackson, P.J., Differen-tial regulation of brain-derived neurotrophic factor and type IIcalciumrcalmodulin-dependent protein kinase messenger RNA ex-
Ž .pression in Alzheimer’s disease, Neuroscience, 60 1994 37–48.w x36 Nawa, H., Bessho, Y., Carnahan, J., Nakanishi, S. and Mizuno, K.,
Regulation of neuropeptide expression in cultured cerebral corticalneurons by brain-derived neurotrophic factor, J. Neurochem., 60Ž .1993 772–775.
w x37 Nawa, H., Pelleymounter, M.A. and Carnahan, J., Intraventricularadministration of BDNF increases neuropeptide expression in new-
Ž .born rat brain, J. Neurosci., 14 1994 3751–3765.w x38 Neeper, S.A., Gomez-Pinilla, F., Choi, J. and Cotman, C., Exercise
Ž .and brain neurotrophins, Nature, 373 1995 109.w x39 Patterson, S.L., Grover, L.M., Schwartzkroin, P.A. and Bothwell,
M., Neurotrophin expression in rat hippocampal slices: a stimulus
paradigm inducing LTP in CA1 evokes increases in BDNF andŽ .NT-3 mRNAs, Neuron, 9 1992 1081–1088.
w x40 Phillips, H.S., Hains, J.M., Armanini, M., Laramee, G.R., Johnson,S.A. and Winslow, J.W., BDNF is decreased in the hippocampus of
Ž .individuals with Alzheimer’s disease, Neuron, 7 1991 695–702.w x41 Pittman, Q.J. and Siggins, G.R., Somatostatin hyperpolarizes hip-
Ž .pocampal pyramidal cells in vitro, Brain Res., 221 1981 402–408.w x Ž .42 Reichlin, S., Somatostatin, N. Engl. J. Med., 309 1983 1495–1501,
1556–1563.w x43 Roberts, G.W., Crow, T.J. and Polak, J.M., Location of neuronal
tangles in somatostatin neurons in Alzheimer’s disease, Nature, 314Ž .1985 92–94.
w x44 Rossor, M.N., Emson, P.C., Mountjoy, C.Q., Roth, M. and Iversen,L.L., Reduced amounts of immunoreactive somatostatin in the tem-poral cortex in senile dementia of Alzheimer type, Neurosci. Lett.,
Ž .20 1980 373–377.w x45 Struble, R.G., Kitt, C.A., Walker, L.C., Cork, L.C. and Price, D.L.,
Somatostatinergic neurites in senile plaques of aged non-humanŽ .primates, Brain Res., 324 1984 394–396.
w x46 Tamminga, C.A., Foster, N.L., Fedio, P., Bird, E.D. and Chase,T.N., Alzheimer’s disease: low cerebral somatostatin levels correlatewith impaired congnitive function and cortical metabolism, Neurol-
Ž .ogy, 37 1987 161–165.w x47 Taniwaki, T. and Schwartz, J.P., Somatostatin enhances neurofila-
ment expression and neurite outgrowth in cultured rat cerebellarŽ .granule cells, DeÕ. Brain Res., 88 1995 109–116.
w x48 Thoenen, H., Neurotrophins and neuronal plasticity, Science, 270Ž .1995 593–598.
w x49 Travis, G.H. and Sutcliffe, J.G., Phenol emulsion-enhanced DNA-driven subtractive cDNA cloning: isolation of low-abundance mon-
Ž .key cortex-specific mRNAs, Proc. Natl. Acad. Sci. USA, 85 19881696–1700.
w x50 Yamashita, A., Hayashi, M., Shimizu, K. and Oshima, K., Ontogenyof somatostatin in cerebral cortex of macaque monkey: an immuno-
Ž .histochemical study, DeÕ. Brain Res., 45 1989 103–111.w x51 Yamashita, A., Hayashi, M., Shimizu, K. and Oshima, K., Neu-
ropeptide-immunoreactive cells and fibers in the developing primateŽ .cerebellum, DeÕ. Brain Res., 51 1990 19–25.
w x52 Zafra F., Hengerer, B., Leibrock, J., Thoenen, H. and Lindholm, D.,Activity dependent regulation of BDNF and NGF mRNAs in the rathippocampus is mediated by non-NMDA glutamate receptors, EMBO
Ž .J., 9 1990 3545–3550.