soil respiration along environmental gradients in olympic national park
TRANSCRIPT

Soil Respiration alongEnvironmental Gradients in
Olympic National Park
Evan S. Kane,1 Kurt S. Pregitzer,1,2* and Andrew J. Burton1
1School of Forestry and Wood Products, Michigan Technological University, Houghton, Michigan 49931, USA; 2USDA ForestService North Central Station, Houghton, Michigan 49931, USA
ABSTRACTAlthough mountainous landscapes dominate largeareas of the Earth, our understanding of how ele-vation and aspect influence soil respiration in com-plex mountainous terrain is very limited. Therefore,we measured soil respiration throughout the grow-ing season in 1999 and 2000 at 11 forested sites inOlympic National Park, Washington, USA along el-evation–climatic gradients. The study sites rangedfrom temperate rain forest to alpine forests neartree line. Soil temperature was a significant predic-tor of soil respiration at all sites, and soil moistureexplained additional variability at three sites (R2
from 0.42 to 0.90, P � 0.01). Soil temperatures atthe highest-elevation sites were 4.5°C cooler thanthose at the lowest elevation, but there were norelationships between soil respiration rates at agiven temperature and elevation or mean annualtemperature that would indicate acclimation of soilrespiration to the cooler temperatures at high-ele-vation sites. Experimental urea additions (1.0 and2.0 g N m�2 y�1) made at seven of the sites had no
consistent effect on soil respiration. Total soil car-bon dioxide (CO2) efflux during the growing season(May–September) varied from 0.34 to 0.75 kg C/m2
and was greater at low-elevation sites with warmersoil temperatures and longer growing seasons. Ele-vation and the length of the frost-free season couldboth be used to predict growing season (r2 � 0.53)and annual (r2 � 0.81) soil CO2 efflux for the 10sites located in steep mountainous terrain. Signifi-cant correlations also existed with mean annualtemperature. These results suggest that warmersoils and a longer snow-free season associated withclimatic warming could cause the mountainousecosystems of the Olympic peninsula to evolve in-creasing amounts of CO2 from all elevations andaspects.
Key words: soil carbon dioxide efflux; soil temper-ature; soil moisture; Pacific Northwest; elevation;aspect; lapse rate; climate change; carbon cycling.
INTRODUCTION
Soil respiration is one of the largest terrestrial car-bon (C) fluxes, and changes in C cycling in forestsoils have the potential to significantly impact theglobal C cycle (Houghton and Woodwell 1989;Raich and Nadelhoffer 1989; Raich and Schlesinger1992). Soil respiration generally increases expo-nentially with temperature and often is limited by
available soil moisture (Ratkowsky and others1982; Orchard and Cook 1983; Carlyle and Than1988; Raich and Schlesinger 1992; Kirschbaum1995; Davidson and others 1998; Savage and Da-vidson 2001); thus, even marginal future changesin global temperatures and precipitation regimesare likely to alter terrestrial C exchange. Tempera-ture and precipitation change with elevation andaspect, but little research has been done to comparesoil respiration among multiple ecosystems alongelevation gradients. This is a matter of concern be-cause nearly one-third of the world’s terrestrial bi-
Received 28 January 2002; accepted 4 September 2002; published onlineMay 6, 2003.*Corresponding author; e-mail: [email protected]
Ecosystems (2003) 6: 326–335DOI: 10.1007/s10021-002-0115-7 ECOSYSTEMS
© 2003 Springer-Verlag
326

omes occur in mountainous regions, and high-ele-vation soils can harbor a considerable amount of C(Prichard and others 2000).
Changes in nitrogen (N) availability can also alterbelowground C allocation and soil carbon dioxide(CO2) efflux (Insam and Palojarvi 1995; King andothers 1996; Zak and others 1996; Ettema and oth-ers 1999; Thirukkumaran and Parkinson 2000).This, coupled with the fact that atmospheric N dep-osition is increasing on a global scale (Vitousek andothers 1997), warrants further study of the impactof N availability on soil respiration.
To gain a better understanding of the factors gov-erning soil respiration in mountainous terrain, wemeasured soil CO2 efflux rates, soil temperature,soil moisture content, and stand and physiographiccharacteristics at 11 sites in Olympic National Park,Washington, USA. Experimental N additions werealso made at seven of the 11 sites. The goals of thestudy were to (a) quantify the effects of soil tem-perature and moisture on soil respiration rates atthe 11 sites; (b) determine the influence of eleva-tion, aspect, and slope on differences among sites insoil temperature, soil moisture availability, and soilCO2 efflux; (c) measure the response of soil respi-ration rates to different levels of N addition; and (d)estimate and contrast seasonal and annual soil CO2
evolution at climatically diverse locations acrossthis complex landscape.
METHODS
Study Sites
The 11 study sites in Olympic National Park in-cluded six sites along an elevation gradient in theDeer Park area, three sites along an elevation gra-dient near Hurricane Ridge, one site in the ElwhaRiver Valley, and one site at the low-elevation HohRainforest (Figure 1). The sites span a distance ofnearly 60 km across the northern portion of thepark and range in elevation from 175 to 1450 m(Table 1). They also encompass a range of moistureavailabilities and temperatures. Annual precipita-tion ranged from approximately 50 cm in the DeerPark area to 334 cm at the Hoh Rainforest, andmean annual air temperature during the study pe-riod ranged from 3.4°C at the north-facing high-elevation site at Deer Park to 7.2°C at the HohRainforest.
The six sites in the Deer Park area were estab-lished at three elevations, denoted as low (DL), mid(DM), and high (DH), on opposing north-(N) andsouth- (S) facing slopes (Table 1). North-facingslopes were characterized by high amounts of
coarse woody debris (CWD) and sparse herbaceouscover. The south-facing slopes had larger-diametertrees, fewer trees per hectare, and greater groundflora coverage (Table 1). Most of the Deer Park siteswere dominated by Douglas-fir (Pseudotsuga men-ziesii Mirbel), but western hemlock (Tsuga hetero-phylla Rafinesque) was also common, especially onN aspects. Subalpine fir (Abies lasiocarpa Hooker)dominated the overstory at the high-elevationnorth-aspect site (Table 1).
The three Hurricane Ridge sites were similarlylocated along an elevation gradient (Table 1). Themid- and low-elevation Hurricane Ridge sites weredominated by Douglas-fir. The high elevation sitehad fairly even proportions of Douglas-fir and sub-alpine fir (Table 1), with bare talus occurring insome places. The Elwha River Valley site (ELW) hadan overstory of mature Douglas-fir and little under-story cover. Soil moisture measurements during thestudy indicated that the SW-facing Elwha site wasthe driest one during the study. The Elwha site islocated near the Hurricane Ridge sites and to adegree represents a lower-elevation S-aspect exten-sion of the Hurricane Ridge elevation gradient (Fig-ure 1).
The Hoh temperate rainforest site (HOH) waslocated approximately 50 km southwest of theother sites. The Hoh site had higher soil water con-tents, much greater annual precipitation, andgreater overstory and understory cover and diver-sity than the other study sites. Sitka spruce (Picea
Figure 1. Location of the 11 study sites in Olympic Na-tional Park, Washington, USA.
Soil Respiration in Olympic National Park 327
sitchensis Bongard) and western redcedar (Thuja pli-cata Donn.) were the dominant overstory species.The Hoh rainforest had a diverse forb communitythat included blueberry shrubs (Vaccinium sp.), vinemaple (Acer circinatum), wood sorrel (Oxalisoregana), and sword fern (Polystichum munitum).This was in contrast to a relatively uniform cover-age of salal (Gaultheria shallon) and Oregon grape(Mahonia nervosa) in the understory at Deer Park,Hurricane Ridge, and Elwha.
Plot Instrumentation and Measurements
At the Hoh and Deer Park areas, four 20 � 20 mplots were established per site: two control plots,and two that were amended with dry urea at ratesof 1.0 and 2.0 g N m�2 y�1, respectively. Nitrogenadditions were applied in five equal incrementsduring the growing season (May–August). At theHurricane Ridge and Elwha areas, only two controlplots were established per site. No additions weremade to the four study sites in these two areas.
Soil temperatures at 1-, 5-, and 20-cm depths andair temperature at 100 cm were monitored contin-uously from May 1999 to August 2000 at each plot.Readings were recorded at 30-min intervals in thesummer and 60-min intervals during the winterusing HOBO H8 loggers (Onset Computer Corpora-
tion, Bourne, MA, USA). Radiation shields wereused to protect air temperature thermistors fromdirect solar radiation. Mean annual temperatureswere calculated from data collected between 5 Au-gust 1999 and 4 August 2000 at 10 of the 11 sites.Equipment failure necessitated the estimation ofmean annual temperature from 350 days of mea-sured data and 15 days of interpolated data for thehigh-elevation Hurricane Ridge site (measured datafrom 22 July 1999 to 5 July 2000).
Soil respiration measurements were made usingan infrared gas analyzer (IRGA), (EGM-2 gas ana-lyzer; PP Systems, Haverhill, MA, USA) with a dy-namic soil respiration chamber (SRC-1; PP Sys-tems). Ten respiration collars were locatedapproximately 2 m inside the plot perimeter, fiveon each of two opposing sides. The respiration col-lars (10.2-cm–diameter schedule 40 polyvinylchlo-ride [PVC]) were inserted to a depth of 2.5 cm atleast 1 week prior to initial measurements and wereleft in place for the duration of the experiment. Atlow- and mid-elevation sites, soil respiration wasmeasured approximately biweekly from Maythrough November in 1999 and June through Au-gust in 2000. At high-elevation sites, soil respirationwas measured from July through November in1999 and June through August in 2000. A portable
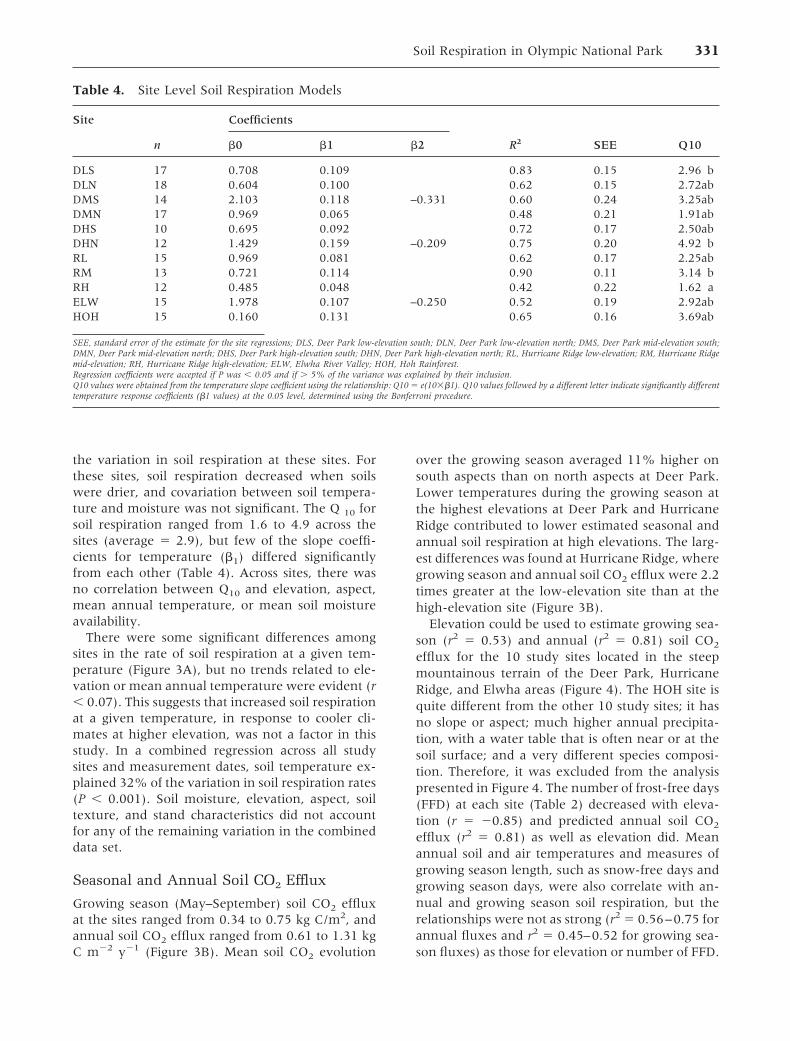
Table 1. Biophysical Properties of the Olympic National Park Study Sites
DLS DLN DMS DMN DHS DHN RL RM RH ELW HOH
Slope (%) 33 14 35 52 71 58 23 60 37 47 0Elevation (m) 710 669 955 898 1423 1450 568 940 1383 480 175Aspect SE N SW N S N N SE SE SW —Age (y) 79 78 221 205 76 73 216 302� 206 183 216Trees ha�1 4700 6450 3475 7875 3950 4075 800 1425 1525 1100 775Basal area
(m2 ha�1)48.0 77.4 69.5 66.8 45.6 62.1 125.8 65.4 60.4 69.6 144.8
Species (% ofbasal area)
PM (71) PM (62) PM (99) PM (47) PM (72) AL (50) PM (77) PM (69) AL (50) PM (96) PS (57)
TH (18) TH (33) — TH (46) PC (6) TH (32) TH (14) TH (29) PM (41) — TP (39)Forb cover (%) 37.8 5.8 54.8 �1 6.3 7.3 27.5 23.0 2.0 17.0 28.0Soil texture
Sand (%) 44 60 63 67 52 54 63 69 82 48 52Silt (%) 35 28 20 20 30 33 23 20 5 31 33Clay (%) 21 12 17 13 18 13 14 10 13 21 15Rock (%) 53 33 54 52 63 54 54 71 48 69 0
DLS, Deer Park low-elevation south; DLN, Deer Park low-elevation north; DMS, Deer Park mid-elevation south; DMN, Deer Park mid-elevation north; DHS, Deer Parkhigh-elevation south; DHN, Deer Park high-elevation north; RL, Hurricane Ridge low-elevation; RM, Hurricane Ridge mid-elevation; RH, Hurricane Ridge high-elevation;ELW, Elwha Ridge Valley; HOH, Hoh Rainforest.The two most dominant overstory species by % basal area are listed for each site using the species codes: AL, Abies lasiocarpa; PC, Pinus contorta; PS, Picea sitchensis; PM,Pseudotsuga menziesii; TP, Thuja plicata; TH, Tsuga heterophyllaBasal areas are based on measurements of all trees with diameter at breast height greater than 10 cm.Mean percent cover of forb species was extrapolated from four 1-m2 vegetation sample plots taken at each site.Mean percentages of sand, silt, and clay are proportions of the nonrock soil fraction.
328 E. S. Kane and others

thermometer was used to measure soil temperatureat 5 cm concurrently with soil respiration measuredat each collar. Barometric pressure readings weretaken during each measurement interval to correctfor pressure broadening of infrared absorptionbands and the effects of atmospheric pressure ongas volumetric concentrations (�mol L �1).
Soil moisture was measured by time domain re-flectometry (TDR cable tester; Tectronix 1502C,Beaverton, OR, USA), using two 30-cm probes atfive sample points per plot (velocity of propagationwas 0.66) (SoilMoisture Equipment Corporation1989). Each pair of probes was located within 30 cmof a respiration collar and was inserted at a 30°angle to avoid channeling of precipitation along theprobe. TDR measurements were taken concurrentlywith soil respiration, enabling direct comparisonsbetween soil volumetric water content and soil res-piration at the plot and site levels. Volumetric watercontent (VWC) was obtained from the TDR wave-forms through the use of coefficients and equations,taken from Gray and Spies (1995), pertaining tosand, silt, and clay fractions present. Two sets of soilcores (5.3-cm diameter, 10 cm deep), from mid andlate growing season, were taken from each plot toobtain soil textures for use in calculating VWC andto determine gravimetric water content of the non-rock fraction for validation of the TDR VWC values.Moisture values from TDR explained 84%–87% (P� 0.001) of the variation in gravimetric water con-tent in the two sets of calibration cores.
Data Analysis
Differences among sites in mean temperature andVWC were tested using Tukey’s studentized rangetest (HSD). The effects of soil temperature, soilmoisture, and N additions across sites were ana-lyzed using a two-factor (study site � N additionrate) analysis of covariance, with temperature andmoisture as covariates and the natural log of soilrespiration as the dependent variable. Plot meansby sample date were used in this analysis.
Multiple regression analysis was used to developempirical relationships for predicting soil CO2 effluxat each site. The natural log of soil respiration wasthe dependent variable, and soil temperature andmoisture were potential predictor variables. Sitemeans for each measurement date were used todevelop the models. Forward selection procedureswere used in accepting predictor variables at �equal to or less than 0.05 and if more than 5% ofthe variance could be explained by adding anotherfactor. The Q 10 of soil respiration was calculated byusing the slope coefficients for temperature (�1),from the linear regressions, in the equation: Q10 �
e(10��1). Differences among sites in �1 were testedusing the Bonferroni procedure (� � 0.05). Site-level correlations of temperature, moisture, andbiophysical characteristics to soil respiration wereexamined through the use of Pearson correlationcoefficients; these variables were used as potentialpredictors of respiration in a combined regressionmodel across all sites where significant.
Seasonal and annual soil CO2 effluxes were esti-mated using individual empirical relationships foreach site, mean daily soil temperatures at 5 cm(measured by HOBO loggers), and VWC (from ac-tual measurements on sample dates and interpo-lated values between sampling dates). Regressionanalyses were used to assess relationships betweenseasonal and annual soil CO2 efflux and elevation,mean annual temperature, average soil moistureavailability, and measures of growing seasonlength. All statistics were performed with PC SAS(Neter and others 1996; Cody and Smith 1997).
RESULTS
Elevation and Aspect Effects on Climate
Air and soil temperatures decreased linearly withelevation (Figure 2 and Table 2), with elevationexplaining 84% (P � 0.001) of the variation in soiltemperatures at 1 cm among all sites. Aspect wasmarginally significant (P � 0.08), explaining anadditional 6% of the variance in soil temperatures.At a given elevation, north-facing slopes at DeerPark had consistently lower temperatures thansouth-facing slopes and fewer snow-free and grow-ing season days (Table 2). Winter (November–Feb-ruary) soil temperature at a depth of 5 cm rangedfrom 0.5°C at DHN to 7.1°C at DLN, with a mean of
Figure 2. Elevational lapse rate for mean annual soiltemperature at 1 cm.
Soil Respiration in Olympic National Park 329

3.0°C for all sites. Snow cover buffered soil temper-atures for at least part of the winter season at thehigh- and mid-elevation sites. Mean soil moistureduring the growing season did not significantly co-vary with elevation or aspect of the sites.
Temperature, Moisture, and N Effects onSoil Respiration Rates
Soil temperature had the greatest influence on soilrespiration rates across sites (Table 3). The responseto N additions varied considerably among sites, withincreases occurring as often as decreases, as indi-cated by the strong interaction between site and N
addition rate (Table 3). Overall, the 2.0 g N m�2 y�1
treatment slightly decreased mean soil respirationrate (circa 8%) relative to the control plots, but thiseffect was driven largely by much lower soil respi-ration rates for this treatment at the HOH site.When the HOH site was excluded from the data set,the N addition rate did not significantly affect meansoil respiration rate for the remaining six sites (P �0.98).
In the multiple-site data set, soil moisture (VWC)was not a significant predictor of soil respirationrates (Table 3); however, regression analyses indi-cate that it was a useful predictor of soil respirationfor some individual sites. In these analyses, theVWC parameter most accurately predicted soil CO2
efflux when inversely transformed. Soil tempera-ture and VWC�1 were used as the driving variablesin predicting respiration rates for each site using thefollowing equation:
NO3 export � FRiverexport[(Sew
� FWSexport (Atm � Fert)]
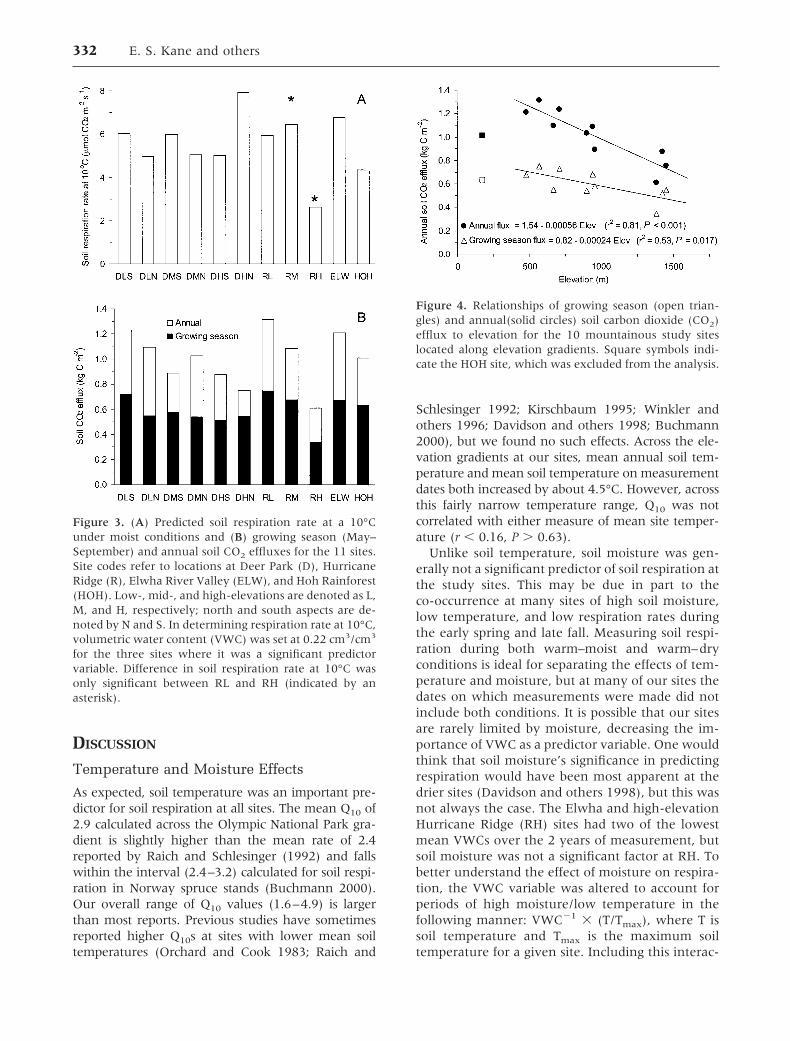
where respiration is in �mol CO2 m�2 s�1, temper-ature is in °C, and VWC is in cm3/cm3. Soil temper-ature was the dominant driver of soil respiration,explaining as much as 90% of the variation amongsample dates in respiration rates at individual sites(Table 4). The moisture parameter significantly im-proved regression models at DMS, DHN, and ELW,explaining 34%, 11%, and 34%, respectively, of
Table 2. Mean Soil Moisture and Soil Temperature Data
SiteMean VWC(cm3/cm3)
Mean Annual Temperature (°C)
FFD SFD GSD
Air Soil
1 m 1 cm 5 cm 20 cm
DLS 0.20 5.8 6.3 6.6 6.3 365 328 204DLN 0.18 5.3 5.7 7.4 6.2 361 312 187DMS 0.20 6.6 6.4 7.0 7.0 341 321 193DMN 0.17 5.7 5.3 5.4 5.5 354 310 171DHS 0.20 4.4 4.6 4.6 4.6 300 229 153DHN 0.22 3.4 3.2 3.1 2.8 233 197 113RL 0.25 6.4 6.8 6.6 6.5 365 338 207RM 0.18 5.1 5.0 4.9 4.9 331 253 166RH 0.18 3.9 4.1 4.0 4.0 224 204 129ELW 0.16 7.1 7.3 7.3 7.4 365 346 218HOH 0.30 7.2 7.6 7.5 7.8 365 344 250
VWC, volumetric water content; FFD, frost-free days, calculated as the number of days annually at or above 0.1 °C at the 1-cm depth; SFD, snow-free days, calculated as daysannually that exhibit diurnal fluctuation in 1 cm soil temperatures indicative of no insulating snow cover; GSD, growing season days, calculated as the number of days annuallywith average soil temperature at or above 5°C at the 1-cm depth. DLS, Deer Park low-elevation south; DLN, Deer Park low-elevation north; DMS, Deer Park mid-elevationsouth; DMN, Deer Park mid-elevation north; DHS, Deer Park high-elevation south; DHN, Deer Park high-elevation north; RL, Hurricane Ridge low-elevation; RM, HurricaneRidge mid-elevation; RH, Hurricane Ridge high-elevation; ELW, Elwha River Valley; HOH, Hoh Rainforest
Table 3. Analysis of Covariance for the Effectsof Study Site, Soil Temperature, Soil Moisture,and Nitrogen (N) Addition on the Natural Log ofSoil Respiration
Source SS df MS F ratio P value
Study site 3.33 6 0.56 7.44 � 0.001N addition rate 0.56 2 0.28 3.78 0.024Study site � N
addition rate 7.68 12 0.64 8.57 � 0.001Soil moisture 0.07 1 0.07 0.88 0.348Soil temperature 17.72 1 17.72 237.07 � 0.001Error 25.48 341 0.08
ss, sum of squares; df, degrees of freedom, ms, mean squareSoil temperature and soil moisture were covariates.
330 E. S. Kane and others
the variation in soil respiration at these sites. Forthese sites, soil respiration decreased when soilswere drier, and covariation between soil tempera-ture and moisture was not significant. The Q 10 forsoil respiration ranged from 1.6 to 4.9 across thesites (average � 2.9), but few of the slope coeffi-cients for temperature (�1) differed significantlyfrom each other (Table 4). Across sites, there wasno correlation between Q10 and elevation, aspect,mean annual temperature, or mean soil moistureavailability.
There were some significant differences amongsites in the rate of soil respiration at a given tem-perature (Figure 3A), but no trends related to ele-vation or mean annual temperature were evident (r� 0.07). This suggests that increased soil respirationat a given temperature, in response to cooler cli-mates at higher elevation, was not a factor in thisstudy. In a combined regression across all studysites and measurement dates, soil temperature ex-plained 32% of the variation in soil respiration rates(P � 0.001). Soil moisture, elevation, aspect, soiltexture, and stand characteristics did not accountfor any of the remaining variation in the combineddata set.
Seasonal and Annual Soil CO2 Efflux
Growing season (May–September) soil CO2 effluxat the sites ranged from 0.34 to 0.75 kg C/m2, andannual soil CO2 efflux ranged from 0.61 to 1.31 kgC m�2 y�1 (Figure 3B). Mean soil CO2 evolution
over the growing season averaged 11% higher onsouth aspects than on north aspects at Deer Park.Lower temperatures during the growing season atthe highest elevations at Deer Park and HurricaneRidge contributed to lower estimated seasonal andannual soil respiration at high elevations. The larg-est differences was found at Hurricane Ridge, wheregrowing season and annual soil CO2 efflux were 2.2times greater at the low-elevation site than at thehigh-elevation site (Figure 3B).
Elevation could be used to estimate growing sea-son (r2 � 0.53) and annual (r2 � 0.81) soil CO2
efflux for the 10 study sites located in the steepmountainous terrain of the Deer Park, HurricaneRidge, and Elwha areas (Figure 4). The HOH site isquite different from the other 10 study sites; it hasno slope or aspect; much higher annual precipita-tion, with a water table that is often near or at thesoil surface; and a very different species composi-tion. Therefore, it was excluded from the analysispresented in Figure 4. The number of frost-free days(FFD) at each site (Table 2) decreased with eleva-tion (r � �0.85) and predicted annual soil CO2
efflux (r2 � 0.81) as well as elevation did. Meanannual soil and air temperatures and measures ofgrowing season length, such as snow-free days andgrowing season days, were also correlate with an-nual and growing season soil respiration, but therelationships were not as strong (r2 � 0.56–0.75 forannual fluxes and r2 � 0.45–0.52 for growing sea-son fluxes) as those for elevation or number of FFD.
Table 4. Site Level Soil Respiration Models
Site Coefficients
n �0 �1 �2 R2 SEE Q10
DLS 17 0.708 0.109 0.83 0.15 2.96 bDLN 18 0.604 0.100 0.62 0.15 2.72abDMS 14 2.103 0.118 –0.331 0.60 0.24 3.25abDMN 17 0.969 0.065 0.48 0.21 1.91abDHS 10 0.695 0.092 0.72 0.17 2.50abDHN 12 1.429 0.159 –0.209 0.75 0.20 4.92 bRL 15 0.969 0.081 0.62 0.17 2.25abRM 13 0.721 0.114 0.90 0.11 3.14 bRH 12 0.485 0.048 0.42 0.22 1.62 aELW 15 1.978 0.107 –0.250 0.52 0.19 2.92abHOH 15 0.160 0.131 0.65 0.16 3.69ab
SEE, standard error of the estimate for the site regressions; DLS, Deer Park low-elevation south; DLN, Deer Park low-elevation north; DMS, Deer Park mid-elevation south;DMN, Deer Park mid-elevation north; DHS, Deer Park high-elevation south; DHN, Deer Park high-elevation north; RL, Hurricane Ridge low-elevation; RM, Hurricane Ridgemid-elevation; RH, Hurricane Ridge high-elevation; ELW, Elwha River Valley; HOH, Hoh Rainforest.Regression coefficients were accepted if P was � 0.05 and if 5% of the variance was explained by their inclusion.Q10 values were obtained from the temperature slope coefficient using the relationship: Q10 � e(10��1). Q10 values followed by a different letter indicate significantly differenttemperature response coefficients (�1 values) at the 0.05 level, determined using the Bonferroni procedure.
Soil Respiration in Olympic National Park 331
DISCUSSION
Temperature and Moisture Effects
As expected, soil temperature was an important pre-dictor for soil respiration at all sites. The mean Q10 of2.9 calculated across the Olympic National Park gra-dient is slightly higher than the mean rate of 2.4reported by Raich and Schlesinger (1992) and fallswithin the interval (2.4–3.2) calculated for soil respi-ration in Norway spruce stands (Buchmann 2000).Our overall range of Q10 values (1.6–4.9) is largerthan most reports. Previous studies have sometimesreported higher Q10s at sites with lower mean soiltemperatures (Orchard and Cook 1983; Raich and
Schlesinger 1992; Kirschbaum 1995; Winkler andothers 1996; Davidson and others 1998; Buchmann2000), but we found no such effects. Across the ele-vation gradients at our sites, mean annual soil tem-perature and mean soil temperature on measurementdates both increased by about 4.5°C. However, acrossthis fairly narrow temperature range, Q10 was notcorrelated with either measure of mean site temper-ature (r � 0.16, P 0.63).
Unlike soil temperature, soil moisture was gen-erally not a significant predictor of soil respiration atthe study sites. This may be due in part to theco-occurrence at many sites of high soil moisture,low temperature, and low respiration rates duringthe early spring and late fall. Measuring soil respi-ration during both warm–moist and warm–dryconditions is ideal for separating the effects of tem-perature and moisture, but at many of our sites thedates on which measurements were made did notinclude both conditions. It is possible that our sitesare rarely limited by moisture, decreasing the im-portance of VWC as a predictor variable. One wouldthink that soil moisture’s significance in predictingrespiration would have been most apparent at thedrier sites (Davidson and others 1998), but this wasnot always the case. The Elwha and high-elevationHurricane Ridge (RH) sites had two of the lowestmean VWCs over the 2 years of measurement, butsoil moisture was not a significant factor at RH. Tobetter understand the effect of moisture on respira-tion, the VWC variable was altered to account forperiods of high moisture/low temperature in thefollowing manner: VWC�1 � (T/Tmax), where T issoil temperature and Tmax is the maximum soiltemperature for a given site. Including this interac-
Figure 3. (A) Predicted soil respiration rate at a 10°Cunder moist conditions and (B) growing season (May–September) and annual soil CO2 effluxes for the 11 sites.Site codes refer to locations at Deer Park (D), HurricaneRidge (R), Elwha River Valley (ELW), and Hoh Rainforest(HOH). Low-, mid-, and high-elevations are denoted as L,M, and H, respectively; north and south aspects are de-noted by N and S. In determining respiration rate at 10°C,volumetric water content (VWC) was set at 0.22 cm3/cm3
for the three sites where it was a significant predictorvariable. Difference in soil respiration rate at 10°C wasonly significant between RL and RH (indicated by anasterisk).
Figure 4. Relationships of growing season (open trian-gles) and annual(solid circles) soil carbon dioxide (CO2)efflux to elevation for the 10 mountainous study siteslocated along elevation gradients. Square symbols indi-cate the HOH site, which was excluded from the analysis.
332 E. S. Kane and others

tion variable explained 12% more of the variance (R2
� 0.54, P � 0.004) than temperature alone at the RHsite. This suggests that the effects of variation in soilmoisture are more important during warm periods.Variations in VWC during drier summer periods mayhave represented larger changes in matric potentialthan variations of similar magnitude during moistspring and fall periods, and thus may have been moresignificant to roots and soil organisms.
In the combined regression across all study sites andmeasurement dates, soil temperature explained only32% of the variation in the overall soil respirationdata set. Much of the remaining variation occurredbetween sites, but it was not related to site differencesin soil moisture, elevation, aspect, or measured over-story stand characteristics. Hanson and others (1993)studied the effects of aspect (northeast versus south-west slope) and topographic position (ridge top versusvalley bottom) on net forest floor CO2 efflux in asmall Tennessee watershed. Their study similarlyfound no consistent relationship between soil respira-tion and aspect or topographic position. Althoughmuch of the site-to-site variability in soil respiration inour study remains unexplained, likely possibilities in-clude differences among sites in soil organic mattercontent, stand-level root respiration, and the quantityand quality of annual above- and belowground litterinputs (Raich and Schlesinger 1992; Boone and oth-ers 1998; Rustad and others 2000; Burton and others2002).
Fertilizer Effects
Several investigators have reported decreases in soilrespiration following fertilizer additions to forest
soils (Insam and Palojarvi 1995; Ettema and others1999; Thirukkumaran and Parkinson 2000; Bow-den and others 2000). Our overall results wereconsistent with these reports, indicating an 8% de-cline in soil respiration at the 2.0 g N m�2 y�1
application rate. However, it must be noted that thisfinding was driven primarily by one fertilized plot atthe HOH site and that relative differences amongtreatments at HOH remained the same throughoutthe experiment, even during initial measurementswhen no N had been applied. The application ofrelatively small amounts of urea (1.0 and 2.0 g Nm�2 y�1) for two growing seasons did not consis-tently affect soil respiration, because increases anddecreases in respiration rates were equally commonon fertilized plots at the other six sites receiving Nadditions. Other investigators have also reportedvarying initial responses of soil respiration to ureaadditions; the long-term effect is typically a reduc-tion (Soderstrom and others 1983; Nohrstedt andBorjesson 1998). The total amounts of N applied inthis study were quite low compared to other studiesshowing a response to N additions (Bowden andothers 2000), and the effects of N additions some-times take several years to become apparent, so it ispossible that continued fertilizer additions wouldeventually lead to a more clearly discernable, con-sistent response.
Seasonal and Annual Soil CO2 Efflux
The range of annual soil respiration fluxes esti-mated in this study generally agrees with resultsfrom previous studies of temperate coniferous eco-systems (Table 5). Our average annual soil CO2
Table 5. Estimated Annual Soil Respiration Rates for Some Common Temperate Forest Ecosystems
Study SitesEstimated Efflux(kg C m�2 y�1) Location Reference
Site range (RH–RL) 0.51–1.31 Olympic National Park, WA, USA Present studyMean of all sites 1.00 Olympic National Park, WA, USA Present studyDouglas-fir 0.49 A. E. Thompson Research Center, WA, USA Vogt and others 1980Western hemlock 0.65 A. E. Thompson Research Center, WA, USA Vogt and others 1980Silver fir 0.62 A. E. Thompson Research Center, WA, USA Vogt and others 1980Red alder 0.57 A. E. Thompson Research Center, WA, USA Vogt and others 1980Coniferous forests 0.68 A. E. Thompson Research Center, WA, USA Raich and Schlesinger 1992Norway spruce 0.71 NE Bavaria, Germany Buchmann 2000Radiata pine 1.00 Victoria, Australia Carlyle and Than 1988Maple swamp 0.53 Harvard Forest, Petersham, MA, USA Davidson and others 1998Hemlock 0.67 Harvard Forest, Petersham, MA, USA Davidson and others 1998Mixed hardwood 0.85 Harvard Forest, Petersham, MA, USA Davidson and others 1998Boreal aspen 0.81–0.91 N Saskatchewan, Canada Russell and Voroney 1998
RH, Hurricane Ridge high-elevation; RL, Hurricane Ridge low-elevationThe respiration rates for the forests at the A. E. Thompson Research Center were estimated by Raich and Schlesinger (1992) using the data of Vogt and others (1980).
Soil Respiration in Olympic National Park 333

efflux was at the upper end of previously reportedvalues. A potential source of error in our annualestimates is the use of empirical relationships de-rived from May to November to predict soil CO2
efflux for the overwinter period, during which soiltemperatures were often lower than those used toderive the relationships. The growing season respi-ration relationships may not hold during wintermonths due to seasonal acclimation to cold temper-atures (Edwards 1991; Bouma and others 1997),higher Q10 values at temperatures close to zero(Schleser 1982; Orchard and Cook 1983; Kirsch-baum 1995; Buchmann 2000), or the influence ofdeep snow cover on soil CO2 losses. Furthermore, itis possible that the active soil microbial pool shiftsfrom mesophiles to psycrophiles in the winter.
For these reasons, our estimates of overwintersoil CO2 efflux may be less reliable than our grow-ing season estimates. However, we think that ourannual flux estimates and the relative differencesamong sites are reasonably accurate because theproportion of the annual flux values attributable todays with temperatures colder than the minimumencountered during our field measurements wasless than 30%. This lessens the impact of overwin-ter errors on our annual estimates. For example,errors as large as 20% in our overwinter estimateswould change the annual values by only 6%.
Elevation was a good predictor of growing seasonand annual soil CO2 efflux at the 10 mountainoussites along the elevation gradients (Figure 4), but itis possible that this elevation–soil respiration rela-tionship would change if regional temperatures in-creased. The ability of factors such as FFD and meanannual temperature to also predict growing seasonand annual soil respiration, and the importance ofsoil temperature at all sites for predicting respirationrates on a given day, all suggest that the amount ofsoil CO2 evolved at a given elevation will increase ifmean annual temperature increases. Warmer soiltemperatures and a longer snow-free season wouldboth likely contribute to this effect. General circu-lation models predict an annual increase in temper-ature of 2.5°C for the Pacific Northwest by the year2050, with little net change in precipitation (MaxPlanck Institute, model ECHAM1-A) (Cubasch andothers 1992). Using the empirical relationships be-tween temperature and soil respiration for our sites,such an increase in soil temperature would result ina mean increase in predicted soil CO2 evolution ofmore than 15% across the Olympic National Parkelevation gradients. At least some of this potentialincrease in soil CO2 loss would likely be offset byincreases in photosynthetic C assimilation over alonger growing season, but any increase in the an-
nual rate of soil CO2 efflux represents a potentialreduction in a forest’s net annual storage of C.
The possibility that roots and microbes might ac-climate to warmer temperatures, respiring less at agiven temperature in a warmer climate, provides ameans by which increased temperature may notresult in changes in soil CO2 efflux as large as thoseour empirical relationships predict. However, wesaw no evidence of warm sites having lower, “ac-climated,” soil respiration rates at a given tempera-ture when comparing sites across the 4.5°C soiltemperature gradient that existed between ourwarmest and coolest sites (Figure 3A). If globaltemperatures increase, warmer soils and longersnow- and frost-free seasons are likely to causemountainous ecosystems, such as those in OlympicNational Park, to evolve increasing amounts of CO2
from the soil at all elevations and aspects.
ACKNOWLEDGMENTS
This research was supported by the US Environ-mental Protection Agency (assistance agreementCR826541-01-0) and the National Park Service. Weare grateful to Jennifer Ashby, Bill Baccus, AnitraBennett, Jiquan Chen, Margaret Gale, Sarah Green,Scott Pugh, Sari Saunders, Leigh Winowiecki, andThomas Wyse for their help and advice.
REFERENCES
Boone RD, Nadelhoffer KJ, Canary JD, Kaye JP. 1998. Rootsexert a strong influence on the temperature sensitivity of soilrespiration. Nature 396:570–72.
Bouma TJ, Nielsen KL, Eissenstat DM, Lynch JP. 1997. Estimat-ing respiration of roots in soil: interactions with soil CO2soiltemperature and soil water content. Plant Soil 195:221–32.
Bowden RD, Rullo G, Steven GR. 2000. Soil fluxes of carbondioxide, nitrous oxide and methane at a productive temperatedeciduous forest. J Environ Qual 29:268–76.
Buchmann N. 2000. Biotic and abiotic factors controlling soilrespiration rates in Picea abiesstands. Soil Biol Biochem 32:1625–1635.
Burton AJ, Pregitzer KS, Ruess RW, Hendrick RL, Allen MF.2002. Root respiration in North American forests: effects ofnitrogen concentration and temperature across biomes. Oeco-logia 131:559–68.
Carlyle JC, Than UB. 1988. Abiotic controls of soil respirationbeneath an eighteen-year-old Pinus radiata stand in south-eastern Australia. J Ecol 76:654–62.
Cody RP, Smith JK. 1997. Applied statistics and the SAS pro-gramming language. Englewood Cliffs (NJ): Prentice-Hall.
Cubasch U, Hasselmann K, Hock H, Maier-Reimer E, Mikola-jewicz U, Santer BD, Sausen R. 1992. Time-dependent green-house warming computations with a coupled ocean–atmo-sphere model. Clim Dynam 9:55–69.
Davidson EA, Belk E, Boone RD. 1998. Soil water content andtemperature as independent or confounded factors controllingsoil respiration in a temperate mixed hardwood forest. GlobalChange Biol 4:217–27.
334 E. S. Kane and others

Edwards NT. 1991. Root and soil respiration responses in Pinustaeda L. seedlings. New Phytol 118:315–21.
Ettema CH, Lowrance R, Coleman DC. 1999. Riparian soil re-sponse to surface nitrogen input: temporal changes in denitri-fication, labile and microbial C and N pools, and bacterial andfungal respiration. Soil Biol Biochem 31:1609–24.
Gray AN, Spies TA. 1995. Water content measurement in forestsoils and decayed wood using time domain reflectometry. CanJ For Res 25:376–85.
Hanson PJ, Wullschleger SD, Bohlman SA, Todd DE. 1993.Seasonal and topographic patterns of forest floor CO2effluxfrom an upland oak forest. Tree Physiol 13:1–15.
Houghton RA, Woodwell GM. 1989. Global climatic change. SciAm 260:36–44.
Insam H, Palojarvi A. 1995. Effects of forest fertilization onnitrogen leaching and soil microbial properties in the northerncalcareous alps of Austria. Plant Soil 168/169:75–81.
King JS, Thomas RB, Strain BR. 1996. Growth and carbon ac-cumulation in root systems of Pinus taeda and Pinus pon-derosa seedlings as affected by varying CO2, temperature andnitrogen. Tree Physiol 16:635–42.
Kirschbaum MUF. 1995. The temperature dependence of soilorganic matter decomposition, and the effect of global warm-ing on soil organic storage. Soil Biol Biochem 27:753–60.
Neter J, Kutner MH, Nachtsheim CJ, Wasserman W. 1996. Ap-plied linear regression models. New York: McGraw-Hill.
Nohrstedt H-O, Borjesson G. 1998. Respiration in a forest soil 27years after fertilization with different doses of urea. Silva Fenn32:383–8.
Orchard VA, Cook FJ. 1983. Relationship between soil respira-tion and soil moisture. Soil Biol Biochem 15:447–53.
Prichard SJ, Peterson DL, Hammer RD. 2000. Carbon distribu-tion in subalpine forests and meadows of the Olympic Moun-tains, Washington. Soil Sci Soc Am J 64:1834–45.
Raich JW, Nadelhoffer KJ. 1989. Belowground carbon allocationin forest ecosystems: global trends. Ecology 70:1346–54.
Raich JW, Schlesinger WH. 1992. The global carbon dioxide flux
in soil respiration and its relationship to vegetation and cli-mate. Tellus 44B:81–99.
Ratkowsky DA, Olley J, McMeekin TA, Ball A. 1982. Relation-ship between temperature and growth rate of bacterial cul-tures. J Bacteriol 149:1–5.
Russell CA, Voroney RP. 1998. Carbon dioxide efflux from thefloor of a boreal aspen forest 1: relationship to environmentalvariables and estimates of C respired. Can J Soil Sci 78:301–10.
Rustad LE, Huntington TG, Boone RD. 2000. Controls on soilrespiration: implications for climate change. Biogeochemistry48:1–6.
Savage KE, Davidson EA. 2001. Interannual variation of soilrespiration in two New England forests. Global BiogeochemCycles 15:337–50.
Schleser GH. 1982. The response of CO2 evolution from soils toglobal temperature changes. Z Naturforsch 37a:287–91.
Soderstrom B, Baath E, Lundgren B. 1983. Decrease in soilmicrobial activity and biomass owing to nitrogen amend-ments. Can J Microbiol 29:1500–6.
Soil Moisture Equipment Corporation. 1989. 6050x1 operatinginstructions manual. Santa Barbara (CA): Soil Moisture.
Thirukkumaran CM, Parkinson D. 2000. Microbial respiration,biomass, metabolic quotient and litter decomposition in alodgepole pine forest floor amended with nitrogen and phos-phorous fertilizers. Soil Biol Biochem 32:59–66.
Vitousek PM, Aber JD, Howarth R, Likens GE, Matson PA,Schindler DW. 1997. Human alteration of the global nitrogencycle: sources and consequences. Ecol Appl 7:737–50.
Vogt KA, Edmonds RL, Antos GC, Vogt DJ. 1980. Relationshipsbetween CO2evolution, ATP concentrations and decomposi-tion in four forest ecosystems in western Washington. Oikos35:72–9.
Winkler JP, Cherry RS, Schlesinger WH. 1996. The Q 10relation-ship of microbial respiration in a temperate forest soil. Soil BiolBiochem 28:1067–72.
Zak DR, Ringelberg DB, Pregitzer KS, Randlett DL, White DC,Curtis PS. 1996. Soil microbial communities beneath Populusgrandidentata grown under elevated atmospheric CO2. EcolAppl 6:257–62.
Soil Respiration in Olympic National Park 335