soil compaction, corn yield response and soil nutrient ... · soil compaction, corn yield response...
TRANSCRIPT

Soil compaction, corn yield response and soil nutrient pool dynamics within an integrated
crop-livestock system in Illinois
Benjamin F. Tracy* and Yan Zhang
Running Head: Soil and crop responses in an integrated crop-livestock system
__________________________
B.F. Tracy, Department of Crop, Soil and Environmental Sciences, Virginia Polytechnic Institute
and State University, Blacksburg, Virginia 24061. Y. Zhang, Department of Crop Sciences,
University of Illinois, Urbana, Illinois 61801
Received: ___________
* Corresponding author ([email protected])

2
ABSRACT
Integrated crop-livestock systems directly link crop and livestock production together to
generate positive economic and environmental outcomes. Some methods used in integrated
systems, like winter grazing on cropland, could negatively affect soil properties and crop
productivity. We compared soil compaction, corn yield and soil nutrient pools between an
integrated crop-livestock system and continuous corn system to address this issue. The study
was conducted near Pana, Illinois, USA between 2002 and 2006. Soil compaction was evaluated
indirectly by measuring soil penetration resistance (PR) and surface CO2 effluxes. Total soil C,
N and microbial biomass C, were measured from 2002 to 2005. Soil PR and CO2 effluxes
showed inconsistent trends related to soil compaction and cattle presence. Corn yield from
2004 to 2006 was higher (P = 0.01) in the integrated system (11.6 Mg ha -1) compared with
continuous corn (10.6 Mg ha-1). Total soil C concentration increased significantly from 2002 to
2005 within components of the integrated system but remained unchanged in continuous corn.
Microbial biomass C was also higher in the integrated system but only in 2005. The study
determined that integration of crops with livestock had generally positive effects on crop yield
and soil organic matter despite the potential for livestock to compact soil during winter grazing.

3
Integrated crop-livestock systems involve linking crop and livestock production together
to generate positive economic and environmental outcomes (Allen et al., 2007). The actual
relationship between crops and livestock can vary in these systems. It may range from relatively
intimate, within-farm integration of crops and livestock (e.g., grazing crop residues after grain
harvest) to more indirect relationships (e.g., shared manure application among crop farms within
a region). Most evidence suggests integrated crop-livestock systems are more environmentally
protective, or restorative, than modern cropping systems that rely on monoculture and heavy
external inputs of fertilizer and pesticide (Allen et al., 2005; Katsvairo et al., 2006, Sulc and
Tracy 2007). Allen et al. (2005) found that an integrated cattle-cotton (Gossypium hirsutum L.)
system with 54% of the land in permanent C4 grass pasture used 23% less irrigation water, 40%
less fertilizer N, less chemical inputs, and was 90% more profitable than the cotton monoculture.
In the northern US Great Plains, diversifying cropping systems with forages was found to
increase grain crop yields, reduce weed pressure, and improve soil quality (Entz et al., 2002).
Integrated systems may also help reduce winter feed costs in temperate regions. Winter feed can
often account for half of the annual production costs in a typical beef cow operation
(Schoonmaker et al., 2003). Cost reductions are often achieved by grazing grain crop residues or
cool-season annual crops after harvest to save on purchased hay or grain (Klopfenstein et al.,
1987; Ward, 1978).
Some integrated systems involve grazing livestock on cropland. A concern is that
livestock presence on cropland would negatively affect soil properties and subsequent crop
productivity. This situation may occur when livestock trample on moist, non-sodbearing
cropland soils causing compaction and subsequent crop yield reductions though alteration of soil
physical and biological properties (Krenzer et al., 1989; Mapfumo et al., 1999; Worrell et al.,

4
1992). Soil compaction was one component addressed in an integrated crop-livestock system
experiment that was initiated in 2002 at the University of Illinois, Dudley Smith Farm near Pana,
IL. In this system, cash crops consisting of corn (Zea mays L.) and oat (Avena sativa L.) were
grown in summer while cattle grazed adjacent perennial pastures (cool and warm-season
grasses). When pastures became dormant in fall, cattle were moved to croplands where they
spent late fall and winter grazing a mixture of cool-season annual cover crops and corn residues.
In spring, cattle were returned to cool-season grass pastures. The objective of this study was to
determine whether cattle presence on cropland would negatively affect soil quality parameters
and subsequent crop yields.
To meet this objective, we compared soil compaction, soil nutrient pools and corn yield
between the integrated crop-livestock system and a continuous corn system between 2002 and
2006. Soil compaction from cattle trampling was evaluated indirectly by measuring soil
penetration resistance (PR) and surface CO2 effluxes. Soil PR is a measure of soil strength and
can be used as an index of soil compaction (Clark et al., 2004; Unger and Kaspar, 1994).
Surface CO2 efflux is a measure of net CO2 production of a soil that includes respiration of roots
and mineralization of organic matter. Soil compaction from machine traffic or cattle trampling
could reduce soil respiration by reducing pore space and limiting O2 diffusion (Conlin and van
den Driessche, 1996; Conlin and van den Driessche, 2000; Shestak and Busse, 2005; Torbert and
Wood, 1992). Reduced soil respiration may indicate less microbial activity and anaerobic
conditions, both of which could negatively affect crop yield. In this study, we used
measurements of surface CO2 efflux during the growing season as an integrative variable to
determine potential persistence of soil compaction from winter grazing. We hypothesized that
cattle trampling on cropland would increase soil compaction and reduce corn yield relative to

5
corn that experienced no winter grazing.
We also measured several soil nutrient pools - total soil C, N and microbial biomass C
and used them as indicators of soil quality. Total C and N closely reflect quantity of soil organic
matter and microbial biomass C is an active pool of organic matter whose size and activity
strongly influences nutrient availability and plant productivity (Paul, 1984, Chapin et al. 2002).
Microbial biomass C is frequently used as an indicator of soil quality since it tends to be more
sensitive to management than total C (Brookes, 1995; Jordan et al., 1995). Because of the
presence of cattle, perennial grasses and cover crops in the integrated system, we hypothesized
that soil organic matter and soil microbial biomass would be greater compared with continuous
corn.

6
MATERIALS AND METHODS
Site Description and Experimental Design
The study site is located on the Dudley Smith research farm near Pana, Illinois, USA
(39.39 N, 89.08 W). The farm had been in corn-soybean (Glycine max (L.) Merr.) rotation for at
least 10 years before it was converted for use in this experiment. In 2002, all 90 hectares of the
farm were converted to a replicated farming systems experiment that integrates cash grain crops
with beef cattle produced on the same land unit. The area has mean annual temperature of 13.4
°C and mean annual precipitation of 1001 mm. Weather data were taken from a nearby station
(~5 km) maintained by Illinois State Climatologist and Illinois State Water Survey. Soils at the
Dudley Smith farm are of the Virden series. They consist of silty, clay loams classified as fine,
smectitic, mesic vertic argiaquolls. Prior to start of the experiment, 84 soil samples were taken
across the farm for baseline data. Soil pH averaged 6.3, while P and K averaged 32 mg kg -1, and
152 mg kg -1, respectively at a 0-15 cm profile depth. Prior to beginning the experiment,
limestone, P and K were applied to individual treatment plots within the farming system using
recommendations based on soil test results.
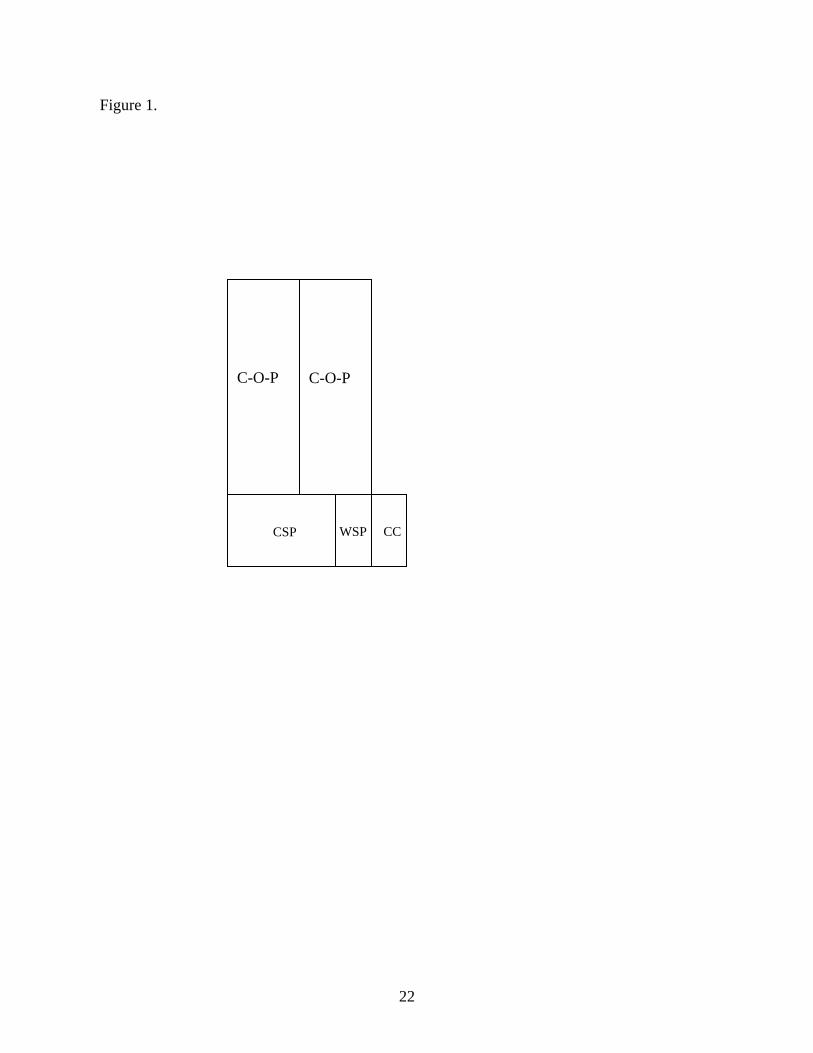
The integrated farming system was replicated three times across the study site. Each
farming system unit consisted of four treatments (Figure 1) that included: 1) corn-oat-pasture
treatment (C-O-P) consisting of equal areas of corn and oats grown in summer and then used for
winter grazing (19 ha), 2) a continuous corn rotation (CC) that was managed exactly as corn in
C-O-P plots except without winter grazing (2 ha), 3) perennial cool-season grass pastures (CSP)
(6 ha), and 4) perennial warm-season grass pastures (WSP) (2 ha). The treatment plots differed
in size because more land was needed to supply winter forage for cattle. Winter pasture cannot

7
be grazed multiple times like summer perennial pasture in rotation and this necessitates need for
more land area. Details on grazing protocols are described below.
Perennial pastures were established in spring of 2002 into a clean seedbed. Within each
farming system, three different cool-season pastures were planted on equal area basis. That is,
three 2 ha pastures within each 6 ha area. The three pastures consisted of: 1) tall fescue (Lolium
arundinaceum (Schreb.) S.J. Darbyshire) sown at 22 kg ha-1, 2) a mixture of orchardgrass
(Dactylis glomerata (L.) and tall fescue sown at 8.5 and 11 kg ha-1, and 3) a mixture of perennial
ryegrass (Lolium perenne (L.), festulolium (Festulolium loliaceum (Hudson) P.V. Fournier]Asch.
& Graebner), tall fescue and orchardgrass sown at 2, 2, 2, and 17 kg ha-1, respectively. Red
clover (Trifolium pretense L.) and white clover (Trifolium repens (L.)) were frost seeded into
pastures in March 2003 at a rate 5 kg ha-1. Warm-season perennial pastures were planted with
Eastern gamagrass (Tripsacum dactyloides (L.) L.), big bluestem (Andropogon gerardii Vitman)
and little bluestem (Schizachyrium scoparium (Michx.) Nash) at a rate of 4.5 kg ha-1,
respectively, and kura clover (Trifolium ambiguum) M. Bieb at 2.7 kg ha-1.
Production of corn and oat started in spring 2003 and continued through 2006. For both
crops, conventional tillage was used for seedbed preparation that included chisel plowing and
field cultivation in spring. In the C-O-P and CC treatments, a Roundup Ready® corn hybrid was
planted each year in mid-late April. Corn was planted to achieve a target population of 75,000
plants ha-1 using 76 cm row spacing. Nitrogen was applied as urea both pre-plant and sidedress
when corn was V6 stage. Nitrogen rates ranged between 150-190 kg N ha-1. Corn was harvested
in mid-late September. Oats planted for grain were sown in early April at a rate of 59 kg ha -1
total weight with a 0.17- m row spacing. Oats were fertilized with urea at a rate of 60 kg N ha-1
at planting. Corn and oats are rotated each year between adjacent fields (Figure 1). Corn was

8
treated with one application of glyphosate isopropylamine salt herbicide (1.12 kg a.i. ha-1) in
June to control weeds. Oats received no herbicide treatment. After oats were harvested in July,
a cool-season annual mixture of oat, cereal rye (Secale cereale L.) and turnip (Brassica spp.)
were sown into a clean, prepared seedbed with seeding rates at 70, 80 and 8 kg ha-1 ,
respectively.
Grazing Management
Grazing on the three replicate farming systems began in fall 2003 and continued through
2006. Cattle grazed cool-season pasture in spring, summer, and fall at a stocking rate of 2.5
cows ha -1 (Figure 1). In each replicate, cool-season pastures were divided into six equal-sized
paddocks for rotational stocking. Warm-season grass pastures were divided into two paddocks
and used to extend the rotation in mid-summer. Paddock residency periods varied between 1 to
6 d during the growing season depending on forage availability. When perennial pastures
become unproductive in late October, we moved cattle to croplands where they grazed cool-
season annuals and corn residues (Figure 1). Pregnant beef cows grazed each cropland area from
November to March with calving occurring in February and March. Stocking rate was 1 cow ha-
1 on croplands. A strip grazing method was used on cropland by moving a single strand of
portable electric fence 25 m approximately every five to ten days depending on forage
availability. Cattle had equal access to corn residues and cool-season annuals in each strip in
addition to a stationary water source. Because cattle needed access to water during the strip
grazing, we could not fence them from previously grazed strips in the field. Cattle were fed hay
and grain as needed when forage became limiting on cropland pastures. Hay and grain were fed
in dry lot areas separate from the treatment plots so did not influence soil properties in sampling
areas. Cattle were given ad libitum access to salt blocks throughout the year.

9
Crop and Soil Sampling
Due to logistic difficulties of sampling the large cropland plots (19 ha), we chose to
confine sampling to smaller, semi-permanent sampling areas that we felt were representative of
each treatment. To do this, we located one 0.5 ha sampling area in each plot such that they
would be grazed at the same time during the strip grazing progression. In this way, each
sampling area across the three replications would be grazed at the same time. Because the strips
were not back-fenced, cattle traveled though plots multiple times to get to water source. Our
placement of sampling areas maximized the likelihood of a similar cattle exposure to all plots
throughout the winter. In the perennial pastures, we choose one random paddock in each
replication for sampling and established a 0.5 ha sampling area in the center. Total soil C, N and
microbial biomass C were measured from soil samples collected in each fall from 2002 to 2005.
Soil penetration resistance, surface CO2 effluxes and corn yield were measured in 2004 and
2005.
Soil PR was measured in late March 2004 and late April 2005 when soils were near or
above field capacity. Twenty electronic penetrometer measurements (1 cm cone diameter, 2 cm
cone length) were taken at a 90° angle to a depth of 46 cm. Measurements were recorded at 2.5
cm intervals and then averaged to obtain two values for 0-23 cm and 24-46 cm depth intervals.
Because soil PR depends on soil moisture, we measured soil moisture gravimetrically from 6
locations in same depth increments. Soils were weighed wet then dried for 48 hr at 50°C and
weighed again. We suspected there might be differences between soil PR under grazed corn
residues and cool-season annuals so we sampled both parts of the C-O-P treatments separately.
Soil PR was measured in perennial pastures, but pastures were much drier than croplands in

10
2005. These differences made it difficult to make a meaningful comparison among the
treatments in 2005 so data from perennial pastures will not be presented.
The yield component method was used to estimate corn grain yield in C-O-P and CC
treatments (Illinois Agronomy Handbook, 2000). In each respective sampling plot, we selected
12 random areas to estimate corn yield. At each sampling point, the number of ears was counted
within a 5.3 m length of row and kernel number estimated for three randomly selected ears.
Those data were used to calculate grain yield. Sampling in 2004, 2005 and 2006 was done in
mid to late August.
For soil variables, 18 soil cores (2.5 cm diameter) were taken from random locations
within each 0.5 ha sampling area to a depth of 15 cm. Sampling was done in each October or
early November after corn harvest. Subsamples were combined and dried for at least 48 hr at
50°C. Roots > 1-mm diameter were removed, and soil finely ground for analyses. Total C and N
concentrations were analyzed using a CHN analyzer (ECS 4010, Costech Analytical
Technologies Inc., Valencia, CA, USA). The chloroform fumigation-incubation method was
used for soil microbial C determination (Jenkinson and Powlson, 1976, Franzleubbers et
al.1996). For processing, ten (10) g of dry soil was placed in a 50 ml glass beaker. Soil moisture
was brought up to ~ 50% of soil water holding capacity. Samples then were pre-incubated in the
dark for 5 days at ~25C in closed mason jars with a few ml of water to maintain humidity. After
pre-incubation, samples were fumigated with chloroform for 24 hr and then transferred to Mason
jars. Each jar contained a 20ml scintillation vial containing 1ml of 2M NaOH to absorb the CO2.
Jars were incubated in the dark at 25C for 10 days. After incubation NaOH was titrated with 1M
HCl, soil microbial biomass C was calculated using a kC value of 0.41 for fumigated samples

11
without subtracting an unfumigated control (Franzluebbers et al., 1999; Voroney and Paul,
1984).
Surface CO2 effluxes were measured using a portable CO2 infrared gas analyzer (LiCor
6400, LiCor Inc., Lincoln, NE, USA) with a closed, static soil respiration chamber.
Measurements were taken from 3 or 4 PVC collars (10 cm diameter × 7 cm) per plot that were
inserted into soil to a depth of 2 cm. Surface CO2 effluxes were measured twice each month
from May to December in 2004 and May to October in 2005. Measurements were conducted
between 9AM to 3PM with exposure time for CO2 efflux measurement averaging about 5
minutes per sampling point depending on soil conditions.
All variables were analyzed using analysis of variance (ANOVA) in the GLM procedure
of SAS (SAS, 2003). Farming system treatments included: 1) corn-oat-pasture treatment(C-O-
P), 2) continuous corn (CC), 3) perennial cool-season grass pasture (CSP) and 4) warm-season
grass pasture (WSP). Subsamples for all variables were pooled within each treatment prior to
analysis to give a sample size of n=3. Treatment and year were used as independent varaibles
and if significant treatment x year interactions were found (P < 0.10), years were analyzed
separately. Corn grain yields were compared between continuous corn (CC) and cropland
pasture (C-O-P). We did not have an ungrazed control for oat grain yield. Total C, N, microbial
biomass C and CO2 effluxes were compared among four main treatments C-O-P, CC, CSP,
WSP. For these variables in the C-O-P treatment, data was pooled from corn residue and cool-
season annual plots. Linear regression was used to determine relationships between total C, N
pools and time over the four year study period. CO2 efflux data collected over each year were
analyzed using a repeated measures ANOVA. Fisher’s LSD test was used to evaluate treatment
differences of significant main effects (P = 0.10).

12
RESULTS AND DISCUSSION
Climatic Conditions
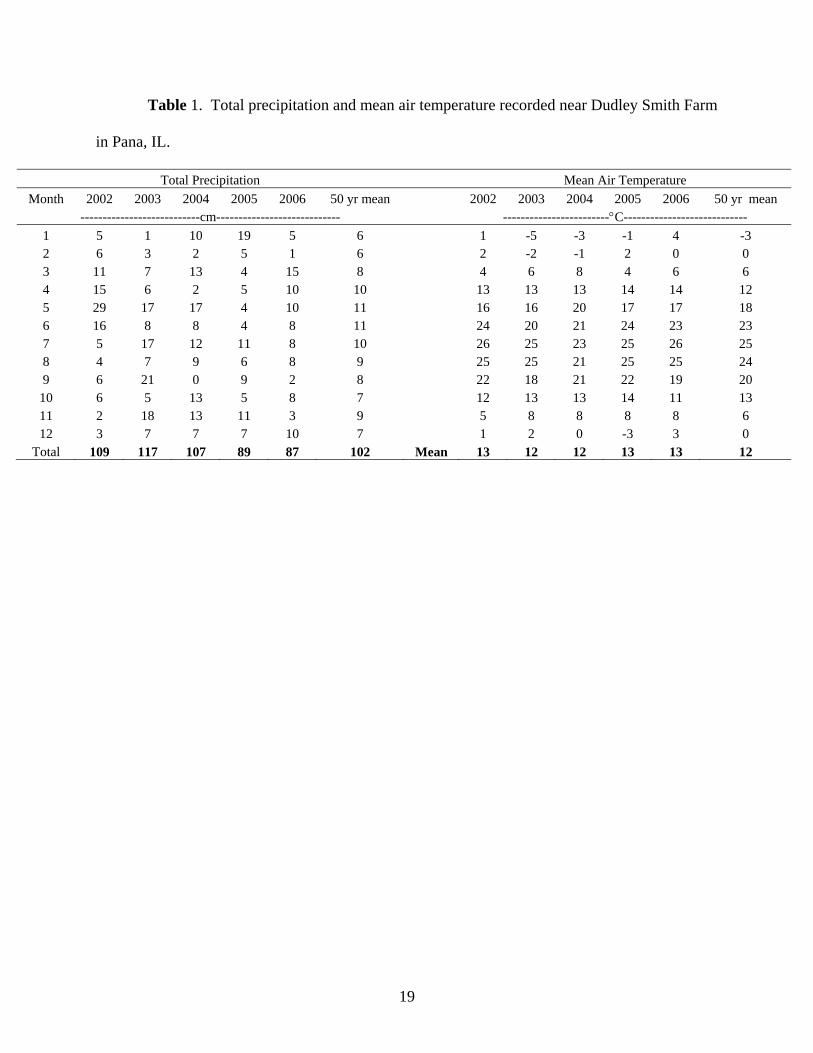
The majority of data in this study was collected between 2004 and 2005. In 2004,
summer was cooler than long-term average data by about 2 ºC from June to August (Table 1).
Temperature in 2005 was closer to the long-term average. Precipitation in 2004 was generally
more variable than 2005 (Table 1). Annual precipitation in 2005 was ~13% below the long-term
(1950-2005) average (1016 mm). The precipitation in spring and early summer (March to June)
in 2005 was drier, ~60% less than long-term average in this period. Because of precipitation and
freeze/thaw events, soils were often wet when cattle were present on croplands during late fall
and winter. Cattle trampling during these times caused severe soil disturbance.
Soil Compaction and Corn Yield
Soil compaction is a complex function of soil texture, moisture, grazing intensity,
vegetation composition and climate (Twerdoff et al., 1999). In this study, we measured soil
compaction indirectly using penetration resistance (PR) and surface CO2 effluxes. Soil
compaction is best measured using both PR and bulk density, but PR should give a good
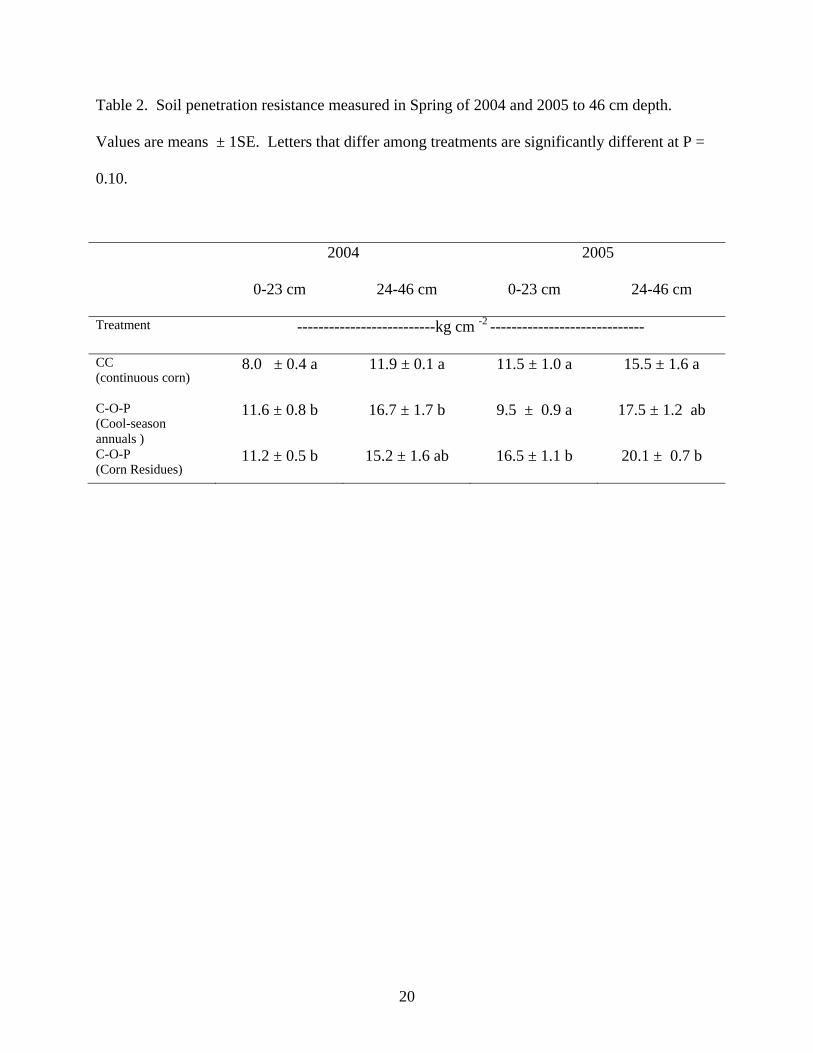
indication of compaction if soil moisture is measured simultaneously. Analysis of PR showed a
significant treatment x year interaction so years were analyzed separately. Soil moisture was
higher in 2004 (281 g kg-1 ) compared with 2005 (242 g kg -1 ) when we measured penetration
resistance. Soil under grazed corn residues tended to be drier (P = 0.13) in 2005 than in 2004,
especially at the 24-26 cm depth (197 g kg-1 vs 237 g kg-1). The dryer soils likely affected soil
PR measurements in 2005 and may explain the higher values under corn residues (Table 2).
Precipitation was also higher in March 2004 (13cm) compared with April 2005 (5cm) when

13
penetration resistance was measured. Overall, PR measurements did not show a consistent trend,
but suggested that cattle presence on grazed cropland may compact soils relative to continuous
corn plots in some years (Table 2). Although PR was measured on perennial pastures, we did
not include this data because pasture soils were much drier than cropland soils in 2005.
Preliminary data collected in 2004 suggested that pasture soils were no more compacted than
cropland, which supports findings from other studies (Mapfumo et al., 1999).
Soil compaction can affect surface CO2 effluxes by reducing air-filled porosity which, in
turn, may restrict oxygen diffusion and increase CO2 accumulation (Conlin and van den
Driessche, 2000; Santruckova et al., 1993). If this situation persists, anaerobic conditions in the
root zone could negatively affect crop growth (Linn and Doran, 1984). The highest mean efflux
rates were reached in late spring or early summer and were close to zero in winter for both
pastures and croplands (data not shown). No differences were found among treatments in 2004,
but perennial pastures showed higher respiration rates (3.6 umol CO2 m-2 s-1) compared with CC
and C-O-P (2.3 umol CO2 m-2 s-1) in 2005 (P < 0.001). Greater fine root biomass and the larger
soil organic C pool, which is a substrate for heterotrophic activity may have contributed to the
higher soil respiration rates measured in pastures in 2005. Similarly, Wagai et al. (1998)
reported a greater soil surface CO2 flux in a native prairie than no-till corn plots in spring and
summer in southern Wisconsin. In forest soils, Shestak and Busse (2005) found that soil
compaction in a clay loam significantly reduced soil respiration as much as 51% on severely
compacted soils. They suggested reduced respiration resulted from less pore space in soils and
not reduced biological activity. Torbert and Wood (1992) found similar results in a laboratory
study using a loamy sand soil. In our study, surface CO2 efflux rates were similar between CC
and C-O-P treatments both years suggesting that if there was soil compaction caused by cattle in

14
winter, it may not have persisted into the growing season. Spring cultivation and tillage in our
study would have likely reduced any shallow compaction that developed over the winter.
Some studies suggest presence of cattle on cropland in winter can compact soils and
reduce crop yield (Krenzer et al., 1989; Mullins and Burmester, 1997; Worrell et al., 1992). Our
results suggest trampling and soil disturbance from cattle presence on cropland had no negative
affect on subsequent corn grain yield and may have helped increase yield over continuous corn
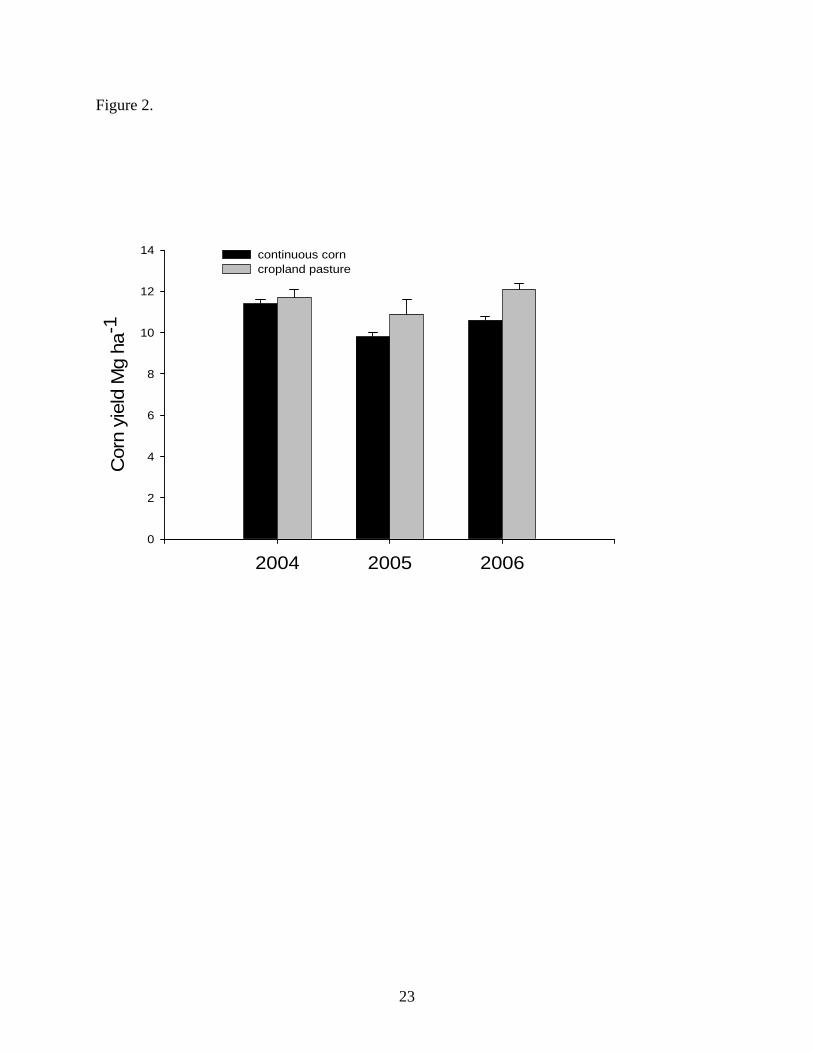
plots. No year x treatment interactions were found and mean corn yield from 2004 to 2006 was
significantly higher (P = 0.01) on C-O-P treatments (11.6 Mg ha -1) compared with continuous
corn (10.6 Mg ha-1). Differences between CC and C-O-P corn grain yield were greater in 2005
and 2006 compared with 2004 suggesting a potential rotational effect (Figure 2). In a study from
Iowa, Clark et al. (2004) showed that winter grazing on corn residues had a minimal effect on
subsequent soybean yield. This was especially true if grazing was restricted to periods when
soils were frozen or if soil was disked before soybean planting. In our study, grazing often
occurred when soils were not frozen. We did, however, use conventional tillage before planting,
and this may have alleviated some compaction. Moreover, we would probably expect to see
greater yield effects related to compaction in a dry year when crops are under stress (Sidhu and
Duiker, 2006; Unger and Kaspar, 1994). In this region, corn is especially sensitive to stress in
July when pollination and grain fill occurs. Rainfall in 2004 and 2005 was above normal in July
and near normal in 2006 so plants were not drought stressed during this critical period (Table 1).
A review by Unger and Kaspar (1994) suggested that PR greater than 20 kg cm-2 in a dry soil
will severely restrict root growth. Our PR measurements approached this threshold in relatively
wet soils (Table 2) so it is possible that PR measured in this study was sufficient to restrict root
growth under prolonged dry conditions.

15
Soil C and N pools
Grasslands generally accumulate more soil organic matter compared with annual
cropland (Bronson et al., 2004; Franzluebbers et al., 2000; Studdert et al., 1997). Microbial
biomass C is an active component of organic matter, and it usually positively associated with
high plant biomass production (Tracy and Frank, 1998) and dense root systems (Stone and
Buttery, 1989). Groffman et al. (1995) observed that microbial biomass C was four times greater
in grazed prairie compared with annual cropland, and this difference was mainly associated with
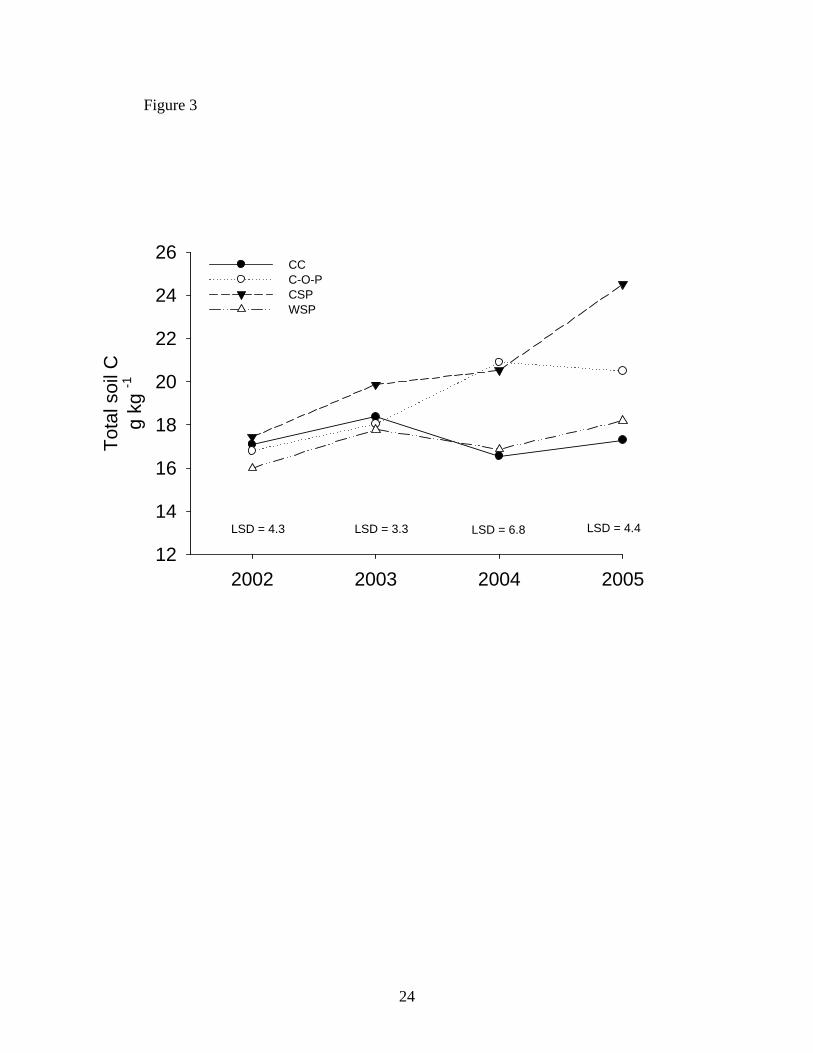
total C inputs. In our study, we found no significant treatment x year interaction for total soil C
or N. Total C increased in CSP and C-O-P treatments, while total C concentrations in CC
remained largely unchanged (Figure 3). Linear regression confirmed that total C concentration
increased in CSP treatments from 2002 to 2005 (R2 = 0.47, P =0.02) and in C-O-P (R2= 0.31,
P=0.07). Total N increased only in the C-O-P treatments – from 1.1 to 1.6 g kg -1 between 2002
and 2005 (Linear regression R2 = 0.47, P=0.02). Total N concentrations in other treatments
remained unchanged over the four years (data not shown). Mean soil C:N ratios ranged from 12
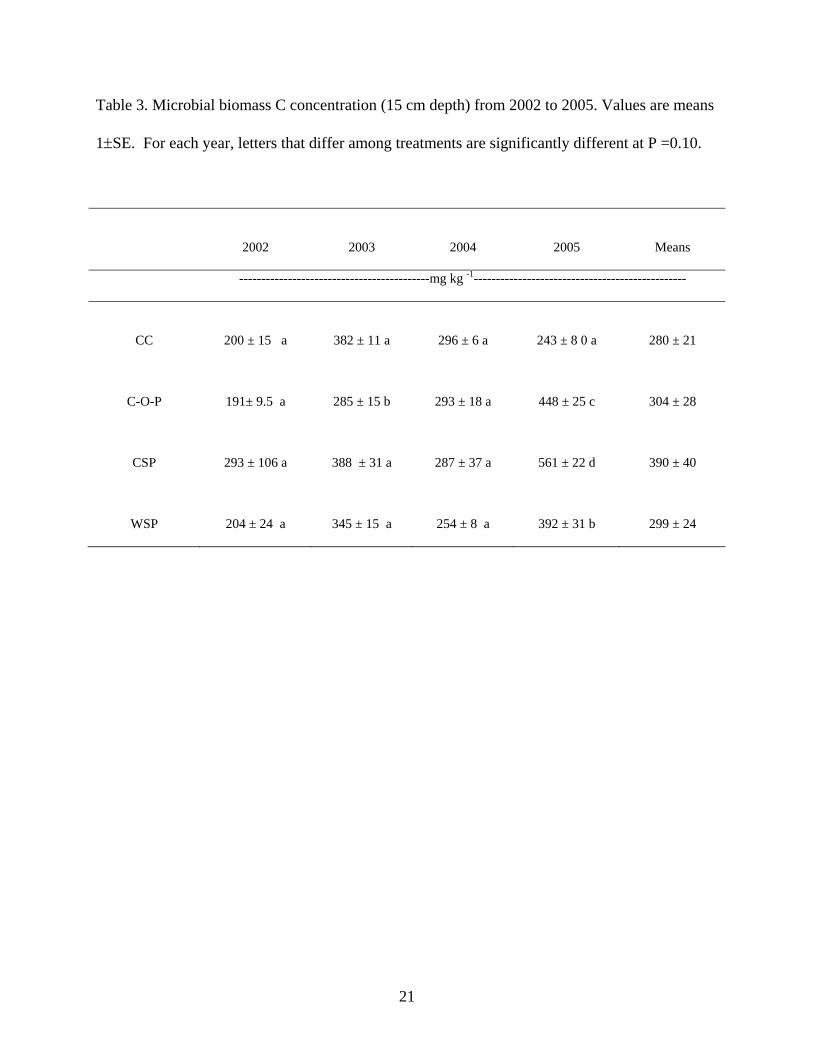
in WSP to 13.3 in C-O-P. Microbial biomass C showed more variation than total C, and we
found a significant treatment x year interaction (P < 0.001). Clear treatment differences were
found for microbial biomass C only in 2005 (Table 3). Microbial biomass concentrations were
highest in CSP, C-O-P treatments and lowest in CC (P < 0.001). These results suggest that CSP
and C-O-P treatments may have built up a greater quantity of labile carbon relative to the other
treatments by 2005. The higher annual variation in microbial biomass may reflect its sensitivity
to management and environmental factors. In terms of soil compaction, it appears to have mixed
effects on soil microbial biomass C with some studies showing no effect (Shestak and Busse,
2005) while others having found reductions in microbial biomass C (Jordan et al., 1999). Cattle

16
presence on cropland appeared to have no detrimental effect on microbial biomass
concentrations in this study.
Our results generally indicate that integration of cattle and pasture within a grain crop
rotation increases the quantity and quality of soil organic matter compared with continuous
cropping. Combining all treatments within the integrated system over fours years revealed that
total C averaged 19.1 g kg -1 while continuous corn averaged 17.3 g kg -1. The biggest
separation in total C concentration occurred in the final year (2005) where total C in the
integrated system averaged 21 g kg -1 and 17.2 g kg-1 in continuous corn. Acosta-Martinez et al.
(2004) compared soil quality parameters in an integrated livestock-cotton production system in
Texas with continuous cotton. They found that soil organic carbon, microbial biomass C and N,
enzyme activities and protozoa populations were higher in the integrated system suggesting that
the quality of soil organic matter was higher in this system compared with continuous cotton.
Like our study, those improvements were largely the result of perennial pasture within the
system. Hoof disturbance from cattle grazing on cropland appeared to have no negative effect on
soil C or N pools relative to continuous corn. In fact, the size of soil C and N pools showed
some of the greatest increases in C-O-P treatments over four years of this experiment. Organic
matter inputs from manure, cool-season annuals and crop residues likely contributed to the
increases in C pools we observed. Only CSP treatments showed greater increases in C pools,
which we expected.
Conclusions
Our data indicated that cattle presence on cropland may have caused soil compaction in
some years, but it had no negative effect on soil properties or corn grain yield. From a whole-
farm perspective, we found significant increases in soil C pools within 5 years of conversion

17
from corn-soybean rotation. These results suggest a farming system integrating cattle with grain
crops can build soil organic matter rapidly, and this should have many positive ramifications for
system productivity overall. A critical component in this system appears to be integration of
perennial pasture and annual cover crops within the system. Cover crop residues, fine root
turnover and manure inputs likely help boost soil organic matter levels and negate soil
compaction that might result from cattle trampling on non-sod bearing soils. Nevertheless,
longer-term evaluation of this integrated system will be required to make robust conclusions
about its impact on soil quality and crop productivity. At this point though, our results suggest
wintering cattle on crop residues and cropland pasture should help reduce winter feeding costs
without negatively affecting crop productivity and soil quality.

18
FIGURE CAPTIONS
Figure 1. Schematic diagram of integrated farming system used in the experiment showing the
four treatments. The farming system is replicated three times across the study site.
Abbreviations are C-O-P – corn-oats-pasture treatment; CSP – cool-season grass pasture; WSP –
warm-season grass pasture; CC – continuous corn.
Figure 2. Estimated corn grain yield from 2004 to 2006 in continuous corn and cropland pasture
(C-O-P) plots.
Figure 3. Trends in total soil C over the four year study period. Abbreviations are C-O-P –
corn-oats-pasture treatment; CSP – cool-season grass pasture; WSP – warm-season grass
pasture; CC – continuous corn. LSD values are given for each year to compare treatments.
19
Table 1. Total precipitation and mean air temperature recorded near Dudley Smith Farm
in Pana, IL.
Total Precipitation Mean Air Temperature Month 2002 2003 2004 2005 2006 50 yr mean 2002 2003 2004 2005 2006 50 yr mean
---------------------------cm---------------------------- ------------------------°C---------------------------- 1 5 1 10 19 5 6 1 -5 -3 -1 4 -3 2 6 3 2 5 1 6 2 -2 -1 2 0 0 3 11 7 13 4 15 8 4 6 8 4 6 6 4 15 6 2 5 10 10 13 13 13 14 14 12 5 29 17 17 4 10 11 16 16 20 17 17 18 6 16 8 8 4 8 11 24 20 21 24 23 23 7 5 17 12 11 8 10 26 25 23 25 26 25 8 4 7 9 6 8 9 25 25 21 25 25 24 9 6 21 0 9 2 8 22 18 21 22 19 20
10 6 5 13 5 8 7 12 13 13 14 11 13 11 2 18 13 11 3 9 5 8 8 8 8 6 12 3 7 7 7 10 7 1 2 0 -3 3 0
Total 109 117 107 89 87 102 Mean 13 12 12 13 13 12
20
Table 2. Soil penetration resistance measured in Spring of 2004 and 2005 to 46 cm depth.
Values are means ± 1SE. Letters that differ among treatments are significantly different at P =
0.10.
2004 2005
0-23 cm 24-46 cm 0-23 cm 24-46 cm
Treatment --------------------------kg cm -2 -----------------------------
CC (continuous corn)
8.0 ± 0.4 a 11.9 ± 0.1 a 11.5 ± 1.0 a 15.5 ± 1.6 a
C-O-P (Cool-season annuals )
11.6 ± 0.8 b 16.7 ± 1.7 b 9.5 ± 0.9 a 17.5 ± 1.2 ab
C-O-P (Corn Residues)
11.2 ± 0.5 b 15.2 ± 1.6 ab 16.5 ± 1.1 b 20.1 ± 0.7 b
21
Table 3. Microbial biomass C concentration (15 cm depth) from 2002 to 2005. Values are means
1±SE. For each year, letters that differ among treatments are significantly different at P =0.10.
2002 2003 2004 2005
Means
-------------------------------------------mg kg -1------------------------------------------------
CC 200 ± 15 a 382 ± 11 a 296 ± 6 a 243 ± 8 0 a
280 ± 21
C-O-P 191± 9.5 a 285 ± 15 b 293 ± 18 a 448 ± 25 c
304 ± 28
CSP 293 ± 106 a 388 ± 31 a 287 ± 37 a 561 ± 22 d
390 ± 40
WSP 204 ± 24 a 345 ± 15 a 254 ± 8 a 392 ± 31 b
299 ± 24
22
Figure 1.
C-O-P
C-O-P
CSP WSP CC
23
Figure 2.
2004 2005 2006
Cor
n yi
eld
Mg
ha-1
0
2
4
6
8
10
12
14 continuous corncropland pasture
24
Figure 3
2002 2003 2004 2005
Tota
l soi
l Cg
kg -1
12
14
16
18
20
22
24
26CCC-O-PCSPWSP
LSD = 4.4LSD = 6.8LSD = 3.3LSD = 4.3

25
REFERENCES
Acosta-Martinez, V., T.M. Zobeck, and V. Allen. 2004. Soil microbial, chemical and physical
properties in continuous cotton and integrated crop-livestock systems. Soil Sci. Soc. Am.
J. 68:1875-1884.
Allen, V.G., C.P. Brown, R. Kellison, E. Segarra, T. Wheeler, P.A. Dotray, J.C. Conkwright,
C.J. Green, and V. Acosta-Martinez. 2005. Integrating cotton and beef production to
reduce water withdrawal from the Ogallala Aquifer in the Southern High Plains. Agron.
J. 97:556-567.
Allen, V. G., M. T. Baker, E. Segarra, C.P. Brown. 2007. Integrated irrigated crop-livestock
systems in dry climates." Agron J. 99: 346-360.
Bronson, K.F., T.M. Zobeck, T.T. Chua, V. Acosta-Martinez, R.S.v. Pelt, and J.D. Booker. 2004.
Carbon and nitrogen pools of southern high plains cropland and grassland soils. Soil Sci.
Soc. Am. J. 68:1695-1704.
Brookes, P.C. 1995. The use of microbial parameters in monitoring soil pollution by heavy
metals. Biol. Fertil. Soils 19:269-279.
Chapin, F.C., H.A. Mooney, and P.A. Matson. 2002. Principles of terrestrial ecosystem ecology.
Springer, New York.
Clark, J.T., J.R. Russell, D.L. Karlen, P.L. Singleton, W.D. Busby, and B.C. Peterson. 2004. Soil
surface property and soybean yield response to corn stover grazing. Agron. J. 96:1364-
1371.
Conlin, T.S.S., and R. van den Driessche. 1996. Short term effects of soil compaction on growth
of Pinus contorta seedlings. Can. J. For. Res. 26:727-739.

26
Conlin, T.S.S., and R. van den Driessche. 2000. Response of soil CO2 and O-2 concentrations to
forest soil compaction at the Long-term Soil Productivity sites in central British
Columbia. Can. J. Soil Sci. 80:625-632.
Entz, M.H., V.S. Baron, P.M. Carr, D.W. Meyer, S.R.S. Jr., and W.P. McCaughey. 2002.
Potential of forages to diversify cropping systems in the Northern Great Plains. Agron. J.
94:240-250.
Franzluebbers, A.J., Haney, R.L., Hons, F.M., Zuberer, D.A., 1996. Determination of microbial
biomass and nitrogen mineralization following rewetting of dried soil. Soil Science
Society of America Journal 60. 1133-1139.
Franzluebbers, A.J., R.L. Haney, and F.M. Hons. 1999. Relationships of chloroform fumigation-
incubation to soil organic pools. Soil Biol Biochem 31:395-405.
Franzluebbers, A.J., J.A. Stuedemann, H.H. Schomberg, and S.R. Wilkinson. 2000. Soil organic
C and N pools under long-term pasture management in the Southern Piedmont USA. Soil
Biol. Biochem. 32:469-478.
Groffman, P.M., C.W. Rice, and J.M. Tiedje. 1995. Denitrification in a tallgrass prairie
landscape. Ecology 74:855-862.
Illinois Agronomy Handbook. 2000. University of Illinois, Urbana, IL.
Jenkinson, D.S., and D.S. Powlson. 1976. The effects of biocide treatments on metabolism in
soil. I. fumigation with chloroform. Soil Biol. Biochem. 8:167-177.
Jordan, D., F. Li, F. Ponder, E.C. Berry, V.C. Hubbard, and K.Y. Kim. 1999. The effects of
forest practices on earthworm populations and soil microbial biomass in a hardwood
forest in Missouri. Appl. Soil Ecol. 13:31-38.

27
Jordan, D.R., W.A. Kremer, K. Bergfield, Y. Kim, and V.N. Cacnio. 1995. Evaluation of
microbial methods as potential indicators of soil quality in historical agricultural fields.
Biol. Fertil. Soils 19:297-302.
Katsvairo, T.W., D.L. Wright, J.J. Marois, D.L. Hartzog, J.R. Rich, and P.J. Wiatrak. 2006. Sod-
livestock integration into the peanut-cotton rotation: a systems farming approach. Agron.
J. 98:1156-1171.
Klopfenstein, T., L. Roth, S. Rivera, and M. Lewis. 1987. Corn residues in beef production
systems. J. Anim. Sci. 65:1139-1148.
Krenzer, E.G., C.F.Chee, and J.F. Stone. 1989. Effects of animal traffic on soil compaction in
wheat pastures. J. Prod. Agric. 2:246-249.
Linn, D.M., and J.W. Doran. 1984. Effect of water-filled pore-space on carbon-dioxide and
nitrous-oxide production in tilled and nontilled soils. Soil Sci. Soc. Am. J. 48:1267-1272.
Mapfumo, E., D.S. Chanasyk, M.A. Naeth, and V.S. Baron. 1999. Soil compaction under grazing
of annual and perennial forages. Can. J. Soil Sci. 79:191-199.
Mullins, G.L., and C.H. Burmester. 1997. Starter fertilizer and the method and rate of potassium
fertilizer effects on cotton grown on soils with and without winter grazing by cattle.
Commun. Soil Sci. Plant Anal. 28:739-746.
Paul, E.A. 1984. Dynamics of organic matter in soils. Plant Soil 76:275-285.
Santruckova, H., O. Heinemeyer, and E.A. Kaiser. 1993. The influence of soil compaction on
microbial biomass and organic-carbon turnover in microaggregates and macroaggregates.
Geoderma 56:587-598.
SAS. 2003. The SAS system for windows. Version 9.1 SAS Inst., Cary, NC.

28
Schoonmaker, J.P., S.C. Loerch, J.E. Rossi, and M.L. Borger. 2003. Stock-piled forage or limit-
fed corn as alternatives to hay for gestating and lactating beef cows. J. Anim. Sci.
81:1099-1105.
Shestak, C.J., and M.D. Busse. 2005. Compaction alters physical but not biological indices of
soil health. Soil Sci. Soc. Am. J.69:236-246.
Sidhu, D., and S.W. Duiker. 2006. Soil compaction in conservation tillage: crop impacts. Agron.
J. 98:1257-1264.
Stone, J.A., and B.R. Buttery. 1989. Nine forages and the aggregation of a clay loam soil. Can. J.
Soil Sci. 69:165-169.
Studdert, G.A., H.E. Echeverria, and E.M. Casanovas. 1997. Crop-pasture rotation for sustaining
quality and productivity of a Typic Argiudoll. Soil Sci. Soc. Am. J. 61:1466-1472.
Sulc, R. M. and B. F. Tracy. 2007. Integrated Crop-Livestock Systems in the U.S. Corn Belt.
Agron J. 99: 335-345.
Torbert, H.A., and C.W. Wood. 1992. Effects of soil compaction and water-filled pore-space on
soil microbial activity and N losses. Commun. Soil Sci. Plant Anal. 23:1321-1331.
Tracy, B.F., and D.A. Frank. 1998. Herbivore influence on soil microbial biomass and nitrogen
mineralization in a northern grassland ecosystem: Yellowstone National Park. Oecologia
114:556-562.
Twerdoff, D.A., D.S. Chanasyk, E. Mapfumo, M.A. Naeth, and V.S. Baron. 1999. Impacts of
forage grazing and cultivation on near-surface relative compaction. Can. J. Soil Sci.
79:465-471.
Unger, P.W., and T.C. Kaspar. 1994. Soil compaction and root-growth - a review. Agron. J.
86:759-766.

29
Voroney, R.P., and E.A. Paul. 1984. Determination of kc and kn in situ for calibration of the
chloroform fumigation incubation method. Soil Biol. Biochem. 16:9-14.
Wagai, R., K.R. Brye, S.T. Gower, J.M. Norman, and L.G. Bundy. 1998. Land use and
environmental factors influencing soil surface CO2 flux and microbial biomass in natural
and managed ecosystems in southern Wisconsin. Soil Biol. Biochem. 30:1501-1509.
Ward, J.K. 1978. Utilization of corn and grain sorghum residues in beef cow forage systems. J
Anim. Sci. 46:831-840.
Wedin, W.F., and T.J. Klopfenstein. 1995. Cropland pastures and crop residues. p. 193-206. In
R. F. Barnes, et al., (ed.). Forages: Volume II. The Science of Grassland Agriculture.
Iowa State University Press, Ames, IA.
Worrell, M.A., D.J. Undersander, and A. Khalilian. 1992. Grazing wheat to different
morphological stages for effects on grain yield and soil compaction. J. Prod. Agric. 5:81-
85.