sequential generation of hydrogen and methane from xylose by two-stage anaerobic fermentation
TRANSCRIPT

ww.sciencedirect.com
i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 7 ( 2 0 1 2 ) 1 3 3 2 3e1 3 3 2 9
Available online at w
journal homepage: www.elsevier .com/locate/he
Sequential generation of hydrogen and methane from xyloseby two-stage anaerobic fermentation
Jun Cheng a,*, Wenlu Song a,b, Ao Xia a, Huibo Su a, Junhu Zhou a, Kefa Cen a
a State Key Laboratory of Clean Energy Utilization, Zhejiang University, Hangzhou 310027, ChinabCommittee of High Technology Development District, Jining 272000, China
a r t i c l e i n f o
Article history:
Received 9 April 2012
Received in revised form
8 June 2012
Accepted 15 June 2012
Available online 15 July 2012
Keywords:
Xylose
Hydrogen
Methane
Fermentation
Energy conversion efficiency
Methanogenesis
* Corresponding author. Tel.: þ86 571 879528E-mail address: [email protected] (J. C
0360-3199/$ e see front matter Copyright ªhttp://dx.doi.org/10.1016/j.ijhydene.2012.06.0
a b s t r a c t
The sequential generation of hydrogen and methane from xylose by two-stage anaerobic
fermentation was investigated for the first time in this study. The effects of substrate
concentration, bacteria domestication and nitrogen source on hydrogen yield were studied
in the first stage. The genetic characterization of the 16S rDNA was used to analyze the
flora of strains domesticated with xylose and glucose. The maximum hydrogen yield is
190.6 ml H2/g xylose when the xylose feedstock concentration is 1% (w/v), hydrogenogens
are domesticated with xylose and yeast extract is used as nitrogen source. The soluble
metabolite byproducts (SMB) from the hydrogen-producing stage were reutilized by
methanogens to produce methane in the second stage. Over 98 wt % of acetate and
butyrate in the SMB are reutilized to give a methane yield of 216.5 ml CH4/g xylose. The
sequential generation of hydrogen and methane from xylose markedly increases the
energy conversion efficiency to 67.5%.
Copyright ª 2012, Hydrogen Energy Publications, LLC. Published by Elsevier Ltd. All rights
reserved.
1. Introduction wastes and obtain a higher energy utilization rate, thus
On account of the fast depletion of fossil fuels and the serious
environmental pollution caused by them, it is necessary to
develop renewable energy resources. Hydrogen produced from
biomass is a promising energy alternative because it is clean,
renewable, having high-energy density, and is compatible with
both electrochemical and combustion processes [1,2].
Compared with other processes, bio-hydrogen production by
dark fermentation has increasingly attracted interest in recent
years because of its low cost, rapid bacteria growth, high
hydrogen-producing capacity, and abundant feedstocks [3e5].
The annual output of renewable straws in theworld reaches
4.5 billion tons [6], among which polysaccharides cellulose and
hemicellulose account for greater than 50% [7]. By using cheap
cellulosic biomass to produce clean gaseous fuels such as
hydrogen andmethane, it is possible to recycle the agricultural
89; fax: þ86 571 87951616heng).2012, Hydrogen Energy P49
promoting the hydrogen industry and the low-carbon
economy. However, the energy conversion efficiency is low by
yielding only hydrogen from fermentation of biomass. For
example, the theoretical energy conversion efficiency of only-
hydrogen production is just 19.1% by dark fermentation from
water hyacinth [8]. During hydrogen production, many soluble
metabolite byproducts (SMB) such as volatile fatty acids (VFAs)
are also produced. It is not only awaste of carbonaceous energy
resources, but also a secondary pollution to the environment.
Our previous study has reported that SMB in the residual of
hydrogen-producing solutions can be reutilized by the metha-
nogen community to further produce methane, thereby
dramatically increasing the energy conversion efficiency and
diminishing the pollution. For example, the theoretical energy
conversion efficiency is promoted to 63.1% by a combination of
hydrogen fermentation and methanogenesis from water
.
ublications, LLC. Published by Elsevier Ltd. All rights reserved.

i n t e rn a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 7 ( 2 0 1 2 ) 1 3 3 2 3e1 3 3 2 913324
hyacinth [8]. So, there is a broad industrial prospect to cogen-
erate hydrogen and methane from cellulosic biomass.
Direct fermentation of raw cellulosic feedstock is generally
inefficient because cellulose and hemicellulose are not
assimilable to most hydrogen-producing bacteria before
hydrolyzing to monomeric sugars [9]. Cellulose is a macro-
molecule made of glucose and some other hexose, while
hemicellulose is mostly composed of pentose (mainly xylose)
and some other monomeric sugars. In straws, the contents of
cellulose and hemicellulose are generally 30e40% and
20e30%, respectively [8,10,11]. In other words, about
333.3e444.4 g hexose and 227.2e340.8 g pentose can be
generated from 1 kg straws [8]. Fermentative hydrogen
production from glucose has beenwell-studied [12,13], andwe
have also reported both theoretical and experimental results
of cogenerating hydrogen and methane from glucose in our
former study [14]. According to the stoichiometric equation:
C6H12O6 þ 2H2O / 4CO2 þ 4H2 þ 2 CH4, 1 mol glucose can
cogenerate 4molH2 and 2mol CH4 theoretically, viz. 1 g glucose
(molecular weight ¼ 180 g/mol) can cogenerate 498 ml H2 and
249 ml CH4. The fermentative experiments show that the
maximum H2 and CH4 yields are 2.75 mol H2/mol glucose and
2.13 mol CH4/mol glucose. Some researchers have investigated
hydrogen production from xylose by fermentation. However,
the low energy conversion efficiencies through hydrogen
fermentation (�31%) [15e19]werenoteconomic,which resulted
in a bottleneck in industrial application. It is quite necessary to
enhance biogas production from xylose. A combination of
hydrogen fermentation and methanogenesis is an effective
methodto improveenergyconversionefficiency.Thesequential
generation of hydrogen andmethane from xylose by two-stage
anaerobic fermentationwas investigatedfor thefirst time inthis
paper. In the first stage, hydrogen was produced from xylose,
and the influences of substrate concentration, bacteria domes-
tication and nitrogen source (yeast extract) on hydrogen
yield were studied. In the second stage, the SMB from the
hydrogen-producing stage were reutilized by the methanogen
community to further produce methane, and consequently the
energy conversion efficiency was remarkably increased.
2. Materials and methods
2.1. Preparation and characterization of seed inocula
2.1.1. Enrichment of hydrogenogensThe anaerobic activated sludge was sampled from a methane
producing fermenter with swinemanure feedstock in Huzhou
Table 1 e Experimental design.
First stage: hydrogen production
No. Feedstock Hydrogenogenic bacteria Yeast
1 Xylose 3 g Domesticated with glucose, 10 ml e
2 Xylose 3 g Domesticated with xylose, 10 ml e
3 Xylose 3 g Domesticated with xylose, 10 ml 0
4 Xylose 6 g Domesticated with xylose, 10 ml e
5 Xylose 6 g Domesticated with xylose, 10 ml 0
marsh gas plant located in Zhejiang Province, China. The
method of pretreatment of the sludge was the same as we
used before [20], in which xylose was used as substrate to
domesticate the hydrogenogens. The preheated solid sludge
was inoculated with xylose feedstock (10 g/l and 20 g/l) to
begin the fermentation reaction. The preheated solid sludge,
which was domesticated with glucose instead of xylose [21]
was used as the control.
2.1.2. Phylogenetic characterization of hydrogenogen-enriched inoculumTotal DNAwas extracted from 0.1ml of preheated solid sludge
enriched in hydrogenogens, as described in the literature
[22]. Bacterial 16S rRNA genes were amplified with
27F (50-AGAGTTTGATCMTGGCTCAG-30) and 1390R (50-ACGGGCGGTGTGTACAA-30) primer set. Primer 519F (50-CAG-CAGCCGCGGTAATAC-30) was used for sequencing. Cloning
and sequencing of hydrogenogenic 16S rDNAwere carried out
as described previously [23].
The 16S rRNA gene sequences were compared with avail-
able database sequences via a BLAST search from GenBank to
determine their phylogenetic positions. The sequences were
grouped into operational taxonomic units (OTUs) based on
nucleic acid sequence similarity within each library. Phylo-
genetic trees were constructed by MEGA software version 3.0
with the neighbor-joining method based on nucleic acid
sequences [24].
The sequences determined in this study are available in
GenBank under HM238272 to HM238282 (16S rRNA gene
sequences of the bacteria).
2.1.3. Enrichment of methanogensThe original activated sludge was domesticated in a tradi-
tional growthmedia [21] to enrich themethanogens, and then
it was acclimated with SMB from the hydrogen-producing
stage to produce methane.
2.2. Fermentation reactor and experiments
A 350-ml glass bottle with a fermentation volume of 300 ml
was used as the fermentation reactor. Five groups of exper-
iments on cogenerating hydrogen and methane from xylose
feedstock were carried out at 37 �C, as shown in Table 1. The
pure xylose chemical (Sinopharm Chemical Reagent Co.Ltd,
China) as the feedstock was inoculated with the preheated
solid sludge enriched in hydrogenogens in the fermenter,
which was purged with N2 gas for 20 min to create the
anaerobic environment. The pH value was adjusted to 6 � 0.1
Second stage: methane production
extract No. Feedstock Methanogenicbacteria
10 Residual of 1 Domesticated with
nitrilotriacetic acid, 10 ml20 Residual of 2
.5 g 30 Residual of 3
40 Residual of 4
.5 g 50 Residual of 5
XY04 (3/58)
XY08 (7/58)
XY24 (2/58)
Clostridium leptum (AJ305238)
Clostridium sporosphaeroides (GQ243738)
XY02 (3/58)
GL05 (5/57)
Clostridium tepidiprofundum (EF197795)
Clostridium neonatale (EU869234)
XY07 (5/58)
Clostridium butyricum (AJ002592)
GL11 (29/57)
Clostridium cochlearium (M59093)
GL37 (2/57)
GL12 (2/57)
XY49 (2/58)
Clostridium ultunense (NR_026531)
XY39 (3/58)
Sphingomonas echinoides (EU730918)
XY35 (2/58)
Bacillus subtilis (EU304958)
XY57 (6/58)
GL50 (4/57)
100
100
100
77
99
95
100
100
100
100
100
86
100
94
86
72
99
99
44
33
0.02
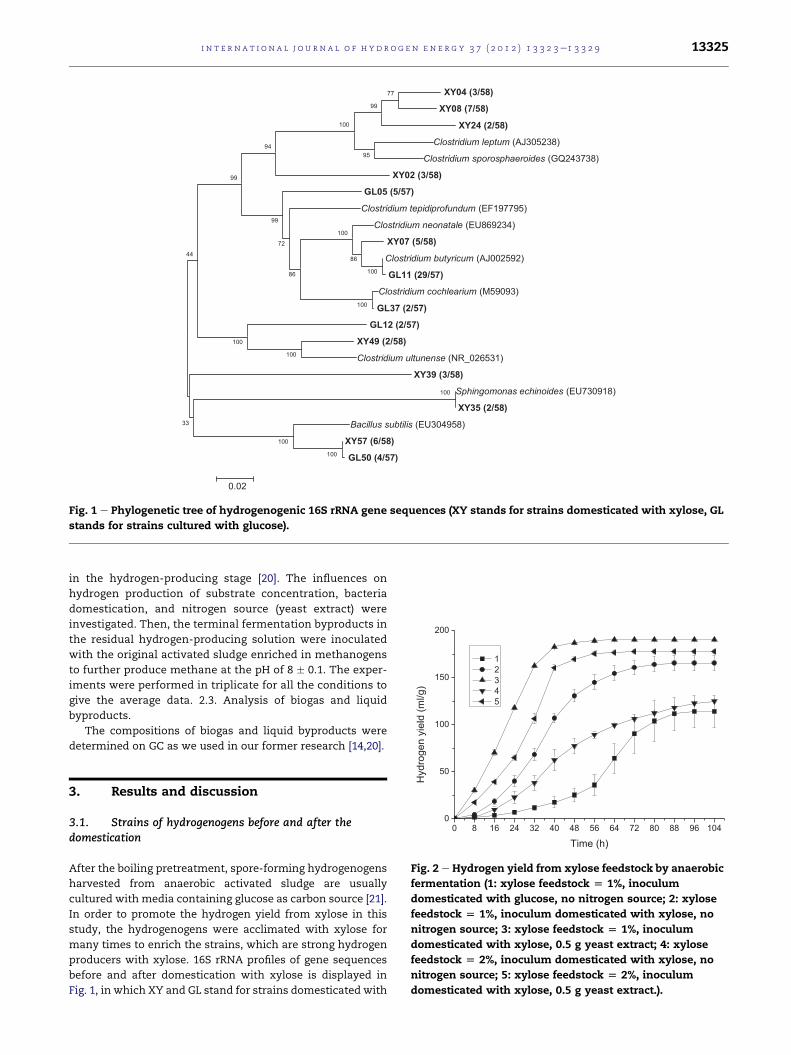
Fig. 1 e Phylogenetic tree of hydrogenogenic 16S rRNA gene sequences (XY stands for strains domesticated with xylose, GL
stands for strains cultured with glucose).
100
150
200
12345
gen
yiel
d (m
l/g)
i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 7 ( 2 0 1 2 ) 1 3 3 2 3e1 3 3 2 9 13325
in the hydrogen-producing stage [20]. The influences on
hydrogen production of substrate concentration, bacteria
domestication, and nitrogen source (yeast extract) were
investigated. Then, the terminal fermentation byproducts in
the residual hydrogen-producing solution were inoculated
with the original activated sludge enriched in methanogens
to further produce methane at the pH of 8 � 0.1. The exper-
iments were performed in triplicate for all the conditions to
give the average data. 2.3. Analysis of biogas and liquid
byproducts.
The compositions of biogas and liquid byproducts were
determined on GC as we used in our former research [14,20].
0 8 16 24 32 40 48 56 64 72 80 88 96 1040
50
Hyd
ro
Time (h)
Fig. 2 e Hydrogen yield from xylose feedstock by anaerobic
fermentation (1: xylose feedstock [ 1%, inoculum
domesticated with glucose, no nitrogen source; 2: xylose
feedstock [ 1%, inoculum domesticated with xylose, no
nitrogen source; 3: xylose feedstock [ 1%, inoculum
domesticated with xylose, 0.5 g yeast extract; 4: xylose
feedstock [ 2%, inoculum domesticated with xylose, no
nitrogen source; 5: xylose feedstock [ 2%, inoculum
domesticated with xylose, 0.5 g yeast extract.).
3. Results and discussion
3.1. Strains of hydrogenogens before and after thedomestication
After the boiling pretreatment, spore-forming hydrogenogens
harvested from anaerobic activated sludge are usually
cultured with media containing glucose as carbon source [21].
In order to promote the hydrogen yield from xylose in this
study, the hydrogenogens were acclimated with xylose for
many times to enrich the strains, which are strong hydrogen
producers with xylose. 16S rRNA profiles of gene sequences
before and after domestication with xylose is displayed in
Fig. 1, in which XY and GL stand for strains domesticated with

0 8 16 24 32 40 48 56 64 72 80 88 96 1040
20
40
60
80
100
120
140
Hyd
roge
n pr
oduc
tion
rate
(ml /
(L·h
))
Time (h)
12345
Fig. 3 e Hydrogen production rate from xylose feedstock by
anaerobic fermentation (1: xylose feedstock [ 1%,
inoculum domesticated with glucose, no nitrogen source;
2: xylose feedstock [ 1%, inoculum domesticated with
xylose, no nitrogen source; 3: xylose feedstock [ 1%,
inoculum domesticated with xylose, 0.5 g yeast extract; 4:
xylose feedstock [ 2%, inoculum domesticated with
xylose, no nitrogen source; 5: xylose feedstock [ 2%,
inoculum domesticated with xylose, 0.5 g yeast extract.).
i n t e rn a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 7 ( 2 0 1 2 ) 1 3 3 2 3e1 3 3 2 913326
xylose and glucose, respectively. There are 58 operative
sequences of XY and the coverage degree is
(3 þ 7þ2 þ 3þ5 þ 2þ3 þ 2þ6)/58 ¼ 57%, while there are 57
operative sequences of GL and the coverage degree is
(5 þ 29þ2 þ 2þ4)/57 ¼ 74%. So the flora is more complex after
domestication with xylose. It can be seen from the phyloge-
netic tree that XY have nine dominant floras, while GL have
five. It is reported that in the spore-forming hydrogenogens
harvested from boiled anaerobic activated sludge, Clostridium
is the dominant flora [25e27]. This study proves that in GL,
Clostridium butyricum is the distinct dominant flora with
a proportion of about 50%. In XY, however, there are only
about 10% of C. butyricum and dominant floras of several other
strains, which aremost related to Clostridium leptum (AJ305238,
similarity of 92%), Clostridium sporosphaeroides (GQ243738,
similarity of 92%), Clostridium ultunense (NR026531, similarity
of 94%) [27], Sphingomonas echinoids (EU730918, similarity of
100%), and Bacillus subtilis (EU304958, similarity of 95%) [28].
Several Clostridium species have been shown to metabolize
Table 2 e Soluble metabolite byproducts from the H2 and CH4-
No.a Ethanol/% (V) Acetate/% (V) Propio
H2 stage CH4 stage H2 stage CH4 stage H2 stage
1/10 0.008 0.0001 0.581 0.005 0.049
2/20 0.011 0.0006 0.626 0.01 0.012
3/30 0.014 0.0002 0.928 0.009 0.027
4/40 0.018 0.0004 1.021 0.012 0.014
5/50 0.019 0.0004 1.254 0.013 0.031
a Conditions of group 1/10e5/50 are the same as Table 1.
xylose by early researches [29e31]. C. butyricum is a strong
hydrogen producer with glucose, but maybe inefficient with
xylose [32]. By enrichment with xylose, strains which
decompose xylose well become dominant in the sludge, and
are expected to give higher hydrogen production rate and
hydrogen yield.
3.2. Hydrogen production from xylose feedstock
The yields (ml/g) and production rates (ml/(L$h)) of hydrogen
fermentation from xylose are shown in Fig. 2 and Fig. 3,
respectively. It is found that when the concentration of xylose
is 1% (w/v), fermentation of strains domesticated with glucose
give a longer lag-phase with a hydrogen yield of
114.1 � 17.2 ml/g (group 1). When strains domesticated with
xylose are used as hydrogenogens and no nitrogen source is
added, the maximum hydrogen production rate is promoted
from 35.8 � 5.5 ml/(L$h) to 48.4 � 1.4 ml/(L$h) and peak time is
advanced from 64 h to 40 h, while the hydrogen yield is
increased from 114.1 � 17.2 ml/g to 165.5 � 7.8 ml/g (group 2).
Themain reason is that the process of domestication changed
the dominant strains in the sludge. Before domestication with
xylose, some genuses of bacteria in the hydrogenogens are
unable to grow with xylose as a sole carbon source due to the
lack of a xylose uptake system [33], resulting in a lower yield of
hydrogen. After the domestication, more hydrogen-producing
bacteria, which could contribute to the degradation of xylose,
reproduce themselves to change the microflora. The results
indicate that the type of feeding carbon substrate is important
in determining the evolution of characteristic hydrogen-
producing communities. The domestication enhances the
microflora’s ability to degrade organics.
The hydrogen production profile of hydrogenogens was
found maximum in the exponential phase of the growth [34],
and most xylose utilizers require yeast extract for growth on
pentose media [35]. In order to get a maximum hydrogen yield
with the bacteria domesticated with xylose, 0.5 g yeast extract
was added into the initial fermentation medium (lack of
nitrogen source) as nitrogen source. It is shown by experiments
that when yeast extract is added as nitrogen source with
bacteria domesticated with xylose, the maximum hydrogen
production rate is promoted from 48.4 � 1.4 ml/(L$h) to
59.6� 6.6 ml/(L$h) and peak time is advanced from 40 h to 24 h,
while the hydrogen yield is further increased to 190.6 � 1.4 ml/
g, which is equivalent to 1.28 � 0.01 mol/mol-xylose (group 3).
The main reason is that yeast extract provides sufficient
producing stages.
nate/% (V) Butyrate/% (V) Valerate/% (V)
CH4 stage H2 stage CH4 stage H2 stage CH4 stage
0.005 0.391 0.001 0.028 0.002
0.004 0.417 0.001 0.014 0.001
0.006 0.411 0.001 0.017 0.001
0.004 0.535 0.001 0.006 0
0.005 0.764 0.002 0.015 0.002

Table 3 e Energy conversion efficiencies in hydrogen and methane sequential generation from xylose.
No.a Hydrogen yield(ml/g)
Methane yield(ml/g)
Efficiency inhydrogen-onlyproduction (%)
Efficiency in hydrogenand methane production (%)
1/10 114.1 � 17.2 173.5 � 3.8 8.5 � 1.3 51.2 � 2.2
2/20 165.5 � 7.8 188.9 � 3.2 12.3 � 0.6 58.8 � 1.4
3/30 190.6 � 1.4 216.5 � 5.7 14.2 � 0.1 67.5 � 1.5
4/40 124.8 � 0.2 172.6 � 9.9 9.3 � 0.01 51.8 � 2.5
5/50 177.8 � 2.2 186.7 � 15.7 13.2 � 0.2 59.2 � 4.0
a Conditions of group 1/10e5/50 are the same as Table 1.
0 1 2 3 4 5 6 7 8 9 10 11 120
50
100
150
200
250
Met
hane
yie
ld (m
l/g)
Time (d)
1' 2' 3' 4' 5'
Fig. 4 e Methane yield from residual hydrogen-producing
solutions of xylose (10: residual of condition 1, xylose
feedstock [ 1%, inoculum domesticated with glucose, no
nitrogen source; 20: residual of condition 2, xylose
feedstock [ 1%, inoculum domesticated with xylose, no
nitrogen source; 30: residual of condition 3, xylose
feedstock [ 1%, inoculum domesticated with xylose, 0.5 g
yeast extract; 40: residual of condition 4, xylose
feedstock [ 2%, inoculum domesticated with xylose, no
nitrogen source; 50: residual of condition 5, xylose
feedstock [ 2%, inoculum domesticated with xylose, 0.5 g
yeast extract.).
i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 7 ( 2 0 1 2 ) 1 3 3 2 3e1 3 3 2 9 13327
nutrition for the growth of the hydrogenogens during
fermentation, resulting in the improvement of hydrogen
production. On one hand, hydrogenogens keep growing with
yeast extract and maintain high metabolic activities; on the
other hand, the growth of hydrogenogens ensures high quan-
tity of hydrogenogenic cells, which is favorable for hydrogen
production.
In this research, the influence of xylose concentration on
hydrogen production was also investigated. When the xylose
concentration (w/v) is increased from 1% to 2%, the maximum
hydrogenproduction rate ispromoted from48.4�1.4ml/(L$h) to
60.0 � 7.0 ml/(L$h), but the hydrogen yield is decreased from
165.5� 7.8 ml/g to 124.8� 0.2 ml/g without yeast extract (group
4); whilewith yeast extract themaximumhydrogen production
rate is greatly promoted from 59.6 � 6.6 ml/(L$h) to 135.3 � 5.5
ml/(L$h),but thehydrogenyield isdecreasedfrom190.6�1.4ml/
g to 177.8 � 2.2 ml/g (group 5). This result may be due to the
excessive xylose concentration resulting in an accumulation of
byproductssuchasVFAsandanincrease inthepartialhydrogen
pressure, in which case the microflora would inhibit hydrogen
production [36]. Thus, a xylose concentration of 1% (w/v) gives
the maximum hydrogen yield, but a higher concentration may
bemore promising to give a higher hydrogen rate.
3.3. Methane production from residual hydrogen-producing solutions
The SMB of hydrogen production from xylose (such as
ethanol, acetate, propionate, butyrate, and valerate) can be
reutilized by the methanogen community to further produce
methane [20]. As shown in Table 2, SMB in residual solutions
after the first hydrogen-producing stage mainly include
acetate and butyrate (more than 90% of the total SMB), and
their concentrations are proportional to the cumulative
hydrogen production (ml) in the first stage. The results imply
that the hydrogen production is through acetate- and
butyrate-type fermentation. In the second stage, metha-
nogens can directly convert acetate to methane through ace-
toclastic pathway. Methanogens split acetate into a methyl
group and an enzyme-bound CO, with the CO subsequently
oxidized to provide electrons for the reduction of the methyl
group to methane [37]. Butyrate and other byproducts can be
converted into acetate by the mixed bacteria, and then
utilized by the methanogens to produce methane. The results
show that after the second stage, over 98 wt % of acetate and
butyrate in residual SMB of the first stage are reutilized by the
methanogen community to produce methane.
In the second stage, the cumulative methane production
(ml) is proportional to the content of SMB produced in the first
stage. The methane yields are shown in Fig. 4. The maximum
methane yield of 216.5 � 5.7 ml/g is obtained when the xylose
feedstock concentration is 1% (w/v), hydrogenogens are
domesticated with xylose, and yeast extract is used as
nitrogen source in the first stage.
3.4. Energy conversion efficiency
The energy conversion efficiency of cogenerating hydrogen and
methane from xylose is calculated based on the heating values
of hydrogen, methane, and raw xylose. The heating value of
xylose is 14.525 kJ/g, which is measured by combustion calo-
rimetry. The heating values of hydrogen and methane are
242 kJ/mol and 801 kJ/mol, respectively. The maximum energy
conversion efficiency of 67.5% (hydrogen yield: 190.6 ml/g;
methane yield: 216.5 ml/g) was obtained (Table 3). A

Table 4 e A comparison of energy conversion efficiencies from xylose in the present study and those reported in relevantstudies.
Microorganism Fermentation type Fermentationtemperature (�C)
Energy conversionefficiency (%)
References
Granulated sludge UASB, hydrogen fermentation 37 8.9 [15]
sewage sludge CSTR, hydrogen fermentation 50 15.5 [16]
Clostridium sp. strain No. 2 Batch, hydrogen fermentation 35 22.9 [17]
Sewage sludge Batch, hydrogen fermentation 35 25.0 [18]
Thermotoga neapolitana
strain DSM 4359
Batch, hydrogen fermentation 75 31.1 [19]
Activated sludge Batch, hydrogen fermentation
and methanogenesis
37 67.5 In the present
study
CSTR, continuously stirred tank reactor.
UASB, upflow anaerobic sludge blanket.
i n t e rn a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 7 ( 2 0 1 2 ) 1 3 3 2 3e1 3 3 2 913328
comparison of energy conversion efficiencies fromxylose in the
present study and those reported in relevant studies is shown
in Table 4. Energy conversion efficiencies ranged from 8.9% to
31.1% through hydrogen fermentation in the literature [15e19].
In the present study, the energy conversion efficiency of 67.5%
through sequential generation of hydrogen and methane was
much higher than those obtained in previous literature.
Up to now, no research about cogenerating hydrogen and
methane from xylose has been reported. Xie et al. obtained
a hydrogen yield of 342.2ml/g and amethane yield of 265.1ml/
g from glucose (energy conversion efficiency: 82.0%) [14].
Cheng et al. obtained a hydrogen yield of 51.7 ml/g and
a methane yield of 143.4 ml/g TVS from water hyacinth
(energy conversion efficiency: 33.2%) [8]. Cheng et al. obtained
a hydrogen yield of 58.0ml/g and amethane yield of 200.9ml/g
TVS from corn straw (energy conversion efficiency: 67.1%) [38].
The energy conversion efficiency of xylose through hydrogen
andmethane cogeneration was lower than that of glucose but
higher than that of cellulosic biomass. This result may be
attributed to two reasons: (1) hexose is more favorable for
bacterial utilization than pentose; (2) some hydrolysis
byproducts from cellulosic biomass inhibit the hydrogen and
methane fermentation.
4. Conclusions
This study demonstrates the feasibility of cogenerating
hydrogenandmethane fromxylose (pentose),which isa typical
hydrolysis product of hemicellulose in biomass, by two-stage
anerobic fermentation. In the first hydrogen production stage,
the maximum hydrogen yield is 190.6 ml H2/g xylose. The SMB
from the hydrogen-producing stage were reutilized by
methanogens to produce methane in the second stage. The
maximum methane yield is 216.5 ml CH4/g xylose. The
sequential generation of hydrogen and methane from xylose
markedly increases the energy conversion efficiency to 67.5%.
Acknowledgments
This study is supportedbyNationalNatural Science Foundation
of China (51176163), International Sci. & Tech. Cooperation
Program of China (2010DFA72730, 2012DFG61770), National
High Technology R&D Program of China (2012AA050101),
National Key Technology R&D Program of China
(2011BAD14B02), Specialized Research Fund for the Doctoral
Program of Higher Education (20110101110021), Program for
New Century Excellent Talents in University (NCET-11-0446),
Key Natural Science Foundation of Zhejiang Province
(Z1090532), Major Sci. & Tech. Special Project of Zhejiang Prov-
ince (2008C13023-3), Fundamental Research Funds for the
Central Universities (2011XZZX007), and Program of Intro-
ducing Talents of Discipline to University (B08026).
r e f e r e n c e s
[1] Lin CY, Wu CC, Wu JH, Chang FY. Effect of cultivationtemperature on fermentative hydrogen production fromxylose by a mixed culture. Biomass and Bioenergy 2008;32(12):1109e15.
[2] Liu Y, Yu P, Song X, Qu YB. Hydrogen production fromcellulose by co-culture of Clostridium thermocellum JN4 andThermoanaerobacterium thermosaccharolyticum GD17.International Journal of Hydrogen Energy 2008;33(12):2927e33.
[3] Logan BE, Oh SE, Kim IS, Van Ginkel S. Biological hydrogenproduction measured in batch anaerobic respirometers.Environmental Science & Technology 2002;36(11):2530e5.
[4] Yokoi H, Maki R, Hirose J, Hayashi S. Microbial production ofhydrogen from starch-manufacturing wastes. Biomass andBioenergy 2002;22(5):389e95.
[5] Yu HQ, Zhu ZH, Hu WR, Zhang HS. Hydrogen productionfrom rice winery wastewater in an upflow anaerobic reactorby using mixed anaerobic cultures. International Journal ofHydrogen Energy 2002;27(11e12):1359e65.
[6] Bi Y, Gao C, Wang Y, Li B. Estimation of straw resources inChina. Nongye Gongcheng Xuebao/Transactions of theChinese Society of Agricultural Engineering 2009;25(12):211e7.
[7] Ren YL, Wang JJ, Liu Z, Ren YL, Li GZ. Hydrogen productionfrom the monomeric sugars hydrolyzed from hemicelluloseby Enterobacter aerogenes. Renew Energ 2009;34(12):2774e9.
[8] Cheng J, Xie B, Zhou J, SongW, Cen K. Cogeneration of H2 andCH4 from water hyacinth by two-step anaerobicfermentation. International Journal of Hydrogen Energy 2010;35(7):3029e35.
[9] Mechichi T, Labat M, Garcia JL, Thomas P, Patel BKC.Characterization of a new xylanolytic bacterium, Clostridiumxylanovorans sp nov. Syst Appl Microbiol 1999;22(3):366e71.

i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 7 ( 2 0 1 2 ) 1 3 3 2 3e1 3 3 2 9 13329
[10] Zhu S, Wu Y, Yu Z, Chen Q, Wu G, Yu F, et al. Microwave-assisted alkali pre-treatment of wheat straw and itsEnzymatic hydrolysis. Biosystems Engineering 2006;94(3):437e42.
[11] Zhu S, Wu Y, Yu Z, Liao J, Zhang Y. Pretreatment bymicrowave/alkali of rice straw and its enzymic hydrolysis.Process Biochemistry 2005;40(9):3082e6.
[12] Li Z, Wang H, Tang Z, Wang X, Bai J. Effects of pH value andsubstrate concentration on hydrogen production from theanaerobic fermentation of glucose. International Journal ofHydrogen Energy 2008;33(24):7413e8.
[13] Mitchell RJ, Kim J-S, Jeon B-S, Sang B-I. Continuous hydrogenand butyric acid fermentation by immobilized Clostridiumtyrobutyricum ATCC 25755: effects of the glucoseconcentration and hydraulic retention time. BioresourceTechnology 2009;100(21):5352e5.
[14] Xie B, Cheng J, Zhou J, Song W, Cen K. Cogeneration ofhydrogen and methane from glucose to improve energyconversion efficiency. International Journal of HydrogenEnergy 2008;33(19):5006e11.
[15] Maintinguer SI, Fernandes BS, Duarte ICS, Saavedra NK,Adorno MAT, Varesche MBA. Fermentative hydrogenproduction with xylose by Clostridium and Klebsiella speciesin anaerobic batch reactors. International Journal ofHydrogen Energy 2011;36(21):13508e17.
[16] Lin C-Y, Wu C-C, Hung C-H. Temperature effects onfermentative hydrogen production from xylose using mixedanaerobic cultures. International Journal of Hydrogen Energy2008;33(1):43e50.
[17] Taguchi F, Mizukami N, Taki TS, Hasegawa K. Hydrogen-production from continuous fermentation of xylose duringgrowth of clostridium sp strain No-2. Canadian Journal ofMicrobiology 1995;41(6):536e40.
[18] Lin CY, Cheng CH. Fermentative hydrogen production fromxylose using anaerobic mixed microflora. InternationalJournal of Hydrogen Energy 2006;31(7):832e40.
[19] Tien Anh N, Tra Huong N, Ha Thi Viet B. Thermophilicfermentative hydrogen production from xylose byThermotoga neapolitana DSM 4359. Renewable Energy 2012;37(1):174e9.
[20] Song W, Cheng J, Zhou J, Xie B, Su H, Cen K. Cogeneration ofhydrogen and methane from protein-mixed food waste bytwo-phase anaerobic process. International Journal ofHydrogen Energy 2010;35:3141e6.
[21] Xie B, Cheng J, Zhou J, Song W, Liu J, Cen K. Production ofhydrogen and methane from potatoes by two-phaseanaerobic fermentation. Bioresource Technology 2008;99(13):5942e6.
[22] Schmidt TM, Delong EF, Pace NR. Analysis of a marinepicoplankton community by 16S rRNA gene cloning andsequencing. Journal of Bacteriology 1991;173(14):4371e8.
[23] Li X, Du B, Fu H, Wang R, Shi J, Wang Y, et al. The bacterialdiversity in an anaerobic ammonium-oxidizing (anammox)reactor community. Systematic and Applied Microbiology2009;32(4):278e89.
[24] Li X, Ma E, Yan L, Meng H, Du X, Zhang S, et al. Bacterial andfungal diversity in the traditional Chinese liquor
fermentation process. International Journal of FoodMicrobiology 2011;146(1):31e7.
[25] Lin PY, Whang LM, Wu YR, Ren WJ, Hsiao CJ, Li SL, et al.Biological hydrogen production of the genus Clostridium:metabolic study and mathematical model simulation.International Journal of Hydrogen Energy 2007;32(12):1728e35.
[26] Murty MVS, Chandra TS. Isolation and characterization ofxylose-utilizing and xylan-utilizing anaerobic-bacteria.Antonie Van Leeuwenhoek Journal of Microbiology 1989;55(2):153e63.
[27] Schnurer A, Schink B, Svensson BH. Clostridium ultunensesp. nov., a Mesophilic bacterium oxidizing acetate insyntrophic association with a hydrogenotrophicmethanogenic bacterium. International Journal ofSystematic Bacteriology 1996;46(4):1145e52.
[28] Koeppel A, Perry EB, Sikorski J, Krizanc D, Warner A,Ward DM, et al. Identifying the fundamental units ofbacterial diversity: a paradigm shift to incorporate ecologyinto bacterial systematics. Proceedings of the NationalAcademy of Sciences U S A 2008;105(7):2504e9.
[29] Fond O, Engasser JM, Mattaelamouri G, Petitdemange H. Theacetone butanol fermentation on glucose and xylose .1.regulation and kinetics in batch cultures. Biotechnology andBioengineering 1986;28(2):160e6.
[30] Gu Y, Ding Y, Ren C, Sun Z, Rodionov DA, Zhang WW, et al.Reconstruction of xylose utilization pathway and regulons infirmicutes. BMC Genomics 2010;11:14.
[31] Ounine K, Petitdemange H, Raval G, Gay R. Regulation andbutanol inhibition OF D-XYLOSE and D-GLUCOSE uptake inClostridium-Acetobutylicum. Applied and EnvironmentalMicrobiology 1985;49(4):874e8.
[32] Lo YC, Chen WM, Hung CH, Chen SD, Chang JS. Dark H-2fermentation from sucrose and xylose using H-2-producingindigenous bacteria: feasibility and kinetic studies. WaterRes 2008;42(4e5):827e42.
[33] Schmiedel D, Hillen W. A Bacillus subtilis 168 mutant withincreased xylose uptake can utilize xylose as sole carbonsource. FEMS Microbiology Letters 1996;135(2e3):175e8.
[34] Dutta T, Das AK, Das D. Purification and characterization of[Fe]-hydrogenase from high yielding hydrogen-producingstrain, Enterobacter cloacae IIT-BT08 (MTCC 5373).International Journal of Hydrogen Energy 2009;34(17):7530e7.
[35] Patel GB. Ethanol production during D-xylose, L-arabinose,and D-ribose fermentation by bacteroides polypragmatus.Applied Microbiology and Biotechnology 1984;20(2):111e7.
[36] Lin CY, Lay CH. Carbon/nitrogen-ratio effect on fermentativehydrogen production by mixed microflora. InternationalJournal of Hydrogen Energy 2004;29(1):41e5.
[37] Galagan JE, Nusbaum C, Roy A, Endrizzi MG, Macdonald P,FitzHugh W, et al. The genome of M-acetivorans revealsextensive metabolic and physiological diversity. GenomeResearch 2002;12(4):532e42.
[38] Cheng X-Y, Li Q, Liu C-Z. Coproduction of hydrogen andmethane via anaerobic fermentation of cornstalk waste incontinuous stirred tank reactor integrated with up-flowanaerobicsludgebed.BioresourceTechnology2012;114:327e33.