selenium and mercury concentrations in harbor seals (phoca vitulina) from central california: health...
TRANSCRIPT

Marine Pollution Bulletin xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Selenium and mercury concentrations in harbor seals (Phoca vitulina)from central California: Health implications in an urbanized estuary
http://dx.doi.org/10.1016/j.marpolbul.2014.04.0310025-326X/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author. Present address: University of California Santa Cruz,Long Marine Laboratory, 100 Shaffer Road, Santa Cruz, CA 95060, USA.Tel.: +1 831 459 2691; fax: +1 831 459 3383.
E-mail address: [email protected] (E.A. McHuron).
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury concentrations in harbor seals (Phoca vitulina) from central California:implications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marpolbul.2014.04.031
Elizabeth A. McHuron a,⇑, James T. Harvey a, J. Margaret Castellini b, Craig A. Stricker c, Todd M. O’Hara d
a Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, CA 95039, USAb Institute of Marine Science, School of Fisheries and Ocean Sciences, University of Alaska Fairbanks, P.O. Box 757220, Fairbanks, AK 99775-7220, USAc U.S. Geological Survey, Fort Collins Science Center, Denver Federal Center, Building 21, MS963, Denver, CO 80225, USAd Department of Veterinary Medicine, University of Alaska Fairbanks, P.O. Box 755940, Fairbanks, AK 99775-7000, USA
a r t i c l e i n f o
Keywords:Trace elementsMarine mammalsHairBloodSan Francisco BayStable isotopes (C, N, S)
a b s t r a c t
We measured total selenium and total mercury concentrations ([TSe] and [THg]) in hair (n = 138) andblood (n = 73) of harbor seals (Phoca vitulina) from California to assess variation by geography and sex,and inferred feeding relationships based on carbon, nitrogen, and sulfur stable isotopes. Harbor sealsfrom Hg-contaminated sites had significantly greater [THg], and lesser [TSe] and TSe:THg molar ratiosthan seals from a relatively uncontaminated site. Males had significantly greater [THg] than females atall locations. Sulfur stable isotope values explained approximately 25% of the variability in [THg], indicat-ing increased Hg exposure for seals with a greater use of estuarine prey species. Decreased [TSe] in harborseals from Hg-contaminated regions may indicate a relative Se deficiency to mitigate the toxic effects ofHg. Further investigation into the Se status and the potential negative impact of Hg on harbor seals fromHg-contaminated sites is warranted.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction 40 years (Grigg et al., 2004), which is in contrast to recoveries of
Marine mammals are considered sentinels of marine ecosystemhealth because they are long-lived, have substantial lipid storesthat concentrate lipophilic contaminants, and many are upper tro-phic level consumers (Pauly et al., 1998; Ross, 2000; Wells et al.,2004). Consequently, many toxicants bioaccumulate and biomag-nify in marine mammals, which may have negative impacts onhealth and population dynamics (Reijnders, 1986; Ross et al.,1996; Ross, 2002; Barron et al., 2003). Species found in urbanizedcoastal areas may be especially susceptible to accumulating con-centrations of toxicants as a result of natural and anthropogenicinputs into nearshore ecosystems.
Harbor seals (Phoca vitulina) are a small, non-migratory phocidfound in coastal areas throughout the northern hemisphere. InCalifornia, harbor seals are distributed along the mainland coast,islands, and bays (Lowry et al., 2008), and are present year-roundin San Francisco Bay (SFB), an urbanized estuary that providescritical habitat for resting ashore (hauling-out), reproduction,social interaction, and feeding (Torok, 1994; Grigg et al., 2004).The number of harbor seals in SFB has not increased in the last
harbor seal populations along the west coast of the United Statesfollowing the passage of the Marine Mammal Protection Act in1972 (Harvey et al., 1990; Jeffries et al., 2003; Brown et al., 2005;Lowry et al., 2008). Hypotheses for the apparent lack of recoveryare primarily focused on the urbanized and industrialized natureof SFB, and include habitat alteration, disturbance, and environ-mental contamination (Grigg et al., 2004; Neale et al., 2005).
Mercury (Hg) is a primary toxicant of concern in SFB that mayhave a negative effect on the reproductive, immunological, andneurological systems of wildlife and humans (Wolfe et al., 1998;Zahir et al., 2005). Because some forms of Hg biomagnify in foodwebs (Dietz et al., 2000b; Campbell et al., 2005) and most of theHg present in fish is in a relatively toxic form (monomethyl Hg;MeHg+; Harris et al., 2003), upper trophic level consumers maybe most impacted by environmental contamination of Hg. Marinemammals may be partially protected against the negative effectsof Hg through a number of mechanisms, including demethylationof Hg, excretion of Hg (e.g., urine, feces, hair), the presence oforganic components such as noted for humans (e.g., polyunsatu-rated fatty acids; Guallar et al., 2002), or interactions with otherelements (Cuvin-Aralar and Furness, 1991; Yang et al., 2008).Selenium (Se), an essential element, is believed to mitigate orameliorate the toxic effects of Hg through a number of directbinding and antioxidant mechanisms, including the formation of
Health

2 E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
a non-toxic Se-Hg complex (Yoneda and Suzuki, 1997; Ikemotoet al., 2004a; Yang et al., 2008; Frouin et al., 2012; Lailson-Britoet al., 2012). Most studies on Se and Hg in marine mammals haveemphasized the beneficial nature of Se, likely because marinemammals tend to accumulate relatively high concentrations ofHg. Selenium, however, can have a negative impact on wildlifehealth at increased concentrations (Hamilton, 2004). Symptomsassociated with Se toxicosis in birds and mammals are consistentamong species and include emaciation, changes to keratinized tis-sues, dyspnea, and death (Yang et al., 1983; Raisbeck et al., 1993;O’Toole and Raisbeck, 1995, 1997). Selenium toxicosis was impli-cated in the deaths of three captive California sea lions (Zalophuscalifornianus; Edwards and Whitenack, 1989), but has not beendefinitively documented in captive or free-ranging marine mam-mals, and is dependent on the form of Se present.
San Francisco Bay has a history of Se and Hg contamination, andconcentrations of concern to wildlife and human health for bothelements have been noted in resident fish and birds (Ohlendorfet al., 1986; Hothem et al., 1998; Stewart et al., 2004; Greenfieldet al., 2005; Schwarzbach et al., 2006; Ackerman et al., 2007).Previous studies on the concentrations of these elements in SFBharbor seals found no significant differences in total Hg concentra-tions ([THg]) among harbor seals in central and northern California(Brookens et al., 2007), but did report that Se concentrations ([Se])in blood of seals from SFB (n = 55) were greater than seals fromPuget Sound (n = 6), Monterey Bay (n = 2), and San Nicolas Island(n = 3; Kopec and Harvey, 1995). This, coupled with the observa-tion that some seals in SFB had hair loss and shortened vibrissae,led Kopec and Harvey (1995) to hypothesize that a proportion ofharbor seals in SFB may suffer from chronic Se toxicosis. Despitethe detection of geographical differences in [Se], the Kopec andHarvey (1995) study suffered from small and unequal sample sizes,seasonal differences in sample collection that may have biasedresults, and lack of consideration of the Se:Hg molar ratio, whichmay be a better indicator of toxicity for some tissues (Ralstonet al., 2008). Seals in SFB continue to display hair loss and short-ened vibrissae (McHuron, 2012), and given that the number of har-bor seals in SFB has not increased following federal protection,evaluation of the Se status of SFB harbor seals is warranted.
The objectives of this study were to provide a more detailed andrecent assessment of total [Se] ([TSe]) in harbor seals from threecentral California locations, and to investigate the relationshipbetween [TSe] and [THg] in these seals. We evaluated whether[TSe], [THg], and the TSe:THg molar ratio differed with locationand sex, and determined the relationship between these elementsand chemical measures of feeding ecology (nitrogen (N), carbon(C), and sulfur (S) stable isotopes). We also examined the relation-ships between [TSe] and [THg] in blood and hair (e.g., THgblood vs.THghair, TSe vs. THg). We hypothesized that [TSe], [THg], and theTSe:THg molar ratio in hair and blood would be greater in sealsfrom SFB than the other two locations. We also hypothesized thatdifferences in [TSe] and [THg] with sex would only exist for [THg](males > females), that there would be a positive relationshipbetween [THg] in hair and blood, and that [THg] would increasewith d15N values (trophic level). Because Se is an essential elementthat is homeostatically regulated, we predicted that there wouldnot be a clear relationship between [TSe] in the two tissue types,nor between [TSe] and chemical measures of feeding ecology.
Fig. 1. Location and number of harbor seals sampled (denoted by black dots sizedrelative to sample size) in central California between August 2009 and February2011. Sample sizes are shown in parentheses following site names.
2. Methods
2.1. Sample collection
Adult and juvenile harbor seals (P1 year of age; n = 138) werecaptured in Elkhorn Slough, SFB, and Tomales Bay between August
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
2009 and February 2011 (Fig. 1). We did not capture any seals dur-ing the time period when females were likely to be nursing adependent pup (March to mid May). Seals were captured usingtangle nets, salmon nets, or a modified beach seine (Jeffries et al.,1993). We assumed that capture method had no biases and thatwe collected a random sample of seals. Standard length (±1 cm;SL), girth (±1 cm), mass (±1 kg), sex, and age class were determinedfor all seals sampled. Age class was determined based upon lengthand mass (Bigg, 1969), body condition, and date of capture. Sealswere manually restrained for sample collection, and when avail-able, were sedated with an intravenous injection of diazepam(0.2 mg/kg). Hair samples were collected from the right lateralneck using an Oster� Pro battery-operated shaver with a size 40cryotech stainless steel blade that had been cleaned in acetone.Samples were stored frozen at �80 �C in polyethylene bags. Wholeblood samples were collected from the extradural intravertebralsinus into trace element clean BD Vacutainer� tubes containingan anticoagulant (K2EDTA). Blood samples were kept on ice untilstored in 20 mL trace metal clean I-Chem™ vials at �80 �C.
Hair was chosen as the primary target tissue for Se and Hg anal-ysis because collection is minimally invasive, it may serve as anexcretory route for excess elements (Wenzel et al., 1993; Saekiet al., 1999; Ikemoto et al., 2004b), and concentrations can repre-sent circulating blood values at the time of hair growth (molt).Because molt in harbor seals occurs over a relatively short timespan (approximately one to two months; Thompson and Rothery,1987; Daniel et al., 2003) and at approximately the same timefor seals in central California, concentrations in hair collected out-side of the molt period are not likely influenced by sampling dateand can be easily compared among locations. Whole blood samples(hereafter referred to as blood) were analyzed from a subset of
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031

E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 3
seals (SFB and Elkhorn Slough; n = 73) to determine the relation-ship between blood and hair concentrations for both elements,and also as an additional matrix to interpret [TSe] and [THg].
2.2. Sample preparation
Hair samples were cleaned before analysis to remove externaldirt, debris, and oil. Samples were soaked in a 1% solution ofTriton� X-100 for 15 min (Airey, 1983), followed by multiple rinseswith ultrapure water (NANOpure Model D4751, Barnstead Interna-tional, Dubuque, Iowa). Hair samples were then frozen and freeze-dried for a minimum of 24 h to remove water before weighing.Samples were analyzed for [TSe] and [THg] in the Wildlife Toxicol-ogy Laboratory at the University of Alaska Fairbanks, and for stableisotopes at the US Geological Survey Stable Isotope Laboratory inDenver, Colorado.
2.3. Selenium analysis
Samples for Se analysis were prepared using a two-step diges-tion in an Anton Paar Multiwave 3000 microwave oven. Approxi-mately 0.1 g of hair and 0.6 g of blood underwent a primarydigestion in 3:1 nitric acid (HNO3; 70%): hydrogen peroxide(H2O2; 30%; v/v), followed by a subsequent digestion in excesshydrochloric acid (HCl; 34–37%; 1:1, v/v) to reduce Se (VI) to Se(IV) as in Knott et al. (2011). Blanks, spikes, duplicates, matrixspikes, and standard reference materials were included in eachdigestion batch for quality control, and a check standard wasanalyzed at the end of each run. Standard reference materialsincluded human hair NIES 13 (1.79 ± 0.17 mg Se/kg, NationalInstitute for Environmental Studies, Japan), dogfish liver DOLT-4(8.3 ± 1.3 mg Se/kg, National Research Council Canada, Institutefor National Measurement Standards, Ottawa, Canada), and wholeblood Seronorm™ (0.260 ± 0.050 mg Se/kg, Sero, Billingstad,Norway). Blank and matrix spikes (Perkin–Elmer, Waltham,Massachusetts) contained 10.0 mg Se/kg.
Selenium concentrations were determined by mercury/hydridesystem-flame ionization atomic spectrometry (MHS-FIAS) on aPerkinElmer AAnalyst 800 atomic absorption spectrometer (AAS)and a six-, seven-, or eight-point calibration curve (0.04–9.8 ng/g). The carrier solution was 10% HCl, and the reductant was sodiumborohydride (0.2% NaBH4 in 0.05% NaOH). The minimum detectionlimit for 0.1 g of hair ranged from 0.084 to 0.21 mg Se/kg. The min-imum detection limit for 0.6 g of blood ranged from 0.076 to0.098 mg Se/kg. The mean percent recoveries (±SD) for hair were:90 ± 8% (blank spikes), 88 ± 12% (matrix spikes), 92 ± 16% (NIES-13), and 91 ± 9% (DOLT-4). The mean percent recoveries for bloodwere: 90 ± 3% (blank spikes), 83 ± 5% (matrix spikes), 86 ± 5%(DOLT-4), and 88 ± 34% (Seronorm™). The mean coefficients of var-iation (CV ± SD) for duplicates were 22 ± 11 (hair) and 9 ± 12(blood). Molar concentrations of Se were calculated by converting[Se] from lg to g and then dividing by the atomic weight of Se(78.96 g/mol).
2.4. Mercury analysis
Total Hg concentrations were measured in approximately 0.01 gof hair and 0.1 g of blood on a DMA-80 Direct Mercury Analyzer(Milestone Inc., Shelton, Connecticut; Castellini et al., 2012). Sam-ples were analyzed in duplicate or triplicate, depending on theamount of sample available, and the mean [THg] was calculatedfor each individual. Quality controls (blanks, liquid standards,and standard reference materials) were analyzed in triplicate atthe beginning of each batch of samples. The liquid standard con-tained 1.0 mg THg/kg (Perkin–Elmer, Waltham, Massachusetts).Standard reference materials included human hair IAEA-086
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
(0.573 mg THg/kg; International Atomic Energy Agency, Vienna,Austria), human hair IAEA-085 (23.2 mg THg/kg), and humanwhole blood (Seronorm™ Trace Elements Whole Blood L-3,0.037 ± 0.0017 mg THg/kg, Sero, Billingstad, Norway). Total Hgconcentrations were determined using a 2-cell, 16-point calibra-tion curve (30.1–452.9 ng). Samples were re-analyzed for sealswith mean [THg]hair greater than 40 lg/g to ensure measurementaccuracy. The mean percent recoveries (±SD) for hair were104 ± 3% (liquid standard) and 101 ± 7% (IAEA-085). The mean per-cent recoveries for blood were 97 ± 2% (liquid standard), 96 ± 5%(IAEA-086), 98 ± 3% (IAEA-085), and 98 ± 5% (Seronorm™). Themean CVs (±SD) for duplicates were 4 ± 3 (hair) and 2 ± 2 (blood).Total Hg concentrations reported are representative of the MeHg+
concentration in each tissue because greater than 80% of Hg pres-ent in hair (Dolbec et al., 2001; Dietz et al., 2011) and blood(Woshner et al., 2008) is in the monomethylated form. Molar con-centrations of THg were calculated by converting [THg] from lg tog and then dividing by the atomic weight of Hg (200.59 g/mol).
2.5. Stable isotope analysis
Hair samples from seals captured in SFB (n = 50) were analyzedfor N, C, and S stable isotopes. Approximately 1.0 mg of sample wasfolded into ultra-light weight tin foil squares (37 � 37 mm; Ssamples were amended with �1.0 mg of V2O5), and analyzed byconventional continuous-flow isotope ratio mass spectrometryusing an elemental analyzer coupled to a mass spectrometer (Fryet al., 1992; Kester et al., 2001). The ratio of stable isotopes isexpressed in delta (d) notation and calculated as:
dX ¼ ðRsample=RstandardÞ � 1� �
� 1000
where X = 15N, 13C, or 34S, and R = 15N/14N, 13C/12C, or 34S/32S in thesample and standard. Primary standards used to normalize isotopedata were: USGS 40 (d13C = �26.24‰ Vienna-Pee Dee Belemniteand d15N = �4.52‰ with respect to air) and USGS 41 (d13C =37.76‰ and 47.57‰), and NBS127 (d34S = 21.1‰ relative toVienna-Canyon Diablo troilite) and IAEA-SO-6 (d34S = �34.05‰).Nearly all of the samples analyzed for N and C were done in dupli-cate, whereas approximately 50% were analyzed in duplicate for S;isotope values were generally within 0.2‰. Accuracy and precisionwere assessed from primary and secondary laboratory standards,and results were generally better than 0.1‰ for nitrogen and car-bon, and 0.3‰ for sulfur.
2.6. Statistical analysis
All statistical analyses were conducted using R version 3.0.1 (RCore Team 2013, http://www.R-project.org/.). Harbor seals fromTomales Bay were excluded from statistical analyses of [TSe]hair
because of poor recoveries, and eight seals were excluded fromd34S statistical analyses because of poor S yield. Two seals wererecaptured twice during the course of this study: one seal twicein the same molt year, and one seal in successive molt years. Weexcluded samples collected from the second capture of both sealsfrom statistical analyses, although for the seal captured in succes-sive years we also conducted the analyses using samples from thesecond capture to ensure that the inclusion of one year over theother did not change our results.
Hair and blood [TSe], [THg], and the TSe:THg molar ratio wereanalyzed using either Analysis of Variance (ANOVA) or Analysisof Covariance (ANCOVA). Samples within locations were pooledbecause independent sample t-tests indicated that there were nodifferences in concentrations between molt years (2009 and2010). Fixed factors included in analyses were location (ElkhornSlough, SFB, and Tomales Bay [only [THg]hair]) and sex, and
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031

4 E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
covariates considered were standard length (SL) and month (bloodonly). Standard length was considered as a covariate to remove anyvariation in [TSe] and/or [THg] associated with age, and monthincluded because blood concentrations of some elements vary withseason (summer vs. winter; Kopec and Harvey, 1995). Ultimately,SL was only significant as a covariate in one analysis ([THg]hair).Month did explain a significant amount of variability in [THg]blood,but was not included as a covariate because of a significant inter-action with location. The exclusion of month as a covariate shouldnot have impacted the results given that the majority of samplesfrom both locations were collected in similar months with minimalsampling in summer months (June and August; n = 16). Data werelog transformed to meet the assumptions of normality and homo-scedasticity, and post hoc comparisons for [THg]hair were madeusing a Ryan’s Q test with a Kramer modification to correct forunequal sample sizes (Day and Quinn, 1989). Means and rangesare presented using untransformed data.
Linear regression was used to determine whether hair wasrepresentative of blood concentrations for each element, and Pear-son’s correlations were used to examine the relationships between[TSe] and [THg] in each tissue type. Total Hg concentrations werelog transformed to meet the assumption of homoscedasticity forthe regression [THg]blood vs. [THg]hair. Stepwise backwards linearregressions were used to determine whether any of the variabilityin [TSe] and [THg] in hair could be explained by d15N, d13C, or d34Svalues, with SL included as an additional predictor variable.
3. Results
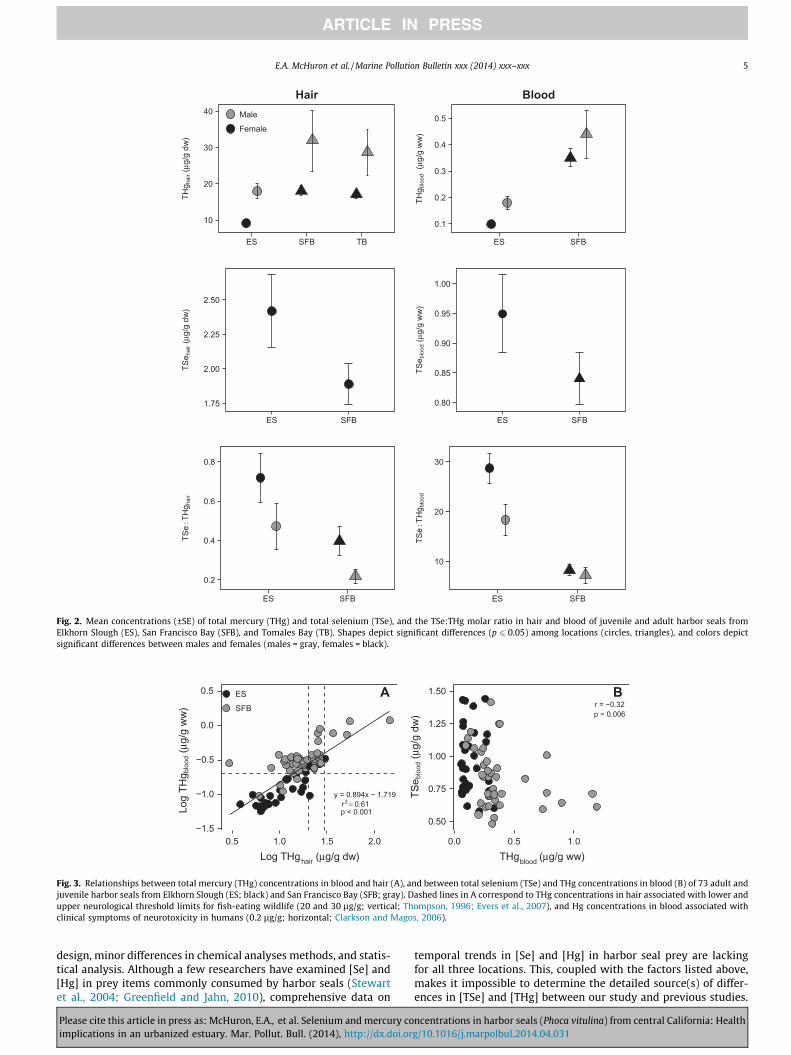
Total selenium concentrations ranged from 0.60 to 6.74 lg/gdry wt in hair, and 0.48 to 1.44 lg/g wet wt in blood (Supplemen-tary Tables 1 and 2). We have not presented [TSe]hair from TomalesBay harbor seals because of poor recoveries in runs that includedthese samples. Harbor seals from SFB had lesser mean [TSe]hair
and [TSe]blood than seals from Elkhorn Slough, although these dif-ferences were only statistically different for blood (hair, F = 2.34,p = 0.13; blood, F = 6.94, p = 0.010; Fig. 2). There were no differ-ences in [TSe] between males and females for either tissue type(hair, F = 0.94, p = 0.33; blood, F = 1.45, p = 0.23; Fig. 2). Therewas no relationship between [TSe]blood and [TSe]hair (r2 = 0.044,p = 0.089).
Total Hg concentrations ranged from 2.96 to 144.31 lg/g dry wtin hair, and 0.057 to 1.19 lg/g wet wt in blood (SupplementaryTables 1 and 2). Harbor seals from SFB and Tomales Bay hadgreater [THg]hair than seals from Elkhorn Slough (F = 13.59,p < 0.001; Fig. 2), and seals from SFB had greater [THg]blood thanseals from Elkhorn Slough (F = 74.84, p < 0.001; Table 2). Males atall locations had greater [THg]hair and [THg]blood than females (hair,F = 10.17, p = 0.002; blood, F = 5.77, p = 0.019; Fig. 2). There was arelatively strong positive relationship between [THg]blood and[THg]hair (r2 = 0.61, p < 0.001; Fig. 3A). Total Hg concentrations insome seals, primarily those from SFB, exceeded concentrationsassociated with neurological effects of Hg toxicosis in both tissuetypes (Fig. 3A).
The TSe:THg molar ratio ranged from 0.049 to 2.33 in hair, andfrom 1.31 to 51.74 in blood (Supplementary Tables 1 and 2). Har-bor seals from Elkhorn Slough had greater TSe:THghair andTSe:THgblood molar ratios than seals from SFB (hair, F = 17.42,p < 0.001; blood, F = 62.86, p < 0.001) Females also had greaterTSe:THg ratios in both tissue types than males (hair, F = 5.08,p = 0.027; blood, F = 5.13, p = 0.027; Fig. 2). There was no correla-tion between [TSe] and [THg] in hair (r = 0.06, p = 0.63), and a weaknegative correlation in blood (r = - 0.32, p = 0.006; Fig. 3B).
Stable isotope (C, N, S) values did not explain a significantamount of variability in [TSe]hair in seals from SFB (p = 0.41). For
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
[THg], only S stable isotope values explained variability in [THg]hair,but the relationship had a relatively poor fit (r2 = 0.15, p = 0.017).This relationship improved when two extreme values wereremoved from the analysis (r2 = 0.27, p = 0.001; Fig. 4).
4. Discussion
4.1. TSe and THg in hair and blood
These are the first published values for [TSe] in hair of harborseals from SFB and Tomales Bay, and the first comprehensive studyof Se for harbor seals in central California. The mean [TSe]hair inharbor seals for all locations (2.10 ± 0.14; lg/g dry wt ± SE) wassimilar (mean within ± 10%) to concentrations reported for Baikal(Pusa sibirica; 2.3 ± 0.16; Ikemoto et al., 2004b) and Caspian seals(Pusa caspica; 2.3 ± 0.48; Ikemoto et al., 2004b), but less than con-centrations reported for Weddell seals (Leptonychotes weddellii;3.12 ± 0.27; Gray et al., 2008), leopard seals (Hydrurga leptonyx;4.13 ± 0.68; Gray et al., 2008), and northern fur seals (Callorhinusursinus; 6.1 ± 1.36; Ikemoto et al., 2004b). Blood [TSe] were similarto concentrations measured in harbor seals from central California(Kopec and Harvey, 1995) and the North Sea (Griesel et al., 2008).The poor relationship between [TSe] in blood and hair might havebeen a result of a number of factors, including homeostatic regula-tion of Se in circulation, that hair is not a major pathway for Seexcretion, or because of temporal differences between blood col-lection and hair formation (i.e., molt occurs from late June to earlyAugust, but most seals were captured from October to February).
Harbor seals from central California had greater [THg] thanpublished values for most pinniped species elsewhere (Fig. 5), withthe maximum measured [THg]hair greater than previously reportedfor any pinniped (144.31 lg/g dry wt). The mean [THg]hair of sealsin our study (19.60 ± 1.44; lg/g dry wt ± SE) was relatively similarto mean concentrations in harbor seals from Greenland (16.3 ± 2.9;Aubail et al., 2011), and ringed seals from Lake Lagoda (17.5 ± 4.0;Pusa hispida ladogensis; Medvedev et al., 1997) and Lake Saimaa(20.7 ± 4.1; P. h. saimensis; Hyvärinen and Sipilä, 1984), but lessthan mean concentrations in dead-stranded harbor seals fromnorthern Germany (33.5 ± 5.6; Wenzel et al., 1993). Blood [THg]were greater than concentrations in harbor seals from the NorthSea (Das et al., 2008) and southern elephant seals from Antarctica(Mirounga leonina; Baraj et al., 2001), but less than lactating north-ern elephant seals from central California (M. angustirostris;Habran et al., 2011). Mercury concentrations in hair are generallygreater in harbor (and ringed) seals compared with other pinnipedspecies, which may be a combination of foraging ecology,differences in the global distribution of atmospheric Hg (northernhemisphere > southern hemisphere; Slemr et al., 2003), and differ-ences in levels of environmental contamination. Harbor sealsinhabit nearshore habitats that often are industrialized and/orurbanized, and these habitats, coupled with a diet comprisedprimarily of fish, make harbor seals susceptible to bioaccumulationand biomagnification of Hg. Hair is an important pathway forelimination of Hg in pinnipeds (Wenzel et al., 1993), and resultsfrom this study indicate it is an appropriate tissue to measure Hgexposure in harbor seals as hair concentrations are representativeof blood concentrations.
In general, harbor seals in this study had lesser [TSe] andgreater [THg] than previously reported for central California(Kopec and Harvey, 1995; Moser, 1996; Brookens et al., 2007).These apparent differences could be the result of a multitude offactors, including temporal differences in foraging behavior (preytype, prey size, location) or environmental concentrations of Seand/or Hg, biological variables (e.g., sex), physiological status(e.g., nutritional status), or an artifact of differences in sampling
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031
10
20
30
40
ES SFB TB
THg h
air (
μg/g
dw
)
Male
Female
Hair
0.1
0.2
0.3
0.4
0.5
ES SFB
THg b
lood
(μg
/g w
w)
Blood
1.75
2.00
2.25
2.50
ES SFB
TSe h
air (
μg/g
dw
)
0.80
0.85
0.90
0.95
1.00
ES SFBTS
e blo
od (μ
g/g
ww
)
0.2
0.4
0.6
0.8
ES SFB
TSe
:TH
g hai
r
10
20
30
ES SFB
TSe
:TH
g blo
od
Fig. 2. Mean concentrations (±SE) of total mercury (THg) and total selenium (TSe), and the TSe:THg molar ratio in hair and blood of juvenile and adult harbor seals fromElkhorn Slough (ES), San Francisco Bay (SFB), and Tomales Bay (TB). Shapes depict significant differences (p 6 0.05) among locations (circles, triangles), and colors depictsignificant differences between males and females (males = gray, females = black).
A
y = 0.894x − 1.719r2 = 0.61p < 0.001
−1.5
−1.0
−0.5
0.0
0.5
Log THghair (μg/g dw)
Log
THg b
lood
(μg/
g w
w)
ES
SFB
Br = −0.32p = 0.006
0.50
0.75
1.00
1.25
1.50
0.5 1.0 1.5 2.0 0.0 0.5 1.0THgblood (μg/g ww)
TSe b
lood
(μg/
g dw
)
Fig. 3. Relationships between total mercury (THg) concentrations in blood and hair (A), and between total selenium (TSe) and THg concentrations in blood (B) of 73 adult andjuvenile harbor seals from Elkhorn Slough (ES; black) and San Francisco Bay (SFB; gray). Dashed lines in A correspond to THg concentrations in hair associated with lower andupper neurological threshold limits for fish-eating wildlife (20 and 30 lg/g; vertical; Thompson, 1996; Evers et al., 2007), and Hg concentrations in blood associated withclinical symptoms of neurotoxicity in humans (0.2 lg/g; horizontal; Clarkson and Magos, 2006).
E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 5
design, minor differences in chemical analyses methods, and statis-tical analysis. Although a few researchers have examined [Se] and[Hg] in prey items commonly consumed by harbor seals (Stewartet al., 2004; Greenfield and Jahn, 2010), comprehensive data on
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
temporal trends in [Se] and [Hg] in harbor seal prey are lackingfor all three locations. This, coupled with the factors listed above,makes it impossible to determine the detailed source(s) of differ-ences in [TSe] and [THg] between our study and previous studies.
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031

y = −4.75x + 103.77r2 = 0.27p = 0.001
EstuarineInshoreBenthic
MarineOffshorePelagic
0
20
40
60
15 16 17 18 19
δ34S (‰)
THg
hair (
μg/g
dw
)
Fig. 4. Relationship between total mercury concentrations (THg) and d34S (‰) inhair of juvenile and adult harbor seals from San Francisco Bay. Two seals wereexcluded because of a very high THg concentration (144 lg/g dw) or a very low d34Svalue (12.3‰). Dashed lines correspond to THg concentrations in hair associatedwith lower and upper neurological threshold limits for fish-eating wildlife (20 and30 lg/g; Thompson, 1996; Evers et al., 2007).
6 E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
4.2. Geographical differences
Fine-scale geographical differences in trace element concentra-tions often are difficult to detect due to a variety of factors, includ-ing movement of animals among regions, small sample sizes,location of capture and relationship with feeding area, and vari-ability with sex and age class (Brookens et al., 2007; Aubail et al.,2011). Despite this, we found differences in [TSe] and [THg]between seals that were captured at locations approximately
Fig. 5. Mean total mercury (THg) concentrations in hair of phocids (n = 19; circles), otarmeasured in pups were included in means only if they could not be removed, and none owhen latitude/longitude coordinates were not given, and the location for one Antarcticwhere concentrations were present for multiple species so that shapes did not complete(1984), Bacher (1985), Wenzel et al. (1993), Fossi et al. (1997), Medvedev et al. (1997), YBrookens et al. (2007), Gray et al. (2008), Agusa et al. (2011) and Aubail et al. (2011).
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
200 km apart. The trend towards lesser [TSe]hair and [TSe]blood inharbor seals from SFB was unexpected because of environmentalSe contamination that has resulted in increased [Se] in residentwildlife (Ohlendorf et al., 1986; Hothem et al., 1998; Stewartet al., 2004; Greenfield et al., 2005), and because Se in circulationis homeostatically regulated. Because Se is an essential element,blood and hair concentrations are likely influenced by factors otherthan just dietary intake, thereby making it difficult to make conclu-sions about the causes of these geographical differences. Potentialcauses include differences in Se exposure (mainly via dietaryintake), forms of Se in the diet, differences in the physiologic needfor Se (e.g., oxidative stress, age-dependent), or retention of Sebecause of increased [Hg] or variations in other constituents (haironly). It also is possible that the differences in [TSe]blood betweenSFB and Elkhorn Slough were statistically, but not biologically sig-nificant. The poor relationship between [TSe] in blood and hairindicates that hair is not an ideal matrix for physiologic (circulat-ing) status determination outside of the molt period, although hairmay still be a useful matrix for detecting geographical differencesin [TSe].
The geographical differences in [THg] were likely a result of Hgcontamination of SFB and Tomales Bay from historical mercury andgold mining activities (Alpers et al., 2005; Johnson et al., 2009;Gehrke et al., 2011). Although atmospheric inputs, storm-waterrunoff, industrial inputs, and geologic weathering also contributeto Hg input, the fluxes of Hg (at least in SFB) are largely dominatedby contamination from historic mining (Conaway et al., 2003).These inputs have resulted in [Hg] in fish from SFB and TomalesBay that exceed concentrations of concern for human health(0.23 lg/g wet wt; Davis et al., 2002; Ridolfi et al., 2010), and con-centrations potentially of concern to wildlife health (Schwarzbachet al., 2006; Ackerman et al., 2007). In contrast, Elkhorn Slough is
iids (n = 6; triangles), and odobenids (n = 1; square; lg/g). Mercury concentrationsf the means represent concentrations solely for pups. Locations were approximatedseal was adjusted to fit on the map. Longitude was adjusted slightly for locations
ly overlay one another. Data from: Freeman and Horne (1973), Hyvärinen and Sipiläediler et al., 1993, Wiig et al. (1999), Beckmen et al. (2002), Ikemoto et al. (2004b),
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031

E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 7
primarily influenced by agricultural inputs and has lesser [Hg] insediment and fish tissues than SFB (Meador et al., 1998, 2005).Seals in Elkhorn Slough primarily forage in nearby Monterey Bay,instead of within the estuary like seals in SFB (Torok, 1994;Oxman, 1995; Eguchi and Harvey, 2005), which also may reducetheir exposure to toxicants.
The similarity in [THg]hair between seals in SFB and Tomales Baymight have been the result of similar levels of environmental con-tamination; however, movement of seals between these regionsalso could have contributed to these similarities. Whereas harborseals exhibit strong site fidelity to several haul-out sites and makepredominantly short-distance movements, long-distance move-ments among sites does occur (Yochem et al., 1987; Suryan andHarvey, 1998). This may result in overlap of foraging habitatbetween seals using these two regions. Although it was possiblefor seals in this study to travel among all site locations, previoustelemetry work indicated that overlap of foraging habitat wasmore likely for seals using SFB and Tomales Bay (Torok, 1994;Oxman, 1995; Nickel, 2003).
4.3. Sex differences
We did not detect any differences in [TSe] between males andfemales, which might have been due to homeostatic regulation ofSe. Although lactation does appear to affect circulating [TSe](Habran et al., 2011), this was not a factor in our study becauseseals were not captured during the lactation period. We did detectdifferences in [THg] between the sexes, with male harbor sealshaving greater [THg] than females in both tissue types. Brookenset al. (2007) also found that adult male harbor seals from centralCalifornia had greater [THg]hair than adult females; however, nosex differences in [THg]blood were observed. Alternatively, an ear-lier study on harbor seals from SFB indicated that mean [THg]blood
were greater in males than females (0.35 vs. 0.23 lg/g wet wt;Kopec and Harvey, 1995). A number of factors might have contrib-uted to the differences in [THg] between males and females,including the ability of females to offload Hg to their developingfetus and pup during lactation (Wagemann et al., 1988), differ-ences in the timing of molt (Thompson and Rothery, 1987;Daniel et al., 2003; Reder et al., 2003), differences in the behaviorof males and females during molt (Thompson et al., 1989; Rederet al., 2003), differences in foraging behavior (Thompson et al.,1998), and physiological variations at the molecular and biochem-ical levels (Woods et al., 2005). Differences in the toxicodistribu-tion of Hg was suggested as a potential reason for [THg]differences between male and female polar bears (Ursus maritimus;Cardona-Marek et al., 2009), though this seems unlikely in ourstudy because there was no consistent pattern with sex in the rela-tionship between [THg] and SL (i.e., [THg] of males > females at allsize classes).
The potential role that sex-specific foraging behaviors may haveplayed in the differential exposure of males and females to Hg inthis study is of particular interest because harbor seals lack strongsexual dimorphism, with males only slightly larger than females.Although the diet of harbor seals has been relatively well studied,few researchers have examined how foraging behavior differsbetween males and females. Thompson et al. (1998) found thatmale harbor seals in Scotland had longer foraging trip durationsand greater ranges than females, which may have exposed themto different prey resources. Male and female harbor seals in ourstudy had similar d13C , d15N, and d34S values, indicating that theyforaged at approximately the same trophic level and in similarregions during the molt period (i.e., inshore-benthic vs. offshore-pelagic; McHuron, 2012; unpublished results); however, this doesnot necessarily indicate they consumed the same prey species. It ispossible that males and females foraged in different locations, on
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
different prey species, or a combination of both, which might haveaffected their dietary intake of Hg. Further research on how harborseal foraging behavior differs with sex is required to better under-stand differential exposure of males and females to Hg and othertoxicants.
4.4. Relationship between selenium and mercury
Selenium may ameliorate the toxic effects of Hg through a num-ber of mechanisms, including antioxidant properties, competitionfor binding sites, and the formation of non-toxic Se–Hg complexes(Cuvin-Aralar and Furness, 1991; Yang et al., 2008). Because oftheir antagonistic interaction (covalent complexation), Hg alsomay provide protection against Se toxicosis at increased [Se]. Themolar ratio of TSe:THg may provide a better indicator of toxicityin some tissues (Ralston et al., 2008), and approaches 1:1 in theliver and kidney of marine mammals as [THg] in those tissuesincreases (Koeman et al., 1973; Dietz et al., 2000a). Deviations fromthis molar ratio have been observed, which were attributed to sev-eral factors, including species- or age-specific differences, and thepresence of alternative detoxification methods (Woshner et al.,2001a, 2001b; Brookens et al., 2007). Although hair and bloodare not considered target or traditional tissues to study for theSe–Hg complex, there still may be a relationship between theseelements as Hg may bind to, and potentially retain, seleno-compounds in circulation (Knott et al., 2011). In general, harborseals in our study had a molar surplus of Hg in hair and a molarsurplus of Se in blood; however, this molar ratio may have beenhigher or lower in other blood compartments (i.e. red blood cells,plasma, serum) because Se and Hg are compartmentalized differ-ently in blood (Lieske et al., 2011; Correa et al., 2013). It remainsunknown how [TSe] and [THg] and the molar ratios of TSe:THgrelate specifically to Hg toxicosis/Se status in free-ranging marinemammals; however, the risk of Hg toxicosis, or Hg-induced Se defi-ciency, likely increase as ratios approach 1:1 (Khan and Wang,2009). Three seals from SFB did have TSe:THg molar ratios in bloodof 1.3, 1.6, and 1.8, and as indicated by the lesser TSe:THgblood
molar ratios of SFB seals, circulating [TSe] did not appear toincrease when [THg] increased. This may indicate that the abilityof seals in this region to increase Se uptake is limited, potentiallyby the amount of bioavailable Se in prey species.
The correlation between [TSe] and [THg] in both tissues wasrelatively weak, especially for hair, which might have been becauseSe is an essential element, whereas Hg has no known biologicalfunction. Because of this, it is likely that there are differences inhow Se and Hg are incorporated into hair, which is supported bythe relatively strong relationship we observed between [THg] inblood and hair, and the absence of a relationship between [TSe]in the two tissues. We did find a weak negative correlationbetween [TSe]blood and [THg]blood that was driven by seals fromSFB that had [THg] greater than 0.4 lg/g wet wt. This result wasinconsistent with results from studies on other marine mammalsthat found positive correlations between [TSe] and [THg] in wholeblood (Knott et al., 2011; Correa et al., 2013), which could be aresult of species-specific differences, or other factors discussedbelow. Given the small number of seals with [THg] greater than0.4 (n = 7), this negative correlation may simply have been due toour inability to accurately represent the variability in [TSe] atincreased [THg]. Alternatively, this correlation could have been areflection of dietary intake because the majority of Se and Hg expo-sure is via the diet, or because the Se-Hg relationship is concentra-tion dependent. Factors that may have influenced the strength ofthis correlation include the homeostatic regulation of Se, the pro-portion of inorganic Hg present in blood, and that Se and Hg arecompartmentalized differently within blood (Lieske et al., 2011;Correa et al., 2013). No data exist for the compartmentalization
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031

8 E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
of Se and Hg in blood of harbor seals; however, Correa et al. (2013)found that a greater proportion of THg was present in packed cells(95%) compared with Se (75%). Because of this, the strength of thecorrelation between [Se] and [THg] varied depending on the bloodcompartments being compared, and was much stronger between[THg]whole blood and [Se]serum (0.781) than [THg]whole blood and[TSe]whole blood (0.433; Correa et al., 2013). This may indicate thatwe would have seen a stronger correlation between [TSe] and[THg] if we had analyzed blood compartments separately.
4.5. Relationship with stable isotopes
There was no relationship between [THg]hair and d15Nhair, indi-cating that trophic level-dependent biomagnification of Hg wasnot a primary factor for the variation in [THg] in SFB harbor seals.Although Hg biomagnifies in food webs (Dietz et al., 2000b;Campbell et al., 2005), all of the harbor seals captured in SFB fedat approximately the same trophic level (McHuron, 2012). Mercuryconcentrations in fish from SFB vary with species and locationwithin the bay (Greenfield and Jahn, 2010), which likely contrib-uted to the lack of a relationship between [THg] and d15N. Previousstudies on the relationship between THg and d15N have yieldedmixed results. Aubail et al. (2011) found a positive correlationbetween THg and d15N in hair of seven species of phocids, althoughthis relationship was only assessed among and not within a spe-cies. This relationship may have been driven primarily by the factthat species were from vastly different locations (Antarctic andArctic) and foraged at a greater range of trophic levels than harborseals in this study, as the relationship within a species appearedmuch less clear or even absent. Cardona-Marek et al. (2009) foundno direct relationship between [THg] in hair of polar bears andd15N, although a model including d15N and d13C was a good predic-tor of [THg] among bears.
Sulfur is not measured routinely in marine mammals (Hoekstraet al., 2002; Barros et al., 2010), and to our knowledge this is thefirst study to measure S stable isotopes in any pinniped. Marineprimary producers have a greater proportion of 34S compared withterrestrial plants (d34S = �21‰ vs. �0–6‰; Rees et al., 1978;Chukhrov et al., 1980) and because different producers often havedistinct signals, the use of S can help discriminate among sourcesof primary productivity in aquatic food webs (Connolly et al.,2004). In our study, the relationship between [THg] and d34S mighthave been the result of geographical foraging differences amongseals, with seals that primarily foraged on estuarine species withinSFB having greater [THg] than seals that also depended on foragingsites outside of SFB. This is supported by the fact that [Hg] in fishfrom SFB were greater in estuarine than offshore species(Greenfield and Jahn, 2010), and telemetry studies that indicatedsome seals in SFB tended to forage almost exclusively within SFB,whereas other seals made more frequent trips outside of the bay(Torok, 1994; Nickel, 2003). McGrew et al. (2014) also found a rela-tionship between [THg] and d34S values in the liver and skeletalmuscle of Alaskan gray wolves (Canis lupus), with greater valuesin coastal wolves with a marine-based diet compared with interiorwolves feeding on terrestrial prey. Results from our study and theMcGrew et al. (2014) study indicate that S should be consideredwhen using stable isotopes to study the foraging ecology and tox-icant exposure of coastal mammals.
5. Conclusions
Results from this study do not lend support to the hypothesisthat increased [Se] in harbor seals from SFB are contributing tothe observed changes to keratinized tissues in these seals. Instead,[TSe] and the TSe:THg molar ratio in hair and blood of seals fromSFB were less than seals from Elkhorn Slough. These results are
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
inconsistent with a previous study from this region (Kopec andHarvey, 1995), which may be due to efforts by the San FranciscoBay Regional Water Quality Board in the last decade to reduce agri-cultural and refinery discharges of Se to SFB, and potential seasonalbiases in sampling design in the Kopec and Harvey study(McHuron, 2012). It may simply be coincidental that seals fromareas with increased [THg] had lesser [TSe]; however, it also is pos-sible that the physiologic need for Se in animals from sites withincreased Hg may be greater because Se is needed to maintain nor-mal selenoenzyme activities and mitigate the toxic effects of Hg.Further studies investigating additional biomarkers of Se statusand the compartmentalization of Se and Hg in blood of harbor sealsfrom SFB are warranted to determine if the decreased [TSe]detected in this study are biologically important (e.g., biochemicalfunction perspective).
Despite the high [THg] in hair and blood, seals in this studyappeared in relatively good health with no noticeable signs of tox-icosis. This indicated that [THg] were not great enough to induceobservable gross health problems, or that Se or other element con-centrations were sufficient to mitigate effects of Hg. Negativehealth effects due to Hg exposure are rarely documented in free-ranging marine mammals; however, Hg remains a potential threatto marine mammal populations, especially for species that forageat a relatively high trophic level and are found in close proximityto industrial or urban areas. Seals in this study had [THg]blood farbelow concentrations in captive seals that died from acute MeHg+
poisoning (Ronald et al., 1977); however, they did have [THg] inboth tissues that exceeded concentrations associated with neuro-logical effects due to Hg toxicosis in other species (Thompson,1996; Evers et al., 2007), which was especially apparent for sealswith lower d34S values. Additionally, increases in [THg] did notresult in corresponding increases in [TSe], indicating that the abil-ity of Se to provide a protective effect against Hg toxicosis may belimited for harbor seals in this region. Pups, which were not sam-pled as part of this study, may be the cohort most likely affected byHg toxicosis because they are exposed in utero and during lactationvia milk, and are unable to demethylate (detoxify) MeHg+ as effi-ciently as adults (van de Ven and Koeman, 1979; Wagemannet al., 1988). These factors, coupled with the apparent temporalincrease in and spatial variation of [THg] in seals, warrant futureinvestigation into the role that Hg may play in the health and pop-ulation dynamics of harbor seals from central California.
Acknowledgements
This study was conducted under NMFS Permit Nos. 555-1870and 373-1686, US Fish and Wildlife Service Permit Nos. 81640-2009-041 and 81640-2011-002, National Park Service PermitPORE-2011-SCI-0003, and San Jose State University IACUC No.933. This study would not have been possible without support fromthe National Parks Service (especially Sarah Allen and Ben Becker),US Fish and Wildlife Service, Don Edwards National Wildlife Refuge,Moss Landing Marine Laboratories, the Wildlife ToxicologyLaboratory, USGS, and all the volunteers that assisted in harbor sealcaptures. Funding for this project was provided by the PADIFoundation, Packard Foundation, Harvey Fellowship, and the Earland Ethel Myers Oceanographic and Marine Biology Trust. Anyuse of trade, firm, or product names is for descriptive purposes onlyand does not imply endorsement by the U.S. Government.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.marpolbul.2014.04.031.
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031

E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 9
References
Ackerman, J.T., Eagles-Smith, C.A., Takekawa, J.Y., Demers, S.A., Adelsbach, T.L.,Bluso, J.D., Miles, A.K., Warnock, N., Suchanek, T.H., Schwarzback, S.E., 2007.Mercury concentrations and space use of pre-breeding American avocets andblack-necked stilts in San Francisco Bay. Sci. Total Environ. 384, 452–466.
Agusa, T., Nomura, K., Kunito, T., Anan, Y., Iwata, H., Tanabe, S., 2011. Accumulationof trace elements in harp seals (Phoca groenlandica) from Pangnirtung in theBaffin Island, Canada. Mar. Pollut. Bull. 63, 489–499.
Airey, D., 1983. Mercury in human hair due to environment and diet: a review.Environ. Health Perspect. 52, 303–316.
Alpers, C.N., Hunerlach, M.P., May, J.T., Hothem, R.L., 2005. Mercury contaminationfrom historical gold mining in California. United States Geological Survey FactSheet 2005–3014.
Aubail, A., Teilmann, J., Dietz, R., Rigét, F., Harkonen, T., Karlsson, O., Rosing-Asvid,A., Caurant, F., 2011. Investigation of mercury concentrations in fur of phocidseals using stable isotopes as tracers of trophic levels and geographical regions.Polar Biol. 34, 1411–1420.
Bacher, G.J., 1985. Mercury concentrations in the Australian fur seal Arctocephaluspusillis from SE Australian waters. Bull. Environ. Contam. Toxicol. 35, 490–495.
Baraj, B., Bianchini, A., Niencheski, L.F.H., Campos, C.C.R., Martinez, P.E., Robaldo,R.B., Muelbert, M.M.C., Colares, E.P., Zarzur, S., 2001. The performance of ZEISSGFAAS-5 instrument on the determination of trace metals in whole bloodsamples of southern elephant seals (Mirounga leonina) from Antarctica.Fresenius Environ. Bull. 10, 859–862.
Barron, M.G., Heintz, R., Krahn, M.M., 2003. Contaminant exposure and effects inpinnipeds: implications for Steller sea lion declines in Alaska. Sci. Total Environ.311, 111–133.
Barros, N.B., Ostrom, P.H., Stricker, C.A., Wells, R.S., 2010. Stable isotopesdifferentiate bottlenose dolphins off west-central Florida. Mar. Mamm. Sci.26, 324–336.
Beckmen, K.B., Duffy, L.K., Zhang, X., Pitcher, K.W., 2002. Mercury concentrations inthe fur of Steller sea lions and northern fur seals from Alaska. Mar. Pollut. Bull.44, 1130–1135.
Bigg, M.A., 1969. The harbour seal in British Columbia. Fish Res. Board Can. Bull.172, 1–33.
Brookens, T.J., Harvey, J.T., O’Hara, T.M., 2007. Trace element concentrations in thePacific harbor seal (Phoca vitulina richardii) in central and northern California.Sci. Total Environ. 372, 676–692.
Brown, R.F., Wright, B.E., Riemer, S.D., Laake, J., 2005. Trends in abundance andcurrent status of harbor seals in Oregon: 1977–2003. Mar. Mamm. Sci. 21, 657–670.
Campbell, L.M., Norstrom, R.J., Hobson, K.A., Muir, D.C.G., Backus, S., Fisk, A.T., 2005.Mercury and other trace elements in a pelagic Arctic marine food web(Northwater Polynya, Baffin Bay). Sci. Total Environ. 351–352, 247–263.
Cardona-Marek, T., Knott, K.K., Meyer, B.E., O’Hara, T.M., 2009. Mercuryconcentrations in Southern Beaufort Sea polar bears: variation based onstable isotopes of carbon and nitrogen. Environ. Toxicol. Chem. 28, 1416–1424.
Castellini, J.M., Rea, L.D., Lieske, C.L., Beckmen, K.B., Fadely, B.S., Maniscalco, J.M.,O’Hara, T.M., 2012. Mercury concentrations in hair from neonatal and juvenileSteller sea lions (Eumetopias jubatus): implications based on age and region inthis northern Pacific marine sentinel piscivore. EcoHealth 9, 267–277.
Chukhrov, F.V., Ermilova, L.P., Churikov, V.S., Nosik, L.P., 1980. The isotopiccomposition of plant sulfur. Org. Geochem. 2, 69–75.
Clarkson, T.W., Magos, L., 2006. The toxicology of mercury and its chemicalcompounds. Crit. Rev. Toxicol. 36, 609–662.
Conaway, C.H., Squire, S., Mason, R.P., Flegal, A.R., 2003. Mercury speciation in theSan Francisco Bay estuary. Mar. Chem. 80, 199–225.
Connolly, R.M., Guest, M.A., Melville, A.J., Oakes, J.M., 2004. Sulfur stable isotopesseparate producers in marine food-web analysis. Oecologia 138, 161–167.
Correa, L., Castellini, J.M., Wells, R.S., O’Hara, T., 2013. Distribution of mercury andselenium in blood compartments of bottlenose dolphins (Tursiops truncatus)from Sarasota Bay, Florida. Environ. Toxicol. Chem. 32, 2441–2448.
Cuvin-Aralar, M.L.A., Furness, R.W., 1991. Mercury and selenium interaction: areview. Ecotox. Environ. Saf. 21, 348–364.
Daniel, R.G., Jemison, L.A., Pendleton, G.W., Crowley, S.M., 2003. Molting phenologyof harbor seals on Tugidak Island, Alaska. Mar. Mamm. Sci. 19, 128–140.
Das, K., Siebert, U., Gillet, A., Dupont, A., Dipoï, C., Fonfara, S., Mazzucchelli, G., DePauw, E., De Pauw-Gillet, M.C., 2008. Mercury immune toxicity in harbour seals:links to in vitro toxicity. Environ. Health 7, 52.
Davis, J.A., May, M.D., Greenfield, B.K., Fairey, R., Roberts, C., Ichikawa, G., Stoelting,M.S., Becker, J.S., Tjeerdema, R.S., 2002. Contaminant concentrations in sportfish from San Francisco Bay, 1997. Mar. Pollut. Bull. 44, 1117–1129.
Day, R.W., Quinn, G.P., 1989. Comparisons of treatments after an analysis ofvariance in ecology. Ecol. Monogr. 59, 433–463.
Dietz, R., Riget, F., Born, E.W., 2000a. An assessment of selenium to mercury inGreenland marine animals. Sci. Total Environ. 245, 15–24.
Dietz, R., Riget, F., Cleemann, M., Aarkrog, A., Johansen, P., Hansen, J.C., 2000b.Comparison of contaminants from different trophic levels and ecosystems. Sci.Total Environ. 245, 221–231.
Dietz, R., Born, E.W., Rigét, F., Aubail, A., Sonne, C., Drimmie, R., Basu, N., 2011.Temporal trends and future predictions of mercury concentrations in northwestGreenland polar bears (Ursus maritimus) hair. Environ. Sci. Technol. 45, 1458–1465.
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
Dolbec, J., Mergler, D., Larribe, F., Roulet, M., Lebel, J., Lucotte, M., 2001. Sequentialanalysis of hair mercury levels in relation to fish diet of an Amazonianpopulation, Brazil. Sci. Total Environ. 271, 87–98.
Edwards, W.C., Whitenack, D.L., 1989. Selenium toxicosis in three California sealions (Zalophus californianus). Vet. Hum. Toxicol. 31, 568–570.
Eguchi, T., Harvey, J.T., 2005. Diving behavior of the Pacific harbor seal (Phocavitulina richardii) in Monterey Bay. California 21, 283–295.
Evers, D.C., Han, Y.J., Driscoll, C.T., Kamman, N.C., Goodale, M.W., Lambert, K.F.,Holsen, T.M., Chen, C.Y., Clair, T.A., Butler, T., 2007. Biological mercury hotspotsin the northeastern United States and southeastern Canada. Bioscience 57, 29–43.
Fossi, M.C., Marsili, L., Junin, M., Castello, H., Lorenzani, J.A., Casini, S., Savelli, C.,Leonzio, C., 1997. Use of nondestructive biomarkers and residue analysis toassess the health status of endangered species of pinnipeds in the south-westAtlantic. Mar. Pollut. Bull. 34, 157–162.
Freeman, H.C., Horne, D.A., 1973. Mercury in Canadian seals. Bull. Environ. Contam.Toxicol. 10, 172–180.
Frouin, H., Loseto, L.L., Stern, G.A., Haulena, M., Ross, P.S., 2012. Mercury toxicity inbeluga whale lymphocytes: limited effects of selenium protection. Aquat.Toxicol. 109, 185–193.
Fry, B., Brand, W., Mersch, F.J., Tholke, K., Garritt, R., 1992. Automated analysissystem for coupled d13C and d15N measurements. Anal. Chem. 64, 288–291.
Gehrke, G.E., Blum, J.D., Marvin-DiPasquale, M., 2011. Sources of mercury to SanFrancisco Bay surface sediment as revealed by mercury stable isotopes.Geochim. Cosmochim. Acta 75, 691–705.
Gray, R., Canfield, P., Rogers, T., 2008. Trace element analysis in the serum and hairof Antarctic leopard seal, Hydrurga leptonyx, and Weddell seal, Leptonychotesweddellii. Sci. Total Environ. 399, 202–215.
Greenfield, B.K., Jahn, A., 2010. Mercury in San Francisco Bay forage fish. Environ.Pollut. 158, 2716–2724.
Greenfield, B.K., Davis, J.A., Fairey, R., Roberts, C., Crane, D., Ichikawa, G., 2005.Seasonal, interannual, and long-term variation in sport fish contamination, SanFrancisco Bay. Sci. Total Environ. 336, 25–43.
Griesel, S., Kakuschke, A., Siebert, A., Prange, A., 2008. Trace element concentrationsin blood of harbor seals (Phoca vitulina) from the Wadden Sea. Sci. Total Environ.392, 313–323.
Grigg, E.K., Allen, S.G., Green, D.E., Markowitz, H., 2004. Harbor seal, Phoca vitulinarichardii, population trends in the San Francisco Bay Estuary, 1970–2002. CalifFish Game 90, 51–70.
Guallar, E., Sanz-Gallardo, I., van’t Veer, P., Bode, P., Aro, A., Gómez-Aracena, J., Kark,J.D., Riemersma, R.A., Martín-Moreno, J.M., Kok, F.J., 2002. Mercury, fish oils, andthe risk of myocardial infarction. N. Engl. J. Med. 347, 1747–1754.
Habran, S., Debier, C., Crocker, D.E., Houser, D.S., Das, K., 2011. Blood dynamics ofmercury and selenium in northern elephant seals during the lactation period.Environ. Pollut. 159, 2523–2529.
Hamilton, S.J., 2004. Review of selenium toxicity in the aquatic food chain. Sci. TotalEnviron. 326, 1–31.
Harris, H., Pickering, I.J., George, G.N., 2003. The chemical form of mercury in fish.Science 301, 1203.
Harvey, J.T., Brown, R.F., Mate, B.R., 1990. Abundance and distribution of harborseals (Phoca vitulina) in Oregon, 1975–1983. Northwestern Nat. 71, 65–71.
Hoekstra, P.F., Dehn, L.A., George, J.C., Solomon, K.R., Muir, D.C.G., O’Hara, T.M.,2002. Trophic ecology of bowhead whales (Balaena mysticetus) compared withthat of other artic marine biota as interpreted from carbon-, nitrogen-, andsulfur-isotope signatures. Can. J. Zool. 80, 223–231.
Hothem, R.L., Lonzarich, D.G., Takekawa, J.E., Ohlendorf, H.M., 1998. Contaminantsin wintering canvasbacks and scaups from San Francisco Bay, California.Environ. Monit. Assess 50, 67–84.
Hyvärinen, H., Sipilä, T., 1984. Heavy metals and high pup mortality in the Saimaaringed seal population in eastern Finland. Mar. Pollut. Bull. 15, 335–337.
Ikemoto, T., Kunito, T., Tanaka, H., Baba, N., Miyazaki, N., Tanabe, S., 2004a.Detoxification mechanism of heavy metals in marine mammals and seabirds:interaction of selenium with mercury, silver, copper, zinc, and cadmium in liver.Arch. Environ. Contam. Toxicol. 47, 402–413.
Ikemoto, T., Kunito, T., Watanabe, I., Yasunaga, G., Baba, N., Miyazaki, N., Petrov, E.A.,Tanabe, S., 2004b. Comparison of trace element accumulation in Baikal seals(Pusa sibirica), Caspian seals (Pusa caspica) and northern fur seals (Callorhinusursinus). Environ. Pollut. 127, 83–97.
Jeffries, S.J., Brown, R.F., Harvey, J.T., 1993. Techniques for capturing, handling, andmarking harbour seals. Aquat. Mamm. 19, 21–25.
Jeffries, S.J., Huber, H., Calambokidis, J., Laake, J., 2003. Trends and status of harborseals in Washington State: 1978–1999. J. Wildl. Manage. 67, 207–218.
Johnson, B.E., Esser, B.K., Whyte, D.C., Ganguli, P.M., Austin, C.M., Hunt, J.R., 2009.Mercury accumulation and attenuation at a rapidly forming delta with a pointsource of mining waste. Sci. Total Environ. 407, 5056–5070.
Kester, C.L., Rye, R.O., Johnson, C.A., Schwartz, C.H., Holmes, C.H., 2001. On-linesulfur isotope analysis of organic material by direct combustion: preliminaryresults and potential applications. Isotopes Environ. Health Stud. 37, 53–65.
Khan, M.A.K., Wang, F., 2009. Mercury–selenium compounds and their toxicologicalsignificance: toward a molecular understanding of the mercury–seleniumantagonism. Environ. Toxicol. Chem. 28, 1567–1577.
Knott, K.K., Schenk, P., Beyerlein, S., Boyd, D., Ylitalo, G.M., O’Hara, T.M., 2011. Blood-based biomarkers of selenium and thyroid status indicate possible adversebiological effects of mercury and polychlorinated biphenyls in SouthernBeaufort Sea polar bears. Environ. Res. 111, 1124–1136.
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031

10 E.A. McHuron et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
Koeman, J.H., Peeters, W.H.M., Koudstaal-Hol, C.H.M., Tjioe, P.S., De Goeij, J.J.M.,1973. Mercury–selenium correlations in marine mammals. Nature 245, 385–386.
Kopec, D.A., Harvey, J.T., 1995. Toxic pollutants, health indices, and populationdynamics of harbor seal in San Francisco Bay, 1989–1992, vol. 96–4. MossLanding Mar Lab Tech Pub.
Lailson-Brito, J., Cruz, R., Dorneles, P.R., Andrade, L., Azevedo, A., Fragoso, A.B., Vidal,L.G., Costa, M.D., Bisi, T.L., Almeida, R., Carvalho, D.P., Bastos, W.R., Malm, O.,2012. Mercury–selenium relationships in liver of Guiana dolphin: the possiblerole of Kupffer cells in the detoxification process by tiemannite formation. PLoSONE. http://dx.doi.org/10.1371/journal.pone.0042162.
Lieske, C.L., Moses, S.K., Castellini, J.M., Klejka, J., Hueffer, K., O’Hara, T.M., 2011.Toxicokinetics of mercury in blood compartments and hair of fish-fed sled dogs.Acta Vet. Scand. 53, 66.
Lowry, M.S., Carretta, J.V., Forney, K.A., 2008. Pacific harbor seal census in Californiaduring May-July 2002 and 2004. Cal. Fish Game 94, 180–193.
McGrew, A.K., Ballweber, L.R., Moses, S.K., Stricker, C.A., Beckmen, K.B., Salman,M.D., O’Hara, T.M., 2014. Mercury in gray wolves (Canis lupus) in Alaska:increased exposure through consumption of marine prey. Sci. Total Environ.468–469, 609–613.
McHuron, E.A., 2012. Health implications of mercury, selenium, and a red pelage inharbor seals (Phoca vitulina richardii) off central California [M.Sc. thesis] San JoseState University, Moss Landing, CA.
Meador, J.P., Robisch, P.A., Clark, R.C., Ernest, D.W., 1998. Elements in fish andsediment from the Pacific Coast of the United States: results from the nationalbenthic surveillance project. Mar. Pollut. Bull. 37, 56–66.
Meador, J.P., Ernest, D.W., Kagley, A.N., 2005. A comparison of the non-essentialelements cadmium, mercury, and lead found in fish and sediment from Alaskaand California. Sci. Total Environ. 339, 189–205.
Medvedev, N., Panichev, N., Hyvärinen, H., 1997. Levels of heavy metals in seals ofLake Lagoda and the White Sea. Sci. Total Environ. 206, 95–105.
Moser, D.G., 1996. Trace element and organochlorine compounds in harbor seals(Phoca vitulina richardsi) along the Pacific Coast [M.Sc. thesis]. San Jose StateUniversity, Moss Landing, CA.
Neale, J.C.C., Gulland, F.M.D., Schmelzer, K.R., Harvey, J.T., Bert, E.A., Allen, S.G.,Greig, D.J., Grigg, E.K., Tjeerdema, R.S., 2005. Contaminant loads andhematological correlates in the harbor seal (Phoca vitulina) of San FrancicoBay, California. J. Toxicol. Environ. Health 68, 617–633.
Nickel, B.A., 2003. Movement and habitat use patterns of harbor seals in the SanFrancisco Estuary, California [M.A. thesis]. San Francisco State University, SanFrancisco, CA.
Ohlendorf, H.M., Lowe, R.W., Kelly, P.R., Harvey, T.E., 1986. Selenium and heavymetals in San Francisco Bay diving ducks. J. Wildl. Manage. 50, 64–70.
O’Toole, D., Raisbeck, M.F., 1995. Pathology of experimentally induced chronicselenosis (alkali disease) in yearling cattle. J. Vet. Diagn. Invest. 7, 364–373.
O’Toole, D., Raisbeck, M.F., 1997. Experimentally induced selenosis of adult mallardducks: clinical signs, lesions, and toxicology. Vet. Path. 34, 330–340.
Oxman, D.S., 1995. Seasonal abundance, movements, and food habits of harbor seals(Phoca vitulina richardsi) in Elkhorn Slough, California [M.Sc. thesis]. CaliforniaState University Stanislaus, Moss Landing, CA.
Pauly, D., Trites, A.W., Capuli, E., Christensen, V., 1998. Diet composition and trophiclevel of marine mammals. ICES J. Mar. Sci. 55, 467–481.
Raisbeck, M.F., Dahl, E.R., Sanchez, D.A., Belden, E.L., O’Toole, D., 1993. Naturallyoccurring selenosis in Wyoming. J. Vet. Diagn. Invest. 5, 84–87.
Ralston, N.V.C., Ralston, C.R., Blackwell III, J.L., Raymond, L.J., 2008. Dietary andtissue selenium in relation to methymercury toxicity. Neuro Toxicol. 29, 802–811.
Reder, S., Lydersen, C., Arnold, W., Kovacs, K.M., 2003. Haulout behaviour of HighArctic harbour seals (Phoca vitulina vitulina) in Svalbard, Norway. Polar Biol. 27,6–16.
Rees, C.E., Jenkins, W.J., Monster, J., 1978. The sulfur isotopic composition of oceanwater sulphate. Geochim. Cosmochim. Acta 42, 377–381.
Reijnders, P.J.H., 1986. Reproductive failure in common seals feeding on fish frompolluted coastal waters. Nature 324, 456–457.
Ridolfi, K., Grenier, L., Melwani, A., McKee, L., Allen, R., Collins, J., Davis, J., 2010.Impairment assessment for mercury in Tomales Bay, CA. ACS/Watersheds/Conservation Biology; 2010 Jul. Contribution No. 614. Sponsored by the SanFrancisco Bay Water Quality Control Board.
Ronald, K., Tessaro, S.V., Uthe, J.F., Freeman, H.C., Frank, R., 1977. Methylmercurypoisoning in the harp seal (Pagophilus groenlandicus). Sci. Total Environ. 8, 1–11.
Ross, P.S., 2000. Marine mammals as sentinels in ecological risk assessment. Hum.Ecol. Risk Assess. 6, 29–46.
Ross, P.S., 2002. The role of immunotoxic environmental contaminants infacilitating the emergence of infectious diseases in marine mammals. Hum.Ecol. Risk Assess. 8, 277–292.
Ross, P., De Swart, R., Addison, R., Van Loveren, H., Vos, J., Osterhaus, A., 1996.Contaminant-induced immunotoxicity in harbour seals: wildlife at risk?Toxicology 112, 157–169.
Please cite this article in press as: McHuron, E.A., et al. Selenium and mercury coimplications in an urbanized estuary. Mar. Pollut. Bull. (2014), http://dx.doi.or
Saeki, K., Nakajima, M., Noda, K., Loughlin, T.R., Baba, N., Kiyota, M., Tatsukawa, R.,Calkins, D.G., 1999. Vanadium accumulation in pinnipeds. Arch. Environ.Contam. Toxicol. 36, 81–86.
Schwarzbach, S.E., Albertson, J.D., Thomas, C.M., 2006. Effects of predation, flooding,and contamination on reproductive success of California Clapper Rails (Ralluslongirostris obsoletus) in San Francisco Bay. Auk 123, 45–60.
Slemr, F., Brunke, E., Ebinghaus, R., Temme, C., Munthe, J., Wängberg, I., Schroeder,W., Steffen, A., Bert, T., 2003. Worldwide trend of atmospheric mercury since1977. Geophys. Res. Lett. 30, 1516.
Stewart, A.R., Luoma, S.N., Schlekat, C.E., Doblin, M.A., Hieb, K.A., 2004. Food webpathway determines how selenium affects aquatic ecosystems: a San FranciscoBay case study. Environ. Sci. Technol. 38, 4519–4526.
Suryan, R.M., Harvey, J.T., 1998. Tracking harbor seals (Phoca vitulina richardsi) todetermine dive behavior, foraging activity, and haul-out site use. Mar. Mamm.Sci. 14, 361–372.
Thompson, D.R., 1996. Mercury in birds and terrestrial mammals. In: Beyer, W.N.,Heinz, G.H., Redmon-Norwood, A.W. (Eds.), Environmental Contaminants inWildlife: Interpreting Tissue Concentrations. Lewis Publishers, Boca Raton, FL,pp. 341–356.
Thompson, P., Rothery, P., 1987. Age and sex differences in the timing of moult inthe common seal, Phoca vitulina. J. Zool. (Lond.) 212, 597–603.
Thompson, P.M., Fedak, M.A., McConnell, B.J., Nicholas, K.S., 1989. Seasonal and sex-related variation in the activity patterns of common seals (Phoca vitulina). J.Appl. Ecol. 26, 521–535.
Thompson, P.M., Mackay, A., Tollit, D.J., Enderby, S., Hammond, P.S., 1998. Theinfluence of body size and sex on the characteristics of harbour seal foragingtrips. Can. J. Zool. 76, 1044–1053.
Torok, M.L., 1994. Movements, daily activity patterns, dive behavior, and foodhabits of harbor seals (Phoca vitulina richardsi) in San Francisco Bay, California[M.Sc. thesis]. California State University Stanislaus, Moss Landing CA.
van de Ven, W.S.M., Koeman, J.H., 1979. Mercury and selenium in wild andexperimental seals. Chemosphere 8, 539–555.
Wagemann, R., Stewart, R.E.A., Lockhart, W.L., Stewart, B.E., Povoledo, M., 1988.Trace metals and methyl mercury: associations and transfer in harp seal (Phocagroenlandica) mothers and their pups. Mar. Mamm. Sci. 4, 339–355.
Wells, R.S., Rhinehart, H.L., Hansen, L.J., Sweeney, J.C., Townsend, F.I., Stone, R.,Casper, D.R., Scott, M.D., Hohn, A.A., Rowles, T.K., 2004. Bottlenose dolphins asmarine ecosystem sentinels: developing a health monitoring system. EcoHealth1, 246–254.
Wenzel, C., Adelung, D., Kruse, H., Wassermann, O., 1993. Trace metal accumulationin hair and skin of the harbour seal, Phoca vitulina. Mar. Pollut. Bull. 26, 152–155.
Wiig, O., Renzoni, A., Gjertz, I., 1999. Levels of cadmium and mercury in the hair ofAtlantic walruses, (Odobenus rosmarus rosmarus) from Svalbard, Norway. PolarBiol. 21, 343–346.
Wolfe, M.F., Schwarzbach, S., Sulaiman, R.A., 1998. Effects of mercury on wildlife: acomprehensive review. Environ. Toxicol. Chem. 17, 146–160.
Woods, J.S., Echeverria, D., Heyer, N.J., Simmonds, P.L., Wilkerson, J., Farin, F.M.,2005. The association between genetic polymorphisms of coproporphyrinogenoxidase and an atypical porphyrinogenic response to mercury exposure inhumans. Toxicol. Appl. Pharm. 206, 113–120.
Woshner, V.M., O’Hara, T.M., Bratton, G.R., Suydam, R.S., Beasley, V.R., 2001a.Concentrations and interactions of selected essential and non-essentialelements in bowhead and beluga whales of arctic Alaska. J. Wildl. Dis. 37,693–710.
Woshner, V.M., O’Hara, T.M., Bratton, G.R., Beasley, V.R., 2001b. Concentrations andinteractions of selected essential and non-essential elements in ringed seals andpolar bears of arctic Alaska. J. Wildl. Dis. 37, 711–721.
Woshner, V., Knott, K., Wells, R., Willetto, C., Swor, R., O’Hara, T., 2008. Mercury andselenium in blood and epidermis of bottlenose dolphins (Tursiops truncatus)from Sarasota Bay, FL: interaction and relevance to life history and hematologicparameters. EcoHealth 5, 360–370.
Yang, G., Wang, S., Zhou, R., Sun, S., 1983. Endemic selenium intoxication of humansin China. Am. J. Clin. Nutr. 37, 872–881.
Yang, D.Y., Chen, Y.W., Gunn, J.M., Belzile, N., 2008. Selenium and mercury inorganisms: interactions and mechanisms. Environ. Rev. 16, 71–92.
Yediler, A., Panou, A., Schramel, P., 1993. Heavy metals in hair samples of theMediterranean monk seal (Monachus monachus). Mar. Pollut. Bull. 26, 156–159.
Yochem, P.K., Stewart, B.S., DeLong, R.L., DeMaster, D.P., 1987. Diel haul-outpatterns and site fidelity of harbor seals (Phoca vitulina richardsi) on San MiguelIsland, California, in autumn. Mar. Mamm. Sci. 3, 323–332.
Yoneda, S., Suzuki, K.T., 1997. Detoxification of mercury by selenium by binding ofequimolar Hg–Se complex to a specific plasma protein. Toxicol. Appl.Pharmacol. 143, 274–280.
Zahir, F., Rizwi, S.J., Haq, S.K., Khan, R.H., 2005. Low dose mercury toxicity andhuman health. Environ. Toxicol. Pharm. 20, 35.
ncentrations in harbor seals (Phoca vitulina) from central California: Healthg/10.1016/j.marpolbul.2014.04.031