selective activation of α7 nicotinic acetylcholine ... · jussara lagrota-candidod, thereza...
TRANSCRIPT

Available online at www.sciencedirect.com
www.elsevier.com/locate/brainres
b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 6
http://dx.doi.org/100006-8993/& 2014 El
nCorrespondenceFax: þ55 21 2629 226
E-mail addressescarmen.perezd@uamlagrota-candido@vm
1Present address
Research Report
Selective activation of α7 nicotinic acetylcholinereceptor (nAChRα7) inhibits muscular degenerationin mdx dystrophic mice
Paulo Emılio Correa Leitea,1, Luıs Gandıab, Ricardo de Pascualb,Carmen Nanclaresb, Ines Colmenab, Wilson C. Santosb,c,Jussara Lagrota-Candidod, Thereza Quirico-Santosa,n
aDepartment of Cellular and Molecular Biology, Fluminense Federal University, Rio de Janeiro, BrazilbInstituto Teófilo Hernando, Department of Pharmacology and Therapeutics, Autonomous University of Madrid,Madrid, SpaincDepartment of Pharmacy Administration, Fluminense Federal University, Rio de Janeiro, BrazildDepartment of Immunobiology, Fluminense Federal University, Rio de Janeiro, Brazil
a r t i c l e i n f o
Article history:
Accepted 3 May 2014
Amount evidence indicates that α7 nicotinic acetylcholine receptor (nAChRα7) activationreduces production of inflammatory mediators. This work aimed to verify the influence of
Available online 14 May 2014
Keywords:
Skeletal muscle
Mdx mice
Muscular dystrophy
Inflammation
Acetylcholine receptor
Cholinergic anti-inflammatory
pathway
.1016/j.brainres.2014.05.00sevier B.V. All rights res
to: Laboratory of Cellular8.: [email protected] (P.E.es (C. Nanclares), ines..uff.br (J. Lagrota-Candid: National Institute of M
a b s t r a c t
endogenous nAChRα7 activation on the regulation of full-blown muscular inflammation in
mdx mouse with Duchenne muscular dystrophy. We used mdx mice with 3 weeks-old at
the height myonecrosis, and C57 nAChRα7þ/þ wild-type and nAChRα7�/� knockout mice
with muscular injury induced with 60 mL 0.5% bupivacaine (bp) in the gastrocnemius
muscle. Pharmacological treatment included selective nAChRα7 agonist PNU282987
(0.3 mg/kg and 1.0 mg/kg) and the antagonist methyllycaconitine (MLA at 1.0 mg/kg)
injected intraperitoneally for 7 days. Selective nAChRα7 activation of mdx mice with
PNU282987 reduced circulating levels of lactate dehydrogenase (LDH, a marker of cell death
by necrosis) and the area of perivascular inflammatory infiltrate, and production of
inflammatory mediators TNFα and metalloprotease MMP-9 activity. Conversely,
PNU282987 treatment increased MMP-2 activity, an indication of muscular tissue remodel-
ing associated with regeneration, in both mdx mice and WTα7 mice with bp-induced
muscular lesion. Treatment with PNU282987 had no effect on α7KO, and MLA abolished the
nAChRα7 agonist-induced anti-inflammatory effect in both mdx and WT. In conclusion,
nAChRα7 activation inhibits muscular inflammation and activates tissue remodeling by
4erved.
Pathology, Institute of Biology, Fluminense Federal University, Niterói, RJ 24020-141, Brazil.
.C. Leite), [email protected] (L. Gandía), [email protected] (R. de Pascual),[email protected] (I. Colmena), [email protected] (W.C. Santos),o), [email protected] (T. Quirico-Santos).etrology, Quality and Technology (INMETRO) DIMAV, Rio de Janeiro, Brazil.

b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 628
increasing muscular regeneration. These effects were not accompanied with fibrosis and/
or deposition of non-functional collagen. The nAChRα7 activation may be considered as a
potential target for pharmacological strategies to reduce inflammation and activate
mechanisms of muscular regeneration.
& 2014 Elsevier B.V. All rights reserved.
1. Introduction
The nervous and immune systems are not fully independent.In order to maintain homeostasis, both produce cytokines,neurotransmitters and/or express receptors that act upon impor-tant physiological functions (Gallowitsch-Puerta and Pavlov,2007; Junger, 2011; Pena et al., 2011; Rosas-Ballina et al., 2011;Tracey, 2002; Ulloa, 2013; Wang et al., 2004). The vagus nervesystem (VNS) exerts an essential role in the regulation ofinflammation via releasing endogenous acetylcholine (ACh) inthe parenchyma of innervated organs. Although predominantlyexpressed in neuronal tissues, several types of immune cells,including macrophages, also express nAChRα7 mRNA (AlSharariet al., 2013), and reduction of inflammatory cytokine productionoften occurs following nAChRα7 macrophage signaling (Berniket al., 2002). Moreover, in vitro treatment of lipopolysaccharide-stimulated humanmacrophages with ACh reduces production ofpro-inflammatory cytokines (TNFα, IL-1β, IL-6, IL-18) but not ofthe anti-inflammatory cytokine IL-10, and direct in vivo VNSstimulation also prevented development of shock (Borovikovaet al., 2000). Likewise, ACh released from activate spleen Tlymphocytes interact with nAChRα7 on macrophages presentin the red pulp and marginal zone inhibiting TNFα production(Andersson and Tracey, 2012; Rosas-Ballina et al., 2011) and B cellantibody secretion (Andersson and Tracey, 2012), but activatingthe release of the anti-inflammatory IL-10 cytokine from Th2cells (Trakhtenberg and Goldberg, 2011).
The potential role of the cholinergic anti-inflammatorypathway via VNS and nAChRs activation has been demon-strated in various experimental models of septic peritonitis(Borovikova et al., 2000); ultraviolet experimentally injured skin(Osborne-Hereford et al., 2008); collagen-induced rheumatoidarthritis (van Maanen et al., 2009), and more recently it wasdemonstrated that nAChRα7-dependent mechanisms and sig-naling are involved in the modulation of chronic inflammatoryneuropathic pain (AlSharari et al., 2013), and also neuroprotec-tion under ischemic conditions by regulation of neuroinflam-mation and oxidative stress (Parada et al., 2013).
Duchenne muscular dystrophy (DMD) is an X-linked pro-gressive fatal myopathy caused by mutations in the geneencoding for the cytoskeletal protein dystrophin, which isimportant for organizing the membrane cytoskeleton, andaggregating ion channels and neurotransmitter receptors(Carlson, 1998; Hoffman et al., 1987). Lack of dystrophincompromises the structural integrity of the cell membraneleading to aberrant intracellular signaling cascades that reg-ulate both inflammatory and immune activities and contributesubstantially to the physiopathology of muscular lesion (Evanset al., 2009; Lagrota-Candido et al., 2002). Mdx mouse, the
animal model of human DMD, develops a benign phenotypewith a multi-staged disorder characterized by intense myone-crosis with scattered inflammatory infiltrate at 4 weeks (4 wks)of age followed by muscular regeneration (12 wks) and laterpersistent fibrosis (24 wks) (Collins and Morgan, 2003; Evanset al., 2009; Lagrota-Candido et al., 2002). Our group observed amodulation of nAChRα7 expression at different stages of themdx muscular dystrophy, and that nicotine treatment atte-nuated muscular inflammation and increased muscle regen-eration (Leite et al., 2010). This work aimed to provideadditional insight on selective activation of nAChRα7 in themdx muscular inflammation. For such purpose, we used theα7 KO mice and pharmacological manipulation (Parada et al.,2013) with PNU282987 and the nAChRα7 antagonist methylly-caconitine (MLA). This approach allowed us to determine aputative role of α7-mediated stimulation or blockade of endo-genous cholinergic activation in the mdx muscular pathology.
2. Results
2.1. Selective nAChRα7 activation reduces serumand muscular inflammation in mdx mice
TNFα circulating levels were determined to verify a putativeeffect of nAChRα7 activation in mdx mice inflammation. Forsuch purpose it was chosen a 7-daily treatment withPNU282987, a selective nAChRα7 agonist used at two differentdoses (0.3 and 1.0 mg/kg); the MLA antagonist (1.0 mg/kg), orboth agonist and MLA at 1.0 mg/kg. Intraperitoneal treatment ofmdx mice with low dose (0.3 mg/kg) PNU282987 reduced TNFαcirculating levels (Fig. 1A), and such effect was even moreevident (2173.3%; po0.005) following treatment with 1.0 mg/kg dose. Conversely, treatment of mdx mice with the compe-titive antagonist MLA at 1mg/kg abolished the effect, beingobserved a slight increase on TNFα circulating levels comparingwith vehicle-treated mouse. Furthermore, in combination withthe agonist (1 mg/kg PNU282987 plus 1mg/kg MLA) completelyabolished the agonist effect, thus indicating a role for nAChRα7in reducing the production of TNFα pro-inflammatory cytokine.
Next, we verified whether nAChRα7 activation could influ-ence in the production and activity of MMPs, a family ofendopeptidases which are important in normal physiologicalprocesses and skeletal tissue remodeling (Bani et al., 2008).Increased MMP-9 activity in the serum reflects a process ofactive ongoing inflammation in several models (Khandogaet al., 2006; Maddahi et al., 2012; Schiøtz Thorud et al., 2005).Treatment with 1mg/kg PNU282987 caused a marked reduction(91716%; po0.005) of MMP-9 activity in the sera and also asignificant reduction (54718%, po0.05) in the gastrocnemius
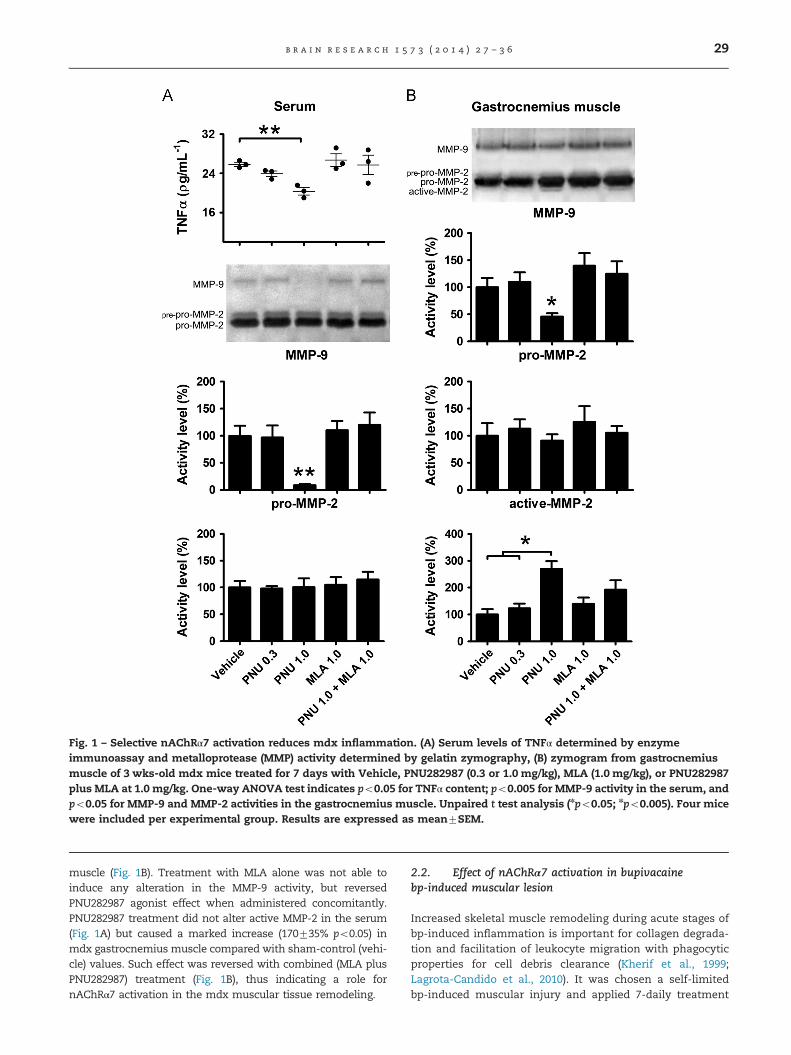
Fig. 1 – Selective nAChRα7 activation reduces mdx inflammation. (A) Serum levels of TNFα determined by enzymeimmunoassay and metalloprotease (MMP) activity determined by gelatin zymography, (B) zymogram from gastrocnemiusmuscle of 3 wks-old mdx mice treated for 7 days with Vehicle, PNU282987 (0.3 or 1.0 mg/kg), MLA (1.0 mg/kg), or PNU282987plus MLA at 1.0 mg/kg. One-way ANOVA test indicates po0.05 for TNFα content; po0.005 for MMP-9 activity in the serum, andpo0.05 for MMP-9 and MMP-2 activities in the gastrocnemius muscle. Unpaired t test analysis (npo0.05; npo0.005). Four micewere included per experimental group. Results are expressed as mean7SEM.
b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 6 29
muscle (Fig. 1B). Treatment with MLA alone was not able toinduce any alteration in the MMP-9 activity, but reversedPNU282987 agonist effect when administered concomitantly.PNU282987 treatment did not alter active MMP-2 in the serum(Fig. 1A) but caused a marked increase (170735% po0.05) inmdx gastrocnemius muscle compared with sham-control (vehi-cle) values. Such effect was reversed with combined (MLA plusPNU282987) treatment (Fig. 1B), thus indicating a role fornAChRα7 activation in the mdx muscular tissue remodeling.
2.2. Effect of nAChRα7 activation in bupivacainebp-induced muscular lesion
Increased skeletal muscle remodeling during acute stages ofbp-induced inflammation is important for collagen degrada-tion and facilitation of leukocyte migration with phagocyticproperties for cell debris clearance (Kherif et al., 1999;Lagrota-Candido et al., 2010). It was chosen a self-limitedbp-induced muscular injury and applied 7-daily treatment

Fig. 2 – nAChRα7 activation influences muscular MMP activities, Zymograms from muscular lesion induced with 60 llbupivacaine (0.5%) injected in the gastrocnemius muscle of nAChRα7þ/þ (A) and nAChRα7�/� (B) mice treated for 4 days withVehicle, 1.0 mg/kg PNU282987, or without any treatment (Control). One-way ANOVA test indicates po0.05 for MMP-9 andMMP-2 activities of nAChRα7þ/þ mice. Unpaired t-test analysis (*po0.05). Four mice were included per experimental group.Results are expressed as mean7SEM.
b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 630

b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 6 31
with drugs in nAChRα7þ/þ and nAChRα7�/� mice. It wasobserved significant reduction (33711%; po0.05) on MMP-9activity, and a marked increase (81713%; po0.05) on active-MMP-2 activity in the wild-type nAChRα7þ/þ (Fig. 2A). Con-versely, treatment with the nAChRα7 selective agonist did notchange MMP-9, pro-MMP-2 and active-MMP-2 activities in thenAChRα7�/� mice (Fig. 2B).
2.3. Selective nAChRα7 activation reduces muscle celldeath
As selective nAChRα7 activation reduced muscular inflam-mation mediated by MMP-9 activity, we considered that
Fig. 3 – Selective nAChRα7 activation reduces muscle cell death. (days with Vehicle, 1.0 mg/kg PNU282987, 1.0 mg/kg MLA, or botnAChRα7þ/þ and nAChRα7�/�mice with 60 ll bp-induced musculwith Vehicle, 1.0 mg/kg PNU282987, or PNU282987 plus MLA atmuscle cells treated with vehicle or PNU282987 at 1.0 mg/kg. Scwere included per experimental group. Results are expressed a
PNU282987 could exert a protective effect on muscular cellmembrane integrity. As an indicator for cytotoxicity damageit was used quantification of LDH release in the serum ofmdx, and bp-induced muscle injury of nAChRα7þ/þ andnAChRα7�/� mice. Intraperitoneal PNU282987 (1 mg/kg)injection during 7 days reduced LDH activity in comparisonto sham-control vehicle (71716%, po0.05). Treatment of mdxmice with 1 mg/kg MLA abolished PNU282987 effect andfurther increased LDH levels in comparison to mdx sham-control (vehicle). Furthermore, treatment of mdx mice withPNU282987 plus MLA both at 1 mg/kg was capable to partlyreverse (po0.05) the protective effect afforded by the selectivenAChRα7 activation (Fig. 3A). Serum of nAChRα7þ/þ mice with
A) Graph depicts LDH serum levels of mdx mice treated for 7h PNU282987 and MLA at 1.0 mg/kg. (B) LDH serum level ofar injury in the gastrocnemius muscle and treated for 4 days1.0 mg/kg. (C) Images of Evans blue incorporation by mdxale bar 100 lm. Unpaired t-test analysis (*po0.05). Five mices mean7SEM.

b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 632
bp-induced muscle injury obtained after treatment withselective nAChRα7 activation showed marked reduction(79730%, po0.05) on LDH activity compared to control-sham mice receiving vehicle. Combined drug treatment(PNU282987 plus MLA) abolished (po0.05) the effect. Conver-sely, drug treatment of nAChRα7�/�KO mice with bp-inducedmuscular lesion did not change the pattern of LDH activity inthe serum (Fig. 3B). To establish a relation of nAChRα7selective activation with reduced LDH serum levels andmyoblast membrane integrity mice received Evans blue dye24 h before sacrifice. Evans blue dye was almost excluded
Fig. 4 – nAChRα7 activation reduces mdx muscular lesion and iIB4 biotin antibody and streptavidin Cy3 conjugate (white arrowblood vessels in the gastrocnemius muscle (white dots) of mdx1.0 mg/kg MLA, or PNU282987 plus MLA at 1.0 mg/kg, in contrashow areas with inflammatory infiltrate and regeneration of md1.0 mg/kg PNU282987. Scale bar 100 lm. (C) Morphometric analycollagen deposition in mdx gastrocnemius muscle. (D) Morphomgastrocnemius muscle of nAChRα7þ/þ and nAChRα7�/� after treUnpaired t test analysis (npo0.05; npo0.005). Seven mdx, and foexperimental group. Results are expressed as mean7SEM.
from mdx skeletal muscle fibers in mdx mice treated with1.0 mg/kg PNU282987 which showed few red-fluorescentmuscle fibers, an indication that treatment exerted a protec-tive effect reducing myoblast necrosis (Fig. 3C).
2.4. Selective nAChRα7 activation reduces mdxmuscular lesion
Extravasation of inflammatory leukocytes from activatedvascular endothelial cells into the site of injury is involvedin the physiopathology of mdx muscular lesion by either
mproves regeneration (A) Depicts blood vessels labeled withs); and area of perivascular inflammatory infiltrate nearbymice treated for 7 days with Vehicle; 1.0 mg/kg PNU282987;st to nAChRα7þ/þ control mice. (B) Histological micrographsx gastrocnemius muscle after treatment with Vehicle orsis of foci with inflammatory infiltrate, regeneration andetric analysis of foci with inflammatory infiltrate in theatment with Vehicle or 1.0 mg/kg PNU282987 for 4 days.ur nAChRα7þ/þ and nAChRα7�/� mice were included per

b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 6 33
promoting myonecrosis or muscular regeneration. IsolectinB4 (IB4) immunolabeling was used as histochemical markerto assess the influence of nAChRα7 selective activation onvascular endothelia of mdx mice.
A slight IB4 immunolabeling was observed in the controlwild-type nAChRα7þ/þ muscular tissues. In contrast, after7-days treatment with vehicle control-sham mdx miceshowed an intense perivascular inflammatory infiltrate(Fig. 4A). Treatment of mdx mice with 1.0 mg/kg PNU282987reduced the area of perivascular inflammatory infiltrate andalso the presence of IB4 positive active endothelia. Treatmentwith the competitive antagonist MLA reversed the anti-inflammatory effect of PNU282987 (Fig. 4A). Morphometricanalysis confirmed that treatment of mdx mice with theselective nAChRα7 agonist PNU282987 reduced the area ofinflammatory infiltrate (49713%, po0.005) and increased(34712%, po0.05) the percentage of regenerating myofiberscharacterized by strong basophilic and centrally locatednuclei. Treatments did not cause significant changes (Fig. 4Band C) in sirius red staining, used as a parameter of collagendeposition in the muscular microenvironment. Histologicalanalysis of gastrocnemius muscle of nAChRα7þ/þ mice withbp-induced muscular lesion showed a significant reduction ofinflammatory infiltrate area (38%711%, po0.05) after selec-tive nAChRα7 activation with PNU282987. In contrast, theexperimentally injured muscle nAChRα7�/� mice did notshow any alteration after treatment (Fig. 4D).
3. Discussion
Excessive inflammatory signaling is the underlying pathogenicmechanism leading to tissue damage in chronic inflammatorydiseases, although activation of distinct anti-inflammatorypathways triggers the process of wound healing and tissuerepair (Murray and Smale, 2012). Indeed, activation of endo-genous pathways responsible for regulating inflammationmayrestraint exacerbated immune response and activate diversemechanisms that promote muscle regeneration in the mdxmouse, a model of inflammatory myopathy (Leite et al., 2010).
Data from various experimental models of inflammationpoint towards the nAChRα7 being crucial for cholinergicmodulation of inflammation (AlSharari et al., 2013; Paradaet al., 2013). Although in vitro response to high nAChRα7agonist concentrations rapidly desensitize the receptor for along period (AlSharari et al., 2013), in vivo treatment withnicotine was capable to significantly reduce mdx inflamma-tory lesion and increase muscle regeneration (Leite et al.,2010). Indeed, the α7 subunit-containing nicotinic acetylcho-line receptor (nAChRα7) is an essential component in thecholinergic anti-inflammatory pathway regulating the levelsof TNF, high mobility group box 1 (HMGB1), and othercytokines during inflammation (Parrish et al., 2008; Pavlovand Tracey, 2012).
To provide additional insight on the role of the α7 nAChRin mdx chronic muscular inflammation, we used the α7 KOmice and in vivo pharmacological manipulation withPNU282987, a highly selective nAChRα7 agonist that causeminor side effects than nicotine, and the MLA antagonist(Wang et al., 2004). The spleen is a primary target for signals
in the efferent pathway of the inflammatory reflex, becausealmost all TNF released systemically in the early phase ofrodent endotoxemia originates in the spleen (Olofsson et al.,2012). We observed that repeated administration of PNU282987markedly reduced circulating levels of proinflammatory TNFαcytokine and MMP-9 activity in the serum, and also in mdxgastrocnemius muscle.
Matrix metalloproteases MMP-2, MMP-9 and also MMP-10are directly involved in muscle plasticity being essential forskeletal muscle maintenance and regeneration in musculardystrophies and sarcolemmal damage associated withmechanical injury (Bani et al., 2008; Bobadilla et al., 2014;Li et al., 2009; Rullman et al., 2007). Since MMP-9 is up-regulated at the height (3 wks-old) of the mdx myonecrosis(Bani et al., 2008; Li et al., 2009), a role for nAChRα7 inreducing mdx muscle inflammation was further confirmedwith experiments showing MLA-induced reversion of theagonist stimulatory effect. Moreover, the nAChRα7 agonistwas also capable to activate mdx muscle regeneration, asin vivo treatment with PNU282987 agonist also increasedMMP-2 activity and the area with basophilic myoblastscharacteristic of muscle regeneration. In addition, treatmentdid not reduce tissue inflammation in nAChRα7�/� mice withbp-induced muscular lesion. This is an indication that selec-tive nAChRα7 activation reduces the inflammatory signalingmediated by metalloprotease MMP-9 important for TNFαcleavage into 17-kDa mature form and the inflammatorypleiotropic actions of NFkB transcription factor signalingpathway. Interestingly NFkB activity in mdx muscle is loca-lized in both dystrophic muscle and immune cells, andpersistent NFkB signaling exacerbates production of pro-inflammatory mediators that promotes muscle necrosis andlimits skeletal muscle fiber regeneration through inhibitoryeffect on muscle progenitor cells (Acharyya et al., 2007).
The abnormal clustering of acetylcholine receptors (AChR) onmdx skeletal muscle fibers compromises the formation ofneuromuscular junction and functional integrity of skeletalmuscles leading to massive infiltration of inflammatory cells,necrosis, and severe muscle degeneration characterized byincreased leakage of cytosolic LDH and creatine kinase enzymes.Treatment with the nAChRα7 agonist PNU282987 reduced LDHrelease in the serum and myofiber necrosis of mdx andnAChRα7þ/þ with bp-muscular lesion. Therefore, selective acti-vation of nAChRα7 in both experimental models was capable toprevent development of muscular degeneration. Moreover, selec-tive nAChRα7 activation also increased MMP-2 activity in skeletalmuscles from both mdx and nAChRα7þ/þ with bp-muscularlesion. This is an indication that nAChRα7 activation directlyinfluences tissue remodeling by increasing muscular regenera-tion without inducing fibrosis and/or deposition of non-functional collagen. The hypothesis that nAChRα7 has a relevantinvolvement as a regulator of muscle inflammation and tissueremodeling is consistent because experimentally injured musclenAChRα7�/� mice did not show reduction on MMP-9 activity ormodulation of active-MMP-2 after PNU282987 treatment. Thesedata strongly suggest that metalloprotease balance is crucial fora switch between pro-inflammatory to tissue remodeling phe-notype, and that nAChRα7 selective activation positively influ-ence this phenomenon. Altogether the results indicate thatnAChRα7 activation may be considered as a potential target for

b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 634
pharmacological strategies to reduce inflammation and activatemechanisms of muscular tissue remodeling.
4. Experimental procedure
4.1. Animal care
The animal use procedures were approved by the institu-tional animal care and used according to the guidance ofEthics Committee for Handling Research Animals of theCommittee of Animal Experimentation from the FluminenseFederal University (CEUA-UFF, Brazil), and of the MedicalSchool of Universidad Autónoma de Madrid (UAM-Spain).Male mdx dystrophic and age-matched wild type non-dystrophic mice (both C57BL/10 background) were main-tained at the Cellular Pathology animal house facilities ofthe Institute of Biology, UFF, and male nAChRα7 knockout(�/� ) and age-matched nAChRα7 wild type (þ/þ) mice (bothC57BL/6 background) at Universidad Autónoma de Madrid.Mice were kept at constant temperature (21 1C) with a lightcycle of 12 h. Mdx and control mice were sacrificed in theperiod of inflammatory prevalence at 4 weeks, andnAChRα7�/� and respective control mice at 6 weeks age.
4.2. Inflammatory model – induction of muscular lesion
nAChRα7þ/þ and nAChRα7�/� (KO) mice at 6 wks of age wereanesthetized by intraperitoneal injection with ketamine(100 mg/Kg) and xylazine (10 mg/Kg). Muscular lesion wasperformed as previously described (Mussini et al., 1987) in thecentral region of gastrocnemius muscles. Ipsilateral musclewas injected with 60 mL 0.5% bupivacaine hydrochloride (bp)(Sigma-Aldrich, Spain), and the contralateral muscle (control-sham) with vehicle.
4.3. In vivo pharmacological treatment
Mdx mice at 3 wks (21 days postnatal – PN) received dailyintraperitoneal injections of 0.3 or 1 mg/kg PNU282987 (N-(3R)-1-Azabicyclo[2.2.2]oct-3-yl-4-chlorobenzamide) (TocrisBioscience, Bristol, UK) or vehicle saline solution during 7days. nAChRα7�/� and nAChRα7þ/þ mice with bp-inducedmuscular lesion were equally treated during 4 days. Groupswere also treated with the nAChRα7 antagonist methyllyca-conitine citrate ([1α,4(S),6β,14α,16β]-20-Ethyl-1,6,14,16-tetra-methoxy-4-[[[2-(3-methyl-2,5-dioxo-1-pyrrolidinyl)benzoyl]oxy]methyl]aconitane-7,8-diol citrate) (MLA 1029, TocrisBioscience, Bristol, UK) at 1 mg/kg or PNU282987 togetherwith MLA (both at 1 mg/kg). Mice were sacrificed at the end oftreatment for blood and tissue collection. At least 4 animalswere included in each group, and experiments were repeatedthree times.
4.4. Quantification of TNFα by ELISA assay
Mice sera used for determination of TNFα levels were quantifiedin triplicate by commercial enzyme-linked immunosorbentassay (ELISA) according to the manufacturer's instructions(PeproTech Inc., NJ, USA). Absorbance was assessed in a
microplate spectrophotometer (Thermo Scientific™ MultiskanFC, USA) at wavelength 450 nm and also 650 nm correction filterafter addition of 1% SDS stopping solution.
4.5. Gelatin zymography for detection of metalloproteaseactivity
Serum was immediately frozen in aliquots and preserved at�80 1C until use. Gastrocnemius muscles were removed,homogenized (TBS 1/10 w/v, 100 mM Tris–HCl pH 7.6,200 mM NaCl, 100 mM CaCl2 and 1% Triton X-100) thenclarified by centrifugation at 15,000g for 10 min at 10 1C, andprotein quantification determined by the Lowry method.Equal amounts of total protein were loaded (60 μg/lane formuscle; 20 μg/lane for serum) and metalloprotease activitydetermined by zymography as previously described (Kherifet al., 1999; Leite et al., 2010). Gelatinase activity was visua-lized as unstained bands on a blue background representingareas of proteolysis. Metalloproteases are secreted in a latentform and require cleavage of a NH2 terminus peptide foractivation. Exposure of proenzymes to SDS during gel separationleads to activation without proteolytic cleavage (Talhouk et al.,1992) with appearance of bands corresponding to 100-kDa (MMP-9), 66-kDa (pre-pro-MMP-2) and 60-kDa (pro-MMP-2), and lessfrequently 55-kDa (active-MMP-2) (Kherif et al., 1999). Semi-quantitative analysis was performed by using image analysissoftware (Scion Image, NIH, Bethesda, Md., USA).
4.6. Histological staining and morphometric analysis
Gastrocnemius muscles were removed and fixed in formalin-buffered Millonig fixative (pH 7.2) for 24 h. Sections (10 μmthick) of tissue embedded in paraplast (Sigma, St. Louis, Mo.,USA) were stained with hematoxylin and eosin and sirius redto analyze histological alterations and collagen depositionrespectively. Degenerating and necrotic fibers were identifiedby homogeneous pale eosinophilic sarcoplasm, whereasregenerating fibers were characterized by strong basophilicand centrally-located nuclei. High-definition whole-areaimages of all cross sections from a single mouse at each timepoint were acquired from individual photomicrographs witha microdigital camera mounted on a Zeiss Axioplan micro-scope (Zeiss, Oberkochen, Germany) by using a 20� objec-tive. Images were blended by using Adobe Photoshop CS3Extended software. Total surface area, areas occupied byinflammatory infiltrate, regenerating myofibers and collagendeposition were determined with Image-Pro 4.5 (MediaCybernetics). For analysis of perivascular inflammatory infil-trate nearby blood vessel, sections were also stained forisolectin B4 (Isolectin GS-IB4 1:100; Vector, Burlingame, CA,USA), and its extension in each image demarcated by whitedots. Results are expressed as percentage of total area in thecross-section.
4.7. Immunofluorescence
10 mm slices were washed with PBS and incubated for 1 h withblocking buffer (0.05% Triton X-100 in PBS, containing 5%normal goat serum). For labeling endothelial cells, sampleswere incubated for 2 h at 4 1C with biotinylated Griffonia

b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 6 35
simplicifolia I isolectin B4 (1:100; Vector, Burlingame, CA, USA),followed by 1 h incubation with streptavidin Cy3-conjugate(1:1000; Jackson Immuno Research; West Grove, PA, USA) atroom temperature and counterstained with Dapi. Imageswere obtained with a Nikon Eclipse TE2000-U microscope(Olympus, Tokyo, Japan) with identical time exposure andimage settings and merged on Adobe Photoshop CS5.
4.8. Assessment of muscle damage
Lactate dehydrogenase (LDH) release was used to verify celldeath by necrosis. LDH activity was measured with CytoTox96s colorimetric, non-radioactive, cytotoxicity assay (Pro-mega Corp. Madison, WI, USA). Briefly, serum was dilutedand incubated in a 1:2 dilution with substrate at roomtemperature for 30 minutes. Stop solution was added to eachwell and absorbance recorded with a 490 nm filter in an ELISAreader. Evans blue dye (10 mg/ml in PBS pH 7.4) injectedintraperitoneally 24 h before sacrifice allows observation ofin vivo myofiber damage (Crawford et al., 2000). At the end ofdrug treatment, mice were sacrificed and muscles wereremoved, frozen in OCT (Tissue-Tek, Elkhard, IN,USA) embed-ding medium, and Evans blue uptake by individual fibers in10 mm-thick frozen sections were fixed in cold acetone forimage analysis in a Nikon Eclipse TE2000-U microscope(Olympus, Tokyo, Japan) with identical time exposure andimage settings.
4.9. Quantitative and statistical analysis
Gelatin zymograms, histological staining and morphometrywere analyzed using Scion Image for Windows software(Scion Corporation, National Institutes of Health; Bethesda,MD). GraphPad Prism 5 (GraphPad software Inc.) was used tocalculate mean and standard errors. One-way ANOVA andunpaired t test were applied to obtain statistical significanceof means. Differences were considered to be statisticallysignificant at the 0.05 level of confidence.
Funding
This research was supported by Brazilian Foundations: Rio deJaneiro Research Foundation (FAPERJ), Coordenação de Aperfei-çoamento de Pessoal do Ensino Superior (CAPES-DGU Program),and REDOXOMA (INCT group of REDOX process in Biomedicine,CNPq/FAPESP), and Spanish Foundations: MINECO (Ministeriode Economía y Competitividad, Spain; SAF-2010-18837 to LG)and CEAL-Santander. The funders had no role in study design,data collection and analysis, decision to publish, or manuscriptpreparation.
Competing interest
The authors declare that have no competing interest.
Acknowledgments
This study was supported by grants from CAPES-DGU pro-gram (Coordenação de Aperfeiçoamento de Pessoal de NívelSuperior), FAPERJ (Fundação de Amparo a Pesquisa do Rio deJaneiro), MEC (Ministerio de Economía y Competitividad,Spain; SAF-2010-18837) and CEAL-Santander.
r e f e r e n c e s
Acharyya, S., Villalta, S.A., Bakkar, N., Bupha-Intr, T., Janssen, P.
M., Carathers, M., Li, Z.W., Beg, A.A., Ghosh, S., Sahenk, Z.,
Weinstein, M., Gardner, K.L., Rafael-Fortney, J.A., Karin, M.,
Tidball, J.G., Baldwin, A.S., Guttridge, D.C., 2007. Interplay of
IKK/NF-kappaB signaling in macrophages and myofibers
promotes muscle degeneration in Duchenne muscular
dystrophy. J. Clin. Invest. 117, 889–901.AlSharari, S.D., Freitas, K., Damaj, M.I., 2013. Functional role of
alpha7 nicotinic receptor in chronic neuropathic and
inflammatory pain: studies in transgenic mice. Biochem.
Pharmacol. 86, 1201–1207.Andersson, U., Tracey, K.J., 2012. Neural reflexes in inflammation
and immunity. J. Exp. Med. 209, 1057–1068.Bani, C., Lagrota-Candido, J., Pinheiro, D.F., Leite, P.E., Salimena,
M.C., Henriques-Pons, A., Quirico-Santos, T., 2008. Pattern of
metalloprotease activity and myofiber regeneration in skeletal
muscles of mdx mice. Muscle Nerve 37, 583–592.Bernik, T.R., Friedman, S.G., Ochani, M., DiRaimo, R., Ulloa, L.,
Yang, H., Sudan, S., Czura, C.J., Ivanova, S.M., Tracey, K.J., 2002.
Pharmacological stimulation of the cholinergic
antiinflammatory pathway. J. Exp. Med. 195, 781–788.Bobadilla, M., Sainz, N., Rodriguez, J.A., Abizanda, G., Orbe, J.,
deMartino, A., Verdugo, J.M.G., Paramo, J.A., Prosper, F., Perez-
Ruiz, A., 2014. MMP-10 is required for efficient muscle
regeneration in mouse models of injury and muscular
dystrophy. Stem cells 32, 447–461.Borovikova, L.V., Ivanova, S., Zhang, M., Yang, H., Botchkina, G.I.,
Watkins, L.R., Wang, H., Abumrad, N., Eaton, J.W., Tracey, K.J.,
2000. Vagus nerve stimulation attenuates the systemic
inflammatory response to endotoxin. Nature 405, 458–462.Carlson, C.G., 1998. The dystrophinopathies: an alternative to the
structural hypothesis. Neurobiol. Dis. 5, 3–15.Collins, C.A., Morgan, J.E., 2003. Duchenne’s muscular
dystrophy: animal models used to investigate
pathogenesis and develop therapeutic strategies. Int. J. Exp.
Pathol. 84, 165–172.Crawford, G.E., Faulkner, J.A., Crosbie, R.H., Campbell, K.P.,
Froehner, S.C., Chamberlain, J.S., 2000. Assembly of the
dystrophin-associated protein complex does not require the
dystrophin COOH-terminal domain. J. Cell Biol. 150,
1399–1410.Evans, N.P., Misyak, S.A., Robertson, J.L., Bassaganya-Riera, J.,
Grange, R.W., 2009. Immune-mediated mechanisms
potentially regulate the disease time-course of duchenne
muscular dystrophy and provide targets for therapeutic
intervention. Phys. Med. Rehabil. 1, 755–768.Gallowitsch-Puerta, M., Pavlov, V.A., 2007. Neuro-immune
interactions via the cholinergic anti-inflammatory pathway.
Life Sci. 80, 2325–2329.Hoffman, E.P., Brown Jr., R.H., Kunkel, L.M., 1987. Dystrophin: the
protein product of the Duchenne muscular dystrophy locus.
Cell 51, 919–928.Junger, W.G., 2011. Immune cell regulation by autocrine
purinergic signalling. Nat. Rev. Immunol. 11, 201–212.

b r a i n r e s e a r c h 1 5 7 3 ( 2 0 1 4 ) 2 7 – 3 636
Khandoga, A., Kessler, J.S., Hanschen, M., Khandoga, A.G.,Burggraf, D., Reichel, C., Hamann, G.F., Enders, G.,Krombach, F., 2006. Matrix metalloproteinase-9 promotesneutrophil and T cell recruitment and migration in thepostischemic liver. J. Leuk. Biol. 79, 1295–1305.
Kherif, S., Lafuma, C., Dehaupas, M., Lachkar, S., Fournier, J.G.,Verdiere-Sahuque, M., Fardeau, M., Alameddine, H.S., 1999.Expression of matrix metalloproteinases 2 and 9 inregenerating skeletal muscle: a study in experimentallyinjured and mdx muscles. Dev. Biol. 205, 158–170.
Lagrota-Candido, J., Vasconcellos, R., Cavalcanti, M., Bozza, M.,Savino, W., Quirico-Santos, T., 2002. Resolution of skeletalmuscle inflammation in mdx dystrophic mouse isaccompanied by increased immunoglobulin and interferon-gamma production. Int. J. Exp. Pathol. 83, 121–132.
Lagrota-Candido, J., Canella, I., Pinheiro, D.F., Santos-Silva, L.P.,Ferreira, R.S., Guimaraes-Joca, F.J., Lannes-Vieira, J., Quirico-Santos, T., 2010. Characteristic pattern of skeletal muscleremodelling in different mouse strains. Int. J. Exp. Pathol. 91,522–529.
Leite, P.E., Lagrota-Candido, J., Moraes, L., D’Elia, L., Pinheiro, D.F.,da Silva, R.F., Yamasaki, E.N., Quirico-Santos, T., 2010.Nicotinic acetylcholine receptor activation reduces skeletalmuscle inflammation of mdx mice. J. Neuroimmunol. 227,44–51.
Li, H., Mittal, A., Makonchuk, D.Y., Bhatnagar, S., Kumar, A., 2009.Matrix metalloproteinase-9 inhibition amelioratespathogenesis and improves skeletal muscle regeneration inmuscular dystrophy. Hum. Mol. Genet. 18, 2584–2598.
Maddahi, A., Povlsen, G.K., Edvinsson, L., 2012. Regulation ofenhanced cerebrovascular expression of proinflammatorymediators in experimental subarachnoid hemorrhage via themitogen-activated protein kinase kinase/extracellular signal-regulated kinase pathway. J. Neuroinflammation 9, 274–290.
Murray, P.J., Smale, S.T., 2012. Restraint of inflammatory signalingby interdependent strata of negative regulatory pathways.Nat. Immunol. 13, 916–924.
Mussini, I., Favaro, G., Carraro, U., 1987. Maturation, dystrophicchanges and the continuous production of fibers in skeletalmuscle regenerating in the absence of nerve. J. Neuropathol.Exp. Neurol. 46, 315–331.
Olofsson, P.S., Rosas-Ballina, M., Levine, Y.A., Tracey, K.J., 2012.Rethinking inflammation: neural circuits in the regulation ofimmunity. Immunol. Rev. 248, 188–204.
Osborne-Hereford, A.V., Rogers, S.W., Gahring, L.C., 2008.Neuronal nicotinic alpha7 receptors modulate inflammatorycytokine production in the skin following ultraviolet radiation.J. Neuroimmunol. 193, 130–139.
Parada, E., Egea, J., Buendia, I., Negredo, P., Cunha, A.C., Cardoso,S., Soares, M.P., Lopez, M.G., 2013. The microglial
α7-acetylcholine nicotinic receptor is a key element in
promoting neuroprotection by inducing heme oxygenase-1via nuclear factor erythroid-2-related factor 2. Antioxid. Redox
Signal. 19, 1135–1148.Parrish, W.R., Rosas-Ballina, M., Gallowitsch-Puerta, M.,
Ochani, M., Ochani, K., Yang, L.H., Hudson, L., Lin, X., Patel, N.,
Johnson, S.M., Chavan, S., Goldstein, R.S., Czura, C.J., Miller, E.
J., Al-Abed, Y., Tracey, K.J., Pavlov, V.A., 2008. Modulation ofTNF release by choline requires alpha7 subunit nicotinic
acetylcholine receptor-mediated signaling. Mol. Med. 14,567–574.
Pavlov, V.A., Tracey, K.J., 2012. The vagus nerve and the
inflammatory reflex-linking immunity and metabolism. Nat.Rev. Endocrinol. 8, 743–754.
Pena, G., Cai, B., Ramos, L., Vida, G., Deitch, E.A., Ulloa, L., 2011.
Cholinergic regulatory lymphocytes re-establishneuromodulation of innate immune responses in sepsis.
J. Immunol. 187, 718–725.Rosas-Ballina, M., Olofsson, P.S., Ochani, M., Valdes-Ferrer, S.I.,
Levine, Y.A., Reardon, C., Tusche, M.W., Pavlov, V.A.,
Andersson, U., Chavan, S., Mak, T.W., Tracey, K.J., 2011.
Acetylcholine-synthesizing T cells relay neural signals in avagus nerve circuit. Science 334, 98–101.
Rullman, E., Rundqvist, H., Wagsater, D., Fischer, H., Eriksson, P.,
Sundberg, C.J., Jansson, E., Gustafsson, T., 2007. A single boutof exercise activates matrix metalloproteinase in human
skeletal muscle. J. Appl. Physiol. 102, 2346–2351.Schiøtz Thorud, H.M., Stranda, A., Birkeland, J.-A., Lunde, P.K.,
Sjaastad, I., Kolset, S.O., Sejersted, O.M., Iversen, P.O., 2005.
Enhanced matrix metalloproteinase activity in skeletalmuscles of rats with congestive heart failure. Am. J. Physiol.
289, R389–R394.Talhouk, R.S., Bissell, M.J., Werb, Z., 1992. Coordinated expression
of extracellular matrix-degrading proteinases and their
inhibitors regulates mammary epithelial function duringinvolution. J. Cell Biol. 118, 1271–1282.
Tracey, K.J., 2002. The inflammatory reflex. Nature 420, 853–859.Trakhtenberg, E.F., Goldberg, J.L., 2011. Neuroimmune
Communication. Science 334, 47–48.Ulloa, L., 2013. The cholinergic anti-inflammatory pathway meets
microRNA. Cell Res. 23, 1249–1250.van Maanen, M.A., Lebre, M.C., van der Poll, T., LaRosa, G.J.,
Elbaum, D., Vervoordeldonk, M.J., Tak, P.P., 2009. Stimulation of
nicotinic acetylcholine receptors attenuates collagen-inducedarthritis in mice. Arthritis Rheum. 60, 114–122.
Wang, H., Liao, H., Ochani, M., Justiniani, M., Lin, X., Yang, L.,
Al-Abed, Y., Metz, C., Miller, E.J., Tracey, K.J., Ulloa, L., 2004.Cholinergic agonists inhibit HMGB1 release and improve
survival in experimental sepsis. Nat. Med. 10, 1216–1221.