sel-t, sel- b dan mhc dosen imunologi fakultas farmasi universitas pancasila jakarta
TRANSCRIPT

SEL-T, Sel- B dan MHC
• Dosen Imunologi• Fakultas Farmasi Universitas
Pancasila• Jakarta
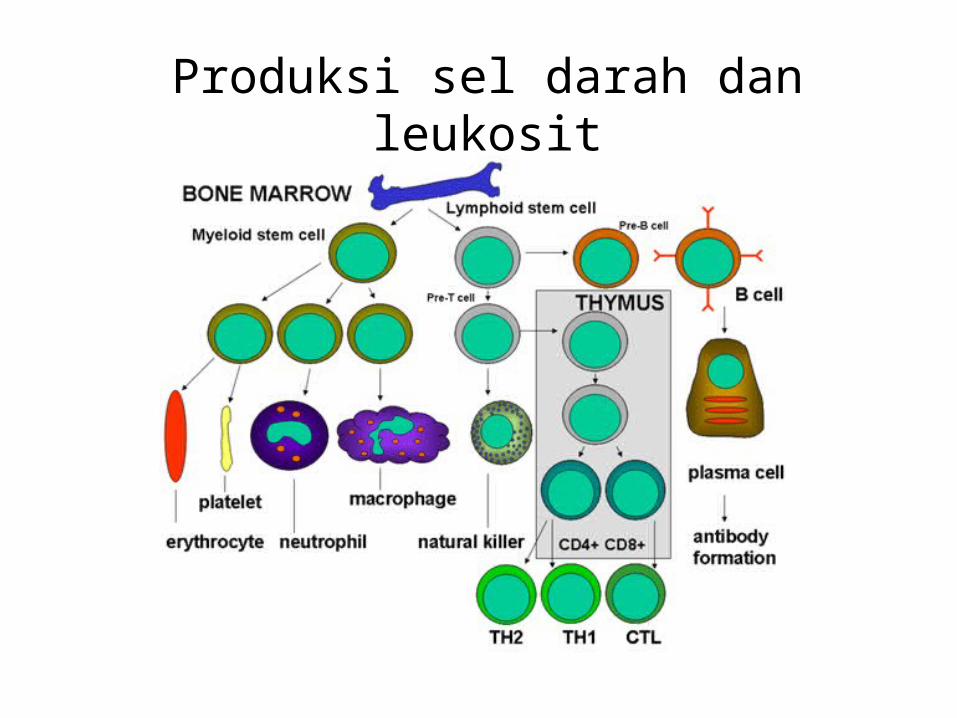
Produksi sel darah dan leukosit

Cell-Mediated Immunity
• Adalah respon adaptiv imuniti yg di mediasi oleh sel sistem imun spesifik:– lymphocytes T Primer (T cells), juga sel macrophages dan
sel NK .– Adalah immunitas yg dapat ditransfer dr satu organisme
ke organisme lain oleh sel limfoid, tetapi tdk dengan antibodi serum.
• Sel-T adalah agen utama dari selular imuniti.y

Sel-T • Adalah koordinator utama dan effektor
dari komponen selular imuniti
• Terciri oleh perkembangannya di dalam Thymus dan adanya T-cell receptor (TCR) complex
CD8+ cytotoxic T cell killing a tumor cell

Sel-T• Ada dua tipe utama:
1. CD4+: Stimulate other immune cells.2. CD8+ Cytotoxic T cells: Kill intracellularly-infected cells.
• Ada dua tipe utama dari CD4+ T cells:1. TH1: Inflammatory T cells -- Stimulate macrophages and promote inflammatory responses.
2. TH2: Helper T cells -- Stimulate B-cells to produce antibodies.
(A third type, TH3, has recently been shown to promote IgA production.)
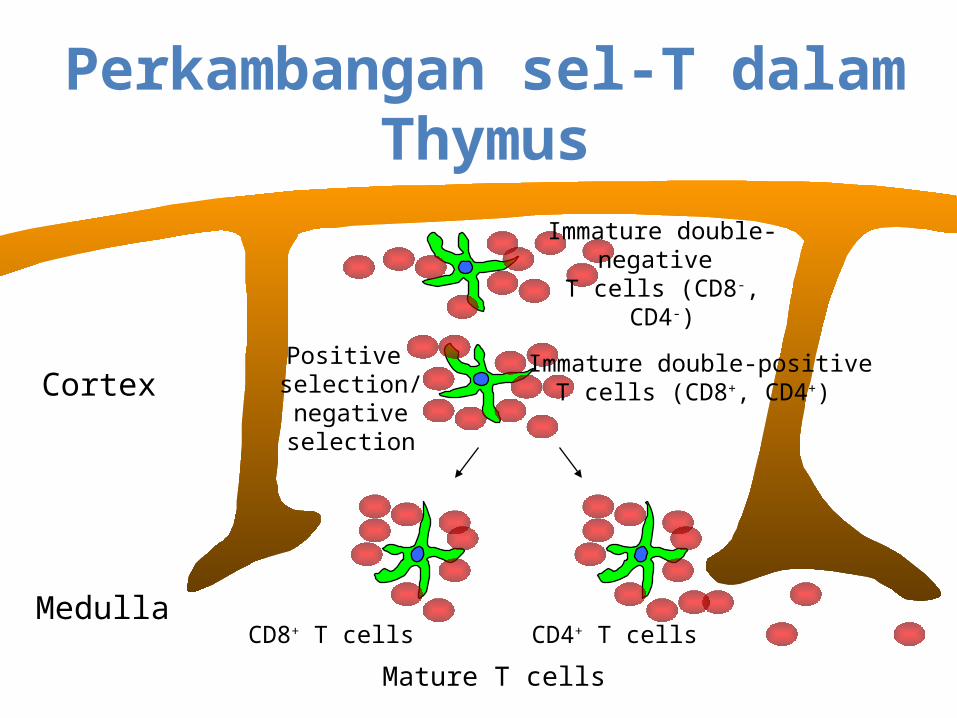
Perkambangan sel-T dalam Thymus
Cortex
Medulla
Immature double-negative
T cells (CD8-, CD4-)
Immature double-positiveT cells (CD8+, CD4+)
Positive selection/negativeselection
CD8+ T cells CD4+ T cells
Mature T cells
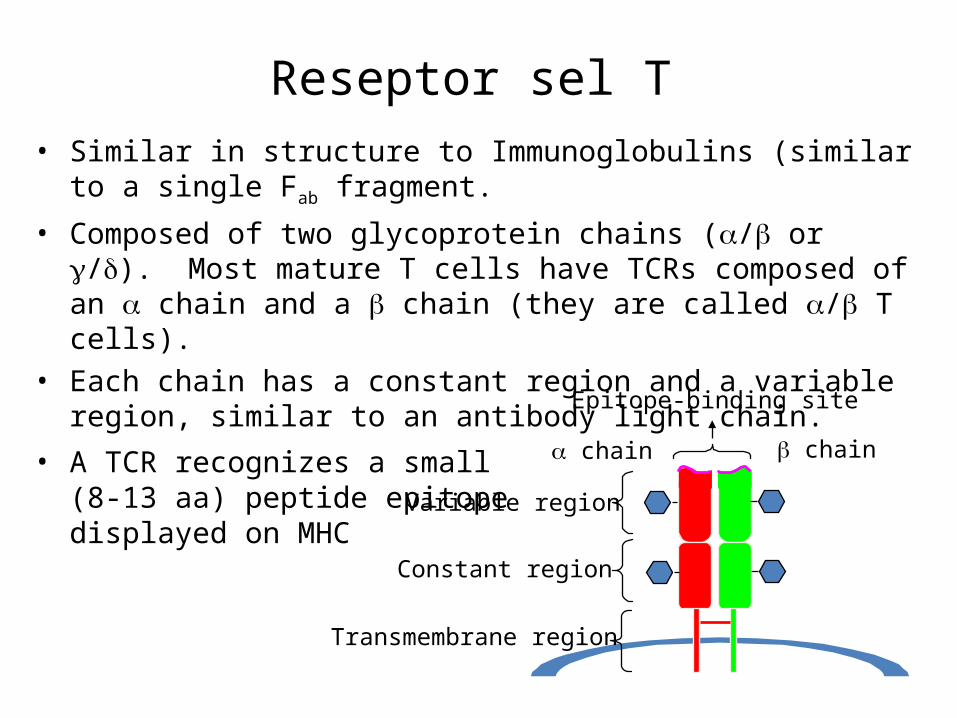
Reseptor sel T• Similar in structure to Immunoglobulins (similar to a single Fab fragment.
• Composed of two glycoprotein chains (/ or /). Most mature T cells have TCRs composed of an chain and a chain (they are called / T cells).
• Each chain has a constant region and a variable region, similar to an antibody light chain.
• A TCR recognizes a small(8-13 aa) peptide epitopedisplayed on MHC chain chain
Epitope-binding site
Variable region
Constant region
Transmembrane region

TCR compared to ImmunoglobulinsSimilarities• Both have specific Antigen-binding region created by the variable regions of two
polypeptide chains.• Both display great potential for diversity via genetic recombination at the
genome level
Differences• A TCR is monovalent (has one binding site). An Ig is bivalent (has two binding
sites).• The TCR has no secreted form. It is always membrane-bound.• The TCR does not recognize free antigen. Antigen must be presented to a T cell
on an MHC molecule • There is no class switching for the TCR. Once made, the TCR does not change.
chain chain
Epitope-binding site
Variable region
Constant region
Transmembrane region
T cell Receptor Immunoglobulin
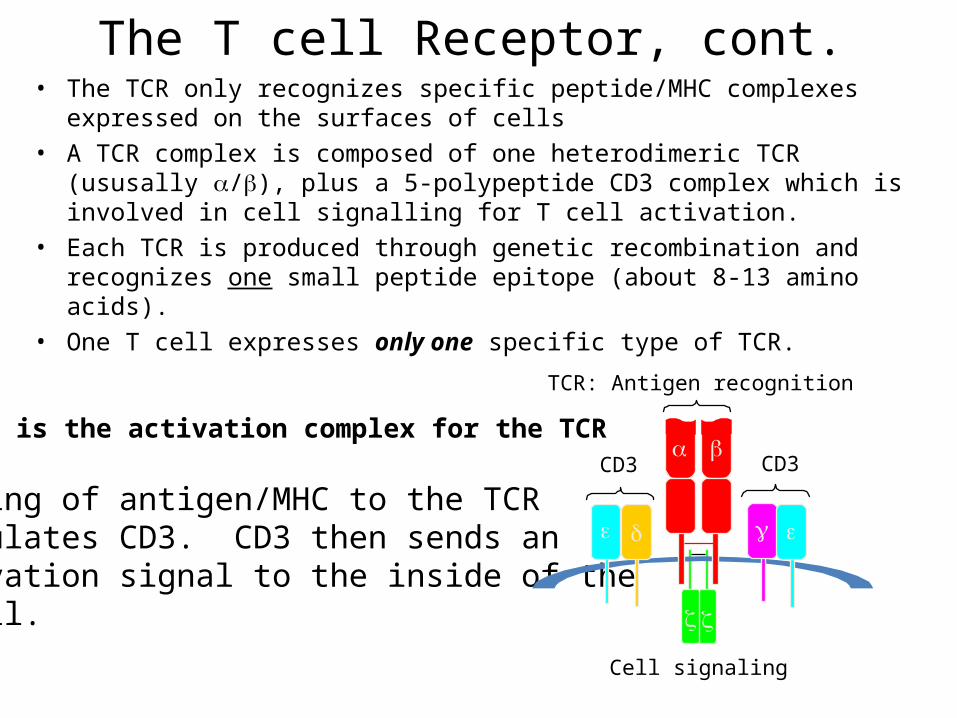
The T cell Receptor, cont.• The TCR only recognizes specific peptide/MHC complexes expressed on the
surfaces of cells• A TCR complex is composed of one heterodimeric TCR (ususally /), plus a
5-polypeptide CD3 complex which is involved in cell signalling for T cell activation.
• Each TCR is produced through genetic recombination and recognizes one small peptide epitope (about 8-13 amino acids).
• One T cell expresses only one specific type of TCR.
CD3 is the activation complex for the TCR
Binding of antigen/MHC to the TCRstimulates CD3. CD3 then sends an activation signal to the inside of theT cell.
TCR: Antigen recognition
CD3CD3
Cell signaling
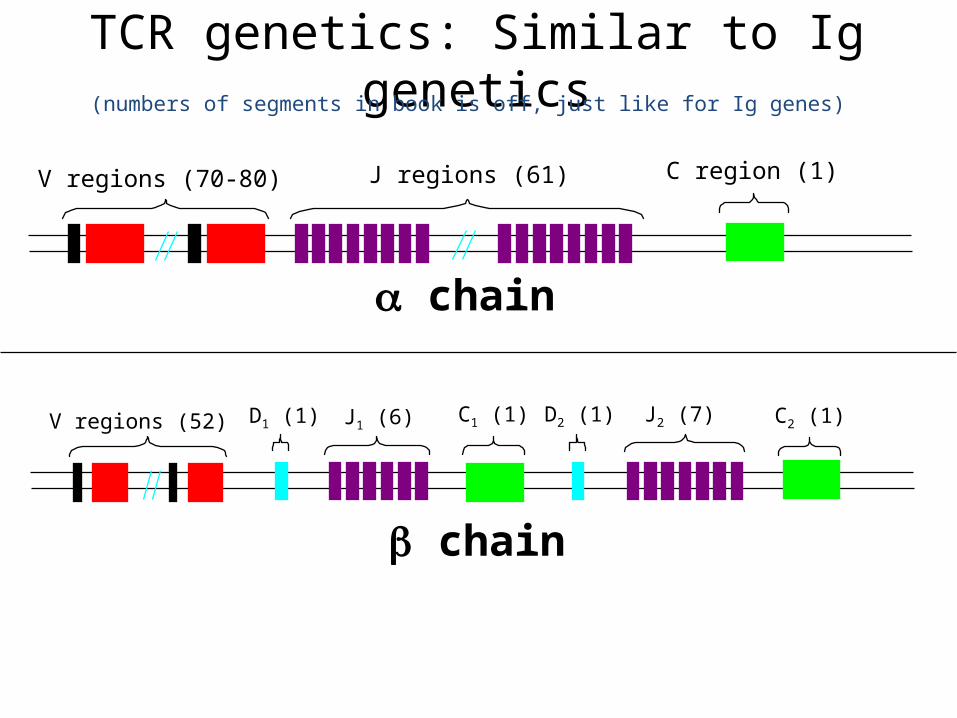
TCR genetics: Similar to Ig genetics
V regions (70-80) J regions (61) C region (1)
V regions (52) J1 (6)D1 (1) C1 (1) D2 (1) J2 (7) C2 (1)
(numbers of segments in book is off, just like for Ig genes)
chain
chain
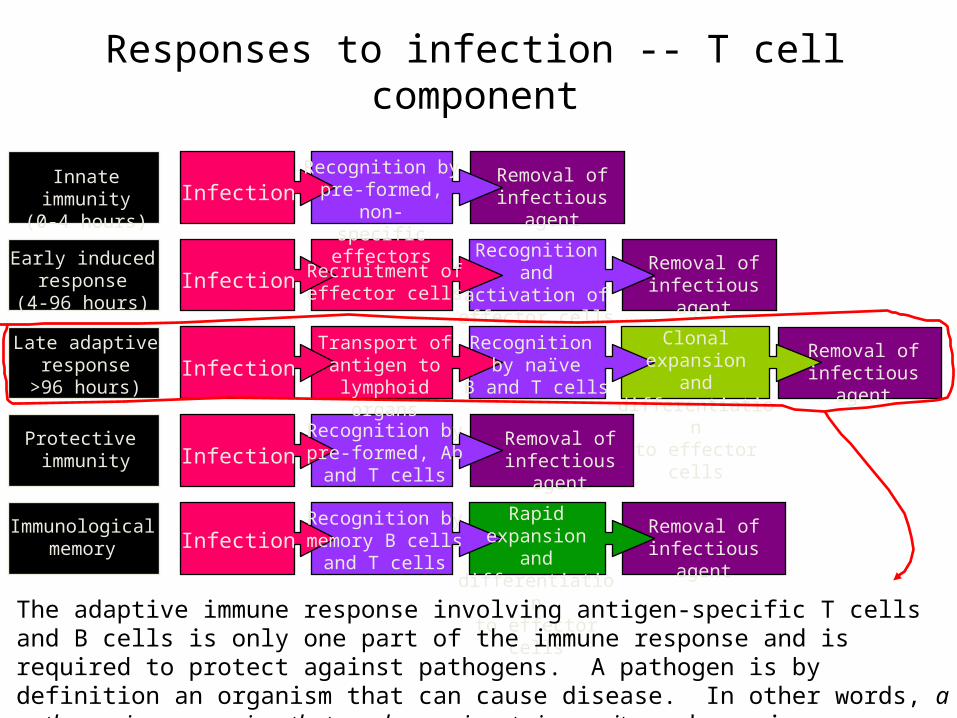
Responses to infection -- T cell component
Infection
Infection
Infection
Infection
Infection
Innate immunity(0-4 hours)
Early inducedresponse
(4-96 hours)
Late adaptiveresponse
>96 hours)
Protective immunity
Immunologicalmemory
Recognition bypre-formed, non-specific effectors
Recruitment ofeffector cells
Transport ofantigen to
lymphoid organs
Recognition bypre-formed, Ab
and T cells
Recognition bymemory B cells
and T cells
Removal ofinfectious agent
Removal ofinfectious agent
Removal ofinfectious agent
Removal ofinfectious agent
Removal ofinfectious agent
Recognition andactivation ofeffector cells
Recognition by naïve
B and T cells
Clonal expansionand differentiation
to effector cells
Rapid expansionand differentiation
to effector cells
The adaptive immune response involving antigen-specific T cells and B cells is only one part of the immune response and is required to protect against pathogens. A pathogen is by definition an organism that can cause disease. In other words, a pathogen is an organism that can bypass innate immunity and requires an adaptive immune response for clearance.

Generation of an adaptive immune response
• During an adaptive immune response,T cells which recognize specific antigen(s) are selected for differentiation into armed effector cells which undergo clonal expansion to produce a battery of antigen-specific cells.
• Clonal expansion refers to the process by which antigen-specific T cells or B cells are stimulated to reproduce clones of themselves to increase the system’s repertoire of antigen-specific effectors.
• Activation of antigen-specific T cells (the initiation of the adaptive response) occurs in the secondary lymph tissues (lymph nodes and spleen).
• This activation depends upon antigen presentation by a professional antigen presenting cell (APC) along with simultaneous co-stimulation. (eg., B7 on the APC, CD28 on the T cell).
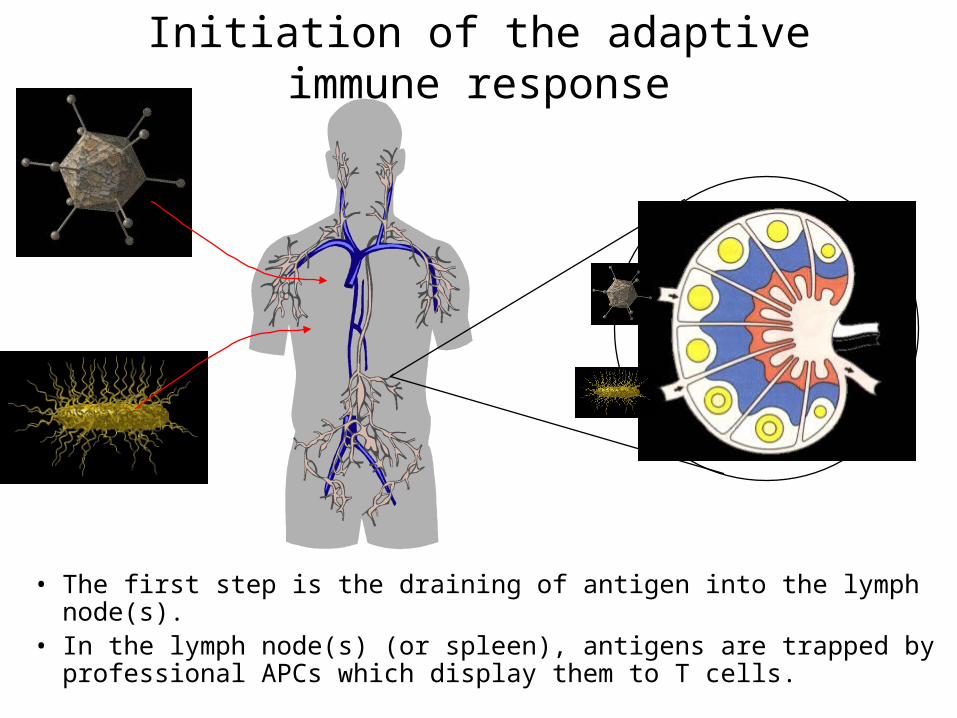
Initiation of the adaptive immune response
• The first step is the draining of antigen into the lymph node(s).• In the lymph node(s) (or spleen), antigens are trapped by
professional APCs which display them to T cells.

The professional Antigen Presenting Cells (APCs)• Three types of APC are found in the lymph nodes:
– Dendritic cells -- constitutively express MHC I and MHC II (can stimulate both CD4+ and CD8+ T cells) as well as B7 (the co-stimulatory signal). Antigen presentation appears to be the sole purpose of dendritic cells, and these cells can be infected by a wide variety of viruses. Dendritic cells are not phagocytic. They can present some viral peptides on their MHC II, and contribute to the induction of antibody against viruses. They are very efficient at stimulation of cytotoxic responses.
– Macrophages -- Resting macrophages express little MHC II or B7, but have receptors for bacterial cell wall components which, upon binding, activate the macrophage to express high levels of B7 and MHC II. Once activated, macrophages are efficient at stimulating CD4+ T cells, both for inflammatory responses and helper (antibody) responses.
– B cells -- B cells express high levels of MHC II, but not B7. Microbial cell wall components can induce B7 expression by B cells (like macrophages). Once induced to express B7, B cells can activate helper T cells. B cells can take up soluble antigen through their Ig receptors (unlike dendritic cells or macrophages).
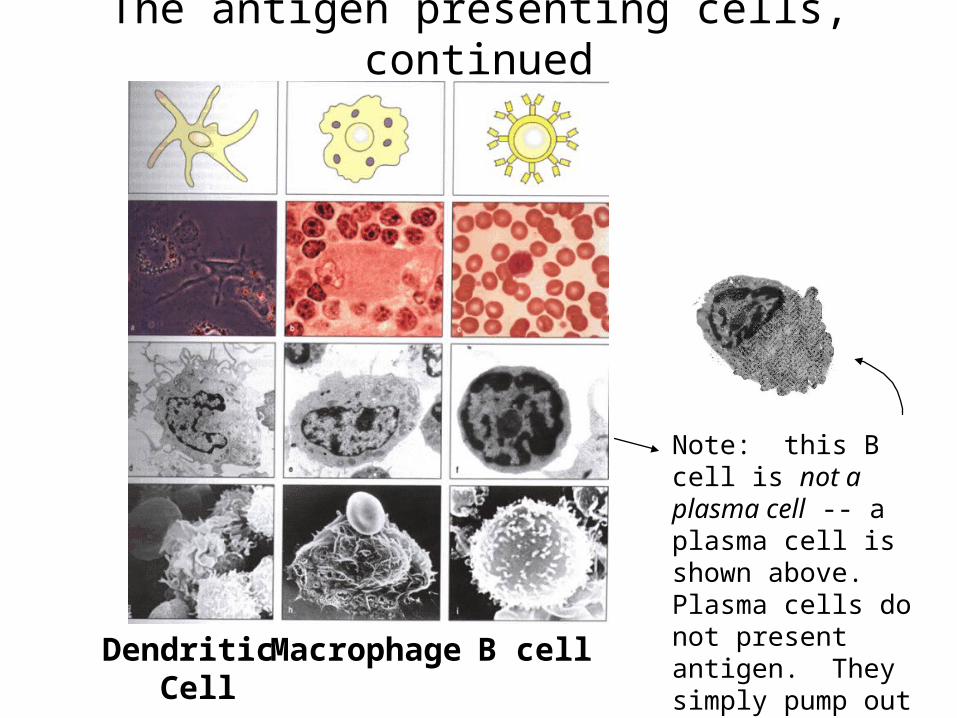
The antigen presenting cells, continued
Dendritic Cell
Macrophage B cell
Note: this B cell is not a plasma cell -- a plasma cell is shown above. Plasma cells do not present antigen. They simply pump out antibody for a few days then die.
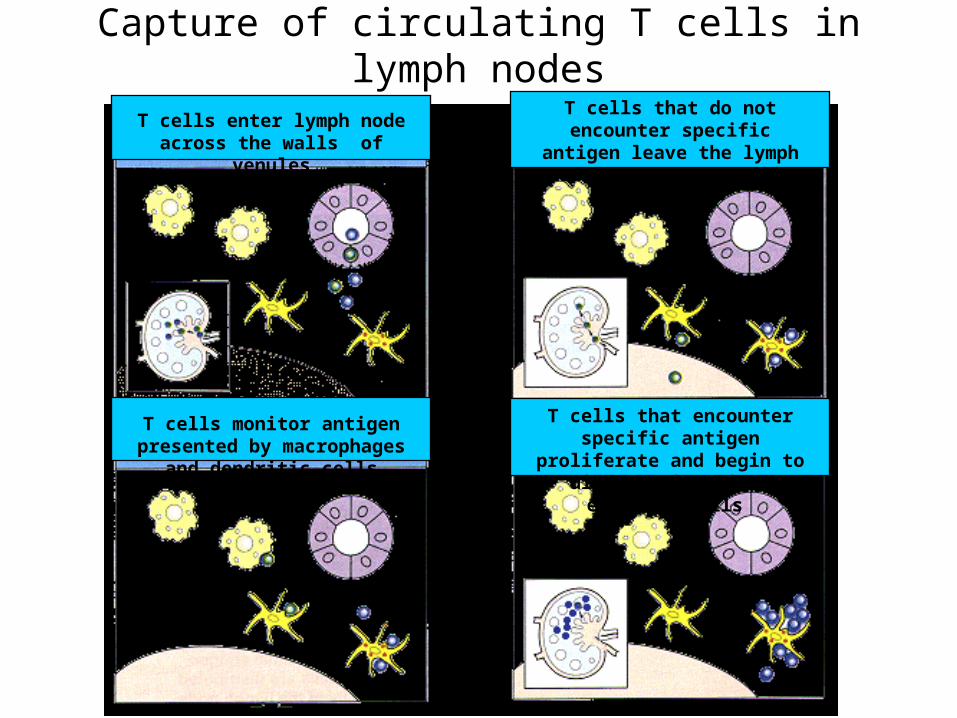
Capture of circulating T cells in lymph nodes
T cells enter lymph node across the walls of venules
T cells monitor antigen presented by macrophages and dendritic cells
T cells that do not encounter specific antigen leave the lymph node through lymphatic vessels
T cells that encounter specific antigen proliferate and begin to differentiate into effector cells
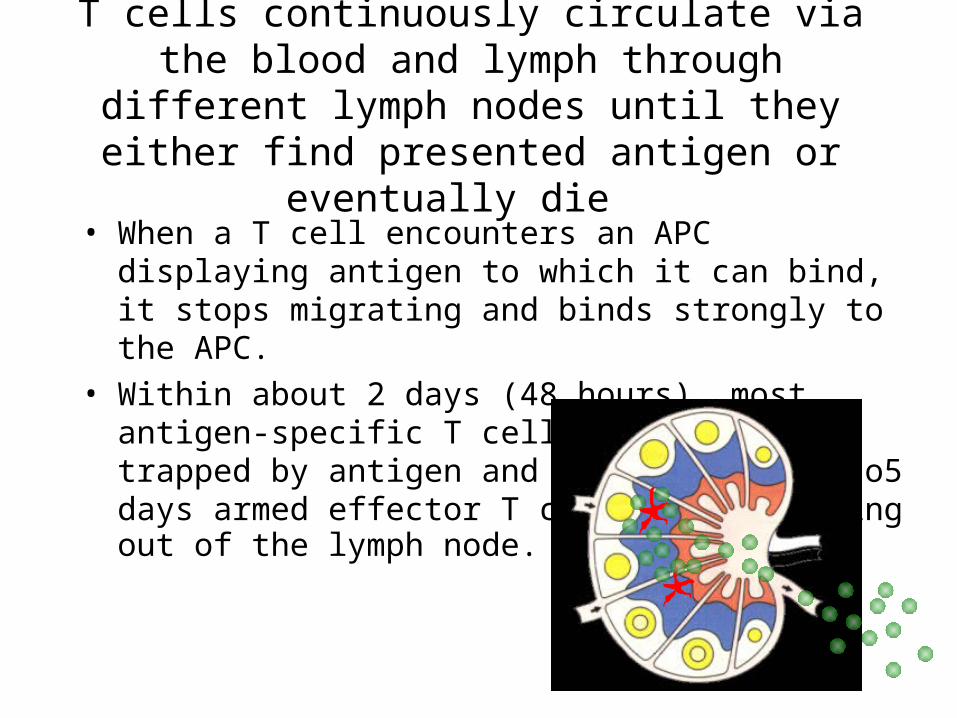
T cells continuously circulate via the blood and lymph through different lymph nodes until they either find presented antigen or eventually die
• When a T cell encounters an APC displaying antigen to which it can bind, it stops migrating and binds strongly to the APC.
• Within about 2 days (48 hours), most antigen-specific T cells have been trapped by antigen and within about 4 to5 days armed effector T cells are migratingout of the lymph node.
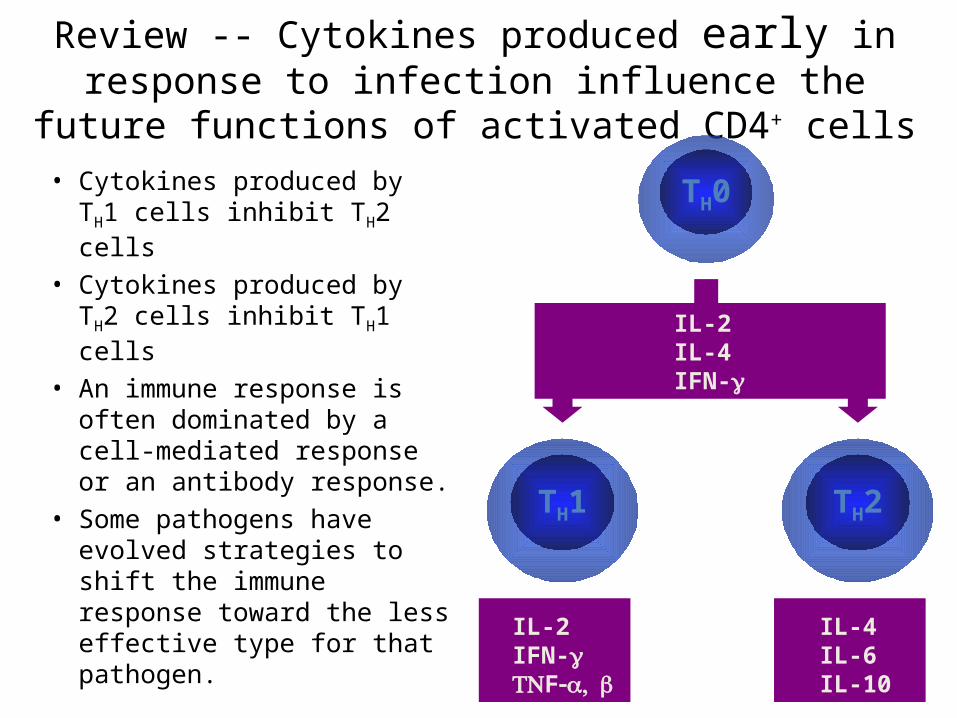
Review -- Cytokines produced early in response to infection influence the future functions of activated CD4+ cells
• Cytokines produced by TH1 cells inhibit TH2 cells
• Cytokines produced by TH2 cells inhibit TH1 cells
• An immune response is often dominated by a cell-mediated response or an antibody response.
• Some pathogens have evolved strategies to shift the immune response toward the less effective type for that pathogen.
TH0
TH2TH1
IL-2IL-4IFN-
IL-2IFN-F
IL-4IL-6IL-10
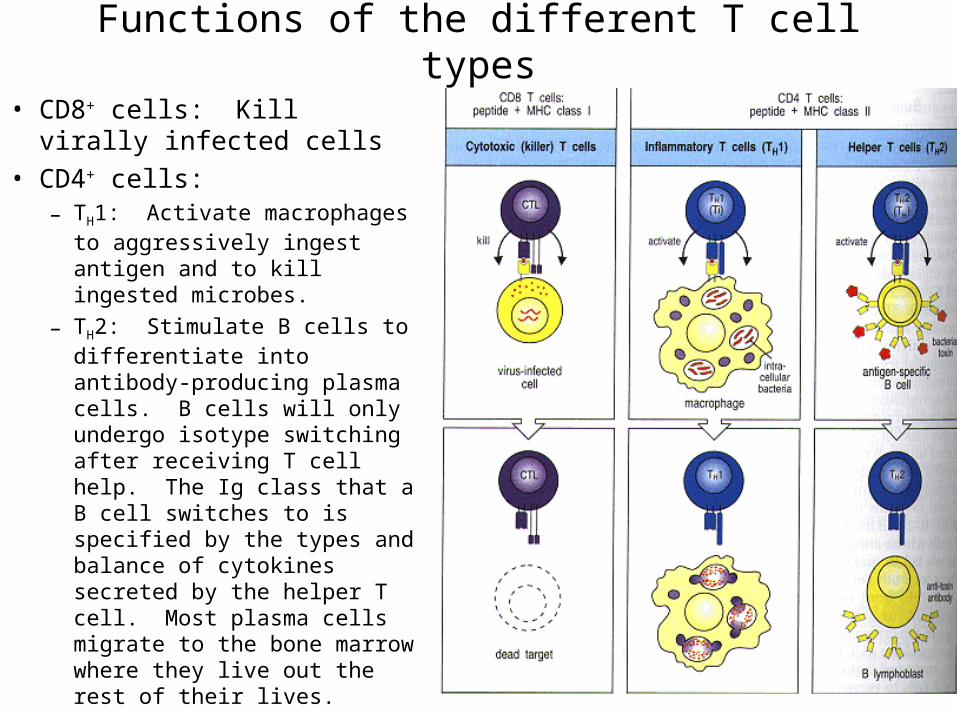
Functions of the different T cell types
• CD8+ cells: Kill virally infected cells
• CD4+ cells: – TH1: Activate macrophages to
aggressively ingest antigen and to kill ingested microbes.
– TH2: Stimulate B cells to differentiate into antibody-producing plasma cells. B cells will only undergo isotype switching after receiving T cell help. The Ig class that a B cell switches to is specified by the types and balance of cytokines secreted by the helper T cell. Most plasma cells migrate to the bone marrow where they live out the rest of their lives.
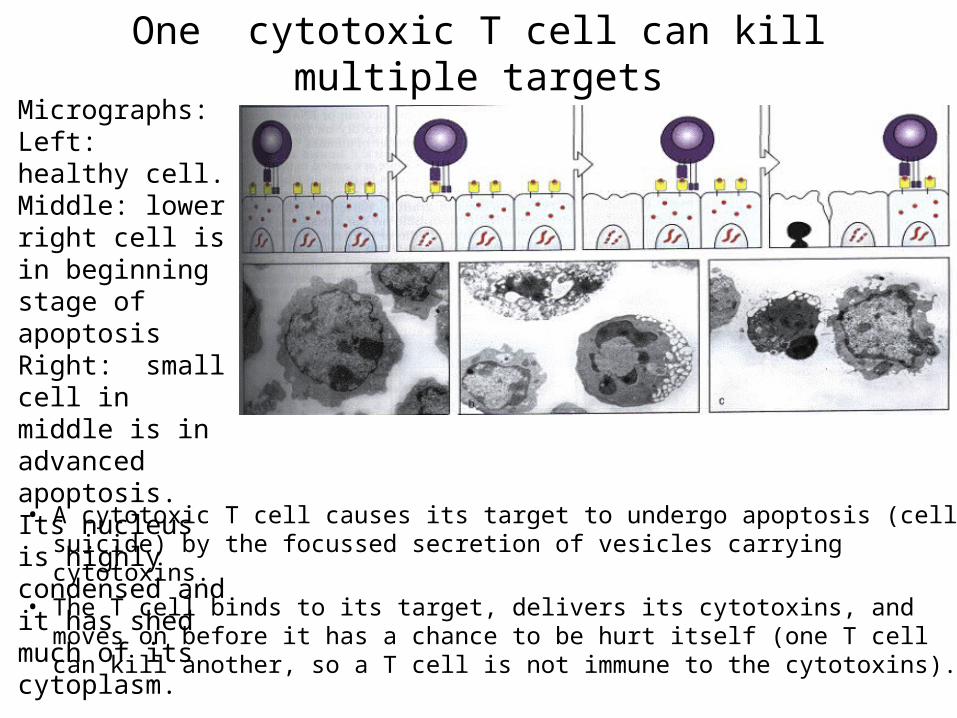
One cytotoxic T cell can kill multiple targets
• A cytotoxic T cell causes its target to undergo apoptosis (cell suicide) by the focussed secretion of vesicles carrying cytotoxins.
• The T cell binds to its target, delivers its cytotoxins, and moves on before it has a chance to be hurt itself (one T cell can kill another, so a T cell is not immune to the cytotoxins).
Micrographs:Left: healthy cell.Middle: lower right cell is in beginning stage of apoptosisRight: small cell in middle is in advanced apoptosis. Its nucleus is highly condensed and it has shed much of its cytoplasm.

Immunological memory
• When B cells are activated to reproduce, some differentiate into plasma cells and some become long-term memory cells.
• An adaptive immune response also produces T cell memory, but the nature of memory T cells is unknown. Two possibilities exist. Memory T cells probably originate from either:– 1. A long-lived subset of effector T cells that differentiates into
memory T cells -- like memory B cells.– 2. The continuous low-level activation of naïve T cells by specific
antigen that is retained in the lymph nodes after an infection. This mechanism would suggest that APCs in the lymph node hold on to antigen on a long-term basis after an infection and continuously stimulate T cells at a low level so there is always a small effector population ready to go.

MHC classes I and IIFunctions:• class I MHC:
– Displays peptides derived from antigen originating inside the cell (endogenous antigen).
– Important in cytotoxic responses (eg, CD8+-killing of virus-infected cells).• Class II MHC:
– Displays antigen derived from ingested antigens (exogenous antigen).– Important in humoral (antibody) responses as well in fighting as some intracellular
parasites (eg. Mycobacterium tuberculosis and M. leprae)
• Locations:– Class I MHC found on all nucleated cells (all cells need to be prepared to be killed in
case of a viral take-over or tumorigenic transformation).– Class II MHC found only on antigen presenting cells (cells that present antigen to
CD4+ T cells --> Macrophages, activated B-cells, dendritic cells.

Antigen Presentation to T cells: MHC
• Antigens are presented to T cells as short peptide fragments bound to Major Histocompatibility (MHC) molecules.
• Two types of MHC in humans and mice:– MHC I: presents an 8-10 amino acid peptide to CD8+ T
cells.– MHC II: presents a longer peptide (13 aa or more) to CD4+
T cells.
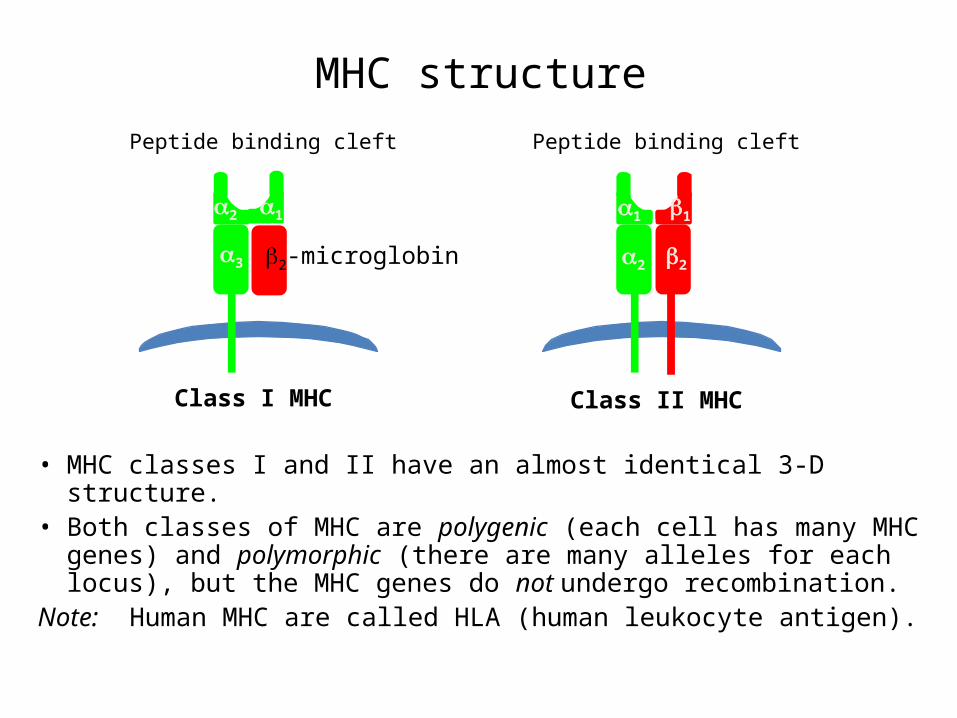
MHC structure
• MHC classes I and II have an almost identical 3-D structure.• Both classes of MHC are polygenic (each cell has many MHC
genes) and polymorphic (there are many alleles for each locus), but the MHC genes do not undergo recombination.
Note: Human MHC are called HLA (human leukocyte antigen).
Peptide binding cleft
1
2 2
1
Class II MHC
Peptide binding cleft
12
3 2-microglobin
Class I MHC
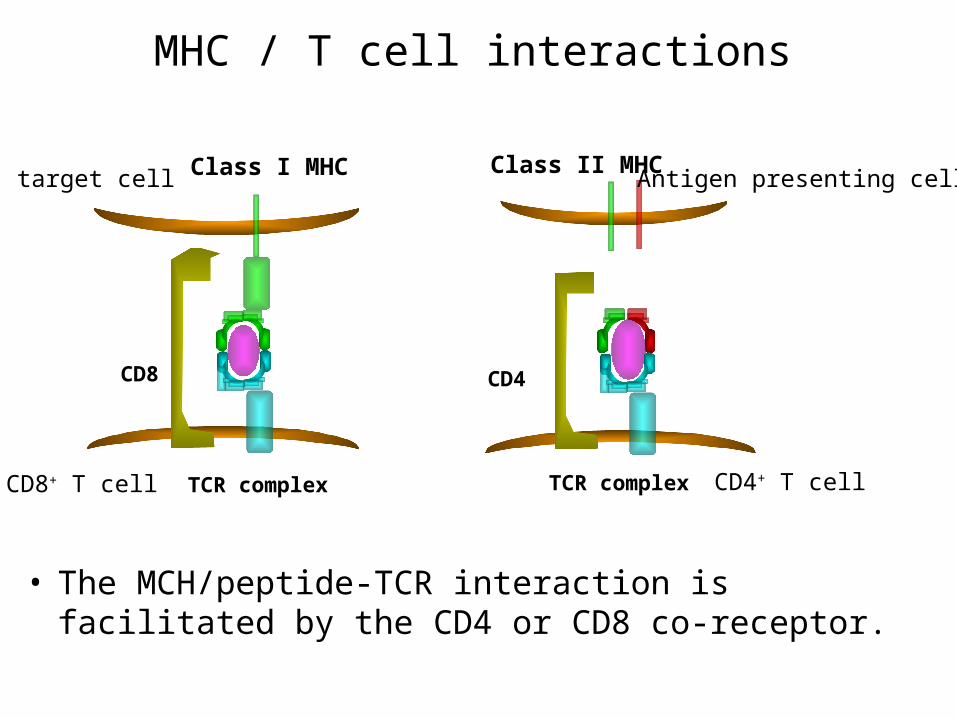
MHC / T cell interactions
• The MCH/peptide-TCR interaction is facilitated by the CD4 or CD8 co-receptor.
Class II MHCClass I MHC
TCR complex
CD8
CD8+ T cell
target cell
CD4
Antigen presenting cell
TCR complex CD4+ T cell
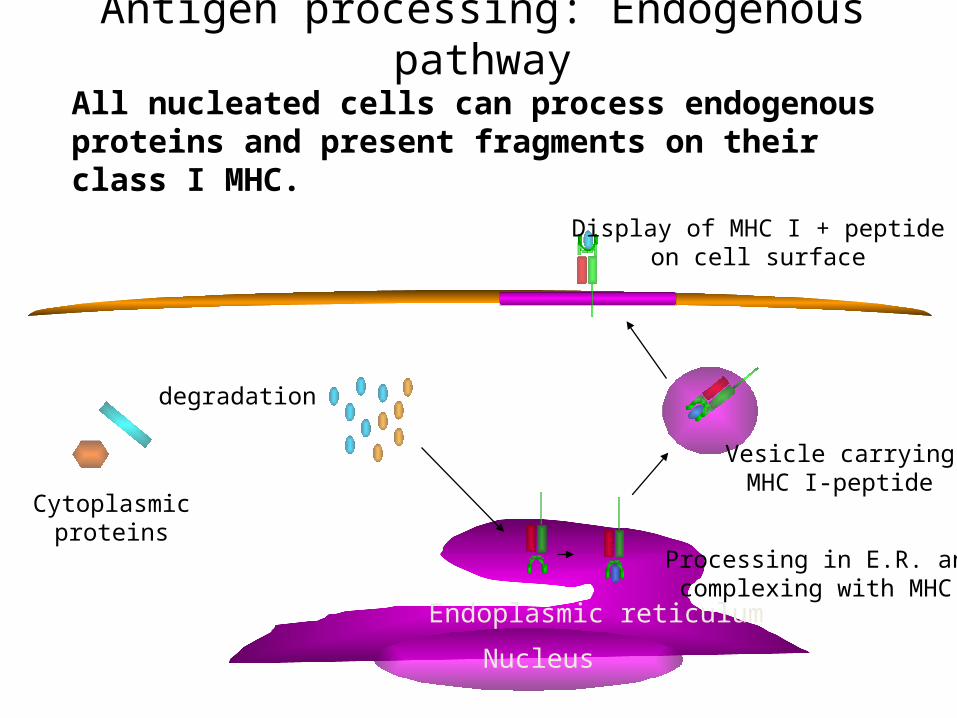
Antigen processing: Endogenous pathway
All nucleated cells can process endogenous proteins and present fragments on their class I MHC.
Endoplasmic reticulum
Nucleus
Cytoplasmicproteins
degradation
Vesicle carryingMHC I-peptide
Processing in E.R. and complexing with MHC I
Display of MHC I + peptideon cell surface
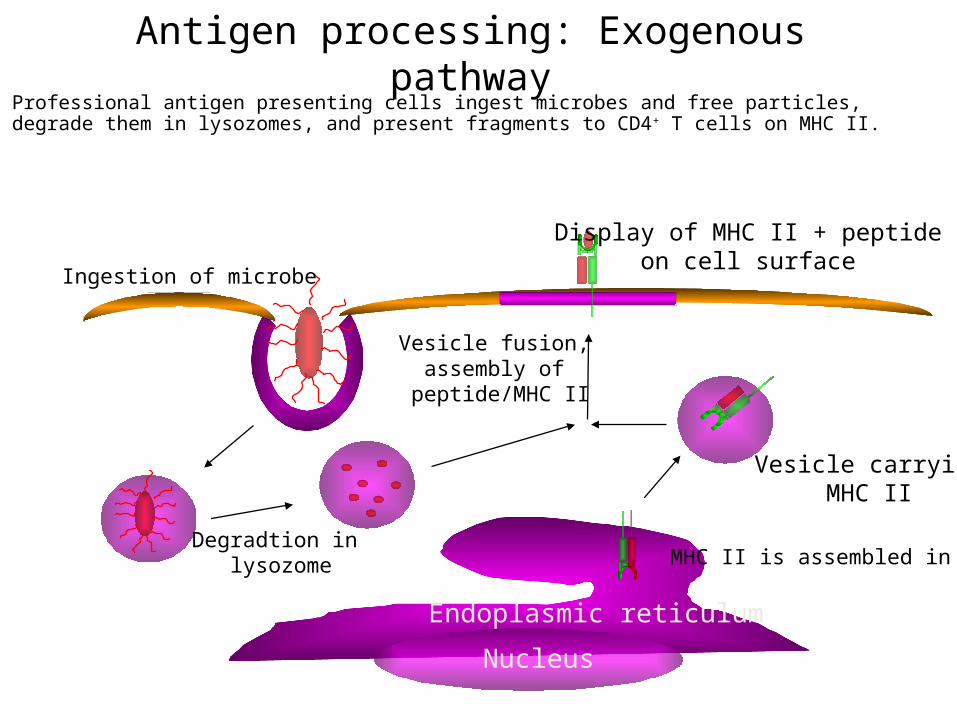
Antigen processing: Exogenous pathwayProfessional antigen presenting cells ingest microbes and free particles, degrade them in lysozomes, and present fragments to CD4+ T cells on MHC II.
Endoplasmic reticulum
Nucleus
Vesicle carryingMHC II
MHC II is assembled in ER
Display of MHC II + peptideon cell surface
Ingestion of microbe
Degradtion in lysozome
Vesicle fusion, assembly of
peptide/MHC II

CD4+ T cell activation
• T cells require co-stimulation for activation -- binding of the TCR to MHC/peptide is not enough to activate a T cell by itself.
• B7 on an APC binds to CD28 on the T cell to deliver a co-stimulatory signal. .
• Activation by peptide/MHC-TCR binding plus a co-stimulatory signal leads to Interleukin-2 (IL-2) release and up-regulation of the IL-2 receptor on the T cell.
• IL-2 stimulates growth and proliferation of T cells.

CD8+ T cell activation
• A naïve circulating CD8+ T cell also requires co-stimulation to become an “armed” effector cell.
• A CD8+ T cell can be activated by an APC displaying MHC I/peptide along with B7 (CD8+ cells also have CD28).
• Activation of the CD8+ cell causes upregulation of the IL-2 receptor and production of IL-2, leading to growth and proliferation.
• An activated CD8+ T cell can sustain itself on its own IL-2 production, once activated.
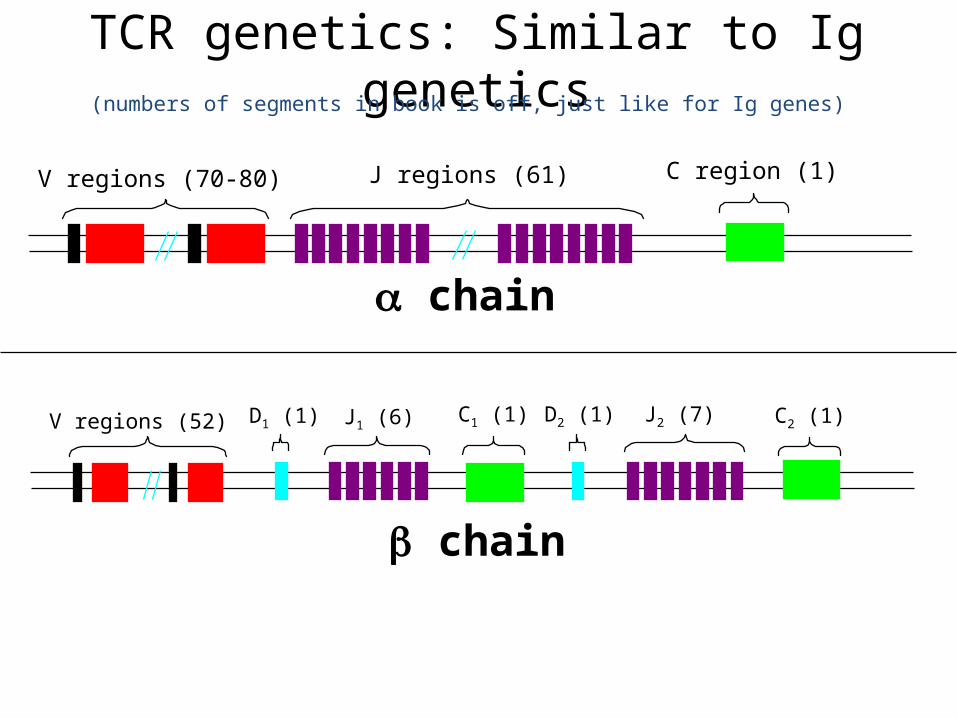
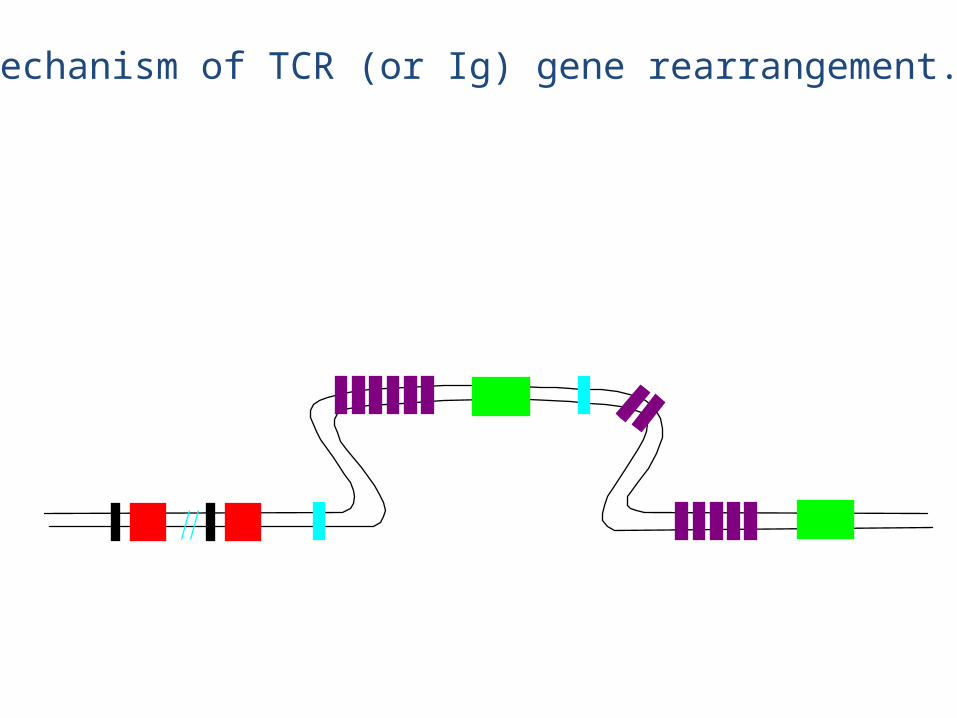
TCR genetics: Similar to Ig genetics
V regions (70-80) J regions (61) C region (1)
V regions (52) J1 (6)D1 (1) C1 (1) D2 (1) J2 (7) C2 (1)
(numbers of segments in book is off, just like for Ig genes)
chain
chain
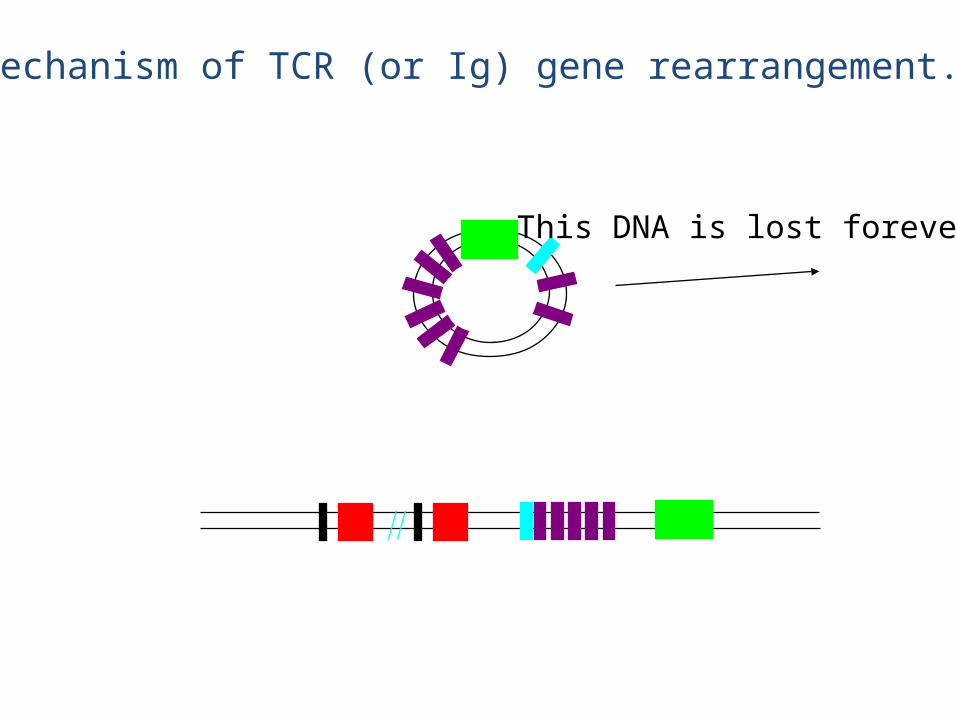
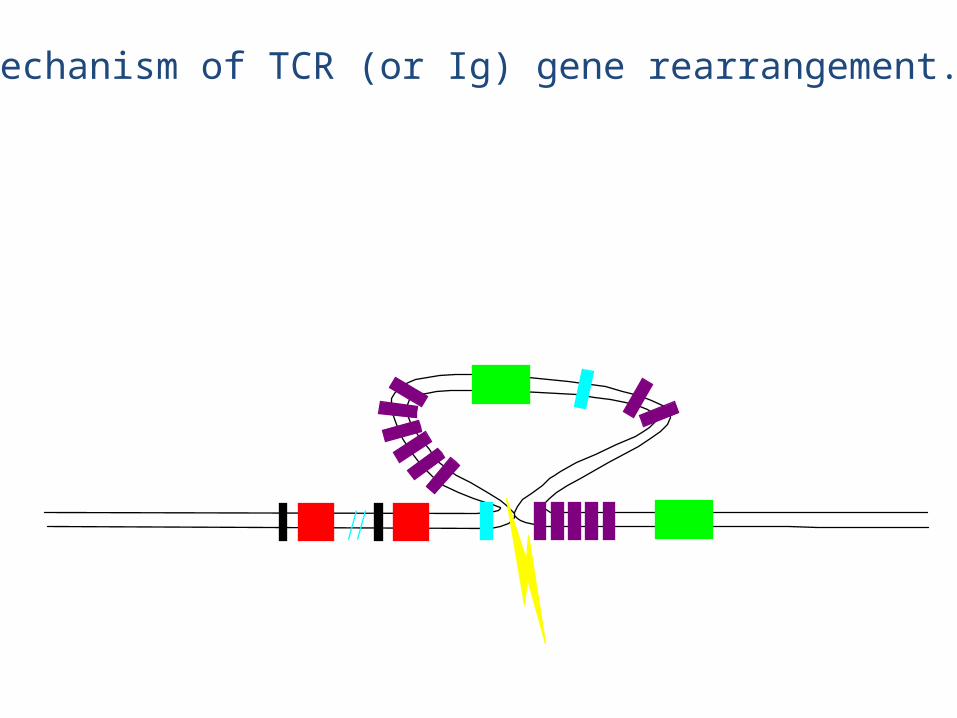
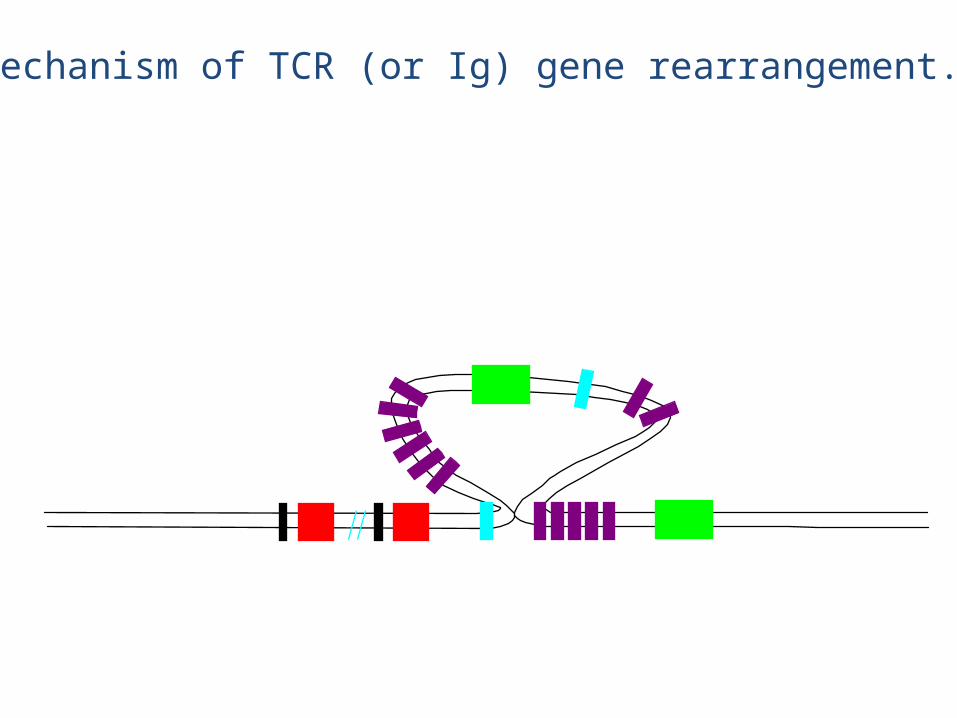
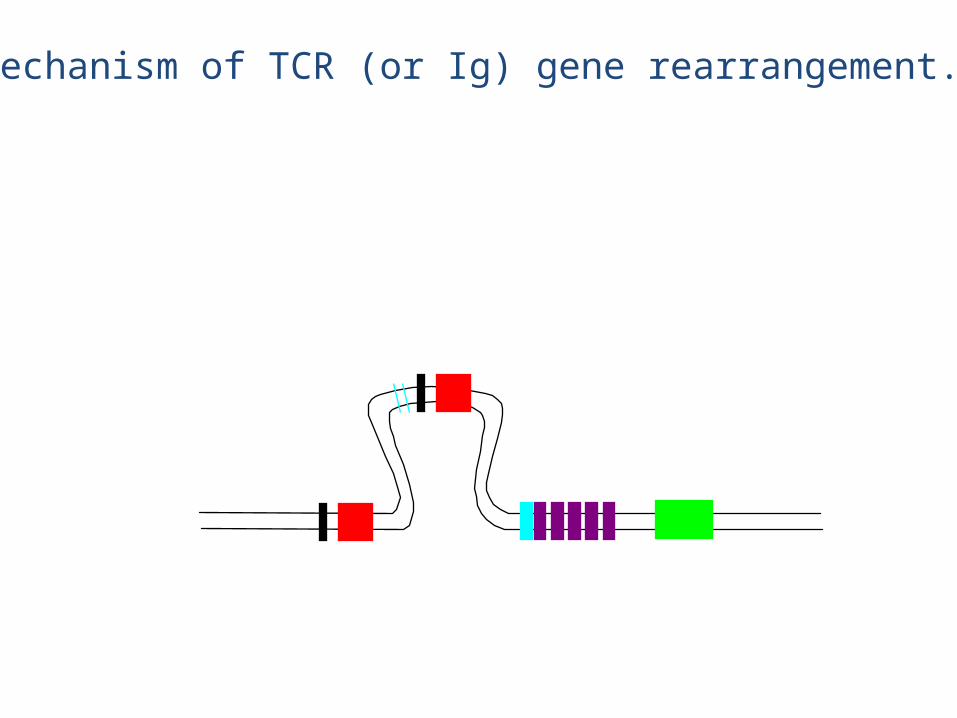
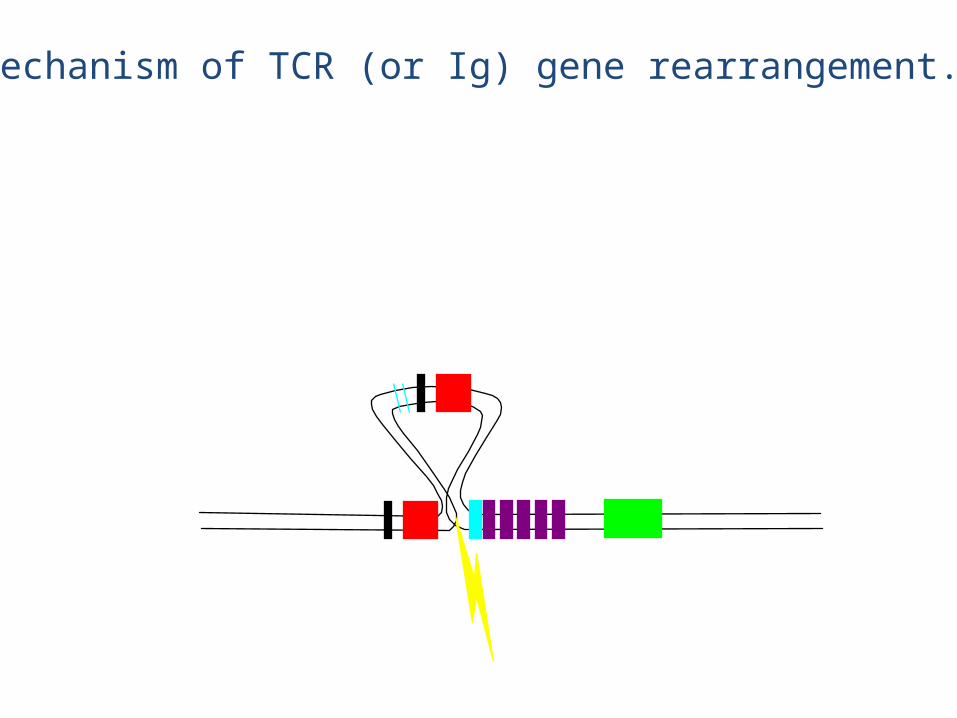
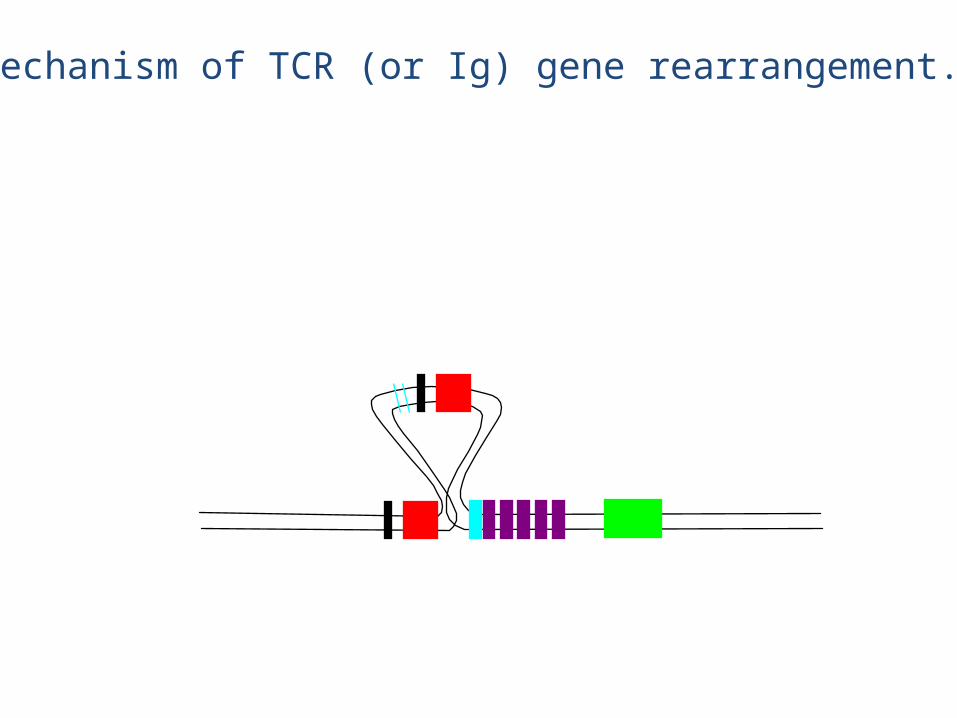
Mechanism of TCR (or Ig) gene rearrangement.
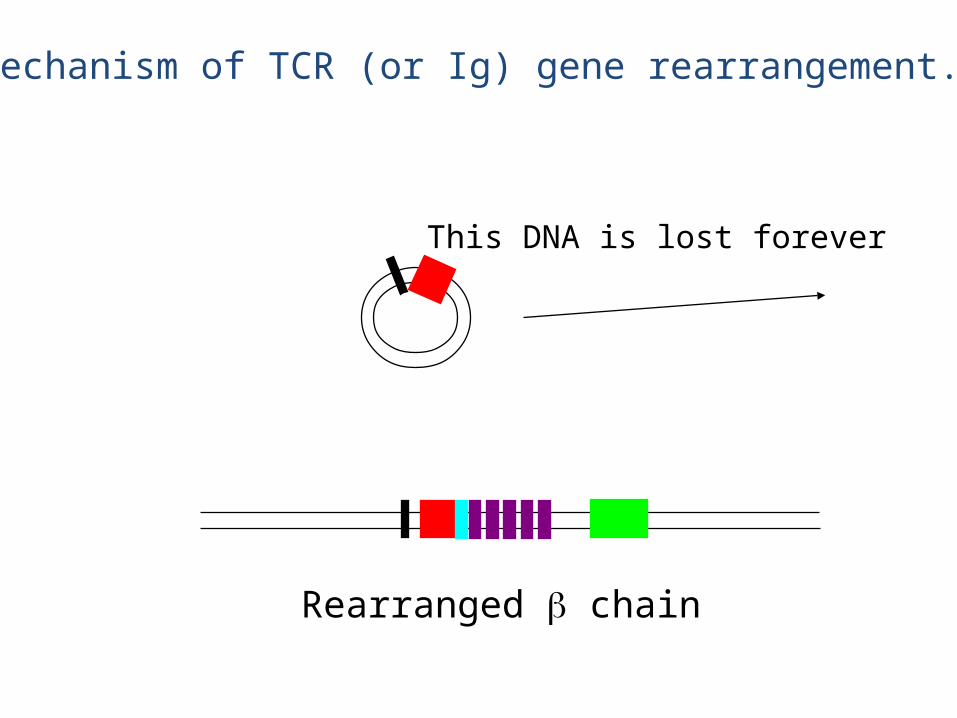
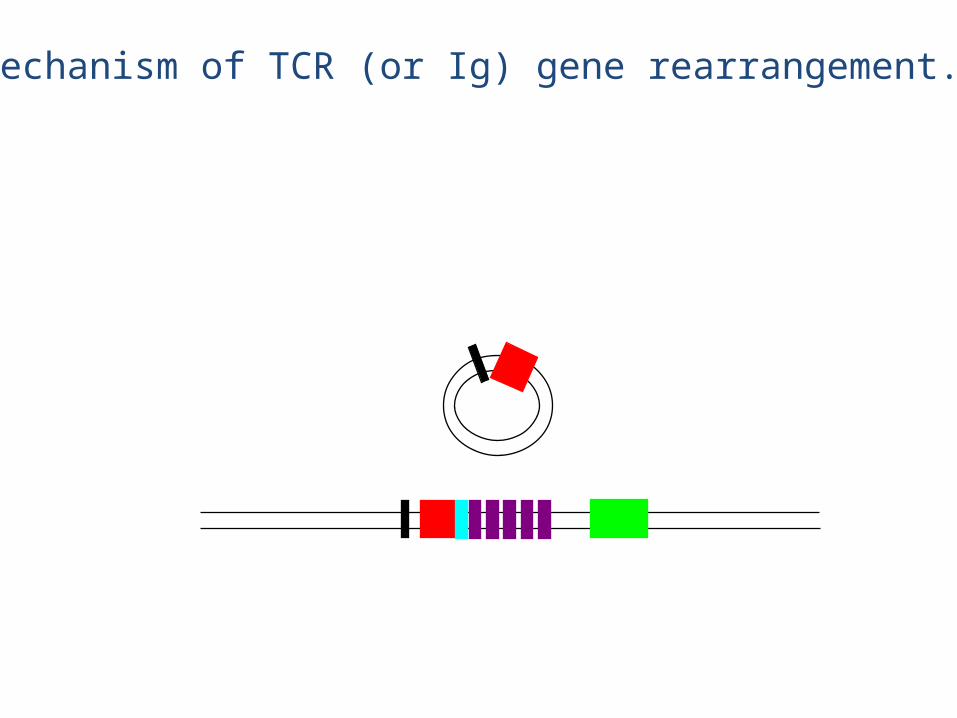
This DNA is lost forever
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
This DNA is lost forever
Rearranged chain
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
Mechanism of TCR (or Ig) gene rearrangement.
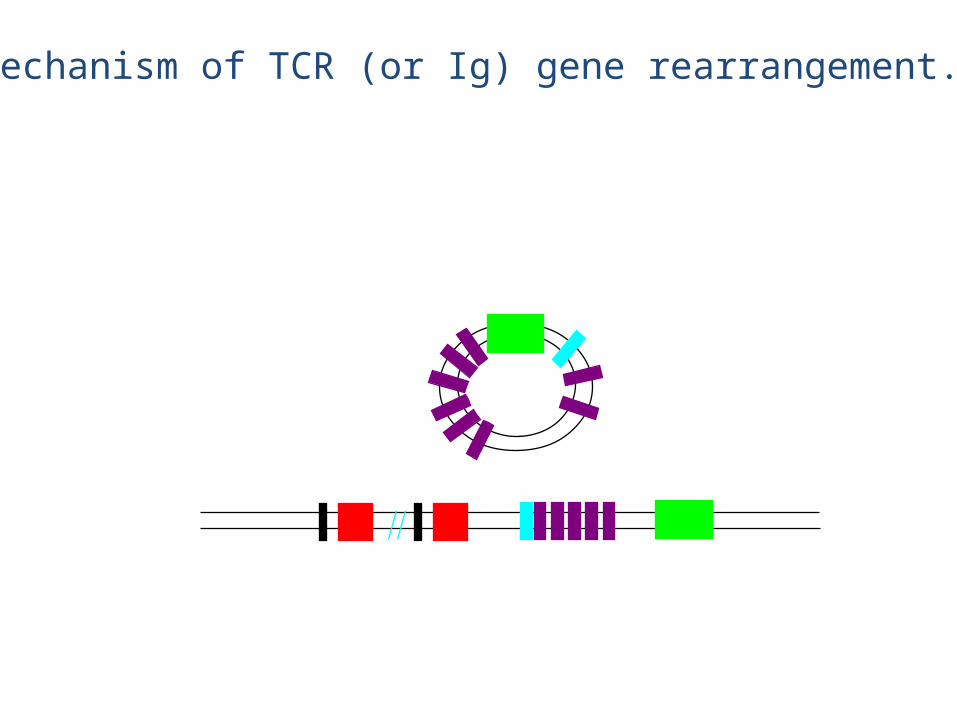
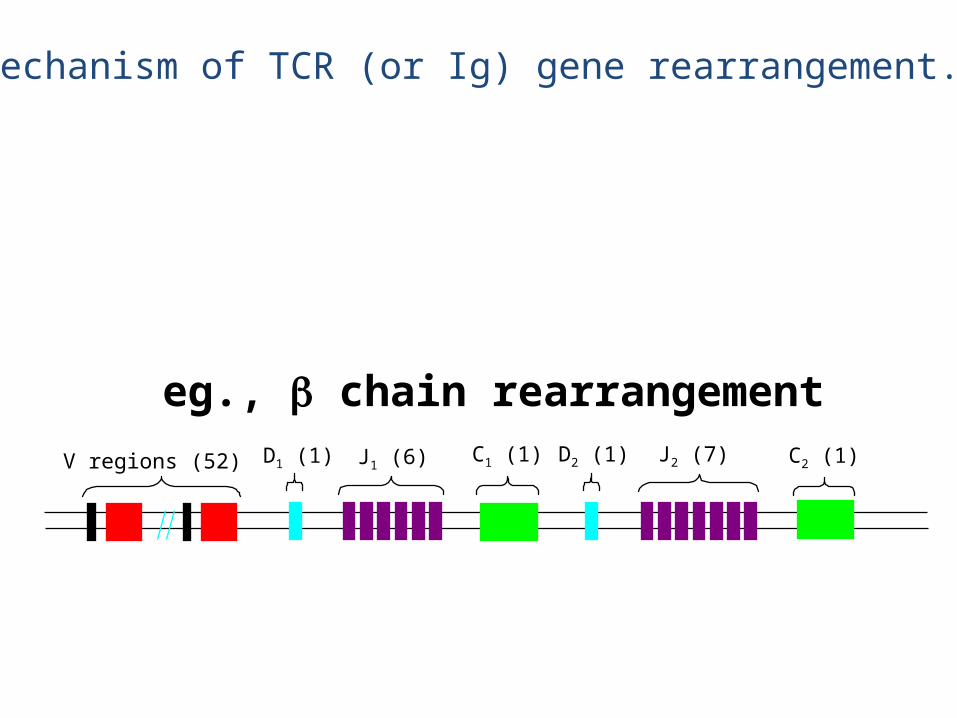
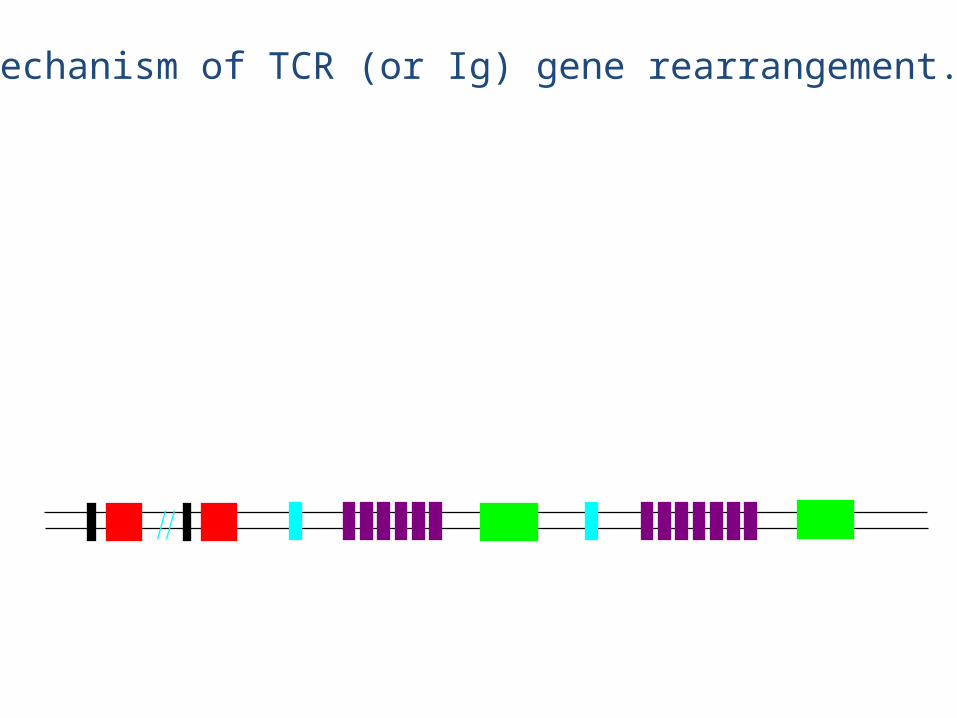
V regions (52) J1 (6)D1 (1) C1 (1) D2 (1) J2 (7) C2 (1)
eg., chain rearrangement
Mechanism of TCR (or Ig) gene rearrangement.

T cells develop in the thymusand undergo positive and negative selection
• Positive selection: T cells which can react to self MHC (major histocompatability complex) carrying peptides are allowed to live. Those that cannot undergo apoptosis (suicide).
• Negative selection: T cells that react strongly to self-antigens on MHC are eliminated.
• Only those T cells that can react to MHC, but do not bind strongly to self-antigens emerge as mature T cells from the thymus.
• Only about 2% of immature T cells make it through positive and negative selection.

–The T cell receptor (TCR): Structure and function–TCR expression•Genetic organization•Gene rearrangement

• The chain rearrangement occurs before chain rearrangement.
• If a functional chain is produced, the chain gene is rearranged.
• If a functional chain is not produced, the pre-T cell dies.• The mechanism of rearrangement is basically the same as for B
cells -- the same enzymes are even used.
• Note: chain rearrangement can occur several times, so once a functional chain is produced, a functional TCR will most likely be produced. (Both TCR chain rearrangements and Ig heavy and light chain rearrangements generally only happen once for each chromosome in each cell).
TCR gene rearrangement, continued