sebenta fisiologia (recovered 1)
DESCRIPTION
fisioTRANSCRIPT

Fisiologia

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
2 | P á g i n a
Módulo 1 - Princípios Gerais de Fisiologia
Homeostasia
“A constância do meio interno é a condição para a vida livre e independente... Todos os mecanismos vitais, independentemente da sua variedade, tem um único objectivo, que é o de preservar constantes as condições de vida no ambiente interno” - Claude Bernard (1813-1878) A palavra homeostasia é usada pelos fisiologistas para designar manutenção das condições constantes (ou estáticas) do meio interno. Em síntese, todos os órgãos e tecidos do corpo exercem funções que ajudam a manter essas condições constantes. Por exemplo, os pulmões fornecem oxigénio para o líquido extracelular para repor o que está sendo consumido pelas células; os rins mantêm constantes as concentrações iónicas e o sistema gastrointestinal fornece nutrientes.
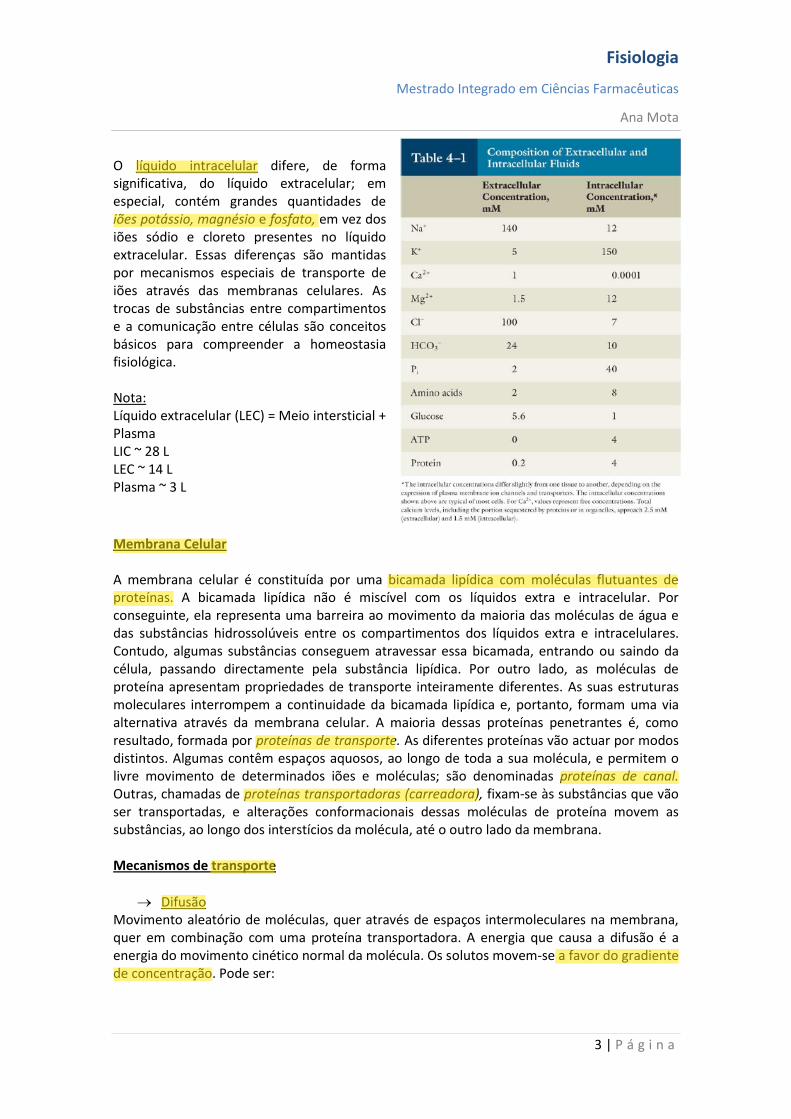
Líquido Extracelular (LEC) Cerca de 60% do corpo humano é composto por líquidos. Embora a maior parte desse líquido fique no interior das células — líquido intracelular —, cerca de um terço ocupa os espaços fora das células e é chamado de líquido extracelular (movimenta-se continuamente por todo o corpo). É transportado rapidamente no sangue circulante e, em seguida, misturado entre o sangue e os líquidos teciduais por difusão através das paredes capilares. No líquido extracelular, estão os iões e os nutrientes necessários às células, para manutenção da vida celular. Por conseguinte, todas as células partilham de um ambiente essencialmente constante, o líquido extracelular, razão por que esse é chamado de meio interno do corpo, ou milieu intérieur, expressão criada pelo fisiologista francês, Claude Bernard. As células são capazes de viver, crescer e desempenhar suas funções específicas enquanto estiverem disponíveis, nesse ambiente interno, as concentrações adequadas de oxigénio, glicose, diversos iões, aminoácidos e outros constituintes.
O líquido extracelular contém grandes quantidades de iões sódio, cloreto e bicarbonato, mais os nutrientes para as células, tais como oxigénio, glicose, ácidos gordos e aminoácidos. Também contém dióxido de carbono que está a ser transportado das células até os pulmões para serem excretados, além de outros produtos celulares que, igualmente, estão a ser transportados para o rim, onde vão ser excretados.
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
3 | P á g i n a
O líquido intracelular difere, de forma significativa, do líquido extracelular; em especial, contém grandes quantidades de iões potássio, magnésio e fosfato, em vez dos iões sódio e cloreto presentes no líquido extracelular. Essas diferenças são mantidas por mecanismos especiais de transporte de iões através das membranas celulares. As trocas de substâncias entre compartimentos e a comunicação entre células são conceitos básicos para compreender a homeostasia fisiológica. Nota: Líquido extracelular (LEC) = Meio intersticial + Plasma LIC ~ 28 L LEC ~ 14 L Plasma ~ 3 L
Membrana Celular A membrana celular é constituída por uma bicamada lipídica com moléculas flutuantes de proteínas. A bicamada lipídica não é miscível com os líquidos extra e intracelular. Por conseguinte, ela representa uma barreira ao movimento da maioria das moléculas de água e das substâncias hidrossolúveis entre os compartimentos dos líquidos extra e intracelulares. Contudo, algumas substâncias conseguem atravessar essa bicamada, entrando ou saindo da célula, passando directamente pela substância lipídica. Por outro lado, as moléculas de proteína apresentam propriedades de transporte inteiramente diferentes. As suas estruturas moleculares interrompem a continuidade da bicamada lipídica e, portanto, formam uma via alternativa através da membrana celular. A maioria dessas proteínas penetrantes é, como resultado, formada por proteínas de transporte. As diferentes proteínas vão actuar por modos distintos. Algumas contêm espaços aquosos, ao longo de toda a sua molécula, e permitem o livre movimento de determinados iões e moléculas; são denominadas proteínas de canal. Outras, chamadas de proteínas transportadoras (carreadora), fixam-se às substâncias que vão ser transportadas, e alterações conformacionais dessas moléculas de proteína movem as substâncias, ao longo dos interstícios da molécula, até o outro lado da membrana. Mecanismos de transporte
Difusão Movimento aleatório de moléculas, quer através de espaços intermoleculares na membrana, quer em combinação com uma proteína transportadora. A energia que causa a difusão é a energia do movimento cinético normal da molécula. Os solutos movem-se a favor do gradiente de concentração. Pode ser:

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
4 | P á g i n a
Difusão simples – as moléculas deslocam-se através da membrana sem se fixarem a proteínas transportadoras. Pode ocorrer através da membrana fosfolipídica ou através de canais iónicos.
Difusão facilitada – Requer uma proteína transportadora, a qual ajuda na passagem de moléculas através da membrana, provavelmente pela combinação química com elas e fazendo-as atravessar a membrana dessa forma.
A velocidade da difusão de uma substancia através da membrana celular é directamente proporcional à sua solubilidade lipídica. A água e outras moléculas insolúveis nos lípidos, difundem-se pelos canais proteicos (ou iónicos) da membrana celular. A maioria (mas não todos) dos canais proteicos é muito selectiva para o transporte de um ou mais iões ou moléculas. Isso resulta das características do próprio canal, tais como seu diâmetro, forma e natureza das cargas eléctricas nas superfícies internas.
Canais de sódio
Com diâmetro calculado de apenas 0,3 por 0,5 nm, tem, nas suas superfícies internas, cargas negativas. Postula-se que essas fortes cargas negativas atraiam os iões sódio, com mais intensidade do que outros iões fisiologicamente importantes, para o interior dos canais, devido ao menor diâmetro iónico do sódio não-hidratado. Uma vez no interior do canal, os iões sódio podem difundir-se em qualquer direcção, segundo as leis da difusão. Por conseguinte, o canal de sódio é especificamente selectivo para a passagem dos iões sódio.
Canais de potássio Por outro lado, outro grupo de canais proteicos é selectivo para o transporte de potássio. Esses canais, com diâmetros calculados menores que os dos canais de sódio, da ordem de 0,3 por 0,3 nm, não contêm cargas negativas. Como resultado, não existem forças atractivas fortes que puxem os iões para o interior dos canais e os iões não são retirados das moléculas de água que os hidratam. A forma hidratada do ião potássio é muito menor que a forma hidratada do ião sódio porque o ião sódio tem todo um conjunto orbital de electrões a menos que o ião potássio, o que permite ao ião sódio atrair número bem maior de moléculas de água do que o potássio. Por conseguinte, os iões hidratados de potássio (menores) podem passar facilmente por esse canal mais estreito, ao passo que os iões sódio são rejeitados, o que, de novo, causa permeabilidade selectiva para um tipo de ião.
Transporte activo
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
5 | P á g i n a
Movimento de substâncias através da membrana, em combinação com uma proteína transportadora, mas também contra um gradiente de energia (isto é, de um estado de baixa concentração para um estado de alta concentração). Requer uma fonte adicional de energia, para além da energia cinética. Os solutos movem-se contra o gradiente de concentração.
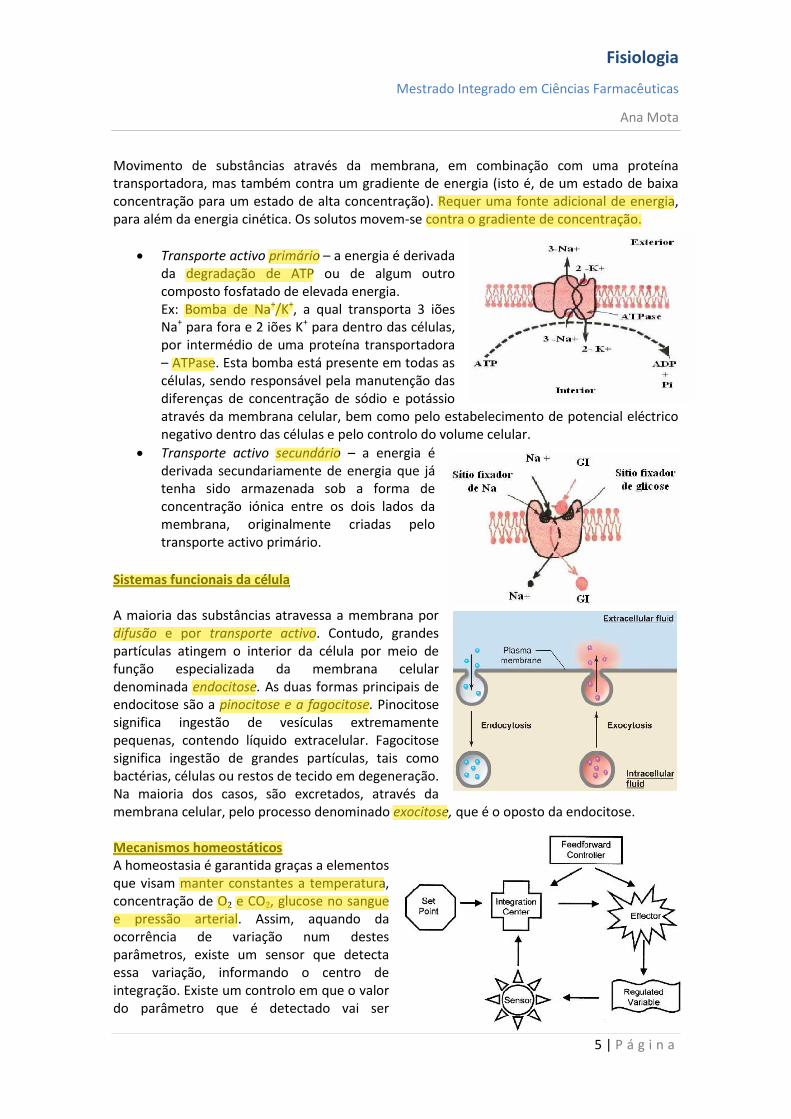
Transporte activo primário – a energia é derivada da degradação de ATP ou de algum outro composto fosfatado de elevada energia. Ex: Bomba de Na+/K+, a qual transporta 3 iões Na+ para fora e 2 iões K+ para dentro das células, por intermédio de uma proteína transportadora – ATPase. Esta bomba está presente em todas as células, sendo responsável pela manutenção das diferenças de concentração de sódio e potássio através da membrana celular, bem como pelo estabelecimento de potencial eléctrico negativo dentro das células e pelo controlo do volume celular.
Transporte activo secundário – a energia é derivada secundariamente de energia que já tenha sido armazenada sob a forma de concentração iónica entre os dois lados da membrana, originalmente criadas pelo transporte activo primário.
Sistemas funcionais da célula A maioria das substâncias atravessa a membrana por difusão e por transporte activo. Contudo, grandes partículas atingem o interior da célula por meio de função especializada da membrana celular denominada endocitose. As duas formas principais de endocitose são a pinocitose e a fagocitose. Pinocitose significa ingestão de vesículas extremamente pequenas, contendo líquido extracelular. Fagocitose significa ingestão de grandes partículas, tais como bactérias, células ou restos de tecido em degeneração. Na maioria dos casos, são excretados, através da membrana celular, pelo processo denominado exocitose, que é o oposto da endocitose. Mecanismos homeostáticos A homeostasia é garantida graças a elementos que visam manter constantes a temperatura, concentração de O2 e CO2, glucose no sangue e pressão arterial. Assim, aquando da ocorrência de variação num destes parâmetros, existe um sensor que detecta essa variação, informando o centro de integração. Existe um controlo em que o valor do parâmetro que é detectado vai ser

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
6 | P á g i n a
comparado com um valor padrão, considerando o normal, logo, se estes não coincidirem, o centro efector desencadeia uma resposta (SNC), estabilizando a variação.
Regulação da Pressão Arterial Vários sistemas distintos contribuem para a regulação da pressão arterial. Um desses, o sistema barorreceptor, é exemplo simples e excelente de um mecanismo de controlo. Nas paredes da bifurcação das artérias carótidas, no pescoço e, também, na crossa da aorta, no toráx, existem muitos receptores neuronais chamados barorreceptores, que são estimulados pela distensão (estiramento) da parede arterial. Quando a pressão arterial se eleva, os barorreceptores são estimulados de forma excessiva, são transmitidos impulsos para o bulbo cefálico. Aí, esses impulsos inibem o centro vasomotor, o que, por sua vez, reduz o número de impulsos transmitidos, pelo sistema nervoso simpático, para o coração e para os vasos sanguíneos. Essa diminuição dos impulsos provoca uma menor actividade de bombeamento pelo coração e maior facilidade para o fluxo de sangue pelos vasos periféricos. Ambos os efeitos provocam o abaixamento da pressão arterial até ao seu valor normal. De modo inverso, a queda da pressão arterial relaxa os receptores de estiramento, permitindo que o centro vasomotor fique mais activo que o usual, o que provoca a elevação da pressão arterial até ao seu valor normal.
Resposta em várias situações -Hipotensão postural -Hemorragia Apresentam oscilações - Pequenas - Grandes Atenuação das oscilações -Sensores diferenciais -Mecanismos de antecipação
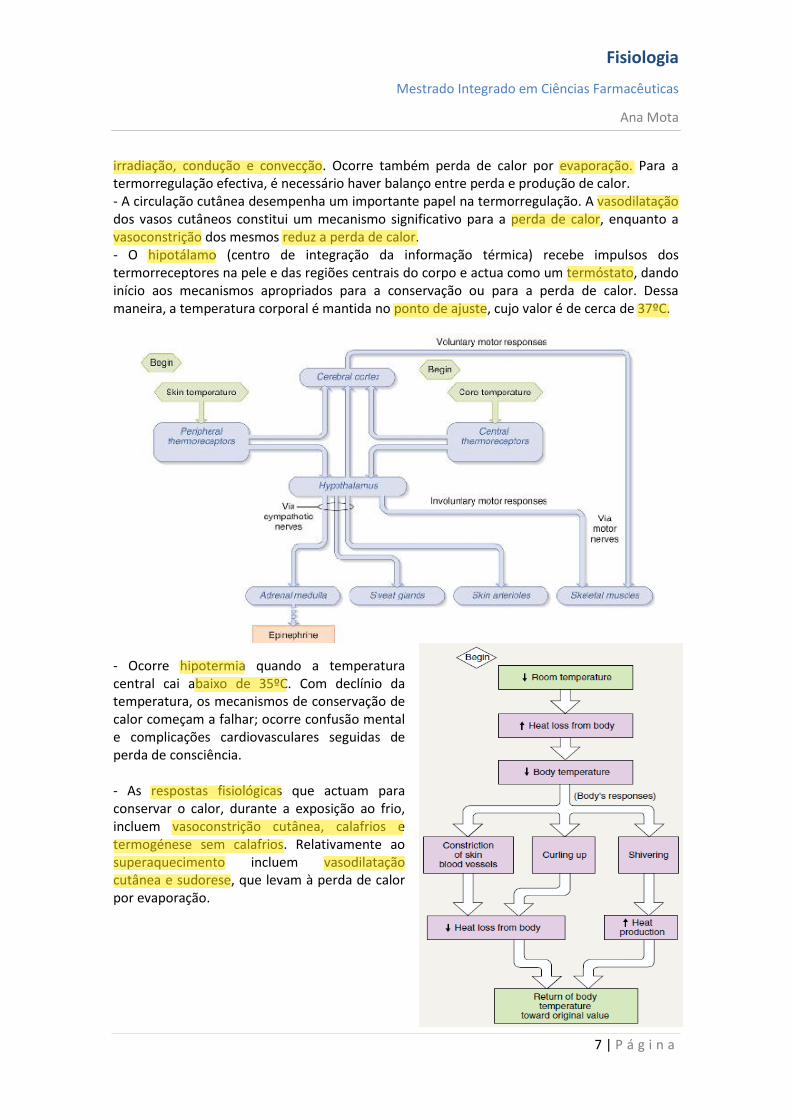
Regulação da temperatura corporal - Para assegurar condições ideais para a actividade das enzimas, os humanos mantêm a temperatura corporal central entre 36 e 38ºC. O calor é produzido no corpo por reacções metabólicas, enquanto é perdido pela superfície corporal para o meio ambiente por
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
7 | P á g i n a
irradiação, condução e convecção. Ocorre também perda de calor por evaporação. Para a termorregulação efectiva, é necessário haver balanço entre perda e produção de calor. - A circulação cutânea desempenha um importante papel na termorregulação. A vasodilatação dos vasos cutâneos constitui um mecanismo significativo para a perda de calor, enquanto a vasoconstrição dos mesmos reduz a perda de calor. - O hipotálamo (centro de integração da informação térmica) recebe impulsos dos termorreceptores na pele e das regiões centrais do corpo e actua como um termóstato, dando início aos mecanismos apropriados para a conservação ou para a perda de calor. Dessa maneira, a temperatura corporal é mantida no ponto de ajuste, cujo valor é de cerca de 37ºC. - Ocorre hipotermia quando a temperatura central cai abaixo de 35ºC. Com declínio da temperatura, os mecanismos de conservação de calor começam a falhar; ocorre confusão mental e complicações cardiovasculares seguidas de perda de consciência. - As respostas fisiológicas que actuam para conservar o calor, durante a exposição ao frio, incluem vasoconstrição cutânea, calafrios e termogénese sem calafrios. Relativamente ao superaquecimento incluem vasodilatação cutânea e sudorese, que levam à perda de calor por evaporação.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
8 | P á g i n a
Ritmos biológicos Servem para manter a constância do meio interno, funcionando sem detectores e regulados internamente – inconscientes. 1. Mecanismos de antecipação 2. Funcionam sem detectores 3. São regulados internamente por pacemakers hipotalâmicos 4. Influenciados por indicadores externos (Ex: luz)
Homeostasia: níveis de regulação 1. Identificar uma variável do meio interno 2. Estabelecer o valor de referência para essa variável 3. Identificar factores que afectam essa variável 4. Analisar a influência desses factores 5. Identificar a forma como o organismo monitoriza essa variável 6. Identificar os efectores que restabelecem o valor normal dessa variável
Comunicação celular
Linguagem química Utiliza substâncias químicas para comunicar. A 1ª célula sintetiza a substância que tem informação, a qual é expulsa para o meio extracelular, atingindo uma célula-alvo. É mais lenta que a linguagem eléctrica. São do tipo:
Endócrina O mensageiro tem capacidade de percorrer longas distâncias. Estes tomam o nome de hormonas, lançadas para a corrente sanguínea e transportadas pelo sistema circulatório.
Sinalização sináptica As células nervosas libertam substâncias químicas (neurotransmissores) nas suas terminações para influenciarem as células com as quais contactam.
Parácrina A substância química utilizada como mensageiro limita-se a comunicar a mensagem a células vizinhas, uma vez que existem mecanismos que destoem esta mensagem, não podendo esta comunicar a grandes distâncias.
Autócrina A substância química é sintetizada e lida na mesma célula, podendo também ser lida nas células vizinhas.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
9 | P á g i n a
Linguagem eléctrica Ocorre apenas quando as células estão muito próximas e existem pontos de ligação entre elas – gap junction. Utilizam iões para se comunicarem, os quais transportam uma mensagem (a descodificação desta mensagem é feita baseada na alteração das cargas da célula, sendo alterada a sua função). Ocorre em neurónios, astrócitos, músculo liso e cardíaco. Receptores celulares - Quase sempre proteínas; - Reconhecem a substância química que possui a mensagem; - Alvos de fármacos; - Permite prever quais as células que comunicam entre si; - Classificados de acordo com o tipo de mensagem que as activa e forma como traduzem a mensagem.
Intracelulares (mensageiros lipossolúveis) Os mensageiros hidrofóbicos necessitam de uma proteína que os transporte no plasma mas atravessam facilmente a membrana das células alvo ligando-se ao seus receptores que tem uma localização citoplasmática ou nuclear. Os mensageiros hidrofóbicos modificam a expressão de genes conduzindo a respostas lentas e sustentadas.
Receptores da membrana (especificidade dos receptores) Os receptores são específicos, no entanto, para cada mensageiro pode ter-se vários receptores. Se o nº de receptores aumenta, a célula fica mais susceptível a pequenas quantidades de mensageiros.
Características dos receptores
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
10 | P á g i n a
Tipos de receptores (Sistemas de Transdução de Sinal)
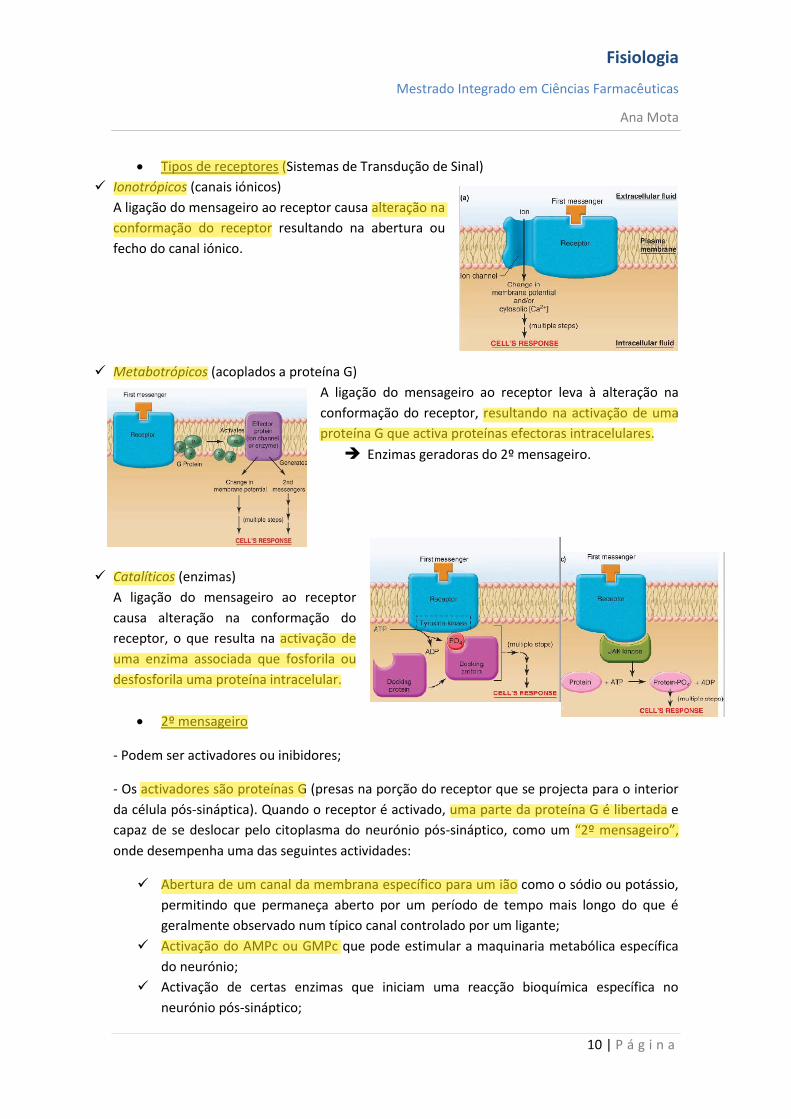
Ionotrópicos (canais iónicos)
A ligação do mensageiro ao receptor causa alteração na
conformação do receptor resultando na abertura ou
fecho do canal iónico.
Metabotrópicos (acoplados a proteína G)
A ligação do mensageiro ao receptor leva à alteração na
conformação do receptor, resultando na activação de uma
proteína G que activa proteínas efectoras intracelulares.
Enzimas geradoras do 2º mensageiro.
Catalíticos (enzimas)
A ligação do mensageiro ao receptor
causa alteração na conformação do
receptor, o que resulta na activação de
uma enzima associada que fosforila ou
desfosforila uma proteína intracelular.
2º mensageiro
- Podem ser activadores ou inibidores;
- Os activadores são proteínas G (presas na porção do receptor que se projecta para o interior
da célula pós-sináptica). Quando o receptor é activado, uma parte da proteína G é libertada e
capaz de se deslocar pelo citoplasma do neurónio pós-sináptico, como um “2º mensageiro”,
onde desempenha uma das seguintes actividades:
Abertura de um canal da membrana específico para um ião como o sódio ou potássio,
permitindo que permaneça aberto por um período de tempo mais longo do que é
geralmente observado num típico canal controlado por um ligante;
Activação do AMPc ou GMPc que pode estimular a maquinaria metabólica específica
do neurónio;
Activação de certas enzimas que iniciam uma reacção bioquímica específica no
neurónio pós-sináptico;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
11 | P á g i n a
Activação da transcrição dos genes, resultando na síntese de proteínas que podem
alterar o metabolismo ou morfologia celular.
- Enzimas que geram segundos mensageiros:
Adenilciclase: AMPc
Guanilciclase: GMc
Fosfolipase C: DAG, IP3, Ca2+
O IP3 promove a saída de cálcio (do RE) para o meio extracelular;
Os segundos mensageiros produzidos pela fosfolipase C são responsáveis pela resposta da
célula à mensagem importada;
Fosfolipase A2: ácido araquidónico, eicosanóides
A fosfolipase A2 transforma os fosfolípidos em ácido araquidónico, o qual será transformado
noutras substâncias, formando a família dos eicosanóides (medidores da inflamação);
A fosfodiasterase degrada o AMPc;
Regulação da Resposta Celular
Remoção do mensageiro extracelular
◦ Difusão para longe do receptor
◦ Captação para dentro da célula e degradação
◦ Degradação enzimática extracelular
Inactivação do 2º mensageiro
◦ Degradação enzimática intracelular
Regulação do número de receptores
Regulação da afinidade dos receptores
Nota: - Concentração do mensageiro e número e afinidade dos receptores -> factores que regulam a intensidade/duração da resposta celular. - O nº de receptores está relacionado com a
concentração do mensageiro no meio extracelular
– up and down regulation
Uma maior densidade de receptores irá
permitir uma resposta mais intensa
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
12 | P á g i n a
Resposta Celular 1. Potencial de Membrana 2. Metabolismo Celular 3. Síntese de Proteínas 4. Libertação de Mensageiros 5. Contracção Muscular 6. Transporte de Iões 7. Proliferação e Diferenciação 8. Transcrição de genes
Módulo 2 – Neurónios e Células Gliais
Neurónios
- Função:
Dendrites – receber informação
Corpo celular – processamento
Axónio – condução
Telodendrites - transmitir
O espaço entre a dendrite de um neurónio e o
axónio de outro denomina-se sinapse.
- Transporte Axonal:
- Classes funcionais de Neurónios:
Sensitivos ou Aferentes: conduzem informação da periferia para o SNC. São sensores
do sistema de homeostasia. No SNC estabelecem sinapse (interneurónios) com
axónios mais curtos.
Interneurónios: localizam-se integralmente no SNC. Conduzem os potenciais de acção
de um neurónio para outro.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
13 | P á g i n a
Motores ou Eferentes: conduzem informação do SNC para a periferia. Direccionam as
respostas de todo o corpo. Têm axónios compridos.
Canais iónicos
Selectivos: seleccionam os iões para passarem com base na carga, tamanho e quanta
água consegue atrair e manter;
Passivos ou activos;
Únicos funcionalmente;
Localizam-se em determinadas regiões.
- Canais iónicos neuronais
Passivos: estão permanentemente abertos e encontram-se distribuídos por toda a
membrana do neurónio.
Activos: abrem perante um estímulo adequado, sendo sensíveis a substâncias químicas
(abrindo quando uma substância química se liga a eles). Podem ser:
Activados por mensageiros:
Localizam-se nas dendrites e corpo celular. Neurotransmissores como a acetilcolina e o GABA
ligam-se aos canais activados por mensageiros provocando a sua abertura, o que permite que
os iões se movimentem ao longo da membrana.
Dependentes da voltagem:
Localizam-se no cone de implantação, ao longo de todo o axónio não mielinizado e nos
nódulos de Ranvier do axónio mielinizado. Durante o potencial de acção, a voltagem ao longo
da membrana varia provocando a abertura de canais dependentes do potencial. Quando o
neurónio está em repouso, estes canais estão fechados.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
14 | P á g i n a
Nota: A
permeabilidade da membrana aos iões depende do número e tipo de canais que estão abertos
em cada momento.
Apenas uma pequena separação de carga é necessária para o estabelecimento do potencial de membrana em repouso.
Potencial de Membrana Permeabilidade aos iões (K+ > Na+)
Gradiente de electroquímico dos iões Bomba Na+/ K+ (3-2)
Os neurónios são selectivamente permeáveis ao potássio, sódio e cloro. As células são
impermeáveis a proteínas carregadas negativamente e a outros grandes aniões. Estes são
demasiado grandes para passar através da membrana celular.
A permeabilidade da membrana aos iões é aumentada, aumentando o nº de canais
iónicos.
A permeabilidade da célula para um dado ião aumenta quando canais iónicos activos estão
abertos para esse ião. Este mecanismo é usado pelo SN para produzir rápidas mudanças na
permeabilidade da membrana.
Potencial de Repouso: A bomba Na+/K+ estabelece os gradientes de concentrações e gera uma pequena diferença de potencial negativa; este mecanismo é responsável por um gasto até 40% do ATP produzido pela célula.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
15 | P á g i n a
Embora existam diferentes concentrações iónicas nos líquidos
intra e extracelular, estes são quase electricamente neutros (têm
um nº praticamente igual de iões carregados positiva e
negativamente).
Existe uma distribuição de carga desigual entre a região
imediatamente adjacente ao interior e exterior da membrana celular. Esta diferença de cargas
eléctricas – diferença de potencial – pode ser medida entre o exterior e o exterior das células.
Sendo este gradiente eléctrico necessário para o transporte de proteínas do citoplasma. O
sistema nervoso faz uso dos potenciais de membrana – mecanismos de comunicação.
A -70 mV (células não estimuladas - potencial de repouso), K+ sai da célula e o Na+ entra na
célula Bomba de Na+/K+ (mantém o potencial de membrana) previne que deixe de haver
gradiente destes iões.
Neurónios em repouso são muito permeáveis ao K+
e ao Cl- e apenas um pouco permeáveis ao Na+, que
entra na célula devido ao gradiente electroquímico,
facto que torna o potencial da membrana
ligeiramente mais positivo.
O potencial de repouso é proporcional à tendência do K+ para se difundir para o
exterior da célula e não ao fluxo real destes.
O potencial de membrana é condicionado por:
Permeabilidade da membrana aos iões - mais permeáveis ao K+ porque o nº de canais
passivos é superior ao de Na+;
Iões deslocam-se por difusão – gradiente electroquímico de iões – Na+ tem tendência a
entrar e K+ a sair;
Saem 3 K+ e entram 2 Na+ - cargas não compensadas;
Dentro das células existem cargas negativas fixas que atraem e retêm cargas positivas
para o centro da célula, desfalcando a membrana.
Bomba Na+/K+
3 Na+ para fora;
2 K+ para dentro;
O nº de iões Cl- que entram e saem é sensivelmente o mesmo, pelo que não interagem
significativamente no potencial.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
16 | P á g i n a
Alterações no Potencial de Membrana Potenciais Gradativos
Despolarização: diminuição do
potencial de membrana
Hiperpolarização/pós-
hiperpolarização: aumento do
potencial de membrana
Potencial de Acção
Os sinais neurais são transmitidos por meio de potenciais de acção, que são variações muito rápidas do potencial de membrana. Cada potencial de acção começa por modificação abrupta do potencial de repouso negativo normal para um potencial positivo e, em seguida, termina com modificação quase tão rápida para o potencial negativo. Para conduzir um sinal neural, o potencial de acção desloca-se ao longo da fibra nervosa até atingir seu término - transferência de cargas positivas para o interior no início e regresso dessas cargas para o exterior no fim. As fases sucessivas do potencial de acção são as seguintes:
Fase de repouso: é o potencial de membrana em repouso, antes que comece o potencial de acção. Diz-se que a membrana está "polarizada" durante esta fase, devido ao elevado potencial de membrana presente.
Fase de despolarização: em determinado momento, a membrana fica, abruptamente, muito permeável aos iões sódio, o que permite a entrada de grande número de iões sódio para o interior do axónio. O estado "polarizado" normal de - 90 mV é perdido, com o potencial variando rapidamente na direcção da positividade. Isso é chamado de despolarização. Nas fibras nervosas de maior calibre, o potencial de membrana "ultrapassa" (overshoots) o valor zero, e fica positivo, mas, nas fibras mais finas e em muitos neurónios do SNC, o potencial apenas fica próximo do valor zero, e não o ultrapassa para ficar positivo.
Fase de repolarização: dentro de poucos décimos milésimos de segundo após a membrana ter ficado extremamente permeável aos iões sódio, os canais de sódio começam a fechar, enquanto os canais de potássio se abrem mais do que o normal. Então, a rápida difusão dos iões potássio para o exterior restaura o potencial de membrana negativo normal do repouso. Isso é chamado de repolarização da membrana.
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
17 | P á g i n a
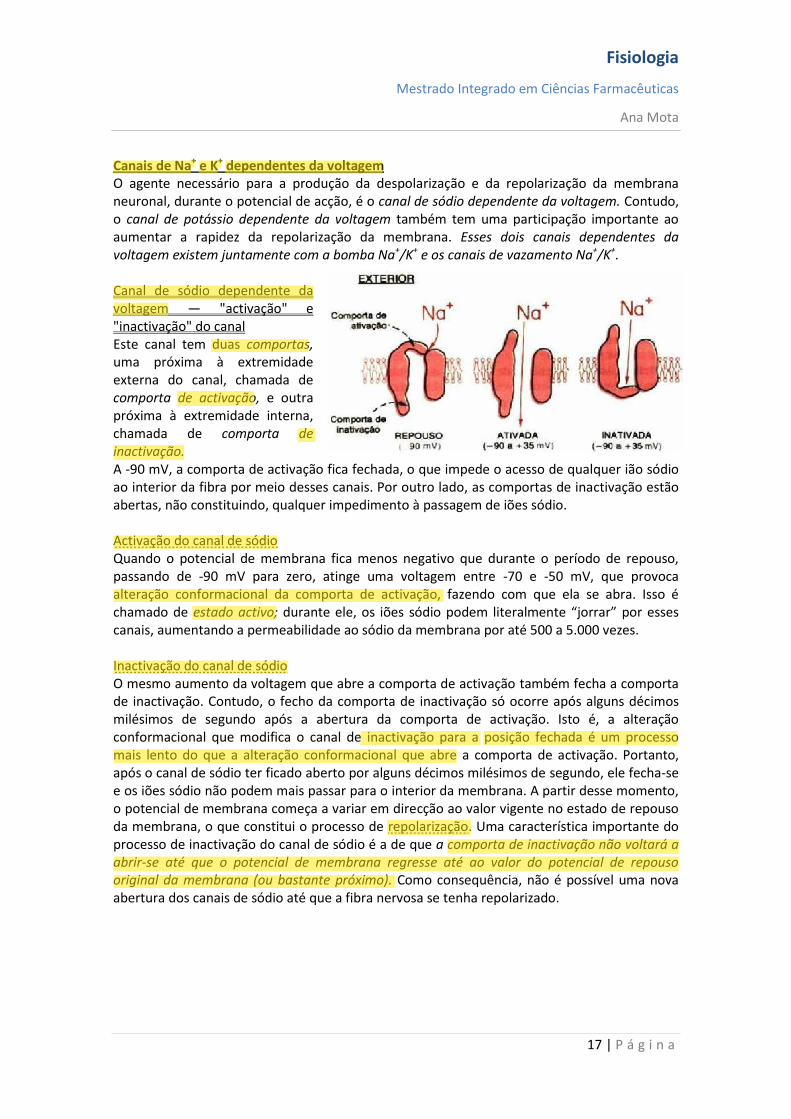
Canais de Na+ e K+ dependentes da voltagem O agente necessário para a produção da despolarização e da repolarização da membrana neuronal, durante o potencial de acção, é o canal de sódio dependente da voltagem. Contudo, o canal de potássio dependente da voltagem também tem uma participação importante ao aumentar a rapidez da repolarização da membrana. Esses dois canais dependentes da voltagem existem juntamente com a bomba Na+/K+ e os canais de vazamento Na+/K+. Canal de sódio dependente da voltagem — "activação" e "inactivação" do canal Este canal tem duas comportas, uma próxima à extremidade externa do canal, chamada de comporta de activação, e outra próxima à extremidade interna, chamada de comporta de inactivação. A -90 mV, a comporta de activação fica fechada, o que impede o acesso de qualquer ião sódio ao interior da fibra por meio desses canais. Por outro lado, as comportas de inactivação estão abertas, não constituindo, qualquer impedimento à passagem de iões sódio. Activação do canal de sódio Quando o potencial de membrana fica menos negativo que durante o período de repouso, passando de -90 mV para zero, atinge uma voltagem entre -70 e -50 mV, que provoca alteração conformacional da comporta de activação, fazendo com que ela se abra. Isso é chamado de estado activo; durante ele, os iões sódio podem literalmente “jorrar” por esses canais, aumentando a permeabilidade ao sódio da membrana por até 500 a 5.000 vezes. Inactivação do canal de sódio O mesmo aumento da voltagem que abre a comporta de activação também fecha a comporta de inactivação. Contudo, o fecho da comporta de inactivação só ocorre após alguns décimos milésimos de segundo após a abertura da comporta de activação. Isto é, a alteração conformacional que modifica o canal de inactivação para a posição fechada é um processo mais lento do que a alteração conformacional que abre a comporta de activação. Portanto, após o canal de sódio ter ficado aberto por alguns décimos milésimos de segundo, ele fecha-se e os iões sódio não podem mais passar para o interior da membrana. A partir desse momento, o potencial de membrana começa a variar em direcção ao valor vigente no estado de repouso da membrana, o que constitui o processo de repolarização. Uma característica importante do processo de inactivação do canal de sódio é a de que a comporta de inactivação não voltará a abrir-se até que o potencial de membrana regresse até ao valor do potencial de repouso original da membrana (ou bastante próximo). Como consequência, não é possível uma nova abertura dos canais de sódio até que a fibra nervosa se tenha repolarizado.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
18 | P á g i n a
Canais de potássio dependentes da voltagem - activação Durante o estado de repouso, o canal de potássio está fechado - os iões potássio são impedidos de passar por esse canal para o exterior. Quando o potencial de membrana começa a aumentar, a partir de -90 mV, em direcção ao zero, essa variação de voltagem provoca uma alteração conformacional, abrindo o canal e permitindo o aumento da difusão do potássio por este. Contudo, devido à lentidão com que estes canais se abrem, ficam abertos apenas a partir do momento em que os canais de sódio começam a ser inactivados (começam a fechar-se). Assim, a diminuição da entrada de sódio para a célula, com aumento simultâneo da saída de potássio, combinam-se para aumentar a velocidade do processo da repolarização, levando, dentro de poucos décimos milésimos de segundo, à recuperação completa do potencial de repouso da membrana. Em suma, durante a fase de repouso (antes do início do potencial de acção), a condutância do potássio é 50 a 100 vezes maior que a do sódio. Isso é causado pelo maior vazamento de iões potássio que de iões sódio pelos canais de vazamento. Todavia, com o início do potencial de acção, os canais de sódio ficam instantaneamente activados, permitindo aumento de 5000 vezes da condutância do sódio. Em seguida, o processo de inactivação fecha os canais de sódio em fracção de milissegundos. O início do potencial de acção também leva à activação, por voltagem, dos canais de potássio, fazendo-os abrir em fracção de milissegundos após a abertura dos canais de sódio. Ao término do potencial de acção, o retorno do potencial de membrana a seu estado negativo faz com que os canais de potássio se fechem, mas, novamente, só após retardo de um milissegundo ou mais. Durante a fase inicial do potencial de acção, um número muito maior de iões sódio flui para o interior da fibra que de iões potássio para o exterior. E isso que faz com que o potencial de acção se torne positivo. Em seguida, os canais de sódio começam a ficar inactivos, enquanto os canais de potássio se abrem, de modo que a proporção entre as condutâncias varia, passando a uma condutância muito maior do potássio que do sódio. Isso permite uma perda rápida de iões potássio para o exterior, enquanto, em essência, não há fluxo de iões sódio para o interior. Como consequência, o potencial de acção retorna rapidamente a sua linha de base.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
19 | P á g i n a
Pós-hiperpolarização O potencial de membrana fica ainda mais negativo que o potencial de membrana de repouso original, durante alguns milissegundos, após o término do potencial de acção. A causa deste pós-potencial é, principalmente, que muitos canais de potássio permanecem abertos durante vários milissegundos após o processo de repolarização da membrana ter-se completado. Isso permite que um excesso de iões potássio se difunda para fora da fibra nervosa, deixando, durante um milissegundo ou mais, um défice extra de iões positivos no interior, o que implica uma maior negatividade intracelular. Nota: O potencial de acção tem a mesma amplitude e duração ao longo do axónio; Consegue percorrer longas distâncias porque não “enfraquece” e é regenerado
continuamente à medida que se movimenta pelo axónio; Despolarização – aumento da permeabilidade da membrana aos iões sódio para
dentro da célula; Repolarização – diminuição da permeabilidade da membrana a iões sódio e aumento
da permeabilidade de iões potássio;
Período refractário absoluto – nenhum potencial de acção pode ser produzido por qualquer estímulo, seja qual for a intensidade;
Período refractário relativo – um estímulo mais forte que o limiar pode produzir um potencial de acção;
A intensidade do estímulo determina a frequência de potenciais de acção; (anexos 3 e 4)
Lei do tudo ou nada Uma vez tendo sido produzido um potencial de acção, em algum ponto da membrana de fibra normal, o processo de despolarização irá propagar-se, se as condições forem adequadas, por toda a membrana e, caso as condições não sejam adequadas, poderá não se propagar. Isso é chamado de “lei do tudo ou nada” e é aplicável a todos os tecidos excitáveis normais. Todavia, ocasionalmente, o potencial de acção poderá atingir uma região da membrana onde não será capaz de gerar voltagem suficiente para estimular a área seguinte da membrana. Quando isso ocorre, a propagação da despolarização cessa. Por conseguinte, para que ocorra a propagação continuada de um impulso, a proporção entre o potencial de acção e o limiar deve ser sempre maior que 1. Isso é chamado de factor de segurança para a propagação. A partir do limiar de estimulação, não
importa a intensidade do estímulo pois o potencial de acção é sempre o mesmo.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
20 | P á g i n a
Condução dos potenciais de acção
Neurónios sem mielina - a condução é feita ponto por ponto, por isso é mais lenta.
Neurónios com mielina – Condução saltatória
- Circundando o axónio existe a bainha de mielina que, muitas vezes, é bem maior que o próprio axónio e que, a intervalos de cerca de 1 a 3 mm, ao longo de toda a extensão do axónio, é interrompida pelos nódulos de Ranvier. A bainha de mielina é formada, em torno do axónio, pelas células de Schwann do seguinte modo: a membrana de uma célula de Schwann, inicialmente, circunda o axónio. Em seguida, essa célula gira em torno do axónio por muitas voltas, depositando múltiplas camadas de sua membrana celular, que contém a substância lipídica esfingomielina. Essa substância é um excelente isolante, capaz de diminuir o fluxo iónico através da membrana por cerca de 5000 vezes, ao mesmo tempo que reduz a capacitância da membrana por 50 vezes. Contudo, no ponto de junção entre duas células de Schwann sucessivas, ao longo do axónio, persiste pequena região não-isolada, com apenas cerca de 2 a 3 μm de extensão, por onde os iões podem fluir, com facilidade, do líquido extracelular para o interior do axónio. Essa região é o nódulo de Ranvier. - Muito embora os iões não possam fluir com intensidade significativa através das espessas bainhas de mielina dos nervos mielinizados, podem fluir com grande facilidade pelos nódulos de Ranvier. Por conseguinte, os potenciais de acção só podem ocorrer nos nódulos. Assim, os potenciais de acção são conduzidos de nódulo para nódulo; esse processo é chamado de condução saltatória. Isto é, a corrente elétrica flui pelos líquidos extracelulares que circundam a fibra, mas também pelo axoplasma, de nódulo a nódulo, excitando sequencialmente os sucessivos nódulos. Assim, o impulso nervoso salta ao longo da fibra, o que deu origem à designação de "saltatória". - A condução saltatória é importante por duas razões: Este mecanismo aumenta de muito a velocidade da transmissão neural nas fibras
mielinizadas por até 5 a 50 vezes. A condução saltatória conserva energia para o axónio, pois apenas os nódulos
despolarizam, permitindo perda de iões cerca de 100 vezes menor do que a que seria

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
21 | P á g i n a
necessária, caso não ocorresse condução saltatória e, como resultado, exigindo pouca actividade metabólica para o restabelecimento das diferenças de concentração de sódio e potássio, através da membrana celular, após uma série de impulsos nervosos.
- O excelente isolamento criado pela membrana de mielina e a redução de 50 vezes da capacitância da membrana permitem que o processo de repolarização ocorra com transferência muito reduzida de iões. Assim, ao término do potencial de acção, quando os canais de sódio começam a fechar, a repolarização ocorre de modo tão rápido que, em geral, os canais de potássio ainda não estão abertos em número significativo. Como resultado, a condução do impulso nervoso por fibra nervosa mielínica é efectuada, quase que inteiramente, pelas variações sequenciais dos canais de sódio dependentes da voltagem, com contribuição muito pequena dos canais de potássio.
A velocidade de condução nas fibras nervosas varia desde o mínimo de 0,5 m/s, nas fibras amielínicas mais delgadas, até cerca de 100 m/s (o comprimento de um campo de futebol em um segundo), nas fibras mielínicas de maior calibre. Em termos aproximados, essa velocidade de condução aumenta em proporção directa com o diâmetro nas fibras mielínicas e com a raiz quadrada do diâmetro da fibra nas amielínicas.
Condução fisiológica dos PAs
Um novo potencial de acção não pode ser produzido enquanto a membrana estiver despolarizada pelo potencial de acção precedente. A razão disso é que, logo depois que se inicia um potencial de acção, os canais de sódio (ou de cálcio, ou os dois) ficam inactivos e qualquer quantidade de sinal excitatório que seja aplicada a esses canais nesta fase não irá abrir as comportas de inactivação. A única condição que as reabrirá é o retorno do potencial de acção ao valor (ou quase) do potencial de membrana em repouso. Então, dentro de uma pequena fracção de segundo, as comportas de inactivação dos canais abrem-se e um novo potencial de acção poderá ser produzido. O intervalo de tempo durante o qual não pode ser produzido outro potencial de acção, mesmo com um estímulo forte, é chamado de período refractário absoluto. Esse período, para as grandes fibras mielínicas, é da ordem de 1/2500 s. Após o período refractário absoluto, existe um período refractário relativo, com duração entre um quarto e um meio da do período

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
22 | P á g i n a
absoluto. Durante este, estímulos mais fortes que os normais são capazes de excitar a fibra. Essa refractariedade relativa tem duas causas:
Alguns canais de sódio ainda não retornaram de seu estado de inactivação; Os canais de potássio ainda estão, em geral, inteiramente abertos, produzindo estado
de hiperpolarização, que dificulta a estimulação da fibra.
PAs conduzidos unidireccionalmente – estado de hiperpolarização da membrana
(aplicar um maior estímulo para que se abram os canais de sódio)
Estado conformacional dos canais – responsável pelo potencial da membrana
Classificação das fibras nervosas
Tipo A: SNC
Tipo B e C: SNA

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
23 | P á g i n a
Transmissão Sináptica
Sinapses são:
Junções especializadas;
Pontos onde as extremidades de neurónios vizinhos se encontram e o estímulo passa
de um neurónio para o seguinte por meio de mediadores químicos, os
neurotransmissores.
Estas ocorrem por contacto das terminações
nervosas (axónios) com as dendrites. Não
existe contacto físico – fenda sináptica.
São libertados dos axónios
neurotransmissores, que atravessam a fenda
e estimulam receptores nas dendrites e
assim transmitem o impulso nervoso de um
neurónio para outro.
Podem ser classificadas como:
Axodendríticas (axónio – dendrite) Axosomátivas (axónio – corpo celular) Axoaxónicas (axónio – axónio) Dendrodendríticas (dendrite – dendrite)

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
24 | P á g i n a
As sinapses formam circuitos entre os neurónios que podem ser:
Lineares-unidireccionais (vias sensitiva e motora)
Divergentes, convergentes, circulares e reverbativas (SNC)
Podem ser:
Sinapse eléctrica
Junções comunicantes contendo pontos de comunicação gap junction que se estendem do citoplasma do neurónio pré-sináptico até ao citoplasma do neurónio pós-sináptico;
A informação é conduzida sob a forma de corrente eléctrica, a qual flui com retardamento mínimo graças à presença dos pontos de comunicação gap junction;
Não existem transmissores químicos – processo bidireccional;
Rápida propagação da actividade de um neurónio para o seguinte assegura que um grupo de neurónios, desempenhando funções idênticas, actue em conjunto – acção sincronizada.
Alguns neurónios, células gliais e musculares podem estar unidos por junções permeáveis; a mesma estrutura pode funcionar como excitatória ou inibitória, funcionando em ambas as direcções.
Sinapse química
Ocorre conversão de energia eléctrica (impulso nervoso ou PA) em energia química (neurotransmissor);
As estruturas que fazem as comunicações estão fisicamente separadas;
Unidireccional;
O sinal que passa de um lado para o outro é excitatório ou inibitório;
Ao chegar à célula pré-sináptica, o impulso provoca a libertação de um neurotransmissor, mediador químico, que se liga a receptores da superfície da célula pós-sináptica
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
25 | P á g i n a
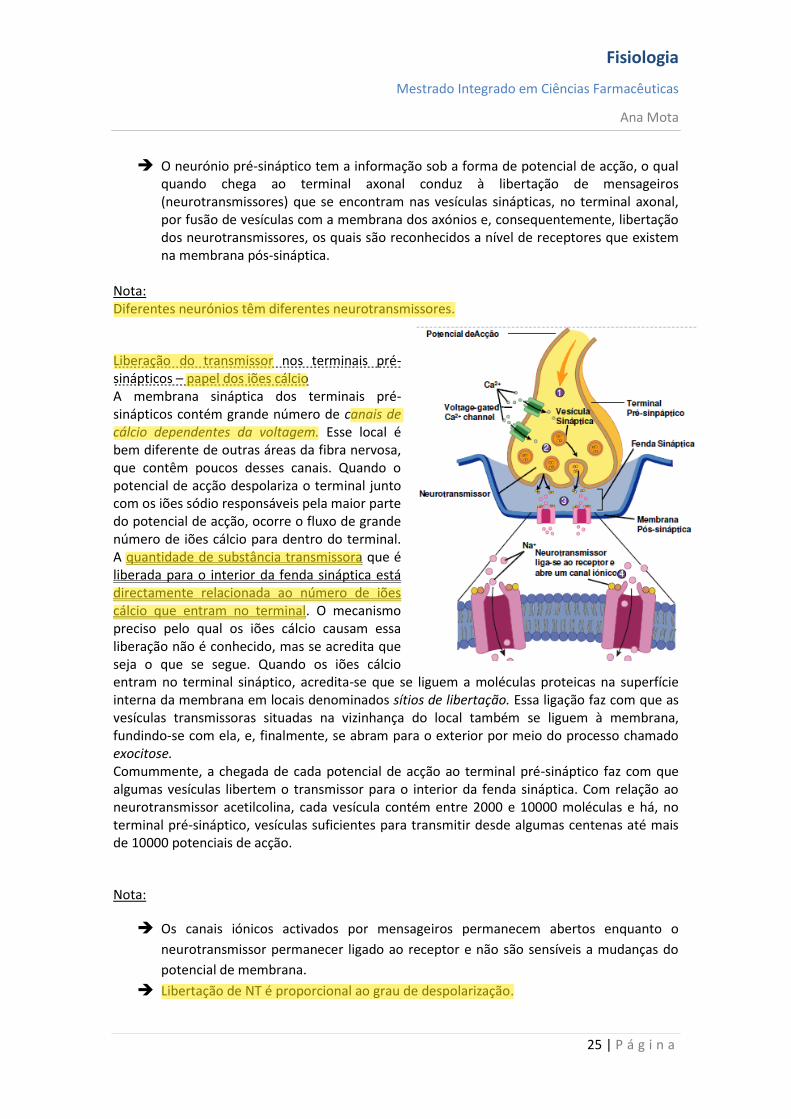
O neurónio pré-sináptico tem a informação sob a forma de potencial de acção, o qual quando chega ao terminal axonal conduz à libertação de mensageiros (neurotransmissores) que se encontram nas vesículas sinápticas, no terminal axonal, por fusão de vesículas com a membrana dos axónios e, consequentemente, libertação dos neurotransmissores, os quais são reconhecidos a nível de receptores que existem na membrana pós-sináptica.
Nota: Diferentes neurónios têm diferentes neurotransmissores.
Liberação do transmissor nos terminais pré-sinápticos – papel dos iões cálcio A membrana sináptica dos terminais pré-sinápticos contém grande número de canais de cálcio dependentes da voltagem. Esse local é bem diferente de outras áreas da fibra nervosa, que contêm poucos desses canais. Quando o potencial de acção despolariza o terminal junto com os iões sódio responsáveis pela maior parte do potencial de acção, ocorre o fluxo de grande número de iões cálcio para dentro do terminal. A quantidade de substância transmissora que é liberada para o interior da fenda sináptica está directamente relacionada ao número de iões cálcio que entram no terminal. O mecanismo preciso pelo qual os iões cálcio causam essa liberação não é conhecido, mas se acredita que seja o que se segue. Quando os iões cálcio entram no terminal sináptico, acredita-se que se liguem a moléculas proteicas na superfície interna da membrana em locais denominados sítios de libertação. Essa ligação faz com que as vesículas transmissoras situadas na vizinhança do local também se liguem à membrana, fundindo-se com ela, e, finalmente, se abram para o exterior por meio do processo chamado exocitose. Comummente, a chegada de cada potencial de acção ao terminal pré-sináptico faz com que algumas vesículas libertem o transmissor para o interior da fenda sináptica. Com relação ao neurotransmissor acetilcolina, cada vesícula contém entre 2000 e 10000 moléculas e há, no terminal pré-sináptico, vesículas suficientes para transmitir desde algumas centenas até mais de 10000 potenciais de acção.
Nota:
Os canais iónicos activados por mensageiros permanecem abertos enquanto o
neurotransmissor permanecer ligado ao receptor e não são sensíveis a mudanças do
potencial de membrana.
Libertação de NT é proporcional ao grau de despolarização.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
26 | P á g i n a
Acção da substância transmissora sobre o neurónio pós-sináptico — função dos receptores Nas sinapses, a membrana do neurónio pós-sináptico contém grande número de proteínas receptoras. Esses receptores têm dois componentes importantes:
Componente de ligação, que se projecta para fora da membrana, no interior da fenda sináptica, onde se fixa o neurotransmissor proveniente do terminal pré-sináptico;
Componente ionóforo, que atravessa toda a membrana para o interior do neurónio pós-sináptico. Por seu lado, o ionóforo pode ser de um dos dois tipos:
Canal iónico – (transmissão rápida) permite a passagem de tipos específicos de iões através da membrana → Receptores Ionotrópicos;
Activador de “2º mensageiro” – (transmissão lenta) não é um canal iónico, mas, em vez disso, é uma molécula que protrui para dentro do citoplasma da célula e activa uma ou mais substâncias dentro do neurónio pós-sináptico. Essas substâncias servem como “2º mensageiro”, para alterar funções internas específicas → Receptores metabotrópicos.
Canais iónicos Os canais iónicos, na membrana neuronal pós-sináptica, são, usualmente, de dois tipos:
Canais catiónicos – permitem que os iões sódio passem, quando estão abertos, mas, às vezes, permitem, também, a passagem de iões potássio e/ou cálcio;
Canais aniónicos – permitem a passagem de iões cloreto, mas, também, de quantidade diminutas de outros iões.
A abertura dos canais de sódio excita o neurónio pós-sináptico. Por isso, a substância transmissora que abre os canais catiónicos é chamada de transmissor excitatório. Por outro lado, a abertura dos canais aniónicos inibe o neurónio (entrada de cargas negativas), e os transmissores que os abrem são denominados transmissores inibitórios. O neurotransmissor libertado liga-se aos receptores da membrana, modificando a
membrana pós-sináptica.
A abertura e fecho dos canais iónicos fornecem um meio para o controlo rápido dos
neurónios pós-sinápticos.
Os receptores pós-sinápticos são proteínas transmembranares que sofrem alterações
configuracionais estereoquímicas quando se unem a um NT.
Dependendo do tipo de canal aberto vai haver alteração do potencial de membrana:
Despolarização: canais permeáveis a cargas positivas;
Hiperpolarização: canais permeáveis a Cl-.
Existem 2 tipos de sinapses químicas:
Excitatórias

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
27 | P á g i n a
Causam uma mudança eléctrica excitatória no potencial pós-sináptico (EPSP), que vai conduzir
a despolarização da membrana, logo dispara um potencial de acção; as sinapses excitatórias
são desencadeadas por NT como o glutamato e o nAChR que tornam a membrana permeável a
catiões (Na+, K+ e Ca2+), promovendo a despolarização da célula pós-sináptica, o que aumenta a
excitabilidade.
Inibitórias
Causam um potencial pós-sináptico inibitório (IPSP), que vai conduzir à hiperpolarização da
membrana e como esta reprime a excitabilidade, torna assim mais difícil alcançar o potencial
de limiar eléctrico; as sinapses inibitórias são desencadeadas por NT como o GABA e a glicina
que tornam a membrana permeável a aniões (Cl-) promovendo a hiperpolarização da célula
pós-sináptica, o que inibe a excitabilidade localizam-se maioritariamente no corpo celular
onde produzem um efeito mais poderoso.
A amplitude de um potencial sináptico varia e está relacionado com o nº de vesículas
sinápticas que libertam o NT relacionado com a quantidade de Ca2+ que entra no
terminal pré-sináptico – potenciais graduais.
Os potenciais sinápticos diminuem à medida que viajam pela sinapse.
Adição de EPSPs
Permite a entrada de mais Ca2+ no terminal;
Aumento do potencial pós-sináptico – potenciação – por um período de tempo.
A adição de vários EPSPs é necessária para despolarizar a célula a um estímulo limiar.
Adição espacial – dois ou mais terminais pré-sinápticos estimulam simultaneamente
um neurónio pós-sináptico – ocorrem simultaneamente numerosas EPSPs em vários
pontos das dendrites. Portanto, o potencial pós-sináptico combinado será maior do
que qualquer um dos potenciais individuais. A adição espacial permite que o EPSPs
tenha magnitude suficiente para exceder o limiar.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
28 | P á g i n a
Adição temporal – vários potenciais de acção chegam em sucessão a um único
terminal pré-sináptico – um segundo potencial pós-sináptico chega antes de a
membrana ter retornado ao nível de repouso. Há tempo suficiente para ocorrerem
várias aberturas de canais ao longo da duração de um só potencial pós-sináptico, não
havendo um período refractário que limite a frequência da produção de EPSPs. Os
efeitos desses 2 potenciais são aditivos (somados no tempo) e constituem um
potencial de despolarização de maior magnitude do que cada um separadamente.
Adição de IPSPs
Manter o potencial da membrana abaixo do estímulo limiar;
Impedir que a célula pós-sináptica gere um potencial de acção;
Também exibem adições temporais e espaciais.
↓
Os IPSPs e os EPSPs também sofrem adição entre si. Quando potenciais inibitórios são
adicionados a potenciais excitatórios, diminuem a magnitude dos EPSPs ou mesmo cancelam o
seu efeito impedindo a formação de um potencial de acção na célula pós-sináptica.
Inibição pré-sináptica
(sinapse axoaxónica)
Diminui a entrada de cálcio no terminal;
Diminui o potencial sináptico subsequente.
Sinapses entre terminação nervosa e corpo celular, uma dendrite ou terminal de outro axónio.
Redução da libertação de NT;
GABA: mediador inibitório do cérebro;
Nas junções sinápticas – não se verificam IPSPs;
GABA aumenta a permeabilidade da membrana a Cl- - abertura dos canais cloro
neutraliza eficazmente as correntes do potencial de acção, abrindo-se um menor nº de
canais Ca2+ dependentes da voltagem.
Em suma, permite o bloqueio selectivo de conexão sináptica específica, sem alterar a
excitabilidade do neurónio pós-sináptico.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
29 | P á g i n a
Inibição pós-sináptica
Actividade neuronal intencional - necessário que conexões sejam interrompidas em
certas circunstâncias, enquanto outras permanecem operacionais;
As correntes hiperpolarizantes geradas nas sinapses inibitórias contrariam a
ocorrência de quaisquer correntes despolarizantes, inibindo a somação destas;
As sinapses inibitórias são estrategicamente colocadas (ponto de vista espacial) de
modo a exercerem um poder bloqueador máximo.
Em suma, modifica a excitabilidade da célula pós-sináptica.
Nota:
Transmissão sináptica rápida: acção da acetilcolina sobre a junção neuromuscular.
Transmissão sináptica lenta: acção excitatória da norepinefrina sobre os receptores α1-
adrenérgicos, nos vasos periféricos.
Neurotransmissores (NT)
Biomoléculas;
Localizados no terminal pré-
sináptico;
Libertados por estimulação celular;
Atingem o terminal pós-sináptico
através da fenda sináptica;
Estimulação da célula pós-
sináptica;
Sintetizados por enzimas da célula
pré-sináptica;
Actividade atenuada por drogas
que inibem o seu metabolismo;
Vários receptores para cada NT.
A acção da célula pós-sináptica depende:
NT envolvido;
Receptor específico envolvido.
Existem 2 grupos de receptores:
Colinérgicos: receptores da acetilcolina;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
30 | P á g i n a
Adrenérgicos: receptores da norepinefrina.
Nota:
Transmissores de baixo peso molecular – acção rápida Transmissores de acção lenta — neuropéptidos
Os transmissores de baixo peso molecular são os que causam a maior parte das respostas agudas do SN, tais como a transmissão dos sinais sensoriais em direcção ao cérebro e sinais motores em direcção aos músculos. Os neuropéptidos, por outro lado, causam comummente acções mais prolongadas, tais como as modificações a longo prazo do número de receptores, o fecho duradouro de certos canais iónicos e, possivelmente, as alterações a longo prazo do número de sinapses.
NT de baixo peso molecular São quase sem excepção sintetizados no citosol do terminal pré-sináptico e absorvidos, por transporte activo, para o interior das vesículas transmissoras aí localizadas. Então, cada vez que um potencial de acção chega ao terminal pré-sináptico, algumas vesículas, por exocitose, liberam, ao mesmo tempo, o transmissor para o interior da fenda sináptica, dentro de 1 milissegundo ou menos. A acção subsequente desses tipos de transmissores sobre os receptores da membrana também ocorre comummente dentro de 1 ms ou menos. Mais comummente, o efeito é aumento da condutância dos canais iónicos; como exemplo temos o aumento da condutância ao sódio (que causa excitação) ou o aumento da condutância ao potássio (que causa inibição). No entanto, os tipos de transmissores de baixo peso molecular podem, ocasionalmente, estimular as enzimas receptores-ativadas, em lugar de abrir canais iónicos, provocando alterações do funcionamento da máquina metabólica intracelular.
Reciclagem das vesículas As vesículas que armazenam e liberam os transmissores de baixo peso molecular são continuamente recicladas, isto é, são usadas várias e várias vezes. Após se fundirem com a membrana sináptica e liberarem o transmissor, as vesículas, em primeira instância, simplesmente se tornam parte da membrana sináptica. No entanto, dentro de alguns segundos ou minutos, a parte da membrana correspondente à vesícula se invagina para o interior do terminal pré-sináptico e se desprende, para formar uma nova vesícula. Essas membranas ainda contêm as proteínas de transporte apropriadas, necessárias para que nova quantidade de substância transmissora se concentre no interior da vesícula.
A acetilcolina é um típico transmissor de baixo peso molecular que obedece os princípios acima com relação à síntese e libertação. Sintetizada no terminal pré-sináptico a partir de acetil-coenzima A (acetil-CoA) e colina
na presença da enzima colina acetiltransferase; Transportada para o interior de vesículas específicas; As vesículas libertam a acetilcolina na fenda sináptica, onde é quebrada em acetato e
colina sob a acção da enzima colinesterase, que está ligada ao retículo proteoglicano que enche o espaço da fenda sináptica;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
31 | P á g i n a
As vesículas são recicladas e a colina é transportada activamente de volta para o interior do terminal para ser usada, novamente, na síntese de uma nova molécula de acetilcolina.
Nota: Para uma substância assumir funções de NT, é necessário: Ser sintetizada por neurónios; Ser sujeita a processos em vesículas, quando armazenadas; Libertação ser exocitótica; Ter receptores para a substância na membrana.
Exemplos de NT de baixo peso molecular
Acetilcolina
- NT do SNP mediador químico do SNA simpático e parassimpático; - Participa em vias motoras e vegetativas; - Função: memória, aprendizagem e controlo do movimento - efeito excitatório e inibitório (em algumas terminações nervosas parassimpáticas periféricas, tais como a inibição do coração pelos nervos vagos); - Localização: → Junção neuromuscular; → Gânglios do SNA; → Terminações do SNP; → SNC (hipocampo, vias reticulares ascendentes, fibras aferentes para sistemas visual e auditivo); - Os receptores da acetilcolina (receptores colinérgicos) podem ser:
Metabotrópicos – receptores muscarínicos (M1, M2 e M3) presentes no músculo liso e
glândulas. Podem ter efeito inibitório ou excitatório;
Ionótropicos – receptores nicotínicos, os quais são activados pela nicotina e pela
acetilcolina. Estes são canais iónicos (presentes nos gânglios simpáticos) selectivos
para catiões (sódio e potássio), ou seja, são permeáveis ao cloro. A acetilcolina liga-se
directamente ao receptor.
Glutamato
- NT ubiquitário – principal NT excitatório no cérebro e medula espinhal; - Captado pelos astrócitos e convertido em glutamina (acção da glutamina sintetase), a qual volta a glutamato nos neurónios excitatórios (acção da glutaminase); - Uma vez formado, pode ser recaptado pelas células gliais ou pelo terminal sináptico; - Os receptores do glutamato podem ser: Metabotrópicos – receptores AMPA; Ionotrópicos – canais iónicos.
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
32 | P á g i n a
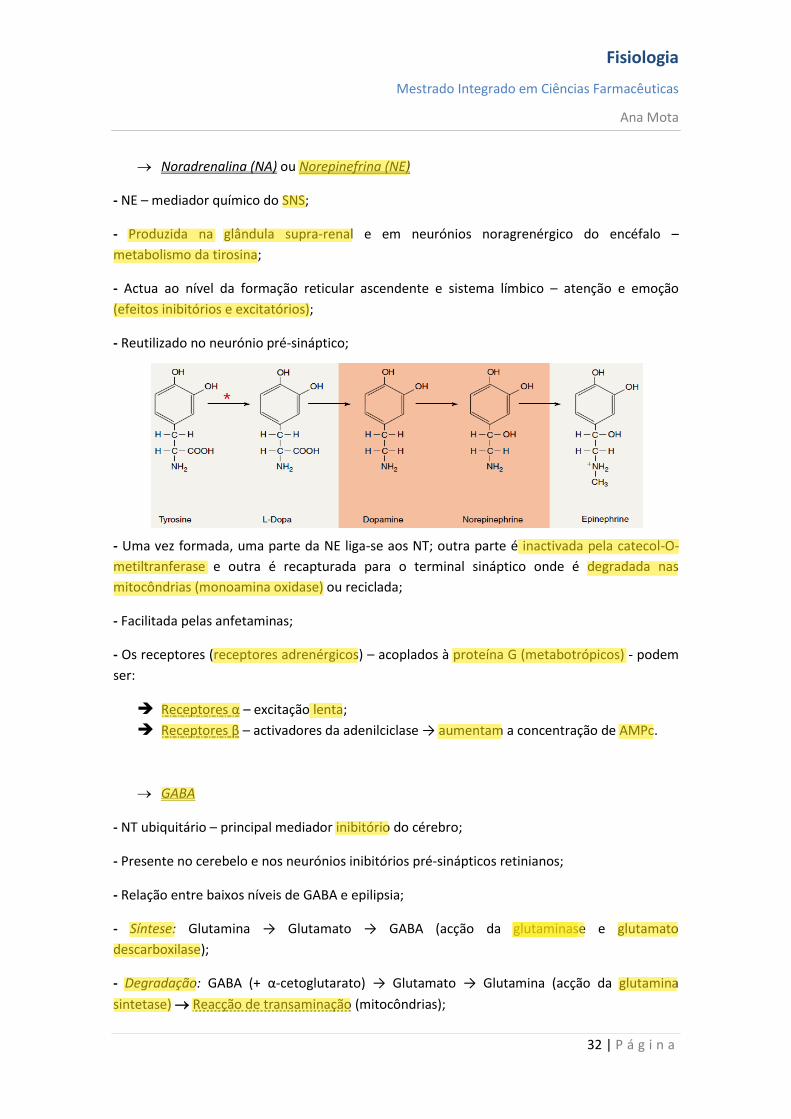
Noradrenalina (NA) ou Norepinefrina (NE)
- NE – mediador químico do SNS;
- Produzida na glândula supra-renal e em neurónios noragrenérgico do encéfalo –
metabolismo da tirosina;
- Actua ao nível da formação reticular ascendente e sistema límbico – atenção e emoção
(efeitos inibitórios e excitatórios);
- Reutilizado no neurónio pré-sináptico;
- Uma vez formada, uma parte da NE liga-se aos NT; outra parte é inactivada pela catecol-O-
metiltranferase e outra é recapturada para o terminal sináptico onde é degradada nas
mitocôndrias (monoamina oxidase) ou reciclada;
- Facilitada pelas anfetaminas;
- Os receptores (receptores adrenérgicos) – acoplados à proteína G (metabotrópicos) - podem
ser:
Receptores α – excitação lenta;
Receptores β – activadores da adenilciclase → aumentam a concentração de AMPc.
GABA
- NT ubiquitário – principal mediador inibitório do cérebro;
- Presente no cerebelo e nos neurónios inibitórios pré-sinápticos retinianos;
- Relação entre baixos níveis de GABA e epilipsia;
- Síntese: Glutamina → Glutamato → GABA (acção da glutaminase e glutamato
descarboxilase);
- Degradação: GABA (+ α-cetoglutarato) → Glutamato → Glutamina (acção da glutamina
sintetase) Reacção de transaminação (mitocôndrias);

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
33 | P á g i n a
- Os receptores podem ser:
Ionotrópicos - aumentam a entrada de Cl-;
Metabotrópicos - diminuem a entrada de Ca2+.
Dopamina
- Produzida na glândula supra-renal e neurónios noradrenérgicos do encéfalo – metabolismo
da tirosina;
- Receptores:
Dopaminérgicos – associados à proteína G, aumentam e diminuem a concentração de
AMPc consoante o subtipo de receptores;
- Após actuar, pode ser recapturada ou catabolizada;
- Acção inibitória ou excitatória – aprendizagem;
- Presente em vias extra-piramidais, eminência mediana, sistema mesocorticolímbico e sistema
límbico.
Serotonina
- Metabolismo do triptofano;
- Produzida nas plaquetas e tracto gastrointestinal, encéfalo e retina;
- 7 tipos de receptores:
Seroroninérgicos;
- Após actuar, pode ser recapturada ou catabolizada;
- Acção inibitória e excitatória – processo emotivos;
- Presente no sistema límbico e núcleos da raphe.
Histamina
- Metabolismo da histidina;
- Sintetizada nos mastócitos da mucosa gástrica e hipófise;
- 3 tipos de receptores:

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
34 | P á g i n a
Histaminérgicos;
- Acção excitatória;
- Presente em neurónios do núcleo tuberomamilar do hipotálamo posterior, sendo daí
distribuída para outras regiões do encéfalo;
- Funções na libido, comportamento sexual, secreção de hormonas, pressão arterial, sede e
limiar da dor.
Glicina
- Aminoácido inibitório;
- Auxilia o glutamato no desempenho da função dos receptores ionotrópicos;
- Receptores inibitórios (abrem canais de Cl-).
Aspartato
- Aminoácido excitatório;
- Células piramidais corticais e estreladas espinhais do córtex visual.
Pode-se ainda identificar:
Polipéptideos: peptídeo opióides, substância P, vasopressina, oxitocina, entre outros;
Transmissores purinérgicos: adenosina e ATP;
Transmissores gasosos: NO e CO;
Transmissor lipídico: anandamina.
Transmissão não-vesicular
A transmissão sináptica é frequentemente mediada por gases voláteis que não necessitam da
exocitose vesicular para saírem do botão terminal. Logo que são sintetizados (NO e CO)
difundem-se em todas as direcções.
NO estimula a produção de GMPc
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
35 | P á g i n a
Convergência e divergência
Integração sináptica
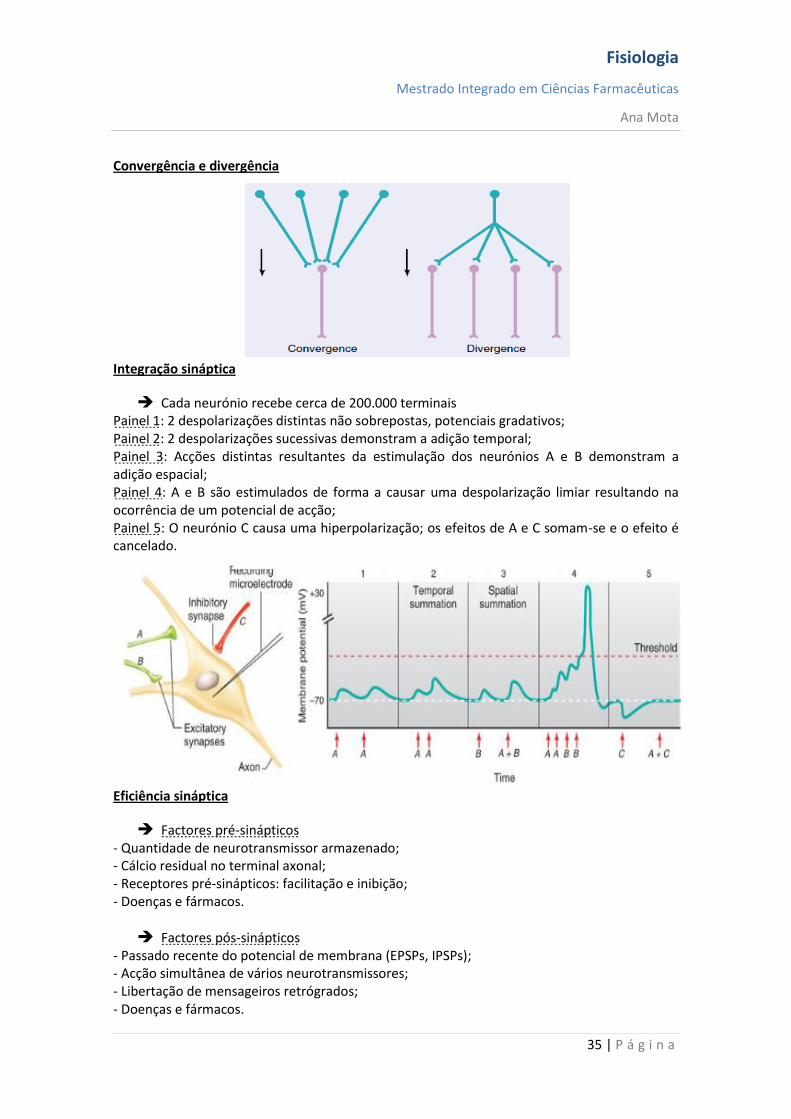
Cada neurónio recebe cerca de 200.000 terminais Painel 1: 2 despolarizações distintas não sobrepostas, potenciais gradativos; Painel 2: 2 despolarizações sucessivas demonstram a adição temporal; Painel 3: Acções distintas resultantes da estimulação dos neurónios A e B demonstram a adição espacial; Painel 4: A e B são estimulados de forma a causar uma despolarização limiar resultando na ocorrência de um potencial de acção; Painel 5: O neurónio C causa uma hiperpolarização; os efeitos de A e C somam-se e o efeito é cancelado.
Eficiência sináptica
Factores pré-sinápticos - Quantidade de neurotransmissor armazenado; - Cálcio residual no terminal axonal; - Receptores pré-sinápticos: facilitação e inibição; - Doenças e fármacos. Factores pós-sinápticos
- Passado recente do potencial de membrana (EPSPs, IPSPs); - Acção simultânea de vários neurotransmissores; - Libertação de mensageiros retrógrados; - Doenças e fármacos.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
36 | P á g i n a
Neuromodulação
Na membrana pós-sináptica podem formar-se neuromensageiros (lipossolúveis) que podem
modificar a resposta do neurónio pré-sináptico.
Acção dos neuromoduladores:
Modulam e modificam a actividade do neurónio pós-sináptico;
Podem coexistir com o NT mas em vesículas distintas;
Os NT têm um efeito rápido directamente sobre a membrana pós-sináptica, mas os
moduladores não têm um efeito directo sobre ela;
A sua função é reforçar, prolongar, inibir ou limitar o efeito do NT;
Actuam por meio de 2º mensageiro, alterando a resposta do receptor do NT;
Grande variedade de respostas.
Alvo de Fármacos
Alvos pré-sinápticos A. Libertação ou degradação do NT no terminal axonal; B. Aumento da libertação de NT; C. Inibição da libertação do NT; D. Inibição da síntese de NT; E. Redução da captação de NT. Alvos pós-sinápticos F. Redução da degradação do NT na fenda sináptica; G. Acção de agonistas ou antagonistas dos receptores; H. Alteração da resposta da célula pós-sináptica.
Regeneração Neuronal
Hipocampo: formação de memória –
origem de neurónios em ritmo reduzido;
Se o corpo celular intacto é possível
recuperar a função do neurónio –
rapidamente antes da acção das células
gliais;
Se o corpo celular lesado – degenera;
Se o axónio lesado – regenera.
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
37 | P á g i n a
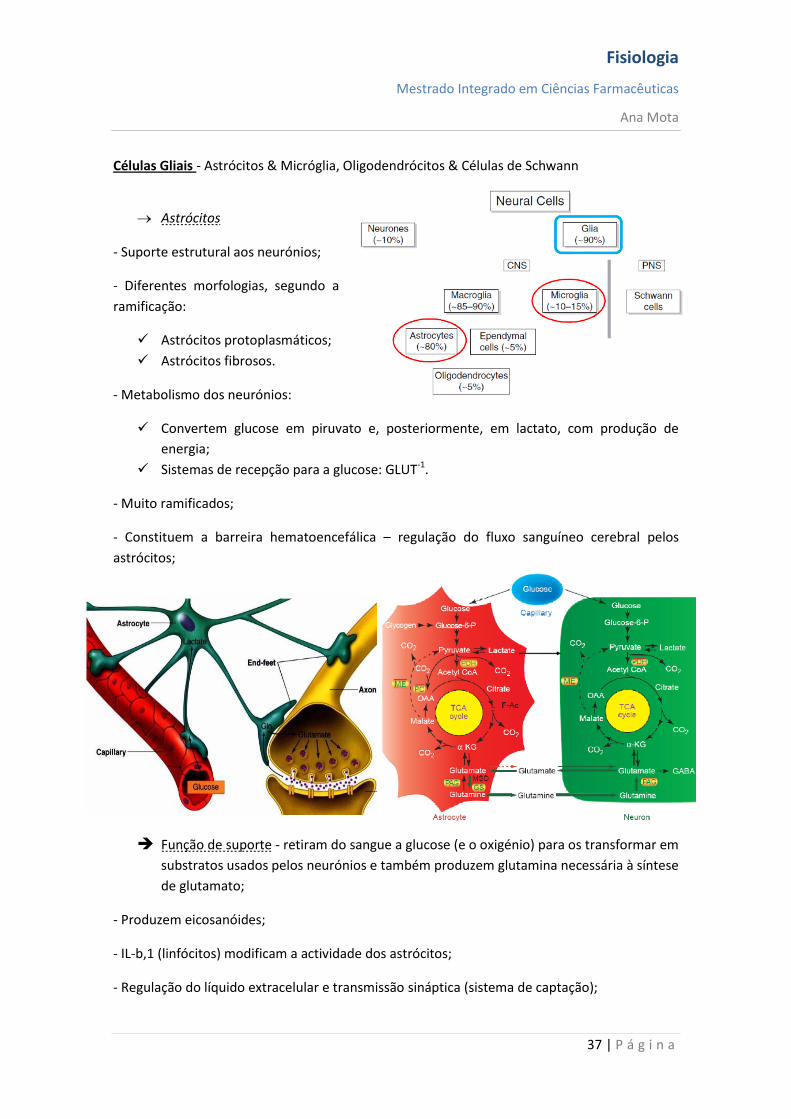
Células Gliais - Astrócitos & Micróglia, Oligodendrócitos & Células de Schwann
Astrócitos
- Suporte estrutural aos neurónios;
- Diferentes morfologias, segundo a
ramificação:
Astrócitos protoplasmáticos;
Astrócitos fibrosos.
- Metabolismo dos neurónios:
Convertem glucose em piruvato e, posteriormente, em lactato, com produção de
energia;
Sistemas de recepção para a glucose: GLUT-1.
- Muito ramificados;
- Constituem a barreira hematoencefálica – regulação do fluxo sanguíneo cerebral pelos
astrócitos;
Função de suporte - retiram do sangue a glucose (e o oxigénio) para os transformar em
substratos usados pelos neurónios e também produzem glutamina necessária à síntese
de glutamato;
- Produzem eicosanóides;
- IL-b,1 (linfócitos) modificam a actividade dos astrócitos;
- Regulação do líquido extracelular e transmissão sináptica (sistema de captação);

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
38 | P á g i n a
- Envolvidos na regulação da atenção e vigília-sono.
Oligodendrócitos e células de Schwann
- Produzem mielina (reveste os axónios) – condução mais eficiente de potenciais;
- ATP induz os oligodendrócitos a produzir a bainha de mielina.
Micróglia
- Células da micróglia:
Importantes na estrutura do tecido nervoso – detectam as substâncias que entram
neste;
Afecta a nível do sistema imunitário, uma vez que intervém na defesa do SN.
- Diferentes ramificações relativamente aos astrócitos
- Funções de defesa do SN.
Funções:
Regulação da composição do LEC;
Suporte energético para os neurónios;
Regulação do fluxo sanguíneo;
Formação da barreira hematoencefálica;
Regulação da transmissão sináptica:
- Concentração sináptica de NT;
- Libertação de neuromodeladores
e gliotransmissores, os quais
regulam a quantidade de NT
libertado – ciclo da glutamina: o
glutamato e o GABA são
convertidos em glutamina, sendo
incorporados pelos astrócitos,
podendo ser libertados.
Desenvolvimento e
plasticidade neuronal:
- Comunicação neurónio – astrócito;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
39 | P á g i n a
- Orientação da migração neuronal;
- Mielinização.
Resposta inflamatória (micróglia e astrócitos):
- Astrogliose reactiva;
- Activação e proliferação da micróglia;
- Neuroprotecção / Neurodegeneração.
Estudo das células gliais: Culturas
LPS modifica a morfologia das células da micróglia, tornando-as mais eficazes no combate às infecções;
Moléculas com funções fisiológicas em microrganismos. Astrócitos/Neurónios
Relação entre a proporção de neurónios e astrócitos em termos filogenéticos.
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
40 | P á g i n a
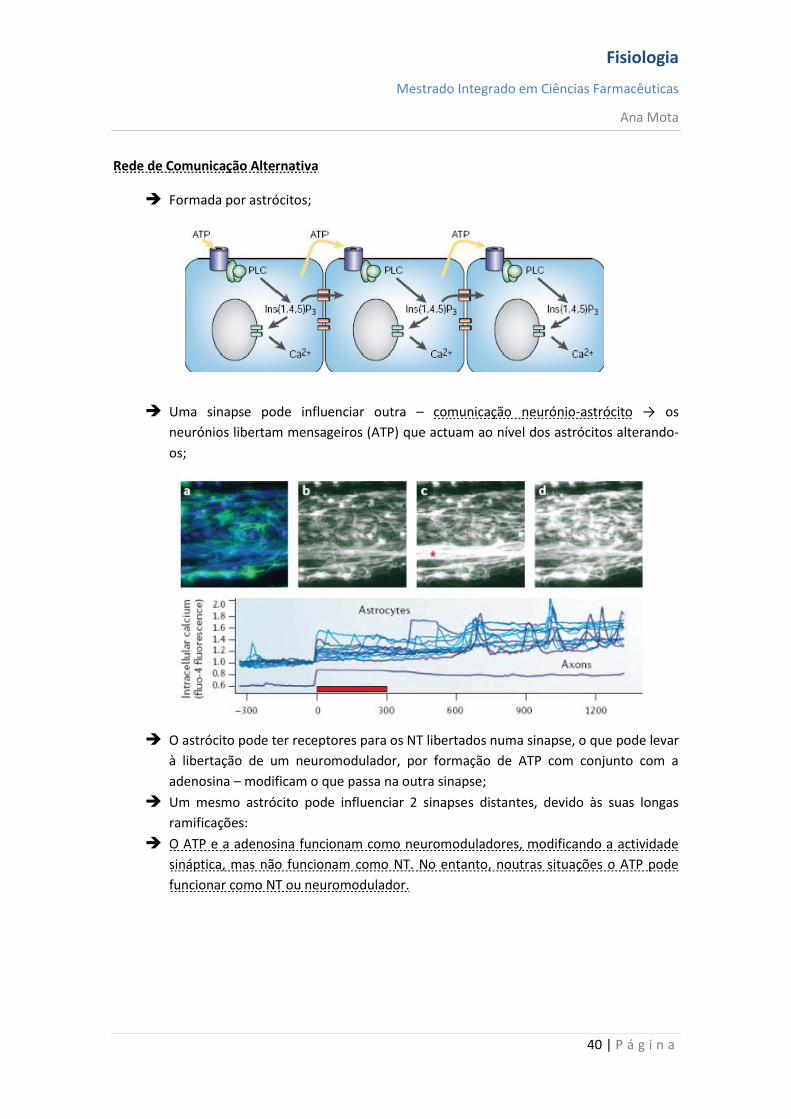
Rede de Comunicação Alternativa
Formada por astrócitos;
Uma sinapse pode influenciar outra – comunicação neurónio-astrócito → os
neurónios libertam mensageiros (ATP) que actuam ao nível dos astrócitos alterando-
os;
O astrócito pode ter receptores para os NT libertados numa sinapse, o que pode levar
à libertação de um neuromodulador, por formação de ATP com conjunto com a
adenosina – modificam o que passa na outra sinapse;
Um mesmo astrócito pode influenciar 2 sinapses distantes, devido às suas longas
ramificações:
O ATP e a adenosina funcionam como neuromoduladores, modificando a actividade
sináptica, mas não funcionam como NT. No entanto, noutras situações o ATP pode
funcionar como NT ou neuromodulador.
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
41 | P á g i n a
Nota:
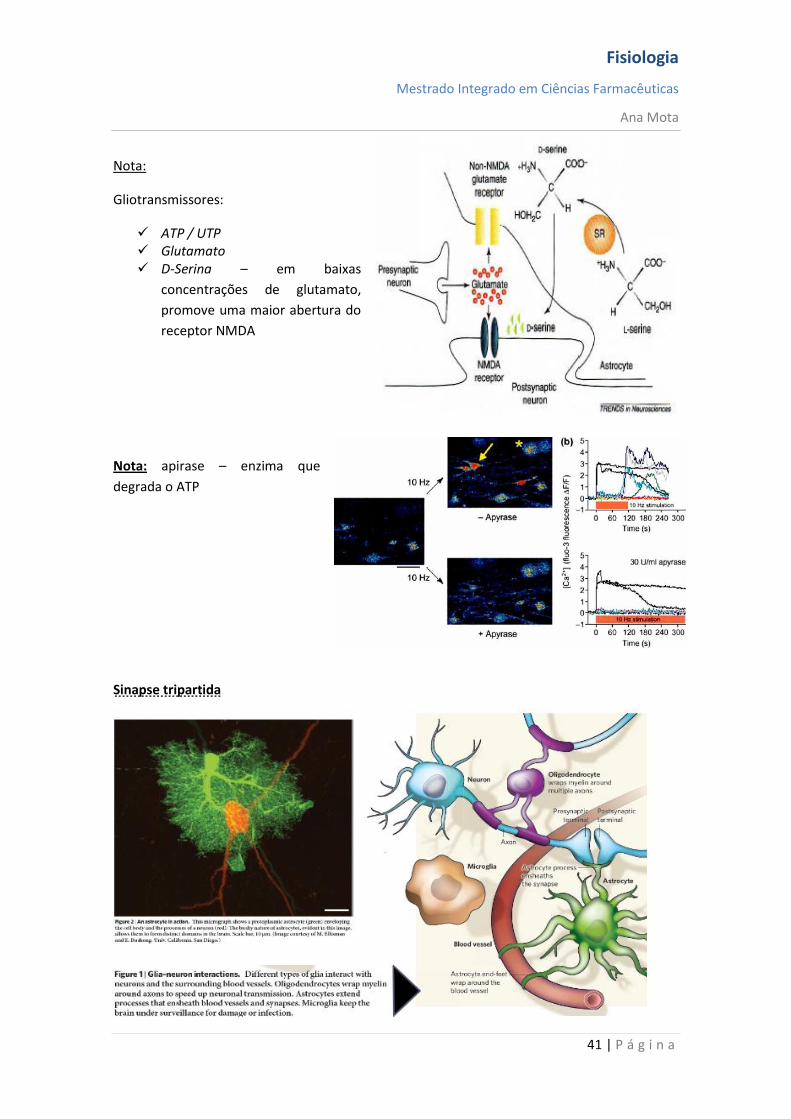
Gliotransmissores:
ATP / UTP Glutamato D-Serina – em baixas
concentrações de glutamato,
promove uma maior abertura do
receptor NMDA
Nota: apirase – enzima que
degrada o ATP
Sinapse tripartida
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
42 | P á g i n a
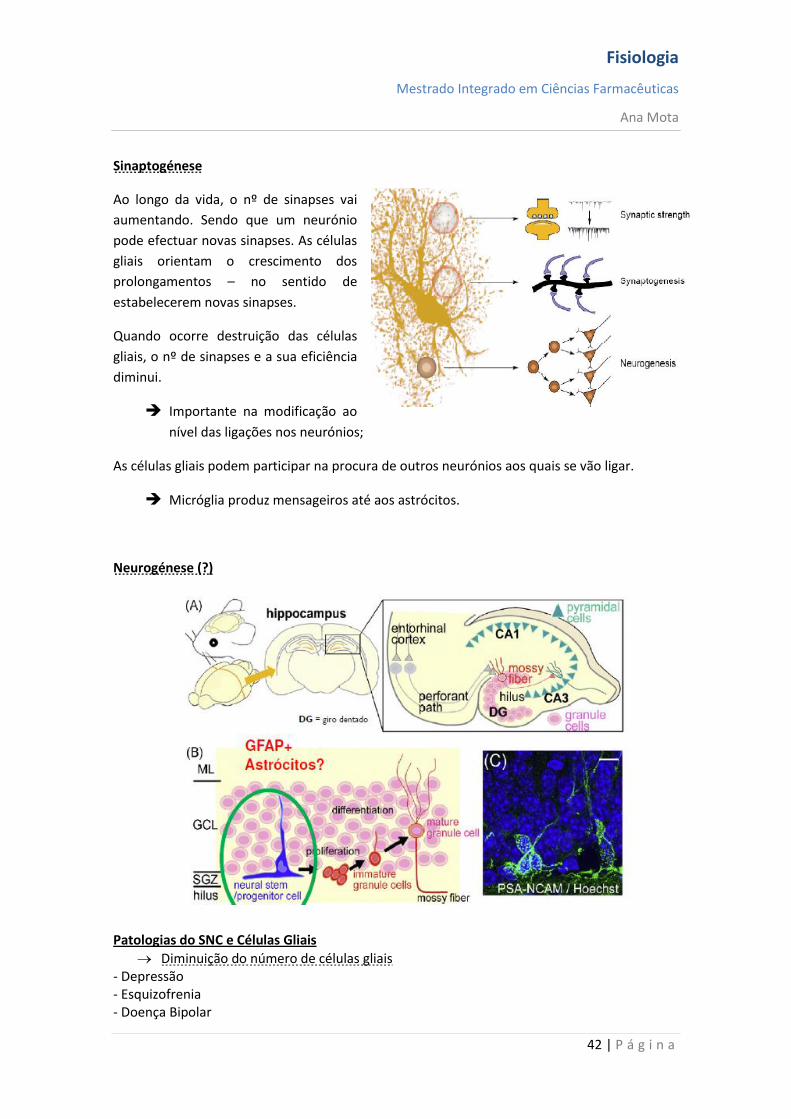
Sinaptogénese
Ao longo da vida, o nº de sinapses vai
aumentando. Sendo que um neurónio
pode efectuar novas sinapses. As células
gliais orientam o crescimento dos
prolongamentos – no sentido de
estabelecerem novas sinapses.
Quando ocorre destruição das células
gliais, o nº de sinapses e a sua eficiência
diminui.
Importante na modificação ao
nível das ligações nos neurónios;
As células gliais podem participar na procura de outros neurónios aos quais se vão ligar.
Micróglia produz mensageiros até aos astrócitos.
Neurogénese (?)
Patologias do SNC e Células Gliais
Diminuição do número de células gliais - Depressão - Esquizofrenia - Doença Bipolar

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
43 | P á g i n a
Participação na resposta inflamatória - Esclerose múltipla - Dor/Hiperalgesia/Alodínia - Alzheimer - Parkinson
DOR / Nocicepção
Conjunto das percepções de dor que somos
capazes de distinguir. Está intimamente ligada
ao sistema emocional, bem como se relaciona
com vários outros sistemas do corpo tais como
o imunitário. Dor pode ser induzida num
receptor de três formas específicas:
Mecânica - Quando ocorre um
estímulo mecânico capaz de excitar
um mecanoreceptor de dor (receptor
estimulado por meios físicos, tais
como, impacto, fricção,
tracionamento, rompimento).
Ex: Corte, pancada, perfuração, abrasão, pressão.
Térmica - Quando ocorre um estímulo térmico que é capaz de excitar um
termoceptor de dor (receptor sensível ao calor ou ao frio). Ex: Queimaduras até
segundo grau.
Obs: no caso das queimaduras pode ocorrer quimiocepção por extravasamento de
fluidos celulares que mediam a dor, mas isso será explicado abaixo.
Química - Quando ocorre estímulo mediado por mecanismo químico em um
quimioceptor (receptor sensível a alterações químicas do meio ou a ligação de um
mediador químico ou neurotransmissor).
Ex: Na inflamação (seja ela aguda ou crónica) ocorre destruição de algumas células ou
mesmo liberação (sem morte) de algumas substâncias que são capazes de estimular
algumas terminações nervosas (Ex: prostaglandina) ou hipersensibilizá-las a outro
estímulo.
Nota: A nocicepção é o componente fisiológico da dor e compreende os processos de
transdução, transmissão e modulação do estímulo nociceptivo; Nociceptor é um receptor sensorial que envia sinal que causa a percepção da dor em
resposta a um estímulo que possui potencial de dano. Nociceptores são terminações nervosas responsáveis pela nocicepção.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
44 | P á g i n a
GLUTI e GLAST – não captam o glutamato e este acumula-se na fenda.
Doenças neurodegenerativas
Esclerose múltipla
- Doença neurológica crónica;
- Causa desconhecida – maior
incidência em indivíduos de
raça caucasiana;
- Ocorre destruição da bainha
de mielina, provavelmente
causada pela destruição, pelo
próprio sistema imunitário,
das células gliais (que
produzem mielina);
- A ausência de isolamento eficaz dos neurónios (e da sua sustentação pelas células gliais) leva
à perda de função e degeneração das mesmas;
- Caracterizada por:
Fraqueza muscular;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
45 | P á g i n a
Rigidez e dor articular;
Descoordenação motora;
Tremores;
Formigueiros em algumas partes do corpo.
Doença de Alzheimer
- Incidência em pessoas idosas;
- Caracterizada por:
Acumulação de placas amilóides em áreas amplamente dispersas do cérebro, incluindo
córtex cerebral, hipocampo e gânglios da base;
- A demência grave poderá estar a perda da entrada colinérgica para o córtex cerebral,
resultante da perda dos neurónios no núcleo basal de Meynert;
- Muitos pacientes exibem uma anomalia genética relacionada com a apoliproteína E, uma
proteína que transporta colesterol;
- O diagnóstico definitivo só pode ser confirmado pela autópsia.
SIDA
3 – Acumulação de Ca2+
intracelular e lesão neuronal
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
46 | P á g i n a
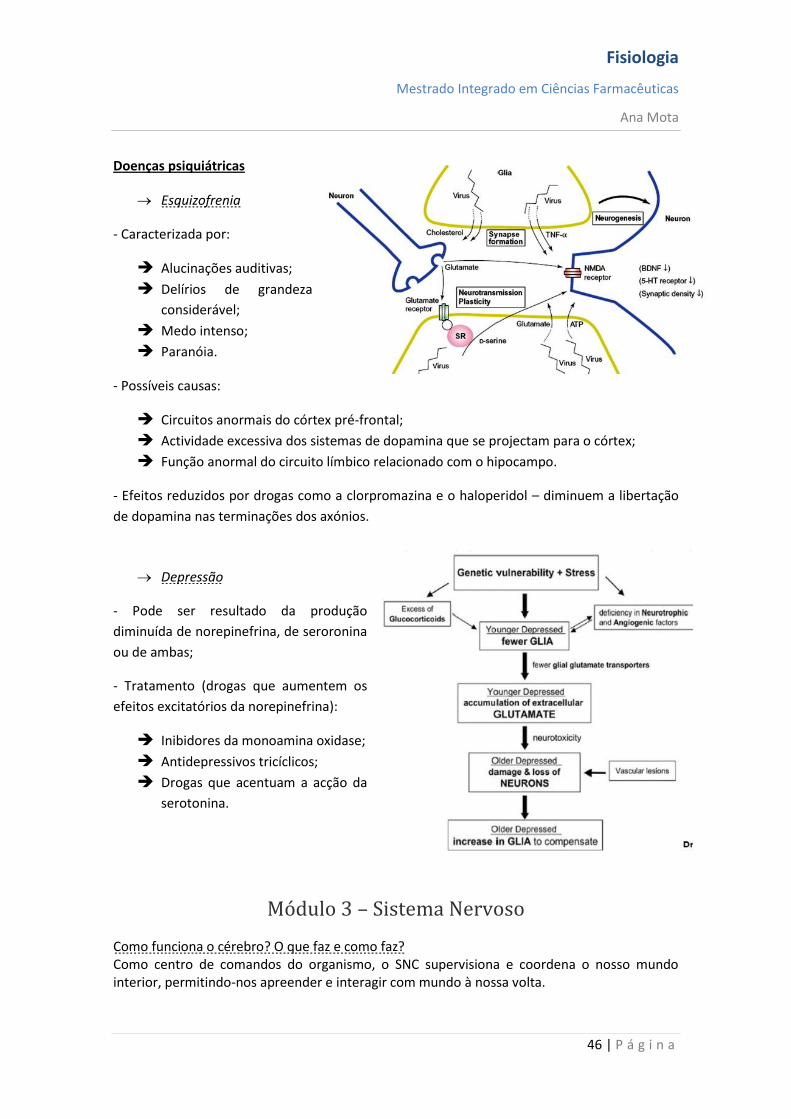
Doenças psiquiátricas
Esquizofrenia
- Caracterizada por:
Alucinações auditivas;
Delírios de grandeza
considerável;
Medo intenso;
Paranóia.
- Possíveis causas:
Circuitos anormais do córtex pré-frontal;
Actividade excessiva dos sistemas de dopamina que se projectam para o córtex;
Função anormal do circuito límbico relacionado com o hipocampo.
- Efeitos reduzidos por drogas como a clorpromazina e o haloperidol – diminuem a libertação
de dopamina nas terminações dos axónios.
Depressão
- Pode ser resultado da produção
diminuída de norepinefrina, de seroronina
ou de ambas;
- Tratamento (drogas que aumentem os
efeitos excitatórios da norepinefrina):
Inibidores da monoamina oxidase;
Antidepressivos tricíclicos;
Drogas que acentuam a acção da
serotonina.
Módulo 3 – Sistema Nervoso
Como funciona o cérebro? O que faz e como faz? Como centro de comandos do organismo, o SNC supervisiona e coordena o nosso mundo interior, permitindo-nos apreender e interagir com mundo à nossa volta.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
47 | P á g i n a
No SNC, é possível identificar dois tipos distintos de tecido, com base no seu aspecto:
Substância cinzenta – corpos celulares dos axónios;
Substância branca – axónios de nervos mielinizados e oligodendrócitos.
Constituição celular do SNC:
Neurónios;
Células gliais: oligodendrócitos, micróglia, células ependimais e astrócitos;
Capilares sanguíneos.
Regiões do cérebro:
Tronco cerebral - Formação reticular - Bolbo raquidiano - Mesencéfalo
Cerebelo
Diencéfalo - Tálamo - Hipotálamo
Telencéfalo - Córtex Cerebral - 2 Hemisférios - 4 Lóbulos

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
48 | P á g i n a
Hemisférios Cerebrais:
Corpo Caloso
Controlo Contralateral
Divisão de tarefas - Linguagem: escrita e fala - Linguagem: compreensão - Cálculo matemático - Orientação espacial - Gerir emoções
Heminegligência - Barbear só metade da face - Ingerir comida de 1 lado do prato - Mover apenas 1 lado do corpo - Sentir apenas 1 lado do corpo
Córtex Cerebral: áreas de Brodmann
Mapa do córtex cerebral humano, mostrando a divisão em cerca de 50 áreas distintas, divisão essa feita em função de diferenças estruturais histológicas, denominadas áreas de Brodmann. O mapa é importante, uma vez que veio a ser utilizado por praticamente todos os neurofisiologistas e neurologistas para se referirem às diferentes áreas funcionais do córtex humano. O "sulco central" estende-se verticalmente pelo cérebro. Em geral, os sinais sensoriais correspondentes a todas as modalidades de sensação terminam no córtex cerebral posterior à fissura central. Mais importante que isso, o córtex somatossensorial situa-se imediatamente atrás do sulco central, localizado principalmente nas áreas 1, 2, 3, 5, 7 e 40 de Brodmann, correspondendo ao lobo parietal do córtex. Além disso, os sinais visuais terminam no lobo occipital e os sinais auditivos, no lobo temporal. A área cortical anterior ao sulco central está envolvida com o controlo motor do corpo e com alguns aspectos do pensamento analítico.
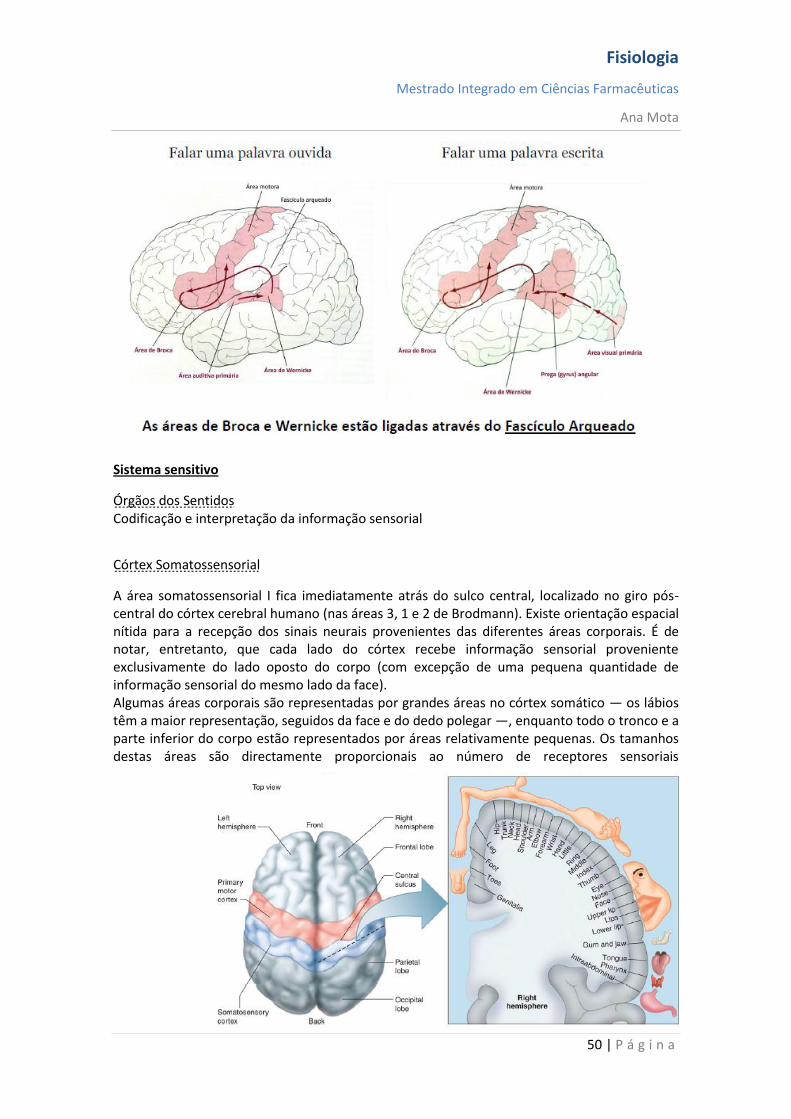
Linguagem
Controlada no lobo temporo-parietal – área
de Wernick;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
49 | P á g i n a
Alterações nesta área provocam a não compreensão das palavras, o sentido, a
coerência do discurso – sabem ouvir e ler mas não entendem aquilo que dizem ou
escrevem;
Visão – córtex visual;
Audição – córtex temporal;
Lateralidade mal definida – a pessoa perde a noção de uma parte do corpo;
Predominante o controlo do hemisfério esquerdo;
Entrada da linguagem – aspecto sensorial;
Saída da linguagem – aspecto motor.
Área de Wernick
- Área parirtoccipitotemporal do córtex (área da associação);
- Transmite informação para a área de Broca, transformando aquilo que foi “pensado” em
palavras;
- Define os neurónios que devem ser activados para movimentar os músculos relacionados
com a fala – córtex motor primário para falar;
- Lesão nesta área resulta em:
Incapacidade de compreender a linguagem falada ou escrita;
Afasia de Wernicke ou afasia sensorial ou receptiva: produção de palavras, escritas e faladas de modo normal; sequência e contexto desfasados - “salada de palavras”;
Se a lesão se estender para além dos limites desta área – incapacidade total de usar a comunicação pela linguagem – afasia global.
Área de Broca
- Área pré-frontal do córtex (área da associação);
- Fornece a saída para o córtex motor próximo que controla os músculos necessários para a
produção da fala;
- Afasia de Broca: produzem poucas palavras, escritas ou faladas e com grande dificuldade; referem apenas as mais importantes, falam de modo “telegráfico”.
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
50 | P á g i n a
Sistema sensitivo
Órgãos dos Sentidos Codificação e interpretação da informação sensorial
Córtex Somatossensorial
A área somatossensorial I fica imediatamente atrás do sulco central, localizado no giro pós-central do córtex cerebral humano (nas áreas 3, 1 e 2 de Brodmann). Existe orientação espacial nítida para a recepção dos sinais neurais provenientes das diferentes áreas corporais. É de notar, entretanto, que cada lado do córtex recebe informação sensorial proveniente exclusivamente do lado oposto do corpo (com excepção de uma pequena quantidade de informação sensorial do mesmo lado da face). Algumas áreas corporais são representadas por grandes áreas no córtex somático — os lábios têm a maior representação, seguidos da face e do dedo polegar —, enquanto todo o tronco e a parte inferior do corpo estão representados por áreas relativamente pequenas. Os tamanhos destas áreas são directamente proporcionais ao número de receptores sensoriais

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
51 | P á g i n a
especializados, existentes em cada região corporal periférica correspondente. Por exemplo, nos lábios e no polegar, há grande número de terminações nervosas especializadas, enquanto na pele do tronco o número dessas terminações é relativamente pequeno. A cabeça está representada na porção mais lateral da área sensorial somática I, enquanto as regiões corporais mais inferiores estão representadas mais medialmente.
Representação da localização e do tamanho relativo das diferentes partes do corpo em função do número de neurónios motores do córtex cerebral que controlam as diferentes regiões corporais.
Categorias Funcionais • Tipo de estímulo adequado (Forma de Energia)
Mecanoreceptores - Constituídos por canais iónicos fixos à membrana pelo citosqueleto;
- Estímulo Alteração na célula Abertura dos canais Fluxo de iões Diferença de
potencial Resposta. Quimioreceptores
- Não são canais iónicos mas estão relacionados;
- Produzem mensageiros que alteram a configuração dos canais iónicos Diferença de
potencial Resposta. Fotoreceptores
- Luz Alteração do receptor Via geradora de 2º mensageiro Abertura do canal; - Canais iónicos. Termoreceptores
• Origem do estímulo adequado (Localização do Receptor)
Exteroreceptores Propioreceptores Interoreceptores
• Experiência sensorial subjectiva Nociceptores (dor) Receptores do frio ou calor
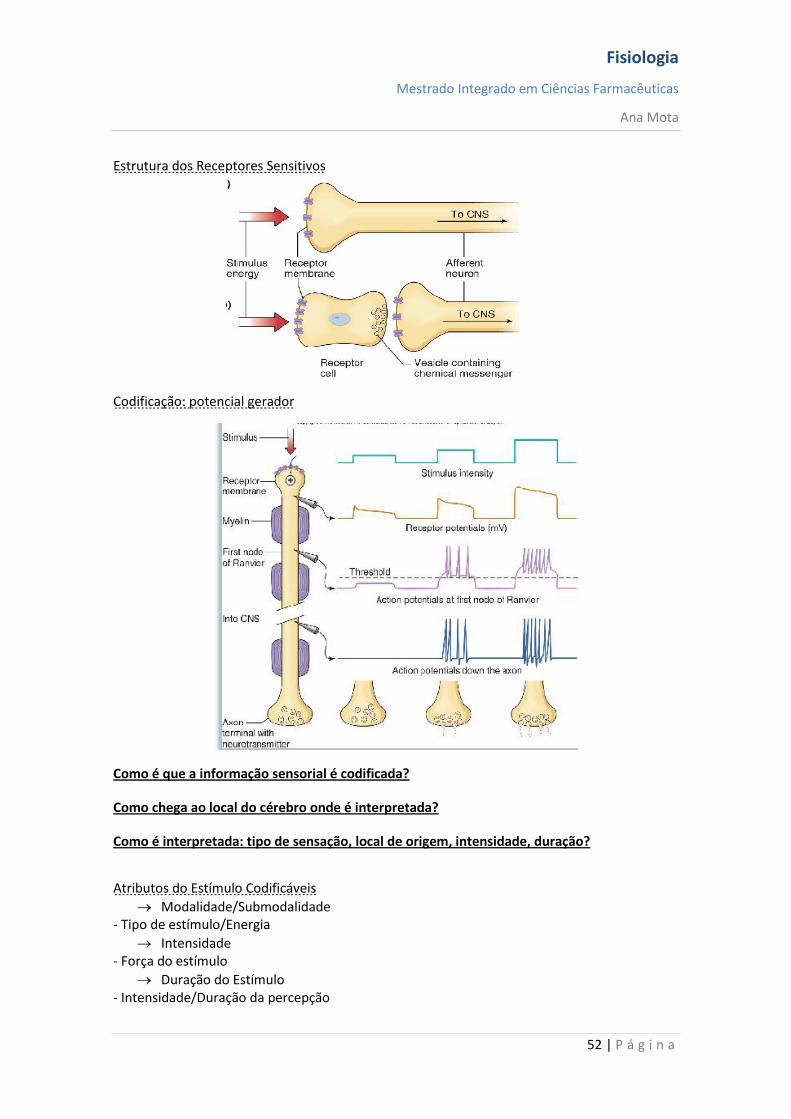
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
52 | P á g i n a
Estrutura dos Receptores Sensitivos
Codificação: potencial gerador
Como é que a informação sensorial é codificada?
Como chega ao local do cérebro onde é interpretada?
Como é interpretada: tipo de sensação, local de origem, intensidade, duração?
Atributos do Estímulo Codificáveis
Modalidade/Submodalidade - Tipo de estímulo/Energia
Intensidade - Força do estímulo
Duração do Estímulo - Intensidade/Duração da percepção

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
53 | P á g i n a
Localização - Local do corpo onde é aplicado - Acuidade sensitiva
Modalidade Sensitiva
Relacionado com o tipo de receptor e a forma de energia que o activa;
Visão: receptores sensíveis a diferentes comprimentos de onda de radiação variável;
Temperatura: receptores sensíveis a diferentes temperaturas;
Pressão/tacto: receptor activado por forças mecânicas.
Lei das Energias Específicas:
A estimulação de um neurónio sensitivo produz apenas um tipo de sensação - Tacto, Pressão, Frio, Dor...
A sensação característica que cada neurónio sensitivo transmite é apenas aquela produzida pelo estímulo que normalmente o activa - Estímulo Adequado. Exemplo: receptores da visão (bastonetes e cones):
Especializadas na recepção da luz;
Luz é o estímulo adequado. Nota: Receptores sensitivos: - Célula sensitiva; - Receptores da membrana (em todas as células e nos receptores sensitivos).
Sensibilidade Somática
São conhecidos pelo menos seis tipos inteiramente diferentes de receptores tácteis, mas há muitos outros tipos semelhantes a eles.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
54 | P á g i n a
Terminações nervosas livres, que são encontradas em toda a pele e em muitos outros tecidos, podem detectar toque e pressão. Como exemplo, podemos citar as sensações de toque e pressão provocadas pelo contacto suave com a córnea do olho, apesar dessa estrutura não ter outras terminações nervosas além das terminações nervosas livres;
Um receptor de toque de sensibilidade especial é o corpúsculo de Meissner, uma terminação nervosa encapsulada alongada que excita uma fibra nervosa sensorial de grande diâmetro (tipo Aβ). Dentro da cápsula há várias espirais de filamentos nervosos terminais. Esses receptores estão presentes na pele que não possui pêlos (a chamada pele glabra) e são particularmente abundantes nas pontas dos dedos, nos lábios e em outras áreas da pele onde está altamente desenvolvida a capacidade de discernir as características espaciais das sensações de toque. Os corpúsculos de Meissner adaptam-se em fracção de segundos após serem estimulados, o que indica que são particularmente sensíveis ao movimento de objectos muito leves sobre a superfície da pele e, também, a vibrações de baixa frequência;
A ponta dos dedos e outras áreas que contêm os corpúsculos de Meissner também contêm grande número de receptores tácteis de ponta expandida, o tipo a que pertencem os discos de Merkel. A parte pilosa da pele também contém uma quantidade moderada de receptores de ponta expandida, apesar de praticamente não serem encontrados nesses locais os corpúsculos de Meissner. Esses receptores diferem dos corpúsculos de Meissner pelo fato de transmitirem um sinal inicialmente forte, mas que se adapta parcialmente, ao qual se segue um sinal contínuo mais fraco, de adaptação muito lenta. Por isso, eles são responsáveis pelo fornecimento de sinais de situações estacionárias, que permitem a detecção do toque continuado de objectos sobre a pele. Os discos de Merkel agrupam-se formando um só órgão receptor, denominado receptor de cúpula de Iggo;
O movimento suave de qualquer pêlo do corpo estimula a fibra nervosa que está entrelaçada em sua base. Assim, cada pêlo tem sua fibra nervosa basal, denominada órgão piloso terminal, que também é um receptor de toque. Esse receptor se adapta facilmente e, da mesma forma que os corpúsculos de Meissner, detecta principalmente os movimentos de objectos sobre a superfície corporal ou o contacto inicial com o corpo;
Localizados nas camadas mais profundas da pele e também nos tecidos mais profundos estão os órgãos terminais de Ruffini, que são terminações encapsuladas e com muitas ramificações. Essas terminações adaptam-se muito pouco e, por isso, são importantes para a detecção de estados de deformação continuada da pele e dos tecidos mais profundos, tais como os sinais de toque e de pressão mais fortes e contínuos. Estão localizados nas cápsulas articulares e ajudam a sinalizar o grau de rotação da articulação;
Corpúsculos de Pacini que se situam imediatamente abaixo da pele e também em regiões mais profundas, nas faseias teciduais do corpo. Só são estimulados por movimentos muito rápidos dos tecidos, pois adaptam-se em centésimos de segundo. Por isso, são particularmente importantes para a detecção da vibração dos tecidos ou de outras modificações extremamente rápidas do estado mecânico tecidual.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
55 | P á g i n a
Paladar
As identidades de determinadas substâncias químicas que estimulam os diferentes receptores gustativos ainda não são conhecidas. No entanto, estudos psicofisiológicos e neurofisiológicos identificaram pelo menos 13 possíveis (ou prováveis) receptores químicos nas células gustativas, como se segue:
2 receptores para o sódio; 2 receptores para o potássio;
1 receptor para cloreto; 1 receptor para adenosina; 1 receptor para inosina; 2 receptores para o sabor doce; 2 receptores para o sabor amargo; 1 receptor para o glutamato; 1 receptor para o ião hidrogénio. Sensibilidade a diferentes tipos de sabores: doce, amargo, salgado e ácido;
Papilas gustativas contêm terminais de neurónios sensitivos;
Ocorre activação de diferentes quimioreceptores para os diferentes tipos de sabores.
Olfacto
Ocorre, resumidamente:
A activação da proteína receptora, pela
substância odorante, activa o complexo de
proteína G;
Segue-se activação de múltiplas moléculas de
adenilil ciclase, no interior da membrana
olfactiva; que por sua vez causam,

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
56 | P á g i n a
Formação de muitas mais moléculas de AMPc;
AMPc abre ainda mais os canais de sódio, permitindo a entrada de grandes
quantidades;
Os iões sódio aumentam o potencial eléctrico na direcção positiva no interior da
membrana celular, excitando o neurónio olfactivo e transmitindo potenciais de acção
para o SNC através de um nervo olfactivo.
Intensidade do estímulo
Força do estímulo;
Capacidade de activar o neurónio sensitivo e gerar potenciais de acção;
Codificada pela frequência com que se geram potenciais de acção do neurónio;
Estímulo:
- Baixa intensidade – alteração do potencial de membrana (despolarização) mas que não é
suficiente para atingir o limiar de disparo – não é transmitido ao córtex;
- Alta intensidade – suficiente para atingir o limiar de disparo desencadear potenciais de
acção.
Adaptação dos
Receptores

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
57 | P á g i n a
Localização do estímulo
A localização do estímulo é codificada pelo local onde se encontra o receptor estimulado, e pelo facto da informação de cada receptor viajar por uma via única até uma região especifica do SNC associada a uma modalidade e localização particular.
O tamanho do campo receptivo influencia a capacidade de localização.
A densidade de receptores (A) e o tamanho do campo receptor de cada unidade sensitiva (B) no local onde é aplicado o estímulo são factores que afectam a precisão ou acuidade sensitiva.
Acuidade sensitiva
A precisão ou acuidade com que é possível localizar e diferenciar 2 estímulos adjacentes depende do grau de convergência dos neurónios sensitivos nas vias neuronais ascendentes que conduzem a informação ao SNC. Capacidade de localizar
estímulos.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
58 | P á g i n a
SOBREPOSIÇÃO DE CAMPOS RECEPTORES
E estimulação de campos receptores vizinhos que se sobrepõem permite fornecer informação geral sobre o estímulo.
O neurónio B responde mais intensamente ao estímulo do que os neurónios A e C porque o estímulo é aplicado no centro do seu campo receptor onde a densidade de receptores é maior.
Inibição Lateral A inibição lateral acentua o contraste entre estímulos adjacentes e o padrão de condução dos potenciais de acção até ao SNC, permitindo uma maior resolução na localização do estímulo.
Área central é mais activada, por
isso mais estimulada mas as
laterais também condicionam,
não podendo, contudo, conduzir
informação.
O SNC pode seleccionar certo tipo de informação sensorial, inibindo neurónios das vias aferentes.
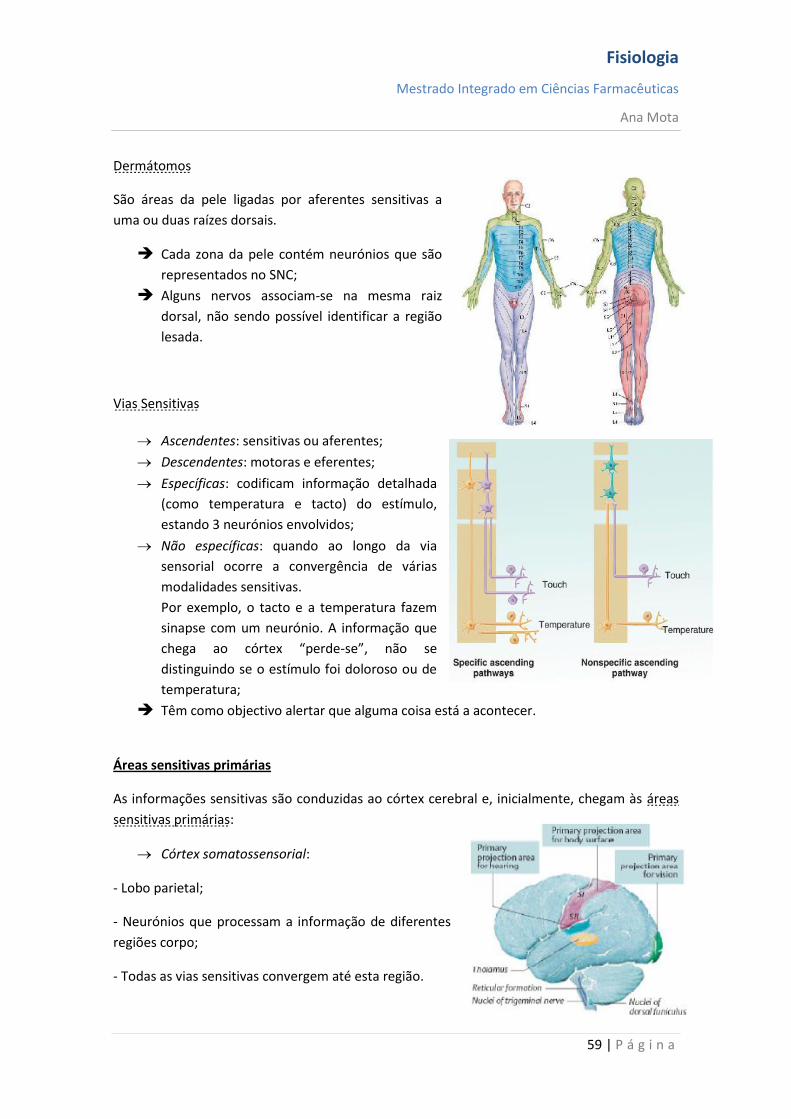
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
59 | P á g i n a
Dermátomos
São áreas da pele ligadas por aferentes sensitivas a
uma ou duas raízes dorsais.
Cada zona da pele contém neurónios que são
representados no SNC;
Alguns nervos associam-se na mesma raiz
dorsal, não sendo possível identificar a região
lesada.
Vias Sensitivas
Ascendentes: sensitivas ou aferentes;
Descendentes: motoras e eferentes;
Específicas: codificam informação detalhada
(como temperatura e tacto) do estímulo,
estando 3 neurónios envolvidos;
Não específicas: quando ao longo da via
sensorial ocorre a convergência de várias
modalidades sensitivas.
Por exemplo, o tacto e a temperatura fazem
sinapse com um neurónio. A informação que
chega ao córtex “perde-se”, não se
distinguindo se o estímulo foi doloroso ou de
temperatura;
Têm como objectivo alertar que alguma coisa está a acontecer.
Áreas sensitivas primárias
As informações sensitivas são conduzidas ao córtex cerebral e, inicialmente, chegam às áreas
sensitivas primárias:
Córtex somatossensorial:
- Lobo parietal;
- Neurónios que processam a informação de diferentes
regiões corpo;
- Todas as vias sensitivas convergem até esta região.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
60 | P á g i n a
Córtex auditivo:
- Lobo temporal;
- Recebe a informação auditiva.
Córtex visual:
- Lobo occipital;
- Recebe a informação visual.
Junto ao córtex somatossensorial:
- Sabores.
Nocicepção
Recepção de sinais no SNC induzidos pela activação de receptores sensitivos especializados que fornecem informação sobre a ocorrência de lesão nos tecidos. Esta informação é codificada por NOCICEPTORES.
Nem todos os estímulos nóxicos originam DOR.
Dor
Resposta subjectiva à informação nociceptiva;
Consiste na percepção de uma sensação desagradável com origem numa região particular do corpo que pode estar associada a uma lesão dos tecidos;
A experiência da dor é modificada por vários factores cognitivos e emocionais: ansiedade, antecipação, experiência passada e influências socioculturais.
A dor é uma resposta subjectiva à informação nociceptiva, não correspondendo a
nenhuma forma de energia.
Nociceptores
Morfologia: Terminais livres de neurónios (dendrites);
Estímulo/Receptor: Mecânicos, Térmicos, Químicos; Limiar mais elevado que outros receptores; Necessários estímulos mais intensos para activar estes receptores – percepcionar a dor Características:
Nociceptores Térmicos e Mecânicos

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
61 | P á g i n a
- Fibras Aδ (5-30 m/s); - Sensação aguda de dor (rápida); - Fibras mielinizadas.
Nociceptores Polimodais - Fibras C (0.5-2 m/s); - Estímulos de várias formas de energia: receptor químico, térmico ou mecânico; - Fibras não mielinizadas; - Dor incomodativa, insustentável.
Os neurónios Aδ e C conduzem informação até à medula espinal (raízes dorsais). Ocorre sinapse com o 2º neurónio da cadeia sensitiva ou então ocorre a sinapse com o interneurónio (excitatório ou inibitório) antes da sinapse com o 2º neurónio.
Substância P (SP)
A sua libertação nas terminações nervosas periféricas produz vasodilatação e aumento da
permeabilidade vascular, estimula a quimiotaxia de leucócitos e a activação local de
neutrófilos e macrófagos, aumenta a produção e a libertação de mediadores inflamatórios,
assim como a desgranulação de mastócitos e a consequente libertação local de histamina,
verificando-se ainda o aumento da libertação de prostaglandina E2, e outros eucosanóides,
que contribuem para a estimulação/sensibilização dos nociceptores.
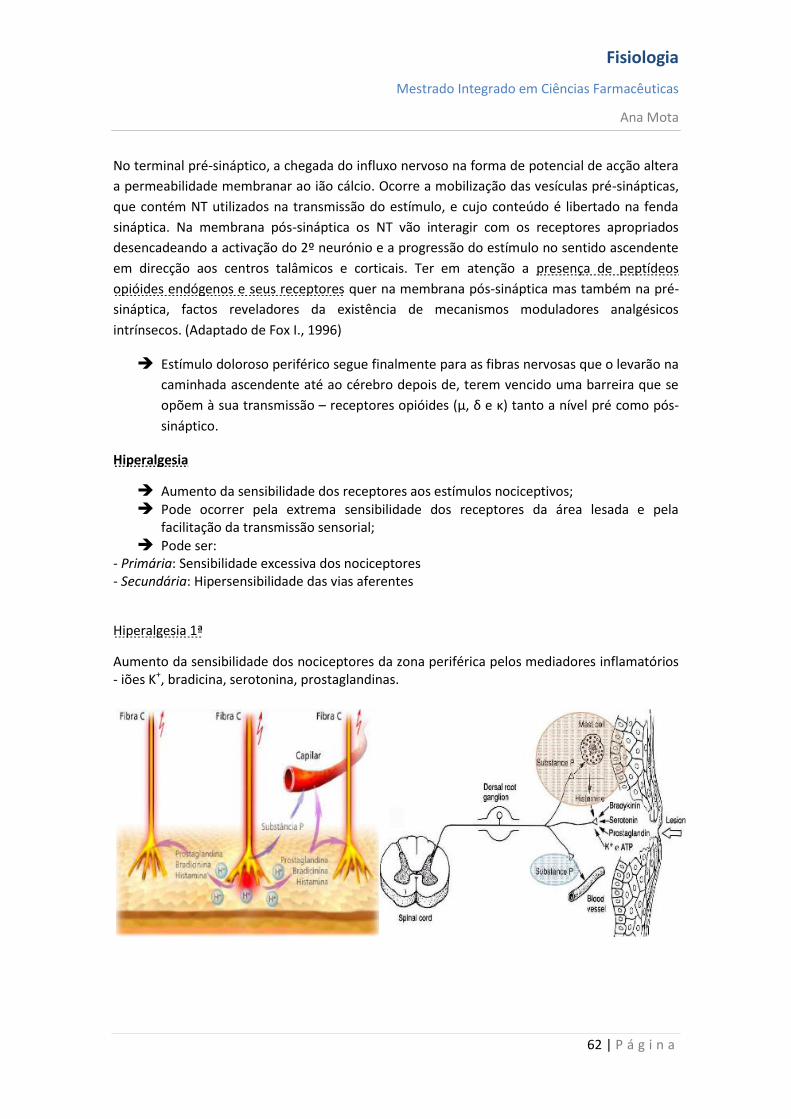
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
62 | P á g i n a
No terminal pré-sináptico, a chegada do influxo nervoso na forma de potencial de acção altera
a permeabilidade membranar ao ião cálcio. Ocorre a mobilização das vesículas pré-sinápticas,
que contém NT utilizados na transmissão do estímulo, e cujo conteúdo é libertado na fenda
sináptica. Na membrana pós-sináptica os NT vão interagir com os receptores apropriados
desencadeando a activação do 2º neurónio e a progressão do estímulo no sentido ascendente
em direcção aos centros talâmicos e corticais. Ter em atenção a presença de peptídeos
opióides endógenos e seus receptores quer na membrana pós-sináptica mas também na pré-
sináptica, factos reveladores da existência de mecanismos moduladores analgésicos
intrínsecos. (Adaptado de Fox I., 1996)
Estímulo doloroso periférico segue finalmente para as fibras nervosas que o levarão na
caminhada ascendente até ao cérebro depois de, terem vencido uma barreira que se
opõem à sua transmissão – receptores opióides (μ, δ e κ) tanto a nível pré como pós-
sináptico.
Hiperalgesia
Aumento da sensibilidade dos receptores aos estímulos nociceptivos; Pode ocorrer pela extrema sensibilidade dos receptores da área lesada e pela
facilitação da transmissão sensorial; Pode ser:
- Primária: Sensibilidade excessiva dos nociceptores - Secundária: Hipersensibilidade das vias aferentes
Hiperalgesia 1ª
Aumento da sensibilidade dos nociceptores da zona periférica pelos mediadores inflamatórios - iões K+, bradicina, serotonina, prostaglandinas.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
63 | P á g i n a
Hiperalgesia 2ª
- Aumenta a frequência dos disparos neuronais. - Activa sinapses salientes o que promove a activação da área espinotalâmica. - Tipo de hiperalgesia que pode levar à dor crónica.
Sensibilização Aferente - Glutamato (AMPA/NMDA/ mGLUR) - Substância P (NK1R) - CGRP, BDNF, ATP (P2X3)
Nota:
GLUTAMATO (receptores AMPA e NMDA)
- Actua instantaneamente e persiste por milissegundos;
- Sensação de dor rápida .
SUBSTÂNCIA P (receptores NK1)
- Liberação lenta e aumento da concentração num período de segundos ou até minutos;
- Sensação de dor lenta
Percepção/Modulação da Dor
A estimulação directa de certas regiões do cérebro causa supressão da dor; Portão da dor – Modulação da DOR; Localização de centros cerebrais sensíveis a morfina; A descoberta de péptidos endógenos com acção analgésica (β-endorfina e encefalina); A caracterização dos receptores dos opióides; Identificação das vias descendentes envolvidas no controlo da nocicepção – Sistema
Anti-Nociceptivo.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
64 | P á g i n a
Analgesia
- Abolição da sensibilidade à dor sem supressão de outras propriedades sensitivas, nem perda de consciência. - Resulta da supressão da entrada de sinais de dor para o sistema nervoso, pelo próprio cérebro. - O sistema de analgesia é formado por três componentes principais: Substância cinzenta periaquedutal e
as áreas periventriculares do mesencéfalo;
Núcleo magno da rafe (formação reticular do bulbo);
Complexo inibitório da dor (localizado nos cornos dorsais da medula espinhal).
Os neurónios da substância cinzenta periaquedutal, das áreas periventriculares do mesencéfalo e da porção superior da ponte em redor do aqueduto de Sylvius enviam sinais para o núcleo magno da rafe e para o núcleo reticular paragigantocelular. Destes núcleos a informação parte para os cornos dorsais da medula espinal, onde se situa o complexo inibitório da dor. Através da libertação de NT a via da dor é bloqueada. Os NT assosiados ao sistema analgésico são: - Encefalinas e serotoninas. - A encefalina causa inibição pré e pós-sináptica das fibras da dor C e Aδ. Pensa-se que tal ocorre pelo bloqueio dos canais de Ca2+, que impedem a libertação dos NT da dor; - As serotoninas são secretadas pelas fibras que provêm do núcleo magno da rafe nos cornos dorsais da medula. Sendo responsáveis pelo estímulo da secreção de encefalinas pelos neurónios locais da espinal medula.
Nota: Apreciação da Dor
Córtex Somatossensorial – fibras Aδ
Córtex Pré-frontral – fibras Aδ
Núcleo Periventricular do Hipotálamo – fibras C - Subst. Cinzenta Periaquedutal do Mesencéfalo (PAG) Estimulação desta área causa analgesia Estimulação inibe a dor cirúrgica
- Núcleo Rafe Magnus (Formação Reticular do Bolbo).
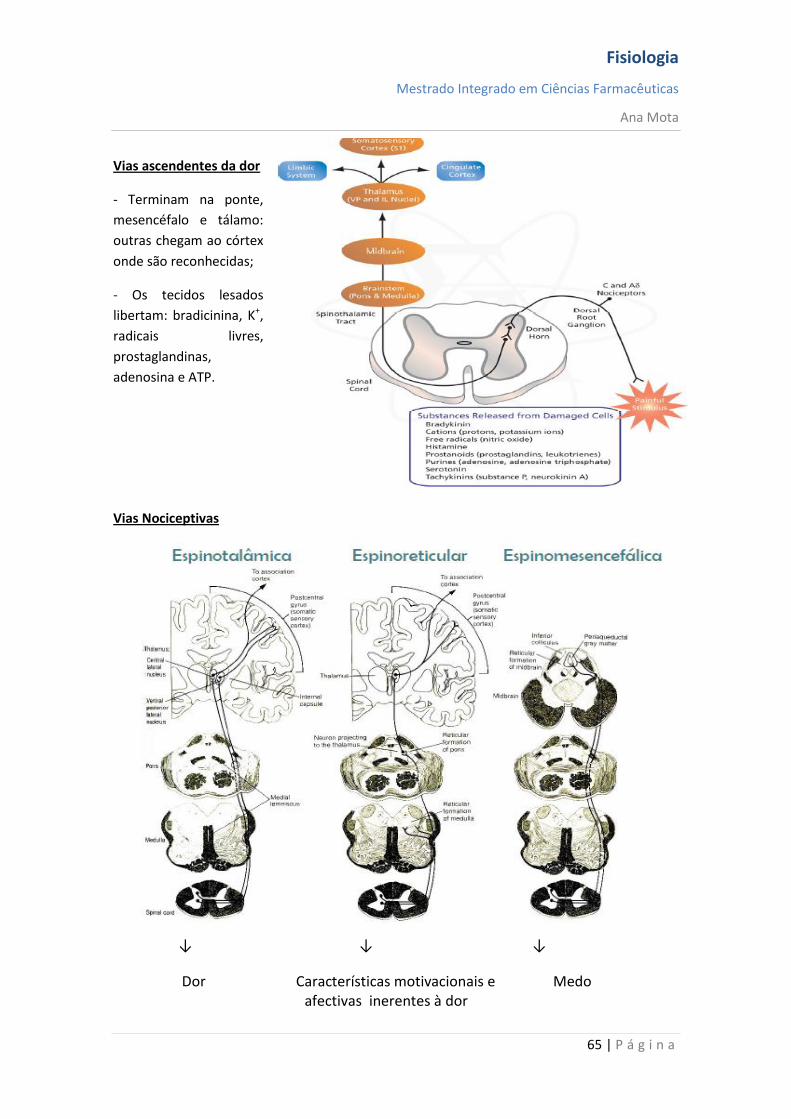
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
65 | P á g i n a
Vias ascendentes da dor
- Terminam na ponte,
mesencéfalo e tálamo:
outras chegam ao córtex
onde são reconhecidas;
- Os tecidos lesados
libertam: bradicinina, K+,
radicais livres,
prostaglandinas,
adenosina e ATP.
Vias Nociceptivas
↓ ↓ ↓
Dor Características motivacionais e Medo afectivas inerentes à dor

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
66 | P á g i n a
Dor referida
- Envolve sinais de dor que se originam num órgão ou tecido interno (visceral);
- Muitas vezes, uma pessoa sente dor em uma parte do corpo consideravelmente afastada dos
tecidos que estão realmente provocando dor. Comummente, a dor inicia-se em um dos órgãos
viscerais e é referida à área sobre a superfície corporal. Ocasionalmente, a dor pode estar
referida a outra área corporal profunda que, no entanto, não coincide exactamente à víscera
que a está a provocar. O conhecimento desses diferentes tipos de dor referida é
extremamente importante no diagnóstico clínico, pois muitas doenças viscerais não causam
outros sinais clínicos excepto a dor referida;
- Mecanismo mais provável para o surgimento da dor referida - ramificações das fibras viscerais de dor fazem junções sinápticas na medula espinhal, com neurónios de segunda ordem, que também recebem sinais de fibras de dor provenientes da pele. Dessa maneira, quando uma fibra de dor localizada na víscera é estimulada, o sinal é conduzido para o SNC pelos mesmos neurónios que conduzem sinais dolorosos provenientes da pele, o que faz com que a pessoa tenha a sensação de que os sinais tenham, na realidade, se originado na própria pele, em lugar da víscera; - Um exemplo comum é o do enfarte agudo do miocárdio, onde a dor é referida à superfície esquerda da mandíbula ou do pescoço, ou também ao braço esquerdo. Em vez de associar esta dor ao coração, o paciente percebe a sensação como proveniente da face ou do braço. O que significa que sinais aferentes viscerais, a partir do coração, convergem sobre os mesmos neurónios da medula espinhal que recebem entradas cutâneas da periferia.
Portão da Dor - Sequência dos neurónios na transmissão
sensitiva que passa pelas fibras Aδ e pelas
fibras C, que conduzem a informação periférica
até à ME. Aqui fazem sinapse com
interneurónios excitatórios e inibitórios que
vão conduzir ou não a informação;
- Neste local passam também neurónios que
conduzem outros tipos de informação (outras
modalidades sensitivas: tacto, pressão, etc). Quando se estimulam simultaneamente as
modalidades pressão, tacto e dor ocorre um bloqueio da transmissão dolorosa e atenuação do
tacto/pressão;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
67 | P á g i n a
- Neurónio sensitivo (fibra) → corno posterior da medula → sinapse com interneurónios
inibitórios → bloqueiam transmissão da dor → estabelecimento de um equilíbrio entre as vias
dolorosas/não dolorosas;
A substância que está envolvida no bloqueio da transmissão noceptora é a encefalina
(neuroreceptor opiáceo).
Em suma: - As fibras A-α e A-β libertam uma grande descarga eléctrica que activa os interneurónios, na Substância Gelatinosa; - Os interneurónios libertam encefalina que inibe a libertação da substância P e o bloqueio das vias da dor; - As fibras C mais finas inactivam os interneurónios e estimulam os neurónios de projecção da dor.
Percepção da Informação Sensorial
Adaptação de receptores; Emoções, a personalidade e a
experiência prévia; Informação é cancelada; Ausência de receptores para
detectar certas formas de energia - raios X, UV, IV;
Neurónios lesionados podem transmitir informação errada – “sintoma do membro fantasma”;
Drogas - ilusões e alucinações
Medula Espinal
É responsável por: - Transmissão de informação (Encéfalo ↔ Sistema Nervoso Periférico); - Integrar a informação que recebe; - Produzir respostas através de mecanismos reflexos (actos involuntários) Num corte transversal: Matéria cinzenta – Situa-se numa
posição central. Constituída por corpos celulares, terminais de axónios, interneurónios e células gliais.
Matéria branca – Situa-se na periferia. É constituída por axónios mielinizados, alguns deles agrupados em tractos nervosos.
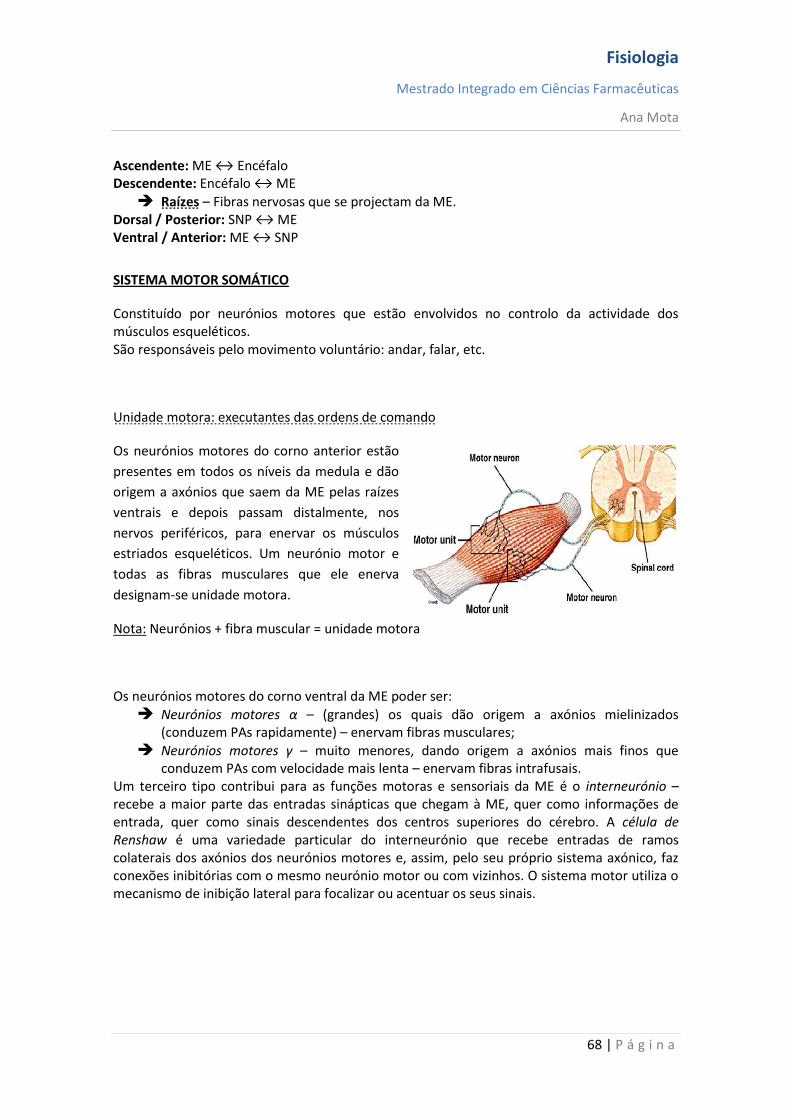
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
68 | P á g i n a
Ascendente: ME ↔ Encéfalo Descendente: Encéfalo ↔ ME
Raízes – Fibras nervosas que se projectam da ME. Dorsal / Posterior: SNP ↔ ME Ventral / Anterior: ME ↔ SNP
SISTEMA MOTOR SOMÁTICO
Constituído por neurónios motores que estão envolvidos no controlo da actividade dos músculos esqueléticos. São responsáveis pelo movimento voluntário: andar, falar, etc.
Unidade motora: executantes das ordens de comando
Os neurónios motores do corno anterior estão
presentes em todos os níveis da medula e dão
origem a axónios que saem da ME pelas raízes
ventrais e depois passam distalmente, nos
nervos periféricos, para enervar os músculos
estriados esqueléticos. Um neurónio motor e
todas as fibras musculares que ele enerva
designam-se unidade motora.
Nota: Neurónios + fibra muscular = unidade motora
Os neurónios motores do corno ventral da ME poder ser:
Neurónios motores α – (grandes) os quais dão origem a axónios mielinizados (conduzem PAs rapidamente) – enervam fibras musculares;
Neurónios motores γ – muito menores, dando origem a axónios mais finos que conduzem PAs com velocidade mais lenta – enervam fibras intrafusais.
Um terceiro tipo contribui para as funções motoras e sensoriais da ME é o interneurónio – recebe a maior parte das entradas sinápticas que chegam à ME, quer como informações de entrada, quer como sinais descendentes dos centros superiores do cérebro. A célula de Renshaw é uma variedade particular do interneurónio que recebe entradas de ramos colaterais dos axónios dos neurónios motores e, assim, pelo seu próprio sistema axónico, faz conexões inibitórias com o mesmo neurónio motor ou com vizinhos. O sistema motor utiliza o mecanismo de inibição lateral para focalizar ou acentuar os seus sinais.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
69 | P á g i n a
Exemplos de categorias de informação que modificam a produção dos comandos motores provenientes do cérebro
Reflexo na ME
Reflexo é uma resposta automática produzida pelo arco reflexo a um estímulo sensorial que ocorre sem pensamento consciente.
Reflexo espinal é qualquer acto reflexo mediado através da medula espinal.
Nota:
A via que medeia um determinado reflexo constitui o seu arco reflexo. O arco reflexo é constituído por um órgão sensitivo, um neurónio aferente, uma ou mais sinapses numa estação de integração, um neurónio eferente e um efector. Nos mamíferos, a conexão entre os neurónios aferentes e eferentes somáticos ocorre geralmente no sistema nervoso central (SNC).
Os neurónios aferentes atingem o SNC através das raízes dorsais ou dos nervos cranianos e têm os seus corpos celulares nos gânglios das raízes dorsais ou nos gânglios dos nervos cranianos. Os neurónios eferentes saem do SNC através das raízes ventrais ou dos nervos cranianos motores correspondentes. A actividade no arco reflexo tem início num receptor sensitivo com um potencial receptor (um dos tipos de potencial gradativo), cuja amplitude é proporcional à intensidade do estímulo. Se o estímulo for suficientemente intenso, gera-se em seguida um potencial de acção no nervo aferente. Os potenciais de acção dos nervos aferentes originam, no SNC, potenciais sinápticos inibitórios ou excitatórios (IPSP e EPSP, respectivamente). No nervo eferente, por sua vez, são gerados novamente potenciais de acção. Quando estes atingem o efector, originam uma resposta (e.g. contracção muscular; secreção glandular). A actividade do reflexo pode também ser modificada através de múltiplas aferências que convergem nos neurónios eferentes. Os músculos esqueléticos possuem uma grande variedade de receptores. Dois deles
são particularmente importantes para o controlo motor: os Fusos Musculares e os Órgãos Tendinosos de Golgi. Os Fusos Musculares são inervados por fibras aferentes do grupo I (fibras mielinizadas de grande diâmetro) e do grupo II (fibras mielinizadas pequenas). Os Órgãos Tendinosos de Golgi são inervados apenas por fibras aferentes
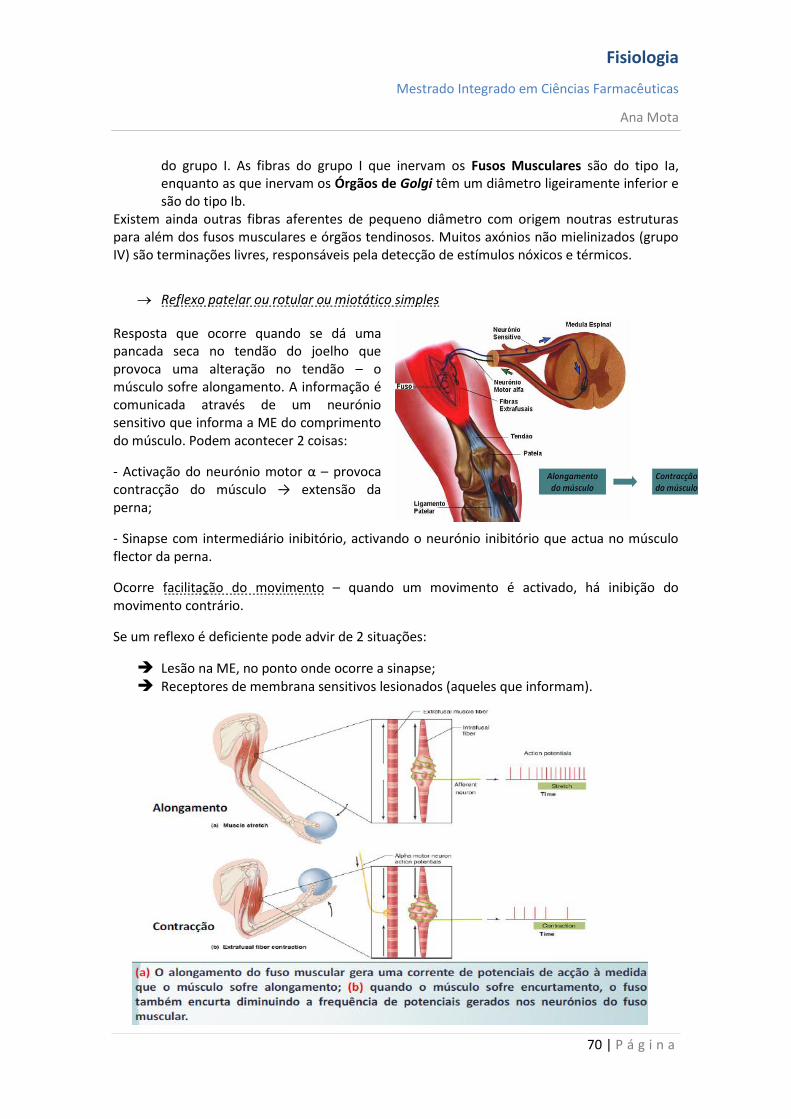
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
70 | P á g i n a
do grupo I. As fibras do grupo I que inervam os Fusos Musculares são do tipo Ia, enquanto as que inervam os Órgãos de Golgi têm um diâmetro ligeiramente inferior e são do tipo Ib.
Existem ainda outras fibras aferentes de pequeno diâmetro com origem noutras estruturas para além dos fusos musculares e órgãos tendinosos. Muitos axónios não mielinizados (grupo IV) são terminações livres, responsáveis pela detecção de estímulos nóxicos e térmicos.
Reflexo patelar ou rotular ou miotático simples
Resposta que ocorre quando se dá uma pancada seca no tendão do joelho que provoca uma alteração no tendão – o músculo sofre alongamento. A informação é comunicada através de um neurónio sensitivo que informa a ME do comprimento do músculo. Podem acontecer 2 coisas:
- Activação do neurónio motor α – provoca contracção do músculo → extensão da perna;
- Sinapse com intermediário inibitório, activando o neurónio inibitório que actua no músculo flector da perna.
Ocorre facilitação do movimento – quando um movimento é activado, há inibição do movimento contrário.
Se um reflexo é deficiente pode advir de 2 situações:
Lesão na ME, no ponto onde ocorre a sinapse; Receptores de membrana sensitivos lesionados (aqueles que informam).

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
71 | P á g i n a
FUSO MUSCULAR A densidade de Fusos Musculares é proporcional à complexidade de movimentos executados pelo músculo. São estruturas encapsuladas e contêm três componentes principais: 1. Um grupo de fibras musculares intrafusais especializadas; 2. Axónios sensitivos que terminam nas fibras musculares; 3. Axónios motores que regulam a sensibilidade do fuso. O centro do fuso está envolvido por uma cápsula de tecido conjuntivo e contém um fluido gelatinoso que facilita o deslizamento das fibras musculares entre si. As fibras musculares especializadas do fuso são as Fibras Intrafusais, por oposição às fibras musculares normais Extrafusais. As Fibras Intrafusais são mais pequenas e não contribuem de forma significativa para a contracção muscular. As suas regiões centrais contêm poucas miofibrilas e são essencialmente não contrácteis; apenas as regiões polares contraem activamente. Existem, por sua vez, dois tipos de Fibras Musculares Intrafusais: 1. Fibras de Cadeia Nuclear; 2. Fibras de Saco Nuclear. Um Fuso Muscular típico contém duas Fibras de Saco Nuclear e um número variável de Fibras de Cadeia Nuclear, normalmente cinco. Os axónios mielinizados que atravessam a cápsula terminam na região central das Fibras Intrafusais. A maioria das terminações aferentes enrola-se à volta destas fibras. Quando as fibras intrafusais são estiradas, as terminações nervosas aumentam a frequência de despolarização. Isto acontece porque o estiramento do fuso também estira a parte central das Fibras Intrafusais, à volta das quais estão enroladas as terminações aferentes. O consequente estiramento das terminações aferentes activa canais sensíveis ao estiramento que despolarizam a membrana e dão origem a potenciais de acção. Quando termina o estiramento, as fibras intrafusais relaxam e a frequência de despolarização diminui. Existem dois tipos de fibras aferentes nos fusos musculares: as Primárias e as Secundárias. Normalmente, existe apenas uma Terminação Primária em cada fuso, constituída por todas as ramificações de um neurónio aferente do grupo Ia. Os aferentes do grupo Ia têm, por sua vez, origem nos dois tipos de fibras intrafusais. As Terminações Secundárias são formadas por

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
72 | P á g i n a
fibras do grupo II que têm origem sobretudo nas fibras de cadeia nuclear. As Terminações Primárias e Secundárias têm diferentes padrões de actividade. Inervação Recíproca A compressão do tendão da patela activa receptores de alongamento do músculo da extensão da perna junto à articulação. AS RESPOSTAS INCLUEM: (A, C) Contracção compensatória no músculo que sofreu alongamento; (B) Relaxamento do músculo flector que se opõe ao movimento; (D) Condução de informação por neurónios aferentes até ao cérebro.
Órgão do Tendão de Golgi
O órgão do tendão de Golgi é um receptor sensorial encapsulado atravessado por pequeno feixe de fibras tendinosas musculares situado imediatamente acima do ponto em que elas se fundem às fibras musculares. Cada órgão do tendão tem, em média, 10 a 15 fibras musculares conectadas em série com este e é estimulado pela tensão produzida por esse pequeno feixe de fibras musculares. Assim, a principal diferença entre o órgão do tendão de Golgi e o fuso muscular é que o fuso muscular detecta o comprimento do músculo e as alterações desse comprimento, enquanto o órgão do tendão detecta a tensão muscular. O órgão do tendão, à semelhança do receptor primário do fuso muscular, tem tanto a resposta dinâmica como a resposta estática. A resposta dinâmica
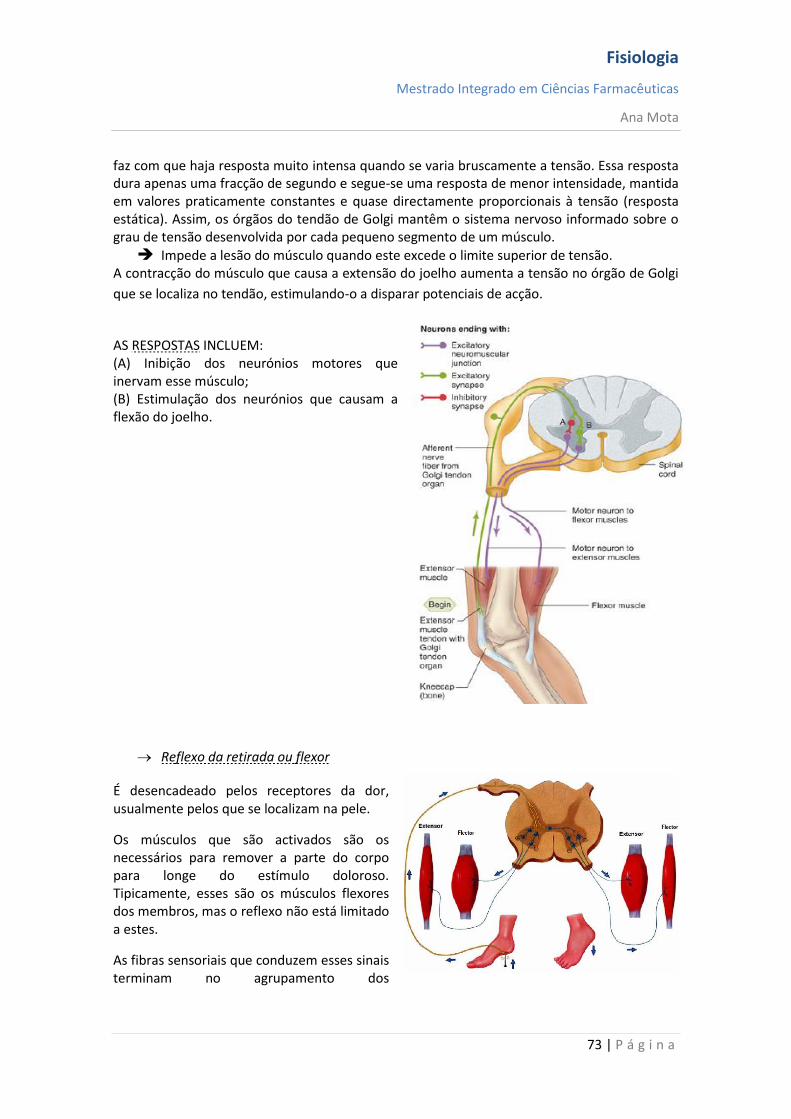
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
73 | P á g i n a
faz com que haja resposta muito intensa quando se varia bruscamente a tensão. Essa resposta dura apenas uma fracção de segundo e segue-se uma resposta de menor intensidade, mantida em valores praticamente constantes e quase directamente proporcionais à tensão (resposta estática). Assim, os órgãos do tendão de Golgi mantêm o sistema nervoso informado sobre o grau de tensão desenvolvida por cada pequeno segmento de um músculo.
Impede a lesão do músculo quando este excede o limite superior de tensão. A contracção do músculo que causa a extensão do joelho aumenta a tensão no órgão de Golgi
que se localiza no tendão, estimulando-o a disparar potenciais de acção.
AS RESPOSTAS INCLUEM: (A) Inibição dos neurónios motores que inervam esse músculo; (B) Estimulação dos neurónios que causam a flexão do joelho.
Reflexo da retirada ou flexor
É desencadeado pelos receptores da dor, usualmente pelos que se localizam na pele.
Os músculos que são activados são os necessários para remover a parte do corpo para longe do estímulo doloroso. Tipicamente, esses são os músculos flexores dos membros, mas o reflexo não está limitado a estes.
As fibras sensoriais que conduzem esses sinais terminam no agrupamento dos

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
74 | P á g i n a
interneurónios da ME, a maioria dos quais fornece entradas excitatórias aos neurónios motores apropriados do corno ventral, enquanto outros inibem neurónios motores que enervam os músculos antagonistas – inibição recíproca.
Reflexo Cutâneo-Plantar – é um Reflexo dos segmentos medulares S1-S2. Pesquisa-se raspando ligeiramente a planta do pé sendo a resposta normal a Flexão Plantar dos dedos do pé. Se a resposta for em extensão – Sinal de Babinsky - indica sempre lesão da Via Piramidal, excepto nas crianças até aos dois anos de idade nas quais é normal.
Programas Motores Redes neuronais que existem entre as principais regiões motoras do córtex cerebral permitem um
controlo preciso do movimento, utilizando informação sensorial e sinais de “intenção” para activar os neurónios motores apropriados e adequar o seu nível de activação.
Mesencéfalo
Vesícula intermédia originária do tubo neural aquando da formação do cérebro
durante o desenvolvimento embrionário;
É anteriormente adjacente à protuberância e posteriormente adjacente ao diencéfalo;
Funções:
- Visão;
- Audição;
- Movimentos do corpo e oculares.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
75 | P á g i n a
Núcleos da Base
- São um aglomerado de
áreas interligadas, que se
localizam profundamente
nos hemisférios cerebrais,
logo abaixo do córtex;
- Papel crucial na iniciação
dos movimentos;
- Podem funcionar como
um filtro complexo,
seleccionando
informação, entre a grande diversidade de sinais que lhes chegam, provenientes da metade
anterior do córtex (regiões sensoriais, motoras, pré-frontais e límbicas). As respostas
provenientes destes são enviadas de volta a áreas do córtex motor;
- Lesões nesta área podem conduzir a patologias como Doença de Parkinson e Doença de Huntington: Lesões nos neurónios do corpo estriado → Coreia de Huntington.
- Incapacidade de coordenar movimentos, apresentando movimentos violentos e involuntários; - Lesões cognitivas surgem tardiamente. Lesões na substância nigra → Doença de Parkinson.
- Pode ser causada pela perda de fibras nervosas secretoras de dopamina - degeneração de neurónios dopaminérgicos; - Caracteriza-se por: Perda progressiva da função motora (50-60 anos); Rigidez em muitos grupos musculares; Tremor presente em repouso, quando nenhum movimento voluntário está a ser
executado; Escrita ilegível; Discurso lento; Movimentos lentos (Bradicinesia); Dificuldade em iniciar um movimento (acinesia).
- Os neurónios da substância nigra libertam dopamina para o corpo estriado. Caso os neurónios dopaminégicos sejam inibidos, deixa de haver um controlo do neurónio estriado; - Tratamentos com fármacos, cirurgia e fisioterapia mas, infelizmente, sem cura.
Cerebelo
1. Coordenação do movimento e postura 2. Elaboração de programas motores 3. Aprendizagem de comportamentos motores 4. Ajuste de programas durante a execução

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
76 | P á g i n a
5. Manutenção do equilíbrio
- É formado por 2 hemisférios cerebelosos e uma parte central, designada verme;
- Localizado ao lado do tronco cerebral;
- Representa 10% do volume total do cérebro mas contém quase metade do nº de neurónios do cérebro;
- As lesões hemisféricas manifestam-se no lado afectado.
Exemplos de lesões no cerebelo: Discurso arrastado; Tremor de intenção;
Incapacidade de travar o movimento; Ataxia: velocidade, força e direcção.
Tronco Cerebral
• Reflexos de posicionamento erecto; • Postura e equilíbrio do corpo; • Tónus muscular; • Coordenação do movimento dos olhos.
- Constituído por: MESENCÉFALO
PONTE BOLBO RAQUIDIANO
Rigidez de “descorticação” Reflexo de postura estática; Descargas a e g exageradas; Reflexos de estiramento facilitados;
Contracção exagerada dos músculos extensores
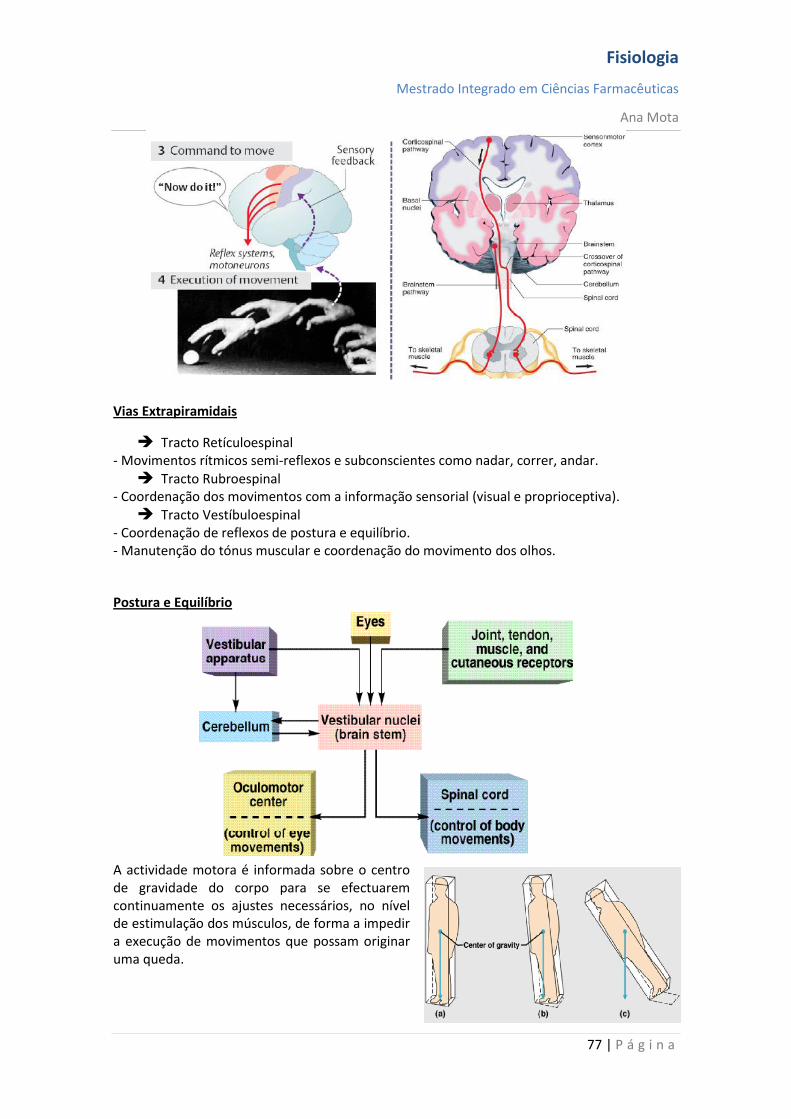
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
77 | P á g i n a
Vias Extrapiramidais
Tracto Retículoespinal - Movimentos rítmicos semi-reflexos e subconscientes como nadar, correr, andar. Tracto Rubroespinal
- Coordenação dos movimentos com a informação sensorial (visual e proprioceptiva). Tracto Vestíbuloespinal
- Coordenação de reflexos de postura e equilíbrio. - Manutenção do tónus muscular e coordenação do movimento dos olhos.
Postura e Equilíbrio
A actividade motora é informada sobre o centro de gravidade do corpo para se efectuarem continuamente os ajustes necessários, no nível de estimulação dos músculos, de forma a impedir a execução de movimentos que possam originar uma queda.
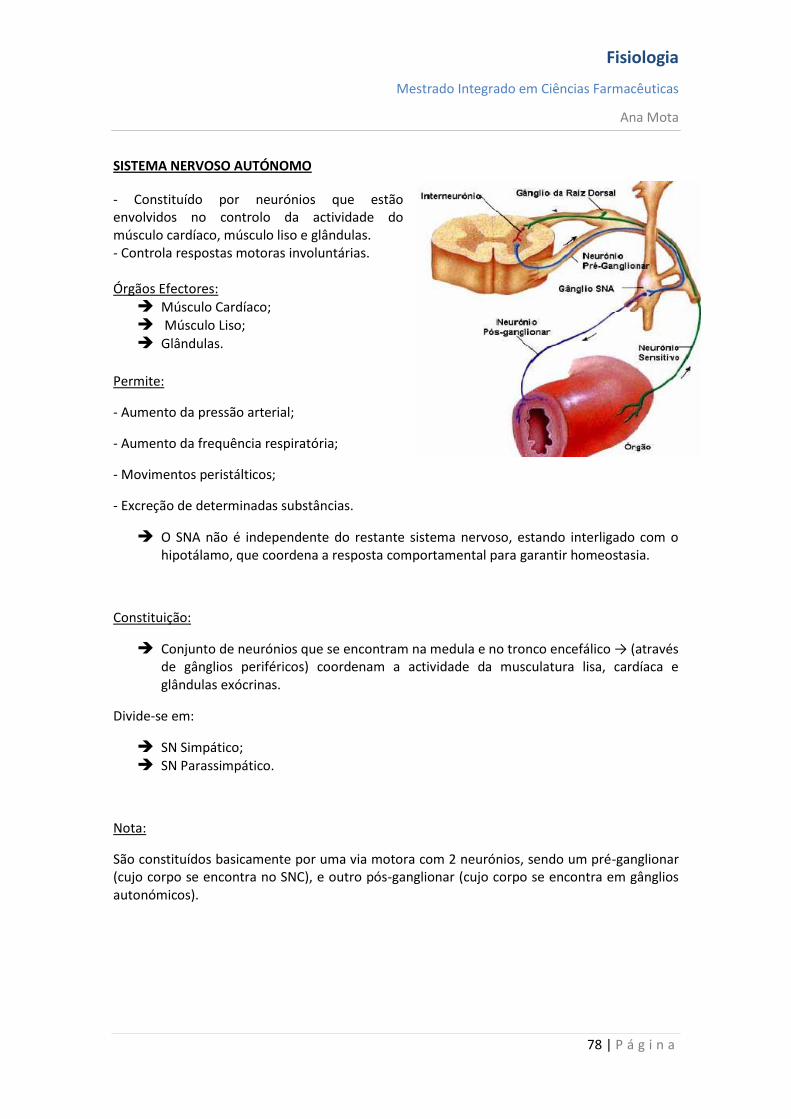
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
78 | P á g i n a
SISTEMA NERVOSO AUTÓNOMO
- Constituído por neurónios que estão envolvidos no controlo da actividade do músculo cardíaco, músculo liso e glândulas. - Controla respostas motoras involuntárias. Órgãos Efectores: Músculo Cardíaco; Músculo Liso; Glândulas.
Permite:
- Aumento da pressão arterial;
- Aumento da frequência respiratória;
- Movimentos peristálticos;
- Excreção de determinadas substâncias.
O SNA não é independente do restante sistema nervoso, estando interligado com o hipotálamo, que coordena a resposta comportamental para garantir homeostasia.
Constituição:
Conjunto de neurónios que se encontram na medula e no tronco encefálico → (através de gânglios periféricos) coordenam a actividade da musculatura lisa, cardíaca e glândulas exócrinas.
Divide-se em:
SN Simpático; SN Parassimpático.
Nota:
São constituídos basicamente por uma via motora com 2 neurónios, sendo um pré-ganglionar (cujo corpo se encontra no SNC), e outro pós-ganglionar (cujo corpo se encontra em gânglios autonómicos).

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
79 | P á g i n a
Neurotransmissão
Os nervos simpáticos diferem dos nervos motores esqueléticos pelo seguinte: cada via simpática que emerge da medula até o tecido estimulado é formada por dois neurónios, um pré-ganglionar e outro pós-ganglionar, o que contrasta com o único neurónio da via motora esquelética. O corpo celular de cada neurónio pré-ganglionar fica situado na ponta intermediolateral da ME e sua fibra passa pela raiz ventral da medula para o nervo espinhal correspondente. Imediatamente após o nervo espinhal ter saído da coluna medular, as fibras simpáticas pré-ganglionares abandonam-na, passando pelo ramo branco para um dos gânglios da cadeia simpática. Em seguida, o trajecto das fibras pode ser um de três:
Podem fazer sinapses com neurónios pós-ganglionares no gânglio onde chegam;
Podem ascender ou descer pela cadeia ganglionar, fazendo sinapses em outros gânglios;
Podem percorrer, por distâncias variáveis, a cadeia ganglionar, saindo por um dos nervos simpáticos que emergem dessa cadeia, indo terminar em um dos gânglios simpáticos periféricos.
Por conseguinte, o neurónio pós-ganglionar origina-se tanto de um dos gânglios da cadeia simpática como de um dos gânglios periféricos. De qualquer uma dessas formas, as fibras pós-ganglionares dirigem-se aos seus destinos nos vários órgãos.
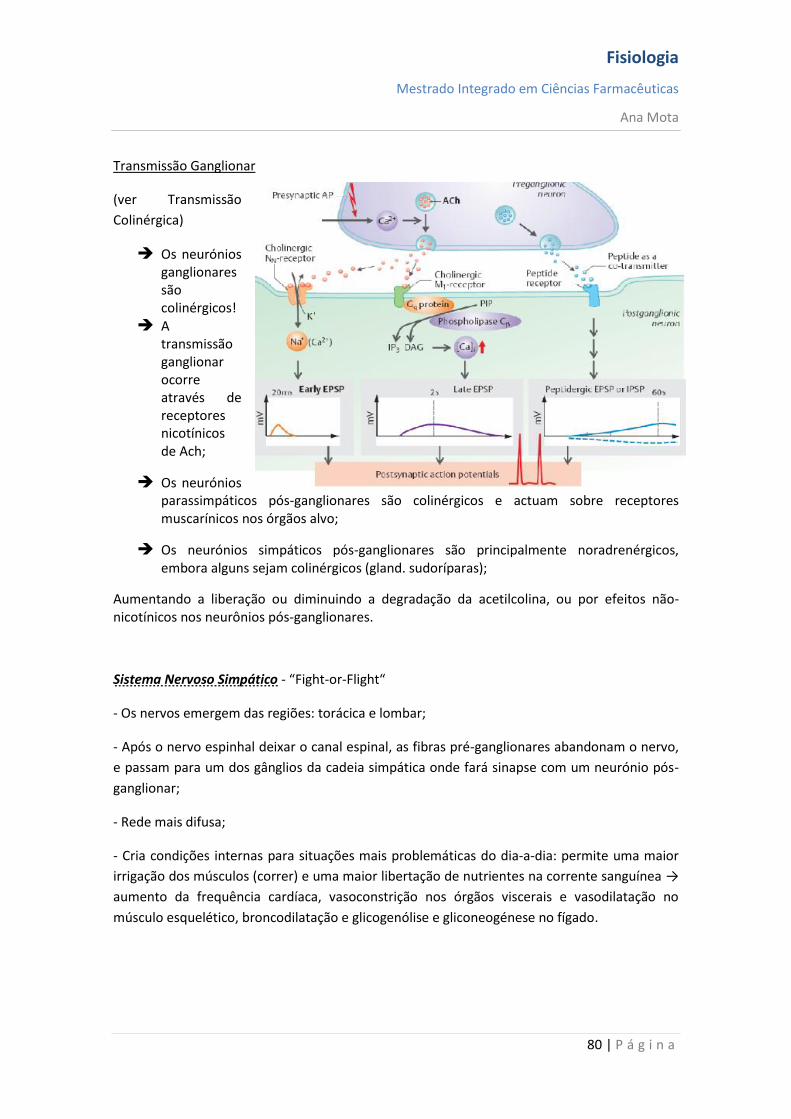
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
80 | P á g i n a
Transmissão Ganglionar
(ver Transmissão
Colinérgica)
Os neurónios ganglionares são colinérgicos!
A transmissão ganglionar ocorre através de receptores nicotínicos de Ach;
Os neurónios parassimpáticos pós-ganglionares são colinérgicos e actuam sobre receptores muscarínicos nos órgãos alvo;
Os neurónios simpáticos pós-ganglionares são principalmente noradrenérgicos, embora alguns sejam colinérgicos (gland. sudoríparas);
Aumentando a liberação ou diminuindo a degradação da acetilcolina, ou por efeitos não-nicotínicos nos neurônios pós-ganglionares.
Sistema Nervoso Simpático - “Fight-or-Flight“
- Os nervos emergem das regiões: torácica e lombar;
- Após o nervo espinhal deixar o canal espinal, as fibras pré-ganglionares abandonam o nervo,
e passam para um dos gânglios da cadeia simpática onde fará sinapse com um neurónio pós-
ganglionar;
- Rede mais difusa;
- Cria condições internas para situações mais problemáticas do dia-a-dia: permite uma maior
irrigação dos músculos (correr) e uma maior libertação de nutrientes na corrente sanguínea →
aumento da frequência cardíaca, vasoconstrição nos órgãos viscerais e vasodilatação no
músculo esquelético, broncodilatação e glicogenólise e gliconeogénese no fígado.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
81 | P á g i n a
Transmissão Adrenérgica
(ver Transmissão Colinérgica)
A L-tirosina, um aminoácido aromático encontrado nos líquidos corporais, é captada pelas células noradrenérgicas por um mecanismo de co-transporte específico com o Na+. No citosol há uma enzima, a tirosina hidroxilase, que é altamente específica e responsável pela conversão de L-Tirosina em DOPA (diidroxifenilalanina). Ela é inibida pelo produto final da via, a noradrenalina, e pelo substrato metilado (a-metiltirosina). Essa etapa é a limitante no processo de síntese de noradrenalina e adrenalina. Ainda no citosol, uma enzima inespecífica, a DOPA descarboxilase é responsável pela conversão de DOPA em dopamina. Essa enzima catalisa também descarboxilações em outros a.a. aromáticos, como o L-triptofano e a L-histidina, precursores respectivamente da serotonina e da histamina. A metildopa é um substrato falso que produz a a-metilnoradrenalina, que compete com a NA na vesícula. Essa substância é resistente à degradação pela MAO (substituição no C-a) e tem uma acção agonista selectiva sobre o receptor α2, diminuindo dessa maneira a liberação de NA. A dopamina-b-hidroxilase (DbH) catalisa a reacção de dopamina para noradrenalina. Essa enzima está presente principalmente na membrana das vesículas, mas há também o componente solúvel dentro das vesículas noradrenérgicas e parte é liberada na junção noradrenérgica, junto com NA. Como é uma enzima que não é degradada rapidamente, a sua concentração pode ser utilizada como meio de mensuração da função simpática. Em algumas células do cérebro e na supra-renal, há a presença da enzima feniletanoamina-N-metil-transferase. Essa catalisa a metilação da noradrenalina formando adrenalina. O armazenamento em vesículas é feito através de um mecanismo que transporta dopamina para dentro da vesícula onde a DbH é capaz de concentrar a noradrenalina a até 1 mol/L. Esse

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
82 | P á g i n a
mecanismo é inibido pela reserpina. As vesículas noradrenérgicas são elétron-densas, esféricas, com 40 a 100 nm, em grande número e a probabilidade da liberação através de estímulos simpáticos é menor do que a probabilidade para a liberação das análogas de ACh nas fibras colinérgicas. Mas como o número de vesículas colinérgicas é menor, se tem aproximadamente o mesmo número de moléculas agindo na sinapse em ambos os casos. Há ainda na vesícula ATP (4 ATP: 1 NA) e uma proteína denominada cromogranina A, que são responsáveis pela neutralidade da carga no interior da vesícula, diminuindo o gradiente electroquímico e estabilizando a NA na vesícula. Vale ressaltar a propriedade agonista simpática que possui o ATP. A liberação dá-se como em outras terminações. Estímulo → Abertura de "portões" de Ca2+ → Fusão de vesículas à membrana → Liberação do conteúdo na junção sináptica.
Autoregulação α2
A regulação da liberação dá-se por meio do receptor α2 que é inibidor da Adenil Ciclase e diminui a [Ca2+]i. O efeito dessa inibição parece ser a supressão de um canal específico de Ca2+ (o canal N). Acredita-se que a liberação de NA ao actuar nos receptores α2 inibem a liberação continuada de até cerca de 90% de noradrenalina (mecanismo auto-inibitório de feedback). A guanetidina, o betrílio e a betanidina são substâncias que inibem a liberação das vesículas pela chegada de um impulso. Simpatomiméticos de acções indirectas estimulam a liberação de vesículas noradrenérgicas, mesmo sem a chegada de um impulso nervoso. Entre os principais temos: tiramina, anfetamina e efedrina.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
83 | P á g i n a
Receptores Adrenérgicos
A norepinefrina (ou a epinefrina) activa sete domínios transmembrana. Estes domínios activam a proteína G heterotrimérica composta de subunidades alfa, beta e gama. As subunidades alfa e beta/gama dissociam e activam a fosfolipase C (PLC)-ß. Esta enzima hidroliza o fosfatidilinositol-4-5-bifosfato (PIP2) em dois produtos: inositol 1,4,5 trisfosfato (IP3) e diacilglicerol (DAG). IP3 liga-se a receptores no retículo endoplasmático, liberando o cálcio intracelular estocado. Sinergicamente com o DAG, a liberação de cálcio activa a PKC (proteína cinase). A PKC fosforila alvos específicos das proteínas celulares, alterando as suas funções. Existem também 2 tipos principais de receptores adrenérgicos, receptores alfa e os receptores beta (os receptores beta são, por sua vez, divididos em dois subtipos, chamados de beta1, e beta2, visto que determinadas substâncias actuam sobre certos receptores beta, mas não sobre outros. Por outro lado, existe distinção menos distinta entre os receptores alfa, também com dois subtipos, alfa1 e alfa2). A norepinefrina e a epinefrina, ambas secretadas pela medula da supra-renal, exercem efeitos diferentes na excitação dos receptores alfa e beta. A norepinefrina activa, principalmente, os receptores alfa, embora tenha efeito
excitador moderado sobre os receptores beta; Por outro lado, a epinefrina excita, com intensidade aproximadamente igual, os dois
tipos de receptores. Por conseguinte, os efeitos relativos da norepinefrina e da epinefrina sobre os diferentes órgãos efectores são determinados pelos tipos de receptores existentes nesses órgãos. Obviamente, se todos eles forem do tipo beta, a epinefrina será o excitante mais potente.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
84 | P á g i n a
- α1: vasoconstrição, midríase, glicogenólise hepática, relaxamento da musculatura lisa do trato gastrointestinal, secreção salivar espessa, secreção de suor nas extremidades (suor frio);
- α2: inibem a liberação do neurotransmissor, actuando como um mecanismo de feedback negativo. Controlam a liberação de insulina pelo pâncreas endócrino;
- β1: taquicardia, lipólise e relaxamento da musculatura lisa do trato gastrointestinal;
- β2: vasodilatação, broncodilatação, relaxamento da musculatura lisa do trato gastrointestinal e glicogenólise hepática;
- β3: lipólise.
Nota 1:
Os receptores alfa e beta não estão necessariamente associados à excitação ou à inibição, mas estão, simplesmente, associados à afinidade da hormona pelos receptores presentes em determinado órgão efector.
Nota 2:
Outra estrutura que recebe a informação do
neurónio pré-ganglionar é a zona medular da
glândula renal. As células desta zona – Células
Cromafins – contêm um NT que é libertado
directamente na corrente sanguínea e, por isso,
afectam todos os órgãos.
Sistema Nervoso Parassimpático - “Rest-and-Digest“
- Os nervos emergem do tronco cerebral ou da espinal medula, da região sacral;
- O 1º neurónio ou neurónio pré-ganglionar faz sinapse com o neurónio próximo do órgão –
neurónios pós-ganglionares;
- Há gânglios que se localizam dentro dos órgãos efectores (músculo cardíaco, liso e glândulas);
- Órgãos individualizados – aquando uma lesão apenas um órgão é afectado;
- Responsável por actividades “normais” do corpo humano → aumento da motilidade e da
secreção GI e diminuição da frequência cardíaca;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
85 | P á g i n a
- O sistema parassimpático, como o simpático, contém neurónios pré e pós-ganglionares. Contudo, excepto no caso de alguns poucos nervos parassimpáticos cranianos, as fibras pré-ganglionares passam, ininterruptas, até o órgão que vão controlar. Então, na parede desse órgão é que ficam situados os neurónios pós-ganglionares. As fibras pré-ganglionares fazem sinapses com essas células, que emitem fibras pós-ganglionares muito curtas, com comprimento variando entre 1 mm a vários centímetros, que se distribuem pela substância do órgão. Essa localização dos neurónios pós-ganglionares simpáticos no próprio órgão visceral é bastante diferente do padrão encontrado nos gânglios simpáticos, dado que os corpos celulares dos neurónios pós-ganglionares simpáticos sempre ficam situados nos gânglios das cadeias simpáticas ou em outros gânglios isolados no abdómen, e não no próprio órgão excitado.
Transmissão Colinérgica
As fibras nervosas simpáticas e parassimpáticas secretam, sem excepção, uma das duas substâncias transmissoras sinápticas, acetilcolina ou norepinefrina. As que secretam acetilcolina são ditas colinérgicas. As que secretam norepinefrina são chamadas de adrenérgicas, termo derivado de adrenalina, o nome britânico para norepinefrina. Todos os neurónios pré-ganglionares são colinérgicos, tanto no SN simpático como no parassimpático. Por conseguinte, a acetilcolina, ou substâncias semelhantes a esta, quando aplicada a um gânglio, irá excitar os neurónios pós-ganglionares simpáticos e parassimpáticos. Os neurónios pós-ganglionares do sistema parassimpático também são todos colinérgicos. Por outro lado, a maioria dos neurónios pós-ganglionares simpáticos é adrenérgica, embora isso não seja inteiramente verdade, visto que as fibras nervosas pós-ganglionares simpáticas que vão para as glândulas sudoríparas, para os músculos piloerectores e para alguns vasos sanguíneos são colinérgicas. Dessa forma, todas as terminações nervosas do sistema parassimpático secretam acetilcolina, enquanto a maioria das terminações nervosas simpáticas secreta norepinefrina. Essas hormonas, por sua vez, actuam sobre os diferentes órgãos, para produzir os efeitos simpáticos e parassimpáticos respectivos. Como resultado, são frequentemente chamados de transmissores simpático e parassimpático.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
86 | P á g i n a
Quando um potencial de acção invade o terminal de uma fibra, o processo de
despolarização aumenta a permeabilidade da membrana da fibra aos iões cálcio, permitindo que se difundam para o interior da terminação nervosa. Aí, esses iões interagem com as vesículas que estão adjacentes à membrana, fazendo com que se fundam à membrana e esvaziem o seu conteúdo para o exterior. Dessa forma, o transmissor é secretado.
A acetilcolina é sintetizada nas terminações nervosas das fibras nervosas colinérgicas. A maior parte de sua síntese ocorre no axoplasma, por fora das vesículas, e, em seguida, a acetilcolina é transportada para o interior dessas vesículas, onde fica armazenada, sob forma altamente concentrada, até ser liberada. Uma vez que a acetilcolina tenha sido secretada pela terminação nervosa colinérgica, ela perdura, no tecido, por apenas alguns segundos; então, a maior parte dela é degradada em ião acetato e em colina, pela enzima acetilcolinesterase, ligada ao colagéneo e aos glicosaminoglicanos do tecido conjuntivo local. Dessa forma, esse é o mesmo mecanismo de destruição que ocorre nas junções neuromusculares das fibras nervosas esqueléticas. A colina que é formada é, por sua vez, transportada de volta para a terminação nervosa, onde vai ser, de novo, utilizada na síntese de nova acetilcolina. Portanto, diversos tipos de receptores foram clonados para determinação da acção da acetilcolina, até ao momento conhece-se os receptores muscarínicos, (subdivididos em 5 tipos - M1, M2, M3, M4, e M5) acoplados à proteína G e os receptores nicotínicos (presentes nas junções neuromusculares e nos gânglios), acoplados a canais iónicos.
Receptores M1 são considerados neurais;
M2 cardíacos;
Receptores M3 estão presentes na musculatura lisa e nas glândulas. Geralmente a acetilcolina interage com este receptor M3 desencadeando contracção muscular lisa, porém as excepções são grandes. Em geral pode dizer-se que os receptores muscarínicos ímpares (M1, M3 e M5) possuem como efector a fosfolipase C que actua na transformação de um fosfolipídio em fosfatidilinositol com produção de diacilglicerol (DAG) e trifosfato de inositol (IP3). Já os receptores muscarínicos pares (M2, M4), actuam inibindo a adenilato ciclase e, portanto, diminuindo a concentração de AMPc intracelular. Os ésteres da colina (agentes muscarínicos) como por exemplo o carbacol actuam nos receptores muscarínicos promovendo uma actividade colinérgica (agonista colinérgico). O carbacol não mostra ser sensível à acção da colinesterase, no entanto suas actividades colinérgicas mais relevantes encontram-se nos músculos lisos gastrointestinais (aumentando sua contracção - peristaltismo), bem como no músculo liso vesical (promovendo o esvaziamento da bexiga urinária - aumento da peristalse uretral, contrai o detrusor da bexiga e aumenta a pressão miccional voluntária máxima) e, também acção sobre os receptores nicotínicos. Poucos efeitos sobre o sistema cardiovascular são verificados com o carbacol. Dentre os antagonistas muscarínicos a atropina é o representante mais conhecido.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
87 | P á g i n a
Os antagonistas dos receptores muscarínicos evitam os afeitos da acetilcolina bloqueando sua ligação com seu receptor colinérgico no músculo liso, cardíaco e também nas células glandulares. Actuam no bloqueio dos receptores muscarínicos presentes no SNC bem como em gânglios periféricos. As junções neuroefectoras parassimpáticas dos diferentes órgãos não têm sensibilidade igual aos antagonistas muscarínicos. Doses pequenas destes agentes, como a atropina, deprimem as secreções salivares, brônquicas e a transpiração. Com doses mais elevadas as pupilas ficam midriáticas, a acomodação do cristalino para a visão de perto é perdida, e os efeitos vagais sobre o coração são bloqueados. Doses ainda maiores inibem o controlo parassimpático sobre a bexiga urinária e sobre o trato gastrointestinal. Bloqueadores dos canais de cálcio. A indicação clínica para administração da nifedipina (um representante do grupo dos agentes bloqueadores dos canais de cálcio) inclui: angina variante, angina de esforço, angina instável, enfarte agudo do miocárdio, entre outros. As concentrações aumentadas do cálcio citosólico provocam aumento da contracção das células dos músculos lisos, cardíacos e vasculares. A entrada de cálcio extracelular é mais importante no início da contracção das células miocárdicas, enquanto a liberação de cálcio dos locais de armazenamento também participa na contracção da musculatura lisa vascular. A entrada de cálcio também deflagra o aumento de cálcio intracelular pelos canais de rianodina, presentes no retículo sarcoplasmático. Os canais de cálcio sensíveis à voltagem contêm domínios de sequências homólogas que são dispostos em fileiras dentro de uma única subunidade. Além da subunidade principal denominada alpha1, os canais de cálcio possuem outras subunidades associadas denominadas alpha2, beta, gama e delta. Dentre os tipos de canais de cálcio descritos temos os canais do tipo L, T e N, sendo que somente os canais do tipo L são sensíveis à acção das diidropiridinas como a nifedipina. Todos os bloqueadores de canal de cálcio aprovados para uso clínico provocam redução da resistência vascular coronária e aumentam o fluxo nestas artérias. As diidropiridonas são mais potentes quando comparada ao verapamil, por exemplo. A nifedipina administrada por via IV aumenta o fluxo sanguíneo no antebraço com pouco efeito pooling venoso, isso indica uma vasodilatação selectiva dos vasos de resistência arterial. Produzem também relaxamento da musculatura lisa vascular, portanto a resistência vascular e a pressão sanguínea diminuem com melhoria da contractilidade e da função ventricular.
A síntese da norepinefrina começa no axoplasma do terminal das fibras nervosas adrenérgicas, mas só é concluída no interior das vesículas.
Na medula supra-renal, essa sequência contínua por mais uma etapa, para transformar cerca de 80% da norepinefrina em epinefrina, da seguinte maneira: Após ter ocorrido a secreção de norepinefrina pela terminação nervosa, ela é removida do local por três meios diferentes: (1) Recaptação pela própria terminação nervosa, por meio de processo de transporte activo - responsável pela remoção de 50 a 80% da norepinefrina secretada;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
88 | P á g i n a
(2) Difusão, para longe da terminação, em direcção aos líquidos corporais circundantes, e deles para o sangue - responsável pela remoção da maior parte da norepinefrina remanescente; (3) Destruição por enzimas (uma delas é a monoaminoxidase, presente nas próprias terminações; outra é catecol-O-metiltransferase, presente, difusamente, em todos os tecidos). Na maioria das vezes, a norepinefrina secretada directamente em um tecido permanece activa por apenas poucos segundos, o que demonstra que sua receptação e difusão são bastante rápidas. Contudo, a norepinefrina e a epinefrina secretadas para o sangue pela medula supra-renal permanecem activas até que se difundam para algum tecido, onde vão ser destruídas pela catecol-O-metiltransferase; em geral, isso ocorre principalmente no fígado. Por conseguinte, quando secretadas para o sangue, a norepinefrina e a epinefrina permanecem activas por 10 a 30 segundos; em seguida, a sua actividade decrescente ainda perdura por 1 a vários minutos.
Nota:
Os canais de sódio e/ou cálcio são muitas vezes abertos, permitindo o influxo rápido dos iões respectivos para a célula, em geral despolarizando a membrana celular e excitando a célula;
A ligação da epinefrina ao seu receptor, na face externa de muitas células, aumenta a actividade da enzima adenilciclase, no interior da célula, e isso, por sua vez, provoca a formação de AMPc. Esse AMPc, então, desencadeia qualquer uma entre várias acções intracelulares, e o efeito exacto depende da maquinaria química da célula efectora.
Receptores da Acetilcolina
A acetilcolina activa dois tipos distintos de receptores. São chamados de receptores muscarínicos e nicotínicos. A razão para essas designações é que a muscarina, um veneno do cogumelo, só activa os receptores muscarínicos, sem qualquer acção sobre os nicotínicos, enquanto a nicotina só activa os receptores nicotínicos; a acetilcolina excita os dois.
Os receptores muscarínicos são encontrados em todas as células efectoras estimuladas pelos neurónios pós-ganglionares do SN parassimpático, bem como os estimulados pelos neurónios pós-ganglionares colinérgicos do sistema simpático;
Os receptores nicotínicos são encontrados nas sinapses entre os neurónios pré e pós-ganglionares dos sistemas: simpático e parassimpático. Também estão presentes nas membranas das fibras musculares esqueléticas, ao nível da junção neuromuscular.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
89 | P á g i n a
Receptores nicotínicos: provocam despolarização;
Receptores muscarínicos: dependendo do mecanismo, podem ter efeitos inibitórios
(hiperpolarização) ou excitatórios (despolarização)
Regulação Recíproca do SNA
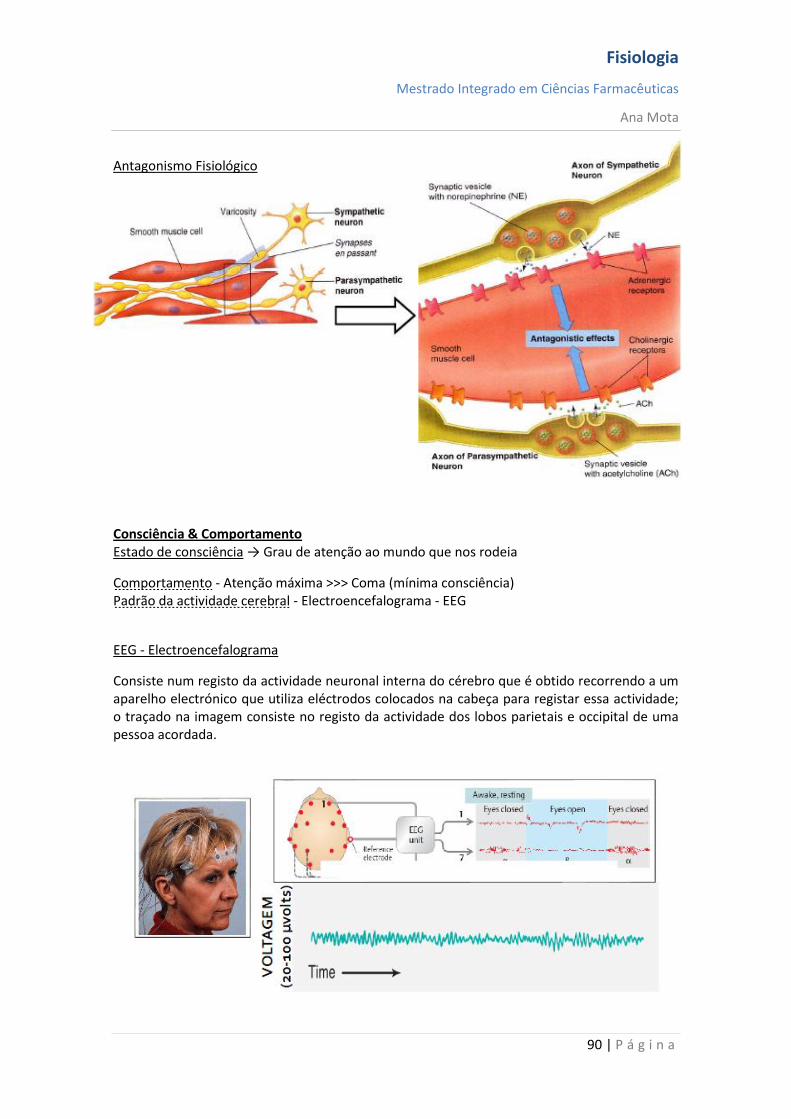
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
90 | P á g i n a
Antagonismo Fisiológico
Consciência & Comportamento Estado de consciência → Grau de atenção ao mundo que nos rodeia
Comportamento - Atenção máxima >>> Coma (mínima consciência) Padrão da actividade cerebral - Electroencefalograma - EEG
EEG - Electroencefalograma
Consiste num registo da actividade neuronal interna do cérebro que é obtido recorrendo a um aparelho electrónico que utiliza eléctrodos colocados na cabeça para registar essa actividade; o traçado na imagem consiste no registo da actividade dos lobos parietais e occipital de uma pessoa acordada.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
91 | P á g i n a
Potenciais Sinápticos
A intensidade é medida em milivolts e corresponde ao somatório dos potenciais sinápticos, os
quais variam em frequência, o que também é registado no EEG.
Ondas do EEG
Amplitude μV (0,5 - 100) - Quantidade de actividade eléctrica do mesmo tipo que ocorre em cada momento. Frequência Hz (1- 40)
- Grau de dessincronização dos neurónios que geram essa actividade eléctrica. Diagnóstico de actividade cerebral anormal
A epilepsia é caracterizada por actividade excessiva e incontrolada de parte do ou de todo o sistema nervoso central. A pessoa predisposta à epilepsia tem crises quando o nível basal da excitabilidade de seu sistema nervoso (ou da parte susceptível ao estado epiléptico) ultrapassa determinado limiar crítico. Mas, enquanto o nível da excitabilidade for mantido abaixo desse limiar, ela não apresentará crises. Em termos básicos, a epilepsia pode ser dividida em três tipos principais: epilepsia tipo grande mal, epilepsia tipo pequeno mal e epilepsia focal.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
92 | P á g i n a
Ondas cerebrais registadas no EEG
As imensidades das ondas cerebrais registadas da superfície do couro cabeludo variam entre 0 e 200 microvolts e as suas frequências oscilam entre uma a cada poucos segundos e 50 ou mais por segundo. A forma das ondas é dependente do grau de actividade do córtex cerebral, as ondas variam acentuadamente entre os estados de vigília, sono e coma. Na maior parte do tempo, essas ondas são irregulares, não sendo possível definir o seu padrão geral no EEG. Todavia, por vezes, podem surgir padrões distintivos. Alguns deles são característicos de anormalidades específicas do encéfalo, como a epilepsia. Outros aparecem, até mesmo, em pessoas normais sadias, podendo ser classificados como ondas alfa, beta, teta e delta.
As ondas alfa são ondas rítmicas, com frequência entre 8 e 13 por segundo, sendo encontradas no EEG de quase todas as pessoas adultas normais, quando acordadas, em estado de cerebração tranquilo e em repouso. Essas ondas são mais intensas na região occipital, mas também podem ser registadas na região do couro cabeludo sobre as regiões parietal e frontal. A sua voltagem é, em média, de 50 microvolts. Durante o sono profundo, as ondas alfa desaparecem completamente; se a atenção da pessoa acordada é direccionada para algum tipo específico de actividade mental, as ondas alfa são substituídas pelas ondas beta, assíncronas, com maior frequência e menor voltagem;
As ondas beta têm frequências acima de 14 ciclos por segundo, podendo atingir 80 por segundo. São as ondas mais frequentemente registadas nas regiões parietal e frontal do couro cabeludo, durante a activação extra do SNC e durante os estados de tensão;
As ondas teta têm frequência entre 4 e 7 ciclos por segundo. Ocorrem, em sua maioria, nas regiões parietal e temporal em crianças e, em alguns adultos, durante stress emocional, em especial durante desapontamentos e frustrações. As ondas teta também ocorrem em muitas doenças cerebrais;
As ondas delta incluem todas as ondas do EEG com frequência abaixo de 3,5 ciclos por segundo e frequentemente têm voltagens que podem ter valores duas ou quatro vezes maiores do que a maioria dos outros tipos de ondas cerebrais. Ocorrem no sono muito profundo, na infância e em doenças orgânicas cerebrais graves. Também ocorrem no córtex de animais que sofreram transecções subcorticais, separando o córtex cerebral do tálamo. Por conseguinte, as ondas delta podem ocorrer, de forma restrita, no córtex, independentemente da actividade nas regiões inferiores do encéfalo.
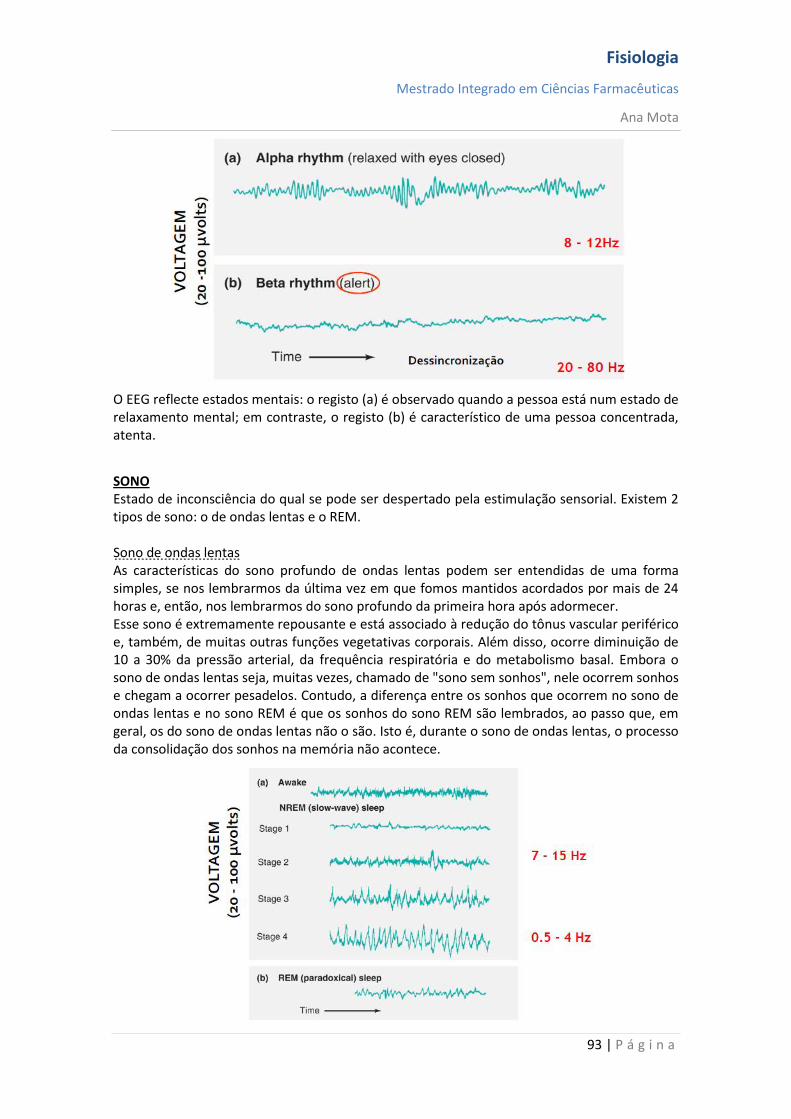
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
93 | P á g i n a
O EEG reflecte estados mentais: o registo (a) é observado quando a pessoa está num estado de relaxamento mental; em contraste, o registo (b) é característico de uma pessoa concentrada, atenta.
SONO Estado de inconsciência do qual se pode ser despertado pela estimulação sensorial. Existem 2 tipos de sono: o de ondas lentas e o REM. Sono de ondas lentas As características do sono profundo de ondas lentas podem ser entendidas de uma forma simples, se nos lembrarmos da última vez em que fomos mantidos acordados por mais de 24 horas e, então, nos lembrarmos do sono profundo da primeira hora após adormecer. Esse sono é extremamente repousante e está associado à redução do tônus vascular periférico e, também, de muitas outras funções vegetativas corporais. Além disso, ocorre diminuição de 10 a 30% da pressão arterial, da frequência respiratória e do metabolismo basal. Embora o sono de ondas lentas seja, muitas vezes, chamado de "sono sem sonhos", nele ocorrem sonhos e chegam a ocorrer pesadelos. Contudo, a diferença entre os sonhos que ocorrem no sono de ondas lentas e no sono REM é que os sonhos do sono REM são lembrados, ao passo que, em geral, os do sono de ondas lentas não o são. Isto é, durante o sono de ondas lentas, o processo da consolidação dos sonhos na memória não acontece.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
94 | P á g i n a
Padrão do EEG sofre modificações durante o sono, reflectindo 4 estádios do sono; a duração de cada série é de ~ 90 minutos, e cada ciclo repete-se 4-8x por noite.
É constituído por 4 fases. Ao longo das 4 fases, a amplitude e a sincronização aumentam e diminui a frequência. O ciclo de 1 a 4 demora cerca de 45 minutos e é repetido várias vezes. O estádio 4 ocorre quando estamos a dormir profundamente e é designado de sono recuperador. Durante a repetição dos ciclos, a fase 4 vai encurtando de tempo e o sono REM vai aumentando de tempo.
Sono REM (sono paradoxal, sono dessincronizado) Em noite normal de sono, episódios de sono REM, cada um durando de 5 a 30 minutos, ocorre, em média, a cada 90 minutos, com o primeiro deles acontecendo dentro de 80 a 100 minutos após o adormecer. Quando a pessoa está extremamente sonolenta, a duração de cada surto de sono REM fica muito curta, podendo, até mesmo, não ocorrer. Por outro lado, à medida que a pessoa fica mais descansada, no correr da noite, a duração dos surtos de sono REM aumenta. Existem diversas características muito importantes do sono REM: Em geral, está associado ao sonhar (sonhos que podem ser lembrados) e a
movimentos musculares corporais activos; A pessoa fica bem mais difícil de ser acordada por meio de estímulos sensoriais do que
durante o sono de ondas lentas e, no entanto, as pessoas acordam, pela manhã, durante um episódio de sono REM, e não do sono de ondas lentas;
O tônus muscular de todo o corpo fica extremamente deprimido, indicando forte inibição das projecções para a medula espinhal das áreas excitatórias do tronco cerebral;
As frequências cardíaca e respiratória ficam irregulares, o que é característico do estado do sonho;
Apesar da intensa inibição dos músculos periféricos, ainda assim surgem poucos movimentos musculares sem regularidade. Esses movimentos incluem, de modo especial, movimentos rápidos dos olhos;
Durante o sono REM, o encéfalo está muito activo, e o metabolismo global encefálico pode aumentar por até 20%;
O EEG regista padrão de ondas cerebrais semelhante ao que é registado na vigília. Por isso, esse tipo de sono foi chamado de sono paradoxal, porque é um paradoxo que a pessoa esteja dormindo, apesar de sua intensa actividade cerebral;
Acentuado por agonistas colinérgicos.
Em resumo, o sono REM é um tipo de sono em que o cérebro está muito activo.
Contudo, a actividade cerebral não é canalizada na direcção apropriada para que a pessoa possa tomar conhecimento do que a rodeia e, por conseguinte, ficar acordada.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
95 | P á g i n a
Fases do Sono
Alterações dos ciclos de sono com a idade:
Nº de horas de sono;
% de horas de sono fase 4;
Duração do sono REM
Alterações de parâmetros fisiológicos observadas durante o sono:
Tensão muscular basal;
Ritmo respiratório;
Ritmo cardíaco;
Pressão Arterial.
Os ciclos de sono variam
ao longo da vida:
Bebés dormem
quase 24 horas;
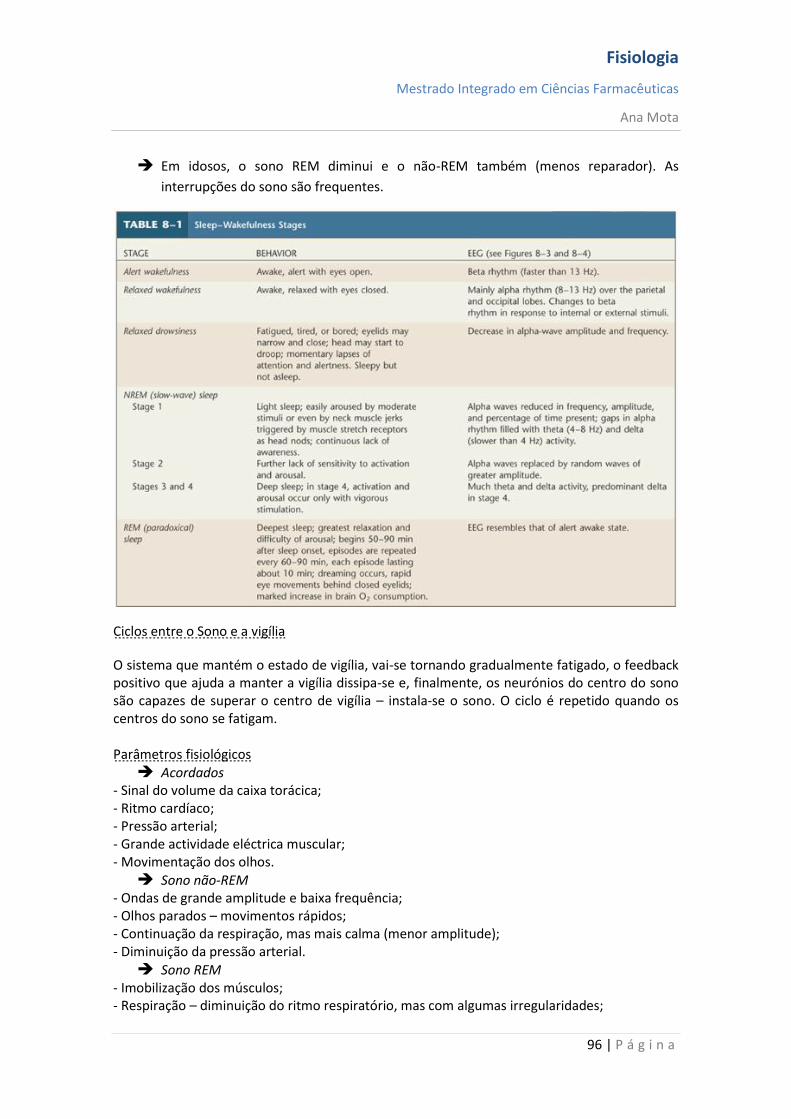
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
96 | P á g i n a
Em idosos, o sono REM diminui e o não-REM também (menos reparador). As
interrupções do sono são frequentes.
Ciclos entre o Sono e a vigília
O sistema que mantém o estado de vigília, vai-se tornando gradualmente fatigado, o feedback positivo que ajuda a manter a vigília dissipa-se e, finalmente, os neurónios do centro do sono são capazes de superar o centro de vigília – instala-se o sono. O ciclo é repetido quando os centros do sono se fatigam. Parâmetros fisiológicos Acordados
- Sinal do volume da caixa torácica; - Ritmo cardíaco; - Pressão arterial; - Grande actividade eléctrica muscular; - Movimentação dos olhos. Sono não-REM
- Ondas de grande amplitude e baixa frequência; - Olhos parados – movimentos rápidos; - Continuação da respiração, mas mais calma (menor amplitude); - Diminuição da pressão arterial. Sono REM
- Imobilização dos músculos; - Respiração – diminuição do ritmo respiratório, mas com algumas irregularidades;
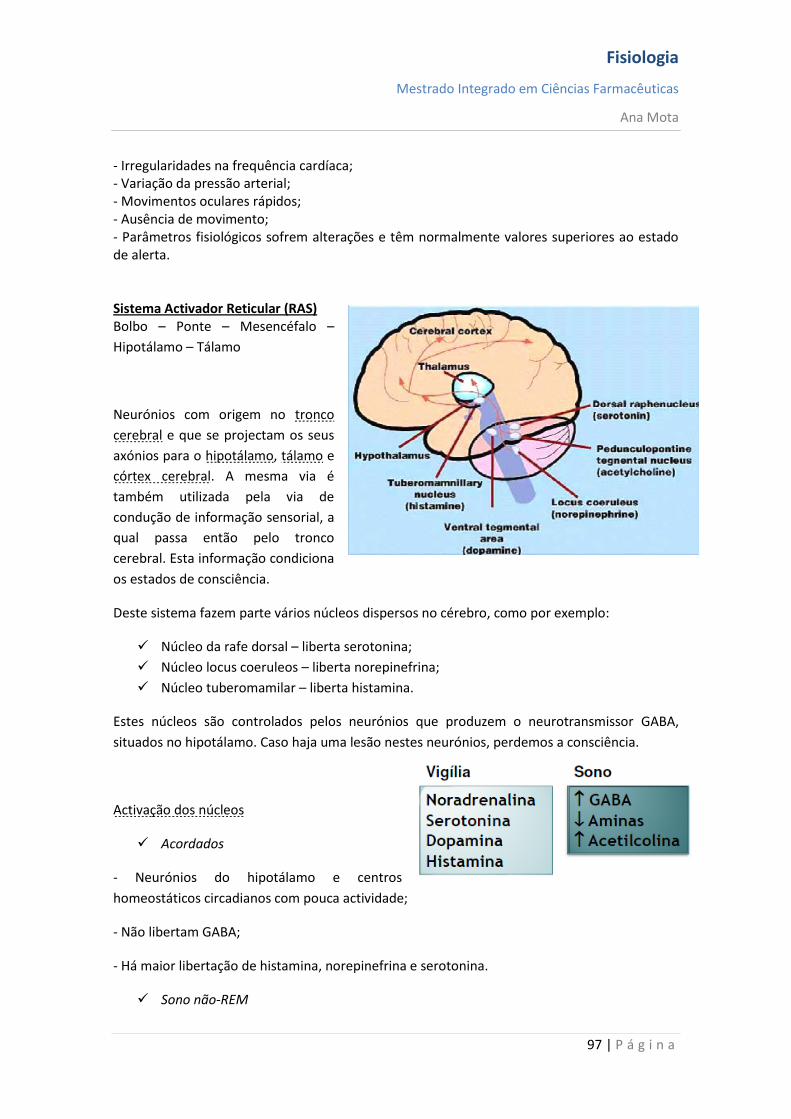
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
97 | P á g i n a
- Irregularidades na frequência cardíaca; - Variação da pressão arterial; - Movimentos oculares rápidos; - Ausência de movimento; - Parâmetros fisiológicos sofrem alterações e têm normalmente valores superiores ao estado de alerta. Sistema Activador Reticular (RAS) Bolbo – Ponte – Mesencéfalo –
Hipotálamo – Tálamo
Neurónios com origem no tronco
cerebral e que se projectam os seus
axónios para o hipotálamo, tálamo e
córtex cerebral. A mesma via é
também utilizada pela via de
condução de informação sensorial, a
qual passa então pelo tronco
cerebral. Esta informação condiciona
os estados de consciência.
Deste sistema fazem parte vários núcleos dispersos no cérebro, como por exemplo:
Núcleo da rafe dorsal – liberta serotonina;
Núcleo locus coeruleos – liberta norepinefrina;
Núcleo tuberomamilar – liberta histamina.
Estes núcleos são controlados pelos neurónios que produzem o neurotransmissor GABA,
situados no hipotálamo. Caso haja uma lesão nestes neurónios, perdemos a consciência.
Activação dos núcleos
Acordados
- Neurónios do hipotálamo e centros
homeostáticos circadianos com pouca actividade;
- Não libertam GABA;
- Há maior libertação de histamina, norepinefrina e serotonina.
Sono não-REM

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
98 | P á g i n a
- Neurónios do hipotálamo e centros homeostáticos circadianos apresentam maior actividade;
- Grande libertação de GABA, o qual inibe a libertação de histamina.
Sono REM
- Neurónios do hipotálamo e centros homeostáticos circadianos apresentam maior actividade;
- Grande libertação de acetilcolina, a qual inibe a libertação de serotonina e norepinefrina.
Fármacos para dormir têm
a mesma acção que o
GABA. Actuam nos
receptores do GABA e
fazem com que os níveis
de noradrenalina,
serotonina e histamina
diminuam.
Coma
- Situação clínica em que o indivíduo não apresenta respostas adequadas a estimulação
ambiental, apresentando-se normalmente de olhos fechados e com comprometimento do
estado de vigília, atenção, linguagem, raciocínio e emoções;
- Caracteriza-se por ser um estado não facilmente reversível da consciência, em consequência
da diminuição da actividade da formação reticular ascendente.
O desencadear de um estado de vigília pode ser atribuído a:
- Estímulo da porção medial da formação reticular, especialmente no mesencéfalo e na parte
superior da ponte, ocasiona intensa vigília;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
99 | P á g i n a
- Estímulo difuso dos nervos sensoriais pelo corpo ocasiona vigília. Estes enviam sinais à porção
mesencefálica do RAS;
- Estímulos do córtex cerebral causam elevado nível de vigília. São enviados fortes sinais para a
porção mesencefálica e talâmica do sistema reticular;
- Estímulo do hipotálamo, especialmente das regiões laterais, pode causar extremos graus de
vigília. Fortes sinais são transmitidos ao sistema reticular.
Ritmo Circadiano
As velocidades de secreção do CRH, ACTH e cortisol estão todas elevadas pela manhã, porém apresentam-se baixas à noite. O nível plasmático de cortisol varia entre cerca de 20 μg/dl 1 hora antes de o indivíduo acordar pela manhã até cerca de 5 μg/dl por volta da meia-noite. Esse efeito decorre da alteração cíclica de 24 horas nos sinais do hipotálamo responsáveis pela secreção de cortisol. Quando o indivíduo modifica os seus hábitos diários de sono, o ciclo muda correspondentemente. Uma das razões pelas quais o ciclo é tão importante é que as determinações dos níveis sanguíneos de cortisol só se mostram válidas quando relacionadas com o momento do ciclo em que foram efectuadas.
ACTH → hormona adrenocorticotrópica;
CRH → hormona de libertação da corticotropina
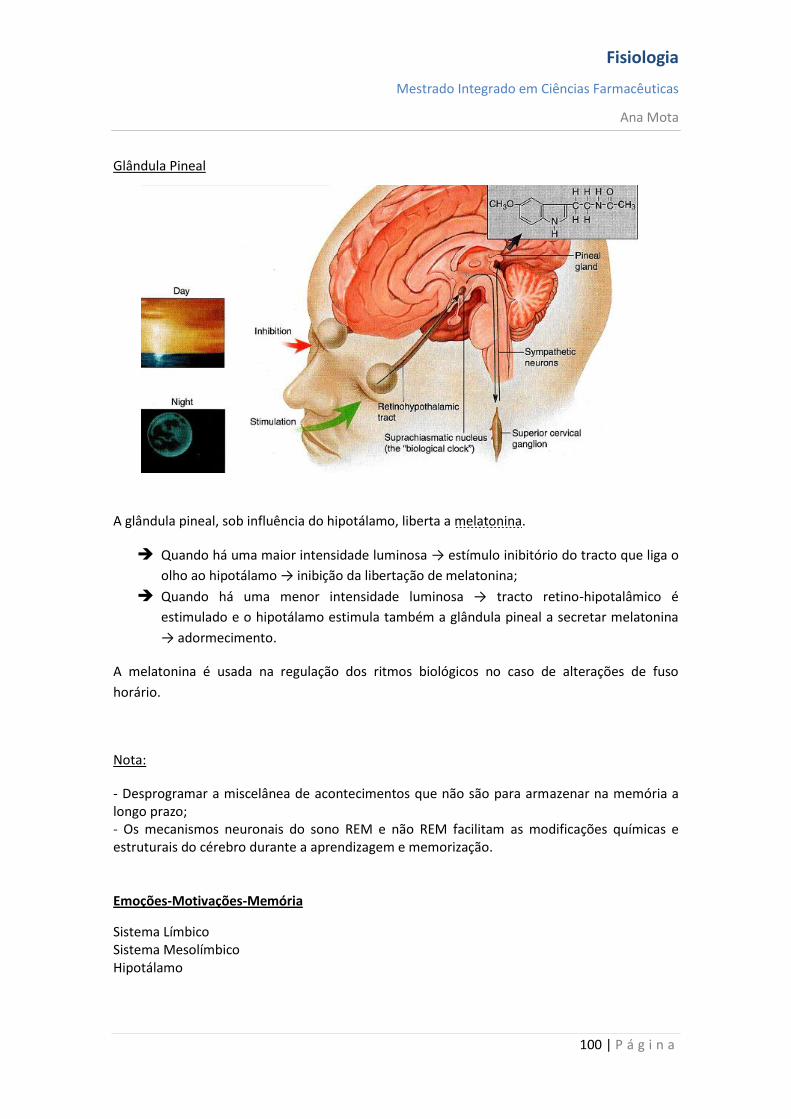
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
100 | P á g i n a
Glândula Pineal
A glândula pineal, sob influência do hipotálamo, liberta a melatonina.
Quando há uma maior intensidade luminosa → estímulo inibitório do tracto que liga o
olho ao hipotálamo → inibição da libertação de melatonina;
Quando há uma menor intensidade luminosa → tracto retino-hipotalâmico é
estimulado e o hipotálamo estimula também a glândula pineal a secretar melatonina
→ adormecimento.
A melatonina é usada na regulação dos ritmos biológicos no caso de alterações de fuso
horário.
Nota:
- Desprogramar a miscelânea de acontecimentos que não são para armazenar na memória a longo prazo; - Os mecanismos neuronais do sono REM e não REM facilitam as modificações químicas e estruturais do cérebro durante a aprendizagem e memorização.
Emoções-Motivações-Memória
Sistema Límbico Sistema Mesolímbico Hipotálamo

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
101 | P á g i n a
Hipotálamo
Esta glândula é importante na coordenação e no controlo de várias tarefas a vários níveis:
Órgãos internos (ligação ao SNA);
Sistema endócrino (reprodução, temperatura, nutrição);
Sistema imunitário (defesa);
Sistema límbico e mesolímbico;
Comportamento, emoções e motivações.
Motivações
Incluem tudo o que fazemos consciente ou inconsciente.
Motivações primárias – satisfação das necessidades básicas de sobrevivência.
Exemplos: ingestão de alimentos e manutenção da temperatura.
Motivações secundárias – influenciadas por hábitos, aprendizagem, intelecto,
emoções.
Sistema mesolímbico
Os conceitos de recompensa e punição são inertentes.
Nestes estão envolvidos neurónios que têm ligação com o hipotálamo, formando o sistema
mesolímbico:
Neurónios que se localizam no locus cerebelus;
Neurónios que se localizam no mesencéfalo;
Neurónios que se localizam no núcleo accumbens, que se projecta para o córtex
frontal.
A dopamina é um NT libertado nestes neurónios, sendo responsável pelo sentimento de
satisfação.
A cocaína estimula a libertação de uma maior quantidade de dopamina, o que aumenta o grau
de satisfação. Da mesma forma, os opiáceos funcionam da mesma maneira – provocando
dependência e alterações comportamentais.
Existindo fármacos que actuam neste sistema.
O sistema mesolímbico dopaminérgico é responsável pela sensação de prazer e recompensa. Alterações neste circuito constituem o principal mecanismo pelo qual as drogas psicoactivas modificam o comportamento.
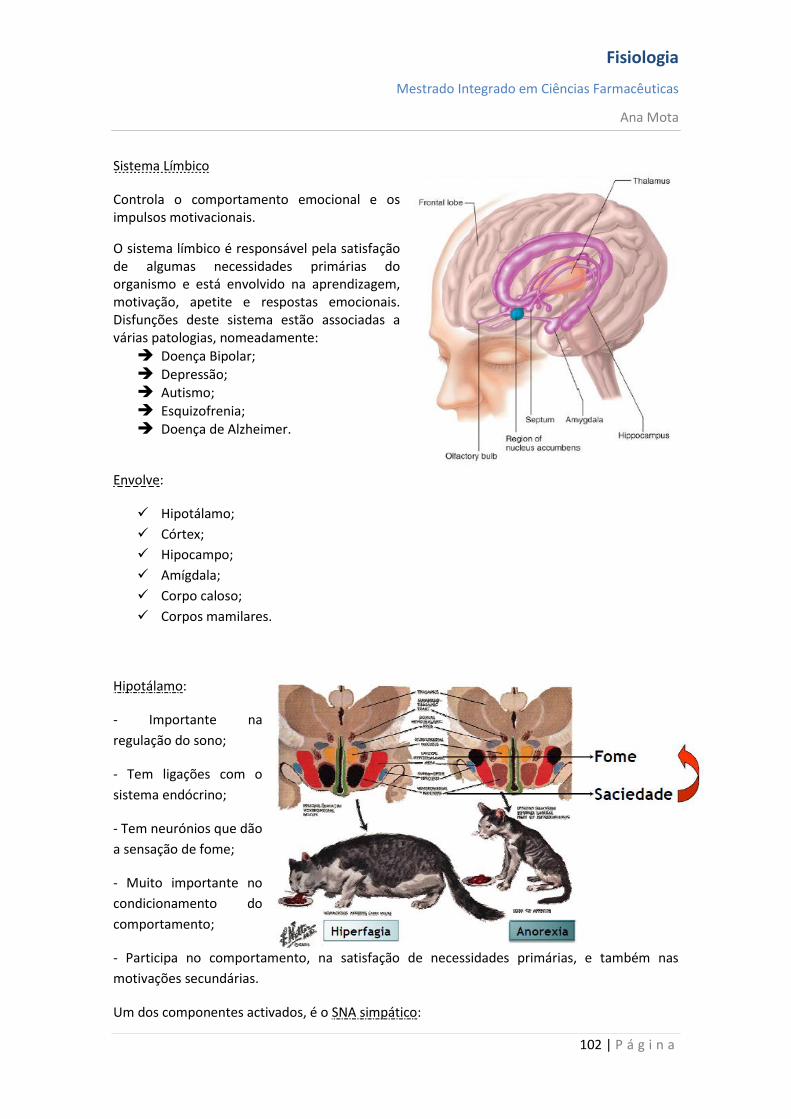
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
102 | P á g i n a
Sistema Límbico
Controla o comportamento emocional e os impulsos motivacionais.
O sistema límbico é responsável pela satisfação de algumas necessidades primárias do organismo e está envolvido na aprendizagem, motivação, apetite e respostas emocionais. Disfunções deste sistema estão associadas a várias patologias, nomeadamente:
Doença Bipolar; Depressão; Autismo; Esquizofrenia; Doença de Alzheimer.
Envolve:
Hipotálamo;
Córtex;
Hipocampo;
Amígdala;
Corpo caloso;
Corpos mamilares.
Hipotálamo:
- Importante na
regulação do sono;
- Tem ligações com o
sistema endócrino;
- Tem neurónios que dão
a sensação de fome;
- Muito importante no
condicionamento do
comportamento;
- Participa no comportamento, na satisfação de necessidades primárias, e também nas
motivações secundárias.
Um dos componentes activados, é o SNA simpático:

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
103 | P á g i n a
Muitas sensações corporais;
Sentimento de emoção;
Participação de neurónios do córtex.
Quando existem anomalias nos neurónios do córtex límbico, modificam-se os circuitos, o que
resulta em doenças:
Autismo
- Não existe manifestação de doenças;
- A pessoa fecha-se no seu próprio mundo.
Esquizofrenia
- Distúrbio manifestado em pessoas que ouvem vozes;
- Delírios de grandeza;
- Medo intenso;
- Paranóia.
Depressão
- Presume-se que, especialmente o sistema da norepinefrina e, talvez, o da serotonina funcionem normalmente para produzir o estímulo ao sistema límbico, para aumentar o sentimento de bem-estar da pessoa, para gerar felicidade, contentamento, bom apetite, desejo sexual adequado e equilíbrio psicomotor, embora uma coisa boa - quando em demasia, possa levar à mania. Em apoio desse conceito existe o fato de que os centros de recompensa e de punição do hipotálamo e áreas circundantes recebem grande número de terminações nervosas do sistema da norepinefrina.
Doença de Alzheimer
- Incide em pessoas idosas;
- Caracterizado pela acumulação de placas amilóides em áreas amplamente dispersas do
cérebro, incluindo o córtex cerebral, o hipocampo e os gânglios da base;
- Muitos pacientes exibem anormalidade genética envolvendo a apolipoproteína E –
transporta colesterol.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
104 | P á g i n a
Emoções
Os neurónios que fazem parte deste sistema têm fortes ligações entre si.
Quando há lesão numa região, ocorre alteração da personalidade da pessoa:
Alteração das competências, mesmo a nível intelectual;
Incapaz de regular emoções → responsável – córtex pré-frontal.
Amígdala – pessoas com lesões nestes neurónios tornam-se agressivas;
Hipocampo – importante na memória. Lesões nesta área conduzem a uma profunda
incapacidade de formar novas memórias – esquecimento de memórias imediatas.
Existem vários tipos de memória, sendo classificada de acordo com a execução de tarefas em
memórias a curto/longo prazo. As memórias a longo prazo ficam gravados no nosso cérebro
para toda a vida.
Aprendizagem e Memória
Além disso, frequentemente as memórias são classificadas de acordo com o tipo de informação que é armazenada. Uma dessas classificações divide a memória em memória declarativa e memória processual, de acordo com o seguinte:
Memória declarativa está associada a acontecimentos actuais baseados na experiência directa da pessoa:
- Memória do ambiente; - Memória das relações temporais; - Memórias das causas da experiência; - Memória do significado da experiência; - Memória das deduções que permaneceram na mente da pessoa. Memória processual consiste no conhecimento de uma sequência de acontecimentos
e das relações entre os acontecimentos.
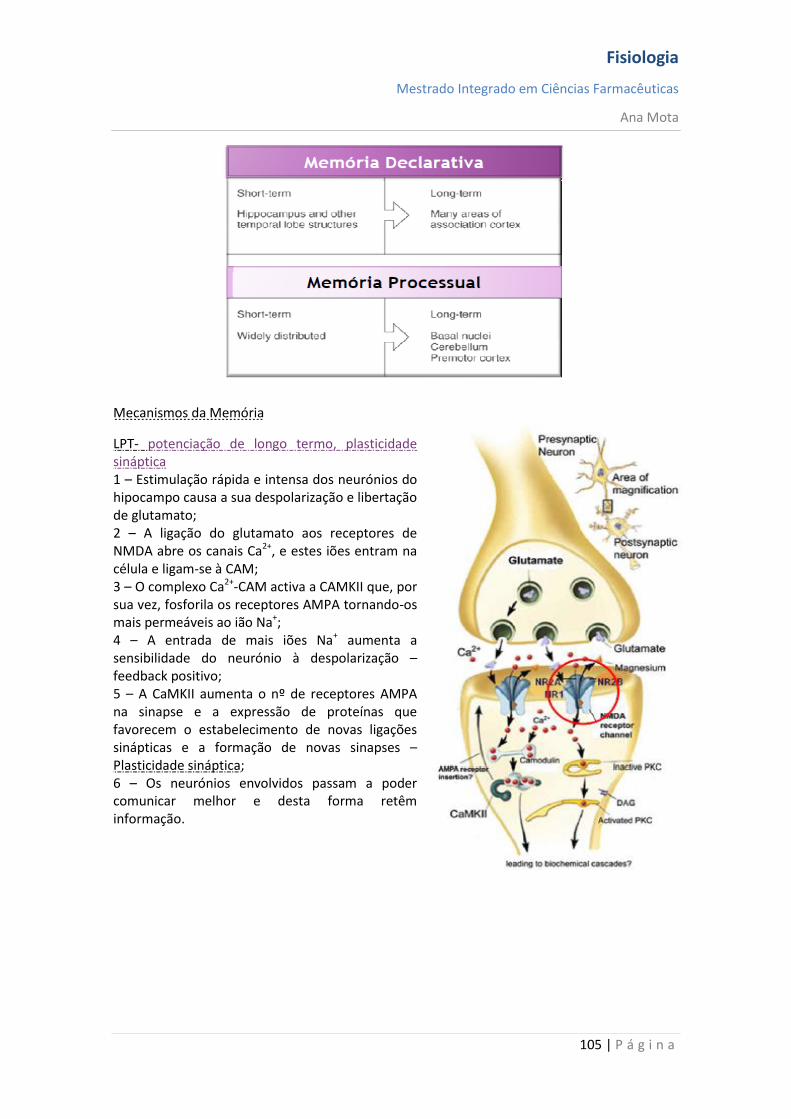
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
105 | P á g i n a
Mecanismos da Memória
LPT- potenciação de longo termo, plasticidade sináptica 1 – Estimulação rápida e intensa dos neurónios do hipocampo causa a sua despolarização e libertação de glutamato; 2 – A ligação do glutamato aos receptores de NMDA abre os canais Ca2+, e estes iões entram na célula e ligam-se à CAM; 3 – O complexo Ca2+-CAM activa a CAMKII que, por sua vez, fosforila os receptores AMPA tornando-os mais permeáveis ao ião Na+; 4 – A entrada de mais iões Na+ aumenta a sensibilidade do neurónio à despolarização – feedback positivo; 5 – A CaMKII aumenta o nº de receptores AMPA na sinapse e a expressão de proteínas que favorecem o estabelecimento de novas ligações sinápticas e a formação de novas sinapses – Plasticidade sináptica; 6 – Os neurónios envolvidos passam a poder comunicar melhor e desta forma retêm informação.
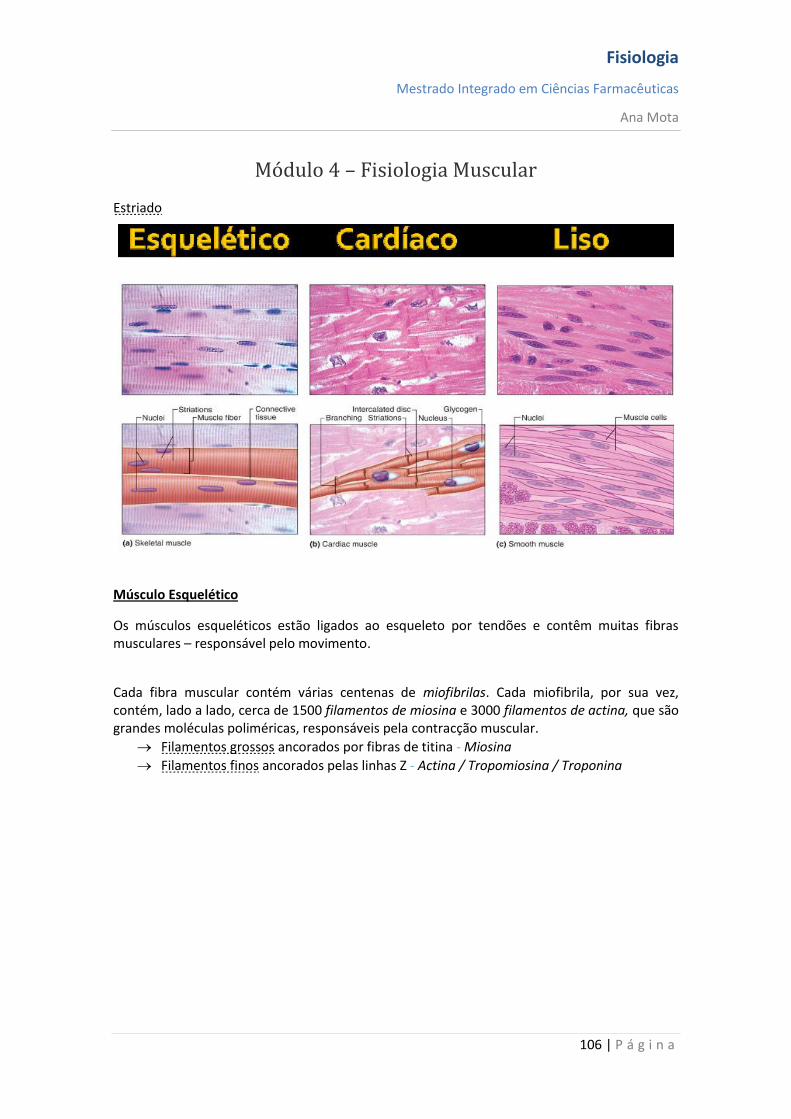
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
106 | P á g i n a
Módulo 4 – Fisiologia Muscular
Estriado
Músculo Esquelético
Os músculos esqueléticos estão ligados ao esqueleto por tendões e contêm muitas fibras musculares – responsável pelo movimento.
Cada fibra muscular contém várias centenas de miofibrilas. Cada miofibrila, por sua vez, contém, lado a lado, cerca de 1500 filamentos de miosina e 3000 filamentos de actina, que são grandes moléculas poliméricas, responsáveis pela contracção muscular.
Filamentos grossos ancorados por fibras de titina - Miosina
Filamentos finos ancorados pelas linhas Z - Actina / Tropomiosina / Troponina

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
107 | P á g i n a
Aparelho contráctil
- São unidades
isoladas, apenas
coladas umas às outras
por tecido conjuntivo;
- As proteínas que o
constituem
determinam a parede
contráctil e formam os
sarcómeros.
O sarcómero é constituído por:

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
108 | P á g i n a
Filamentos grosso – localizados numa zona mais central (filamentos de miosina).
Têm 2 porções:
- Globular – “cabeça de miosina”, com actividade ATPásica, libertando energia para a
contracção; importante nas ligações aos filamentos finos;
- Filamentosa.
Filamentos finos – constituídos por 3 proteínas: troponina, actina e tropomiosina, que
formam um arranjo que regula as interacções entre os 2 tipos de filamentos e estão
envolvidos na contracção.
A actina forma um cordão com vários locais activos. É nestes locais que a miosina (porção
globular) se vai ligar para a activação do mecanismo de contracção.
Em repouso, os locais activos da actina são cobertos através da acção de 2 proteínas:
Troposiosina – impede
acesso aos locais de actina
pela miosina;
Troponina – apresenta na
sua constituição 3 locais: T
(regulação), I (inibição da
interacção com os locais
activos e a manutenção do
posicionamento) e C (à qual
se liga o ião Ca2+ - ião
desencadeador da
contracção muscular). A troponina “tapa” os locais de actina, mas quando se afasta
permite a contracção muscular.
Contracção muscular
Resulta da formação de pontes de ligação entre os filamentos de actina e miosina, ocorrendo
um deslizamento de duas moléculas.
1. Um potencial de acção percorre um axónio motor até às suas terminações nas fibras musculares; 2. Em cada terminação, há secreção de pequena quantidade da substância neurotransmissora, chamada acetilcolina; 3. A acetilcolina actua sobre área localizada da membrana da fibra muscular, abrindo numerosos canais proteicos acetilcolina dependentes;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
109 | P á g i n a
4. A abertura desses canais dependentes da acetilcolina permite o influxo de grande quantidade de iões sódio para o interior da membrana da fibra muscular, no ponto da terminação nervosa. Isso produz um potencial de acção na fibra muscular; 5. O potencial de acção propaga-se ao longo da membrana da fibra muscular do mesmo modo como o faz nas membranas neurais; 6. O potencial de acção despolariza a membrana da fibra muscular e também penetra profundamente no interior dessa fibra. Aí, faz com que o retículo sarcoplasmático liberte, para as miofibrilas, grande quantidade de iões cálcio, que ficam armazenadas em seu interior; 7. Os iões cálcio geram forças atractivas entre os filamentos de actina e de miosina, fazendo com que deslizem um em direcção ao outro, o que constitui o processo contráctil. 8. Após uma fracção de segundo, os iões cálcio são bombeados de volta para o retículo sarcoplasmático, onde permanecem armazenados até que ocorra novo potencial de acção muscular - termina a contracção muscular. Nota: O comprimento dos filamentos não sofre alteração.
Ciclo das Pontes Cruzadas
1. Durante a contracção de um músculo, os iões de cálcio ligam-se à troponina, provocando a exposição dos locais activos nos miofilamentos de actina;
2. As moléculas de miosina fixam-se aos locais activos expostos nos miofilamentos de actina, formando pontes, e o fósforo (P) é libertado da cabeça de miosina;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
110 | P á g i n a
3. A energia armazenada na cabeça dos miofilamentos de miosina é utilizada para mover a cabeça da molécula de miosina. O movimento da cabeça leva o miofilamento de actina a deslizar ao longo do miofilamento de miosina. O ADP é libertado da cabeça de miosina;
4. Uma molécula de ATP liga-se à cabeça de miosina, o que faz libertar a actina da miosina;
5. A molécula de ATP desdobra-se em ADP e fosfato, que permanecem ligados à cabeça de miosina; a molécula da cabeça de miosina volta à sua posição de repouso e a energia é armazenada na cabeça da molécula de miosina. Se o Ca2+ ainda estiver ligado à troponina, a formação e movimento das pontes repete-se (retorno ao passo 1). Este ciclo ocorre muitas vezes durante a contracção muscular.
Nota:
Os ciclos das pontes cruzadas fazem com que o sarcómero encurte;
A tropomiosina move-se (rodando), para expôr os locais da actina onde se liga a miosina;
A miosina liga-se à actina e arrasta-a, causando aproximação das linhas Z e reduzindo a largura das bandas I.
Função do Ca2+
Músculo Relaxado - Tropomiosina bloqueia o local de ligação da miosina à actina; Contracção Muscular - Ca2+ liga-se à Troponina; este complexo afasta a Tropomiosina expondo os locais activos da actina e permitindo as pontes cruzadas.
Funções do ATP
Fonte imediata de energia para a contracção muscular.
A energia necessária para a sua formação provém de quatro fontes essenciais: - Respiração aeróbia; - Respiração anaeróbia; - Fosforilação oxidativa: a capacidade de produzir ATP pela via oxidativa é suportada pela disponibilidade de oxigénio que se encontra armazenado ligado à Mioglobina; - Hidrólise de creatina fosfato.
Processos que utilizam ATP no músculo esquelético Hidrólise do ATP pela ATPase da miosina permite a ligação desta à actina; Durante o movimento das pontes cruzadas a miosina está fortemente ligada à actina.
Esta ligação tem de ser quebrada de forma a permitir que a PT se possa voltar a ligar a um novo monómero de actina de forma a reiniciar o ciclo. Essa ligação é quebrada quando o ATP se liga de novo à miosina. Após essa separação, o ATP que se ligou à miosina é hidrolisado e a energia livre resultante dessa hidrólise é transferida para a miosina, activando-a;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
111 | P á g i n a
O relaxamento do músculo resulta do transporte activo de iões Ca2+ de volta ao retículo sarcoplasmático, o qual requer ATP.
Rigor mortis
- Rigidez cadavérica que resulta da cessação da produção de ATP suficiente pelas células;
- Não ocorre desligamento das pontes – contracção contínua;
- Quando ocorre degradação dos tecidos – deixa de haver rigidez.
Nota:
1 Unidade Motora = 1 Neurónio + Grupo de fibras musculares associadas.
TRANSMISSÃO DOS IMPULSOS DOS NERVOS PARA AS FIBRAS MUSCULARES ESQUELÉTICAS: A JUNÇÃO NEUROMUSCULAR As fibras musculares esqueléticas são inervadas por fibras mielínicas grossas, originadas nos grandes neurónios motores da ponta anterior da medula espinhal. Cada uma dessas fibras nervosas em geral ramifica-se extensamente e estimula de três a várias centenas de fibras musculares esqueléticas. A terminação nervosa forma uma junção, chamada de junção neuromuscular, e o potencial de acção na fibra muscular propaga-se nas duas direcções, dirigindo-se para as suas extremidades. Com excepção de cerca de 2% das fibras musculares, só existe uma dessas junções em cada fibra muscular. Na terminação nervosa existem muitas mitocôndrias que fornecem energia, principalmente para a síntese do transmissor excitatório acetilcolina que, por sua vez, excita a fibra muscular. A acetilcolina é sintetizada no citoplasma da terminação, sendo rapidamente absorvida para o interior de numerosas e pequenas vesículas sinápticas; nas condições normais, existem cerca de 300.000 dessas vesículas em cada terminação do axónio de placa motora. Fixada à matriz da lâmina basal existe grande quantidade da enzima acetilcolinesterase, que é capaz de destruir a acetilcolina.
A junção neuromuscular é a sinapse entre o terminal do neurónio motor e a fibra muscular que este controla;
Os potenciais de acção ao chegarem ao terminal do axónio causam a libertação de acetilcolina na fenda sináptica. A contracção muscular é accionada pela ligação da acetilcolina a receptores nicotínicos musculares.
Secreção de acetilcolina pelas terminações nervosas Quando um impulso nervoso invade a junção neuromuscular, cerca de 300 vesículas de acetilcolina são libertadas pelas terminações axónicas no sulco sináptico. Existem na superfície interna da membrana neural barras densas lineares. De cada lado de uma barra densa existem partículas proteicas que atravessam toda a membrana, e que são

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
112 | P á g i n a
consideradas como formando canais de cálcio dependentes da voltagem. Quando o potencial de acção se propaga por toda a terminação, esses canais abrem-se, permitindo a difusão de grande quantidade de cálcio para o interior da terminação. Os iões cálcio, por sua vez, exercem influência atractiva sobre as vesículas de acetilcolina, puxando-as para a membrana neural adjacente às barras densas. Algumas dessas vesículas fundem-se com a membrana neural e esvaziam seu conteúdo de acetilcolina no sulco sináptico pelo mecanismo de exocitose. O estímulo efectivo para fazer com que a acetilcolina seja libertada das vesículas é o
influxo de iões cálcio; O esvaziamento das vesículas ocorre na membrana adjacente às barras densas.
Efeito da acetilcolina para abrir os canais iónicos dependentes da acetilcolina Os receptores são canais iónicos dependentes da acetilcolina, localizados, na sua quase totalidade, próximo à entrada das pregas subneurais, situadas imediatamente abaixo da área das barras densas, onde a acetilcolina é liberada na fenda sináptica. Cada receptor é um grande complexo proteico, com peso molecular total de 275.000. O complexo é formado por cinco subunidades proteicas, que atravessam toda a espessura da membrana, uma ao lado da outra, formando um círculo que circunda um canal tubular. Esse canal permanece contraído até que a acetilcolina se fixe a uma de suas subunidades → provoca alteração conformacional → abertura do canal. Quando aberto, o canal de acetilcolina tem diâmetro de cerca de 0,65 nm, suficientemente grande para permitir a passagem de todos os iões positivos importantes — sódio (Na+), potássio (K+) e cálcio (Ca2+) — com muita facilidade. Por outro lado, os iões negativos, como os iões cloreto, não passam por ele, devido às fortes cargas negativas presentes em sua abertura externa. Contudo, na prática, uma quantidade maior de iões sódio do que de qualquer outro ião passa pelos canais de acetilcolina, por duas razões. Só existem dois iões positivos em
concentração suficientemente alta para terem importância: os iões sódio, no líquido extracelular, e os iões potássio, no líquido intracelular;
O potencial muito negativo vigente na face interna da membrana muscular, de cerca de - 80 a - 90 mV, puxa os iões sódio, com carga positiva, para o interior da fibra, ao mesmo tempo que impede o efluxo dos iões potássio, quando estes tentam sair.
Por conseguinte, o resultado efectivo da abertura dos canais dependentes da acetilcolina é o de permitir a passagem de grande número de iões sódio para o interior da fibra, carregando com eles muitas cargas positivas. Isso gera um potencial local no interior da fibra, chamado de potencial da placa, que leva a um potencial de acção na membrana muscular, produzindo, assim, a contracção muscular. O "potencial da placa" e a excitação da fibra muscular esquelética O influxo abrupto dos iões sódio para o interior da fibra muscular, consequente à abertura dos canais de acetilcolina, faz com que o potencial de membrana, na área localizada da placa motora, varie, em direcção à positividade, por até 50 a 75 mV, gerando um potencial locai,
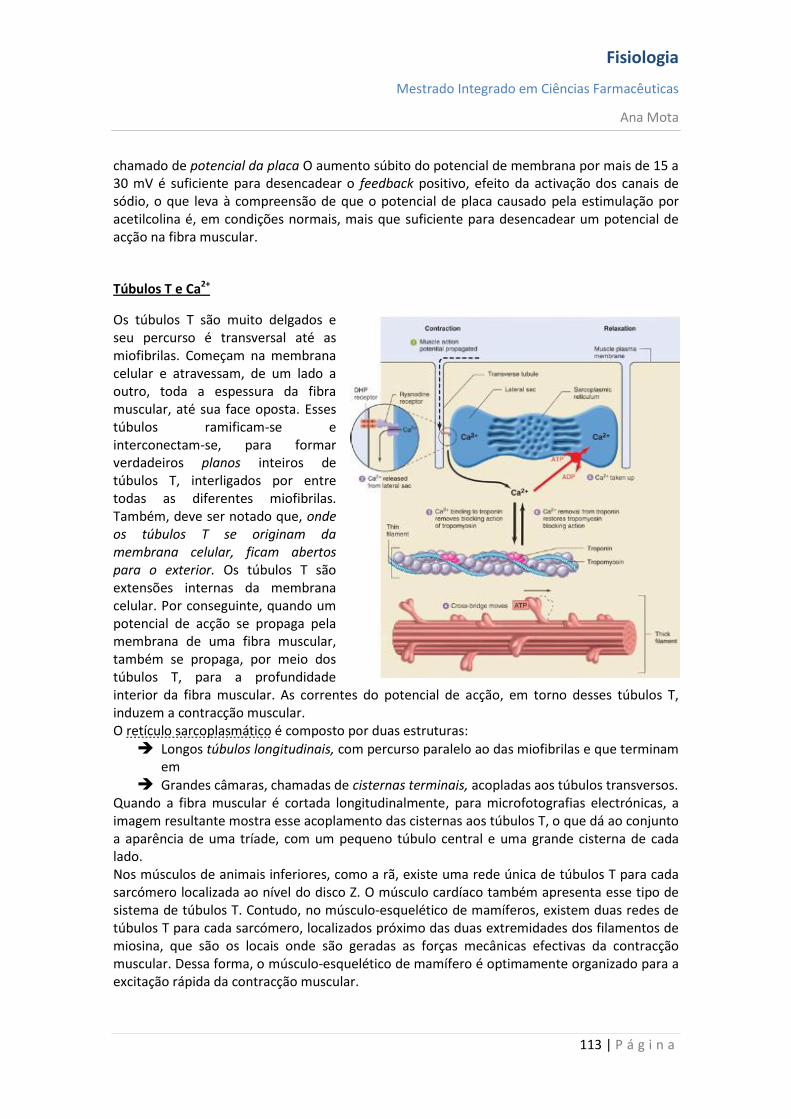
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
113 | P á g i n a
chamado de potencial da placa O aumento súbito do potencial de membrana por mais de 15 a 30 mV é suficiente para desencadear o feedback positivo, efeito da activação dos canais de sódio, o que leva à compreensão de que o potencial de placa causado pela estimulação por acetilcolina é, em condições normais, mais que suficiente para desencadear um potencial de acção na fibra muscular.
Túbulos T e Ca2+
Os túbulos T são muito delgados e seu percurso é transversal até as miofibrilas. Começam na membrana celular e atravessam, de um lado a outro, toda a espessura da fibra muscular, até sua face oposta. Esses túbulos ramificam-se e interconectam-se, para formar verdadeiros planos inteiros de túbulos T, interligados por entre todas as diferentes miofibrilas. Também, deve ser notado que, onde os túbulos T se originam da membrana celular, ficam abertos para o exterior. Os túbulos T são extensões internas da membrana celular. Por conseguinte, quando um potencial de acção se propaga pela membrana de uma fibra muscular, também se propaga, por meio dos túbulos T, para a profundidade interior da fibra muscular. As correntes do potencial de acção, em torno desses túbulos T, induzem a contracção muscular. O retículo sarcoplasmático é composto por duas estruturas: Longos túbulos longitudinais, com percurso paralelo ao das miofibrilas e que terminam
em Grandes câmaras, chamadas de cisternas terminais, acopladas aos túbulos transversos.
Quando a fibra muscular é cortada longitudinalmente, para microfotografias electrónicas, a imagem resultante mostra esse acoplamento das cisternas aos túbulos T, o que dá ao conjunto a aparência de uma tríade, com um pequeno túbulo central e uma grande cisterna de cada lado. Nos músculos de animais inferiores, como a rã, existe uma rede única de túbulos T para cada sarcómero localizada ao nível do disco Z. O músculo cardíaco também apresenta esse tipo de sistema de túbulos T. Contudo, no músculo-esquelético de mamíferos, existem duas redes de túbulos T para cada sarcómero, localizados próximo das duas extremidades dos filamentos de miosina, que são os locais onde são geradas as forças mecânicas efectivas da contracção muscular. Dessa forma, o músculo-esquelético de mamífero é optimamente organizado para a excitação rápida da contracção muscular.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
114 | P á g i n a
Uma das características especiais do retículo sarcoplasmático é que ele contém iões cálcio em concentrações muito elevadas, e muitos desses iões são liberados quando o túbulo T adjacente é excitado. O potencial de acção do túbulo T produz fluxo de corrente para dentro das cisternas do retículo sarcoplasmático, onde se confinam com o túbulo T. Qualquer que seja a natureza desse sinal, ele provoca a abertura rápida de muitos canais de cálcio nas membranas das cisternas e nos túbulos longitudinais do retículo sarcoplasmático que as continuam. Esses canais permanecem abertos por poucos milissegundos; durante esse período, os iões cálcio, responsáveis pela contracção muscular, são libertados no sarcoplasma que banha as miofibrilas. Os iões cálcio que são, assim, liberados pelo retículo sarcoplasmático difundem-se até as miofibrilas adjacentes, onde vão se fixar fortemente à troponina C e isso induz à contracção muscular.
Nota:
O cálcio liga-se à troponina, esta liga-se à tropomiosina → actina activa – contracção
muscular;
O cálcio é indispensável para marcar o ciclo das pontes cruzadas.
Registo de contracções
Muitas características da contracção muscular podem ser especialmente demonstradas pela produção de abalos musculares isolados. Isso pode ser conseguido por estimulação breve do nervo que vai para o músculo ou pela passagem de estímulo eléctrico de curta duração pelo próprio músculo, o que produz contracção única e abrupta do músculo, que dura fracção de segundo. A resposta ao potencial de membrana inicia-se
um pouco depois do potencial de acção terminal.
A tensão é tanto maior quanto maior for o nº de
pontes estabelecidas entre a actina e a miosina e
é proporcional à concentração de cálcio. A partir de uma certa altura, o músculo começa a
relaxar, uma vez que o estímulo que causou a contracção desaparece – a acetilcolina é
degradada, a bomba de cálcio repõe os níveis de cálcio no retículo e a concentração de cálcio
no citoplasma diminui.
Basta retirar cálcio do citoplasma para que não se estabeleçam as pontes de ligação. O
desfasamento entre o processo de activação da contracção e a contracção
propriamente dita faz com que haja uma graduação na força de contracção.
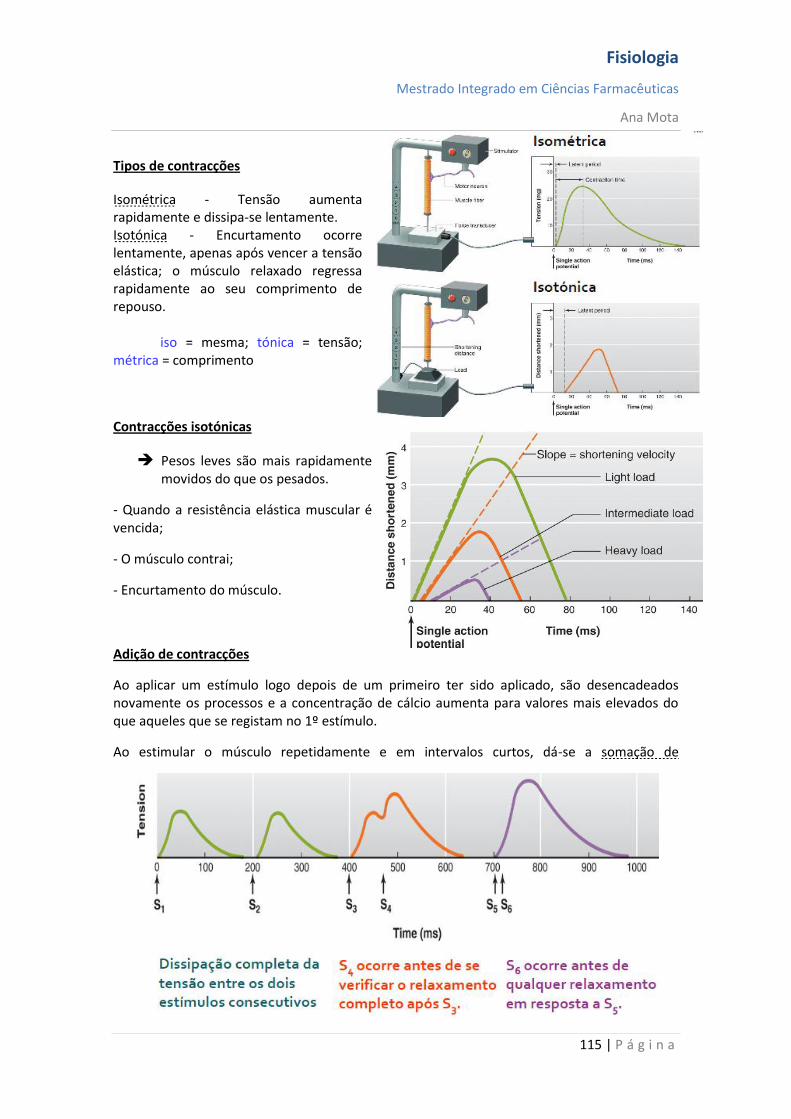
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
115 | P á g i n a
Tipos de contracções
Isométrica - Tensão aumenta rapidamente e dissipa-se lentamente. Isotónica - Encurtamento ocorre lentamente, apenas após vencer a tensão elástica; o músculo relaxado regressa rapidamente ao seu comprimento de repouso.
iso = mesma; tónica = tensão;
métrica = comprimento
Contracções isotónicas
Pesos leves são mais rapidamente movidos do que os pesados.
- Quando a resistência elástica muscular é vencida;
- O músculo contrai;
- Encurtamento do músculo.
Adição de contracções
Ao aplicar um estímulo logo depois de um primeiro ter sido aplicado, são desencadeados novamente os processos e a concentração de cálcio aumenta para valores mais elevados do que aqueles que se registam no 1º estímulo.
Ao estimular o músculo repetidamente e em intervalos curtos, dá-se a somação de

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
116 | P á g i n a
contracções, o que permite um aumento da força de contracção.
É a fase da actividade mecânica do músculo em que há adição de todas as contracções individuais estimuladas que fazem com que a contracção seja mais forte do que aquela que é provocada por um único estímulo.
Se essa frequência de estímulos for muito grande, ocorre tensão máxima do músculo e pode levar à situação de tétano – não é muito sustentável porque ao fim de algum tempo, o músculo vai perdendo força e entra num estado de fadiga (metabolismo da célula e a disponibilidade de ATP estão entre essas causas).
Tétano Muscular É o estado atingido quando ocorre estimulação contínua, isto é, somação de estímulos
com frequências elevadas em que pode ou não ocorrer relaxamento do músculo.
Resulta da somação de ondas múltiplas;
A tetania incompleta ocorrer quando a frequência dos potenciais de acção é suficientemente baixa para permitir o relaxamento parcial das fibras musculares;
A tetania completa ocorre quando a frequência dos potenciais de acção é suficientemente alta para não haver relaxamento das fibras musculares.
Fadiga muscular
Contracções fortes e prolongadas de um músculo levam ao estado bem conhecido de fadiga muscular. Estudos em atletas mostraram que a fadiga muscular aumenta quase em proporção directa

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
117 | P á g i n a
com a intensidade da depleção do glicogénio muscular. Por conseguinte, a maior parte da fadiga resulta, com muita probabilidade, simplesmente da incapacidade dos processos contrácteis e metabólicos das fibras musculares de produzir, de modo contínuo, a mesma quantidade de trabalho. Todavia, a transmissão do sinal neural através da placa motora pode ficar diminuída após actividade muscular prolongada, o que diminuiria ainda mais a contracção muscular. A interrupção do fluxo sanguíneo para um músculo em contracção produz fadiga muscular quase total em um minuto ou pouco mais, devido à perda óbvia do fornecimento de nutrientes - em especial, a falta de oxigénio.
Relação Comprimento/Força
À direita são apresentados diferentes graus de superposição dos filamentos de actina e de miosina, em diversos comprimentos do sarcómero. Nesse esquema, o ponto D marca o afastamento do filamento de actina além da extremidade do filamento de miosina, sem qualquer superposição. Nesse ponto, a tensão desenvolvida pelo músculo activado é zero. Em seguida, à medida que o sarcómero se encurta e os filamentos de actina começam a se sobrepor aos de miosina, começa o desenvolvimento de tensão, com aumento progressivo, até que o comprimento do sarcómero diminua para cerca de 2,2 μm. Nesse ponto, o filamento de actina já se sobrepôs a todas as pontes cruzadas do filamento de miosina, mas ainda não atingiu o centro desse filamento. Com encurtamento ainda maior, o sarcómero mantém tensão máxima até o ponto B, com comprimento desse sarcómero de cerca de 2,0 μm. Nesse ponto, as extremidades dos filamentos de actina começam a se sobrepor umas às outras, além de estarem sobrepostas aos filamentos de miosina. Quando o comprimento do sarcómero cai de 2,0 μm para cerca de 1,65 micrómetros, no ponto A, a força da contracção diminui. É nesse ponto que os dois discos Z do sarcómero entram em contacto com as extremidades dos filamentos de miosina. Então, à medida que a contracção prossegue, com comprimentos do sarcómero ainda menores, as extremidades dos filamentos de miosina são dobradas e a força da contracção diminui abrupta e aceleradamente. A contracção máxima ocorre quando existe grau máximo de sobreposição entre os
filamentos de actina e as pontes cruzadas dos filamentos de miosina;
Quanto maior for o número de pontes cruzadas a puxar o filamento de actina, maior será a força de contracção.
Fontes de Energia
ATP – produzido através da célula:

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
118 | P á g i n a
Glicose:
Do sangue; Reservas de glicogénio do músculo; Algumas situações da degradação de ácidos gordos.
A partir da glicose podem ocorrer 2 processos:
Glicólise
- Pouco rentável;
- Origina ácido láctico que quando se acumula, altera o pH da célula, resultando numa disfunção – diminuição da actividade muscular.
Fosforilação oxidativa
- Rentável e necessita de O2;
- Produção de 36 ATP que são armazenados/transportados para os reservatórios de fosfocreatina, que é a reserva de energia (ATP) muscular.
A capacidade de produzir ATP pela via oxidativa é suportada pela disponibilidade de oxigénio que se encontra armazenado ligado à Mioglobina. A mioglobina fixa o O2 dentro da célula. Quanto maior for a quantidade de O2 mais pode se utilizar na via oxidativa, para obtenção de mais energia.
Utilização das Fontes:
Em exercício de curta duração (30 min):
- Fonte primária de energia é a glicose do sangue;
- 2ª fonte – ácidos gordos;
- 3ª fonte – gorduras do músculo.
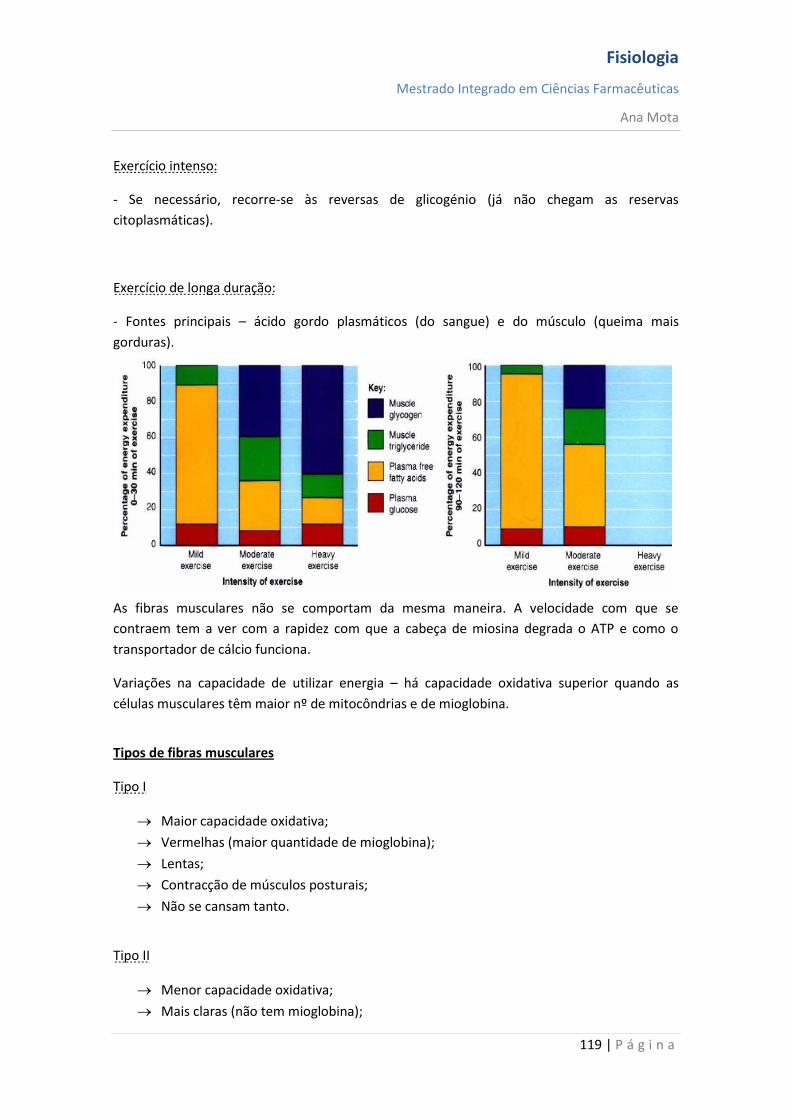
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
119 | P á g i n a
Exercício intenso:
- Se necessário, recorre-se às reversas de glicogénio (já não chegam as reservas
citoplasmáticas).
Exercício de longa duração:
- Fontes principais – ácido gordo plasmáticos (do sangue) e do músculo (queima mais
gorduras).
As fibras musculares não se comportam da mesma maneira. A velocidade com que se
contraem tem a ver com a rapidez com que a cabeça de miosina degrada o ATP e como o
transportador de cálcio funciona.
Variações na capacidade de utilizar energia – há capacidade oxidativa superior quando as
células musculares têm maior nº de mitocôndrias e de mioglobina.
Tipos de fibras musculares
Tipo I
Maior capacidade oxidativa;
Vermelhas (maior quantidade de mioglobina);
Lentas;
Contracção de músculos posturais;
Não se cansam tanto.
Tipo II
Menor capacidade oxidativa;
Mais claras (não tem mioglobina);
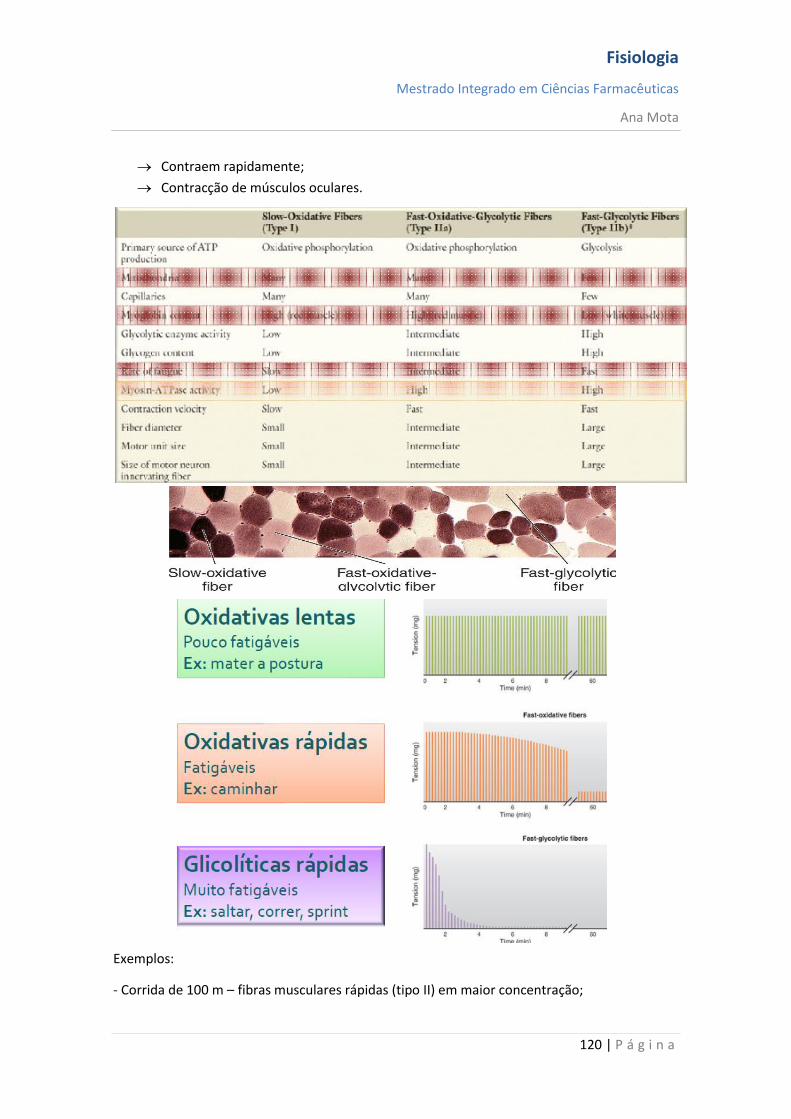
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
120 | P á g i n a
Contraem rapidamente;
Contracção de músculos oculares.
Exemplos:
- Corrida de 100 m – fibras musculares rápidas (tipo II) em maior concentração;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
121 | P á g i n a
- Maratona – fibras musculares lentas (tipo I) em maior concentração.
Tensão muscular: factores
Tensão desenvolvida por cada fibra - Frequências dos potenciais de acção; - Comprimento da fibra; - Diâmetro da fibra; - Fadiga.
Número de fibras activas - Número de fibras por unidade motora; - Número de unidades motoras activas.
Gradação da actividade muscular
Mecânica corporal
Gradação da actividade muscular - Tamanho das unidades motoras activadas (quanto maior for a unidade motora, maior o nº de unidades motoras); - Número de unidades motoras activadas (quanto maior o nº de unidades motoras, maior o grau de contracção mucular); - Frequência do estímulo (maior frequência → somação de contracções); - Recrutamento de unidades motoras.
Comprimento basal óptimo – contracção seja máxima. Os fusos musculares (com neurónios sensitivos) controlam este factor.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
122 | P á g i n a
Contracções breves protegem de fadiga – contracções breves protegem os músculos
da fadiga e há uma actividade muscular mais duradoura.
Músculo cardíaco
Bioquímica da Contracção
Ciclo das Pontes Cruzadas
Semelhante Músculo-Esquelético Actividade ATPásica da Miosina
Lenta Síntese de ATP
Metabolismo aeróbio (O2) Automaticidade
Células Pacemaker
Tecido Nodal Regulação
Sistema Nervoso Autónomo
Estrias transversais;
Discos intercalares – estruturas com junções GAP que estabelecem a comunicação
entre as células. No músculo esquelético não há esta comunicação, cada célula é
independente.
O coração é constituído de três tipos principais de músculo cardíaco: músculo auricular, músculo ventricular e fibras musculares condutoras e excitatórias especializadas. Os tipos auricular e ventricular de músculo contraem-se da mesma maneira que o músculo-esquelético, excepto que a duração da contracção é muito maior. Por outro lado, as fibras condutoras e excitatórias especializadas contraem-se apenas fracamente, por conterem poucas fibrilas contrateis; em vez disso, elas apresentam ritmicidade e velocidades variáveis de condução, proporcionando um sistema excitatório para o coração e um sistema de transmissão para a condução controlada do sinal excitatório cardíaco por todo o coração. O músculo cardíaco é estriado; O músculo cardíaco tem miofibrilas típicas, que contêm filamentos de actina e miasma
quase idênticos aos encontrados no músculo-esquelético, e esses filamentos interdigitam-se e deslizam uns sobre os outros durante o processo de contracção da mesma maneira como ocorre nos músculos esqueléticos
Músculo cardíaco: potencial de acção
O potencial de membrana em repouso do músculo cardíaco normal é de aproximadamente -85 a -95 milivolts (mV) e de cerca de -90 a -100 mV nas fibras de condução especializadas, as fibras de Purkinje.
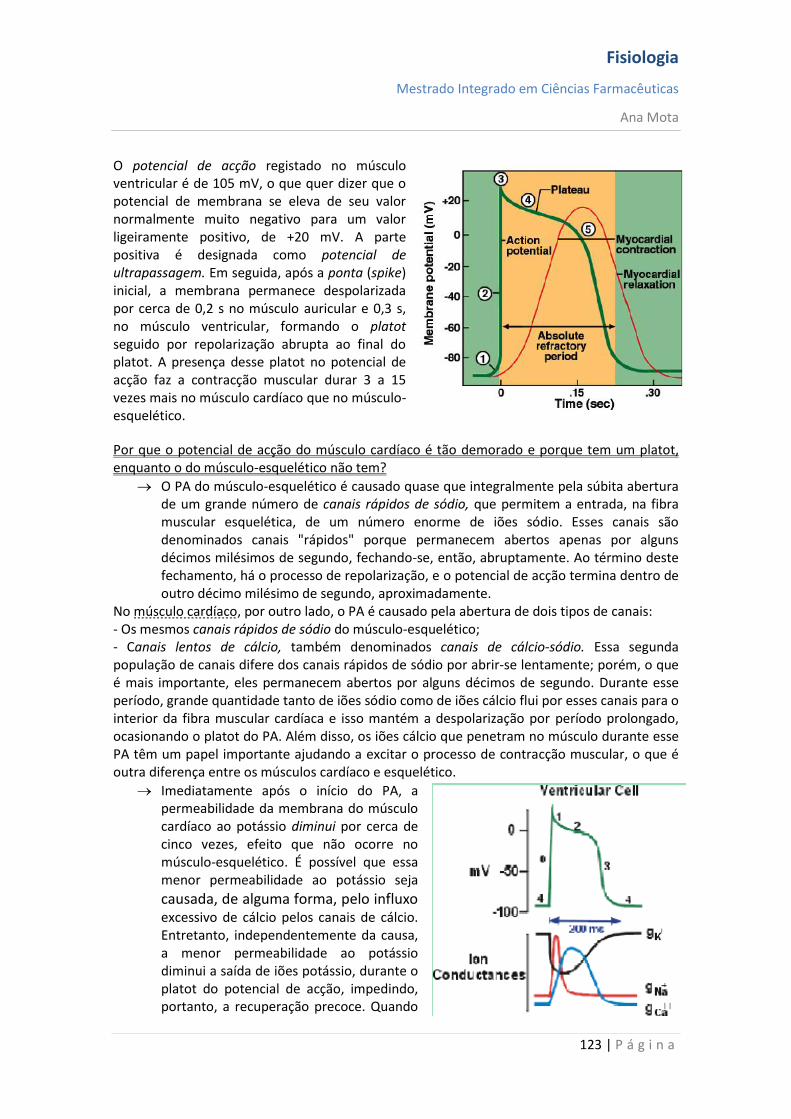
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
123 | P á g i n a
O potencial de acção registado no músculo ventricular é de 105 mV, o que quer dizer que o potencial de membrana se eleva de seu valor normalmente muito negativo para um valor ligeiramente positivo, de +20 mV. A parte positiva é designada como potencial de ultrapassagem. Em seguida, após a ponta (spike) inicial, a membrana permanece despolarizada por cerca de 0,2 s no músculo auricular e 0,3 s, no músculo ventricular, formando o platot seguido por repolarização abrupta ao final do platot. A presença desse platot no potencial de acção faz a contracção muscular durar 3 a 15 vezes mais no músculo cardíaco que no músculo-esquelético.
Por que o potencial de acção do músculo cardíaco é tão demorado e porque tem um platot, enquanto o do músculo-esquelético não tem?
O PA do músculo-esquelético é causado quase que integralmente pela súbita abertura de um grande número de canais rápidos de sódio, que permitem a entrada, na fibra muscular esquelética, de um número enorme de iões sódio. Esses canais são denominados canais "rápidos" porque permanecem abertos apenas por alguns décimos milésimos de segundo, fechando-se, então, abruptamente. Ao término deste fechamento, há o processo de repolarização, e o potencial de acção termina dentro de outro décimo milésimo de segundo, aproximadamente.
No músculo cardíaco, por outro lado, o PA é causado pela abertura de dois tipos de canais: - Os mesmos canais rápidos de sódio do músculo-esquelético; - Canais lentos de cálcio, também denominados canais de cálcio-sódio. Essa segunda população de canais difere dos canais rápidos de sódio por abrir-se lentamente; porém, o que é mais importante, eles permanecem abertos por alguns décimos de segundo. Durante esse período, grande quantidade tanto de iões sódio como de iões cálcio flui por esses canais para o interior da fibra muscular cardíaca e isso mantém a despolarização por período prolongado, ocasionando o platot do PA. Além disso, os iões cálcio que penetram no músculo durante esse PA têm um papel importante ajudando a excitar o processo de contracção muscular, o que é outra diferença entre os músculos cardíaco e esquelético.
Imediatamente após o início do PA, a permeabilidade da membrana do músculo cardíaco ao potássio diminui por cerca de cinco vezes, efeito que não ocorre no músculo-esquelético. É possível que essa menor permeabilidade ao potássio seja
causada, de alguma forma, pelo influxo excessivo de cálcio pelos canais de cálcio. Entretanto, independentemente da causa, a menor permeabilidade ao potássio diminui a saída de iões potássio, durante o platot do potencial de acção, impedindo, portanto, a recuperação precoce. Quando

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
124 | P á g i n a
os canais lentos de cálcio-sódio fecham ao fim de 0,2 a 0,3 s, interrompendo o influxo de iões cálcio e sódio, a permeabilidade da membrana ao potássio aumenta rapidamente, e a rápida perda de potássio pela fibra faz o potencial de membrana retornar a seu nível de repouso, terminando, assim, o PA.
Origem do Cálcio
Nos cardiomiócitos, o Ca2+ provém do meio extracelular, ao contrário do que acontece no músculo esquelético, em que o Ca2+ provém do retículo sarcoplasmático.
Músculo liso
O músculo liso de cada órgão difere da maioria dos
outros órgãos pelas suas dimensões físicas, pela
organização em feixes ou lâminas, pelas respostas aos
diferentes tipos de estímulos, pelas características da
enervação e pela função.
Proteínas contrácteis
CAM → Calmodulina (Homóloga funcional da Troponina C) MLCK → Cinase da Cadeia Leve de Miosina MP → Fosfatase da Miosina
Como é válido para o músculo-esquelético, o factor desencadeante na maioria das

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
125 | P á g i n a
contracções do músculo liso é um aumento da concentração intracelular de iões cálcio. Esse aumento pode ser causado por estimulação da fibra nervosa para a fibra muscular lisa, por estimulação hormonal, por estiramento da fibra e, até mesmo, por alteração do ambiente químico da fibra. Todavia, o músculo liso não contém troponina, a proteína reguladora que é activada pelos iões cálcio para promover a contracção do músculo-esquelético. Pelo contrário, a contracção do músculo liso é activada por um mecanismo inteiramente diferente. Combinação dos iões cálcio com a "calmodulina"— Activação da cinase de miosina e fosforilação da cabeça da miosina Em lugar da troponina, as células do músculo liso contêm grande quantidade de outra proteína reguladora, denominada calmodulina. Embora essa proteína seja semelhante à troponina, por reagir com quatro iões cálcio, difere no modo como desencadeia a contracção. A CAM activa as pontes cruzadas de miosina. Essa activação e a contracção subsequente ocorrem nas seguintes etapas: 1. Os iões cálcio fixam-se a CAM; 2. A combinação CAM-Ca2+ fixa-se, então, e activa a MLCK (cinase de miosina), uma enzima fosforilativa. 3. Uma das cadeias leves de cada cabeça de miosina, chamada de cadeia reguladora, fica fosforilada, em resposta à cinase de miosina. Quando essa cadeia não está fosforilada, não ocorre o ciclo de fixação-desligamento da cabeça. Mas quando a cadeia reguladora está fosforilada, a cabeça adquire a capacidade de se fixar ao filamento de actina e a seguir por todo o processo do ciclo, o que resulta em contracção muscular.
Fim da contracção — papel da " fosfatase da miosina" (MP) Quando a concentração de cálcio cai abaixo do seu nível crítico, todas as etapas descritas acima invertem-se de modo automático, excepto pela fosforilação da cabeça de miosina. Essa inversão exige acção de outra enzima, a fosfatase da miosina, que remove o fosfato da cadeia leve reguladora. Então, o ciclo é interrompido e cessa a contracção. O tempo necessário para o relaxamento da contracção muscular é, portanto, determinado, em grande parte, pela quantidade de fosfatase da miosina activa na célula. Desfosforilação da miosina - MP
Pontes cruzadas
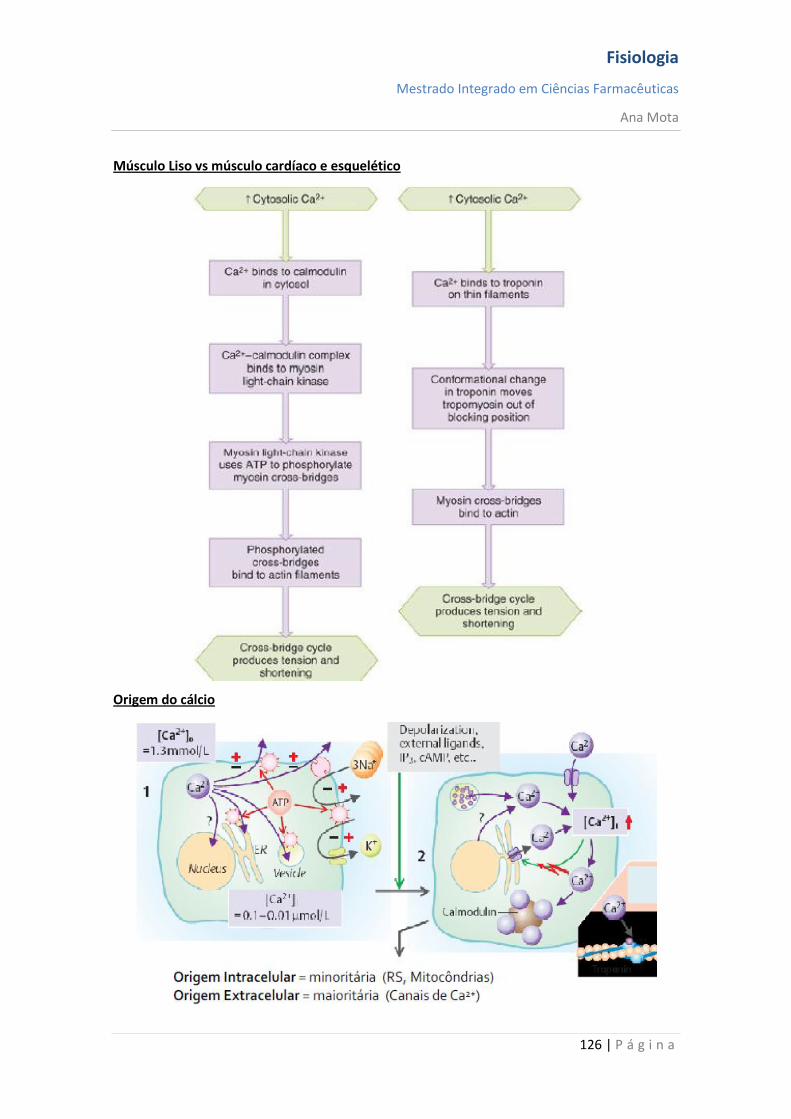
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
126 | P á g i n a
Músculo Liso vs músculo cardíaco e esquelético
Origem do cálcio
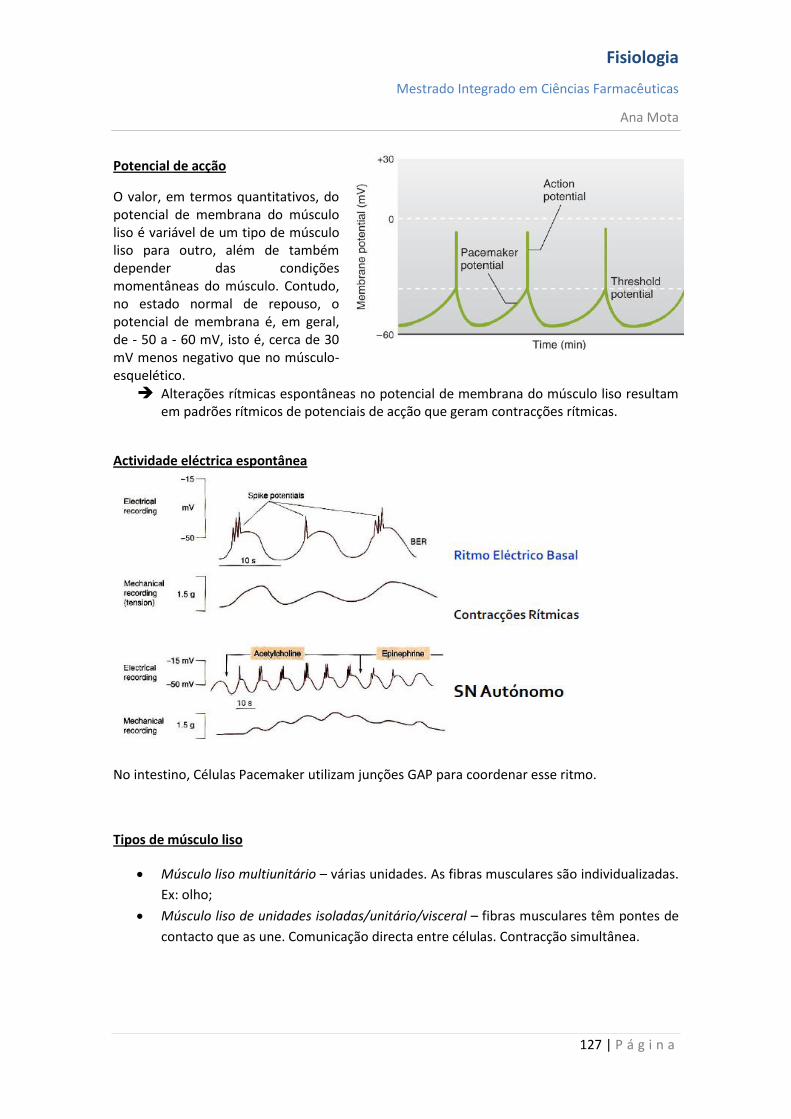
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
127 | P á g i n a
Potencial de acção
O valor, em termos quantitativos, do potencial de membrana do músculo liso é variável de um tipo de músculo liso para outro, além de também depender das condições momentâneas do músculo. Contudo, no estado normal de repouso, o potencial de membrana é, em geral, de - 50 a - 60 mV, isto é, cerca de 30 mV menos negativo que no músculo-esquelético. Alterações rítmicas espontâneas no potencial de membrana do músculo liso resultam
em padrões rítmicos de potenciais de acção que geram contracções rítmicas.
Actividade eléctrica espontânea
No intestino, Células Pacemaker utilizam junções GAP para coordenar esse ritmo.
Tipos de músculo liso
Músculo liso multiunitário – várias unidades. As fibras musculares são individualizadas.
Ex: olho;
Músculo liso de unidades isoladas/unitário/visceral – fibras musculares têm pontes de
contacto que as une. Comunicação directa entre células. Contracção simultânea.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
128 | P á g i n a
Regulação do músculo liso
Actividade eléctrica espontânea (visceral)
Neurotransmissores libertados pelo SNA - NA, ACH, ATP, VIP, NO
Hormonas - Adrenalina, Angiotensina II
Estiramento (visceral)
Alterações locais na composição do LEC - Agentes parácrinos - Acidez, O2, osmolaridade, concentração de iões
Nota:
O óxido nítrico (NO) é um dos agentes parácrinos mais comummente encontrados que produz
relaxamento dos músculos lisos.
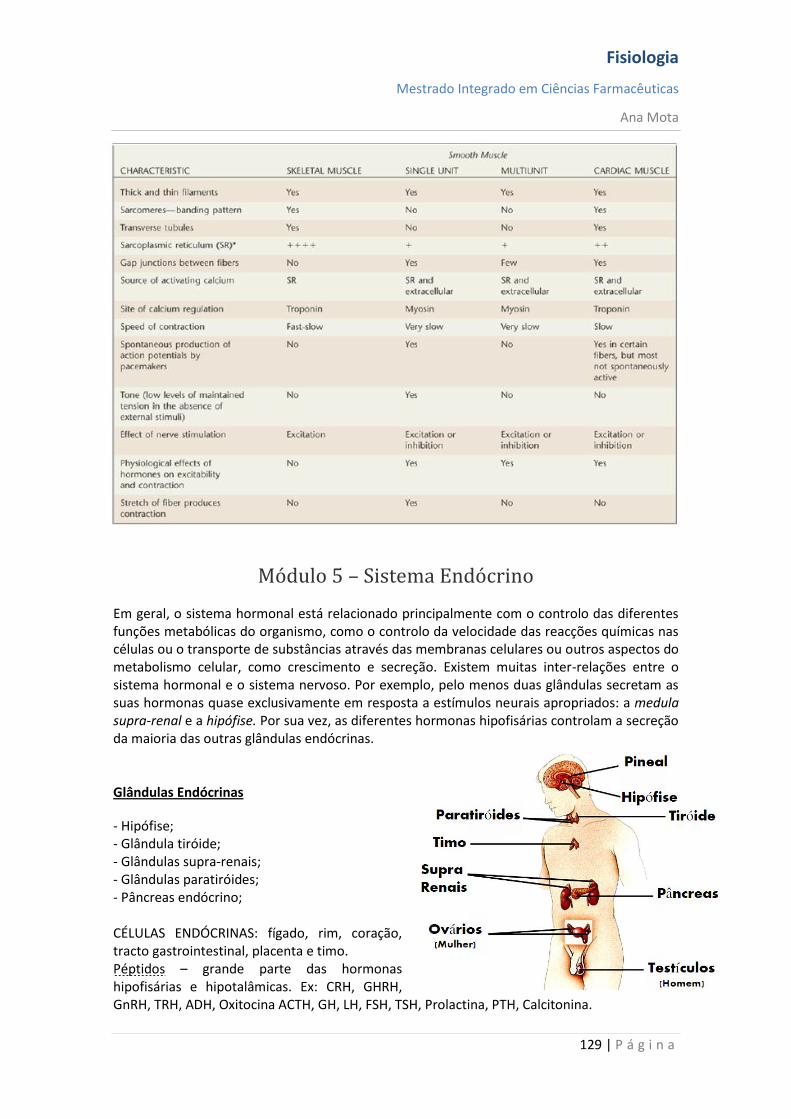
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
129 | P á g i n a
Módulo 5 – Sistema Endócrino
Em geral, o sistema hormonal está relacionado principalmente com o controlo das diferentes funções metabólicas do organismo, como o controlo da velocidade das reacções químicas nas células ou o transporte de substâncias através das membranas celulares ou outros aspectos do metabolismo celular, como crescimento e secreção. Existem muitas inter-relações entre o sistema hormonal e o sistema nervoso. Por exemplo, pelo menos duas glândulas secretam as suas hormonas quase exclusivamente em resposta a estímulos neurais apropriados: a medula supra-renal e a hipófise. Por sua vez, as diferentes hormonas hipofisárias controlam a secreção da maioria das outras glândulas endócrinas.
Glândulas Endócrinas
- Hipófise; - Glândula tiróide; - Glândulas supra-renais; - Glândulas paratiróides; - Pâncreas endócrino; CÉLULAS ENDÓCRINAS: fígado, rim, coração, tracto gastrointestinal, placenta e timo. Péptidos – grande parte das hormonas hipofisárias e hipotalâmicas. Ex: CRH, GHRH, GnRH, TRH, ADH, Oxitocina ACTH, GH, LH, FSH, TSH, Prolactina, PTH, Calcitonina.
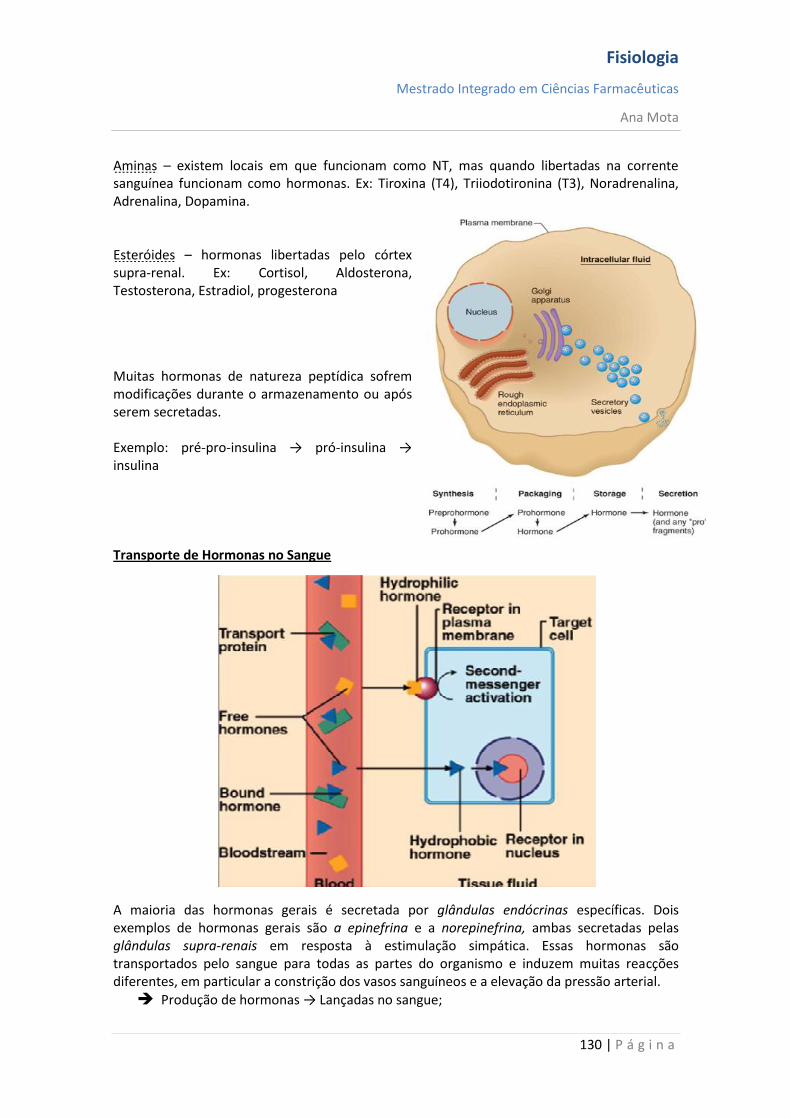
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
130 | P á g i n a
Aminas – existem locais em que funcionam como NT, mas quando libertadas na corrente sanguínea funcionam como hormonas. Ex: Tiroxina (T4), Triiodotironina (T3), Noradrenalina, Adrenalina, Dopamina. Esteróides – hormonas libertadas pelo córtex supra-renal. Ex: Cortisol, Aldosterona, Testosterona, Estradiol, progesterona
Muitas hormonas de natureza peptídica sofrem modificações durante o armazenamento ou após serem secretadas. Exemplo: pré-pro-insulina → pró-insulina → insulina
Transporte de Hormonas no Sangue
A maioria das hormonas gerais é secretada por glândulas endócrinas específicas. Dois exemplos de hormonas gerais são a epinefrina e a norepinefrina, ambas secretadas pelas glândulas supra-renais em resposta à estimulação simpática. Essas hormonas são transportados pelo sangue para todas as partes do organismo e induzem muitas reacções diferentes, em particular a constrição dos vasos sanguíneos e a elevação da pressão arterial. Produção de hormonas → Lançadas no sangue;
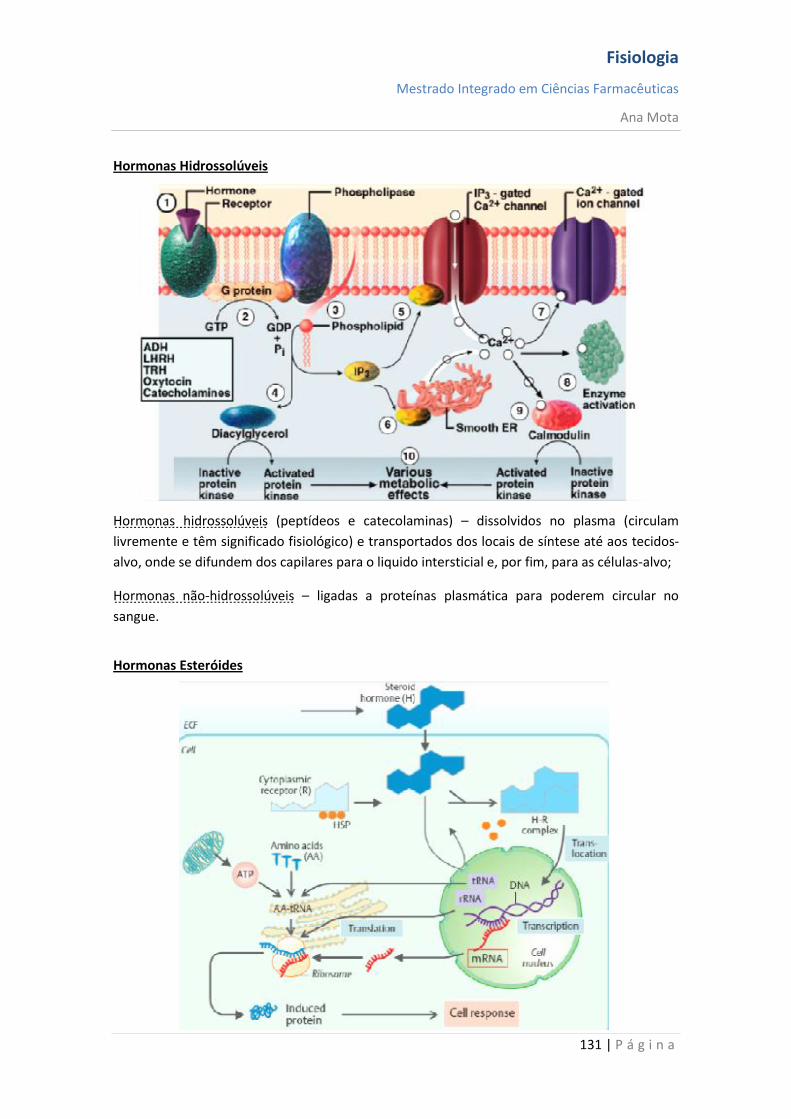
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
131 | P á g i n a
Hormonas Hidrossolúveis
Hormonas hidrossolúveis (peptídeos e catecolaminas) – dissolvidos no plasma (circulam
livremente e têm significado fisiológico) e transportados dos locais de síntese até aos tecidos-
alvo, onde se difundem dos capilares para o liquido intersticial e, por fim, para as células-alvo;
Hormonas não-hidrossolúveis – ligadas a proteínas plasmática para poderem circular no
sangue.
Hormonas Esteróides
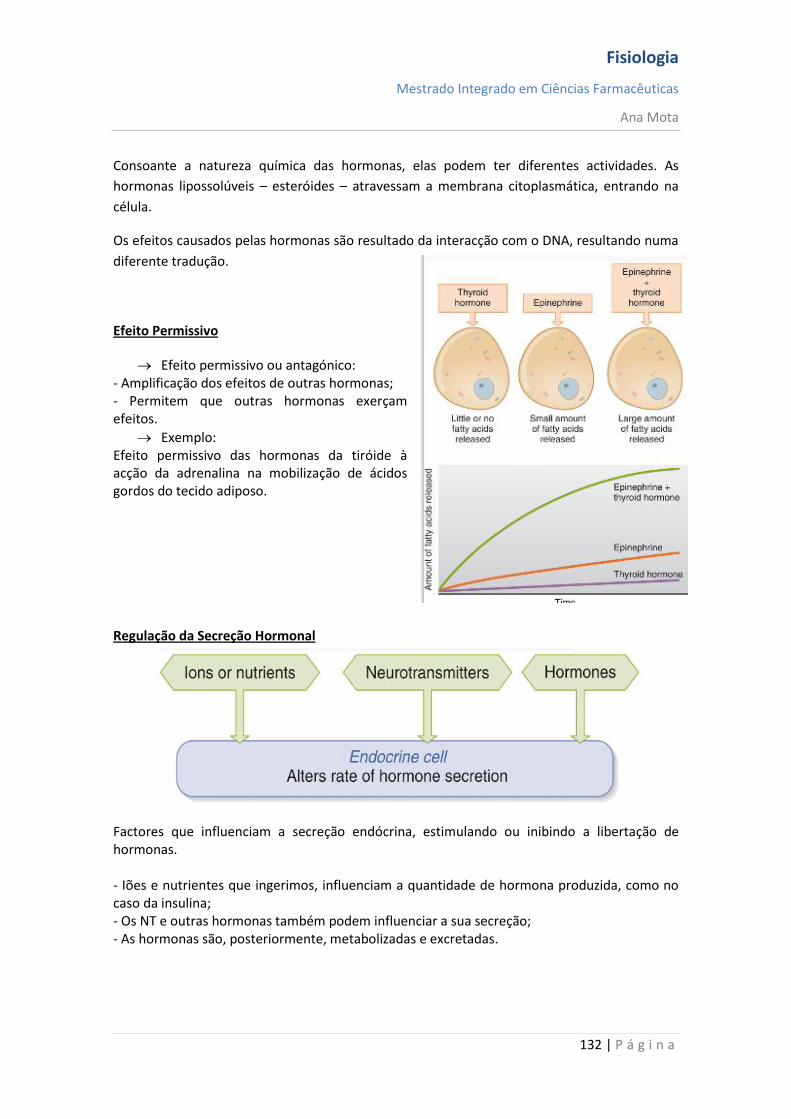
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
132 | P á g i n a
Consoante a natureza química das hormonas, elas podem ter diferentes actividades. As
hormonas lipossolúveis – esteróides – atravessam a membrana citoplasmática, entrando na
célula.
Os efeitos causados pelas hormonas são resultado da interacção com o DNA, resultando numa
diferente tradução.
Efeito Permissivo
Efeito permissivo ou antagónico: - Amplificação dos efeitos de outras hormonas; - Permitem que outras hormonas exerçam efeitos.
Exemplo: Efeito permissivo das hormonas da tiróide à acção da adrenalina na mobilização de ácidos gordos do tecido adiposo.
Regulação da Secreção Hormonal
Factores que influenciam a secreção endócrina, estimulando ou inibindo a libertação de hormonas. - Iões e nutrientes que ingerimos, influenciam a quantidade de hormona produzida, como no caso da insulina; - Os NT e outras hormonas também podem influenciar a sua secreção; - As hormonas são, posteriormente, metabolizadas e excretadas.
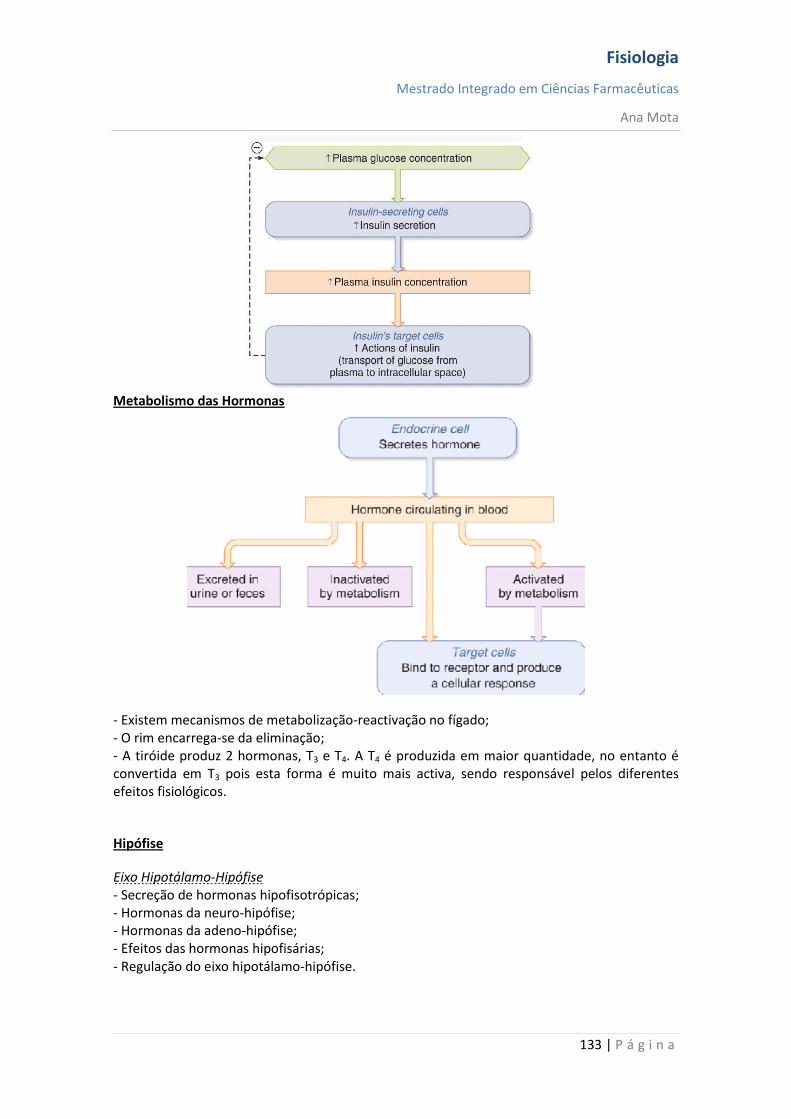
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
133 | P á g i n a
Metabolismo das Hormonas
- Existem mecanismos de metabolização-reactivação no fígado; - O rim encarrega-se da eliminação; - A tiróide produz 2 hormonas, T3 e T4. A T4 é produzida em maior quantidade, no entanto é convertida em T3 pois esta forma é muito mais activa, sendo responsável pelos diferentes efeitos fisiológicos.
Hipófise
Eixo Hipotálamo-Hipófise - Secreção de hormonas hipofisotrópicas; - Hormonas da neuro-hipófise; - Hormonas da adeno-hipófise; - Efeitos das hormonas hipofisárias; - Regulação do eixo hipotálamo-hipófise.
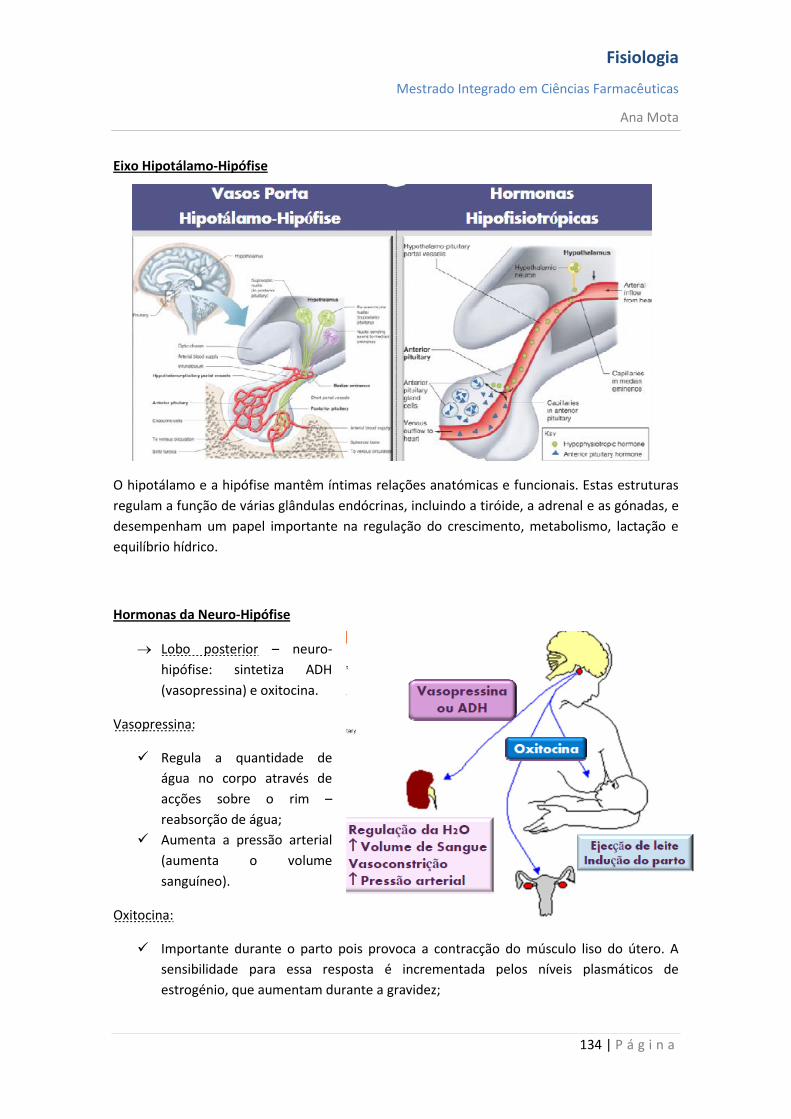
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
134 | P á g i n a
Eixo Hipotálamo-Hipófise
O hipotálamo e a hipófise mantêm íntimas relações anatómicas e funcionais. Estas estruturas
regulam a função de várias glândulas endócrinas, incluindo a tiróide, a adrenal e as gónadas, e
desempenham um papel importante na regulação do crescimento, metabolismo, lactação e
equilíbrio hídrico.
Hormonas da Neuro-Hipófise
Lobo posterior – neuro-
hipófise: sintetiza ADH
(vasopressina) e oxitocina.
Vasopressina:
Regula a quantidade de
água no corpo através de
acções sobre o rim –
reabsorção de água;
Aumenta a pressão arterial
(aumenta o volume
sanguíneo).
Oxitocina:
Importante durante o parto pois provoca a contracção do músculo liso do útero. A
sensibilidade para essa resposta é incrementada pelos níveis plasmáticos de
estrogénio, que aumentam durante a gravidez;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
135 | P á g i n a
Importante na amamentação pois estimula as células mioepiteliais dos alvéolos das
glândulas mamárias e, consequentente, a ejecção de leite.
Lobo anterior – adenohipófise: sintetiza hormonas pépticas.
Hormonas Hipofisiotrópicas
Hormonas da Adenohipófise

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
136 | P á g i n a
Alvos das Hormonas da Adenohipófise
Regulação do Eixo Hipotálamo-Hipófise
Sem excepção, a velocidade de secreção de todos as hormonas é controlada, com muita precisão, por um sistema de controlo interno. Na maioria dos casos, esse controlo é exercido por meio de um mecanismo de feedback negativo, que funciona da seguinte maneira: 1. A glândula endócrina tem tendência natural para secretar a sua hormona em excesso; 2. Devido a essa tendência, a hormona exerce cada vez mais o seu efeito de controlo sobre o órgão-alvo. 3. O órgão-alvo desempenha, por sua vez, sua função. 4. Entretanto, quando a função é excessiva, algum factor relacionado a essa função passa a exercer, em geral, acção de feedback sobre a glândula endócrina, produzindo nela um efeito negativo, no sentido de reduzir a velocidade de secreção. Dessa maneira, a função da hormona é monitorizada, e essa informação, por sua vez, actua como controle por feedback negativo sobre a velocidade de secreção da glândula.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
137 | P á g i n a
Glândula Tiróide
- Triiodotironina (T3); - Tetraidotironina (T4); - Calcitonina.
A glândula tiróide localiza-se no pescoço, por baixo da laringe rodeando a traqueia. É constituída por 2 lóbulos ligados por uma ponte, o istmo. É uma glândula muito bem irrigada.
Hormonas da Tiróide
Tretraiodotironina (T4); Triiodotironina (T3).
A tiróide é constituída por um
grande número de folículos e cada folículo está envolvido por uma camada de células e
preenchido por material proteíco, chamado colóide.
São os folículos que produzem as hormonas T3 e T4.
O constituinte primário do colóide é uma grande glicoproteína, a tiroglobina, que contém as
hormonas da tiróide na sua molécula.
BOMBA DE IODETO (CAPTAÇÃO DE IODETO) A primeira etapa da síntese das hormonas tiróideias consiste no transporte dos iodetos do líquido extracelular para as células glandulares e os folículos da tiróide. A membrana basal da célula tiróideia tem capacidade específica de bombear activamente o iodeto para o interior da célula. Esse processo é denominado captação de iodeto. Na glândula normal, a bomba de iodeto concentra o iodeto por cerca de 30 vezes sua concentração no sangue. Todavia, quando a actividade da glândula tiróide se torna máxima, a relação entre essas concentrações pode aumentar por até 250 vezes.
Síntese de T3 e T4
1 - Oxidação do I- a I2, pela peroxidase da tiróide – membrana apical das células foliculares; 2 - Síntese de Tiroglobulina, nas células foliculares e secretada para o colóide por exocitose; 3-5 - Iodinação Tiroglobulina, ; 6 - Endocitose do colóide;
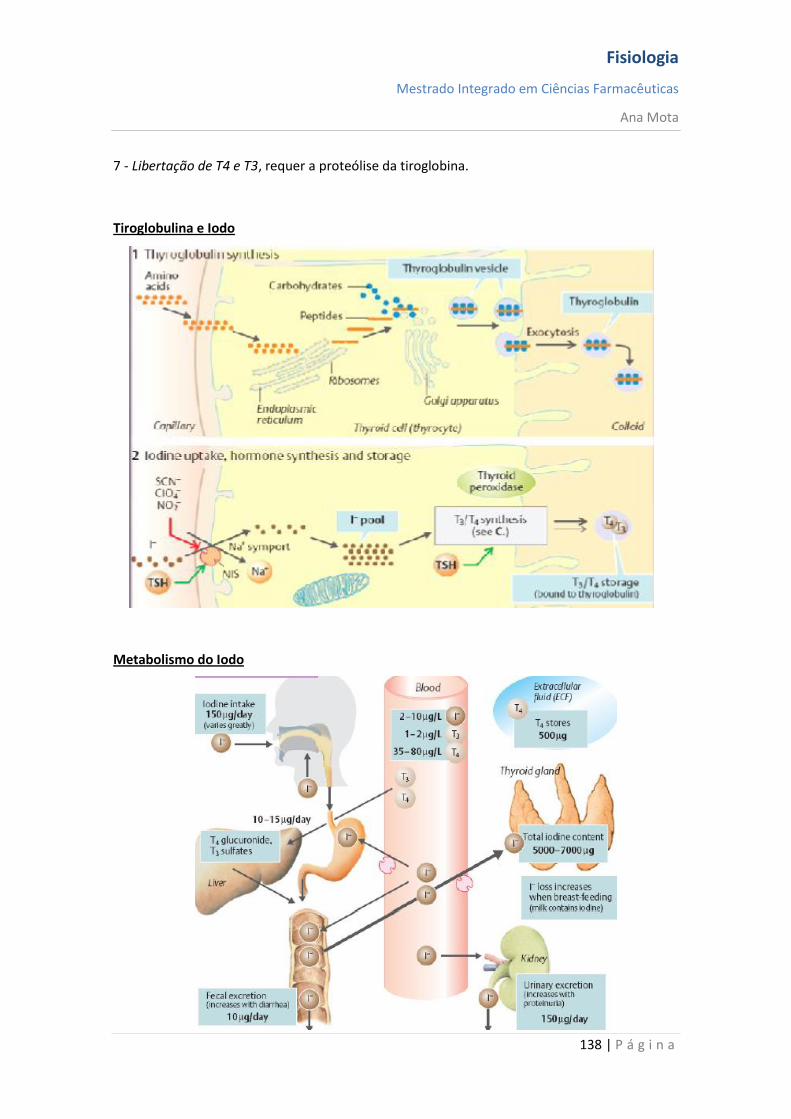
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
138 | P á g i n a
7 - Libertação de T4 e T3, requer a proteólise da tiroglobina.
Tiroglobulina e Iodo
Metabolismo do Iodo

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
139 | P á g i n a
Iodinação e Acoplamento
Quando o I- é oxidado a I2, acopla-se rapidamente à posição 3 das moléculas de tirosina da
tiroglobina, gerando a MIT (monoiodotirosina). Em seguida, a MIT é iodetada na posição 5,
produzindo DIT (diiodotirosina). Após esta etapa, 2 moléculas de DIT ligam-se e formam a
tiroxina – hormona T4; também pode ocorrer que uma molécula de MIT e uma de DIT se
liguem e formem a triiodotironina – hormona T3.
Secreção de T3 e T4
T3 = MIT + DIT T4 = DIT + DIT
A libertação de T3 e T4 no sangue requer a proteólise da tiroglobina. Na superfície das células foliculares, o colóide é incorporado, por endocitose, no lúmen dos folículos. Então, as vesículas coloidais migram na membrana celular apical para a basal e fundem-se com os lisossomas. As proteases lisossómicas libertam T3 e T4 que, assim, deixam a célula.
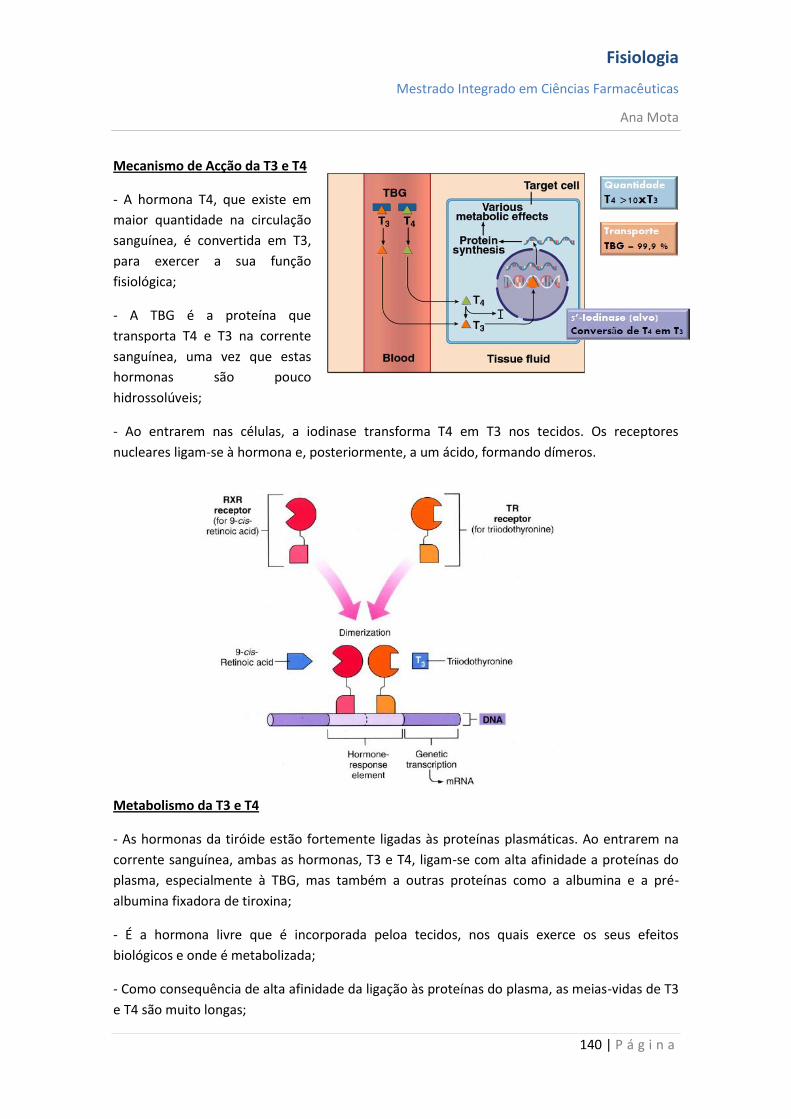
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
140 | P á g i n a
Mecanismo de Acção da T3 e T4
- A hormona T4, que existe em
maior quantidade na circulação
sanguínea, é convertida em T3,
para exercer a sua função
fisiológica;
- A TBG é a proteína que
transporta T4 e T3 na corrente
sanguínea, uma vez que estas
hormonas são pouco
hidrossolúveis;
- Ao entrarem nas células, a iodinase transforma T4 em T3 nos tecidos. Os receptores
nucleares ligam-se à hormona e, posteriormente, a um ácido, formando dímeros.
Metabolismo da T3 e T4
- As hormonas da tiróide estão fortemente ligadas às proteínas plasmáticas. Ao entrarem na
corrente sanguínea, ambas as hormonas, T3 e T4, ligam-se com alta afinidade a proteínas do
plasma, especialmente à TBG, mas também a outras proteínas como a albumina e a pré-
albumina fixadora de tiroxina;
- É a hormona livre que é incorporada peloa tecidos, nos quais exerce os seus efeitos
biológicos e onde é metabolizada;
- Como consequência de alta afinidade da ligação às proteínas do plasma, as meias-vidas de T3
e T4 são muito longas;
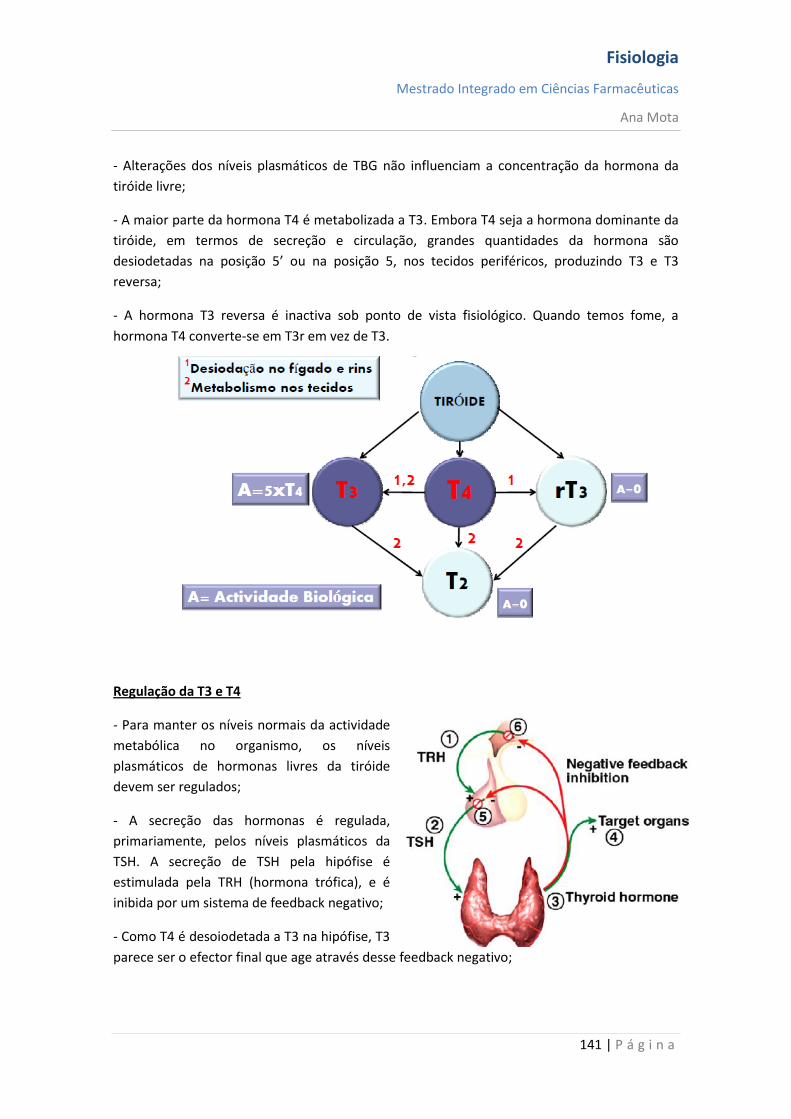
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
141 | P á g i n a
- Alterações dos níveis plasmáticos de TBG não influenciam a concentração da hormona da
tiróide livre;
- A maior parte da hormona T4 é metabolizada a T3. Embora T4 seja a hormona dominante da
tiróide, em termos de secreção e circulação, grandes quantidades da hormona são
desiodetadas na posição 5’ ou na posição 5, nos tecidos periféricos, produzindo T3 e T3
reversa;
- A hormona T3 reversa é inactiva sob ponto de vista fisiológico. Quando temos fome, a
hormona T4 converte-se em T3r em vez de T3.
Regulação da T3 e T4
- Para manter os níveis normais da actividade
metabólica no organismo, os níveis
plasmáticos de hormonas livres da tiróide
devem ser regulados;
- A secreção das hormonas é regulada,
primariamente, pelos níveis plasmáticos da
TSH. A secreção de TSH pela hipófise é
estimulada pela TRH (hormona trófica), e é
inibida por um sistema de feedback negativo;
- Como T4 é desoiodetada a T3 na hipófise, T3
parece ser o efector final que age através desse feedback negativo;
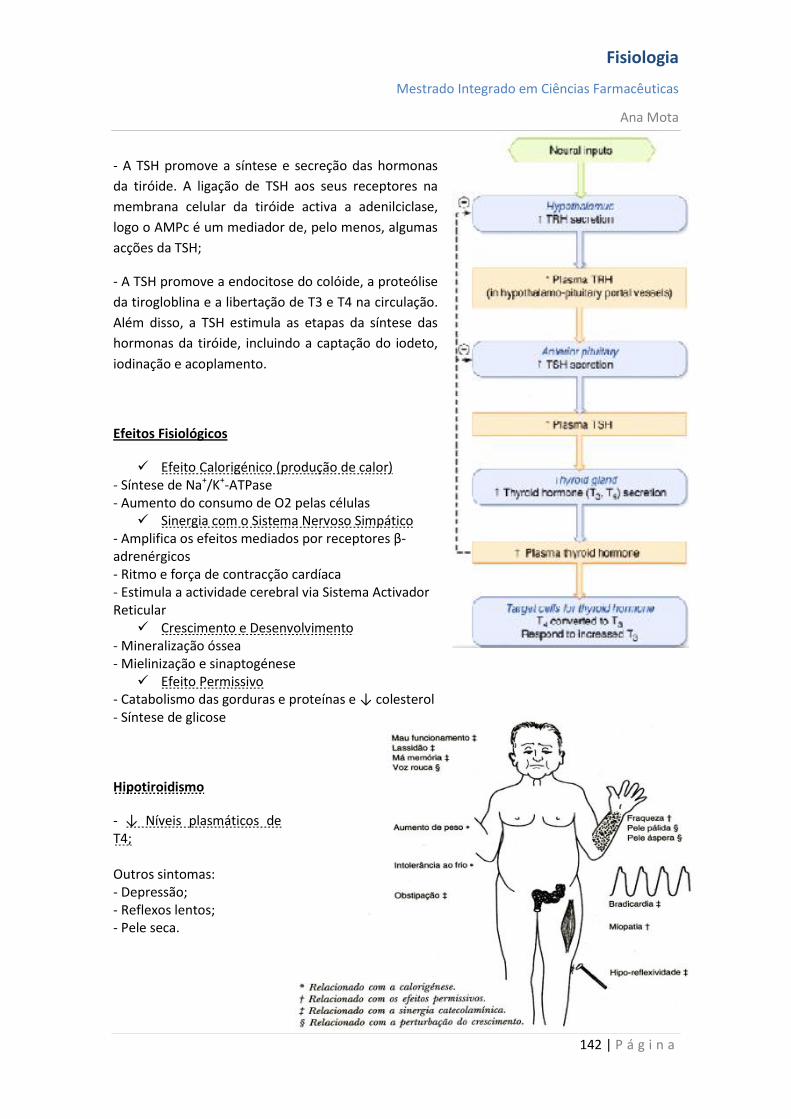
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
142 | P á g i n a
- A TSH promove a síntese e secreção das hormonas
da tiróide. A ligação de TSH aos seus receptores na
membrana celular da tiróide activa a adenilciclase,
logo o AMPc é um mediador de, pelo menos, algumas
acções da TSH;
- A TSH promove a endocitose do colóide, a proteólise
da tirogloblina e a libertação de T3 e T4 na circulação.
Além disso, a TSH estimula as etapas da síntese das
hormonas da tiróide, incluindo a captação do iodeto,
iodinação e acoplamento.
Efeitos Fisiológicos
Efeito Calorigénico (produção de calor) - Síntese de Na+/K+-ATPase - Aumento do consumo de O2 pelas células Sinergia com o Sistema Nervoso Simpático
- Amplifica os efeitos mediados por receptores β-adrenérgicos - Ritmo e força de contracção cardíaca - Estimula a actividade cerebral via Sistema Activador Reticular Crescimento e Desenvolvimento
- Mineralização óssea - Mielinização e sinaptogénese Efeito Permissivo
- Catabolismo das gorduras e proteínas e ↓ colesterol - Síntese de glicose
Hipotiroidismo
- ↓ Níveis plasmáticos de T4; Outros sintomas: - Depressão; - Reflexos lentos; - Pele seca.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
143 | P á g i n a
Cretinismo
Hipotiroidismo no início do desenvolvimento; Manifestações:
- Atraso crescimento e desenvolvimento do SNC; - Sonolência; - Letargia; - Apatia; - Protusão da língua; - Diminuição apetite; - Intolerância ao frio.
Bócio por Carência de Iodo
Em alguns casos, o hipotiroidismo origina o
aparecimento de bócio, o qual diz respeito à
carência de iodo.
Na ausência de iodo, não há produção de T3
e T4 pelas células foliculares, havendo por
isso uma diminuição dos níveis plasmáticos
destas hormonas. Através de um mecanismo
de feedback negativo, há estimulação da
hipófise a produzir TSH, a qual estimula a
produção de T3 e T4. Como não existe iodo
para a produção dessas hormonas, o
feedback vai-se repetindo e forma-se cada
vez mais TSH. Esta, sendo uma hormona
trófica, estimula a proliferação celular,
levando a um aumento do volume da
tiróide.
Hipertiroidismo
- Níveis plasmáticos de T4 e T3;
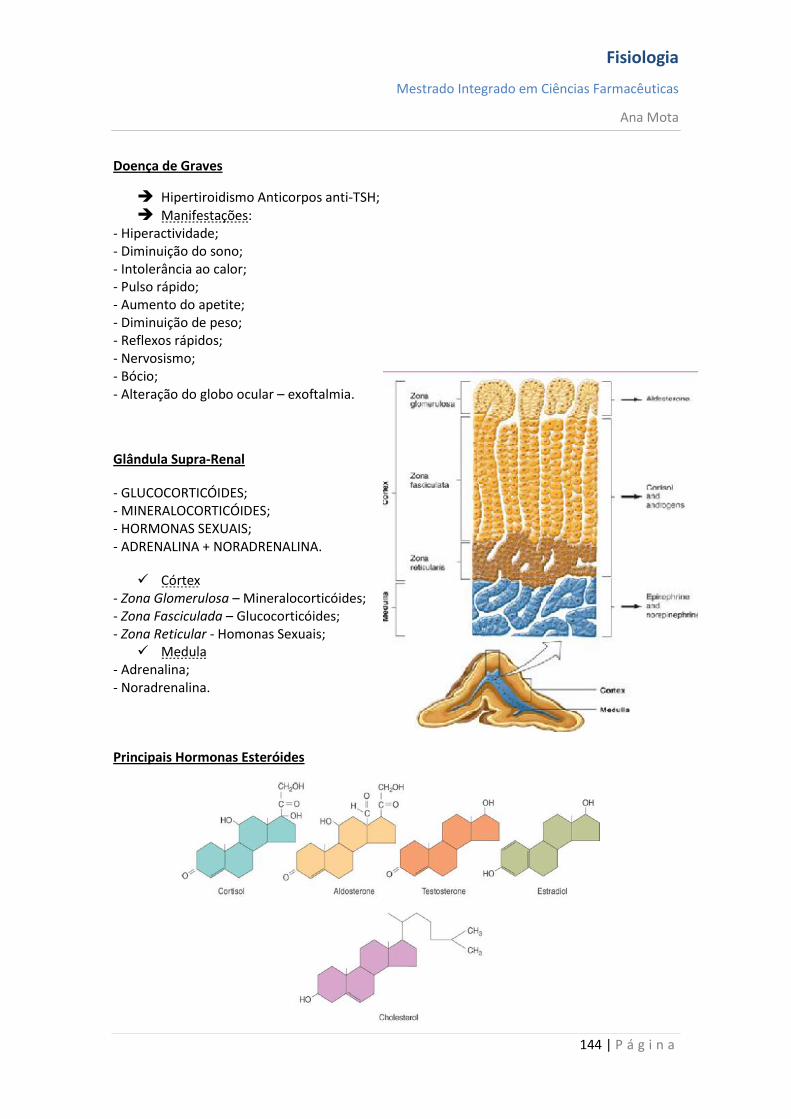
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
144 | P á g i n a
Doença de Graves
Hipertiroidismo Anticorpos anti-TSH; Manifestações:
- Hiperactividade; - Diminuição do sono; - Intolerância ao calor; - Pulso rápido; - Aumento do apetite; - Diminuição de peso; - Reflexos rápidos; - Nervosismo; - Bócio; - Alteração do globo ocular – exoftalmia.
Glândula Supra-Renal
- GLUCOCORTICÓIDES; - MINERALOCORTICÓIDES; - HORMONAS SEXUAIS; - ADRENALINA + NORADRENALINA.
Córtex - Zona Glomerulosa – Mineralocorticóides; - Zona Fasciculada – Glucocorticóides; - Zona Reticular - Homonas Sexuais; Medula
- Adrenalina; - Noradrenalina.
Principais Hormonas Esteróides
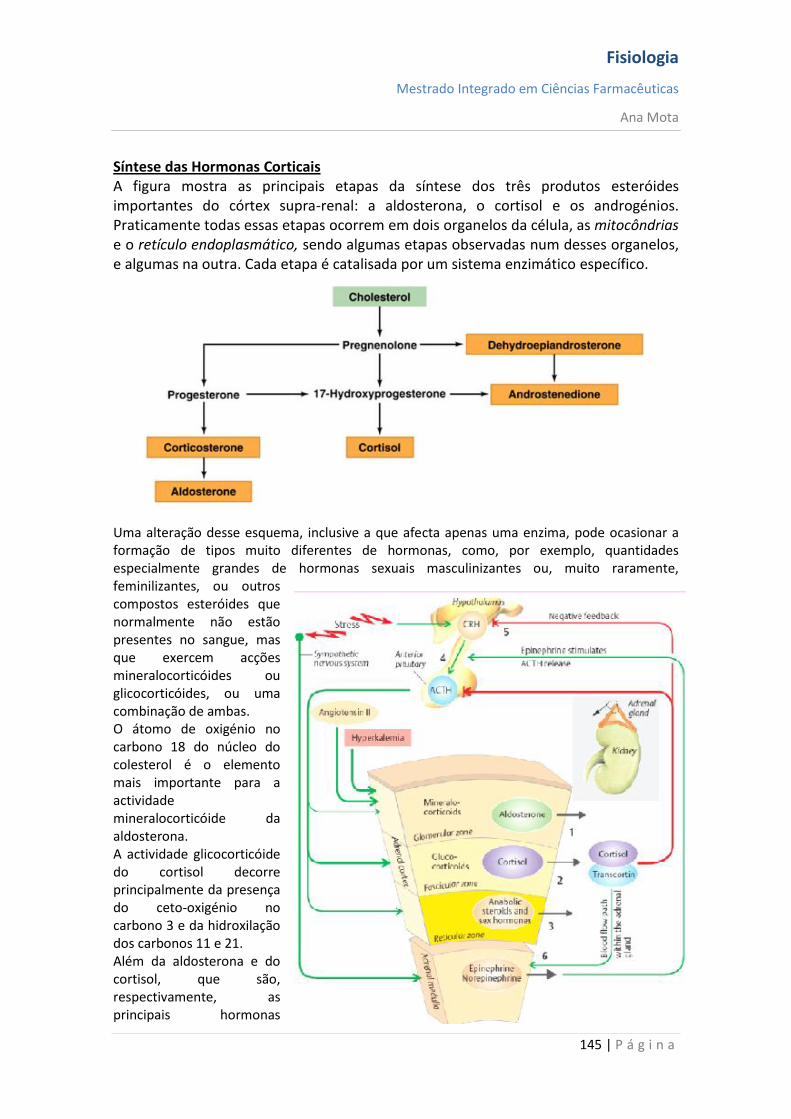
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
145 | P á g i n a
Síntese das Hormonas Corticais A figura mostra as principais etapas da síntese dos três produtos esteróides importantes do córtex supra-renal: a aldosterona, o cortisol e os androgénios. Praticamente todas essas etapas ocorrem em dois organelos da célula, as mitocôndrias e o retículo endoplasmático, sendo algumas etapas observadas num desses organelos, e algumas na outra. Cada etapa é catalisada por um sistema enzimático específico.
Uma alteração desse esquema, inclusive a que afecta apenas uma enzima, pode ocasionar a formação de tipos muito diferentes de hormonas, como, por exemplo, quantidades especialmente grandes de hormonas sexuais masculinizantes ou, muito raramente, feminilizantes, ou outros compostos esteróides que normalmente não estão presentes no sangue, mas que exercem acções mineralocorticóides ou glicocorticóides, ou uma combinação de ambas. O átomo de oxigénio no carbono 18 do núcleo do colesterol é o elemento mais importante para a actividade mineralocorticóide da aldosterona. A actividade glicocorticóide do cortisol decorre principalmente da presença do ceto-oxigénio no carbono 3 e da hidroxilação dos carbonos 11 e 21. Além da aldosterona e do cortisol, que são, respectivamente, as principais hormonas

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
146 | P á g i n a
mineralocorticóides e glicocorticóides, outros esteróides, que exibem uma ou ambas as actividades, são normalmente secretadas em pequenas quantidades pelo córtex supra-renal. Por outro lado, várias outras hormonas esteróides potentes, normalmente não formadas pelas glândulas supra-renais, têm sido sintetizadas e são utilizados em várias formas de terapia.
Actividades Relativas das Hormonas
Cortisol
Glucorticóide, produzido na zona fasciculada da supra-renal;
Efeitos são mediados pela sua ligação a receptores intracelulares nos tecidos-alvo e
pela sua indução (ou repressão) da transcrição de genes;
Efeitos no metabolismo de HC, gorduras e proteínas:
- Reduz as reservas proteicas nos tecidos extra-hepáticos;
- Aumentar a concentração de glucose no sangue;
- Mobilização de ácidos gordos do tecido adiposo – degradação de lípidos.
Transporte do Cortisol
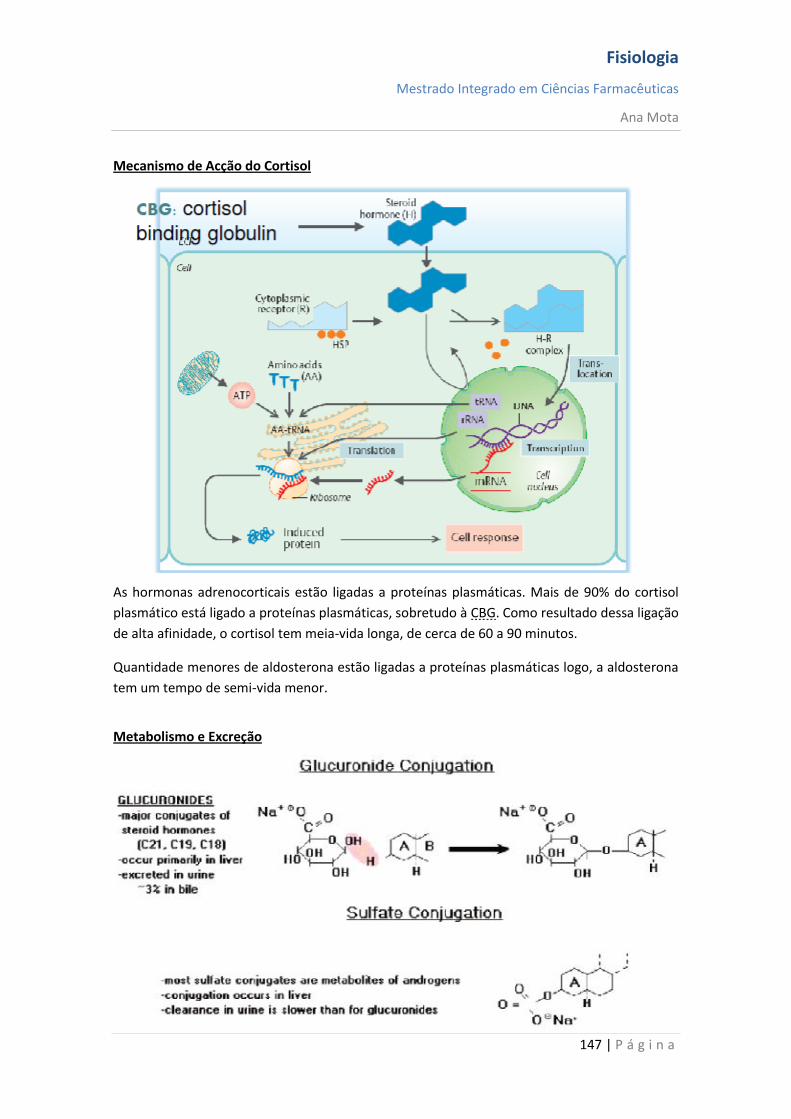
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
147 | P á g i n a
Mecanismo de Acção do Cortisol
As hormonas adrenocorticais estão ligadas a proteínas plasmáticas. Mais de 90% do cortisol
plasmático está ligado a proteínas plasmáticas, sobretudo à CBG. Como resultado dessa ligação
de alta afinidade, o cortisol tem meia-vida longa, de cerca de 60 a 90 minutos.
Quantidade menores de aldosterona estão ligadas a proteínas plasmáticas logo, a aldosterona
tem um tempo de semi-vida menor.
Metabolismo e Excreção
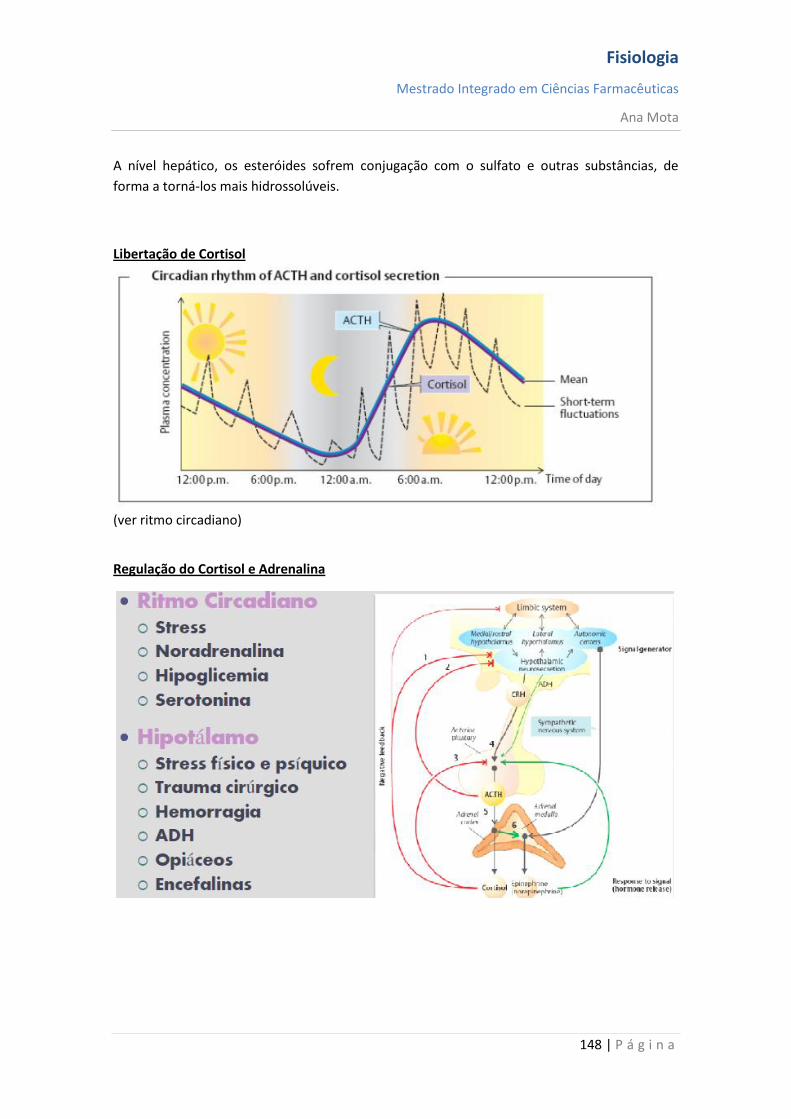
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
148 | P á g i n a
A nível hepático, os esteróides sofrem conjugação com o sulfato e outras substâncias, de
forma a torná-los mais hidrossolúveis.
Libertação de Cortisol
(ver ritmo circadiano)
Regulação do Cortisol e Adrenalina

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
149 | P á g i n a
Resposta ao stress
A secreção de cortisol é importante na
resistência ao stress. Stress físico ou mental
aumenta a secreção de ACTH que, por sua vez,
estimula o córtex adrenala secretar cortisol.
Efeitos do Cortisol
Resposta à Tensão e Resistência ao Stress
- Potencia o efeito das catecolaminas; - Catabolismo das proteínas e lípidos; - Síntese de glicose; - Excitabilidade neuronal; - Reactividade vascular e cardíaca. Efeito Permissivo
- Reactividade vascular às catecolaminas; - Resistência à fadiga (músculo cardíaco); - Efeito da aldosterona, ADH e GH; - Gluconeogénese hepática; - Produção de calor mediada pela T3; - Produção de células do sangue. Regulação da Resposta Inflamatória
- Reduz o número de linfócitos B e T; - Imunossupressão.
Efeitos Metabólicos
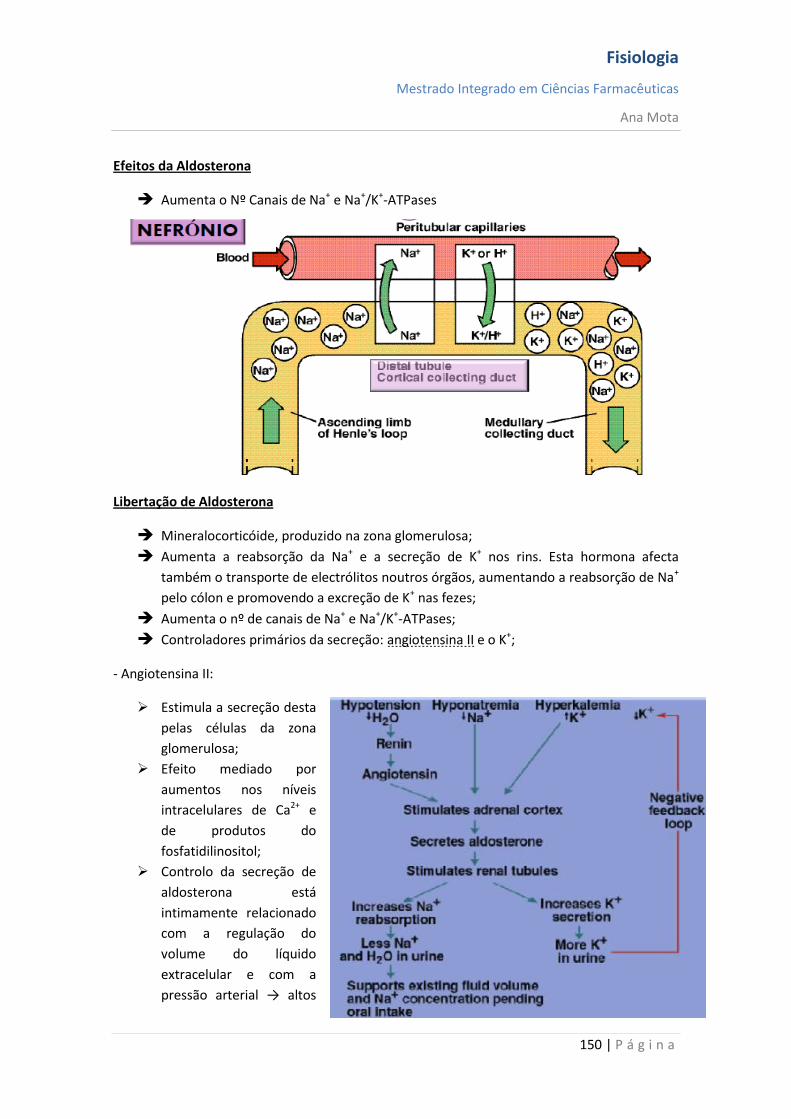
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
150 | P á g i n a
Efeitos da Aldosterona
Aumenta o Nº Canais de Na+ e Na+/K+-ATPases
Libertação de Aldosterona
Mineralocorticóide, produzido na zona glomerulosa;
Aumenta a reabsorção da Na+ e a secreção de K+ nos rins. Esta hormona afecta
também o transporte de electrólitos noutros órgãos, aumentando a reabsorção de Na+
pelo cólon e promovendo a excreção de K+ nas fezes;
Aumenta o nº de canais de Na+ e Na+/K+-ATPases;
Controladores primários da secreção: angiotensina II e o K+;
- Angiotensina II:
Estimula a secreção desta
pelas células da zona
glomerulosa;
Efeito mediado por
aumentos nos níveis
intracelulares de Ca2+ e
de produtos do
fosfatidilinositol;
Controlo da secreção de
aldosterona está
intimamente relacionado
com a regulação do
volume do líquido
extracelular e com a
pressão arterial → altos

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
151 | P á g i n a
níveis plasmáticos de angiotensina II estimulam a secreção de aldosterona;
Actividade do sistema renina-angiotensina é inversamente proporcional à ingestão de
Na+ na dieta;
K+ estimula a secreção de aldosterona. Aumentos da concentração plasmática de K+
aumentam a secreção de aldosterona por despolarização da membrana celular,
abertura dos canais de Ca2+ e aumento da concentração intracelular de Ca2+. Em
resposta a este mecanismo, a secreção de aldosterona aumenta como resultado das
mesmas etapas biossintéticas e tardias afectadas pela angiotensina II;
Altos níveis plasmáticos de K+, aumentam a secreção de aldosterona que, por sua vez,
estimula a secreção tubular de K+. Quando as concentrações plasmáticas de K+ voltam
ao normal, o estímulo para a secreção de aldosterona é removido;
ACTH desempenha um papel permissivo na regulação da secreção de aldosterona.
Desde que existam níveis normais de ACTH no plasma, mantêm-se a resposta
glomerulosa aos seus reguladores principais, a angiotensina II e K+. Por outro lado, se o
ACTH está deficiente, a resposta da aldosterona diminui.
DESTRUIÇÃO DA SUPRA-RENAL Doença de Addison
Outras alterações:
Personalidade; ↓Concentração;
Apreensão; Irritabilidade.
HIPERCORTISOLISMO
Síndrome Cushing
Outras alterações: Personalidade;
Apetite; Insónia; Euforia; Psicose.
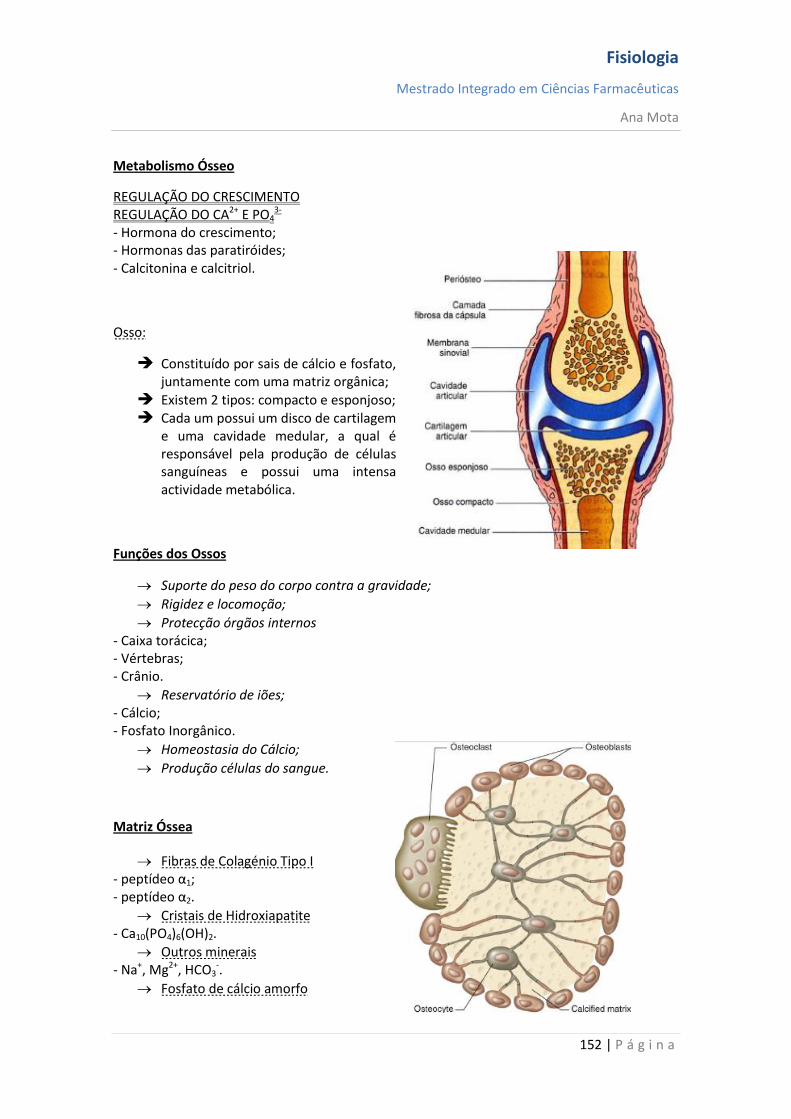
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
152 | P á g i n a
Metabolismo Ósseo
REGULAÇÃO DO CRESCIMENTO REGULAÇÃO DO CA2+ E PO4
3- - Hormona do crescimento; - Hormonas das paratiróides; - Calcitonina e calcitriol.
Osso:
Constituído por sais de cálcio e fosfato, juntamente com uma matriz orgânica;
Existem 2 tipos: compacto e esponjoso; Cada um possui um disco de cartilagem
e uma cavidade medular, a qual é responsável pela produção de células sanguíneas e possui uma intensa actividade metabólica.
Funções dos Ossos
Suporte do peso do corpo contra a gravidade;
Rigidez e locomoção;
Protecção órgãos internos - Caixa torácica; - Vértebras; - Crânio.
Reservatório de iões; - Cálcio; - Fosfato Inorgânico.
Homeostasia do Cálcio;
Produção células do sangue.
Matriz Óssea
Fibras de Colagénio Tipo I - peptídeo α1; - peptídeo α2.
Cristais de Hidroxiapatite - Ca10(PO4)6(OH)2.
Outros minerais - Na+, Mg2+, HCO3
-.
Fosfato de cálcio amorfo
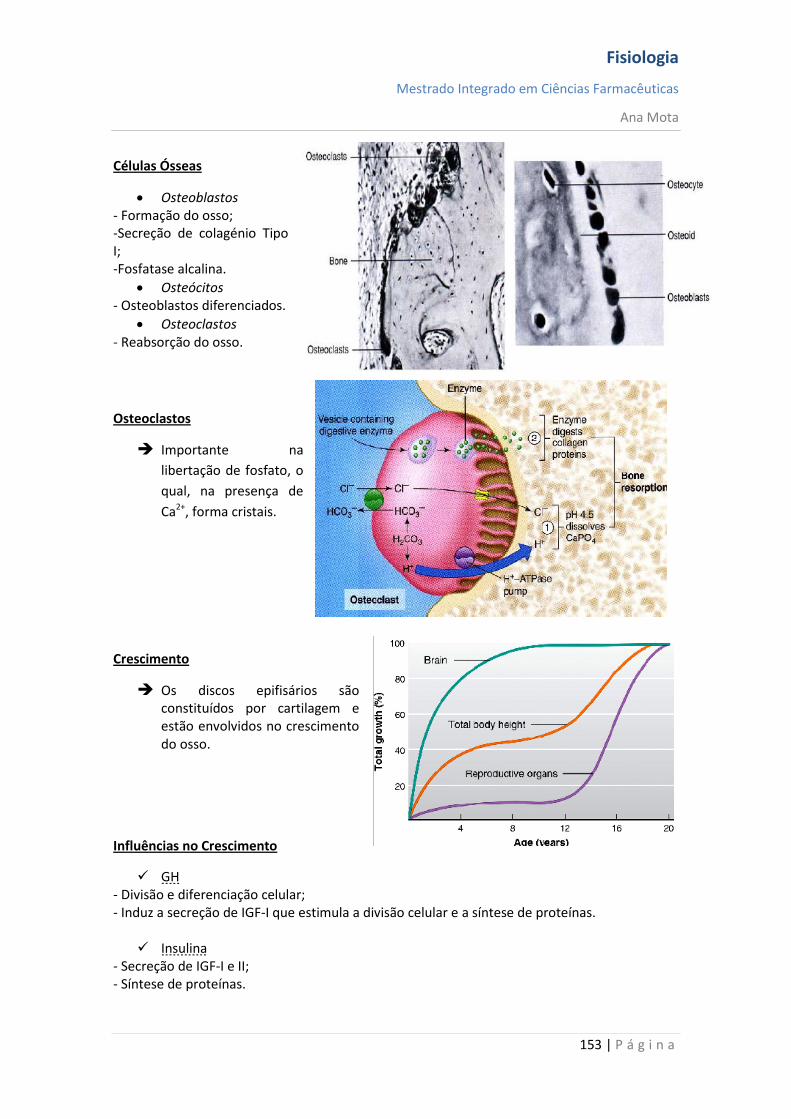
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
153 | P á g i n a
Células Ósseas
Osteoblastos - Formação do osso; -Secreção de colagénio Tipo I; -Fosfatase alcalina.
Osteócitos - Osteoblastos diferenciados.
Osteoclastos - Reabsorção do osso.
Osteoclastos
Importante na
libertação de fosfato, o
qual, na presença de
Ca2+, forma cristais.
Crescimento
Os discos epifisários são constituídos por cartilagem e estão envolvidos no crescimento do osso.
Influências no Crescimento
GH - Divisão e diferenciação celular; - Induz a secreção de IGF-I que estimula a divisão celular e a síntese de proteínas. Insulina
- Secreção de IGF-I e II; - Síntese de proteínas.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
154 | P á g i n a
T3 e T4 - Efeito permissivo à acção da GH. Hormonas Sexuais
- Secreção de GH e IGF-I; - Fecho das epífises (Testosterona); - Síntese de proteínas.
Hormona do Crescimento = GH
Produzida pela adeno-hipófise e é controlada por 2 factores hipotalâmicos (estimulador/inibidor);
Efeito no metabolismo é indirecto, uma vez que depende do factor IGF-I, o qual é produzido nos ossos, cartilagem e fígado;
Estimula o tecido adiposo a libertar ácidos gordos e, noutros tecidos, provoca uma diminuição da captação da glucose pelas células;
Factor IGF-I provoca a divisão dos condrócitos da cartilagem e um aumento da massa muscular, síntese de proteínas e crescimento de vários órgãos;
Regulação da Secreção de GH
Factores que accionam a sua libertação: exercício físico, stress, jejum, sono, etc.;
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
155 | P á g i n a
Nanismo – Gigantismo – Acromegalia
Hipoprodução de GH ou IGF-I → Nanismo; Hiperprodução de GH ou IGF-I → Gigantismo; Acromegalia → as epífises encontram-se fechadas → alterações da estrutura óssea
(rosto, mãos e pés).
Remodelação Óssea
Formação do Osso – Massa óssea - Insulina e IGF-I; - GH; - Testosterona e Estrogénios; - Calcitriol e Calcitonina.
Reabsorção do Osso – ↓ Massa óssea - PTH (elevações crónicas); - Cortisol; - Hormonas da tiróide (T4 e T3).
Índice de Massa Óssea
Distribuição do Cálcio
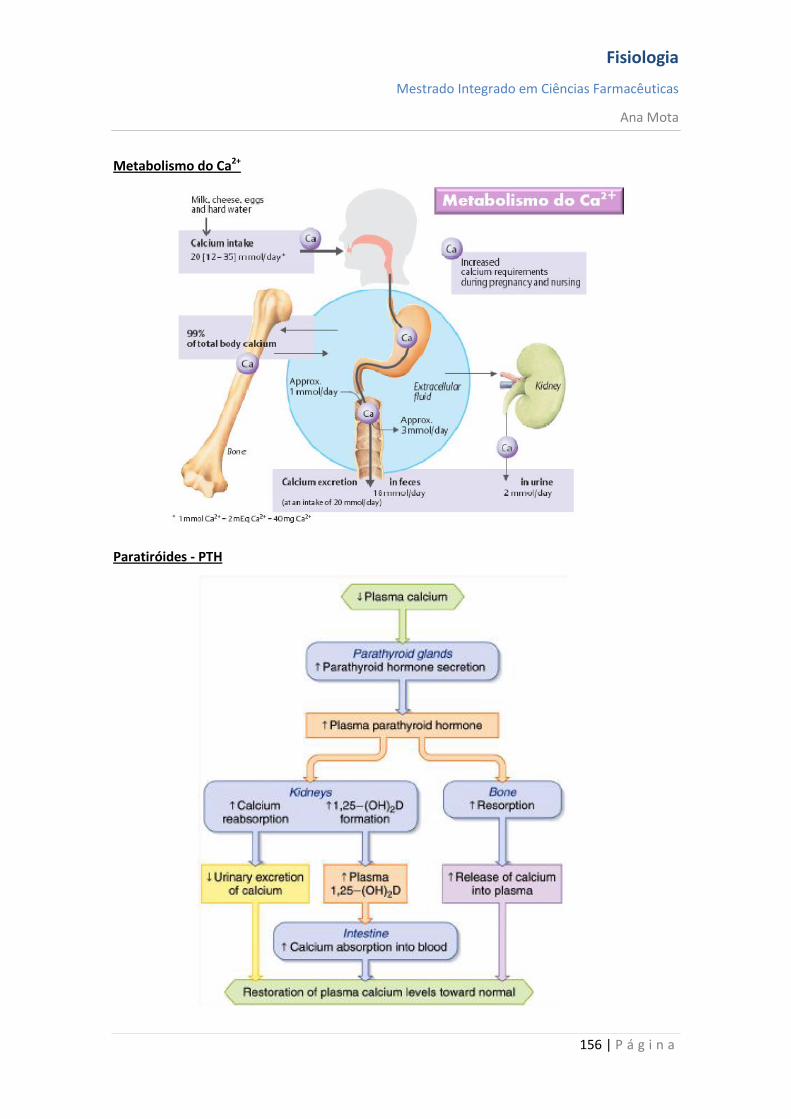
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
156 | P á g i n a
Metabolismo do Ca2+
Paratiróides - PTH

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
157 | P á g i n a
O cálcio e o fosfato presentes no osso funcionam como reservatórios para iões do LEC. Se a
concentração de cálcio no LEC cai abaixo do normal, os iões cálcio movem-se do osso para o
LEC. A distribuição entre o osso e o LEC é afectada pela hormona PTH, que estimulam o
movimento do cálcio e fosfato do osso para o LEC, e pela calcitonina, que tem o efeito oposto.
Inversamente, quando a concentração de cálcio no LEC aumenta acima do normal, o cálcio
pode ser depositado.
A secreção de PTH aumenta em resposta à redução da concentração de cálcio
extracelular. Pequenas reduções na concentração do ião resultam em grandes
aumentos da formação de PTH.
Aumento na concentração de PTH diminui a excreção renal de cálcio e aumenta a
excreção de fosfato. A PTH inibe a reabsorção de fosfato no túbulo proximal.
Calcitriol – 1,25-(OH)2D3
A PTH contribui para a formação de calcitriol (derivafo da vit. D), o qual é um
estimulante à absorção intestinal de Ca2+ e fosfato orgânico;
No nosso organismo existe um composto que, por acção do sol, se transforma em
vitamina D3 → vai para a circulação e chega até ao fígado onde sofre hidroxilação →
fica na forma de outro composto → conduzido até ao rim e sofre hidroxilação →
Calcitriol
O calcitriol actua de acordo com a presença/ausência de PTH. Na presença de PTH há
um aumento da reabsorção do osso;
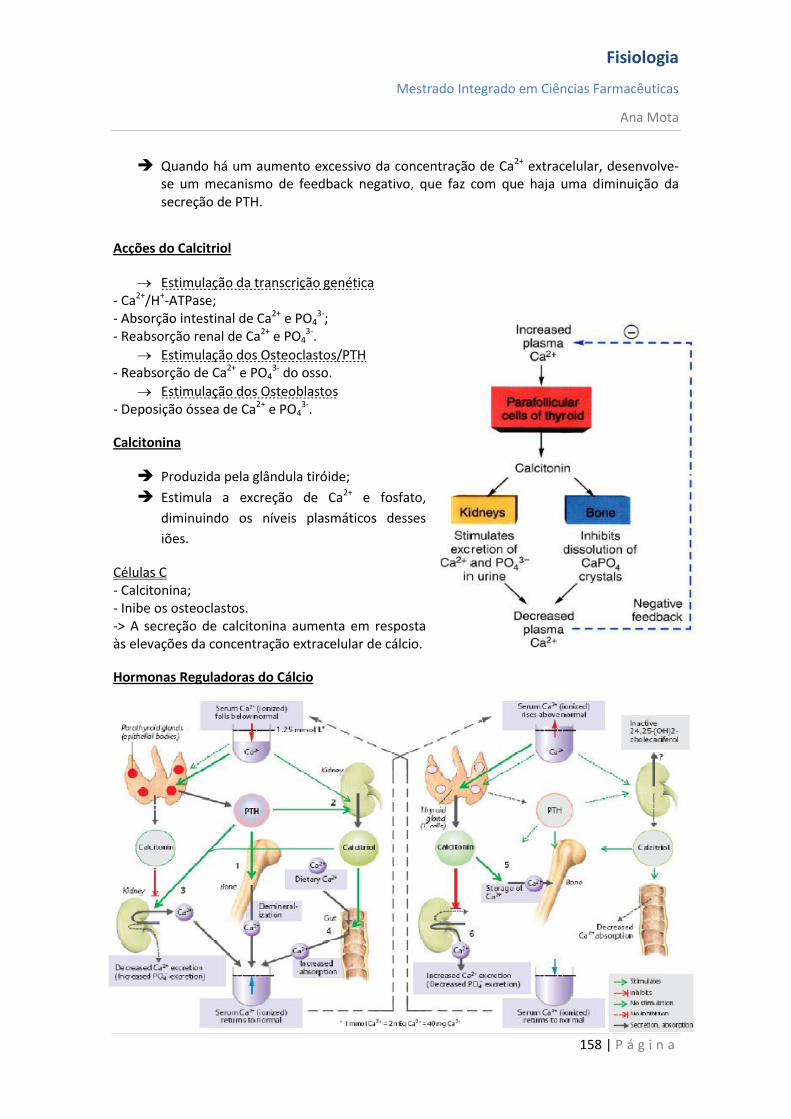
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
158 | P á g i n a
Quando há um aumento excessivo da concentração de Ca2+ extracelular, desenvolve-se um mecanismo de feedback negativo, que faz com que haja uma diminuição da secreção de PTH.
Acções do Calcitriol
Estimulação da transcrição genética - Ca2+/H+-ATPase; - Absorção intestinal de Ca2+ e PO4
3-; - Reabsorção renal de Ca2+ e PO4
3-.
Estimulação dos Osteoclastos/PTH - Reabsorção de Ca2+ e PO4
3- do osso.
Estimulação dos Osteoblastos - Deposição óssea de Ca2+ e PO4
3-.
Calcitonina
Produzida pela glândula tiróide;
Estimula a excreção de Ca2+ e fosfato,
diminuindo os níveis plasmáticos desses
iões.
Células C - Calcitonina; - Inibe os osteoclastos. -> A secreção de calcitonina aumenta em resposta às elevações da concentração extracelular de cálcio.
Hormonas Reguladoras do Cálcio

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
159 | P á g i n a
Ca2+ - PTH; - Calcitriol; - GH.
Ca2+ - Calcitonina; - Cortisol.
Pretende-se que os níveis de cálcio
plasmáticos sejam constantes, uma vez que
este ião modifica actividades
musculares/neurotransmissores.
Outras Hormonas Reguladoras do Ca2+
Cortisol
- Absorção intestinal de Ca2+ e PO43-;
- Excreção renal de Ca2+ e PO43-;
- Inibição dos osteoblastos (a longo prazo); - Osteoporose (a longo prazo).
GH
- Absorção intestinal de Ca2+ e PO43-.
Ca2+ - Desregulação dos níveis plasmáticos
[Ca2+]plasma
Hipocalcemia
- da excitabilidade neuronal;
- da excitabilidade muscular; - Espasmos dos músculos esqueléticos.
[Ca2+]plasma
Hipercalcemia - Arritmias cardíacas;
- da excitabilidade nervosa;
- da excitabilidade muscular.
Doenças Metabólicas Ósseas
RAQUITISMO (crianças) ou OSTEOMALÁCIA (adultos)
- Mineralização deficiente da matriz - Calcitriol. OSTEOPOROSE
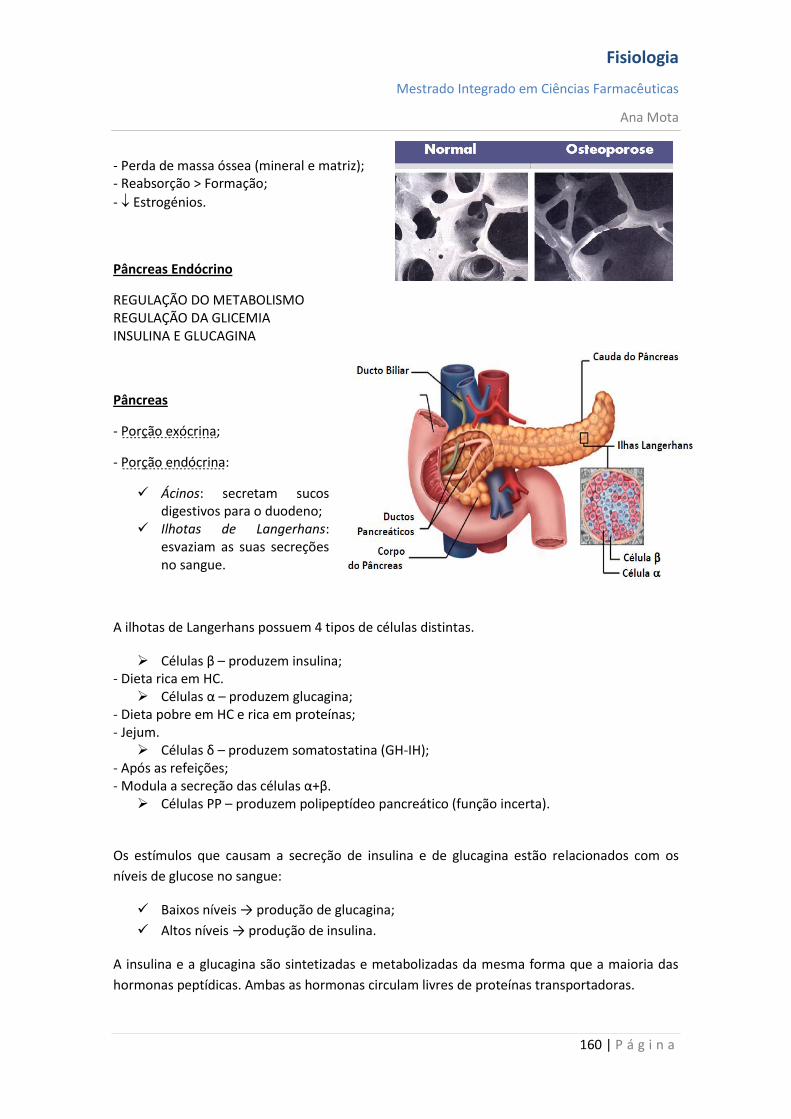
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
160 | P á g i n a
- Perda de massa óssea (mineral e matriz); - Reabsorção > Formação;
- Estrogénios.
Pâncreas Endócrino
REGULAÇÃO DO METABOLISMO REGULAÇÃO DA GLICEMIA INSULINA E GLUCAGINA
Pâncreas
- Porção exócrina;
- Porção endócrina:
Ácinos: secretam sucos digestivos para o duodeno;
Ilhotas de Langerhans: esvaziam as suas secreções no sangue.
A ilhotas de Langerhans possuem 4 tipos de células distintas.
Células β – produzem insulina; - Dieta rica em HC. Células α – produzem glucagina;
- Dieta pobre em HC e rica em proteínas; - Jejum. Células δ – produzem somatostatina (GH-IH);
- Após as refeições; - Modula a secreção das células α+β. Células PP – produzem polipeptídeo pancreático (função incerta).
Os estímulos que causam a secreção de insulina e de glucagina estão relacionados com os
níveis de glucose no sangue:
Baixos níveis → produção de glucagina;
Altos níveis → produção de insulina.
A insulina e a glucagina são sintetizadas e metabolizadas da mesma forma que a maioria das
hormonas peptídicas. Ambas as hormonas circulam livres de proteínas transportadoras.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
161 | P á g i n a
Fontes de glucose
Insulina
Principal controlador do metabolismo orgânico; Essencial para o consumo de HC, síntese de proteínas e armazenamento de lípidos;
Capacidade de o transporte de glucose através da maioria das membranas celulares, logo sempre que os níveis de glicemia aumentam, é libertada insulina, a qual estabiliza rapidamente esses níveis;
Após uma refeição: níveis de glucose , estimulando as células β a produzirem insulina e inibindo as células α;
Transporte de glucose ocorre por difusão facilitada e termina quando a concentração de glucose intracelular iguala a extracelular;
Após exercício físico prolongado: níveis de insulina tendem a baixar ao longo do tempo, apesar das concentrações plasmáticas de glucose se manterem relativamente estáveis. Isto pode ser explicado pelo facto de nº e disponibilidade dos receptores para a insulina aumentar com o exercício;
Através da insulina, a glicose pode então entrar nas células e produzir energia (ATP).
Libertação de Insulina
O ATP fecha os canais de K+, ficando os iões K+ dentro da célula, despolarizando a
membrana, abrindo os canais de Ca2+ dependentes da voltagem. O Ca2+ entra e reduz a
endocitose da insulina.
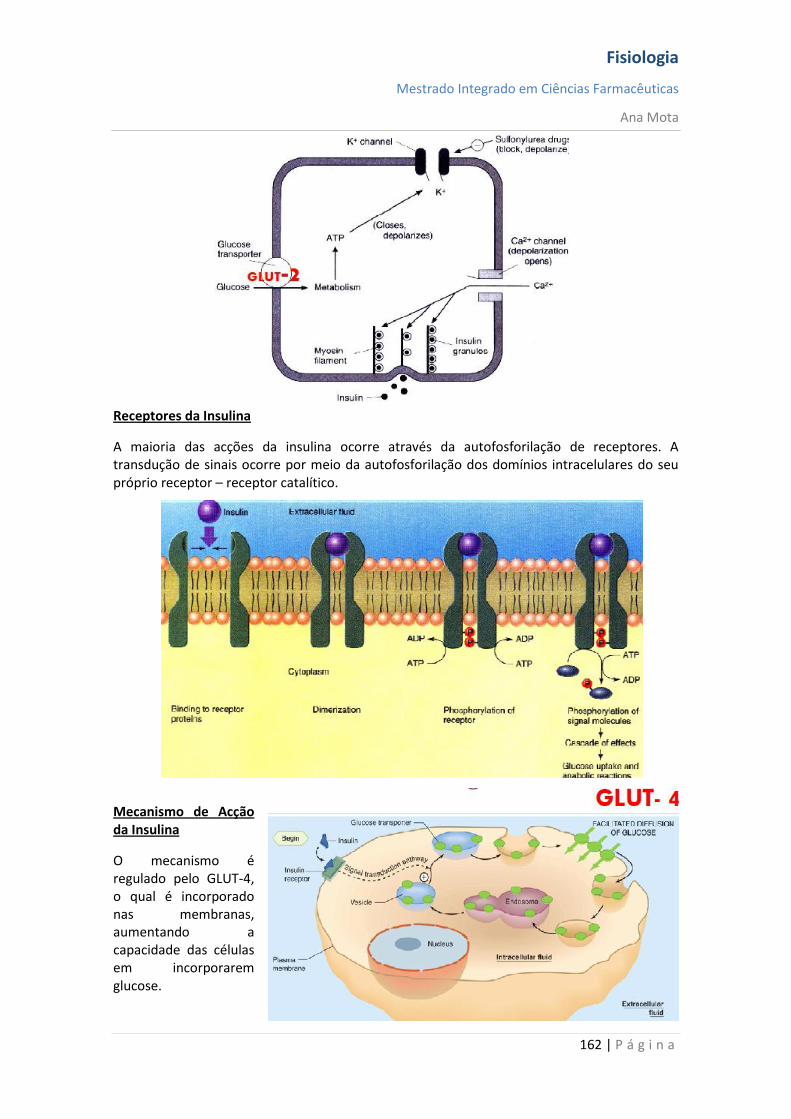
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
162 | P á g i n a
Receptores da Insulina
A maioria das acções da insulina ocorre através da autofosforilação de receptores. A transdução de sinais ocorre por meio da autofosforilação dos domínios intracelulares do seu próprio receptor – receptor catalítico.
Mecanismo de Acção da Insulina
O mecanismo é regulado pelo GLUT-4, o qual é incorporado nas membranas, aumentando a capacidade das células em incorporarem glucose.
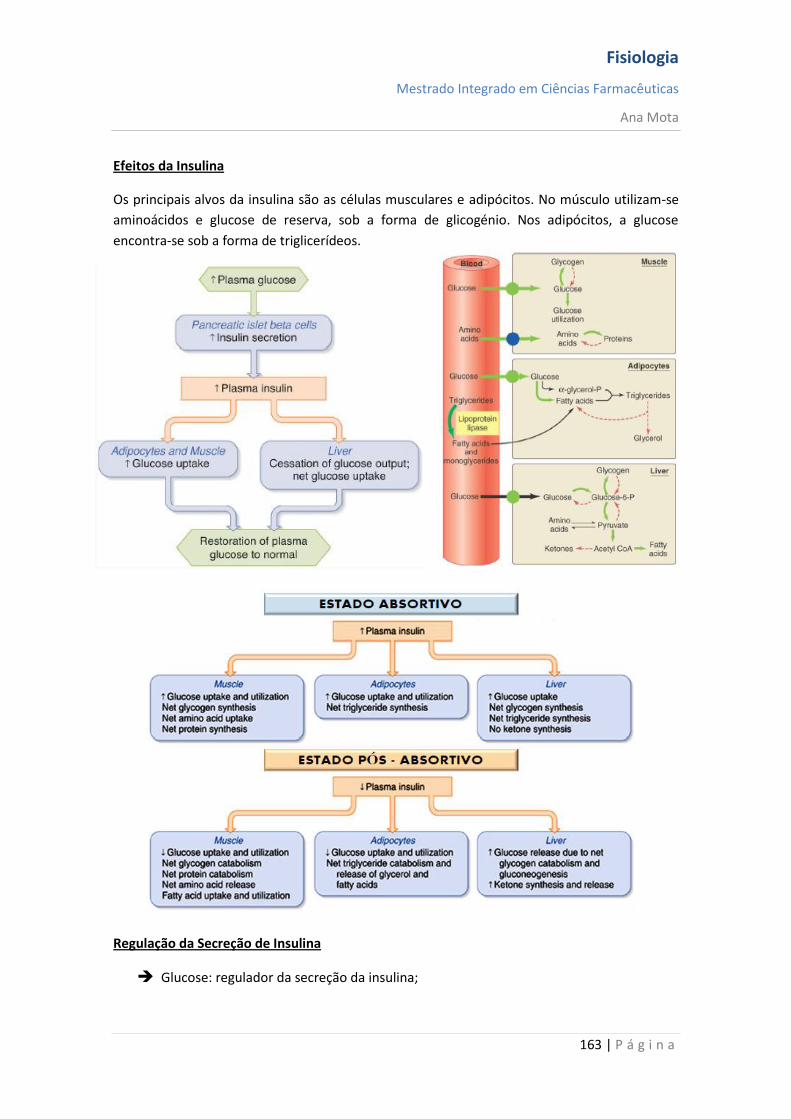
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
163 | P á g i n a
Efeitos da Insulina
Os principais alvos da insulina são as células musculares e adipócitos. No músculo utilizam-se
aminoácidos e glucose de reserva, sob a forma de glicogénio. Nos adipócitos, a glucose
encontra-se sob a forma de triglicerídeos.
Regulação da Secreção de Insulina
Glucose: regulador da secreção da insulina;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
164 | P á g i n a
Feedback da glucose no sangue nas células β do pâncreas. Quando a concentração de
glucose no sangue sobe acima dos níveis de jejum, a secreção de insulina aumenta.
Como resultado dos efeitos da insulina, estimulando a incorporação de glucose pelo
fígado e pelos tecidos periféricos, a concentração de glucose no sangue regressa aos
valores normais;
Além da hiperglicemia, outros estímulos aumentam a secreção de insulina:
- Aminoácidos – os a.a. da dieta são retirados do sangue e usados pelas células para sintetizar
proteínas;
- Hormonas gastrointestinais;
- Cortisol e Hormona do Crescimento – antagonizam os efeitos da insulina na incorporação de
glicose nos tecidos periféricos, levando a um aumento da concentração de glicose no sangue;
- SNA – Parassimpático: estimula a secreção pelas células β;
- Simpático: inibe a secreção pelas células β em situações em que há necessidade de
mais energia, logo de mais glucose;
- GIP – avisa o pâncreas que os níveis de glicemia vão aumentar.
Glucagina
A maioria das acções ocorre pela activação da adenilciclase. A ligação da glucagina a
receptores hepáticos produz a activação da adenilciclase e gera-se um 2º mensageiro,
AMPc que, por sua vez, activa a proteína cinase A, levando a fosforilação que
produzem activação e de várias enzimas;
A glucagina promove hiperglicemia de vários modos:
- Estimula a glicogenólise;
- Inibe a glicólise;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
165 | P á g i n a
- Estimula a gliconeogénese;
- Aumento da concentração da glucose no sangue diminui a secreção de glucagina.
Glucose é o regulador mais importante da secreção de glucagina, exercendo efeitos
opostos ao da secreção de insulina;
A hipoglicemia aumenta a secreção de glucagina:
- Concentração de glucose no sangue retorna ao normal.
Em jejum, ocorre estimulação de células α para produzirem glucagina e inibição das
células β;
Os a.a., especialmente a arginina e a alanina, estimulam a secreção de glucagina;
Jejum e o exercício físico estimulam a secreção desta hormona;
A secreção de glucagina pode ser mediada, pela estimulação do SNA simpático;
Quando não existe glicogénio suficiente, os a.a. são metabolizados a corpos cetónicos,
capazes de fornecer ATP (no fígado).
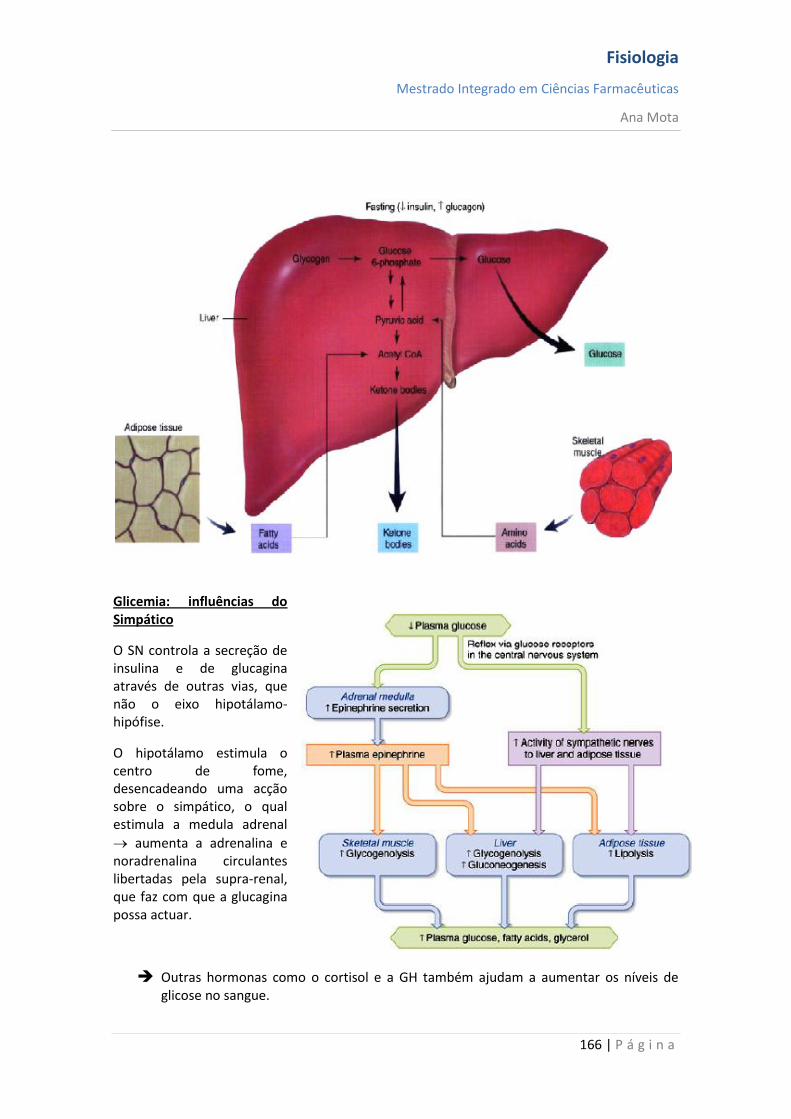
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
166 | P á g i n a
Glicemia: influências do Simpático
O SN controla a secreção de insulina e de glucagina através de outras vias, que não o eixo hipotálamo-hipófise.
O hipotálamo estimula o centro de fome, desencadeando uma acção sobre o simpático, o qual estimula a medula adrenal
aumenta a adrenalina e noradrenalina circulantes libertadas pela supra-renal, que faz com que a glucagina possa actuar.
Outras hormonas como o cortisol e a GH também ajudam a aumentar os níveis de glicose no sangue.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
167 | P á g i n a
Glicemia: influência do Cortisol
Glicemia: controlos de contra-regulação
Diabetes Mellitus
A Diabetes mellitus e, fundamentalmente, o resultado da secreção inadequada de insulina ou da incapacidade dos tecidos para responderem a insulina.
TIPO 1 – 5.10% - Destruição das células β (auto-imune); - Juvenil, adultos (ocasional).
TIPO 2 - 90% - Secreção e/ou Resistência à insulina; - Adultos (~> 40 anos); - Factores de risco: hereditariedade, obesidade.
POLIÚRIA – POLIDIPSIA – POLIFAGIA HIPERGLICEMIA – GLICOSÚRIA – CETONÚRIA

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
168 | P á g i n a
Diagnóstico da Diabetes Mellitus
Medir os níveis de glucose a qualquer altura do dia. No entanto, devem fazer-se testes mais
específicos:
1. Medir a glucose em jejum (< 110 mg/mL);
2. Ingerir 75g glicose pura e medir a glicemia 2h após este níveis (< 140 mg/mL).
Deficiência de Insulina I. Hiperglicemia II. Lipólise III. Cetonémia IV. Cetonúria V. Acidose VI. Hipotensão VII. Disfunção cerebral VIII. Coma MORTE
Num diabético com
hiperglicemia muito elevada
por falta de insulina, há
diminuição da captação de
glicose e aumento da
glicogenólise e da
gliconeogénese.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
169 | P á g i n a
Os corpos cetónicos são ácidos e causam acidose no plasma, ocorrendo diurese osmótica
(perda de líquidos). Juntamente com água, perdem-se também iões Na+, fazendo com que haja
uma redução do volume plasmático e o sangue não chega aos neurónios para os alimentar –
disfunção cerebral.
Módulo 6 – Sistema Cardiovascular
ORGANIZAÇÃO GERAL:
- Coração; - Circulação; - Sangue.
Funções do Sistema Cardiovascular
Rede de Transporte - Nutrientes, hormonas, metabolitos, gases; Regulação da temperatura corporal
- Distribuição de sangue; Protecção e defesa
- Células do sangue; - Mecanismo de coagulação.
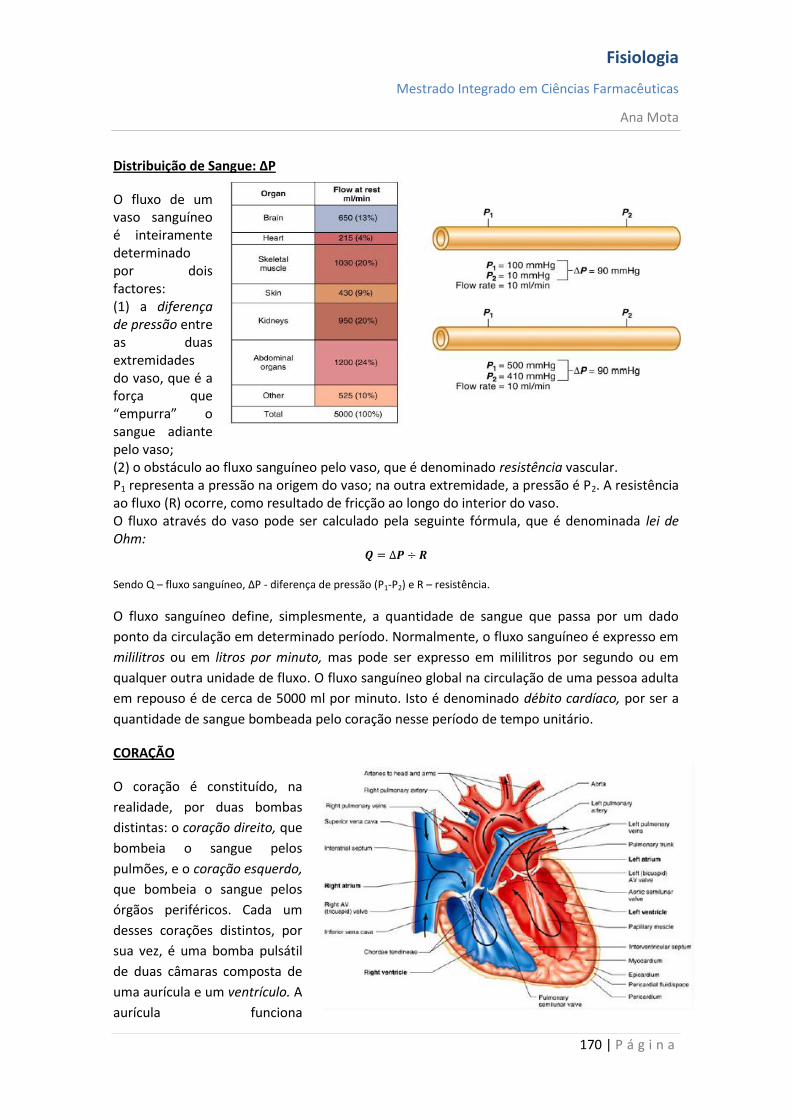
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
170 | P á g i n a
Distribuição de Sangue: ΔP
O fluxo de um vaso sanguíneo é inteiramente determinado por dois factores: (1) a diferença de pressão entre as duas extremidades do vaso, que é a força que “empurra” o sangue adiante pelo vaso; (2) o obstáculo ao fluxo sanguíneo pelo vaso, que é denominado resistência vascular. P1 representa a pressão na origem do vaso; na outra extremidade, a pressão é P2. A resistência ao fluxo (R) ocorre, como resultado de fricção ao longo do interior do vaso. O fluxo através do vaso pode ser calculado pela seguinte fórmula, que é denominada lei de Ohm:
Sendo Q – fluxo sanguíneo, ΔP - diferença de pressão (P1-P2) e R – resistência.
O fluxo sanguíneo define, simplesmente, a quantidade de sangue que passa por um dado
ponto da circulação em determinado período. Normalmente, o fluxo sanguíneo é expresso em
mililitros ou em litros por minuto, mas pode ser expresso em mililitros por segundo ou em
qualquer outra unidade de fluxo. O fluxo sanguíneo global na circulação de uma pessoa adulta
em repouso é de cerca de 5000 ml por minuto. Isto é denominado débito cardíaco, por ser a
quantidade de sangue bombeada pelo coração nesse período de tempo unitário.
CORAÇÃO
O coração é constituído, na
realidade, por duas bombas
distintas: o coração direito, que
bombeia o sangue pelos
pulmões, e o coração esquerdo,
que bombeia o sangue pelos
órgãos periféricos. Cada um
desses corações distintos, por
sua vez, é uma bomba pulsátil
de duas câmaras composta de
uma aurícula e um ventrículo. A
aurícula funciona
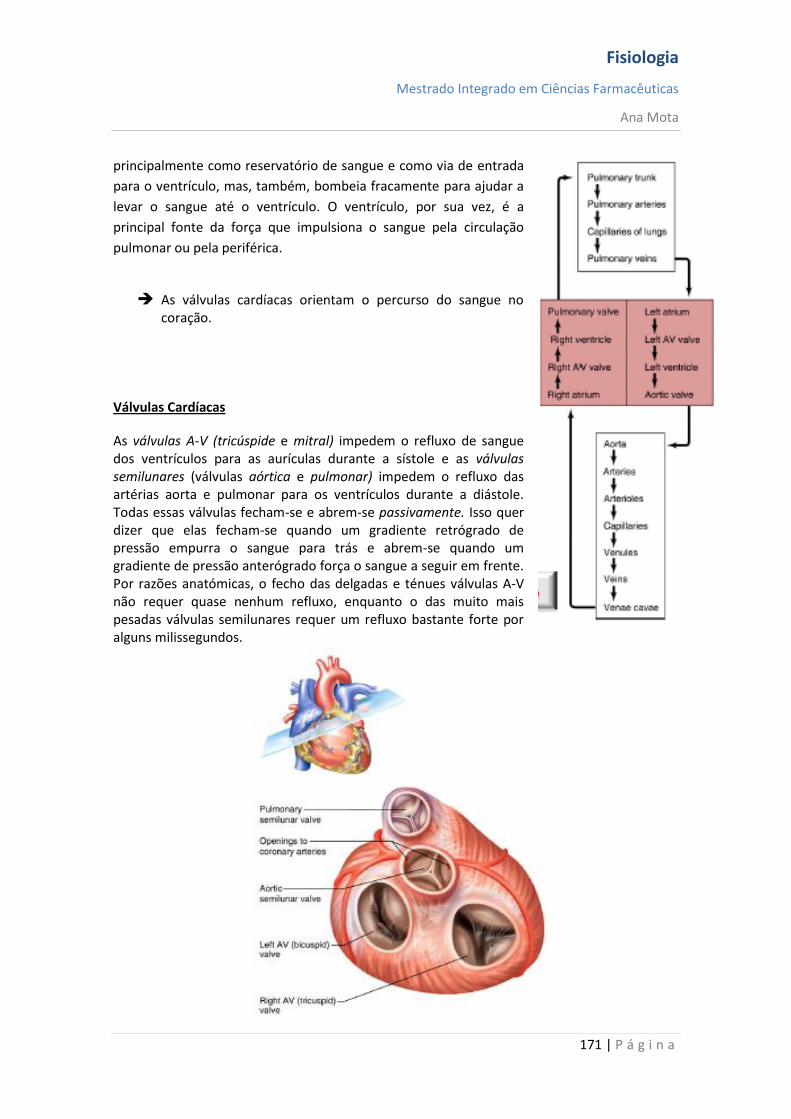
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
171 | P á g i n a
principalmente como reservatório de sangue e como via de entrada
para o ventrículo, mas, também, bombeia fracamente para ajudar a
levar o sangue até o ventrículo. O ventrículo, por sua vez, é a
principal fonte da força que impulsiona o sangue pela circulação
pulmonar ou pela periférica.
As válvulas cardíacas orientam o percurso do sangue no
coração.
Válvulas Cardíacas
As válvulas A-V (tricúspide e mitral) impedem o refluxo de sangue dos ventrículos para as aurículas durante a sístole e as válvulas semilunares (válvulas aórtica e pulmonar) impedem o refluxo das artérias aorta e pulmonar para os ventrículos durante a diástole. Todas essas válvulas fecham-se e abrem-se passivamente. Isso quer dizer que elas fecham-se quando um gradiente retrógrado de pressão empurra o sangue para trás e abrem-se quando um gradiente de pressão anterógrado força o sangue a seguir em frente. Por razões anatómicas, o fecho das delgadas e ténues válvulas A-V não requer quase nenhum refluxo, enquanto o das muito mais pesadas válvulas semilunares requer um refluxo bastante forte por alguns milissegundos.
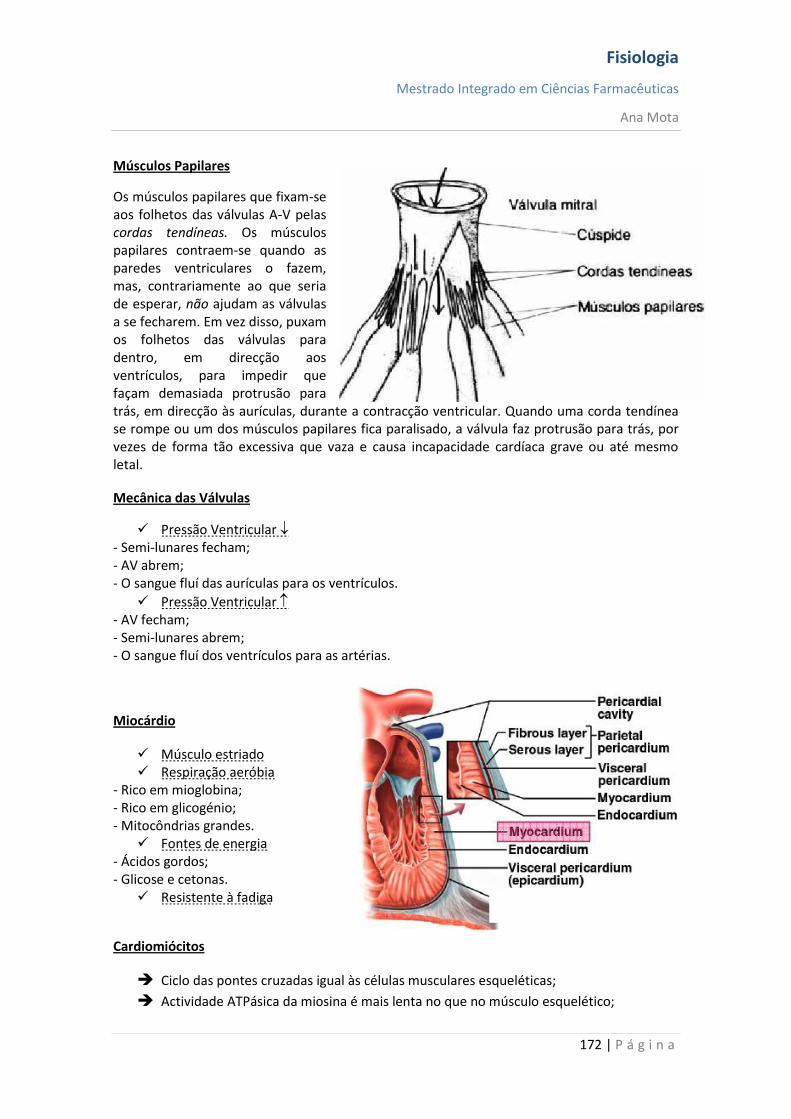
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
172 | P á g i n a
Músculos Papilares
Os músculos papilares que fixam-se aos folhetos das válvulas A-V pelas cordas tendíneas. Os músculos papilares contraem-se quando as paredes ventriculares o fazem, mas, contrariamente ao que seria de esperar, não ajudam as válvulas a se fecharem. Em vez disso, puxam os folhetos das válvulas para dentro, em direcção aos ventrículos, para impedir que façam demasiada protrusão para trás, em direcção às aurículas, durante a contracção ventricular. Quando uma corda tendínea se rompe ou um dos músculos papilares fica paralisado, a válvula faz protrusão para trás, por vezes de forma tão excessiva que vaza e causa incapacidade cardíaca grave ou até mesmo letal.
Mecânica das Válvulas
Pressão Ventricular - Semi-lunares fecham; - AV abrem; - O sangue fluí das aurículas para os ventrículos.
Pressão Ventricular - AV fecham; - Semi-lunares abrem; - O sangue fluí dos ventrículos para as artérias.
Miocárdio
Músculo estriado Respiração aeróbia
- Rico em mioglobina; - Rico em glicogénio; - Mitocôndrias grandes. Fontes de energia
- Ácidos gordos; - Glicose e cetonas. Resistente à fadiga
Cardiomiócitos
Ciclo das pontes cruzadas igual às células musculares esqueléticas;
Actividade ATPásica da miosina é mais lenta no que no músculo esquelético;
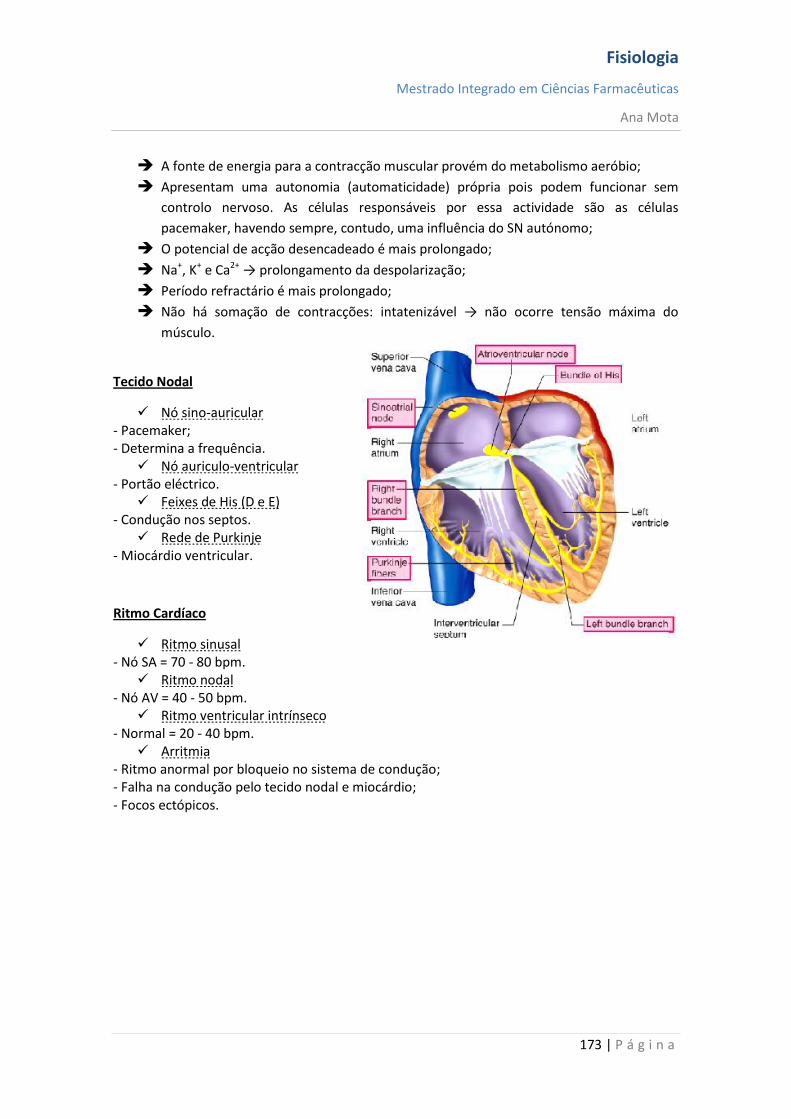
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
173 | P á g i n a
A fonte de energia para a contracção muscular provém do metabolismo aeróbio;
Apresentam uma autonomia (automaticidade) própria pois podem funcionar sem
controlo nervoso. As células responsáveis por essa actividade são as células
pacemaker, havendo sempre, contudo, uma influência do SN autónomo;
O potencial de acção desencadeado é mais prolongado;
Na+, K+ e Ca2+ → prolongamento da despolarização;
Período refractário é mais prolongado;
Não há somação de contracções: intatenizável → não ocorre tensão máxima do
músculo.
Tecido Nodal
Nó sino-auricular - Pacemaker; - Determina a frequência. Nó auriculo-ventricular
- Portão eléctrico. Feixes de His (D e E)
- Condução nos septos. Rede de Purkinje
- Miocárdio ventricular.
Ritmo Cardíaco
Ritmo sinusal - Nó SA = 70 - 80 bpm. Ritmo nodal
- Nó AV = 40 - 50 bpm. Ritmo ventricular intrínseco
- Normal = 20 - 40 bpm. Arritmia
- Ritmo anormal por bloqueio no sistema de condução; - Falha na condução pelo tecido nodal e miocárdio; - Focos ectópicos.
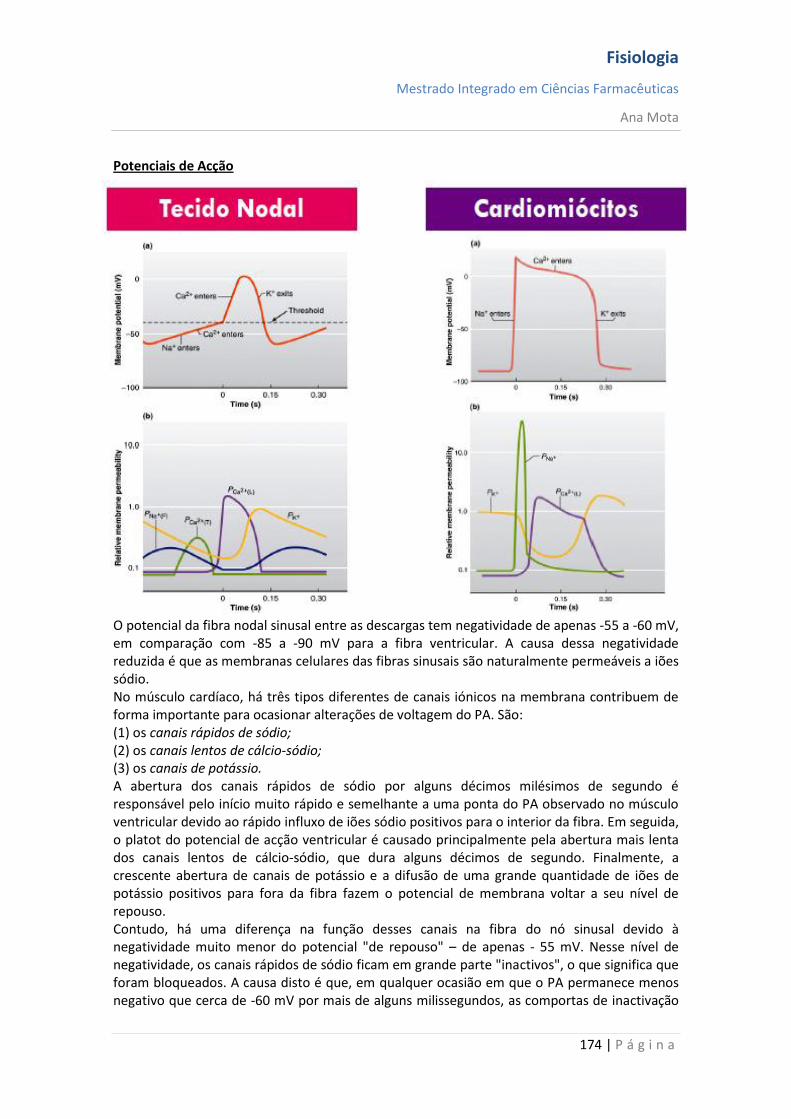
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
174 | P á g i n a
Potenciais de Acção
O potencial da fibra nodal sinusal entre as descargas tem negatividade de apenas -55 a -60 mV, em comparação com -85 a -90 mV para a fibra ventricular. A causa dessa negatividade reduzida é que as membranas celulares das fibras sinusais são naturalmente permeáveis a iões sódio. No músculo cardíaco, há três tipos diferentes de canais iónicos na membrana contribuem de forma importante para ocasionar alterações de voltagem do PA. São: (1) os canais rápidos de sódio; (2) os canais lentos de cálcio-sódio; (3) os canais de potássio. A abertura dos canais rápidos de sódio por alguns décimos milésimos de segundo é responsável pelo início muito rápido e semelhante a uma ponta do PA observado no músculo ventricular devido ao rápido influxo de iões sódio positivos para o interior da fibra. Em seguida, o platot do potencial de acção ventricular é causado principalmente pela abertura mais lenta dos canais lentos de cálcio-sódio, que dura alguns décimos de segundo. Finalmente, a crescente abertura de canais de potássio e a difusão de uma grande quantidade de iões de potássio positivos para fora da fibra fazem o potencial de membrana voltar a seu nível de repouso. Contudo, há uma diferença na função desses canais na fibra do nó sinusal devido à negatividade muito menor do potencial "de repouso" – de apenas - 55 mV. Nesse nível de negatividade, os canais rápidos de sódio ficam em grande parte "inactivos", o que significa que foram bloqueados. A causa disto é que, em qualquer ocasião em que o PA permanece menos negativo que cerca de -60 mV por mais de alguns milissegundos, as comportas de inactivação

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
175 | P á g i n a
no interior da membrana celular que fecham esses canais fecham-se e permanecem fechadas. Por esta razão, só os canais lentos de cálcio-sódio podem abrir-se (ou seja, podem ficar "activos") e, portanto, causar o potencial de acção. O PA tem, pois, desenvolvimento mais lento que o do músculo ventricular e também se recupera por diminuição lenta do potencial, em vez da recuperação abrupta que ocorre no caso da fibra ventricular. Contracção dos Cardiomiócitos
O músculo cardíaco, como todos os tecidos excitáveis, é refractário a reestimulação durante o PA. Por esta razão, o período refractário do coração é o intervalo de tempo durante o qual um impulso cardíaco normal não pode reexcitar uma área já excitada do músculo cardíaco. O período refratário normal do ventrículo é de 0,25 a 0,3 s, que é aproximadamente a duração do PA. Há um período refractário relativo adicional de cerca de 0,5 s, durante o qual o músculo é mais difícil de excitar que o normal mas, ainda assim, pode ser excitado. O período refractário do músculo auricular é muito mais curto que o dos ventrículos (cerca de 0,15 s) e o período refractário relativo tem mais de 0,03 s. Assim sendo, a frequência rítmica de contracção das aurículas pode ser muito mais rápida que a dos ventrículos.
Condução dos Potenciais de Acção
O músculo cardíaco começa a contrair alguns milissegundos após o início de um PA, continuando a contrair-se por alguns milissegundos após o término desse potencial. Por esta razão, a duração de contracção do músculo cardíaco é função principalmente da duração do PA — cerca de 0,2 s no músculo auricular e 0,3 s no músculo ventricular.
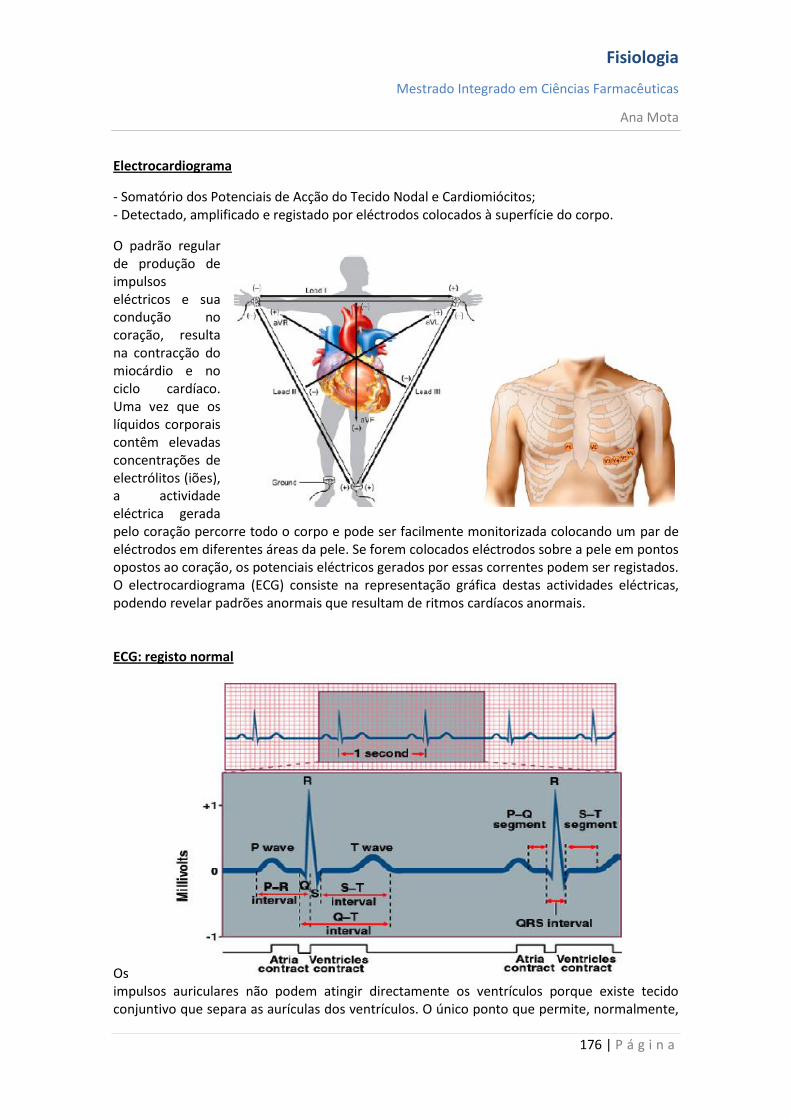
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
176 | P á g i n a
Electrocardiograma
- Somatório dos Potenciais de Acção do Tecido Nodal e Cardiomiócitos; - Detectado, amplificado e registado por eléctrodos colocados à superfície do corpo.
O padrão regular de produção de impulsos eléctricos e sua condução no coração, resulta na contracção do miocárdio e no ciclo cardíaco. Uma vez que os líquidos corporais contêm elevadas concentrações de electrólitos (iões), a actividade eléctrica gerada pelo coração percorre todo o corpo e pode ser facilmente monitorizada colocando um par de eléctrodos em diferentes áreas da pele. Se forem colocados eléctrodos sobre a pele em pontos opostos ao coração, os potenciais eléctricos gerados por essas correntes podem ser registados. O electrocardiograma (ECG) consiste na representação gráfica destas actividades eléctricas, podendo revelar padrões anormais que resultam de ritmos cardíacos anormais.
ECG: registo normal
Os impulsos auriculares não podem atingir directamente os ventrículos porque existe tecido conjuntivo que separa as aurículas dos ventrículos. O único ponto que permite, normalmente,

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
177 | P á g i n a
a passagem do impulso é o nó aurículo-ventricular, localizado na aurícula direita junto da válvula tricúspide. A partir deste nó o impulso atravessa o feixe de His. As propriedades electrofisiológicas do nó AV são semelhantes às do nó sinusal. O nó AV pode funcionar como um pacemaker, quando o nó sinusal deixa de funcionar. Outra função do nó AV é atrasar a velocidade com que o impulso eléctrico chega aos ventrículos, assegurando que estes estão relaxados quando se dá a contracção auricular. O impulso despolariza, então, o nó AV, o feixe de His, os ramos do feixe, a rede de Purkinje e o miocárdio ventricular. A propagação do impulso através do nó AV e feixe de His/sistema de Purkinje não é registada no ECG e ocorre durante o segmento isoeléctrico PR. A despolarização do músculo ventricular produz o complexo QRS. As células presentes no feixe de His e seus ramos são as características células de Purkinje. Estas estão adaptadas para a condução rápida do impulso eléctrico. A despolarização auricular é seguida pela sua repolarização (onda Tp ou Ta), mas esta normalmente não é evidente no electrocardiograma. Contudo, a repolarização ventricular, que se segue ao complexo QRS, é claramente registada no ECG, originando a onda T. Deve notar-se que a despolarização eléctrica do miocárdio auricular e ventricular não é sinónima de contracção auricular e ventricular. Em geral, a despolarização destas estruturas deve preceder a contracção mecânica correspondente.
Onda P Despolarização das aurículas Complexo QRS Despolarização dos ventrículos Repolarização das aurículas Onda T Repolarização dos ventrículos
ECG: registos anormais
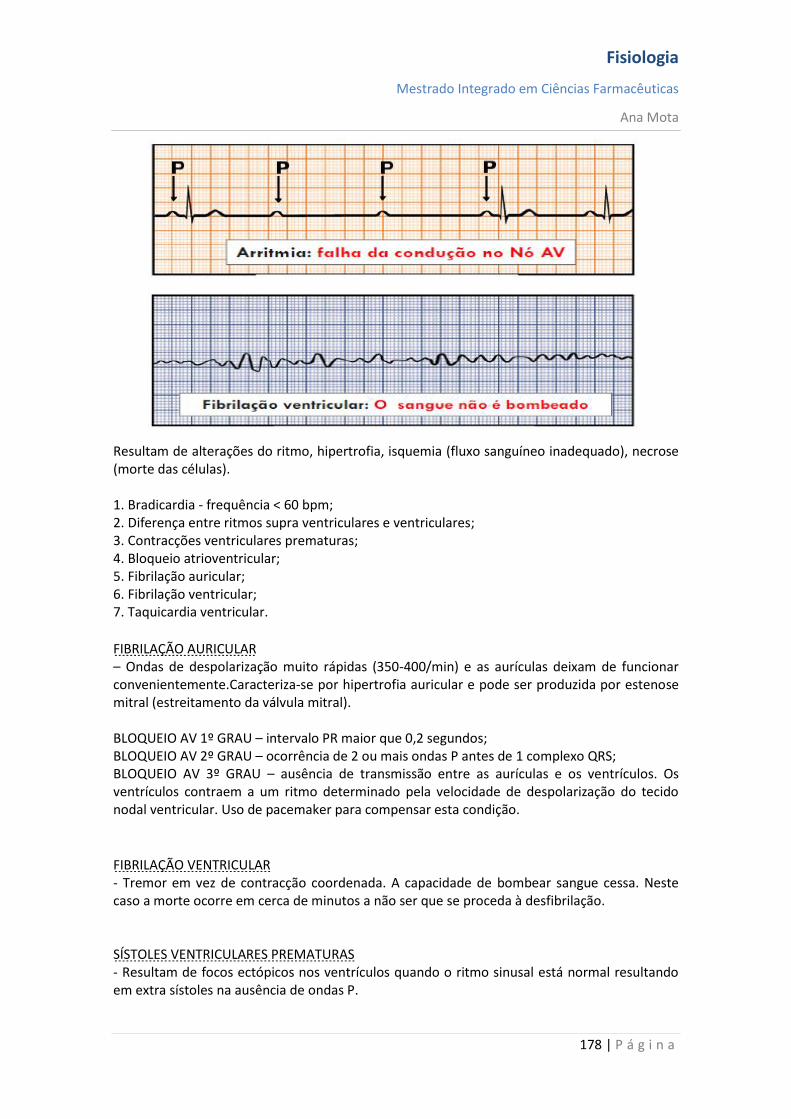
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
178 | P á g i n a
Resultam de alterações do ritmo, hipertrofia, isquemia (fluxo sanguíneo inadequado), necrose (morte das células). 1. Bradicardia - frequência < 60 bpm; 2. Diferença entre ritmos supra ventriculares e ventriculares; 3. Contracções ventriculares prematuras; 4. Bloqueio atrioventricular; 5. Fibrilação auricular; 6. Fibrilação ventricular; 7. Taquicardia ventricular.
FIBRILAÇÃO AURICULAR – Ondas de despolarização muito rápidas (350-400/min) e as aurículas deixam de funcionar convenientemente.Caracteriza-se por hipertrofia auricular e pode ser produzida por estenose mitral (estreitamento da válvula mitral). BLOQUEIO AV 1º GRAU – intervalo PR maior que 0,2 segundos; BLOQUEIO AV 2º GRAU – ocorrência de 2 ou mais ondas P antes de 1 complexo QRS; BLOQUEIO AV 3º GRAU – ausência de transmissão entre as aurículas e os ventrículos. Os ventrículos contraem a um ritmo determinado pela velocidade de despolarização do tecido nodal ventricular. Uso de pacemaker para compensar esta condição.
FIBRILAÇÃO VENTRICULAR - Tremor em vez de contracção coordenada. A capacidade de bombear sangue cessa. Neste caso a morte ocorre em cerca de minutos a não ser que se proceda à desfibrilação. SÍSTOLES VENTRICULARES PREMATURAS - Resultam de focos ectópicos nos ventrículos quando o ritmo sinusal está normal resultando em extra sístoles na ausência de ondas P.
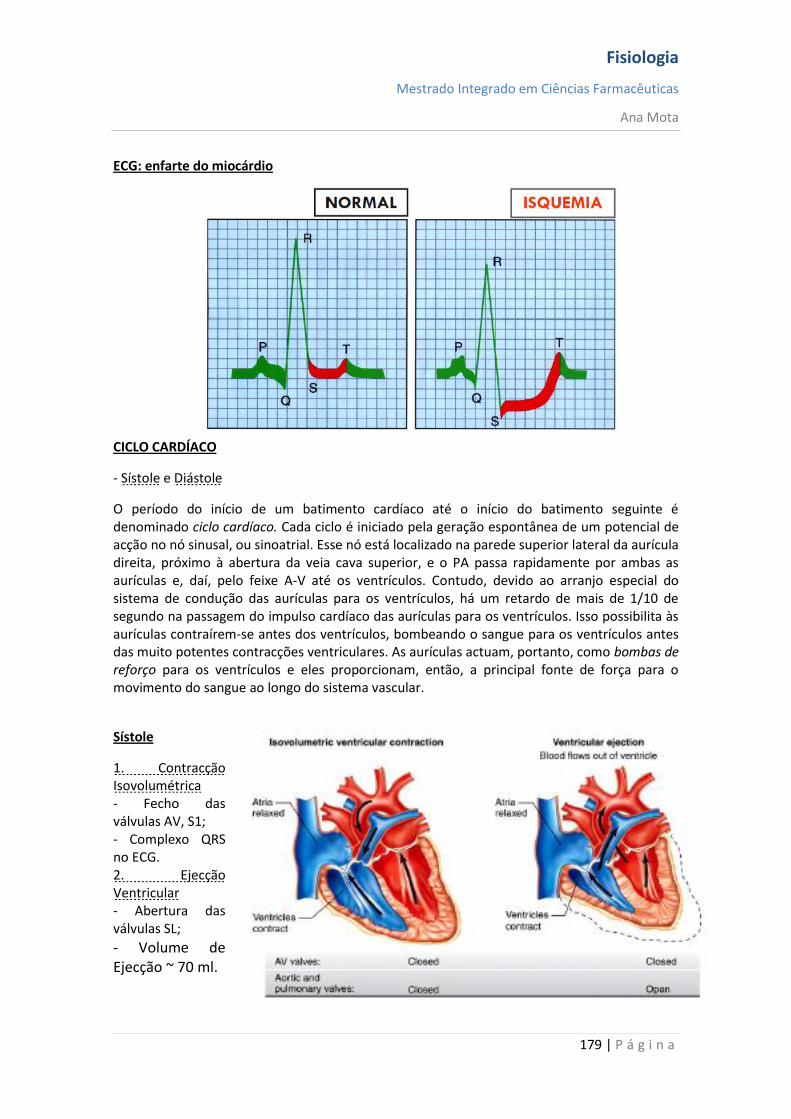
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
179 | P á g i n a
ECG: enfarte do miocárdio
CICLO CARDÍACO
- Sístole e Diástole
O período do início de um batimento cardíaco até o início do batimento seguinte é denominado ciclo cardíaco. Cada ciclo é iniciado pela geração espontânea de um potencial de acção no nó sinusal, ou sinoatrial. Esse nó está localizado na parede superior lateral da aurícula direita, próximo à abertura da veia cava superior, e o PA passa rapidamente por ambas as aurículas e, daí, pelo feixe A-V até os ventrículos. Contudo, devido ao arranjo especial do sistema de condução das aurículas para os ventrículos, há um retardo de mais de 1/10 de segundo na passagem do impulso cardíaco das aurículas para os ventrículos. Isso possibilita às aurículas contraírem-se antes dos ventrículos, bombeando o sangue para os ventrículos antes das muito potentes contracções ventriculares. As aurículas actuam, portanto, como bombas de reforço para os ventrículos e eles proporcionam, então, a principal fonte de força para o movimento do sangue ao longo do sistema vascular.
Sístole
1. Contracção Isovolumétrica - Fecho das válvulas AV, S1; - Complexo QRS no ECG. 2. Ejecção Ventricular - Abertura das válvulas SL;
- Volume de Ejecção ~ 70 ml.
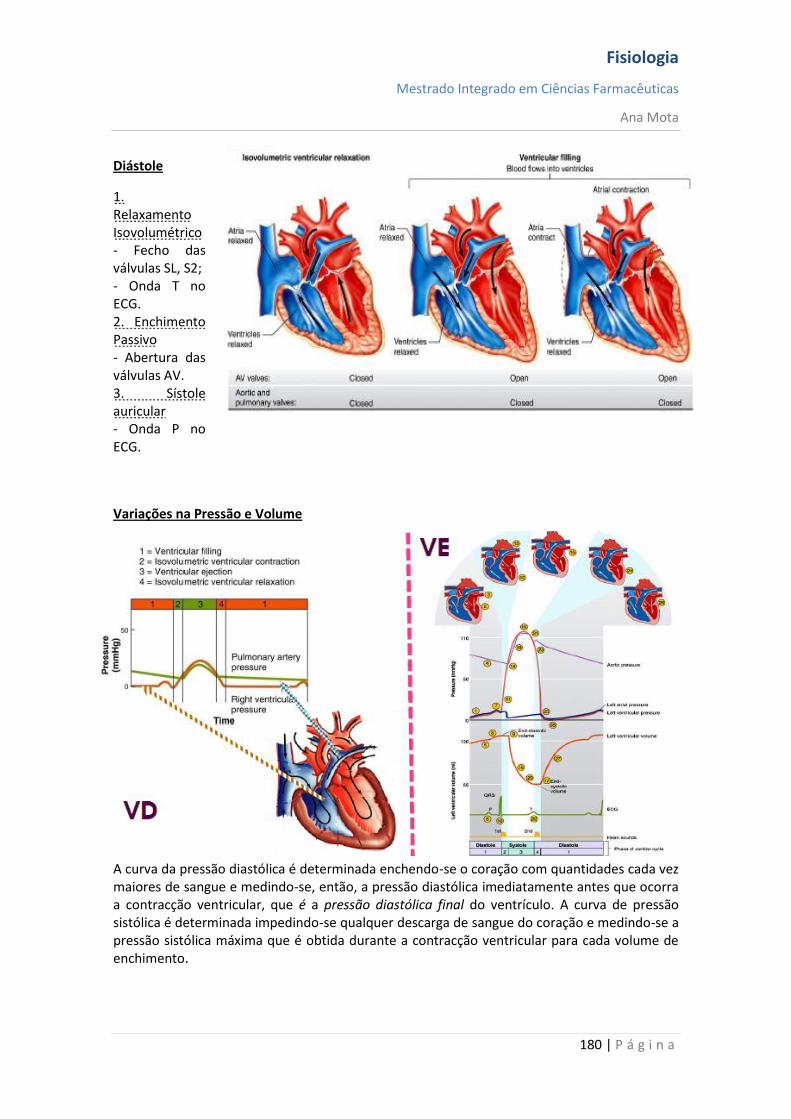
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
180 | P á g i n a
Diástole
1. Relaxamento Isovolumétrico - Fecho das válvulas SL, S2; - Onda T no ECG. 2. Enchimento Passivo - Abertura das válvulas AV. 3. Sístole auricular - Onda P no ECG.
Variações na Pressão e Volume
A curva da pressão diastólica é determinada enchendo-se o coração com quantidades cada vez maiores de sangue e medindo-se, então, a pressão diastólica imediatamente antes que ocorra a contracção ventricular, que é a pressão diastólica final do ventrículo. A curva de pressão sistólica é determinada impedindo-se qualquer descarga de sangue do coração e medindo-se a pressão sistólica máxima que é obtida durante a contracção ventricular para cada volume de enchimento.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
181 | P á g i n a
Fica muito claro que a pressão diastólica não aumenta muito até que o volume ventricular se eleve acima de 150 ml, aproximadamente. Até este volume, portanto, o sangue pode fluir facilmente da aurícula para o ventrículo. Acima de 150 ml, a pressão diastólica passa a aumentar rapidamente, em parte porque o tecido fibroso do coração não se distende mais e, em parte, porque o pericárdio que circunda o coração fica distendido praticamente até o seu limite. Durante a contracção ventricular, a pressão sistólica aumenta rapidamente com volumes ventriculares crescentes, atingindo seu máximo com o volume ventricular de 150 a 170 ml. Em seguida, quando o volume aumenta ainda mais, a pressão sistólica em algumas condições até diminui, porque nesses volumes muito grandes os filamentos de actina e miosina das fibras musculares cardíacas encontram-se de facto separados o suficiente para que a força de contracção das fibras fique menos que óptima. A pressão sistólica máxima para o ventrículo esquerdo normal não estimulado está
entre 250 e 300 mm Hg, mas isso varia muito com a força do coração. Para o ventrículo direito normal, fica entre 60 e 80 mm Hg.
Circulação Coronária
As principais artérias coronárias encontram-se na superfície do coração e só as pequenas artérias penetram na massa muscular cardíaca. É quase totalmente por essas artérias que o coração recebe seu suprimento sanguíneo nutriente. Apenas o décimo de milímetro mais interior da superfície endocárdica podem obter nutrição significativa diretamente do sangue nas câmaras cardíacas. A artéria coronária esquerda supre principalmente as partes anterior e lateral do ventrículo esquerdo, enquanto a artéria coronária direita supre a maior parte do ventrículo direito, assim como a parte posterior do ventrículo esquerdo em 80 a 90% de todas as pessoas. Em cerca de metade dos seres humanos, há um maior fluxo sanguíneo pela artéria coronária direita que

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
182 | P á g i n a
pela esquerda; em cerca de 30% deles, as artérias são aproximadamente iguais; e em 20% a artéria esquerda predomina. A maior parte do fluxo sanguíneo venoso do ventrículo esquerdo sai por meio do seio coronário — o que constitui cerca de 75% do fluxo sanguíneo coronário total — e a maior parte do sangue venoso do ventrículo direito flui pelas pequenas veias cardíacas anteriores directamente até à aurícula direita, não tendo ligação com o seio coronário. Uma pequena quantidade do sangue coronário reflui para o coração pelas diminutas veias tebésicas, que desembocam directamente em todas as câmaras cardíacas.
Fluxo Coronário
O fluxo sanguíneo coronário em repouso nos seres humanos é, em média, cerca de 225 ml/min, o que representa cerca de 0,7 a 0,8 ml/g de músculo cardíaco ou 4 a 5% do débito cardíaco total. Nos exercícios vigorosos, o coração aumenta seu débito cardíaco até quatro a sete vezes, bombeando o sangue contra uma pressão arterial maior que a normal. Por conseguinte, o trabalho produzido pelo coração em condições severas pode aumentar por até seis a oito vezes. O fluxo sanguíneo coronário aumenta três a quatro vezes para suprir os nutrientes adicionais necessários ao coração. Evidentemente, esse aumento não é tão grande quanto o aumento da carga de trabalho, o que significa que a proporção entre o fluxo sanguíneo coronário e o gasto de energia pelo coração diminui. Entretanto, a "eficiência" da utilização cardíaca de energia aumenta para contrabalançar esta deficiência relativa do suprimento sanguíneo.
O fluxo sanguíneo no ventrículo esquerdo cai para um valor baixo durante a sístole, o contrário do fluxo em outros leitos vasculares do corpo. A razão disso é a forte compressão do músculo ventricular esquerdo em volta dos vasos intramusculares durante a sístole. Durante a diástole, o músculo cardíaco se relaxa totalmente e não mais obstrui o fluxo sanguíneo pelos capilares do ventrículo esquerdo, de modo que o sangue flui rapidamente durante toda a diástole. O fluxo sanguíneo pelos capilares coronários do ventrículo direito também sofre alterações fásicas, durante o ciclo cardíaco, mas, como a força de contracção do ventrículo direito é bem menor que a do ventrículo esquerdo, as alterações fásicas inversas são apenas parciais, em contraste com as no ventrículo esquerdo.
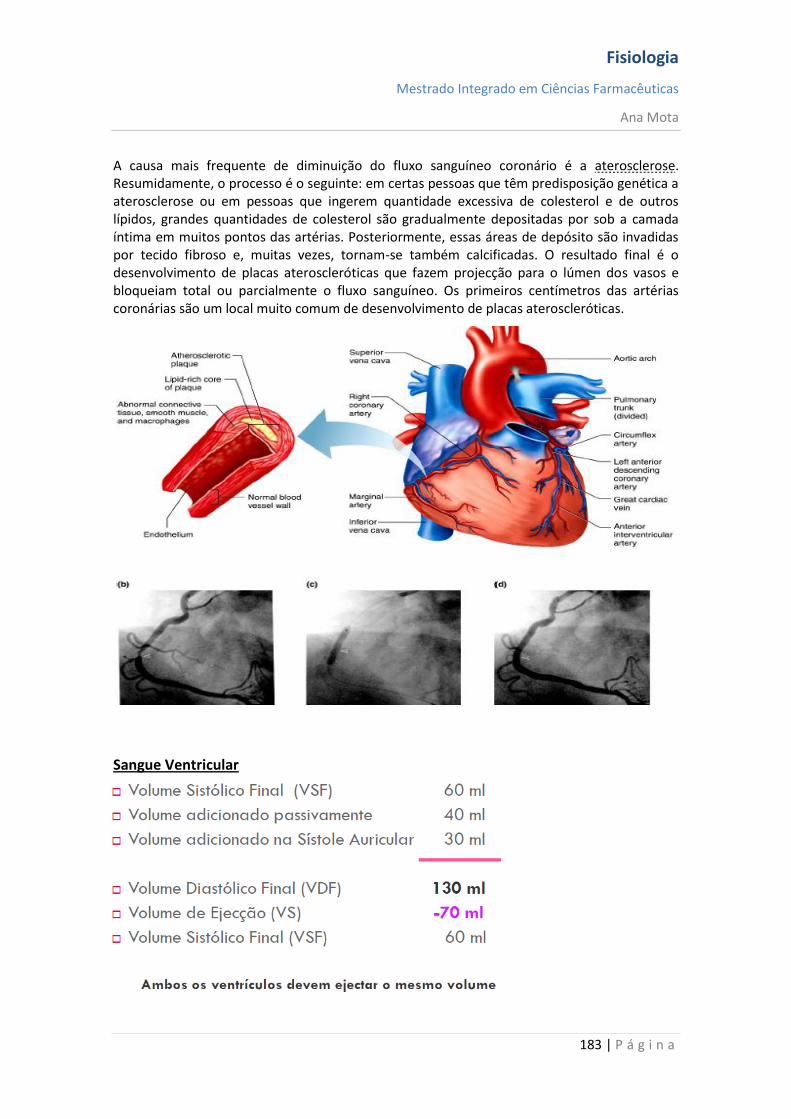
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
183 | P á g i n a
A causa mais frequente de diminuição do fluxo sanguíneo coronário é a aterosclerose. Resumidamente, o processo é o seguinte: em certas pessoas que têm predisposição genética a aterosclerose ou em pessoas que ingerem quantidade excessiva de colesterol e de outros lípidos, grandes quantidades de colesterol são gradualmente depositadas por sob a camada íntima em muitos pontos das artérias. Posteriormente, essas áreas de depósito são invadidas por tecido fibroso e, muitas vezes, tornam-se também calcificadas. O resultado final é o desenvolvimento de placas ateroscleróticas que fazem projecção para o lúmen dos vasos e bloqueiam total ou parcialmente o fluxo sanguíneo. Os primeiros centímetros das artérias coronárias são um local muito comum de desenvolvimento de placas ateroscleróticas.
Sangue Ventricular
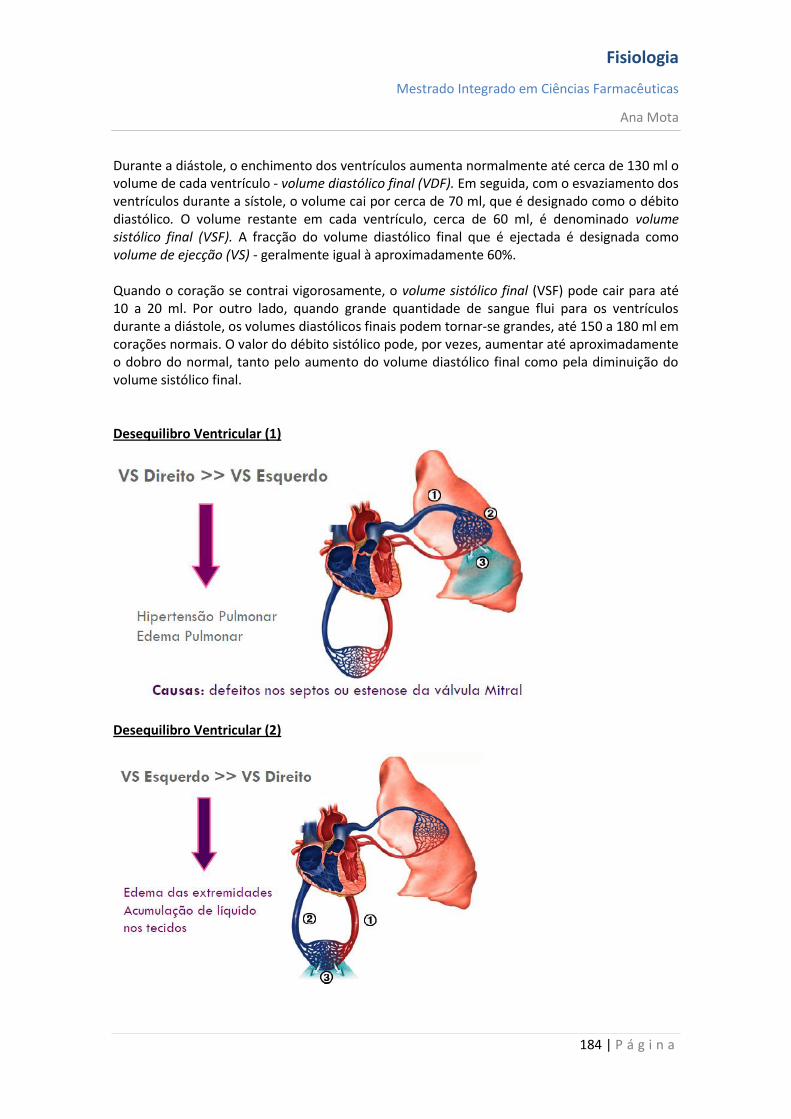
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
184 | P á g i n a
Durante a diástole, o enchimento dos ventrículos aumenta normalmente até cerca de 130 ml o volume de cada ventrículo - volume diastólico final (VDF). Em seguida, com o esvaziamento dos ventrículos durante a sístole, o volume cai por cerca de 70 ml, que é designado como o débito diastólico. O volume restante em cada ventrículo, cerca de 60 ml, é denominado volume sistólico final (VSF). A fracção do volume diastólico final que é ejectada é designada como volume de ejecção (VS) - geralmente igual à aproximadamente 60%. Quando o coração se contrai vigorosamente, o volume sistólico final (VSF) pode cair para até 10 a 20 ml. Por outro lado, quando grande quantidade de sangue flui para os ventrículos durante a diástole, os volumes diastólicos finais podem tornar-se grandes, até 150 a 180 ml em corações normais. O valor do débito sistólico pode, por vezes, aumentar até aproximadamente o dobro do normal, tanto pelo aumento do volume diastólico final como pela diminuição do volume sistólico final.
Desequilibro Ventricular (1)
Desequilibro Ventricular (2)
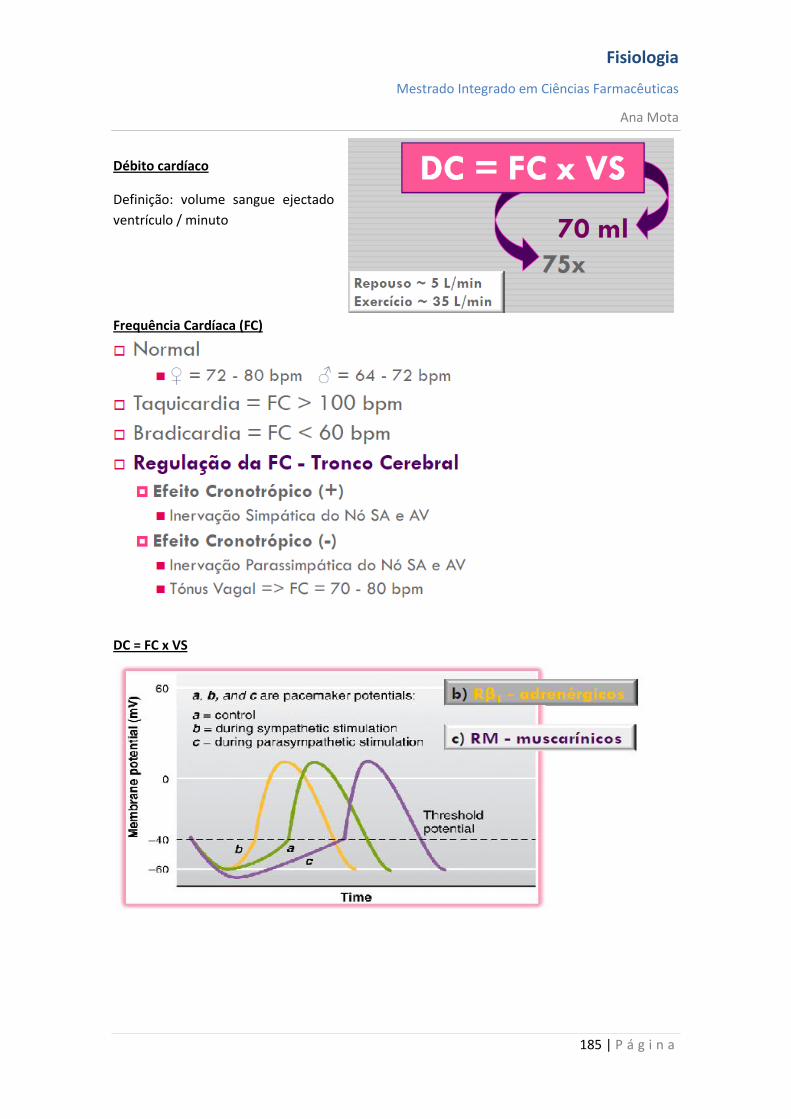
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
185 | P á g i n a
Débito cardíaco
Definição: volume sangue ejectado
ventrículo / minuto
Frequência Cardíaca (FC)
DC = FC x VS
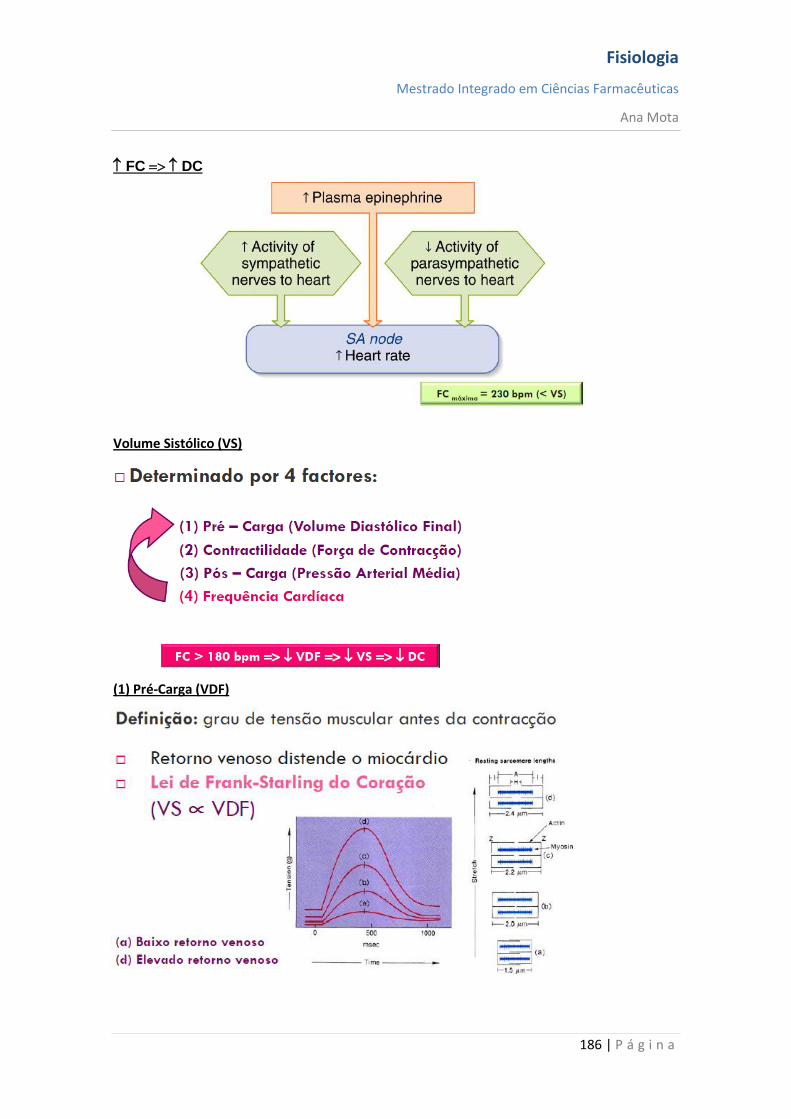
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
186 | P á g i n a
FC DC
Volume Sistólico (VS)
(1) Pré-Carga (VDF)

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
187 | P á g i n a
Lei de Frank-Starling
A quantidade de sangue bombeada pelo coração a cada minuto é determinada pela intensidade do fluxo sanguíneo das veias para o coração, o que é denominado retorno venoso. Isso quer dizer que cada tecido periférico do corpo controla seu próprio fluxo sanguíneo, e a soma de todos os fluxos sanguíneos locais por todos os tecidos periféricos volta à aurícula direita por meio das
veias. O coração, por sua vez, bombeia, automaticamente, esse sangue que chega para as artérias sistémicas, de modo que ele possa fluir novamente pelo circuito. Essa capacidade intrínseca de adaptação do coração à alteração no volume de sangue que entra é denominada mecanismo de Frank-Starling do coração. Basicamente, o mecanismo de Frank-Starling indica que quanto mais o coração se enche, durante a diástole, maior vai ser a quantidade de sangue bombeada para a aorta. Outra forma de expressar isto é: dentro dos limites fisiológicos, o coração bombeia todo o sangue que chega até ele, sem permitir acúmulo excessivo de sangue nas veias.
(2) Contractilidade
Contractilidade VS e DC

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
188 | P á g i n a
Stroke volume is increased at any given end-diastolic volume; that is, the sympathetic stimulation has increased ventricular contractility.
Efeitos do Simpático

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
189 | P á g i n a
(3) Pós-Carga (PAM)
CIRCULAÇÃO
- Circulação Arterial e Arteriolar - Circulação Capilar e Trocas de Substâncias - Circulação Venosa e Retorno Venoso
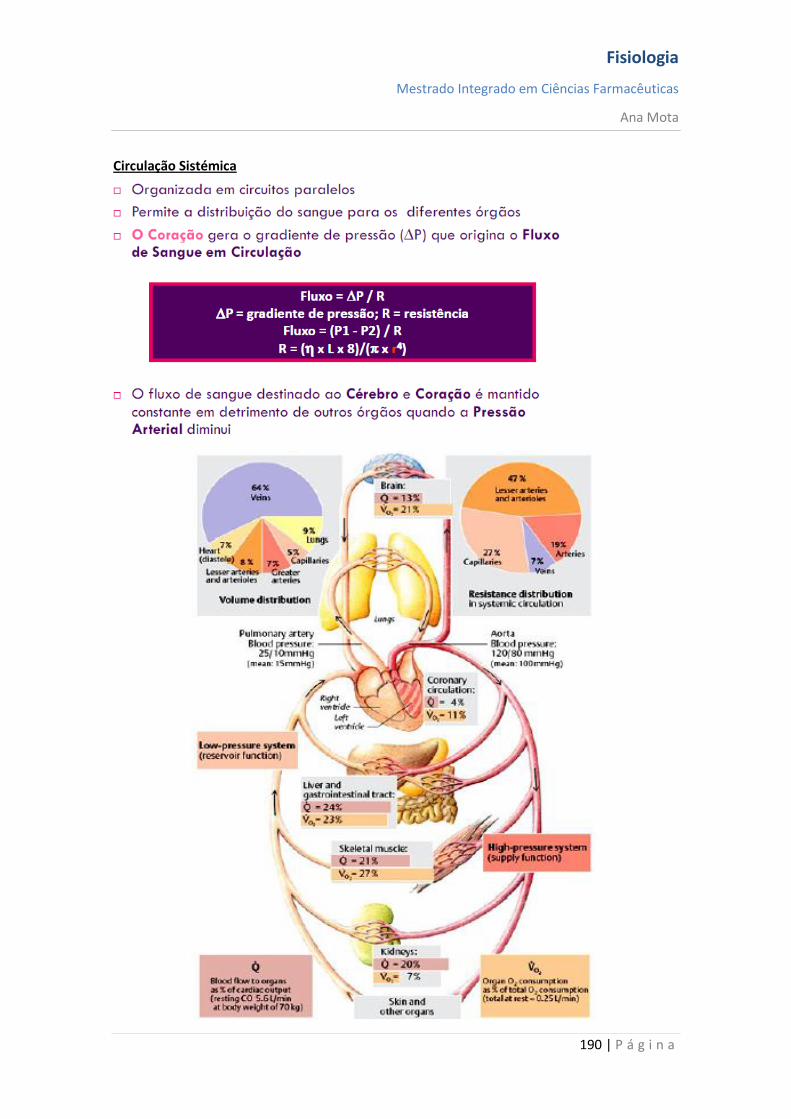
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
190 | P á g i n a
Circulação Sistémica
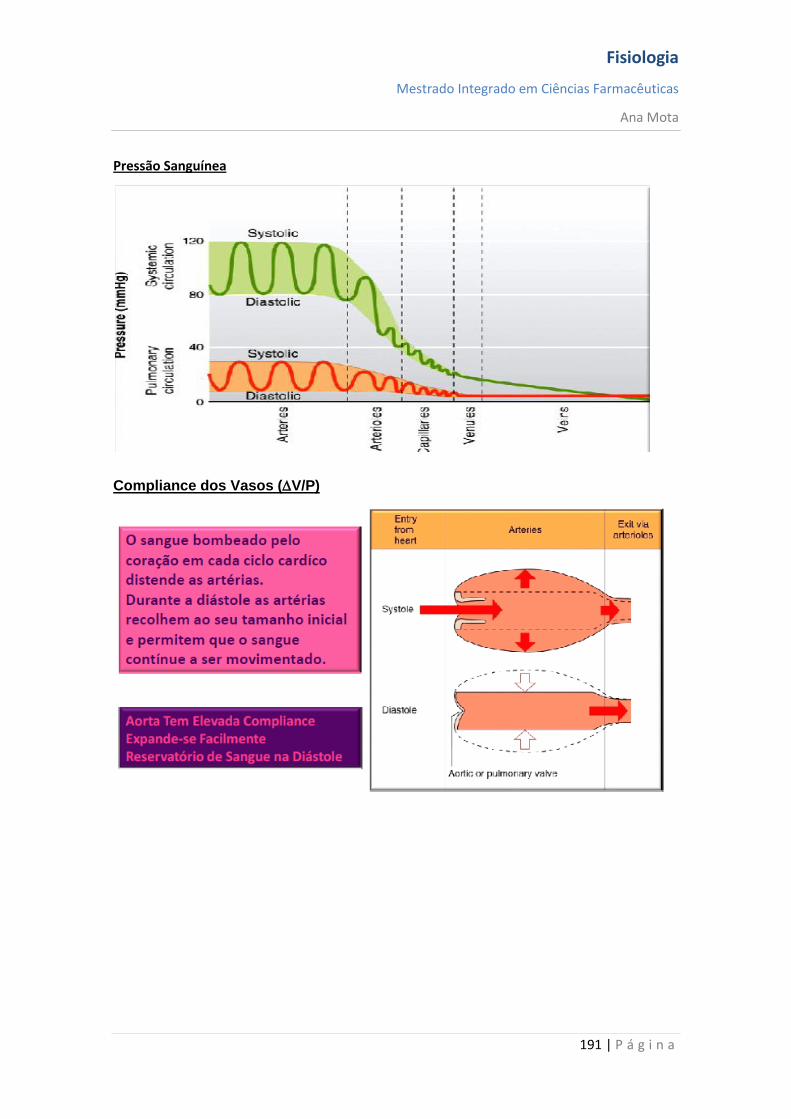
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
191 | P á g i n a
Pressão Sanguínea
Compliance dos Vasos (V/P)

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
192 | P á g i n a
Pressão Arterial
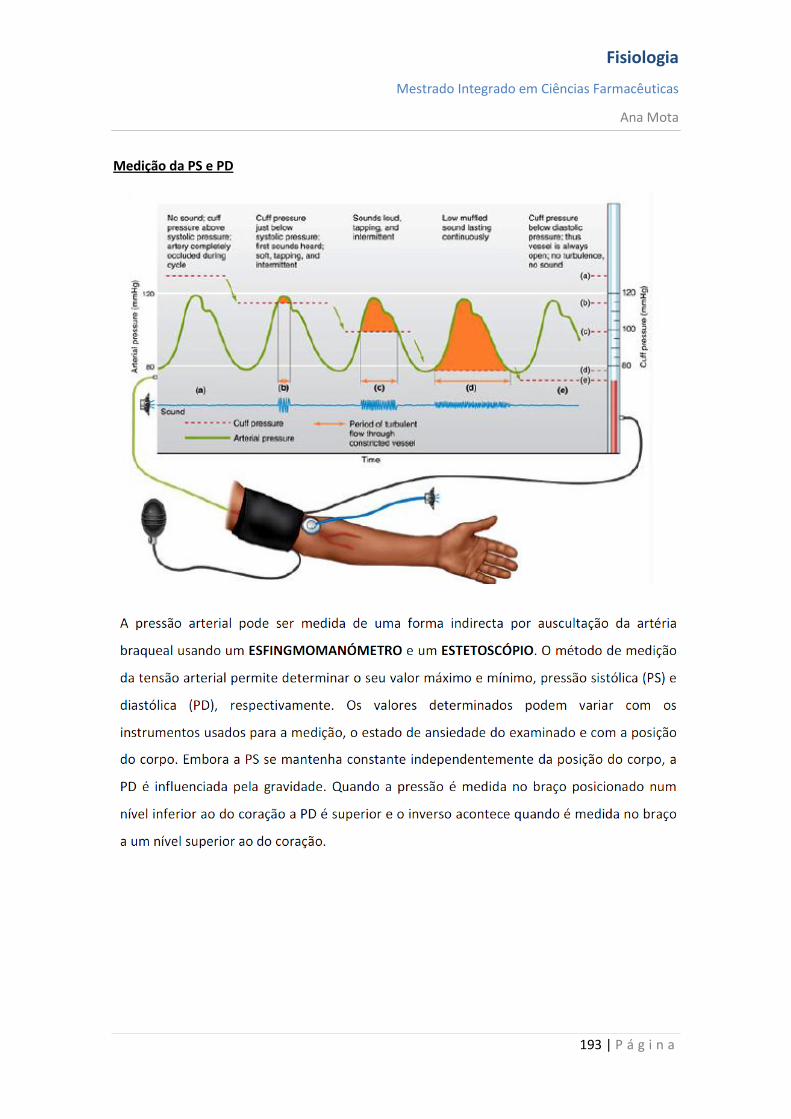
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
193 | P á g i n a
Medição da PS e PD
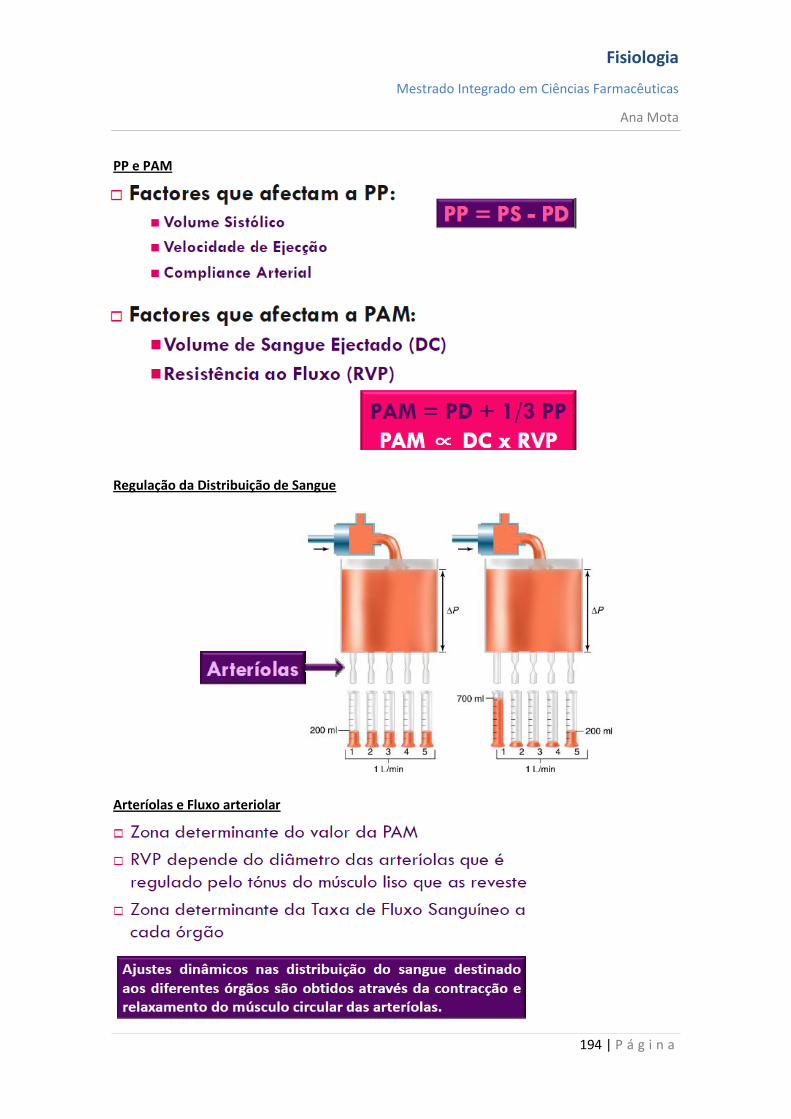
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
194 | P á g i n a
PP e PAM
Regulação da Distribuição de Sangue
Arteríolas e Fluxo arteriolar
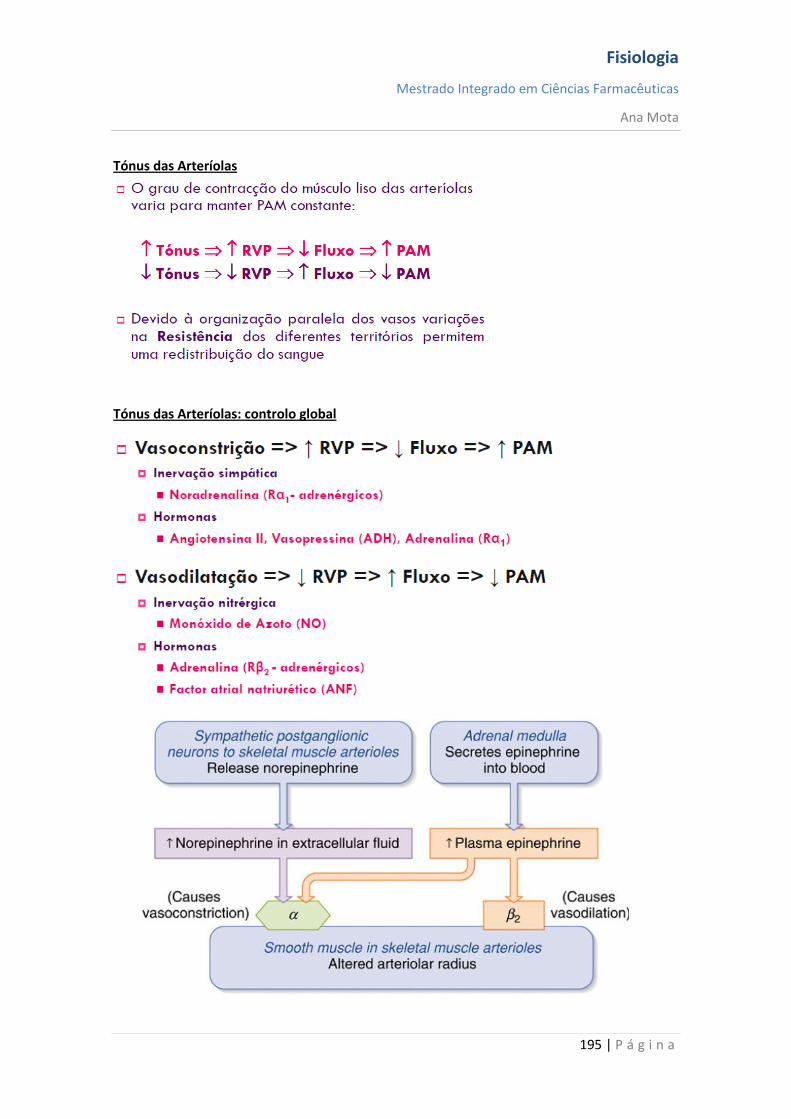
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
195 | P á g i n a
Tónus das Arteríolas
Tónus das Arteríolas: controlo global

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
196 | P á g i n a
Tónus das Arteríolas: controlo local

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
197 | P á g i n a
Agentes Autócrinos/Parácrinos
Endotélio: produção de NO

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
198 | P á g i n a
Regulação da RVP: territórios vasculares
Capilares
Rede Capilar

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
199 | P á g i n a
Fluxo Capilar
Transferência de Substâncias
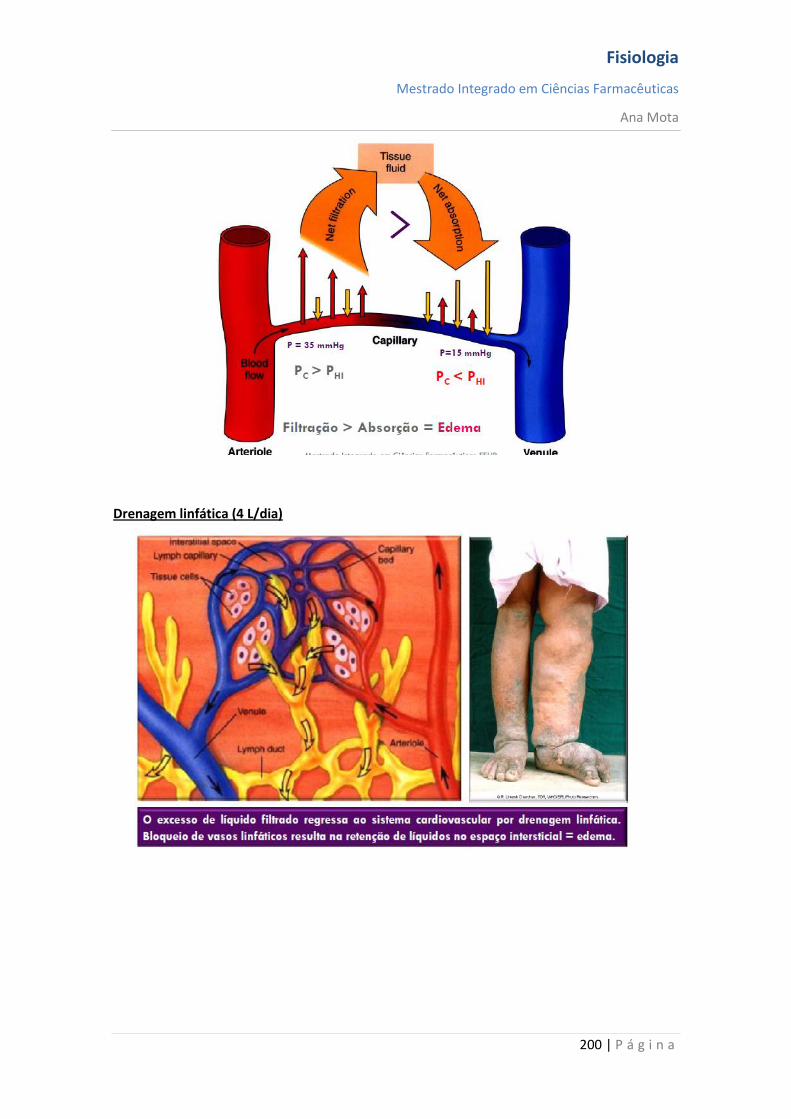
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
200 | P á g i n a
Drenagem linfática (4 L/dia)
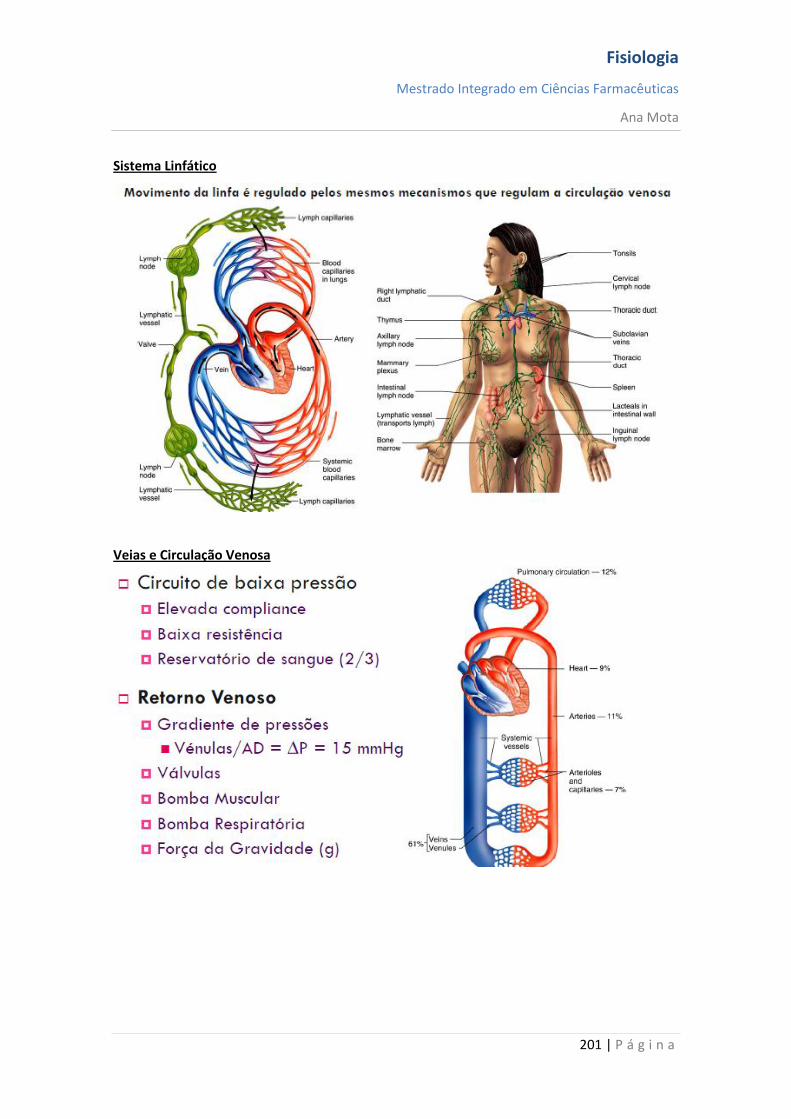
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
201 | P á g i n a
Sistema Linfático
Veias e Circulação Venosa

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
202 | P á g i n a
Retorno Venoso (VDF)
REGULAÇÃO CARDIOVASCULAR
Regulação da PAM - Mecanismos a curto prazo
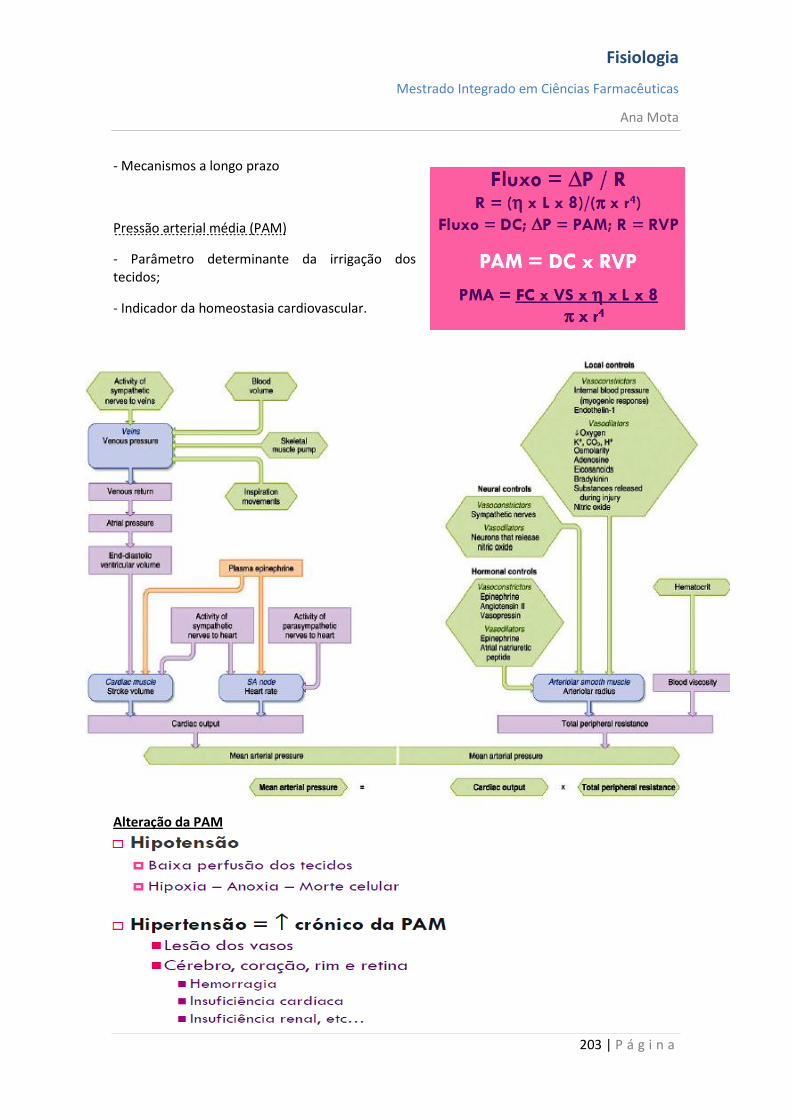
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
203 | P á g i n a
- Mecanismos a longo prazo
Pressão arterial média (PAM)
- Parâmetro determinante da irrigação dos tecidos;
- Indicador da homeostasia cardiovascular.
Alteração da PAM
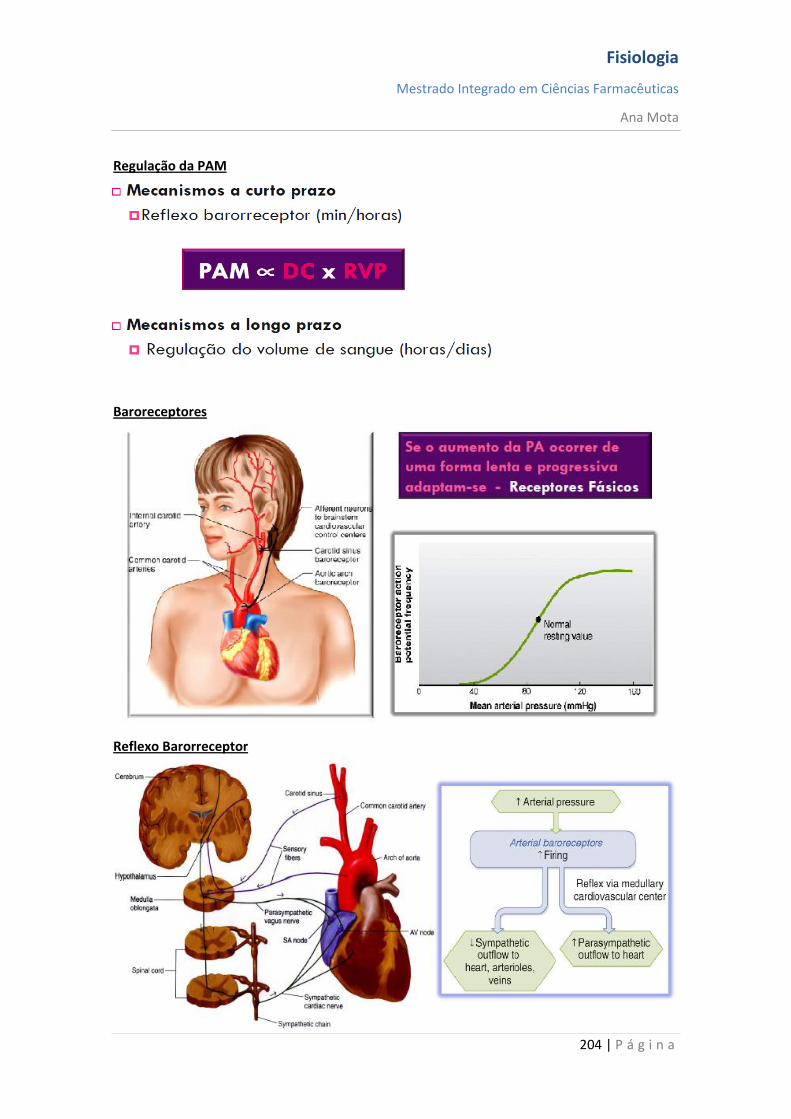
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
204 | P á g i n a
Regulação da PAM
Baroreceptores
Reflexo Barorreceptor
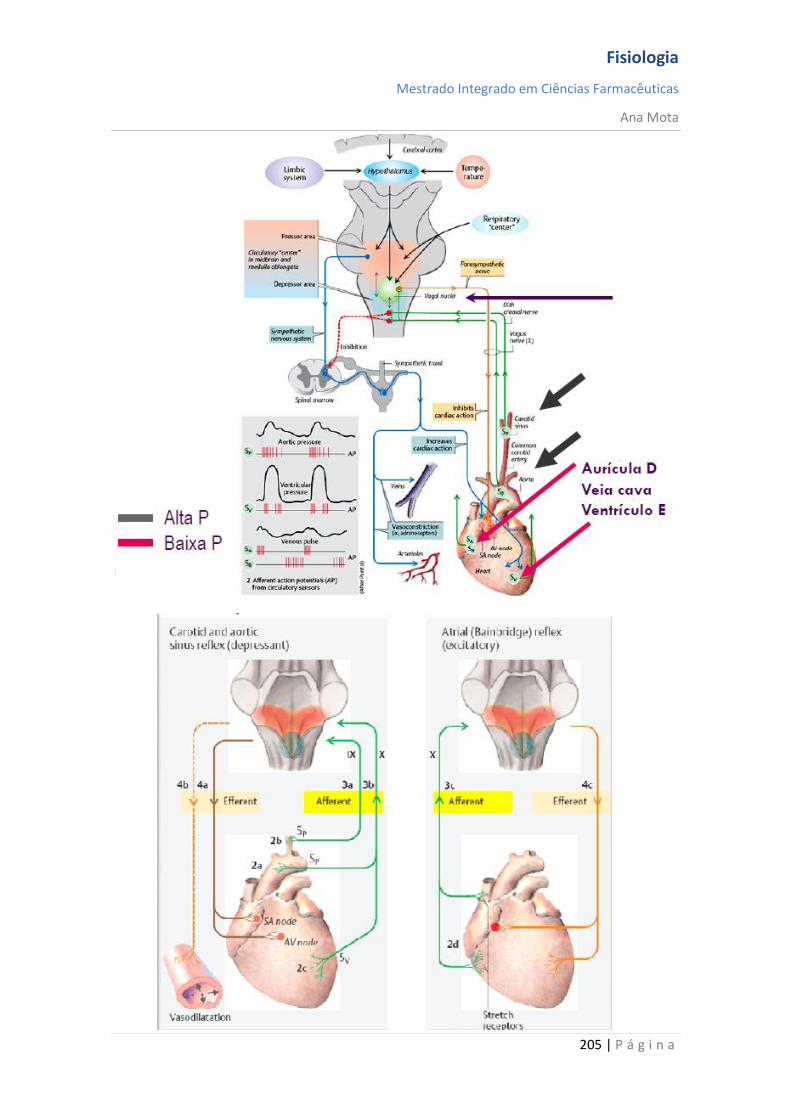
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
205 | P á g i n a

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
206 | P á g i n a
Regulação do Volume de Sangue
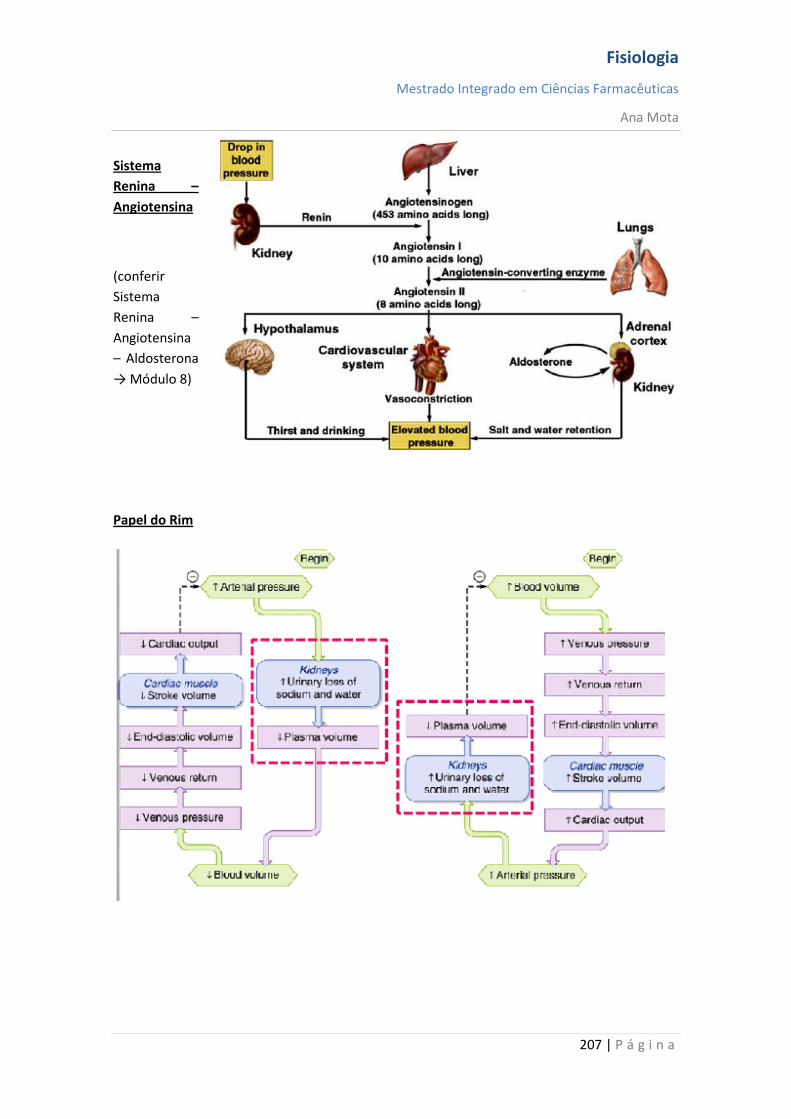
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
207 | P á g i n a
Sistema
Renina –
Angiotensina
(conferir
Sistema
Renina –
Angiotensina
– Aldosterona
→ Módulo 8)
Papel do Rim

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
208 | P á g i n a

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
209 | P á g i n a
Hipertensão Crónica
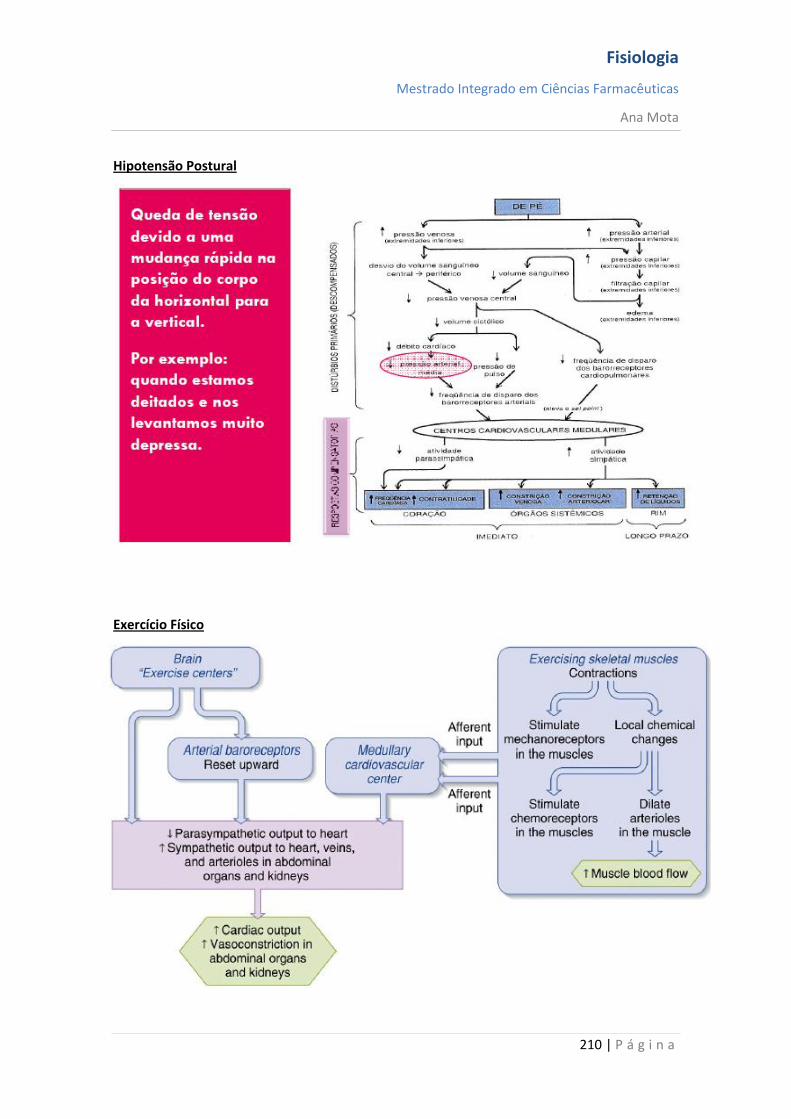
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
210 | P á g i n a
Hipotensão Postural
Exercício Físico
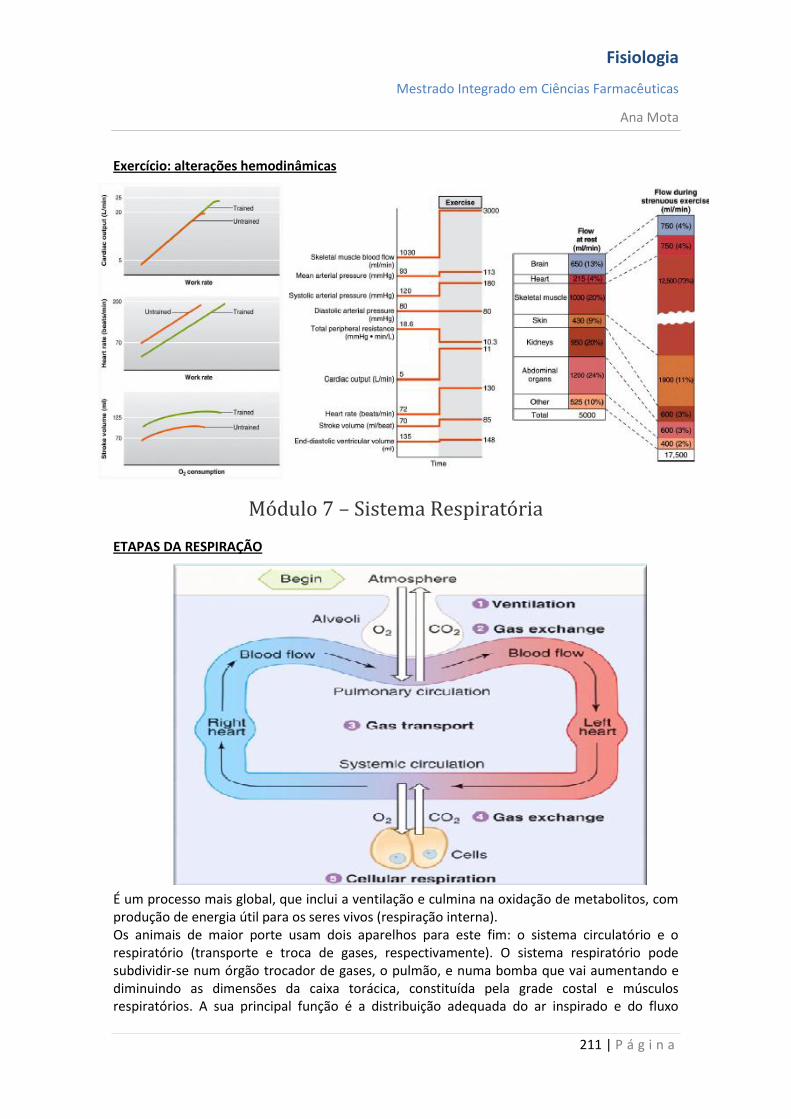
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
211 | P á g i n a
Exercício: alterações hemodinâmicas
Módulo 7 – Sistema Respiratória
ETAPAS DA RESPIRAÇÃO
É um processo mais global, que inclui a ventilação e culmina na oxidação de metabolitos, com produção de energia útil para os seres vivos (respiração interna). Os animais de maior porte usam dois aparelhos para este fim: o sistema circulatório e o respiratório (transporte e troca de gases, respectivamente). O sistema respiratório pode subdividir-se num órgão trocador de gases, o pulmão, e numa bomba que vai aumentando e diminuindo as dimensões da caixa torácica, constituída pela grade costal e músculos respiratórios. A sua principal função é a distribuição adequada do ar inspirado e do fluxo

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
212 | P á g i n a
sanguíneo pulmonar, para que as trocas gasosas se realizem com um mínimo de consumo energético.
Sistema Respiratório: funções
Sistema Respiratório: Anatomia
Os pulmões de um ser humano adulto pesam cerca de 1 Kg. No final da expiração normal, contêm um volume de ar (cerca de 2,4L) designado por Capacidade Residual Funcional (CRF). Após uma inspiração forçada, têm um volume máximo (cerca de 6L) que se designa por Capacidade Pulmonar Total (CPT).
Pulmões: constituição e funções

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
213 | P á g i n a
Vias Respiratórias
Da traqueia, via respiratória única, aos alvéolos, há 23 ramificações, sendo as 16 primeiras condutoras e as 7 últimas transicionais e de trocas. A estas ramificações, corresponde um aumento de área de secção transversa do sistema respiratório e uma consequente lentificação da velocidade do fluxo aéreo (velocidade de fluxo= débito/área de secção transversa). As vias mais centrais, condutoras, não participam nas trocas, constituindo o espaço morto anatómico, que é cerca de 30% em cada ventilação. Entre elas, encontram-se vias de maior calibre envolvidas por tecido cartilaginoso, que impede o seu colapso, e vias de menor calibre com um forte componente muscular, que lhes permite uma dilatação e constrição independente do volume pulmonar - os brônquios. Com as sucessivas ramificações seguem-se vias de calibre inferior a 1mm, que deixam de ter cartilagem - os bronquíolos. Estes encontram-se incrustados na rede de tecido conjuntivo pulmonar, dependendo o seu calibre do volume pulmonar, o que constitui uma importante diferença funcional em relação aos anteriores. Os bronquíolos respiratórios, cujas paredes são indefinidas, correspondem às aberturas dos alvéolos, sendo por isso designados ductos alveolares.
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
214 | P á g i n a
Vias Respiratórias: zona condutora
O epitélio das vias aéreas tem um componente ciliar e secretor ausente a partir dos bronquíolos.
Vias Respiratórias: zona respiratória
300 milhões de alvéolos
É adaptada para tornar a troca gasosa o mais eficiente possível.
As diferentes camadas que compõem a membrana respiratória são:
• Uma camada de líquido que recobre o alvéolo e contém surfactante, o qual reduz a tensão superficial do líquido alveolar;
• O epitélio alveolar, composto de delgadas células epiteliais;
• A membrana basal do epitélio;
• Um estreito espaço intersticial, situado entre o epitélio alveolar e o endotélio capilar;
Ventilação Pulmonar
- Inspiração e Expiração; - Alterações do Volume Pulmonar na Respiração; - Músculos Respiratórios.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
215 | P á g i n a
Lei de Boyle
As substâncias têm tendência a deslocarem-se da zona onde se encontram em maior concentração para as zonas onde se encontram em menor concentração. O mesmo se passa com o ar que se movimenta das zonas de maior pressão para as zonas de menor pressão. Pela lei de Boyle é também possível prever que um aumento de volume causa uma diminuição da pressão e vice-versa.
O ar movimenta-se de zonas de maior pressão para as zonas de menor pressão.
Usam-se 3 convenções para exprimir as pressões no sistema respiratório.
1ª – Pressão atmosférica, que corresponde à pressão atmosférica fora do corpo, à qual é atribuída o valor 0 cm de H2O;
2ª – Pequenas pressões na respiração fisiológica são normalmente expressas em centímetros de H2O (1cm de H2O = 0.74 cm Hg);
3ª – As restantes pressões são avaliadas em referência à pressão atmosférica.
Ventilação Pulmonar

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
216 | P á g i n a
Entende-se por ventilação o processo automático e rítmico que gera movimentos de entrada e saída de ar nas Unidades Respiratórias Terminais (vide infra).É regulado a nível central e depende da contracção e relaxamento de vários músculos: diafragma, músculos da grade costal e caixa torácica e músculos abdominais. Este processo permite a absorção de O2 e a remoção de CO2, pelo organismo, e é, por isso, designado respiração externa por muitos autores.
Pleuras: constituição e funções
A pleura é uma membrana dupla serosa que envolve os pulmões e é constituída por dois
folhetos:
Pleura parietal que recobre internamente a parede costal da cavidade, sendo
subdividida em quatro partes:
A pleura costal que cobre as faces internas da parede torácica;
A pleura mediastinal que cobre as faces laterais do mediastino;
A pleura diafragmática que cobre a face superior do diafragma;
A cúpula pleural que recobre a o ápex pulmonar.
Pleura visceral que recobre os pulmões. Entre os dois folhetos existe uma cavidade denominada cavidade pleural. Esta cavidade possui um líquido, líquido pleural que desempenha 3 funções essenciais:
Actua como lubrificante, permitindo o deslizamento das membranas, à medida que os pulmões e o tórax modificam a sua forma durante a respiração;
Ajuda a manter as membranas pleurais juntas. Provoca uma pressão negativa na cavidade pleural, o que é necessário para
manter os pulmões distendidos no seu nível de repouso;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
217 | P á g i n a
Deste modo, os dois pulmões aderem à parede torácica como se estivessem colados, embora possam deslizar livremente, quando bem lubrificados, à medida que o tórax se expande e se retrai.
Pressões Pulmonares
A pressão intrapleural refere-se à pressão existente no estreito espaço entre a pleura pulmonar e a pleura da parede torácica – a cavidade pleural. Nessa cavidade verifica-se uma pressão negativa. No início da inspiração, a pressão pleural normal é de aproximadamente -5 cm de
água, que é a quantidade de sucção necessária para manter os pulmões abertos em seu nível de repouso. A seguir, durante a inspiração normal, a expansão da caixa torácica traciona a superfície dos pulmões com maior força e cria pressão ainda mais negativa, atingindo valor médio de cerca de -7,5 cm de água. Conclui-se assim que durante a inspiração ocorre um decréscimo da pressão intrapleural que atinge valores mais negativos devido ao aumento do volume torácico.
A diminuição da pressão intrapleural durante a inspiração deve-se: Ao efeito da modificação do volume na pressão, quando o volume da cavidade
torácica aumenta a pressão pleural diminuiu; À medida que a cavidade torácica se expande, os pulmões acompanham-na, porque se
encontram aderidos à parede interna do tórax pela pleura; À medida que os pulmões se expandem, a tendência destes para se retrair aumenta,
gerando um maior efeito de sucção e uma diminuição da pressão intrapleural. Durante a expiração, pelo contrário verifica-se um aumento da pressão pleural que
passa de -7,5 para -5 cm de H2O devido à diminuição do volume torácico e à diminuição da retracção pulmonar.
Pressão transpulmonar É dada pela diferença entre pressão alveolar e a pressão intrapleural. É proporcional ao grau de expansão do pulmão. No entanto, a maneira como a pressão transpulmonar expende os pulmões depende da sua elasticidade. Esta elasticidade é designada de compliância e traduz a

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
218 | P á g i n a
magnitude de mudança de volume no pulmão produzida por uma mudança na pressão transpulmonar. Deste modo quanto maior a compliância do pulmão, mais fácil é a expansão deste a qualquer pressão transpulmonar. Pressão alveolar A pressão alveolar refere-se à pressão existente no interior dos alvéolos pulmonares. Quando a glote está aberta, e não ocorre fluxo de ar para dentro ou para fora dos pulmões, as pressões em todas as partes da árvore respiratória, ao longo dos alvéolos, são exactamente iguais à pressão atmosférica, considerada como 0 centímetro de água. Para provocar a entrada de ar durante a inspiração, a pressão nos alvéolos deve cair para um valor ligeiramente inferior à pressão atmosférica. Essa pressão negativa muito pequena é, entretanto, suficiente para determinar a entrada de cerca de 0,5 L de ar nos pulmões nos 2 segundos necessários para a inspiração. Durante a expiração, ocorrem eventos opostos: a pressão alveolar eleva-se para valores superiores à pressão atmosférica, forçando a saída do 0,5 L de ar inspirado dos pulmões durante os 2 a 3 segundos da expiração.
Variações nas Pressões
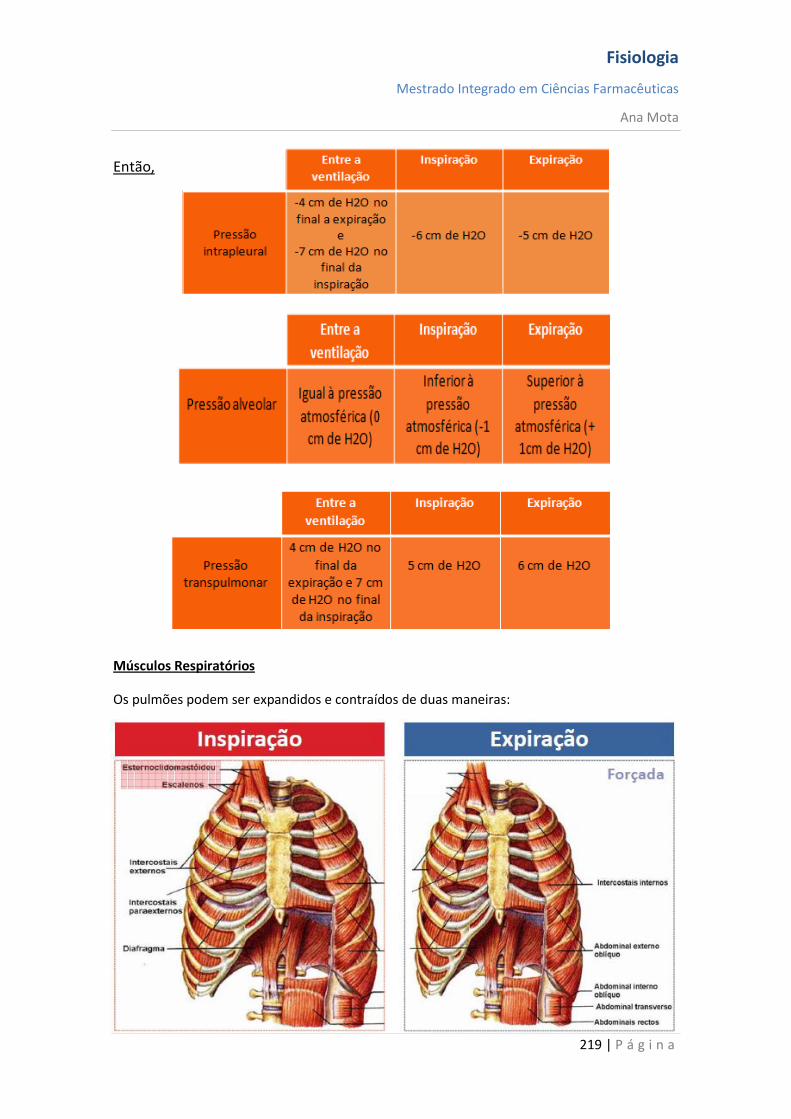
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
219 | P á g i n a
Então,
Músculos Respiratórios
Os pulmões podem ser expandidos e contraídos de duas maneiras:

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
220 | P á g i n a
(1) pelo movimento do diagrama para baixo e para cima, alongando ou encurtando a cavidade
torácica;
(2) pela elevação e depressão das costelas, aumentando e diminuindo o diâmetro antero-posterior da cavidade torácica.
Em repouso, a respiração normal ocorre quase totalmente pelo primeiro desses dois mecanismos, isto é, pela movimentação do diafragma. Durante a inspiração, a contracção do diafragma traciona para baixo a superfície inferior dos pulmões. Em seguida durante a expiração, o diafragma simplesmente relaxa, e a retracção elástica dos pulmões, da parede torácica e das estruturas abdominais comprime os pulmões. Contudo, durante a respiração intensa, as forças elásticas não são suficientes para produzirem a respiração rápida necessária, e a força adicional é obtida, principalmente, pela contracção dos músculos abdominais, que empurra o conteúdo abdominal cima, contra a base do diafragma.
Músculos Inspiração: Esternocleidomastoideo; Escalenos; Intercostais externos.
Músculos Expiração:
Intercostais internos; Abdominal oblíquo externo; Abdominal oblíquo interno; Abdominal transverso; Abdominais rectos.
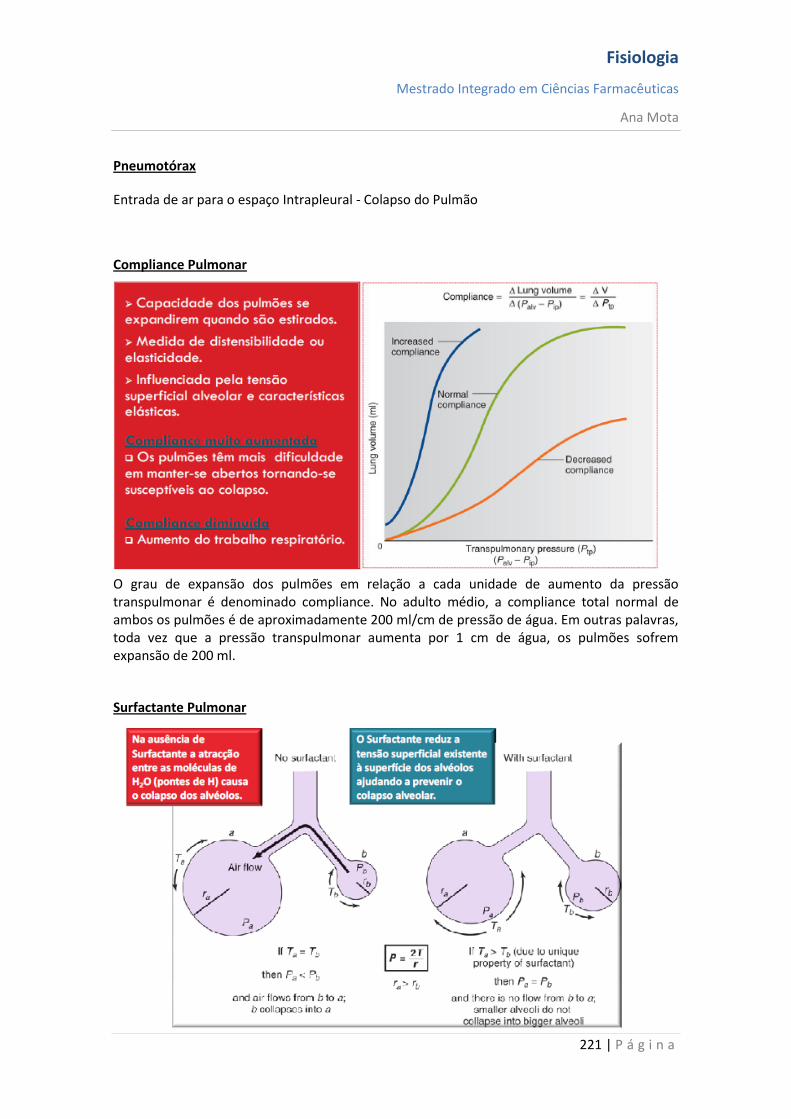
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
221 | P á g i n a
Pneumotórax
Entrada de ar para o espaço Intrapleural - Colapso do Pulmão
Compliance Pulmonar
O grau de expansão dos pulmões em relação a cada unidade de aumento da pressão transpulmonar é denominado compliance. No adulto médio, a compliance total normal de ambos os pulmões é de aproximadamente 200 ml/cm de pressão de água. Em outras palavras, toda vez que a pressão transpulmonar aumenta por 1 cm de água, os pulmões sofrem expansão de 200 ml.
Surfactante Pulmonar
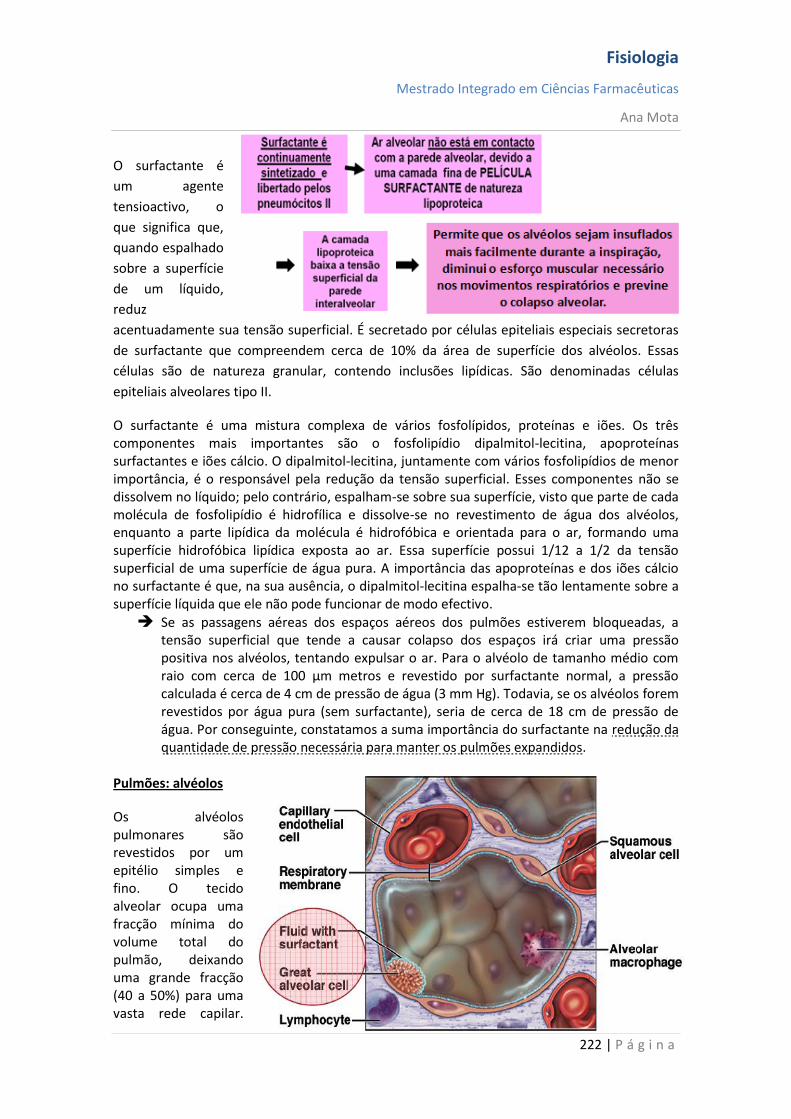
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
222 | P á g i n a
O surfactante é
um agente
tensioactivo, o
que significa que,
quando espalhado
sobre a superfície
de um líquido,
reduz
acentuadamente sua tensão superficial. É secretado por células epiteliais especiais secretoras
de surfactante que compreendem cerca de 10% da área de superfície dos alvéolos. Essas
células são de natureza granular, contendo inclusões lipídicas. São denominadas células
epiteliais alveolares tipo II.
O surfactante é uma mistura complexa de vários fosfolípidos, proteínas e iões. Os três componentes mais importantes são o fosfolipídio dipalmitol-lecitina, apoproteínas surfactantes e iões cálcio. O dipalmitol-lecitina, juntamente com vários fosfolipídios de menor importância, é o responsável pela redução da tensão superficial. Esses componentes não se dissolvem no líquido; pelo contrário, espalham-se sobre sua superfície, visto que parte de cada molécula de fosfolipídio é hidrofílica e dissolve-se no revestimento de água dos alvéolos, enquanto a parte lipídica da molécula é hidrofóbica e orientada para o ar, formando uma superfície hidrofóbica lipídica exposta ao ar. Essa superfície possui 1/12 a 1/2 da tensão superficial de uma superfície de água pura. A importância das apoproteínas e dos iões cálcio no surfactante é que, na sua ausência, o dipalmitol-lecitina espalha-se tão lentamente sobre a superfície líquida que ele não pode funcionar de modo efectivo. Se as passagens aéreas dos espaços aéreos dos pulmões estiverem bloqueadas, a
tensão superficial que tende a causar colapso dos espaços irá criar uma pressão positiva nos alvéolos, tentando expulsar o ar. Para o alvéolo de tamanho médio com raio com cerca de 100 μm metros e revestido por surfactante normal, a pressão calculada é cerca de 4 cm de pressão de água (3 mm Hg). Todavia, se os alvéolos forem revestidos por água pura (sem surfactante), seria de cerca de 18 cm de pressão de água. Por conseguinte, constatamos a suma importância do surfactante na redução da quantidade de pressão necessária para manter os pulmões expandidos.
Pulmões: alvéolos
Os alvéolos pulmonares são revestidos por um epitélio simples e fino. O tecido alveolar ocupa uma fracção mínima do volume total do pulmão, deixando uma grande fracção (40 a 50%) para uma vasta rede capilar.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
223 | P á g i n a
Como resultado, a distância média entre o gás alveolar e a hemoglobina nos eritrócitos é de apenas 1,5 μm, o que torna bastante eficientes as trocas. Independentemente deste reduzido volume, a área de superfície alveolar interna é aproximadamente 1 m2/Kg de peso corporal. A unidade funcional do pulmão, para efeitos de trocas, recebe a designação de Unidade Respiratória Terminal. É composta por um bronquíolo terminal e respectivos ductos alveolares (bronquíolos respiratórios) e alvéolos. Existirão cerca de 60000 destas unidades, cada qual com 250 ductos alveolares e 5000 alvéolos anatómicos.
Resistência ao Ar
Tónus do Músculo Brônquico
As fibras musculares brônquicas são predominantemente inervadas pelo parassimpático, que tem acção constritora ligeira a moderada. Podem ser activadas por reflexos com origem pulmonar (reflexos da tosse e do espirro), quando há irritação das vias respiratórias. Estes reflexos poderão estar hiperactivos nos doentes asmáticos. O simpático exerce um fraco controlo directo, visto que há poucas terminações nervosas deste sistema a nível brônquico. Contudo, há receptores β2-adrenérgicos que respondem a catecolaminas circulantes. O simpático enerva directamente as glândulas submucosas, os gânglios parassimpáticos e o músculo liso vascular. Está descrita, também, uma inervação não-colinérgica e não-adrenérgica com acção broncodilatadora mediada pelo VIP (vasoactive intestinal peptide).
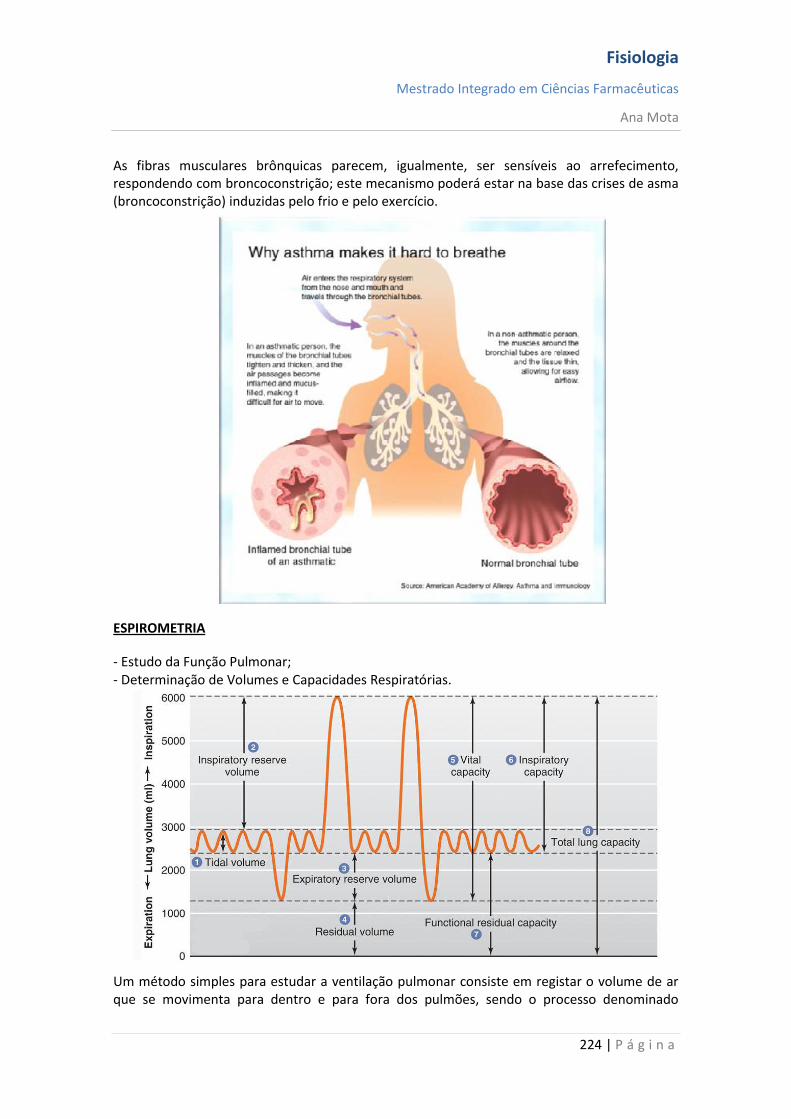
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
224 | P á g i n a
As fibras musculares brônquicas parecem, igualmente, ser sensíveis ao arrefecimento, respondendo com broncoconstrição; este mecanismo poderá estar na base das crises de asma (broncoconstrição) induzidas pelo frio e pelo exercício.
ESPIROMETRIA
- Estudo da Função Pulmonar; - Determinação de Volumes e Capacidades Respiratórias.
Um método simples para estudar a ventilação pulmonar consiste em registar o volume de ar que se movimenta para dentro e para fora dos pulmões, sendo o processo denominado

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
225 | P á g i n a
espirometria. A figura ilustra um espirograma, mostrando as alterações do volume pulmonar em diferentes condições de respiração. Para facilitar a descrição dos eventos da ventilação pulmonar, o ar nos pulmões foi subdividido, em diferentes pontos desse diagrama, em quatro volumes e quatro capacidades diferentes. Numa expiração forçada, os pulmões não expelem todo o ar até ao seu colapso, porque a rigidez da parede torácica limita o esvaziamento completo. Assim, fica sempre algum volume de ar nos pulmões: é o volume residual. Numa expiração normal, fica no pulmão um volume de ar, que é a capacidade residual funcional (CRF), determinada, fundamentalmente, pelas relações mecânicas passivas entre a parede torácica e os pulmões (ponto de equilíbrio). Esta capacidade funcional é um sistema tamponador contra grandes variações da PAo2 em cada ventilação. Se o volume em questão fosse muito pequeno, a PAo2 tornar-se-ia muito próxima da Pvo2, durante a expiração, e muito próxima da Po2 no ar inspirado, durante a inspiração. Como a CRF (2,4 L) corresponde a 40% da capacidade pulmonar total (CPT; 6L), sendo muito superior ao volume de cada ciclo respiratório não há grandes variações nas pressões alveolares de O2 (≈4mmHg) e de CO2 (≈3mmHg). De facto, em termos práticos, pode considerar-se que não há variações ao longo do ciclo respiratório, numa pessoa normal, o mesmo acontecendo com as respectivas pressões parciais no sangue arterial. Além dos volumes referidos, considera-se ainda um volume de reserva inspiratória (volume adicional mobilizado por inspiração forçada) e o volume de reserva expiratória (volume adicional expulso numa expiração forçada). Estes volumes, adicionados ao volume corrente, constituem a capacidade vital. Há várias capacidades pulmonares obtidas por somatório de volumes.
Volume Tidal (VT) – volume de gás inspirado ou expirado durante um ciclo de ventilação normal.
Capacidade de inspiração (CI) – quantidade máxima de gás que pode ser inalada no final de uma expiração normal.
Ventilação Alveolar

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
226 | P á g i n a
A ventilação alveolar corresponde ao volume disponível para as trocas gasosas por minuto. No entanto, para a eficácia das trocas gasosas é necessário avaliar tanto o volume alveolar como o fluxo sanguíneo pulmonar, tendo em conta as diferenças de concentrações de O2 e CO2 entre ar inspirado e expirado. Pode considerar-se que a ventilação alveolar é a parte da ventilação total obtida após a exclusão do espaço morto anatómico.
VA = FR (VC - VEM)
O ar inspirado tem que atravessar as vias aéreas superiores, nas quais não há trocas de gases significativas. Logo, há um volume de ar ventilado que não participa nas trocas: é o espaço morto anatómico. Os seus valores normais rondam os 2 mL/Kg do peso corporal ideal do indivíduo (≅150 mL). No final da expiração, as vias aéreas superiores, condutoras, têm uma Po2 = 102 mmHg e Pco2 = 40 mmHg, semelhantes às alveolares. Na inspiração seguinte, este ar passa para os alvéolos em primeiro lugar e só depois entra ar fresco. Assim, a ventilação alveolar é menos eficaz que a ventilação total; perde-se eficácia devido ao espaço morto anatómico. O pulmão ideal teria um acoplamento perfeito entre a ventilação e o fluxo sanguíneo (V/Q) em todas as regiões, mas não é isto que se verifica, mesmo no indivíduo normal. Há um desvio em relação ao ideal, que é normal e fisiológico. Há áreas em que o acoplamento ventilação/perfusão não é perfeito. Logo, os volumes ventilatórios alveolares não entram totalmente em contacto com capilares pulmonares e, portanto, não efectuam trocas gasosas. O somatório deste volume desperdiçado com o espaço morto anatómico constitui o espaço morto fisiológico, e é este que se subtrai à ventilação total para obter a ventilação alveolar. É este espaço morto que tem correlação fisiológica e, a partir deste ponto, a designação espaço morto deve ser tomada como espaço morto fisiológico.
Trocas e Transporte de Gases
- Trocas de gases entre os alvéolos e o sangue; - Transporte de O2 e CO2 no sangue; - Trocas de gases entre o sangue e os tecidos.
Lei de Henry
A pressão de um gás
em solução é
determinada, não
apenas pela sua
concentração, mas
também pelo
coeficiente de
solubilidade. Isto é,
alguns tipos de
moléculas, em
particular dióxido de
carbono, são física ou

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
227 | P á g i n a
quimicamente atraídas por moléculas de água, enquanto outras são repelidas. Naturalmente,
quando as moléculas são atraídas, maior quantidade delas pode dissolver-se sem criar pressão
excessiva dentro da solução. Por outro lado, no caso das moléculas que são repelidas,
desenvolvem-se pressões excessivas para um menor número de moléculas dissolvidas. Essas
relações podem ser expressas pela seguinte fórmula, que constitui a lei de Henry:
Quando a pressão é expressa em atmosferas e a concentração em volume de gás dissolvido
por cada volume de água, os coeficientes de solubilidade para os gases respiratórios
importantes na temperatura corporal são os seguintes:
Oxigénio 0,024
Dióxido de carbono 0,57
Monóxido de carbono 0,018
Azoto 0,012
Hélio 0,008
Com base nesses valores, verifica-se que o dióxido de carbono é mais de 20 vezes mais solúvel
que o oxigénio, enquanto este é moderadamente mais solúvel do que qualquer um dos outros
três gases.
Gradiente de Pressões dos Gases

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
228 | P á g i n a
Composição do Ar
Trocas de Gases: pulmões
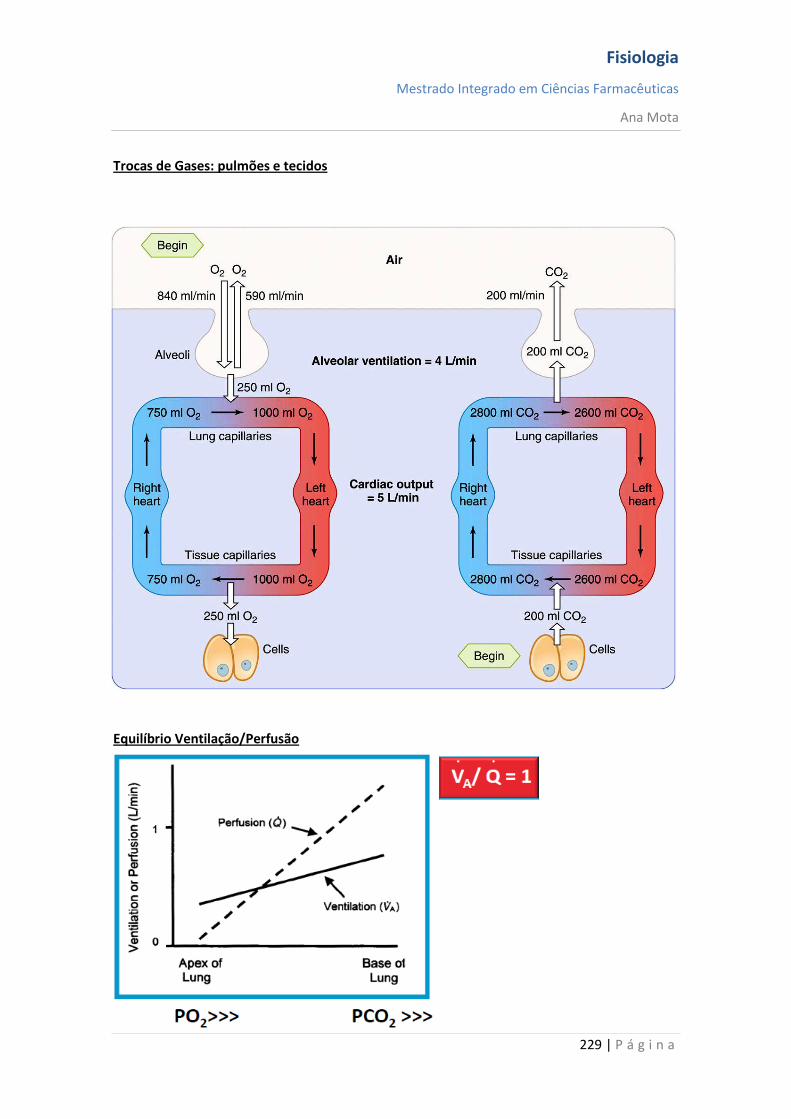
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
229 | P á g i n a
Trocas de Gases: pulmões e tecidos
Equilíbrio Ventilação/Perfusão

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
230 | P á g i n a
Esta relação mede a funcionalidade do sistema respiratório;
Deve haver um equilíbrio entre a capacidade de ventilação e a perfusão sanguínea
alveolar, para que as trocas gasosas ocorram de forma adequada.
Quando VA/Q = 1 _(normal)
Ventilação alveolar e fluxo sanguíneo são normais;
As trocas de O2 e CO2 aproximam-se do ideal (=1).
Desequilíbrios V/Q: shunts
Quando VA/Q = 0 - Shunt
O alvéolo não recebe nenhuma ventilação;
O fluxo sanguíneo permanece normal.
Quando VA/Q = infinito – espaço morto
Não há fluxo sanguíneo capilar nem para levar o O2 para as células nem para trazer o CO2 para os pulmões;
A concentração de O2 nos alvéolos torna-se idêntica à do ar inspirado, uma vez que o ar inspirado não perde nenhum O2 nem recebe nenhum CO2.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
231 | P á g i n a
Nota:
Shunt = sangue desviado não-oxigenado.
Ajustes na Perfusão
A perfusão corresponde ao fluxo sanguíneo pulmonar. Perfusão = FR × volume de ejecção
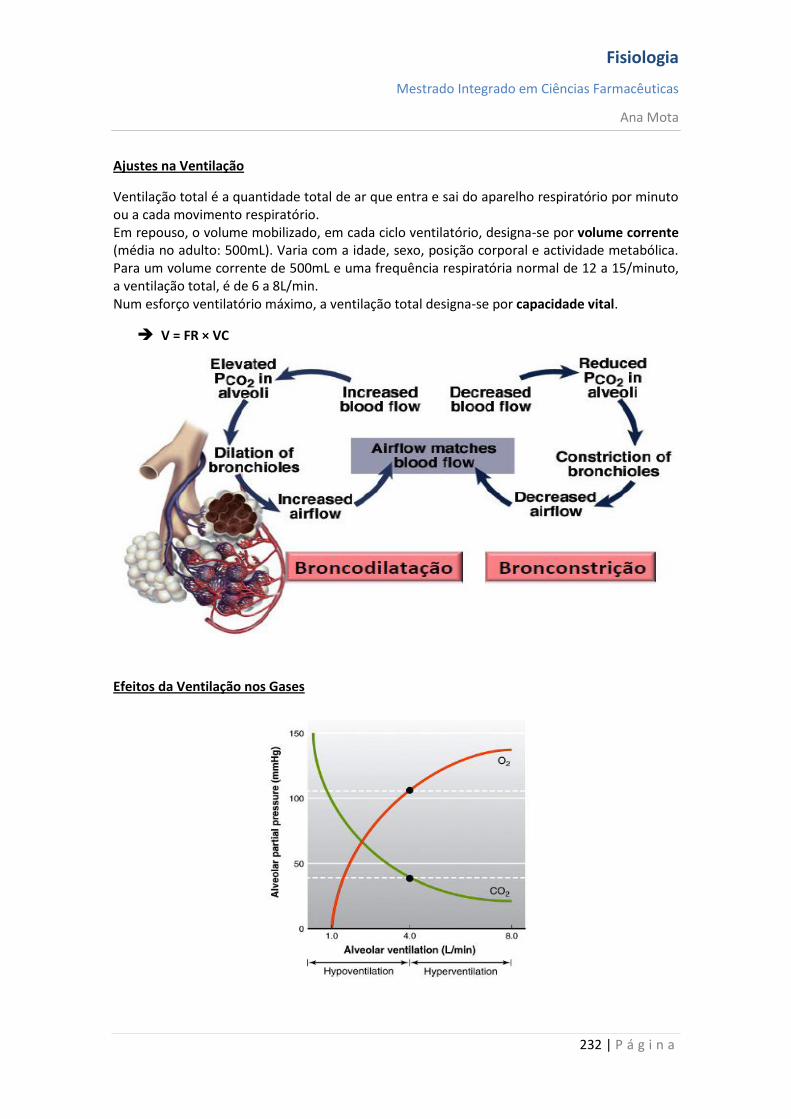
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
232 | P á g i n a
Ajustes na Ventilação
Ventilação total é a quantidade total de ar que entra e sai do aparelho respiratório por minuto ou a cada movimento respiratório. Em repouso, o volume mobilizado, em cada ciclo ventilatório, designa-se por volume corrente (média no adulto: 500mL). Varia com a idade, sexo, posição corporal e actividade metabólica. Para um volume corrente de 500mL e uma frequência respiratória normal de 12 a 15/minuto, a ventilação total, é de 6 a 8L/min. Num esforço ventilatório máximo, a ventilação total designa-se por capacidade vital.
V = FR × VC
Efeitos da Ventilação nos Gases
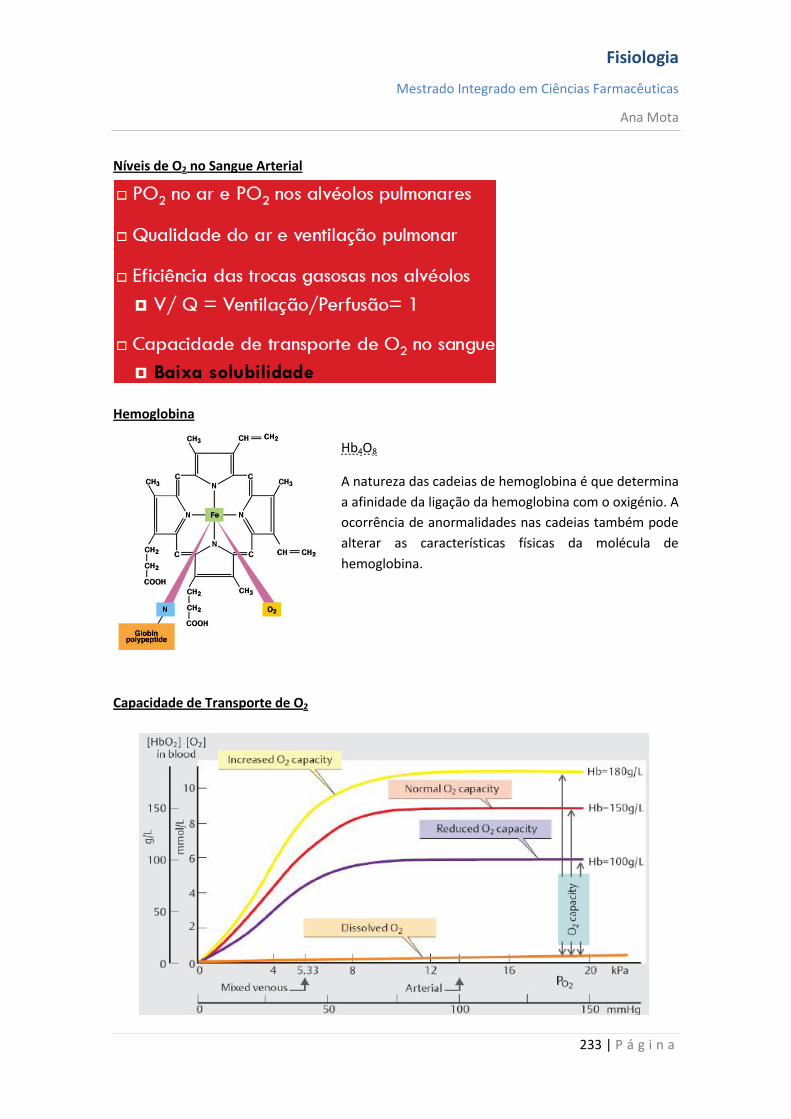
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
233 | P á g i n a
Níveis de O2 no Sangue Arterial
Hemoglobina
Hb4O8
A natureza das cadeias de hemoglobina é que determina
a afinidade da ligação da hemoglobina com o oxigénio. A
ocorrência de anormalidades nas cadeias também pode
alterar as características físicas da molécula de
hemoglobina.
Capacidade de Transporte de O2

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
234 | P á g i n a
O O2 é muito pouco solúvel na água. É por intermédio da hemoglobina (proteína que se liga de forma rápida e reversível ao O2, no interior dos eritrócitos) que é possível o transporte de quantidades significativas de O2 no sangue. Apenas 25% de O2 ligado à hemoglobina (Hb) sofre trocas em cada passagem circulatória pelos tecidos o que permite manter uma reserva que poderá ser usada numa emergência, quer por paragem circulatória, quer por aumento da actividade tecidual. Adicionalmente, mantém um gradiente de pressões que força a difusão. O débito cardíaco, mesmo no exercício máximo, não consegue ultrapassar um valor triplo do normal. Porém, o consumo de O2 pode aumentar até seis vezes o seu valor basal. Isto acontece pela mobilização da referida reserva de O2 associado à Hb. A hemoglobina consiste em quatro núcleos heme (anéis porfirínicos com ferro ferroso-
Fe2+) que se ligam ao O2. Cada grupo heme combina-se com uma cadeia de globina (porção proteica). Assim, a hemoglobina é um tetrâmero. Esta estrutura tetramérica, bem como a sua conformação espacial, conferem à Hb as suas propriedades fisiológicas. A hemoglobina (Hb) liga-se rápida (milissegundos) e reversivelmente ao O2, designando-se esta forma como oxihemoglobina (HbO2).
Efeito da Hemoglobina
A característica mais importante da molécula de hemoglobina reside na sua capacidade de se combinar frouxamente e de modo reversível com o oxigénio. A principal função da hemoglobina no organismo depende da sua capacidade de combinar-se com o oxigénio nos pulmões e, a seguir, liberá-lo rapidamente nos capilares teciduais, onde a tensão gasosa do oxigénio é muito menor do que nos pulmões. O oxigénio não se combina com as duas ligações positivas do ferro da molécula de hemoglobina. Na verdade, liga-se fracamente a uma das seis ligações de "coordenação" do átomo de ferro. Trata-se de uma ligação extremamente fraca, de modo que a combinação é facilmente reversível. Além disso, o oxigénio não se transforma em oxigénio iónico, mas é transportado sob a forma de oxigénio molecular para os tecidos, onde, devido a sua combinação fraca e rapidamente reversível, é liberado nos líquidos teciduais sob a forma de oxigénio molecular dissolvido, e não sob a forma iónica.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
235 | P á g i n a
Curva de Dissociação da OxiHb
Aumento progressivo da percentagem de hemoglobina que se liga ao oxigénio à medida que a Po2 aumenta.
Esse processo é denominado percentagem de saturação da hemoglobina. Como o sangue nas artérias possui geralmente Po2 de cerca de 95 mm Hg, podemos verificar, com base na curva de dissociação, que a saturação habitual do oxigénio do sangue arterial situa-se em torno de 97%. Por outro lado, no sangue venoso normal que retorna dos tecidos, a Po2 é de cerca de 40 mm Hg, e a saturação da hemoglobina é de cerca de 75%. A forma da curva é sigmóide, o que reflecte a interacção dos quatro grupos heme- cinética de ligação cooperativa. Exemplificando, quando três grupos heme já se ligaram ao O2, a capacidade de ligação do restante está aumentada. Esta propriedade permite a ligação a uma quantidade máxima de O2 no pulmão e entrega máxima nos tecidos. O significado funcional da parte plana superior da curva é o de uma margem de segurança nas quedas de PaO2. A saturação arterial de oxigénio não se altera muito até que a pressão parcial de O2 desça abaixo dos 70 mmHg. Todavia, convém ter presente que em qualquer ponto da relação há permuta de gases. Designa-se por P50 a pressão parcial de O2 que corresponde a metade da saturação da hemoglobina. O seu valor é variável, mas sempre superior à PO2 tecidual.
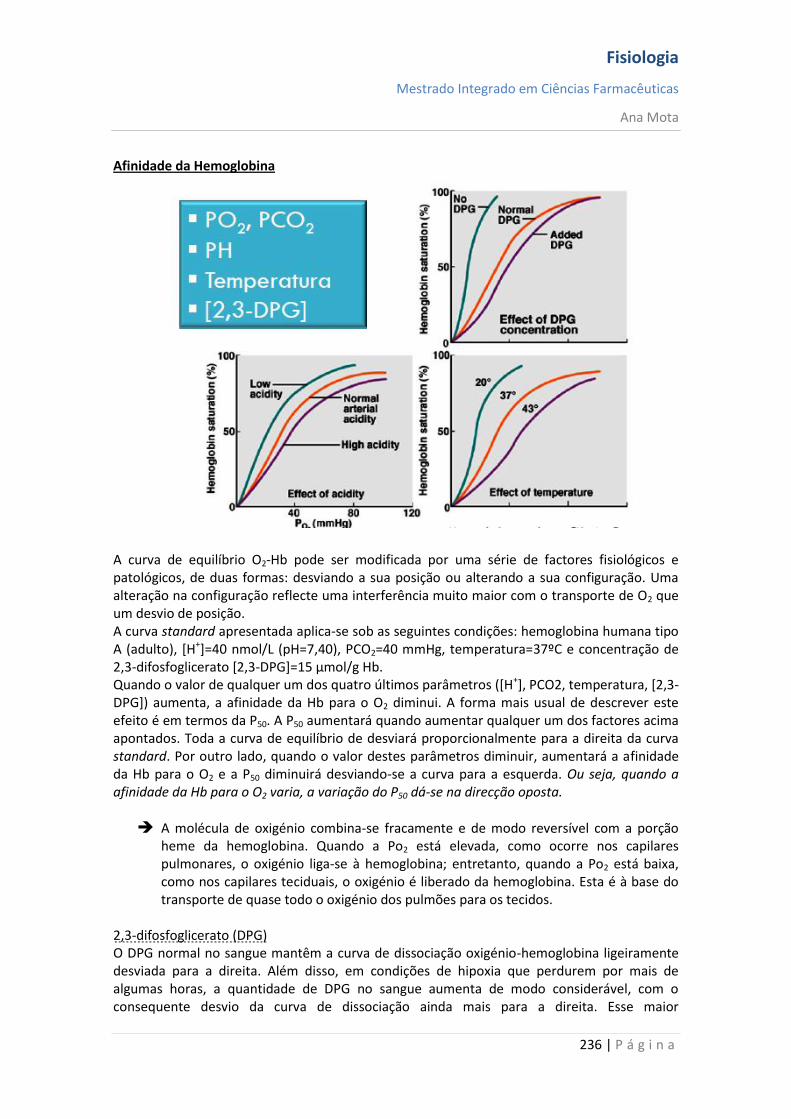
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
236 | P á g i n a
Afinidade da Hemoglobina
A curva de equilíbrio O2-Hb pode ser modificada por uma série de factores fisiológicos e patológicos, de duas formas: desviando a sua posição ou alterando a sua configuração. Uma alteração na configuração reflecte uma interferência muito maior com o transporte de O2 que um desvio de posição. A curva standard apresentada aplica-se sob as seguintes condições: hemoglobina humana tipo A (adulto), [H+]=40 nmol/L (pH=7,40), PCO2=40 mmHg, temperatura=37ºC e concentração de 2,3-difosfoglicerato [2,3-DPG]=15 μmol/g Hb. Quando o valor de qualquer um dos quatro últimos parâmetros ([H+], PCO2, temperatura, [2,3-DPG]) aumenta, a afinidade da Hb para o O2 diminui. A forma mais usual de descrever este efeito é em termos da P50. A P50 aumentará quando aumentar qualquer um dos factores acima apontados. Toda a curva de equilíbrio de desviará proporcionalmente para a direita da curva standard. Por outro lado, quando o valor destes parâmetros diminuir, aumentará a afinidade da Hb para o O2 e a P50 diminuirá desviando-se a curva para a esquerda. Ou seja, quando a afinidade da Hb para o O2 varia, a variação do P50 dá-se na direcção oposta. A molécula de oxigénio combina-se fracamente e de modo reversível com a porção
heme da hemoglobina. Quando a Po2 está elevada, como ocorre nos capilares pulmonares, o oxigénio liga-se à hemoglobina; entretanto, quando a Po2 está baixa, como nos capilares teciduais, o oxigénio é liberado da hemoglobina. Esta é à base do transporte de quase todo o oxigénio dos pulmões para os tecidos.
2,3-difosfoglicerato (DPG) O DPG normal no sangue mantêm a curva de dissociação oxigénio-hemoglobina ligeiramente desviada para a direita. Além disso, em condições de hipoxia que perdurem por mais de algumas horas, a quantidade de DPG no sangue aumenta de modo considerável, com o consequente desvio da curva de dissociação ainda mais para a direita. Esse maior

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
237 | P á g i n a
deslocamento determina a liberação de oxigénio para os tecidos com pressão de oxigénio 10 mm Hg maior do que a que ocorreria sem esse aumento do DPG. Por conseguinte, afirmava-se no passado que esse poderia constituir um mecanismo importante de adaptação à hipoxia. Todavia, a presença de DPG em excesso também dificulta a combinação da hemoglobina com o oxigénio nos pulmões quando a Po2 alveolar está reduzida, causando quase sempre tanto prejuízo quanto benefício. Por isso, é questionável o fato de o desvio da curva de dissociação pelo DPG ser tão benéfico na hipoxia quanto se acreditava. Um desvio da curva para a direita (diminuição da afinidade para o O2) indica que a
quantidade de O2 que pode ser captada à passagem pelos capilares pulmonares está diminuída, a qualquer PaO2. Todavia, como a curva é quase plana para PO2 acima dos 70mmHg, a concentração arterial de oxigénio não se modifica muito por intervenções que desviem a curva para a direita. Porém, um desvio desta natureza indica que a quantidade de oxigénio que pode ser libertada (dissociada da Hb) quando este passa nos capilares sistémicos está aumentada. Este efeito é benéfico na melhoria da entrega de O2 aos tecidos para uma dada PaO2. O aumento da PCO2 e [H+] nos capilares sistémicos reflecte-se por um pequeno desvio para a direita (P50 aumenta cerca de 3mmHg).
Um desvio para a esquerda (que traduz aumento da afinidade para o O2) indica que a ligação do O2 pela Hb está aumentada a cada PaO2. Esta modificação favorece a captação de O2 a partir dos capilares pulmonares.
Movimento do O2: pulmões => tecidos
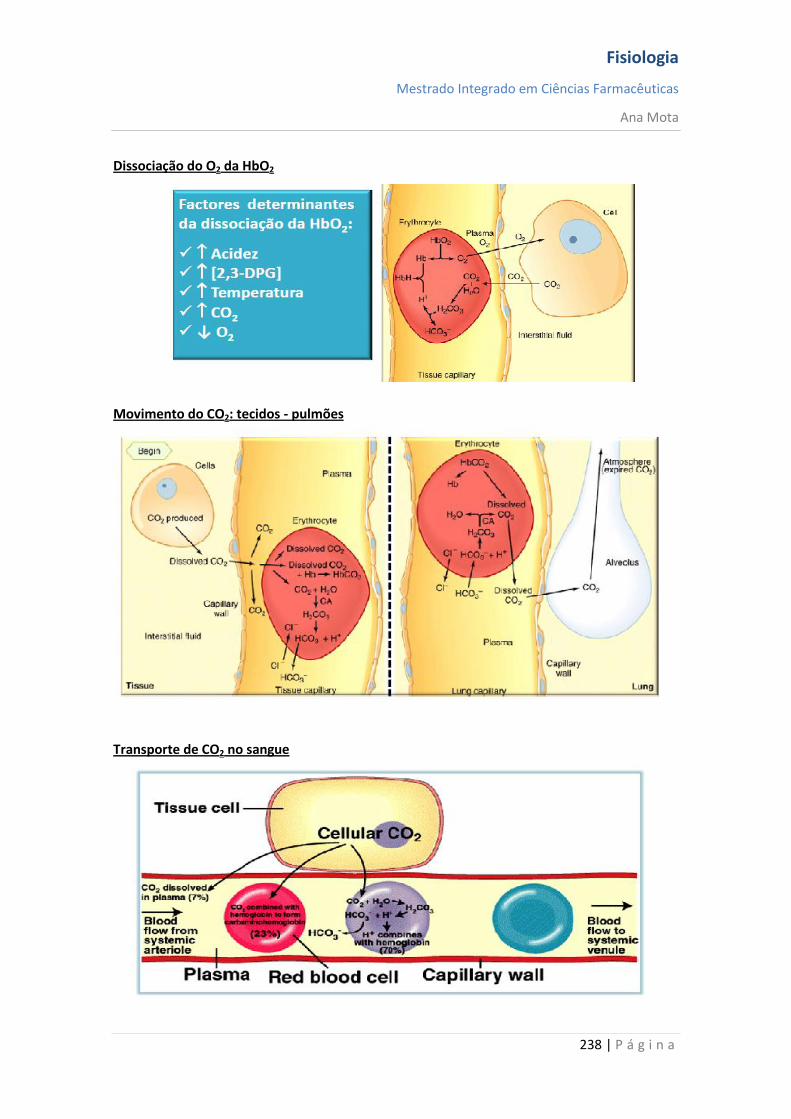
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
238 | P á g i n a
Dissociação do O2 da HbO2
Movimento do CO2: tecidos - pulmões
Transporte de CO2 no sangue

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
239 | P á g i n a
Um dos principais produtos do metabolismo celular é o ácido carbónico (H2CO3) excretado através dos pulmões sob a forma de CO2. A quantidade de CO2 transportada no sangue venoso até aos pulmões ronda os 200 mL/min, no adulto em repouso, mas pode aumentar seis vezes durante o exercício físico. Para uma PaCO2 de 40 mmHg, a concentração total de CO2 no sangue arterial é de cerca de 480 mL/L; no sangue venoso, esta concentração é de cerca de 520 ml/L. A grande quantidade de CO2 no sangue e outros fluidos corporais é necessária para a manutenção do pH (sistema tamponador). À medida que o CO2 é formado nas células, a PCO2 tecidual (que em repouso é, em média, 50 mmHg) aumenta acima dos valores do sangue arterial. Sendo vinte vezes mais solúvel na água que o O2, o CO2 difunde-se para os capilares a favor do gradiente de pressão parcial. O seu transporte no sangue é feito sob três formas: 1. dissolvido (do qual uma pequena porção é convertida em ácido carbónico por hidratação: CO2 + H2O ↔ H2CO3 (7%) 2. ião bicarbonato- HCO3
- (70%) 3. carbamino-CO2 (formas ligada ao componente amino- de proteínas: hemoglobina e outras proteínas plasmáticas) (23%).
Como foi dito, uma pequena porção do CO2 dissolvido é convertida em ácido carbónico segundo a reacção química:
CO2 + H2O ↔ H2CO3 Esta reacção, fisiologicamente lenta (alguns segundos) no plasma, está muito acelerada (microssegundos) no interior do eritrócito pela presença da enzima anídrase carbónica. No plasma e líquidos intersticiais, este equilíbrio químico está geralmente bastante desviado para a esquerda, ou seja, no sentido do CO2 dissolvido. Assim, existe muito pouco ácido carbónico em solução nos líquidos orgânicos. O ácido carbónico que se forma é rapidamente dissociado:
H2CO3 ↔ H+ + HCO3-
Esta reacção é espontânea e muito rápida, visto que os hidrogeniões são rapidamente removidos do local da reacção química, ligando-se à hemoglobina e, em menor extensão, às proteínas plasmáticas. A queda da saturação de O2 da Hb, à medida que passa nos capilares teciduais, melhora a sua capacidade tamponadora, visto que a hemoglobina desoxigenada se combina mais com o H+ que a oxihemoglobina. Os iões bicarbonato difundem-se para fora do eritrócito trocando com o Cl- (chloride shift).

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
240 | P á g i n a
Regulação da Respiração
- Quimioreceptores Centrais e Periféricos; - Centro Respiratório do Bolbo Raquidiano; - Influências dos Núcleos Pontinos e
Córtex Cerebral.
Centro Respiratório
A respiração é controlada automaticamente por um centro nervoso localizado no bulbo.
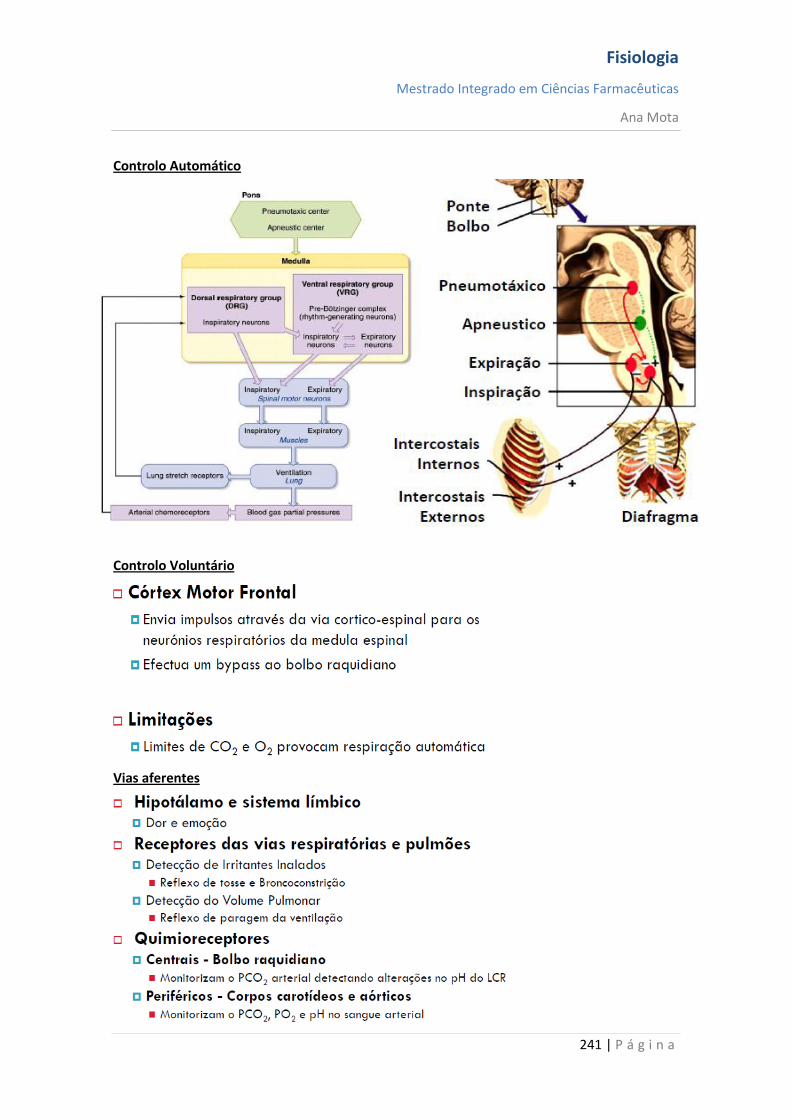
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
241 | P á g i n a
Controlo Automático
Controlo Voluntário
Vias aferentes
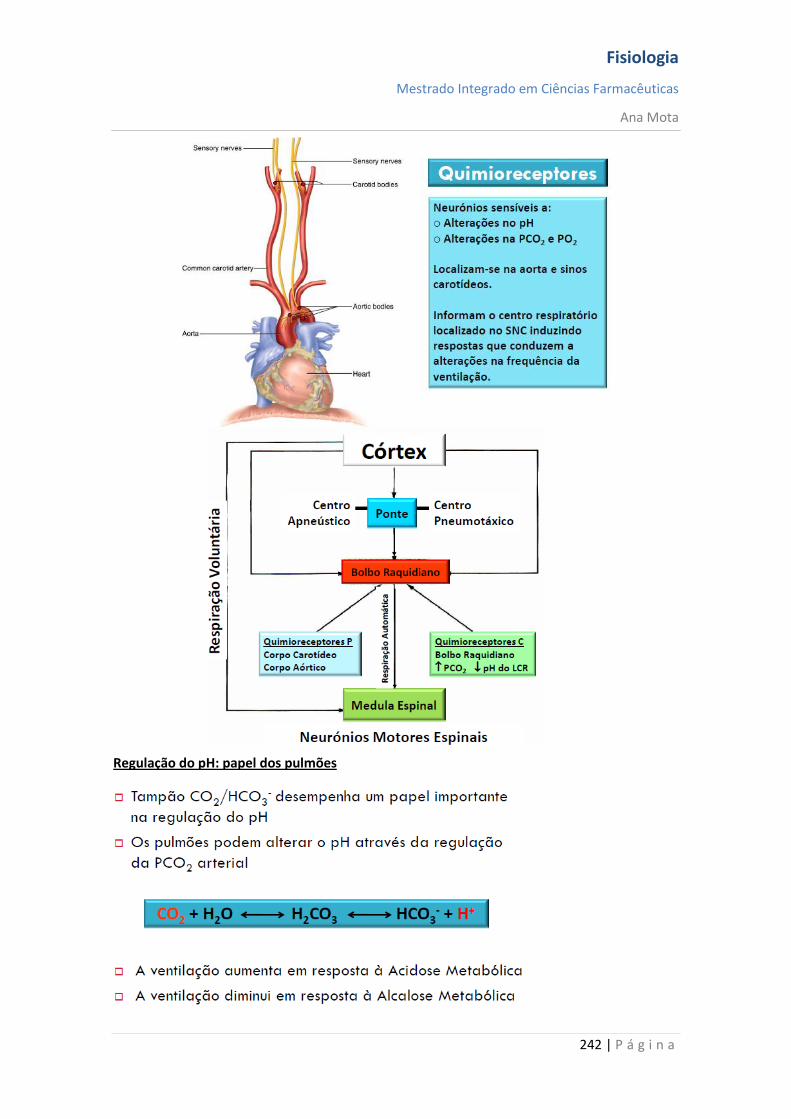
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
242 | P á g i n a
Regulação do pH: papel dos pulmões

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
243 | P á g i n a
Quimioreceptores que respondem a uma diminuição dos níveis de O2 informam o centro de controlo da ventilação para que este aumente a ventilação – Hiperventilação.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
244 | P á g i n a
Efeito do CO2 e pH no LCR
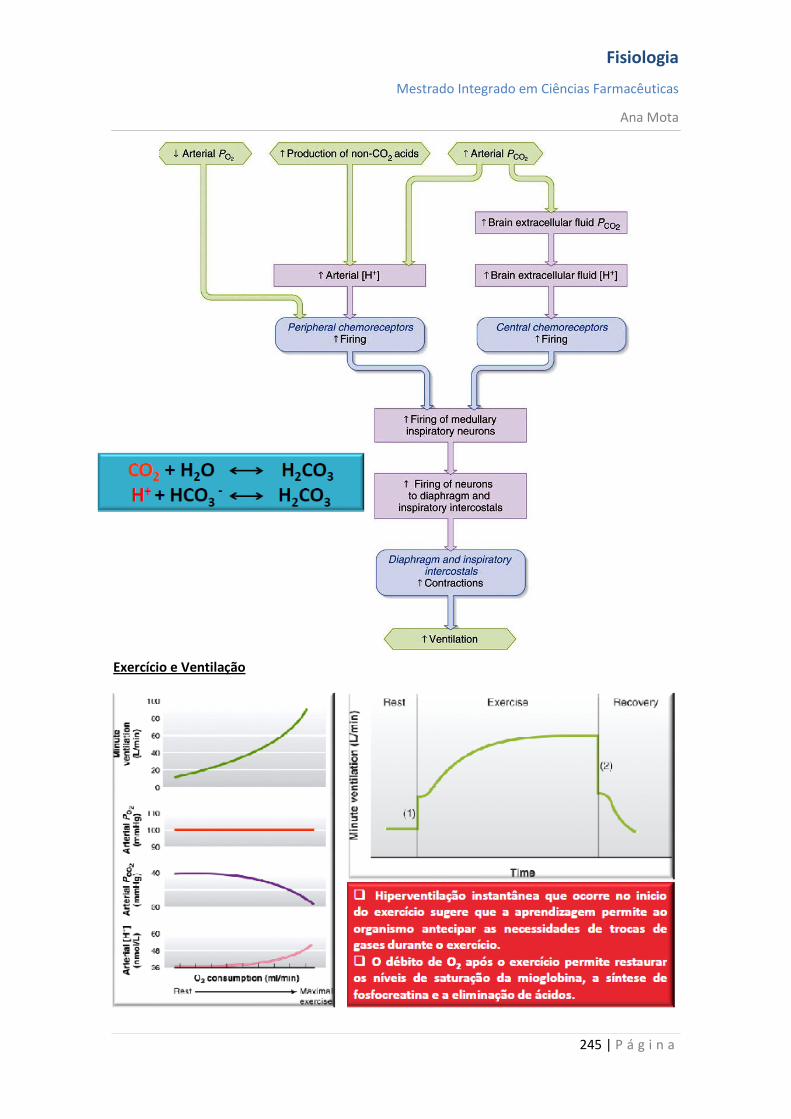
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
245 | P á g i n a
Exercício e Ventilação

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
246 | P á g i n a
Influências na Ventilação
Hipóxia: causas
ACLIMATIZAÇÃO À HIPOXIA DA ALTITUDE
Adaptação da ventilação pulmonar, sistema respiratório e sistema cardiovascular em resposta à diminuição da PO2 no ar inalado que se verifica a elevadas altitudes.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
247 | P á g i n a
Módulo 8 – Sistema urinário e equilíbrio hidro-electrolítico
ORGANIZAÇÃO GERAL
Sistema Urinário: rins, ureteres, bexiga e uretra.
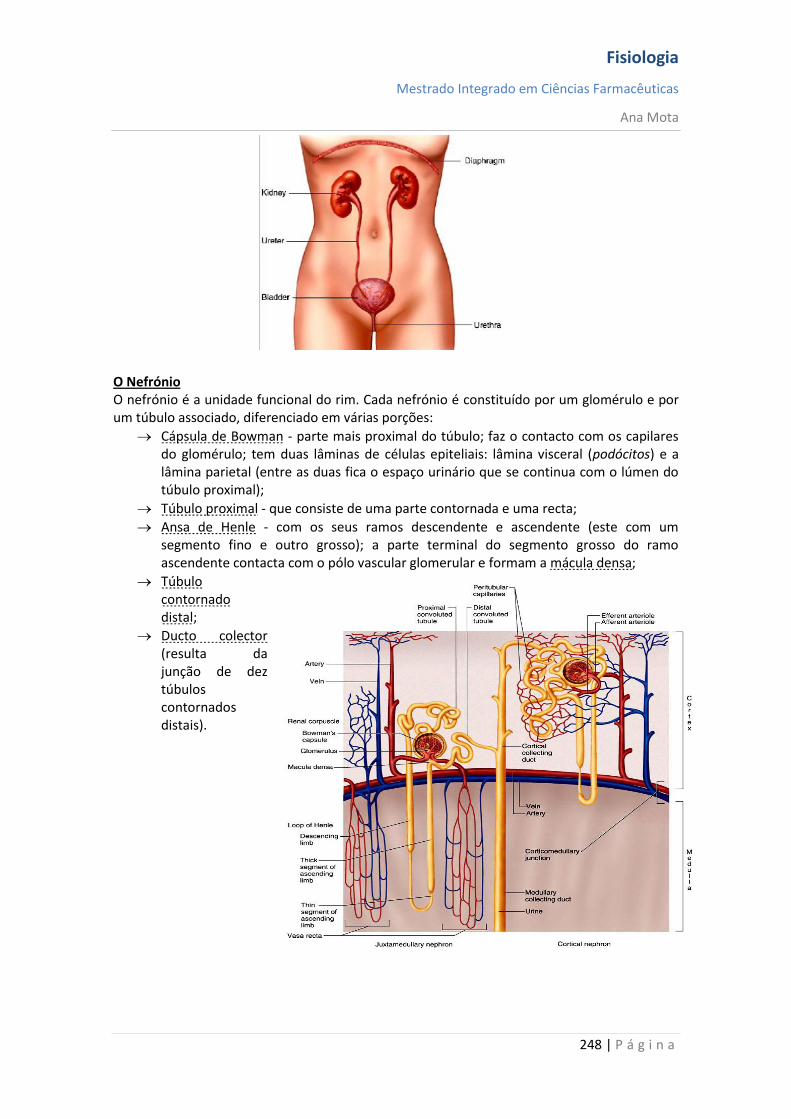
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
248 | P á g i n a
O Nefrónio O nefrónio é a unidade funcional do rim. Cada nefrónio é constituído por um glomérulo e por um túbulo associado, diferenciado em várias porções:
Cápsula de Bowman - parte mais proximal do túbulo; faz o contacto com os capilares do glomérulo; tem duas lâminas de células epiteliais: lâmina visceral (podócitos) e a lâmina parietal (entre as duas fica o espaço urinário que se continua com o lúmen do túbulo proximal);
Túbulo proximal - que consiste de uma parte contornada e uma recta;
Ansa de Henle - com os seus ramos descendente e ascendente (este com um segmento fino e outro grosso); a parte terminal do segmento grosso do ramo ascendente contacta com o pólo vascular glomerular e formam a mácula densa;
Túbulo contornado distal;
Ducto colector (resulta da junção de dez túbulos contornados distais).
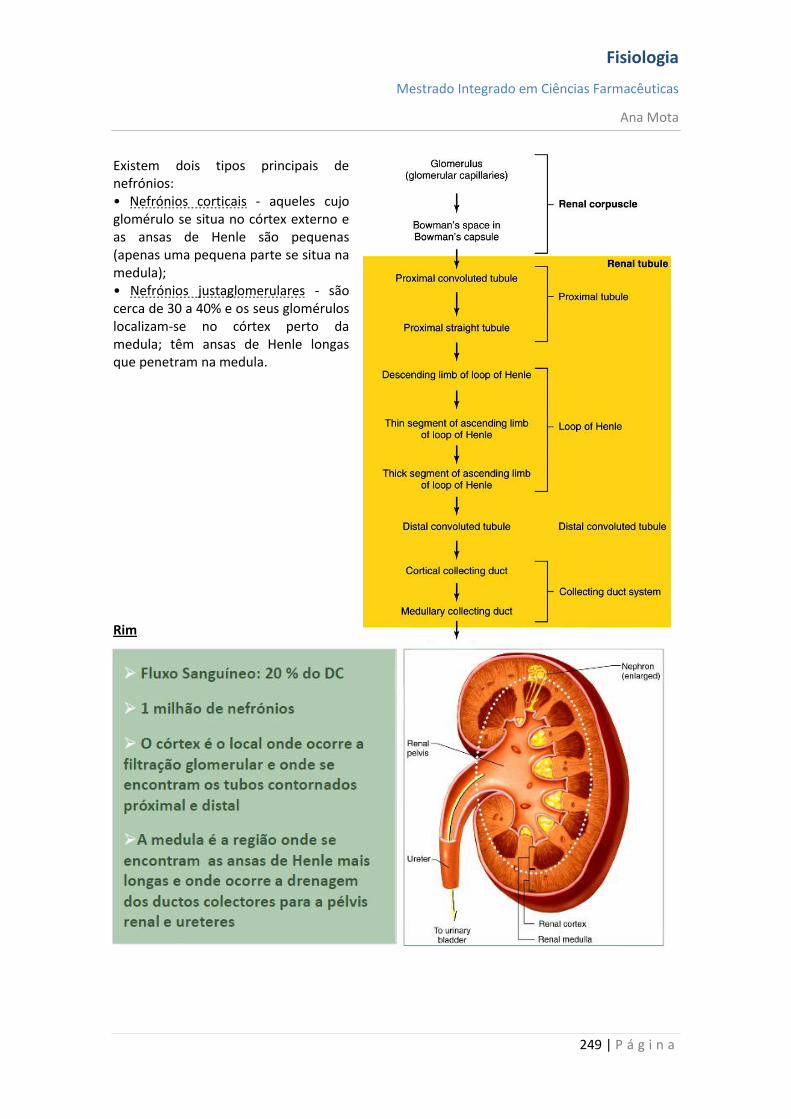
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
249 | P á g i n a
Existem dois tipos principais de nefrónios: • Nefrónios corticais - aqueles cujo glomérulo se situa no córtex externo e as ansas de Henle são pequenas (apenas uma pequena parte se situa na medula); • Nefrónios justaglomerulares - são cerca de 30 a 40% e os seus glomérulos localizam-se no córtex perto da medula; têm ansas de Henle longas que penetram na medula.
Rim

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
250 | P á g i n a
Os rins são órgãos pares localizados retroperitonealmente, ligados funcionalmente ao organismo pelo hilo, local por onde passam os vasos sanguíneos e linfáticos, os nervos e o uréter (que conduz a urina do rim para a bexiga). Em corte coronal podemos distinguir duas regiões principais: o córtex (região externa) e a medula (região interna). A medula divide-se em regiões cónicas (pirâmides renais) que terminam na papila (extremidade do cone) que se projecta para a pelve renal. Os rins recebem, em condições normais, cerca de 20% do débito cardíaco, as artérias renais ramificam-se em artérias interlobares, artérias arqueadas, artérias interlobulares, as quais dão origem às arteríolas aferentes. As arteríolas aferentes ramificam-se em capilares que constituem o glomérulo renal; o sangue continua pelas arteríolas eferentes que dão origem aos capilares peritubulares, os quais drenam para veias de calibre crescente. Como os glomérulos só existem no córtex, os vasos que irrigam a medula originam-se das arteríolas eferentes justaglomerulares e designam-se por vasa recta (descendentes e ascendentes). Os linfáticos também se distribuem apenas ao córtex. A inervação do rim consiste quase exclusivamente de fibras simpáticas eferentes que inervam os vasos e os túbulos renais.
Aparelho Justaglomerular
A Renina é uma enzima sintetizada e libertada pelas células granulares (células musculares lisas modificadas localizadas na média das arteríolas aferentes renais) que integram o complexo justaglomerular. A Renina degrada o Angiotensinogénio (proteína circulante produzida no fígado e rins) em Angiotensina I (decapeptídeo) que, por sua vez, é convertida em Angiotensina II (octapeptídeo activo) pela enzima de conversão da angiotensina (ACE). O nível plasmático da Angiotensina II é determinado pelo nível de Renina plasmática. A secreção de Renina é estimulada por 3 mecanismos principais: a) Aumento da actividade simpática (através de receptores β presentes nas células granulares); b) Redução da pressão arteriolar aferente (mecanismo ainda desconhecido); c) Diminuição da concentração de NaCl na mácula densa: o oposto também se verifica, isto é, a secreção de Renina também é inibida por um aumento do NaCl na mácula densa. A concentração de NaCl neste local é dependente do sódio corporal total e é influenciada pela GFR (taxa de filtração glomerular) e pela reabsorção tubular proximal, as duas variáveis que são modificadas pelo ECV (volume de fluído extracelular). A Angiotensina II estimula directamente a reabsorção de Na+ em vários segmentos tubulares, particularmente no túbulo proximal, onde activa o trocador Na+ / H+. Em concentrações mais elevadas, diminui a GFR por vasoconstrição da arteríola aferente e pela diminuição de Kf (coeficiente de filtração). A produção de angiotensina II plasmática é auto-limitada porque a sua formação inibe a secreção de Renina. A Aldosterona é uma hormona esteróide sintetizada na zona glomerulosa do córtex da glândula suprarrenal. A sua produção é estimulada por: 1. Angiotensina II; 2. Diminuição da concentração plasmática de Na+; 3. Aumento de concentração plasmática de K+; 4. ACTH;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
251 | P á g i n a
5. Diminuição dos níveis plasmáticos do peptídeo natriurético auricular. O factor principal no controlo de secreção de Aldosterona é a Angiotensina II plasmática. A aldosterona estimula a reabsorção de Na+ renal actuando sobre as células principais do ducto colector: atravessa a membrana desta célula e liga-se a um receptor citosólico; o complexo aldosterona – receptor é então translocado para o núcleo onde activa a transcrição do mRNA; as proteínas translocadas – AIP (Aldoterone Induced Proteins) incluem canais de Na+, bomba Na+/K+ e enzimas mitocondriais que aumentam a disponibilidade de ATP para a bomba.
Funções do Rim
SUBSTÂNCIAS TÓXICAS
Compostos nitrogenados resultantes do metabolismo das proteínas, ácidos nucléicos, creatina muscular, hemoglobina e outros resultantes do metabolismo…
Bilirrubina, Fármacos, Pesticidas, Etc....
Produção de Hormonas
Eritropoietina - estimula a produção de eritrócitos pela medula óssea
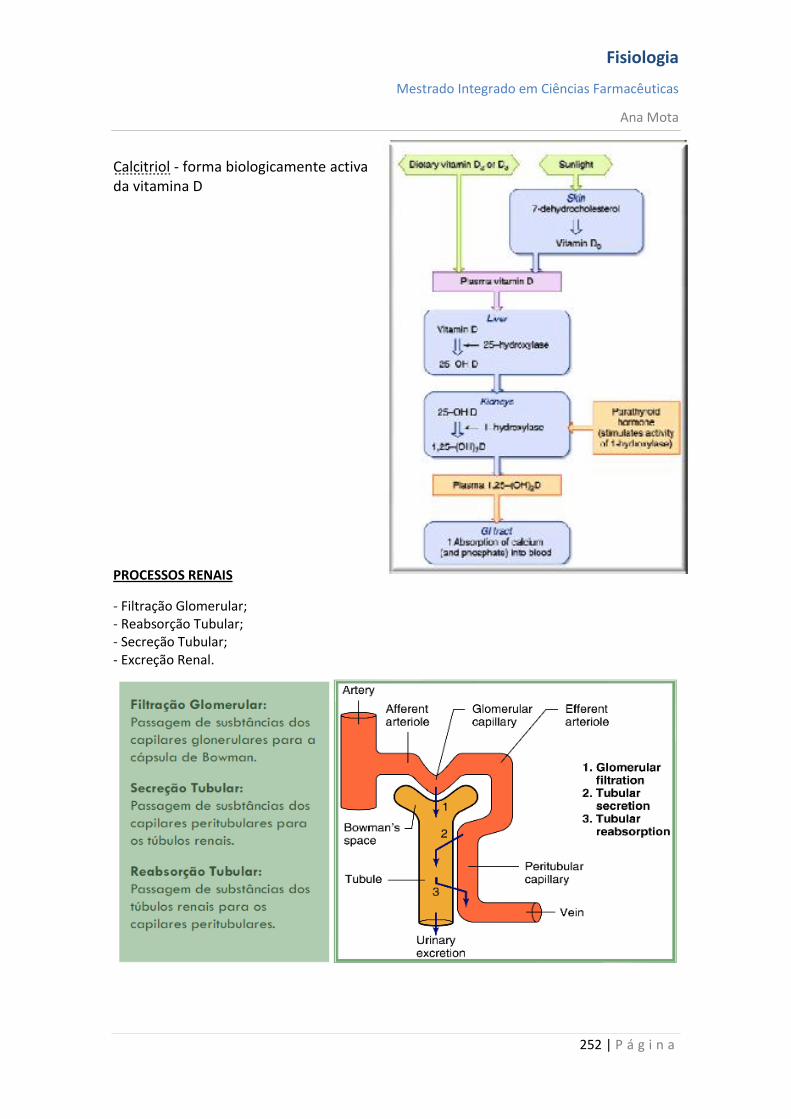
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
252 | P á g i n a
Calcitriol - forma biologicamente activa da vitamina D
PROCESSOS RENAIS
- Filtração Glomerular; - Reabsorção Tubular; - Secreção Tubular; - Excreção Renal.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
253 | P á g i n a
Excreção Renal: reflexo de micção
A micção é um reflexo autonómico controlado voluntariamente; caracteriza-se pela contracção do músculo detrusor (por estimulação dos neurónios parassimpáticos) sincronizada com o relaxamento completo dos dois esfíncteres (por inibição dos neurónios simpáticos e somáticos), o que permite o esvaziamento vesical completo. O controlo voluntário da micção é um comportamento adquirido. Na criança (antes do controlo esfincteriano) o esvaziamento vesical reflecte apenas o componente reflexo da micção desencadeado por estímulos sensitivos a partir dos receptores de estiramento. O esvaziamento voluntário está também perdido nos pacientes com lesão da espinal medula. Neste caso, o que acontece é a falta de sincronização entre a activação reflexa do músculo detrusor e o relaxamento esfincteriano.
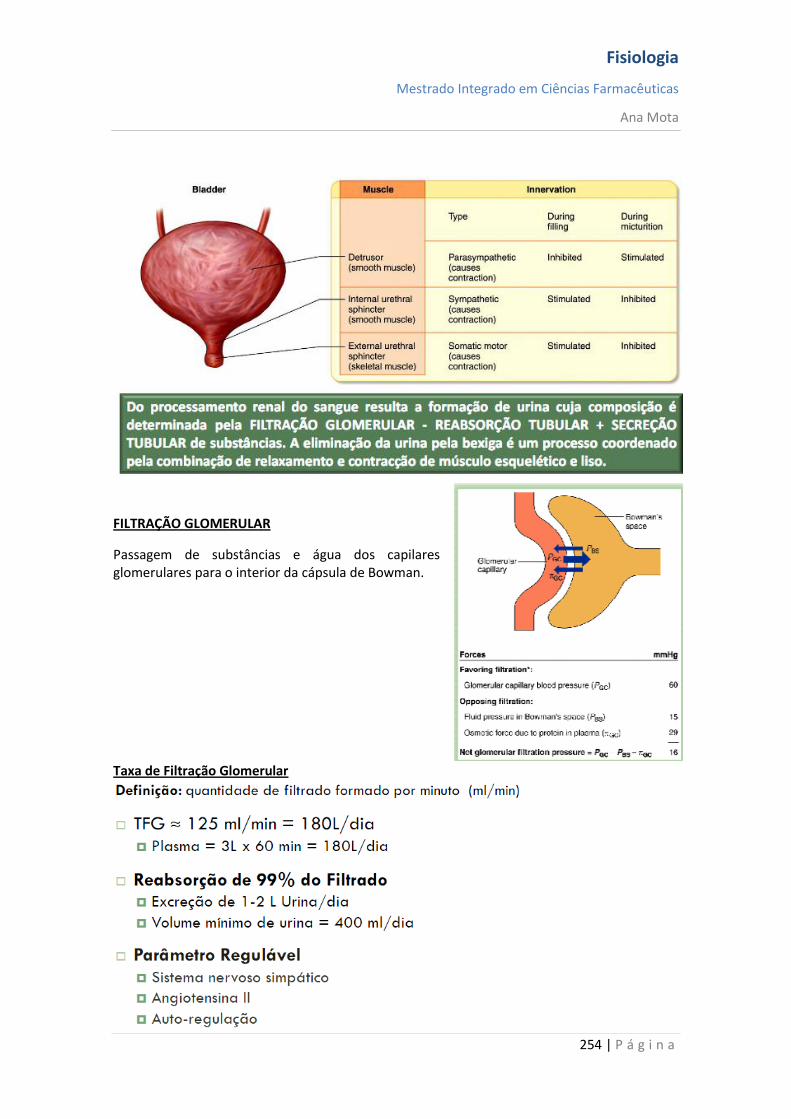
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
254 | P á g i n a
FILTRAÇÃO GLOMERULAR
Passagem de substâncias e água dos capilares glomerulares para o interior da cápsula de Bowman.
Taxa de Filtração Glomerular
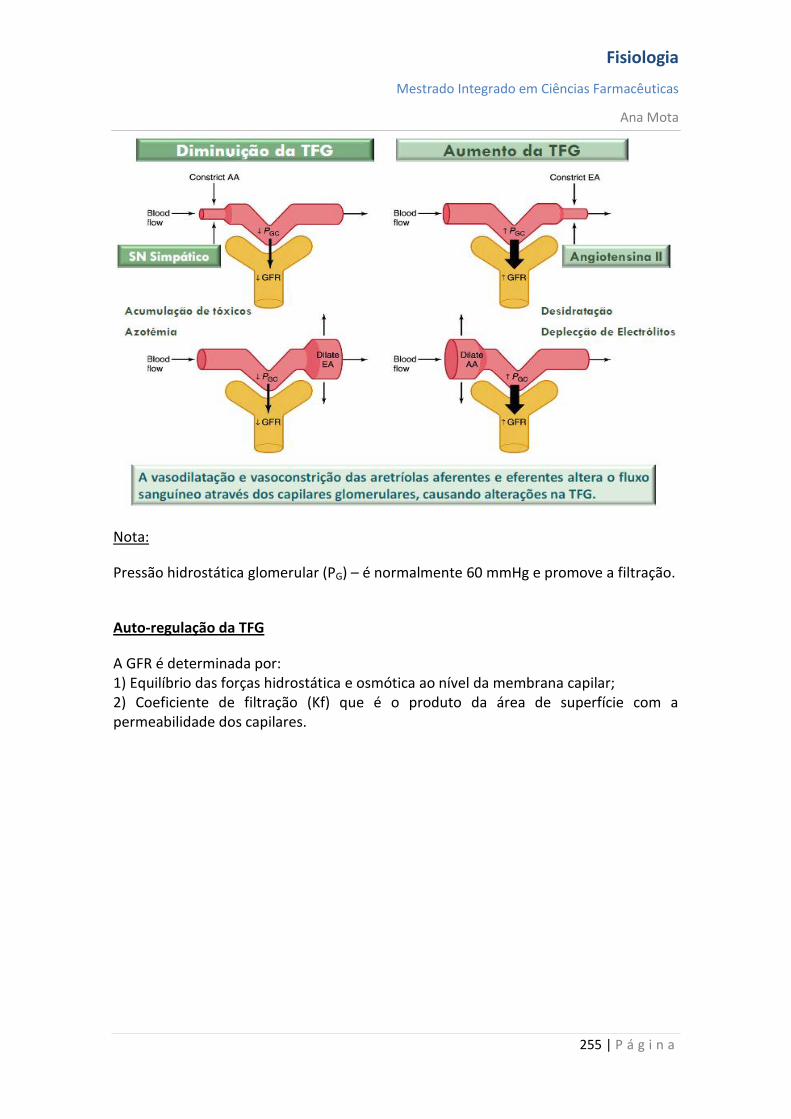
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
255 | P á g i n a
Nota:
Pressão hidrostática glomerular (PG) – é normalmente 60 mmHg e promove a filtração.
Auto-regulação da TFG
A GFR é determinada por: 1) Equilíbrio das forças hidrostática e osmótica ao nível da membrana capilar; 2) Coeficiente de filtração (Kf) que é o produto da área de superfície com a permeabilidade dos capilares.
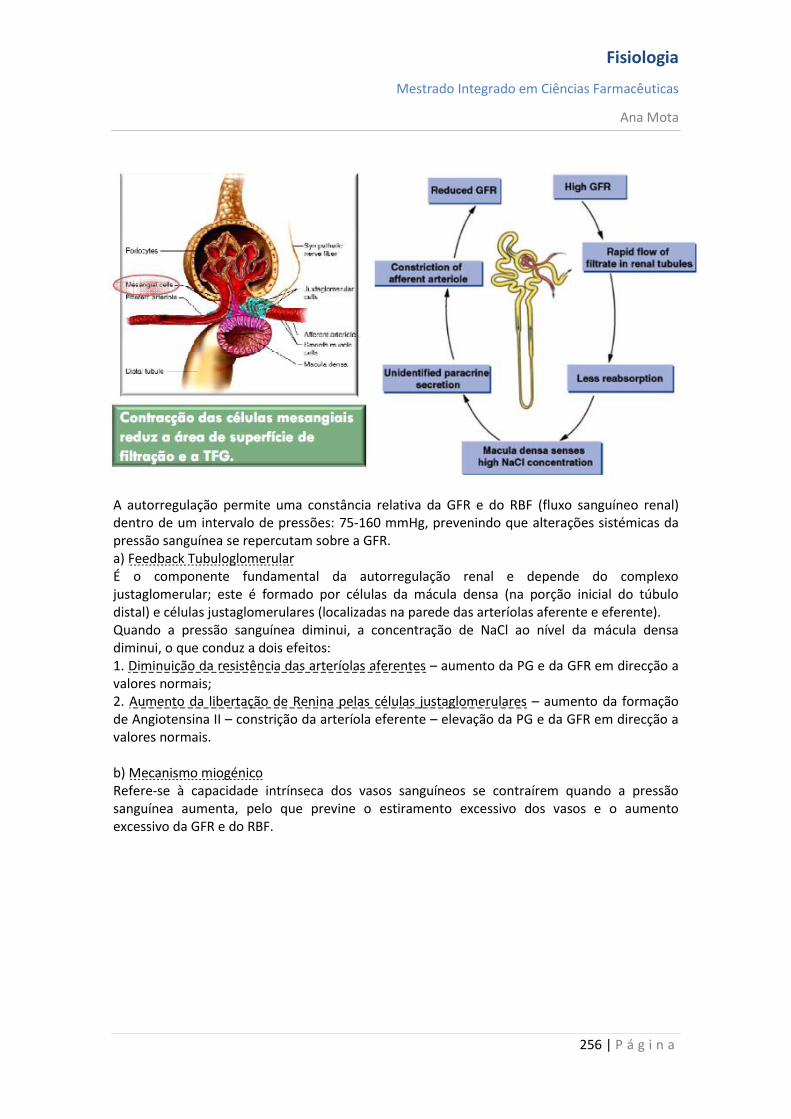
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
256 | P á g i n a
A autorregulação permite uma constância relativa da GFR e do RBF (fluxo sanguíneo renal) dentro de um intervalo de pressões: 75-160 mmHg, prevenindo que alterações sistémicas da pressão sanguínea se repercutam sobre a GFR. a) Feedback Tubuloglomerular É o componente fundamental da autorregulação renal e depende do complexo justaglomerular; este é formado por células da mácula densa (na porção inicial do túbulo distal) e células justaglomerulares (localizadas na parede das arteríolas aferente e eferente). Quando a pressão sanguínea diminui, a concentração de NaCl ao nível da mácula densa diminui, o que conduz a dois efeitos: 1. Diminuição da resistência das arteríolas aferentes – aumento da PG e da GFR em direcção a valores normais; 2. Aumento da libertação de Renina pelas células justaglomerulares – aumento da formação de Angiotensina II – constrição da arteríola eferente – elevação da PG e da GFR em direcção a valores normais. b) Mecanismo miogénico Refere-se à capacidade intrínseca dos vasos sanguíneos se contraírem quando a pressão sanguínea aumenta, pelo que previne o estiramento excessivo dos vasos e o aumento excessivo da GFR e do RBF.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
257 | P á g i n a
Regulação TFG e FSR
A pressão hidrostática glomerular e a pressão oncótica glomerular são os determinantes da GFR mais susceptíveis de controlo fisiológico, nomeadamente por intermédio do sistema nervoso simpático (SNS), hormonas e autacóides (substâncias vasoactivas libertadas pelo rim) e outros mecanismos de feedback intarrenal. 1) A activação do sistema nervoso simpático diminui a GFR – uma activação forte do SNS leva à constrição das arteríolas renais, diminuindo o fluxo sanguíneo renal e a GFR. ex: isquemia cerebral ou hemorragia grave; 2) As hormonas e autacóides controlam a GFR e o fluxo sanguíneo renal (RBF):
– Dieta rica em proteínas – aumento da GFR e do RBF (por estimulação do crescimento dos rins e por redução de resistência vascular renal); – Hiperglicemia (a glicose é cotransportada, tal como os aa, com o sódio no túbulo proximal); – Glucocorticóides – diminuem a resistência vascular renal (aumentam GFB e RBF); – Idade – diminui GFR e RBF por redução do número de nefrónios funcionantes (diminuem 10% por década a partir dos 40 anos).

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
258 | P á g i n a
Clearance Renal
Avaliação da TFG
A substância marcadora ideal da taxa de filtração seria uma substância não absorvida nem segregada pelos túbulos renais e não metabolizada nem produzida pelos rins.
Nota:
O clearance de uma substância que nem é reabsorvida nem segregada é igual à GFR.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
259 | P á g i n a
Clearance da Inulina
Clearances inferiores aos da inulina indicam reabsorção e clearances superiores secreção.
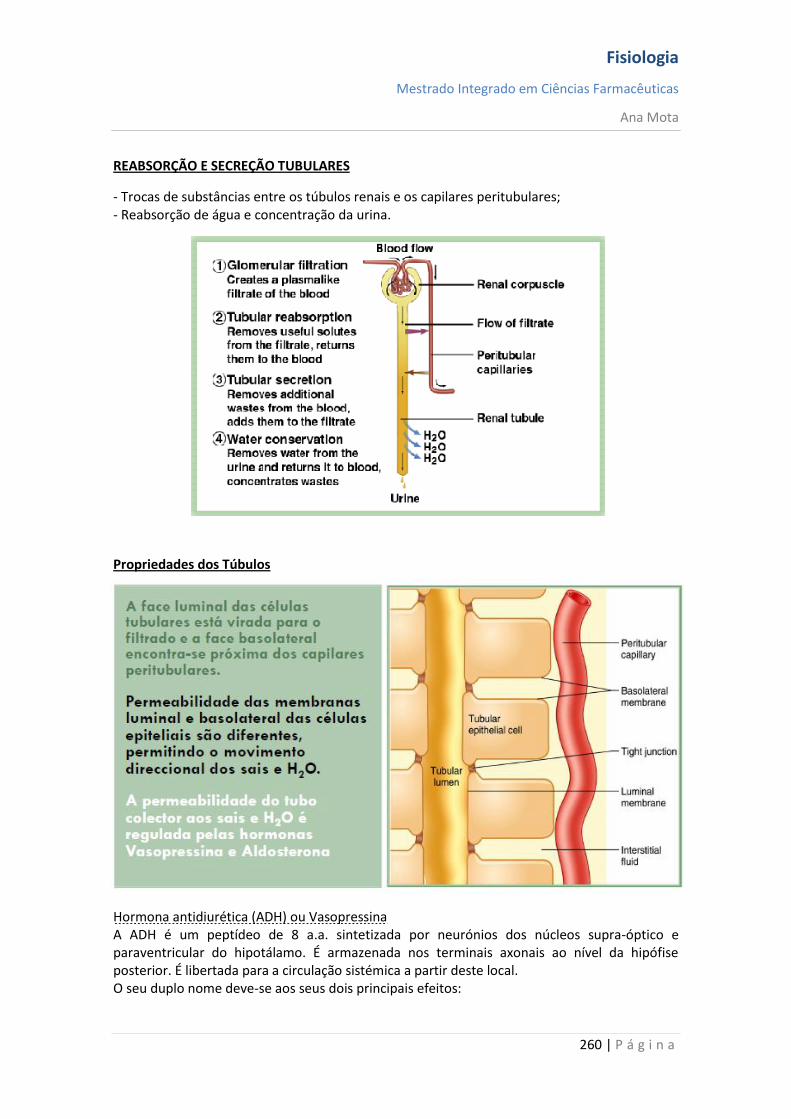
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
260 | P á g i n a
REABSORÇÃO E SECREÇÃO TUBULARES
- Trocas de substâncias entre os túbulos renais e os capilares peritubulares; - Reabsorção de água e concentração da urina.
Propriedades dos Túbulos
Hormona antidiurética (ADH) ou Vasopressina A ADH é um peptídeo de 8 a.a. sintetizada por neurónios dos núcleos supra-óptico e paraventricular do hipotálamo. É armazenada nos terminais axonais ao nível da hipófise posterior. É libertada para a circulação sistémica a partir deste local. O seu duplo nome deve-se aos seus dois principais efeitos:

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
261 | P á g i n a
a) efeito antidiurético - ocorre via receptores V2 e com concentrações plasmáticas relativamente baixas; b) efeito vasoconstritor - é mediado por receptores V1 e ocorre com concentrações plasmáticas superiores. A sua libertação é controlada por osmorreceptores (presentes no hipotálamo) e por barorreceptores periféricos. Um aumento da osmolalidade plasmática acima dos 280 mOsm/Kg provoca um aumento nos níveis plasmáticos da ADH. A sede é estimulada a osmolalidades superiores como segunda linha de defesa contra a hiperosmolalidade. Efeitos da ADH: 1) Redução do fluxo urinário e aumento da osmolalidade da urina (faz a ligação fisiológica entre as osmolalidades plasmática e urinária) 2) Aumento da permeabilidade à água do epitélio do ducto colector (10 a 20 vezes). Ao nível do ducto colector, a ADH liga-se ao receptor V2 presente na membrana basolateral, o qual conduz à formação de AMPc que activa a proteína cínase A; esta leva à produção de aquaporinas tipo 2 (por intermédio da fosforilação de proteínas ainda desconhecidas) que são inseridas na membrana apical aumentando assim a permeabilidade à água das células epiteliais do ducto colector.
Tubo Contornado Proximal
- 65% do Na+ e água filtrados são reabsorvidos ao longo do túbulo proximal; - A concentração de Na+ no fluido tubular permanece constante até ao final do túbulo proximal; - A osmolalidade do fluído tubular proximal diminui ligeiramente relativamente ao plasma; - A concentração de Cl- aumenta e a de HCO3 diminui ao longo de túbulo proximal.
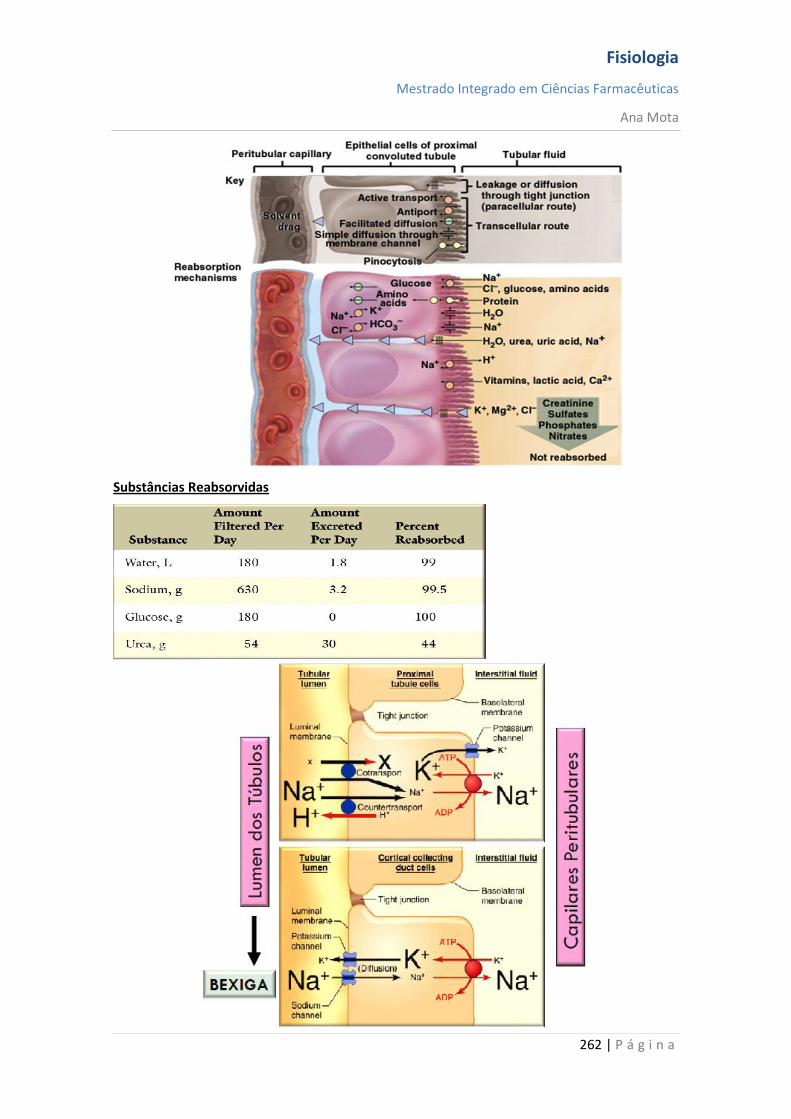
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
262 | P á g i n a
Substâncias Reabsorvidas

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
263 | P á g i n a
A maior parte do potássio (K+) é armazenado no interior das células e apenas 1 a 2% do K+ total existe no espaço extracelular. A principal via de excreção do K+ é a renal com uma contribuição intestinal reduzida (secreção de K+ pelo cólon). A concentração extracelular de K+ necessita de ser regulada com precisão porque é um determinante crítico do potencial de membrana das células excitáveis: quando a concentração plasmática de K+ diminui, há hiperpolarização e quando a sua concentração aumenta há despolarização das membranas. Estas alterações do potencial de repouso alteram a excitabilidade e a condução de impulsos nos nervos e células musculares. As consequências mais graves verificam-se a nível cardíaco: arritmias, fibrilação auricular e ventricular.
A Aldosterona, para além de reduzir a excreção de Na+, estimula a excreção renal de K+. A sua libertação é estimulada pelo aumento da concentração plasmática de K+. Os mecanismos celulares pelos quais a aldosterona actua são: a) aumento do gradiente electroquímico (pelo aumento da permeabilidade apical ao sódio); b) estimulação da ATPase Na+/K+; c) aumento da permeabilidade da membrana apical ao potássio.
A baixa concentração de K+ no lúmen aumenta o gradiente químico, favorecendo a secreção de K+. A Hormona antidiurética estimula também a secreção de K+ nos ductos colectores: aumentando a permeabilidade da membrana apical ao Na+, aumenta a secreção de K+ por despolarização da membrana apical e por estimulação da ATPase Na+/K+.
Movimento da H2O
A água movimenta-se por osmose. É o movimento da água segundo o gradiente de actividade da água, isto é, o fluxo osmótico da água ocorre dos locais de baixa concentração de partículas (alta concentração de água) para os locais de alta concentração de partículas (baixa concentração de água). É determinado pela área da membrana, pela permeabilidade à água e pela diferença da concentração da partícula impermeável. Em parte, a permeabilidade à água é determinada pelas aquaporinas, proteínas transmembranares desenhadas especificamente para a passagem da água. Este movimento

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
264 | P á g i n a
osmótico de água através dos canais pode arrastar substâncias dissolvidas de um modo indiscriminado (“Solvent drag”).
Ansa de Henle
- A reabsorção de água ocorre no ramo descendente da ansa; o ramo ascendente é impermeável à água; - De modo contrário, a reabsorção de Na+ não ocorre no ramo descendente mas 20 a 25% da reabsorção de Na+ ocorre no ramo ascendente. Por isso, o fluido tubular na parte final da ansa é sempre hipotónico.
Ansa Ascendente

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
265 | P á g i n a
Os transportadores apicais que promovem o uptake de Na+ e/ou de Cl- variam com o segmento tubular. Muitas vezes o uptake de Na+ é que possibilita o movimento de outros solutos, como glicose, a.a., H+.
Sistema Multiplicador de Contracorrente
A capacidade do rim em concentrar a urina baseia-se no princípio físico de multiplicação em contracorrente o qual tem por base anatómica a disposição em contracorrente dos ramos ascendente e descendente da ansa e a sua associação com os ductos colectores. A multiplicação em contracorrente é o mecanismo pelo qual o conteúdo de dois tubos adjacentes se desloca em direcções opostas originando concentração de solutos

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
266 | P á g i n a
progressivamente maiores. O gradiente de concentração entre os dois ramos da ansa é estabelecido com gasto de energia e tem por base 3 propriedades: 1) Transporte activo de sódio e cotransporte de potássio, cloreto e outros iões ao longo dos ramos ascendentes (porção grossa); 2) Baixa permeabilidade à água dos ramos ascendentes; 3) Alta permeabilidade à água dos ramos descendentes.
Vasos Rectos
O fluxo sanguíneo medular é relativamente baixo quando comparado com o cortical. Os capilares medulares (vasa recta) originam-se das arteríolas eferentes dos nefrónios justamedulares e dispõem-se em contracorrente: descem para a medula e ascendem em direcção ao córtex. Ao longo dos capilares descendentes o sangue fica progressivamente mais concentrado, uma vez que os vasa recta são altamente permeáveis à água e solutos. Nos ascendentes, o sangue torna-se progressivamente menos concentrado (à medida que os solutos voltam para o interstício e a água entra para os capilares. O resultado final é a manutenção da alta osmolalidade e tonicidade da medula renal. Ureia e Sistema de Contracorrente A nível renal, o sistema de contracorrente assume algumas particularidades: a) A multiplicação em contracorrentes ocorre apenas na medula externa. A concentração intersticial da NaCl aumenta em direcção à interface medula externa / medula interna (resultado do transporte de NaCl e da reduzida permeabilidade à água na porção grossa do ramo ascendente) – o aumento de pressão osmótica intersticial provoca concentração do fluido tubular do ramo descendente pela saída de água para o interstício; b) Na presença de ADH, o aumento da pressão osmótica intersticial causa também reabsorção de água nos ductos colectores ao nível da medula externa; c) As porções cortical e medular externa dos ductos colectores são impermeáveis à ureia pelo que a sua concentração aumenta proporcionalmente à reabsorção de água; d) Deste modo, é criado um gradiente de reabsorção de ureia na porção medular interna dos ductos colectores, altamente permeável à ureia. A ADH aumenta também a permeabilidade à ureia nesta região do ducto colector;
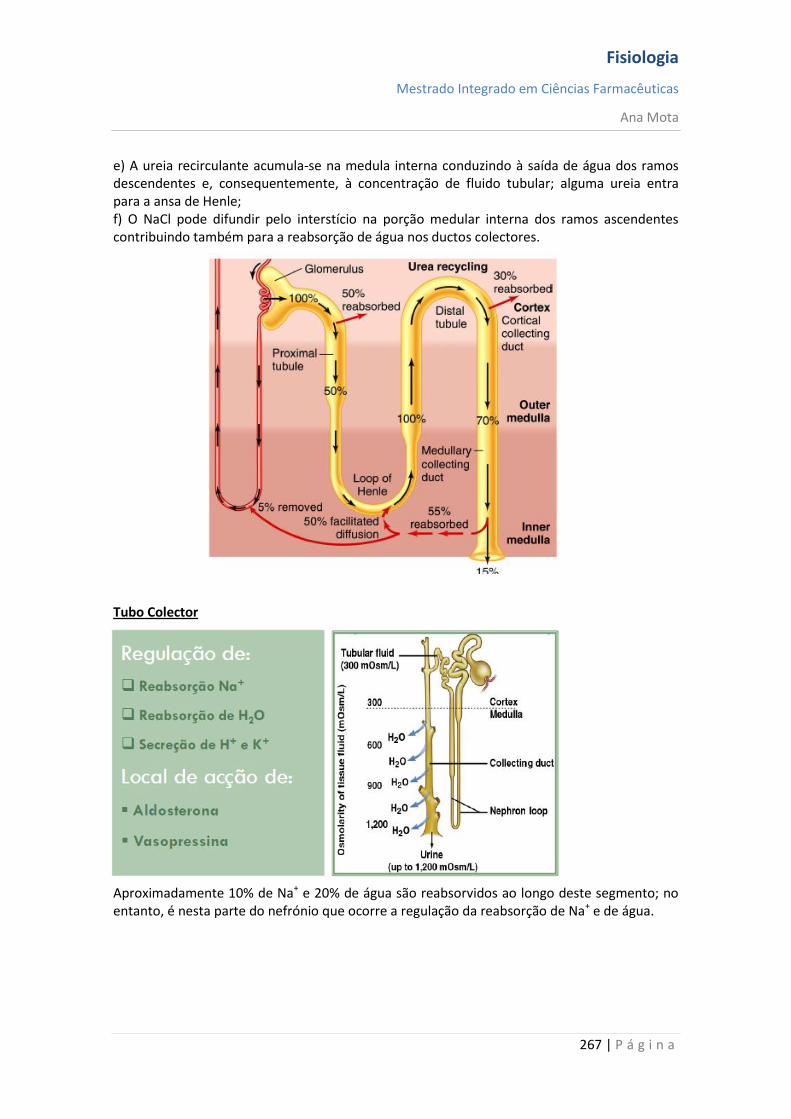
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
267 | P á g i n a
e) A ureia recirculante acumula-se na medula interna conduzindo à saída de água dos ramos descendentes e, consequentemente, à concentração de fluido tubular; alguma ureia entra para a ansa de Henle; f) O NaCl pode difundir pelo interstício na porção medular interna dos ramos ascendentes contribuindo também para a reabsorção de água nos ductos colectores.
Tubo Colector
Aproximadamente 10% de Na+ e 20% de água são reabsorvidos ao longo deste segmento; no entanto, é nesta parte do nefrónio que ocorre a regulação da reabsorção de Na+ e de água.
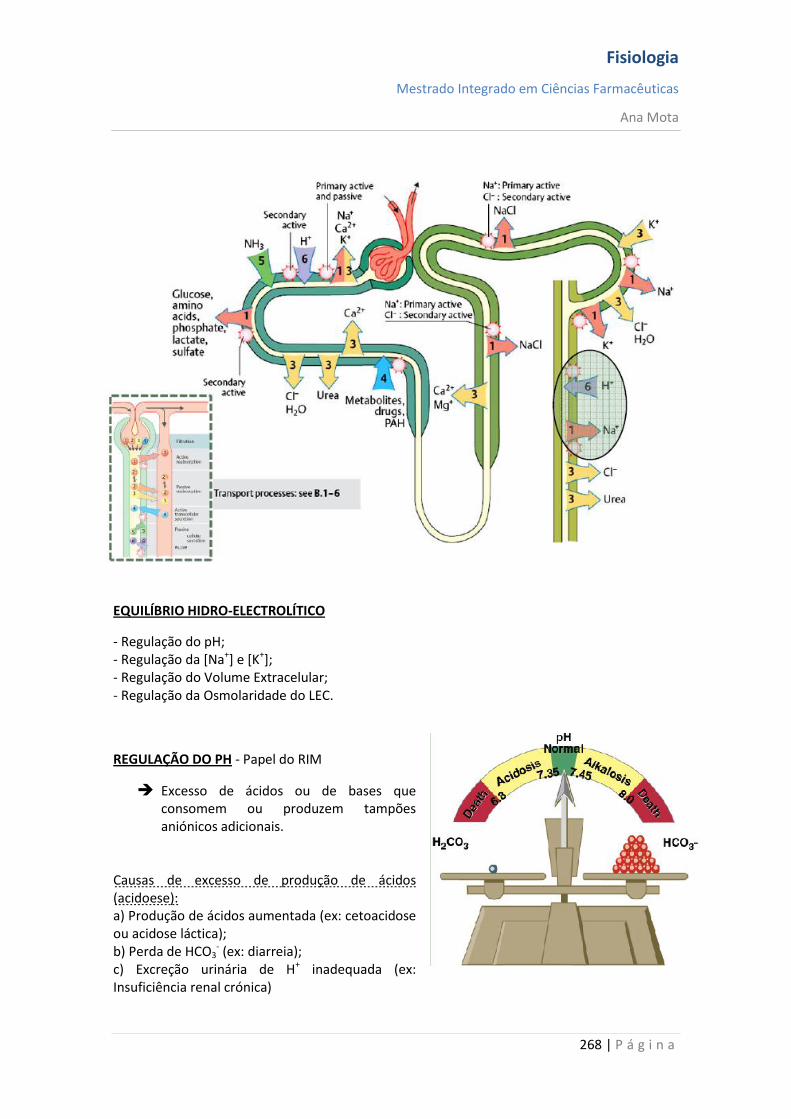
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
268 | P á g i n a
EQUILÍBRIO HIDRO-ELECTROLÍTICO
- Regulação do pH; - Regulação da [Na+] e [K+]; - Regulação do Volume Extracelular; - Regulação da Osmolaridade do LEC.
REGULAÇÃO DO PH - Papel do RIM
Excesso de ácidos ou de bases que consomem ou produzem tampões aniónicos adicionais.
Causas de excesso de produção de ácidos (acidoese): a) Produção de ácidos aumentada (ex: cetoacidose ou acidose láctica); b) Perda de HCO3
- (ex: diarreia); c) Excreção urinária de H+ inadequada (ex: Insuficiência renal crónica)

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
269 | P á g i n a
Causas de excesso de produção de bases (alcalose): a) Produção de bases aumentada (ex: dieta vegetariana); b) Perda de iões H+ (ex: vómito); c) Excreção urinária de HCO3
- inadequada (ex: Hiperaldosteronismo). A alteração primária neste tipo de desequilíbrios é a modificação da concentração de HCO3
-. Quando a concentração de HCO3- diminui, o ratio HCO3
- / Pco2 diminui (<20) e o pH diminui também, originando uma Acidose Metabólica. Na Alcalose Metabólica há aumento da concentração de HCO3
- e do pH.
Acidose e Alcalose
Acidose Metabólica Neste caso, a resposta renal lógica é a reabsorção completa de HCO3
-. A secreção renal de H+ é estimulada pelos níveis plasmáticos elevados de Aldosterona que, deste modo, auxilia a reabsorção de todo o bicarbonato bem como a titulação de tampões que não o bicarbonato. Este mecanismo de secreção de aldosterona em resposta à acidose é, em parte, dependente da secreção de renina. Assim, a angiotensina II, pelo seu efeito sobre o trocador Na+/H+, contribui directamente para o aumento da secreção de H+. O aumento da produção e excreção de amónio também contribui para o aumento da excreção de ácidos. Este mecanismo, ao contrário da formação de ácidos titulados, tem grande capacidade de reserva. Assim, primeiramente, há um aumento da excreção de amónio e, algum tempo depois, da síntese de NH4
+.
Alcalose Metabólica Teoricamente, o aumento da excreção da concentração plasmática seria facilmente corrigida pela excreção urinária do excesso de HCO3
-. No entanto, na prática clínica, este desequilíbrio é acompanhado, muitas vezes, de complicações que tendem a perpetuá-lo porque impedem a excreção eficiente de HCO3-. Este problema pode ser exemplificado com a perda de ácidos através do vómito. Nesta situação, para além da perda de iões H+, há também perda de outros iões como o Na+ e o Cl- e depleção do ECV. Inicialmente, há aumento da excreção HCO3
-. No entanto, devido às alterações concomitantes, há diminuição da GFR e aumento da reabsorção de HCO3
- o que impede a sua excreção apropriada. Deste modo, enquanto o volume não for corrigido, a alcalose metabólica irá persistir. Este distúrbio ácido-base é sempre acompanhado por depleção de K+. Processos que conduzam à depleção de potássio, como o Hiperaldosteronismo, o uso excessivo de diuréticos e a ingestão reduzida de K+, podem originar uma alcalose metabólica.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
270 | P á g i n a
Sistema Tampão
Regulação Renal do PH
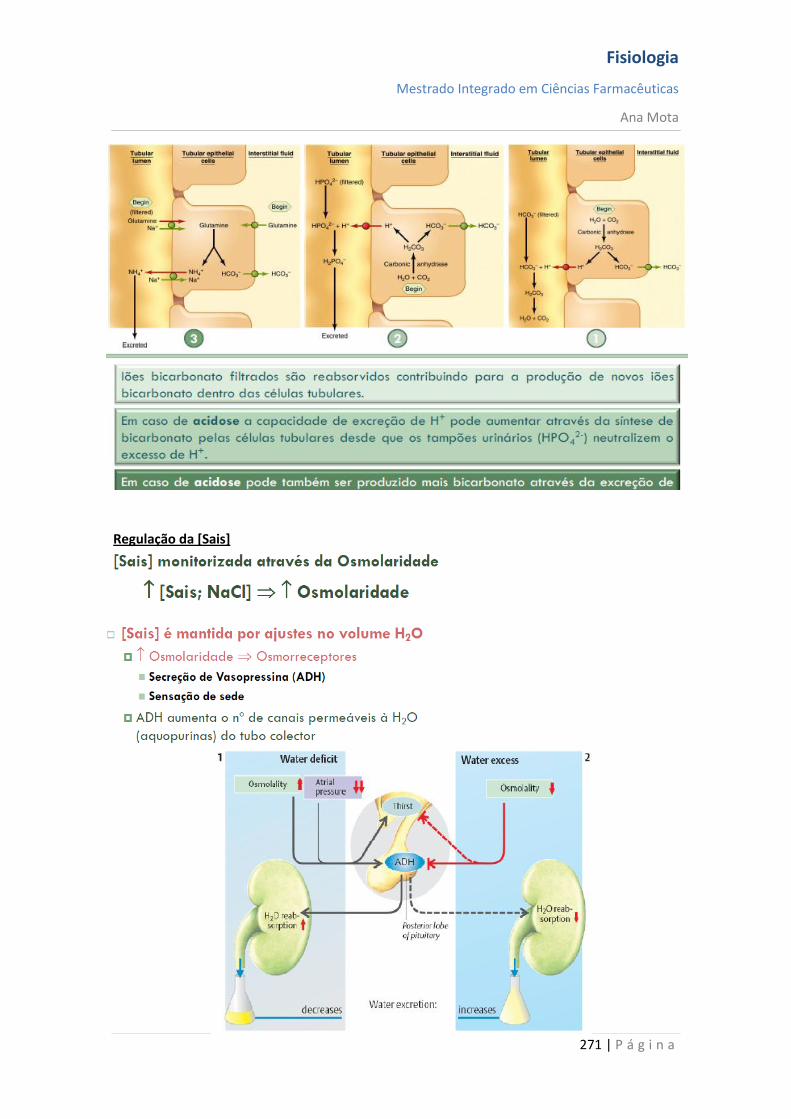
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
271 | P á g i n a
Regulação da [Sais]
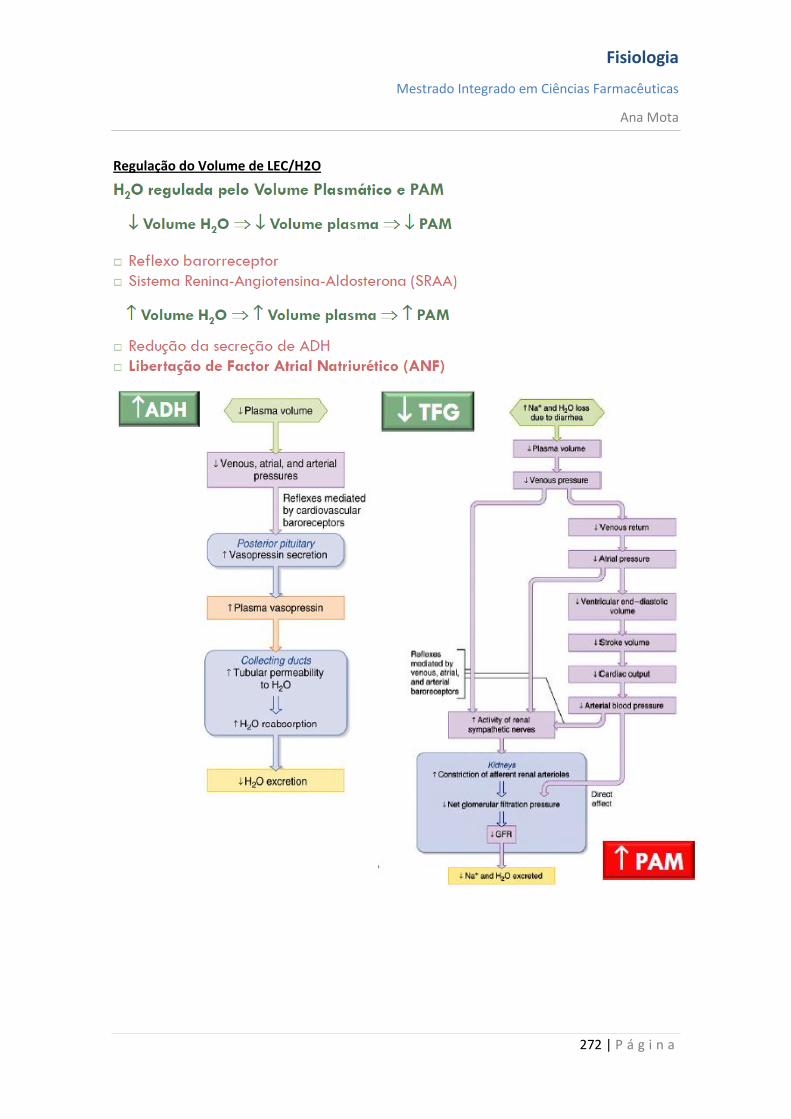
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
272 | P á g i n a
Regulação do Volume de LEC/H2O

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
273 | P á g i n a

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
274 | P á g i n a
Ingestão de água: desidratação
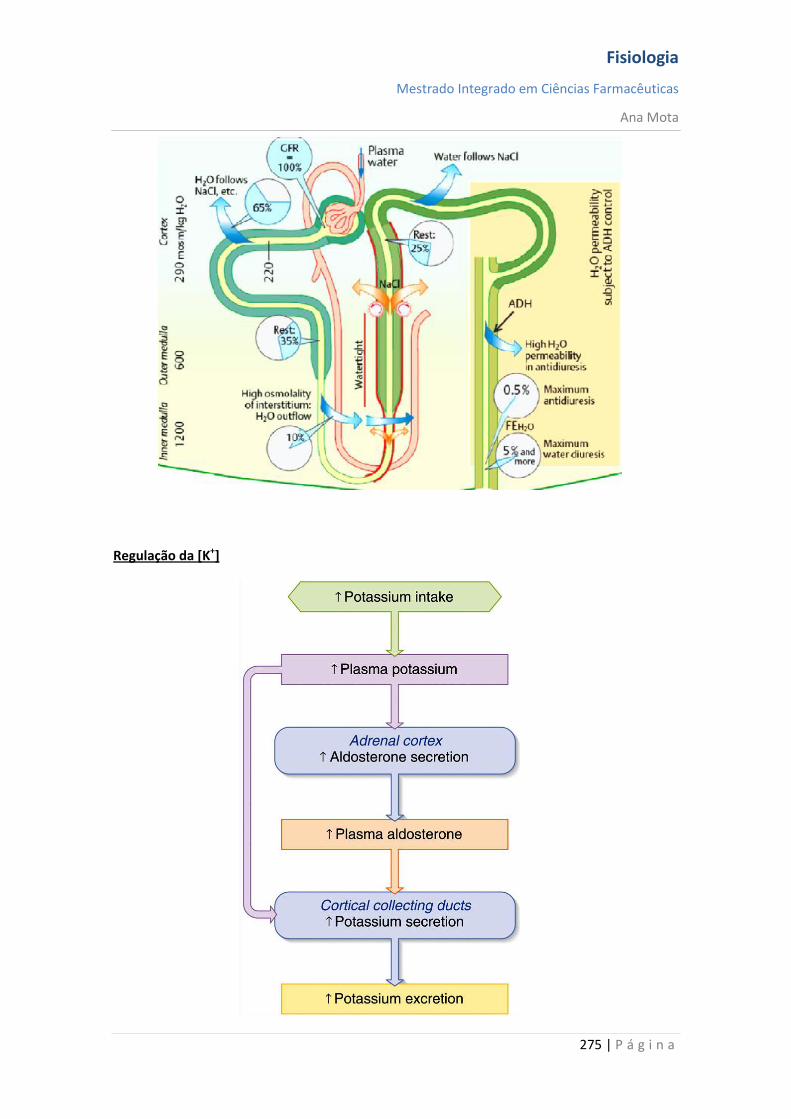
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
275 | P á g i n a
Regulação da [K+]

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
276 | P á g i n a
Falência Renal
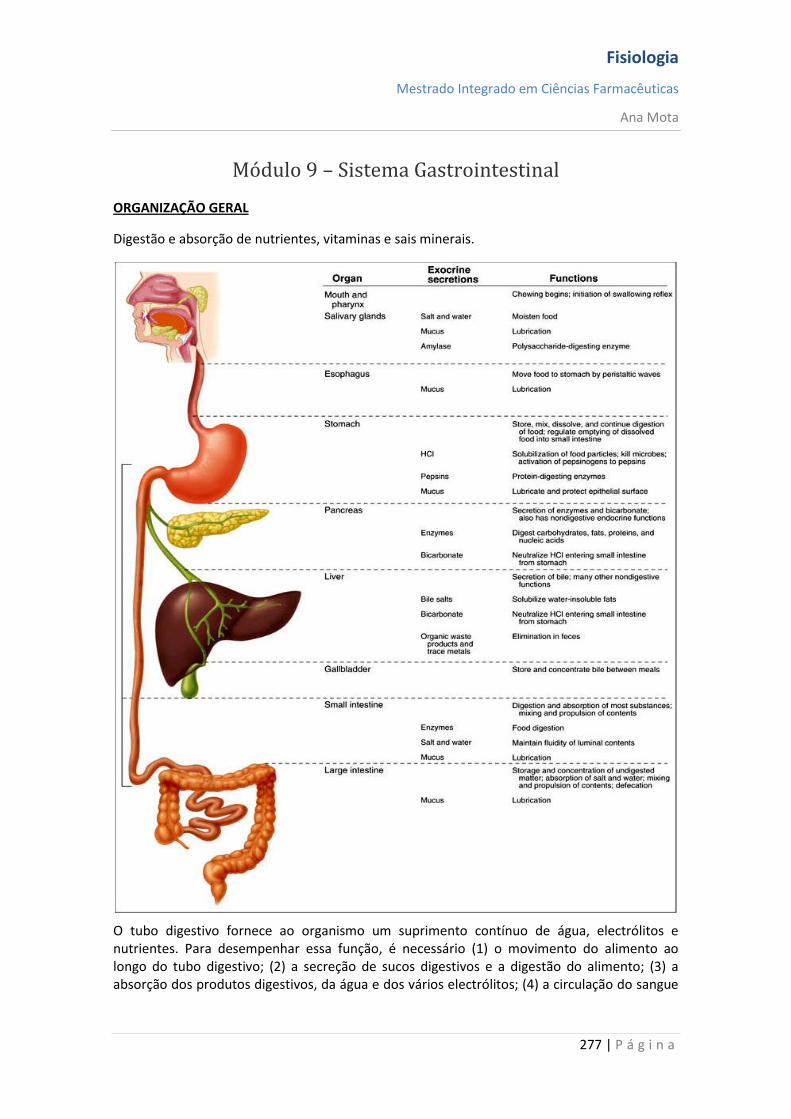
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
277 | P á g i n a
Módulo 9 – Sistema Gastrointestinal
ORGANIZAÇÃO GERAL
Digestão e absorção de nutrientes, vitaminas e sais minerais.
O tubo digestivo fornece ao organismo um suprimento contínuo de água, electrólitos e nutrientes. Para desempenhar essa função, é necessário (1) o movimento do alimento ao longo do tubo digestivo; (2) a secreção de sucos digestivos e a digestão do alimento; (3) a absorção dos produtos digestivos, da água e dos vários electrólitos; (4) a circulação do sangue
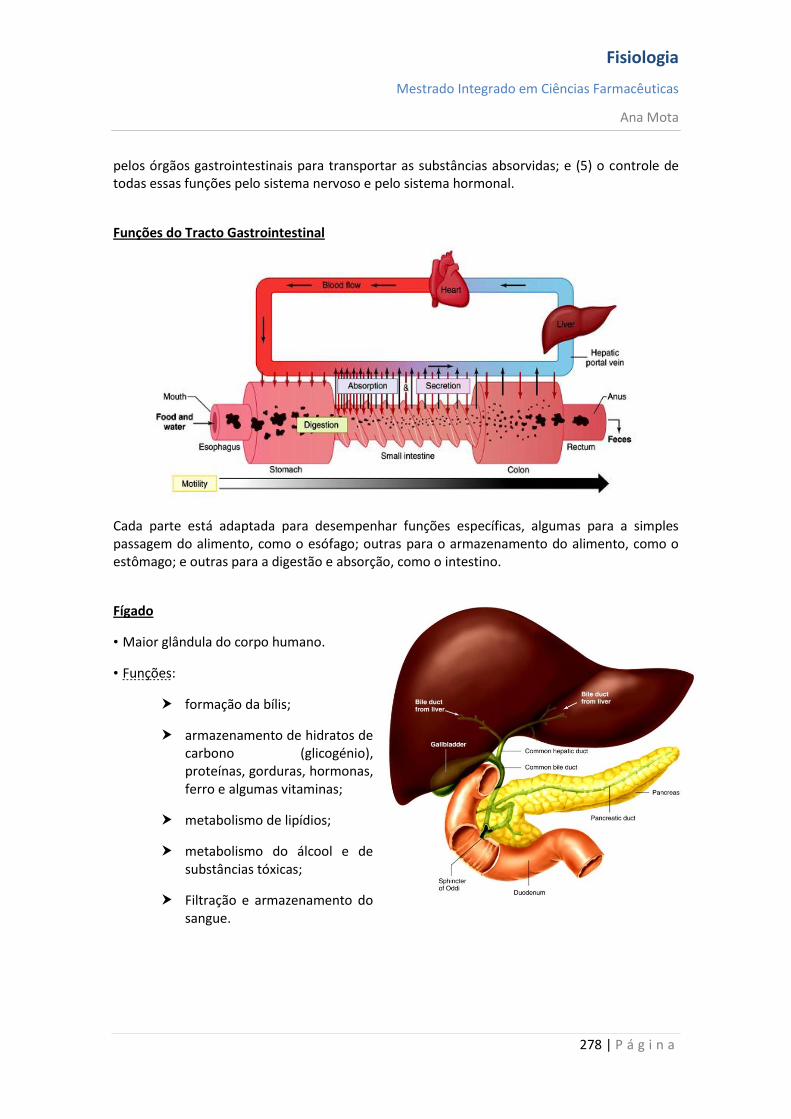
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
278 | P á g i n a
pelos órgãos gastrointestinais para transportar as substâncias absorvidas; e (5) o controle de todas essas funções pelo sistema nervoso e pelo sistema hormonal.
Funções do Tracto Gastrointestinal
Cada parte está adaptada para desempenhar funções específicas, algumas para a simples passagem do alimento, como o esófago; outras para o armazenamento do alimento, como o estômago; e outras para a digestão e absorção, como o intestino.
Fígado
• Maior glândula do corpo humano.
• Funções:
formação da bílis;
armazenamento de hidratos de carbono (glicogénio), proteínas, gorduras, hormonas, ferro e algumas vitaminas;
metabolismo de lipídios;
metabolismo do álcool e de substâncias tóxicas;
Filtração e armazenamento do sangue.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
279 | P á g i n a
Bilís sintetizada pelo fígado e armazenada na vesícula biliar emulsiona gorduras, facilitando a acção das lipases.
Secreções Digestivas
As secreções digestivas são compostas na sua maioria por água. Apenas 100 ml são excretados nas fezes. Os mecanismos de absorção de água são extremamente eficazes e cruciais para a Homeostasia. As secreções hepáticas e pancreáticas são lançadas no intestino passando através do esfíncter de ODDI.
Mucosa Intestinal
Serosa; Camada muscular longitudinal; Camada muscular circular;
Submucosa; Mucosa.
Além disso, existe uma camada difusa de fibras musculares lisas, a muscular da mucosa, localizada nas camadas mais profundas da mucosa. As funções motoras do intestino são executadas pelas diferentes camadas de músculo liso.
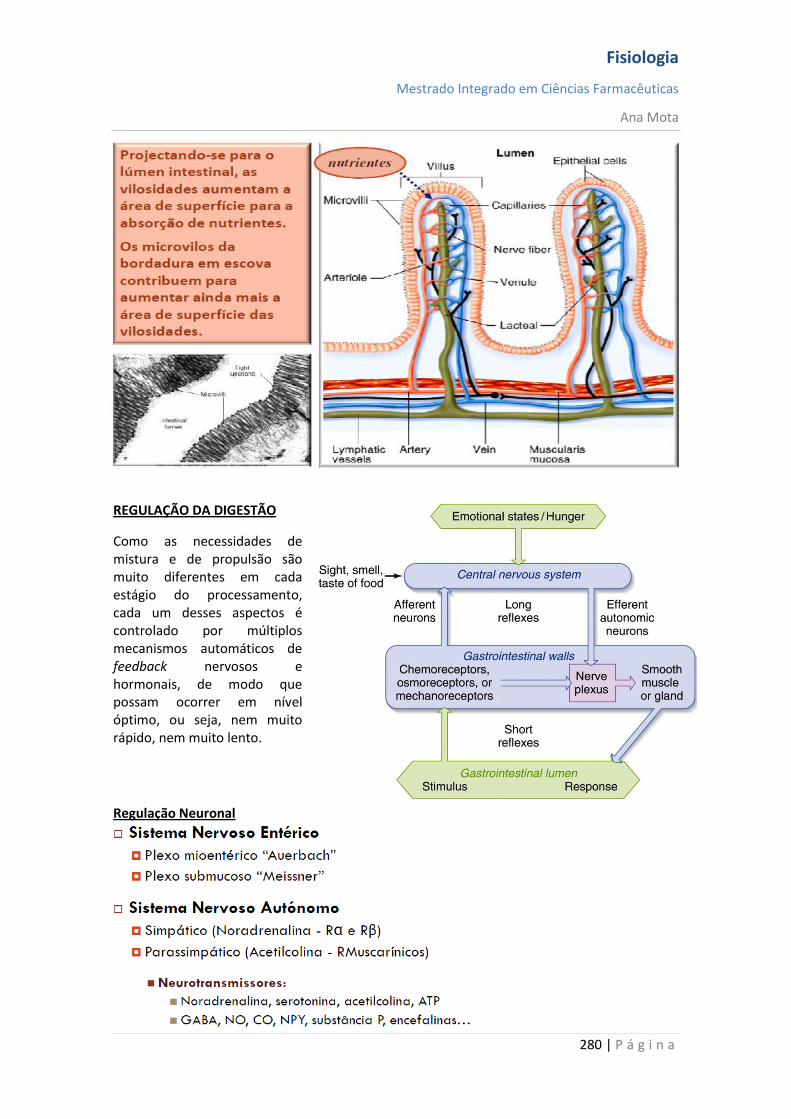
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
280 | P á g i n a
REGULAÇÃO DA DIGESTÃO
Como as necessidades de mistura e de propulsão são muito diferentes em cada estágio do processamento, cada um desses aspectos é controlado por múltiplos mecanismos automáticos de feedback nervosos e hormonais, de modo que possam ocorrer em nível óptimo, ou seja, nem muito rápido, nem muito lento.
Regulação Neuronal

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
281 | P á g i n a
Plexos Nervosos
Actividade Eléctrica e Motilidade
Os sucos digestivos do estômago são secretados pelas glândulas gástricas e entram imediatamente em contacto com o alimento armazenado, situado na superfície mucosa do estômago; quando o estômago está cheio, ondas constritoras peristálticas fracas, também denominadas ondas de mistura, movem-se em direcção ao antro, ao longo da parede do estômago, com frequência aproximada de uma a cada 20 segundos. Essas ondas são iniciadas pelo ritmo eléctrico basal (BER) que consiste em "ondas lentas" eléctricas que ocorrem espontaneamente na parede do estômago. Na maioria das partes do tubo gastrintestinal, essas ondas não são fortes o suficiente para causar contracções, a não ser que desencadeiem inicialmente potenciais de acção; todavia, no estômago, os picos positivos elevam-se acima do limiar para a excitação, até mesmo na ausência de potenciais de acção.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
282 | P á g i n a
Regulação Hormonal
Uma hormona GI é uma substância produzida por um tipo de célula endócrina dispersa por uma região relativamente bem definida do tracto GI proximal. Daqui é libertada para o sangue por um estímulo específico (actividade nervosa, distensão, estimulação química), atingindo o órgão alvo, onde regula a motilidade ou a secreção ou até a proliferação das suas células. A secretina, gastrina e CCK são as hormonas GI clássicas. A nova biologia das hormonas GI continua a afirmar que estas hormonas são peptídeos libertados para o sangue por um estímulo apropriado. Contudo, surgiram novas características reveladas pelas modernas técnicas de biologia molecular: 1. existem múltiplas hormonas agrupadas em diversas famílias; 2. cada gene tem diversos fenótipos; 3. diversos órgãos, para além do tracto GI, exprimem os genes destas hormonas, nomeadamente células neuroendócrinas (da adeno-hipófise, glândula tiróide ou supra-renal), células do sistema nervoso central (córtex cerebral, cerebelo, hipotálamo) ou dos tractos respiratório e urogenital, entre outros; 4. as células processam as pró-hormonas de forma específica, reflectindo a possibilidade de utilizar um gene para a síntese de diversos peptídeos bioactivos.
Como já foi referido, existem cinco hormonas GI que têm uma função perfeitamente estabelecida: secretina, colecistocinina (CCK), gastrina, peptídeo gástrico inibitório (GIP) e motilina. Os outros peptídeos são hormonas candidatas.
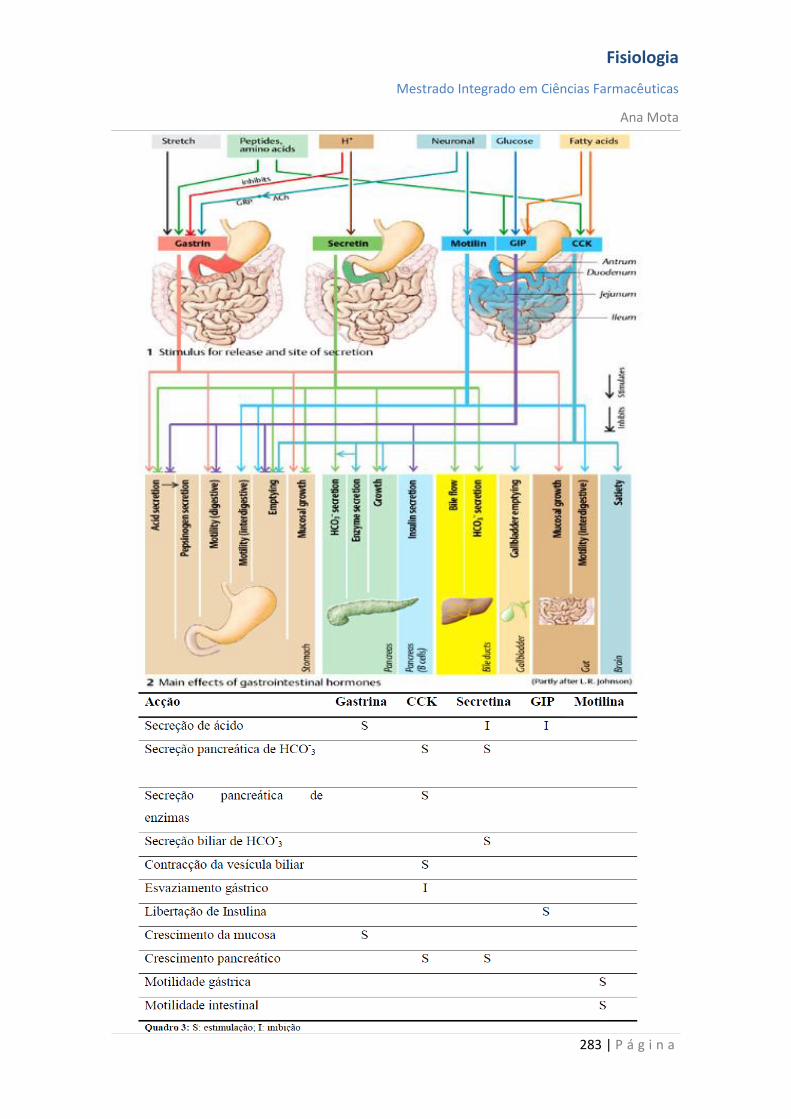
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
283 | P á g i n a
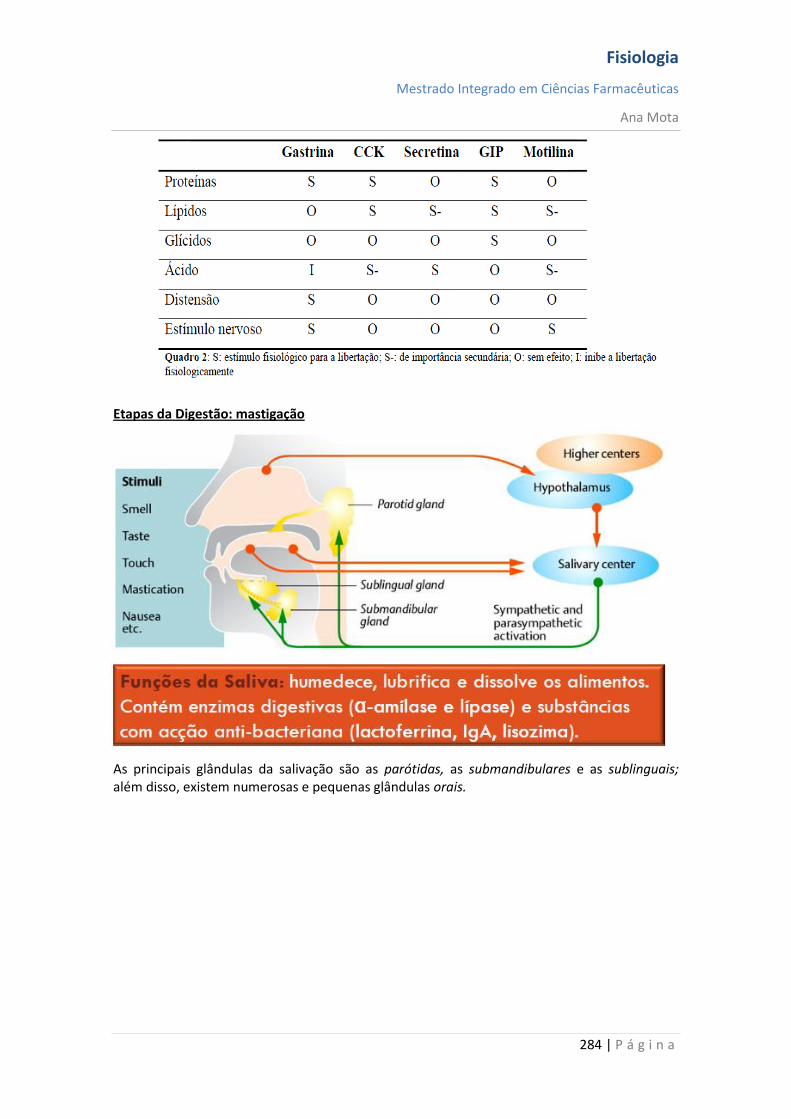
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
284 | P á g i n a
Etapas da Digestão: mastigação
As principais glândulas da salivação são as parótidas, as submandibulares e as sublinguais; além disso, existem numerosas e pequenas glândulas orais.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
285 | P á g i n a
Secreções Salivares
A saliva contém quantidades especialmente grandes de iões potássio e bicarbonato. Por outro lado, as concentrações de iões sódio e cloreto são consideravelmente menores na saliva do que no plasma. Os iões sódio são activamente reabsorvidos dos ductos salivares, enquanto os iões potássio sofrem secreção activa, porém com menor velocidade, em troca do sódio. Por conseguinte, a concentração de sódio da saliva fica muito reduzida, enquanto a concentração de iões potássio aumenta. O excesso de reabsorção de sódio em relação à secreção de potássio cria negatividade de cerca de -70 mV nos ductos salivares, ocasionando reabsorção passiva dos iões cloreto. Por conseguinte, a concentração de iões cloreto cai para níveis muito baixos, juntamente com a redução da concentração de iões sódio. Os iões bicarbonato são secretados pelo epitélio ductal para o lúmen do ducto. Essa secreção é, pelo menos em parte, causada pela troca de bicarbonato por iões cloreto, mas também pode resultar, em parte, de um processo secretor activo. O resultado final desses processos de transporte activo é que, em condições de repouso, as concentrações de iões sódio e cloreto na saliva são de apenas cerca de 15 mEq/l cada uma, ou seja, aproximadamente um sétimo a um décimo de suas concentrações no plasma. Por outro lado, a concentração de iões potássio é de cerca de 30 mEq/l, ou seja, sete vezes maior que sua concentração no plasma, e a concentração de iões bicarbonato é de 50 a 70 mEq/l, representando cerca de duas a três vezes sua concentração plasmática. Durante a salivação máxima, as concentrações salivares de iões modificam-se de modo considerável, uma vez que a velocidade de formação da secreção primária pelos ácinos pode aumentar por até 20 vezes. Como consequência, a secreção flui pelos ductos com tanta rapidez que a reconstituição da secreção por esses ductos fica muito reduzida. Por conseguinte, quando são secretadas quantidades copiosas de saliva, a concentração de cloreto de sódio eleva-se de cerca da metade a dois terços da concentração observada no plasma, enquanto a concentração de potássio cai para apenas quatro vezes a do plasma.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
286 | P á g i n a
Na presença de secreção excessiva de aldosterona, a reabsorção de sódio e cloreto e a secreção de potássio aumentam acentuadamente, de modo que a concentração de cloreto de sódio na saliva está algumas vezes reduzida para quase zero, enquanto a concentração de potássio aumenta ainda mais. Em qualquer estado anormal em que ocorra perda de saliva para o exterior do organismo durante longos períodos de tempo, o indivíduo pode apresentar grave depleção de iões potássio no organismo, devido à elevada concentração desse ião na saliva, resultando, eventualmente, em hipocalemia grave e paralisia.
A acetilcolina estimula a secreção ácida através da ligação a receptores muscarínicos M3 na membrana basal da célula parietal. Esta ligação abre canais de Ca2+, permitindo a entrada de Ca2+ para dentro da célula, o que estimula a secreção de HCl. Tal como a gastrina, a acetilcolina também estimula a libertação de histamina a partir das células enterochromaffin-like (ECL).
Receptores M1: efeitos excitatórios. Ex: secreção de ácido gástrico que ocorre após estimulação do nervo vago é mediada por estes receptores;
Receptores M3: efeitos excitatórios. Ex: no músculo liso visceral, a activação dos receptores pela acetilcolina resulta em contracção.
Etapas da Digestão: deglutição
Foram já identificados diversos neurotransmissores nos interneurónios do sistema nervoso entérico, como acetilcolina, serotonina, peptídeo intestinal vasoactivo (VIP), NO e somatostatina. A acetilcolina e as taquicininas (como a substância P) foram identificadas em neurónios que exercem uma acção excitatória sobre as células musculares; o VIP e o NO em neurónios inibitórios para o músculo.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
287 | P á g i n a
A deglutição é um mecanismo complicado, principalmente porque a faringe exerce, na maioria das vezes, várias outras funções além da deglutição, sendo transformada, por apenas alguns segundos de cada vez, em via para a propulsão do alimento. É muito importante que a respiração não fique comprometida devido à deglutição. Pode ser dividida em:
Fase voluntária, que inicia o processo de deglutição;
Fase faríngea, que é involuntária e consiste na passagem do alimento pela faringe até o esófago;
Fase esofágica, outra fase involuntária que promove a passagem do alimento da faringe para o estômago.
Fase voluntária da deglutição Quando o alimento está pronto para ser deglutido, é "voluntariamente" comprimido ou empurrado para trás, na faringe, como consequência da pressão da língua para cima e para trás contra o palato. Daí em diante, o processo da deglutição torna-se quase totalmente automático e, em geral, não pode ser interrompido. Fase faríngea da deglutição Quando o bolo alimentar penetra na faringe, ele estimula áreas receptoras da deglutição situadas em torno da abertura da faringe, sobretudo nos dois pilares das amígdalas, e impulsos dessas áreas dirigem-se até o tronco cerebral para iniciar uma série de contracções musculares automáticas da faringe, da seguinte maneira: 1. O palato mole é empurrado para cima, a fim de fechar a parte posterior das narinas, impedindo, dessa maneira, o refluxo do alimento para as cavidades nasais; 2. As pregas palatofaríngeas de cada lado da faringe são puxadas medialmente e aproximam-se uma da outra. Dessa maneira, essas dobras formam uma fenda sagital através da qual o alimento deve passar para a faringe posterior. Essa fenda também exerce acção selectiva, permitindo que o alimento adequadamente mastigado passe com facilidade, enquanto a passagem de grandes objectos é impedida. Como essa fase da deglutição tem duração de menos de 1 segundo, qualquer objecto grande tem geralmente sua passagem impedida da faringe para o esófago; 3. As cordas vocais da laringe aproximam-se estreitamente, e a laringe é puxada para cima e para a frente pelos músculos do pescoço. Essa acção, combinada com a presença de ligamentos que impedem o movimento da epiglote para cima, faz com que ela se dobre para trás por sobre a abertura da laringe. Esses dois efeitos impedem a passagem do alimento para a traqueia. A aproximação das cordas vocais é muito importante, mas a epiglote ajuda a evitar que o alimento chegue até as cordas vocais. A destruição das cordas vocais ou dos músculos que as aproximam pode provocar estrangulamento. Por outro lado, a remoção da epiglote não costuma produzir dano sério para a deglutição; 4. O movimento da laringe para cima também aumenta a abertura do esófago. Ao mesmo tempo, os 3 a 4 centímetros superiores da parede muscular esofágica, uma área denominada esfíncter esofágico superior ou esfíncter faringoesofágico, relaxam-se, permitindo que o alimento se mova com facilidade e sem restrição da faringe posterior para a parte superior do esófago. Esse esfíncter, entre as deglutições, permanece fortemente contraído, impedindo,

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
288 | P á g i n a
assim, que o ar penetre no esófago durante a respiração. O movimento da laringe para cima também eleva a glote, retirando-a da corrente principal do fluxo alimentar, de modo que o alimento passe geralmente em ambos os lados da epiglote, e não sobre sua superfície; essa manobra constitui outra protecção contra a passagem do alimento para a traqueia; 5. Ao mesmo tempo que a laringe é elevada e o esfíncter faringoesofágico é relaxado, toda a parede muscular da faringe se contrai, começando na parte superior da faringe e propagando-se para baixo sob forma de onda peristáltica rápida ao longo dos músculos faríngeos médio c inferior e, daí, para o esófago, propelindo o alimento para esse órgão. Em resumo, a mecânica da fase faríngea da deglutição inclui o fechamento da traqueia, a abertura do esófago e o aparecimento de onda peristáltica rápida que se origina na faringe, forçando o bolo alimentar para o esófago superior. Todo o processo ocorre em 1 a 2 segundos. Fase esofágica da deglutição A principal função do esófago é a de conduzir o alimento da faringe para o estômago, sendo que seus movimentos estão especificamente organizados para a execução dessa função. Em condições normais, o esófago exibe dois tipos de movimentos peristálticos - o peristaltismo primário e o peristaltismo secundário. O peristaltismo primário refere-se simplesmente à continuação da onda peristáltica que começa na faringe e se propaga para o esófago durante a fase faríngea da deglutição. Essa onda passa da faringe para o estômago em aproximadamente 8 a 10 segundos. Todavia, o alimento deglutido por uma pessoa na posição erecta é geralmente transferido para a porção inferior do esófago com muito mais rapidez do que a própria onda peristáltica, isto é, em cerca de 5 a 8 segundos, devido ao efeito adicional da gravidade que empurra o alimento para baixo. Se a onda peristáltica primária for insuficiente para movimentar todo o alimento que penetrou no esófago em direcção ao estômago, surgem ondas peristálticas secundárias, que resultam da distensão do esófago pelo alimento retido. Essas ondas, que prosseguem até que o alimento chegue ao estômago, são iniciadas, em parte, por circuitos neurais intrínsecos do sistema nervoso entérico do esófago e, em parte, por reflexos transmitidos por fibras aferentes vagais do esófago para o bulbo e, daí, novamente, de volta para o esófago por fibras eferentes vagais. A musculatura da faringe e do quarto superior do esófago consiste em músculo estriado. Por conseguinte, as ondas peristálticas nessas regiões são controladas apenas por impulsos nervosos esqueléticos nos nervos glossofaríngeo e vago. Nos dois terços inferiores do esófago, a musculatura é lisa, mas essa porção do esófago também é fortemente controlada pelos nervos vagos que actuam por meio de suas conexões com o sistema nervoso entérico. Todavia, quando os nervos vagos para o esófago são seccionados, o plexo nervoso mioentérico do esófago torna-se excitável o suficiente, depois de vários dias, para gerar fortes ondas peristálticas secundárias, mesmo sem o apoio dos reflexos vagais. Por conseguinte, mesmo após paralisia do reflexo da deglutição, o alimento forçado de alguma outra forma no esófago superior e, a seguir, empurrado pela gravidade até o esófago inferior ainda passa facilmente para o estômago. Acalasia é uma condição em que o esfíncter esofágico inferior é incapaz de se relaxar durante o mecanismo da deglutição. Em consequência, a passagem do alimento do esófago para o estômago é impedida. Estudos patológicos sugeriram que a base fisiológica dessa condição reside em alguma patologia ou ausência de função do plexo mioentérico dos dois terços

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
289 | P á g i n a
inferiores do esófago. A musculatura da porção inferior do esófago permanece, então, contraída de maneira incoordenada, e o plexo mioentérico perde a capacidade de transmitir um sinal para causar o "relaxamento receptivo" do esfíncter gastroesofágico à medida que o alimento se aproxima dessa área durante o processo de deglutição.
Etapas da Digestão: estômago
Além das células secretoras de muco que revestem toda a superfície do estômago, a mucosa gástrica possui dois tipos importantes de glândulas tubulares: as glândulas oxínticas (ou gástricas) e as glândulas pilóricas.
As glândulas oxínticas (que formam ácido) secretam ácido clorídrico, pepsinogénio, factor intrínseco e muco, enquanto as glândulas pilóricas secretam principalmente muco para protecção da mucosa pilórica, bem como algum pepsinogénio e, sobretudo, a hormona gastrina;
As glândulas oxínticas localizam-se nas superfícies internas do corpo e do fundo do estômago, ocupando os 80% proximais do estômago. As glândulas pilóricas localizam-se no antro do estômago.
À medida que a onda peristáltica esofágica se propaga para o estômago, uma onda de relaxamento, transmitida por neurónios inibitórios mioentéricos, precede o peristaltismo. Além disso, todo o estômago e, em menor grau, o duodeno relaxam-se à medida que essa onda alcança a extremidade inferior do esófago, de modo que ficam preparados antecipadamente para receber o alimento propelido para o esófago durante o acto da deglutição.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
290 | P á g i n a
Basicamente, o esvaziamento gástrico é promovido pela intensidade das contracções peristálticas do antro. Ao mesmo tempo, o esvaziamento é dificultado por graus variáveis de resistência à passagem do quimo no piloro. Na maioria das vezes, as contracções peristálticas do antro são fracas e funcionam principalmente para misturar o alimento com as secreções gástricas. Todavia, cerca de 20% das vezes, enquanto o alimento ainda está no estômago, essas contracções ficam muito intensas, começando ao nível da incisura angular do estômago e, a seguir, propagando-se por todo o antro, não mais como contracções fracas de mistura, porém sob forma de poderosas constrições peristálticas semelhantes a anéis. Com o esvaziamento progressivo do estômago, essas contracções começam cada vez mais distantes no corpo do estômago, isolando gradualmente as porções mais baixas do alimento armazenado e juntando esse alimento ao quimo no antro. Essas contracções peristálticas intensas geram quase sempre até 50 a 70 cm de pressão de água, o que corresponde a cerca de seis vezes o tipo habitual de ondas peristálticas de mistura. Por conseguinte, a intensidade desse peristaltismo antral constitui o principal factor que determina a velocidade do esvaziamento gástrico. Quando o tônus pilórico está normal, cada uma dessas poderosas ondas peristálticas antrais força vários mililitros de quimo para o interior do duodeno. Por conseguinte, as ondas peristálticas exercem uma acção bombeadora, que costuma ser denominada "bomba pilórica".

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
291 | P á g i n a
A abertura distal do estômago é o piloro. Nessa região, o músculo circular torna-se 50 a 100% mais espesso do que nas porções iniciais do antro gástrico. Além disso, o piloro permanece em ligeira contracção tónica quase todo o tempo. Por conseguinte, o músculo circular pilórico é quase sempre denominado esfíncter pilórico. Apesar da contracção tónica do esfíncter pilórico, o piloro em geral permanece ligeiramente aberto, o suficiente para permitir que a água e outros líquidos deixem o estômago com facilidade. Por outro lado, a constrição costuma impedir a passagem da maioria das partículas alimentares até haver mistura com o quimo, adquirindo uma consistência quase líquida. Todavia, o grau de constrição do piloro pode aumentar ou diminuir sob a influência de sinais nervosos e humorais provenientes do estômago e do duodeno. Por conseguinte, o piloro, bem como o antro gástrico, também actua no controle do esvaziamento gástrico.
Não apenas os reflexos nervosos do duodeno para o estômago inibem o esvaziamento gástrico, como também as hormonas libertadas da porção proximal do intestino exercem o mesmo efeito. O estímulo para a produção das hormonas provém principalmente da gordura que chega ao duodeno, embora outros tipos de alimentos possam aumentar as hormonas em menor grau. Ao penetrar no duodeno, a gordura extrai várias hormonas diferentes do epitélio duodenal e jejunal, ligando-se a "receptores" nas células epiteliais ou através de algum outro mecanismo. Por sua vez, as hormonas são transportados pelo sangue até o estômago onde: (1) inibem a actividade da bomba pilórica e, ao mesmo tempo, (2) aumentam ligeiramente a força de contracção do esfíncter pilórico. Esses efeitos são importantes, visto que a digestão das gorduras é muito mais lenta do que a da maioria dos outros alimentos. Infelizmente, as hormonas precisas que provocam inibição hormonal do estômago por feedback não estão totalmente definidas. Todavia, o mais potente deles parece ser a colecistocinina (CCK), que é liberada pela mucosa do jejuno em resposta a substâncias gordurosas presentes no quimo. Essa hormona actua como inibidor competitivo, bloqueando a maior motilidade gástrica causada pela gastrina. Outra hormona é a secretina, que é libertada principalmente pela mucosa duodenal em resposta ao ácido gástrico liberado do estômago pelo piloro. Essa hormona exerce efeito geral, porém apenas fraco, ao reduzir a motilidade gastrintestinal. Por fim, a hormona denominada peptídio inibidor gástrico (GIP), libertada pela porção superior do intestino delgado em resposta principalmente à presença de gorduras no quimo, mas não a hidratos de carbono, também é conhecido por sua capacidade de inibir a motilidade gástrica em certas condições.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
292 | P á g i n a
Produção de HCl
Quando estimuladas, as células parietais secretam uma solução ácida contendo cerca de 160 mmol de HCl por litro, que é quase isotónica em relação aos líquidos corporais. O pH desse ácido é de cerca de 0,8, ilustrando a sua extrema acidez. Nesse pH, a concentração de iões hidrogénio é cerca de 3 milhões de vezes a do sangue arterial. E, para concentrar os iões hidrogénio, essa enorme quantidade requer mais de 1500 calorias de energia por litro de suco gástrico. O ácido clorídrico é formado nas membranas semelhantes a vilosidades dos canalículos e, a seguir, é conduzido para o exterior. Foram formuladas diferentes hipóteses para o mecanismo preciso da formação de ácido clorídrico. Um desses mecanismos propostos é: a. O ião cloreto é activamente transportado do citoplasma da célula parietal para o lúmen do canalículo, enquanto os iões sódio são activamente transportados para fora do lúmen. Esses dois efeitos combinados criam um potencial negativo de -40 a -70 mV no canalículo, o que, por sua vez, determina a difusão passiva de iões potássio de carga positiva, bem como de pequeno número de iões sódio do citoplasma celular para o interior dos canalículos. Assim, com efeito, o cloreto de potássio e quantidades muito menores de cloreto de sódio penetram nos canalículos; b. A água dissocia-se em iões hidrogénio e iões hidroxilo no citoplasma celular. Os iões hidrogénio são, então, activamente secretados para o interior dos canalículos em troca de iões potássio, sendo esse processo de troca activa catalisado pela H+/K+-ATPase. Além disso, os iões sódio sofrem reabsorção activa por uma bomba de sódio distinta. Por conseguinte, a maior parte dos iões potássio e iões sódio que se difundiram para os canalículos é reabsorvida, enquanto os iões hidrogénio tomam o seu lugar, formando uma solução muito forte de ácido clorídrico; c. A água atravessa a célula e penetra nos canalículos por osmose. Por conseguinte, a secreção final que penetra nos canalículos contém ácido clorídrico na concentração de 155 mEq/1, cloreto de potássio na concentração de 15 mEq/1 e uma quantidade muito pequena de cloreto de sódio; d. Por fim, o dióxido de carbono, formado durante o metabolismo no interior da célula ou proveniente do sangue, combina-se, sob a influência da anidrase carbónica, com iões hidroxilo (formados na etapa a, quando ocorreu a dissociação da água), formando iões bicarbonato. A seguir, esses iões bicarbonato difundem-se para fora da célula, pelo interior do líquido extracelular, em troca de iões cloreto que penetram na célula e que mais tarde serão secretados nos canalículos. A importância do dióxido de carbono nas reacções químicas para a

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
293 | P á g i n a
formação de ácido clorídrico é ilustrada pelo fato de a inibição da anidrase carbónica pela acetazolamida diminuir a formação de ácido clorídrico.
Regulação da produção de HCl
Gastrina Os estímulos nervosos (vagais ou locais), para além de uma estimulação directa das células parietais, conduzem também à libertação de gastrina a partir das células G antrais e da região pilórica. A gastrina é libertada para a corrente sanguínea e acaba por atingir as glândulas da mucosa gástrica (acção endócrina). A gastrina não tem uma acção excitatória directa nas células parietais muito potente. As suas acções devem-se sobretudo a um efeito indirecto sobre a libertação de histamina. Liga-se a receptores CCK-B que aumentam a concentração intracelular de Ca2+.
Histamina A histamina é o principal mediador fisiológico da secreção de HCl. É libertada por células enterochromaffin-like (ECL) localizadas na mucosa gástrica. Tem uma acção parácrina. As células ECL libertam histamina sobretudo após estimulação pela gastrina e pela acetilcolina. A histamina liga-se a receptores H2 na membrana celular da célula parietal que aumentam a concentração intracelular de AMPc, estimulando a secreção ácida. Estimulação nervosa A estimulação nervosa da secreção gástrica pode ter início a partir de estímulos provenientes do cérebro ou do estômago. Os sinais com origem no estômago activam dois tipos de reflexos: - reflexos vagovagais longos, que são transmitidos desde a mucosa gástrica até ao tronco cerebral e daí até ao estômago pelo nervo vago; - reflexos curtos, com origem local e que se localizam inteiramente no sistema nervoso entérico. Os estímulos que podem desencadear estes reflexos incluem a distensão gástrica, estímulos tácteis na mucosa gástrica e a presença de estímulos químicos (nomeadamente aminoácidos, peptídeos e ácido). O neurotransmissor libertado por todas as terminações nervosas é a acetilcolina, com uma excepção. As terminações que enervam as células G libertam GRP (gastrinreleasing peptide). A acetilcolina estimula a secreção ácida através da ligação a receptores muscarínicos M3 na membrana basal da célula parietal. Esta ligação abre canais de Ca2+, permitindo a entrada de Ca2+ para dentro da célula, o que estimula a secreção de HCl. Tal como a gastrina, a acetilcolina também estimula a libertação de histamina a partir das células enterochromaffin-like (ECL). Efeito multiplicativo dos mediadores Nenhum dos mediadores atrás descritos é eficaz quando actua isoladamente. De facto, para que exista uma secreção substancial de ácido é necessária a presença simultânea dos três. Existe uma secreção basal, não estimulada, de pequenas quantidades histamina. Quando existe uma estimulação nervosa (vagal ou local), que liberta Acetilcolina e GRP, todos os mediadores passam a estar presentes, o que origina uma secreção abundante de ácido pelas células parietais (acção de potenciação).
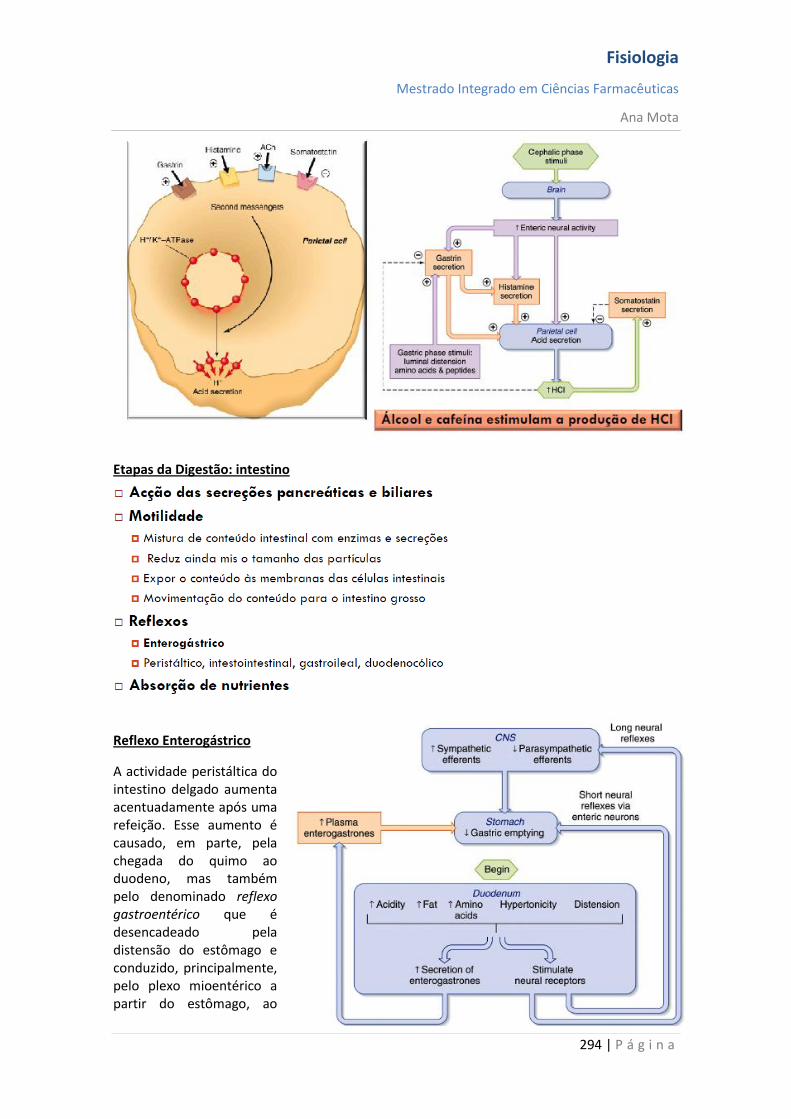
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
294 | P á g i n a
Etapas da Digestão: intestino
Reflexo Enterogástrico
A actividade peristáltica do intestino delgado aumenta acentuadamente após uma refeição. Esse aumento é causado, em parte, pela chegada do quimo ao duodeno, mas também pelo denominado reflexo gastroentérico que é desencadeado pela distensão do estômago e conduzido, principalmente, pelo plexo mioentérico a partir do estômago, ao

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
295 | P á g i n a
longo da parede do intestino delgado. Além dos sinais nervosos que afectam o peristaltismo do intestino delgado, também é afectado por diversos factores hormonais. Essas hormonas incluem gastrina, colecistocinina, insulina e serotonina, que aumentam a motilidade intestinal e são secretadas durante as várias fases do processamento do alimento. Por outro lado, a secretina e o glucagon inibem a motilidade do intestino delgado. Infelizmente, a importância quantitativa de cada um desses factores hormonais no controle da motilidade ainda é questionável. A função das ondas peristálticas no intestino delgado não consiste apenas em causar progressão do quimo em direcção à válvula ileocecal, mas também em espalhar o quimo ao longo da mucosa intestinal. Quando o quimo proveniente do estômago penetra no intestino e provoca a distensão inicial do intestino proximal, as ondas peristálticas desencadeadas começam imediatamente a espalhar o quimo ao longo do intestino, sendo que esse processo se intensifica à medida que maior quantidade de quimo penetra no duodeno. Ao alcançar a válvula ileocecal, o quimo é por vezes bloqueado durante várias horas, até que a pessoa faça outra refeição, quando um novo reflexo gastroentérico (também denominado gastroileal) intensifica o peristaltismo do íleo e força o restante do quimo através da válvula ileocecal para o cego.
Secreções Pancreáticas
O pâncreas, situado paralelamente e abaixo do estômago, é uma grande glândula composta, cuja estrutura interna se assemelha à das glândulas salivares. Além da secreção de insulina pelas ilhotas de Langerhans, os acinos do pâncreas também secretam enzimas digestivas, e grandes volumes de solução de bicarbonato de sódio são secretados pelos pequenos dútulos e pelos ductos maiores que se originam dos acinos. O produto combinado flui, então, pelo longo ducto pancreático que habitualmente se une ao ducto hepático imediatamente antes de desaguar no duodeno pelo esfíncter de Oddi. O suco pancreático é secretado em grandes quantidades em resposta à presença de quimo nas porções superiores do intestino delgado; as características desse suco são determinadas, em certo grau, pelos tipos de alimentos contidos no quimo.
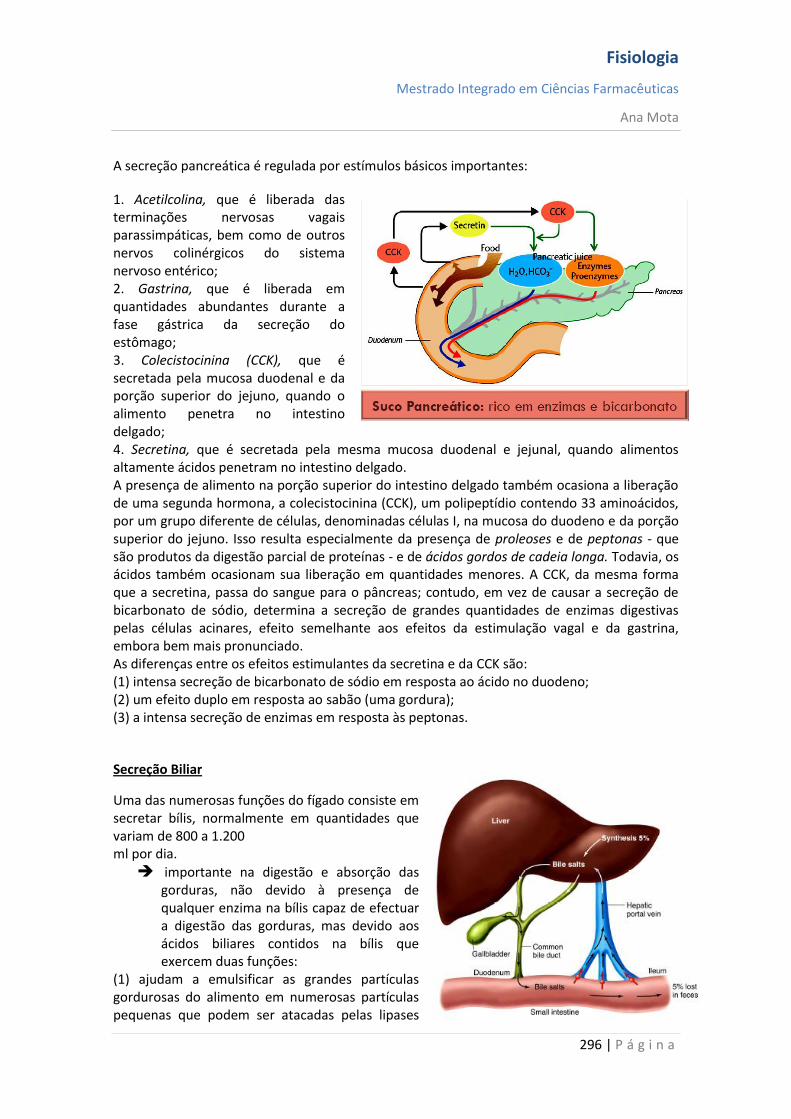
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
296 | P á g i n a
A secreção pancreática é regulada por estímulos básicos importantes:
1. Acetilcolina, que é liberada das terminações nervosas vagais parassimpáticas, bem como de outros nervos colinérgicos do sistema nervoso entérico; 2. Gastrina, que é liberada em quantidades abundantes durante a fase gástrica da secreção do estômago; 3. Colecistocinina (CCK), que é secretada pela mucosa duodenal e da porção superior do jejuno, quando o alimento penetra no intestino delgado; 4. Secretina, que é secretada pela mesma mucosa duodenal e jejunal, quando alimentos altamente ácidos penetram no intestino delgado. A presença de alimento na porção superior do intestino delgado também ocasiona a liberação de uma segunda hormona, a colecistocinina (CCK), um polipeptídio contendo 33 aminoácidos, por um grupo diferente de células, denominadas células I, na mucosa do duodeno e da porção superior do jejuno. Isso resulta especialmente da presença de proleoses e de peptonas - que são produtos da digestão parcial de proteínas - e de ácidos gordos de cadeia longa. Todavia, os ácidos também ocasionam sua liberação em quantidades menores. A CCK, da mesma forma que a secretina, passa do sangue para o pâncreas; contudo, em vez de causar a secreção de bicarbonato de sódio, determina a secreção de grandes quantidades de enzimas digestivas pelas células acinares, efeito semelhante aos efeitos da estimulação vagal e da gastrina, embora bem mais pronunciado. As diferenças entre os efeitos estimulantes da secretina e da CCK são: (1) intensa secreção de bicarbonato de sódio em resposta ao ácido no duodeno; (2) um efeito duplo em resposta ao sabão (uma gordura); (3) a intensa secreção de enzimas em resposta às peptonas.
Secreção Biliar
Uma das numerosas funções do fígado consiste em secretar bílis, normalmente em quantidades que variam de 800 a 1.200 ml por dia. importante na digestão e absorção das
gorduras, não devido à presença de qualquer enzima na bílis capaz de efectuar a digestão das gorduras, mas devido aos ácidos biliares contidos na bílis que exercem duas funções:
(1) ajudam a emulsificar as grandes partículas gordurosas do alimento em numerosas partículas pequenas que podem ser atacadas pelas lipases

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
297 | P á g i n a
secretadas no suco pancreático; (2) ajudam no processo de transporte e absorção dos produtos terminais da gordura digerida através da membrana da mucosa intestinal.
Meio de excreção do sangue para vários produtos de degradação importantes. Incluem especialmente a bilirrubina, um produto terminal da destruição da hemoglobina e excesso de colesterol, sintetizado pelas células hepáticas.
A bílis é secretada pelo fígado em duas etapas: (1) inicialmente, a bílis é secretada pelos hepatócitos; essa secreção inicial contém grandes quantidades de ácidos biliares, colesterol e outros constituintes orgânicos, que são secretadas para os diminutos canalículos biliares localizados entre as células hepáticas nas lâminas hepáticas; (2) A seguir, a bílis flui perifericamente para os septos interlobulares, onde os canalículos desaguam nos autos biliares terminais; a seguir, flui por ductos progressivamente maiores até alcançar finalmente o ducto hepático e o ducto colédoco, pelo qual a bílis desagua directamente no duodeno ou é desviada para a vesícula biliar por meio do ducto cístico. Em seu trajecto por esses ductos biliares, a bílis inicial recebe uma secreção secundária. Essa secreção adicional consiste em solução aquosa de iões sódio e bicarbonato, que, algumas vezes, aumenta a quantidade total de bílis por até 100%. A secreção secundária é estimulada pela secretina, resultando em quantidades aumentadas de ião bicarbonato que suplementam as secreções pancreáticas na neutralização do ácido proveniente do estômago.
Ácidos Biliares
Os ácidos biliares são compostos esteróides anfipáticos derivados do catabolismo enzimático do colesterol pelo fígado por acção da 7-α-hidroxilase. Os ácidos biliares primários (essencialmente os ácidos cólico e quenodesoxicólico) são sintetizados nos hepatócitos e conjugados no terminal carboxilo com taurina ou com glicina. Esta amidação aumenta o carácter anfipático e diminui as constantes de ionização dos ácidos biliares, tornando--os mais hidrofílicos e mais facilmente excretáveis para a bile. Os glicuronoconjugados, que predominam em humanos, e os tauroconjugados, predominantes em roedores, têm valores de Ka de aproximadamente 4 e 2, respectivamente e existem predominantemente na forma de sais aniónicos, a pH fisiológico. Assim, são chamados de sais biliares.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
298 | P á g i n a
O estímulo mais potente que ocasiona as contracções da vesícula biliar é a hormona CCK. Trata-se da mesma colecistocinina que determina a secreção aumentada de enzimas pelas células acinares do pâncreas. O estímulo para sua liberação no sangue a partir da mucosa duodenal consiste principalmente na entrada de alimentos gordurosos no duodeno. Todavia, além da CCK, a vesícula biliar também é estimulada, ainda que menos acentuadamente, por fibras nervosas colinérgicas dos vagos e do sistema nervoso entérico. Trata-se dos mesmos nervos que promovem a motilidade e a secreção em outras partes do tubo GI superior. Até mesmo com a ocorrência de contracções relativamente fortes da vesícula biliar, o esvaziamento ainda pode ser difícil porque o esfíncter de Oddi permanece normalmente num estado de contracção tónica. Por conseguinte, para que haja esvaziamento, o esfíncter também deve estar relaxado. Pelo menos três factores diferentes ajudam nesse processo de relaxamento:
1. A CCK, em vez de estimular o esfíncter de Oddi, possui fraco efeito relaxante; todavia, esse efeito em si não costuma ser suficiente para permitir a ocorrência de esvaziamento significativo;
2. As contracções rítmicas da vesícula biliar transmitem ondas peristálticas ao longo do ducto colédoco em direcção ao esfíncter de Oddi, causando uma onda de relaxamento que inibe parcialmente o esfíncter adiante da onda peristáltica. Mas isso também não costuma ser suficiente para permitir o esvaziamento de grandes quantidades;
3. Quando as ondas peristálticas intestinais propagam-se pela parede do duodeno, a fase de relaxamento de cada uma dessas ondas relaxa acentuadamente o esfíncter de Oddi, juntamente com o relaxamento da parede intestinal. Esse parece ser incontestavelmente o mais potente de todos os efeitos relaxantes sobre o esfíncter de Oddi. Como consequência, a bile geralmente penetra no duodeno sob forma de “esguichos” que são sincronizados com as contracções peristálticas do duodeno.
Em suma, a vesícula biliar lança seu depósito de bile concentrada no duodeno, principalmente em resposta ao estímulo da CCK. Quando não existe gordura na refeição, a vesícula biliar esvazia-se pouco; entretanto, na presença de quantidades adequadas de gordura, a vesícula biliar esvazia-se por completo em cerca de 1 hora.
As células hepáticas formam cerca de 0,5 g de sais biliares diariamente. O precursor dos sais biliares é o colesterol, que é fornecido na dieta ou sintetizado nas células hepáticas durante o metabolismo das gorduras, sendo então convertido em ácido eólico ou ácido quenodesoxicólico, em quantidades aproximadamente iguais. A seguir, esses ácidos combinam-se principalmente com a glicina e, em menor grau, com a taurina, formando os ácidos biliares glico e tauroconjugados. Os sais desses ácidos são secretados na bile. Os sais biliares exercem duas acções importantes no tubo intestinal:
1. Possuem acção detergente sobre as partículas de gordura do alimento, porque diminuem a tensão superficial das partículas e permitem que a agitação no tubo intestinal desintegre os glóbulos de gordura em partículas com pequenas dimensões. Esse processo é conhecido como função emulsificadora ou detergente dos sais biliares;
2. Ajudam na absorção de ácidos gordos, monoglicerídios, colesterol e outros lipídios do tubo intestinal. Exercem essa função pela formação de minúsculos complexos com esses lípidos.
Esses complexos, denominados micelas, são altamente solúveis devido às cargas eléctricas dos sais biliares. Os lípidos são transportados nessa forma até a mucosa, onde são então
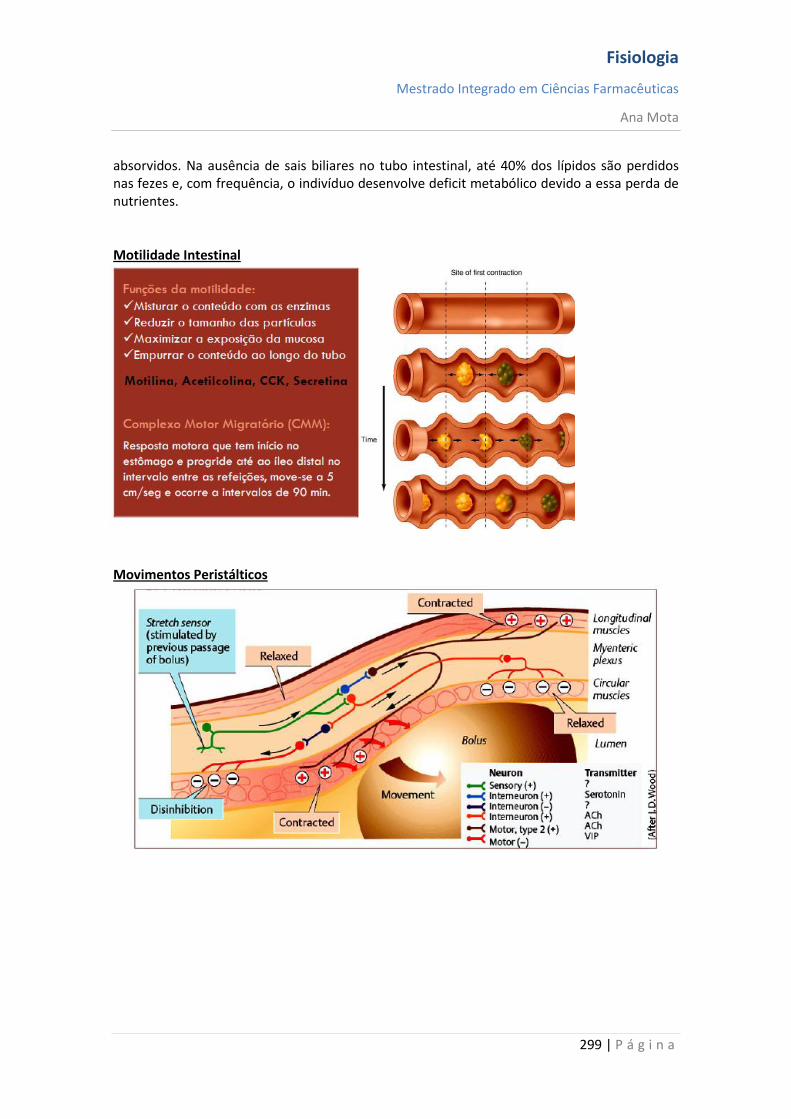
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
299 | P á g i n a
absorvidos. Na ausência de sais biliares no tubo intestinal, até 40% dos lípidos são perdidos nas fezes e, com frequência, o indivíduo desenvolve deficit metabólico devido a essa perda de nutrientes.
Motilidade Intestinal
Movimentos Peristálticos
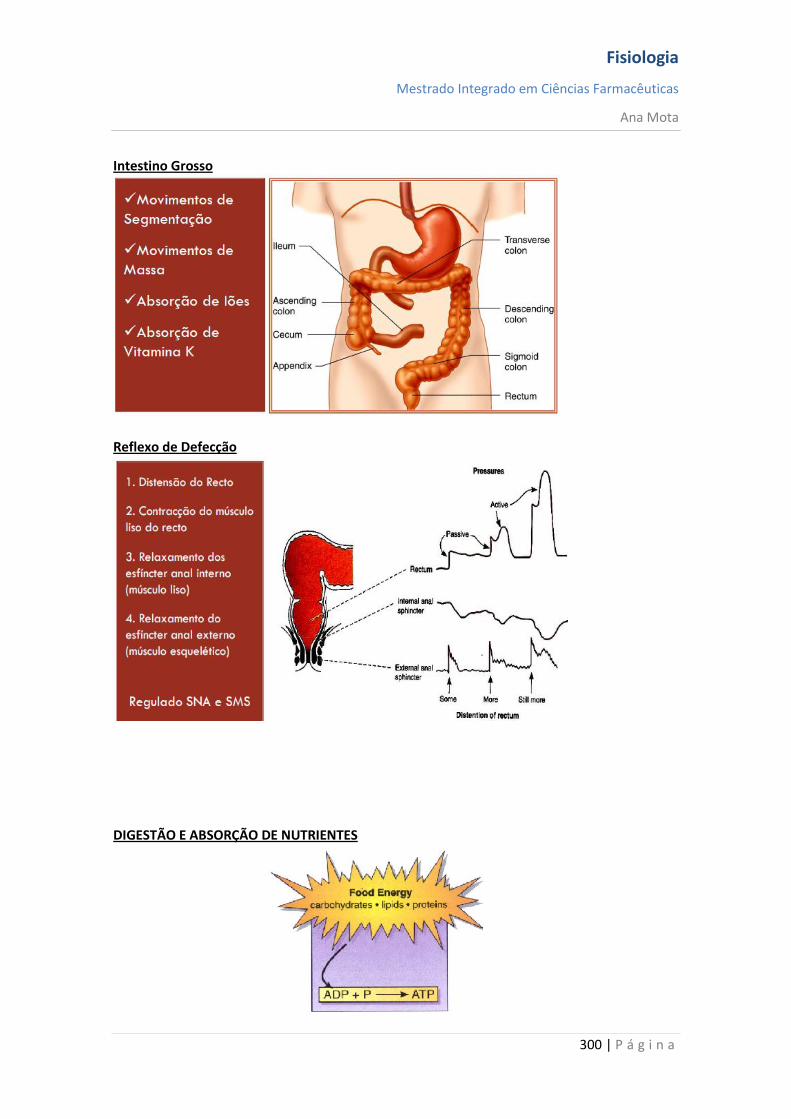
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
300 | P á g i n a
Intestino Grosso
Reflexo de Defecção
DIGESTÃO E ABSORÇÃO DE NUTRIENTES

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
301 | P á g i n a
HIDRATOS DE CARBONO
Principais H.C ingeridos na dieta Enzimas digestivas Local de digestão e absorção
HIDRATOS DE CARBONO DA DIETA
Apenas os Monossacarídeos podem
ser directamente absorvidos.
Ligações Glicosídicas
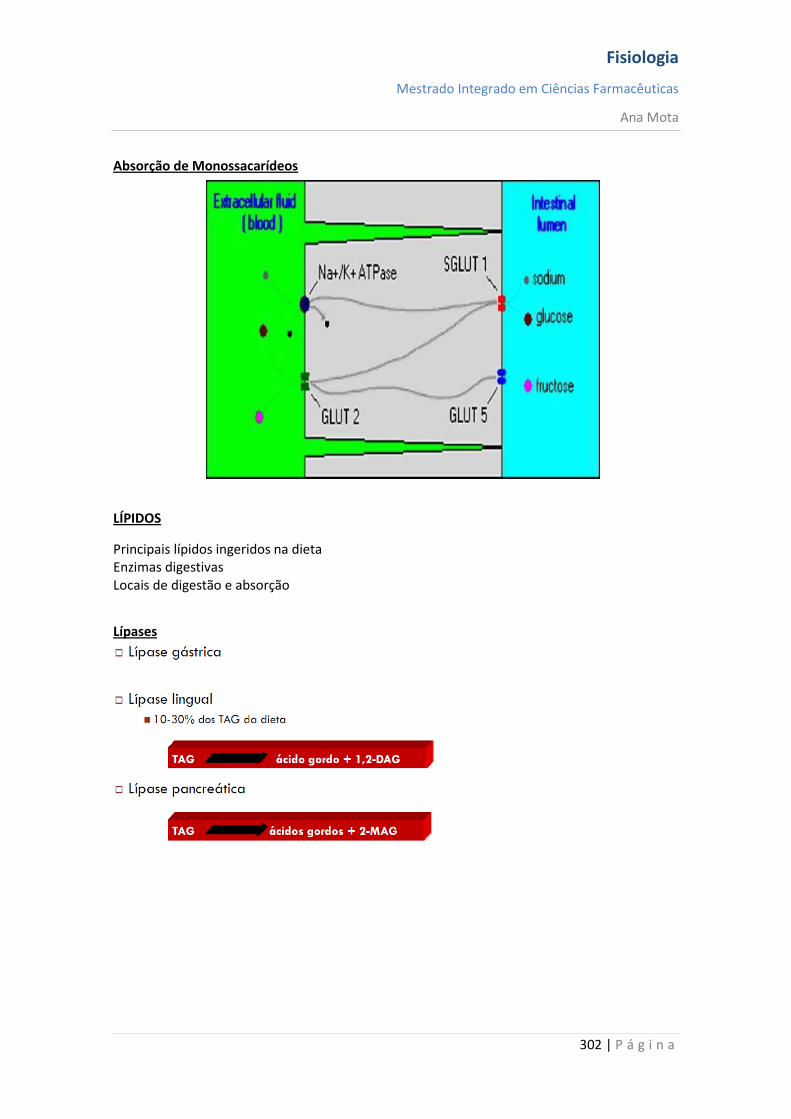
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
302 | P á g i n a
Absorção de Monossacarídeos
LÍPIDOS
Principais lípidos ingeridos na dieta Enzimas digestivas Locais de digestão e absorção
Lípases
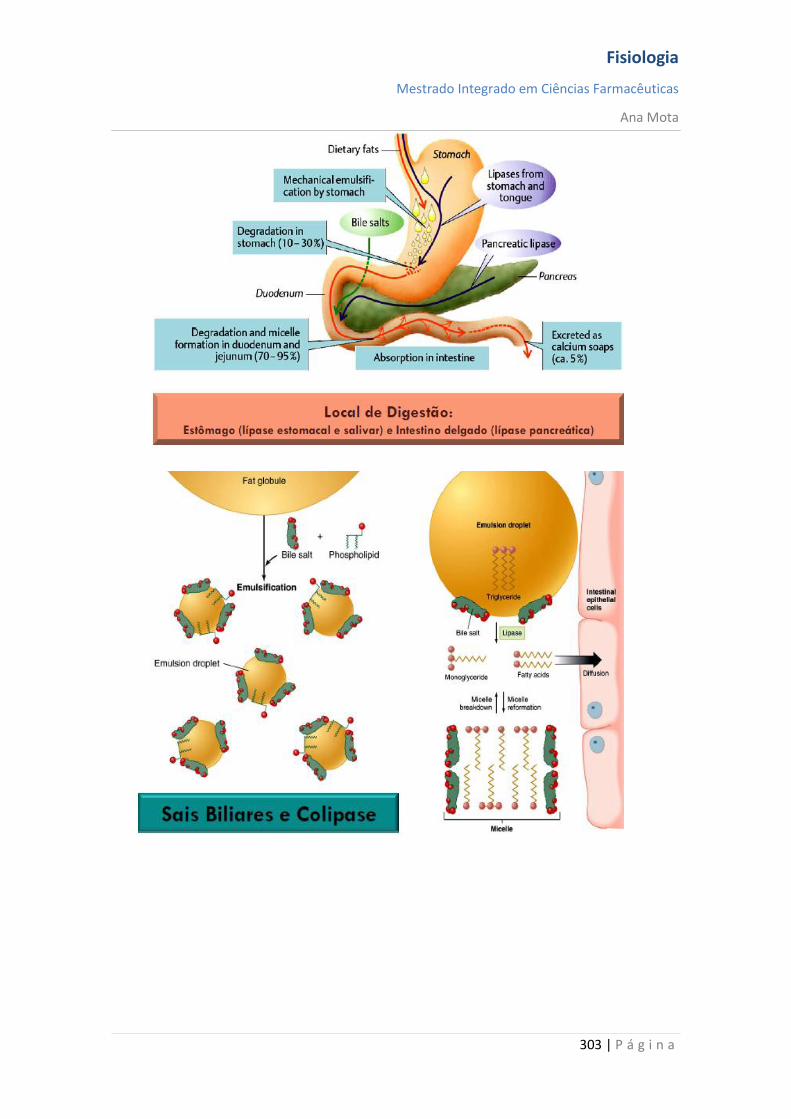
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
303 | P á g i n a
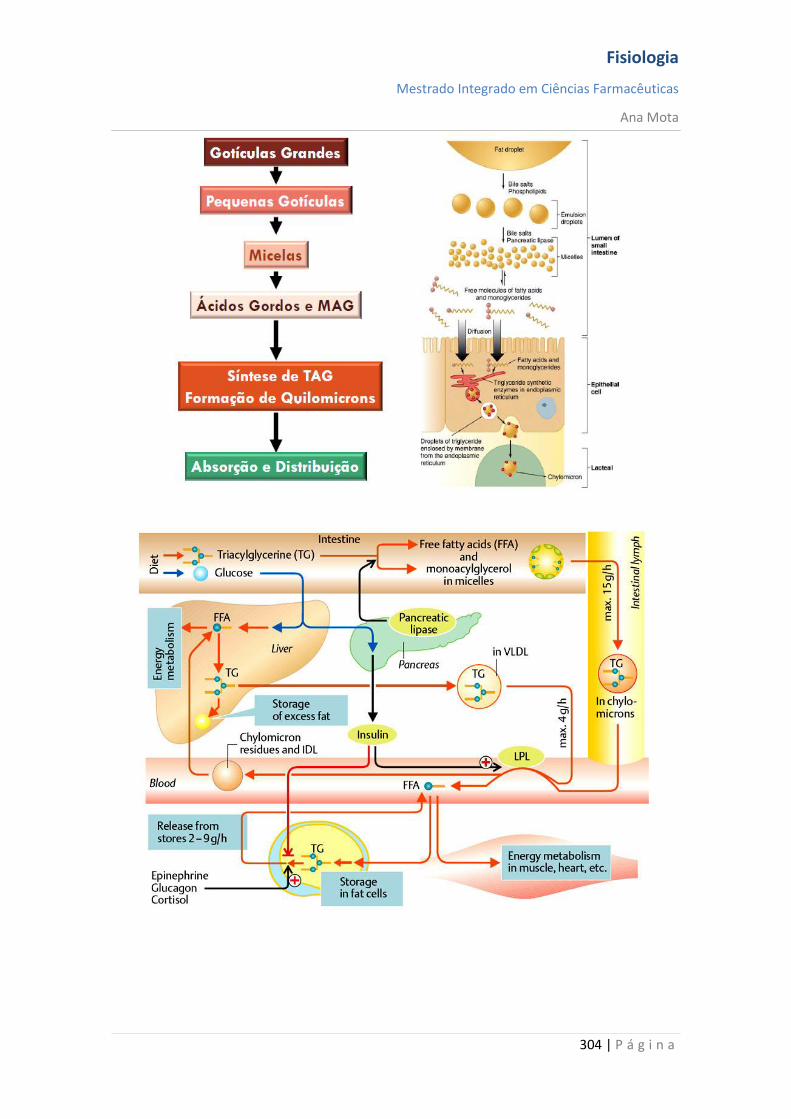
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
304 | P á g i n a

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
305 | P á g i n a
Lipoproteínas
PROTEÍNAS
Enzimas digestivas Locais de digestão Absorção de aminoácidos e péptidos
Peptidases
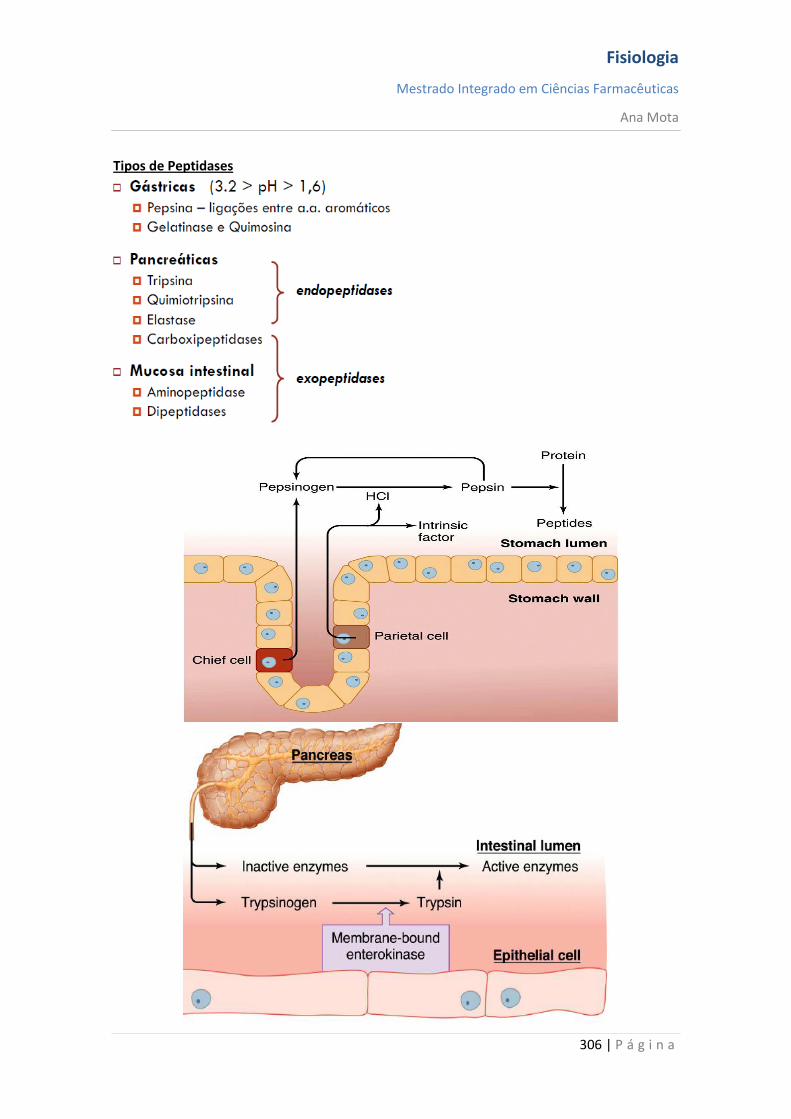
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
306 | P á g i n a
Tipos de Peptidases
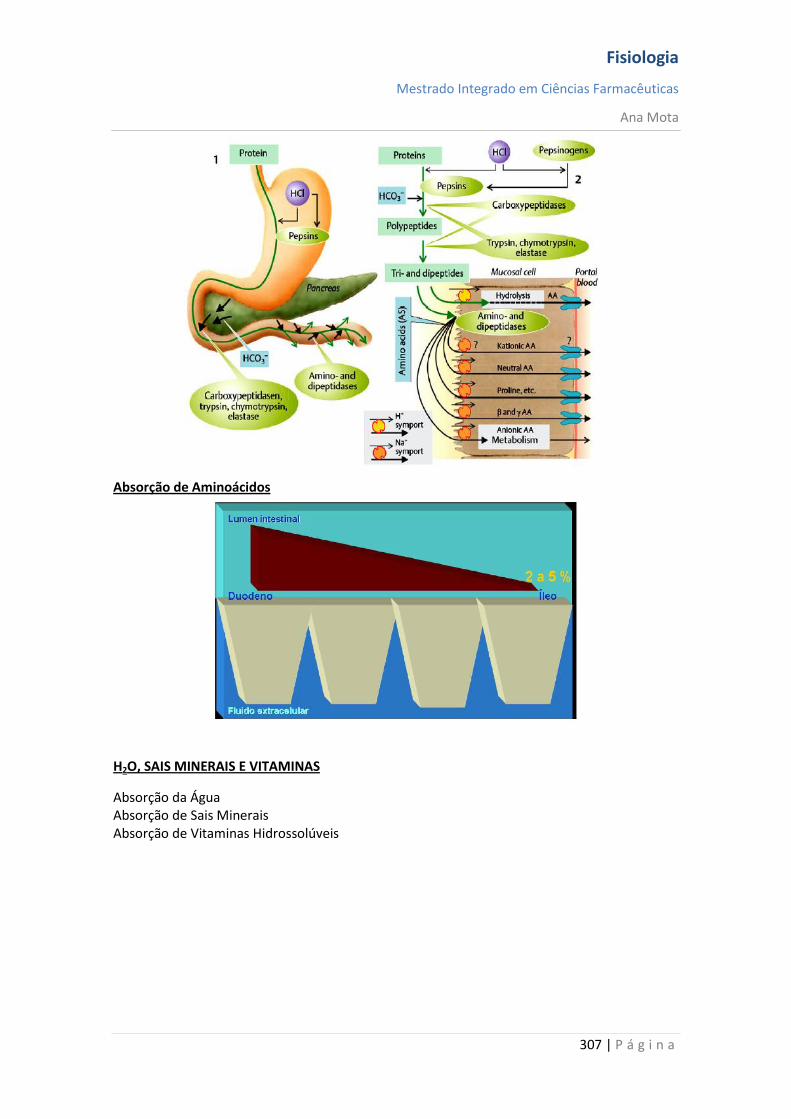
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
307 | P á g i n a
Absorção de Aminoácidos
H2O, SAIS MINERAIS E VITAMINAS
Absorção da Água Absorção de Sais Minerais Absorção de Vitaminas Hidrossolúveis
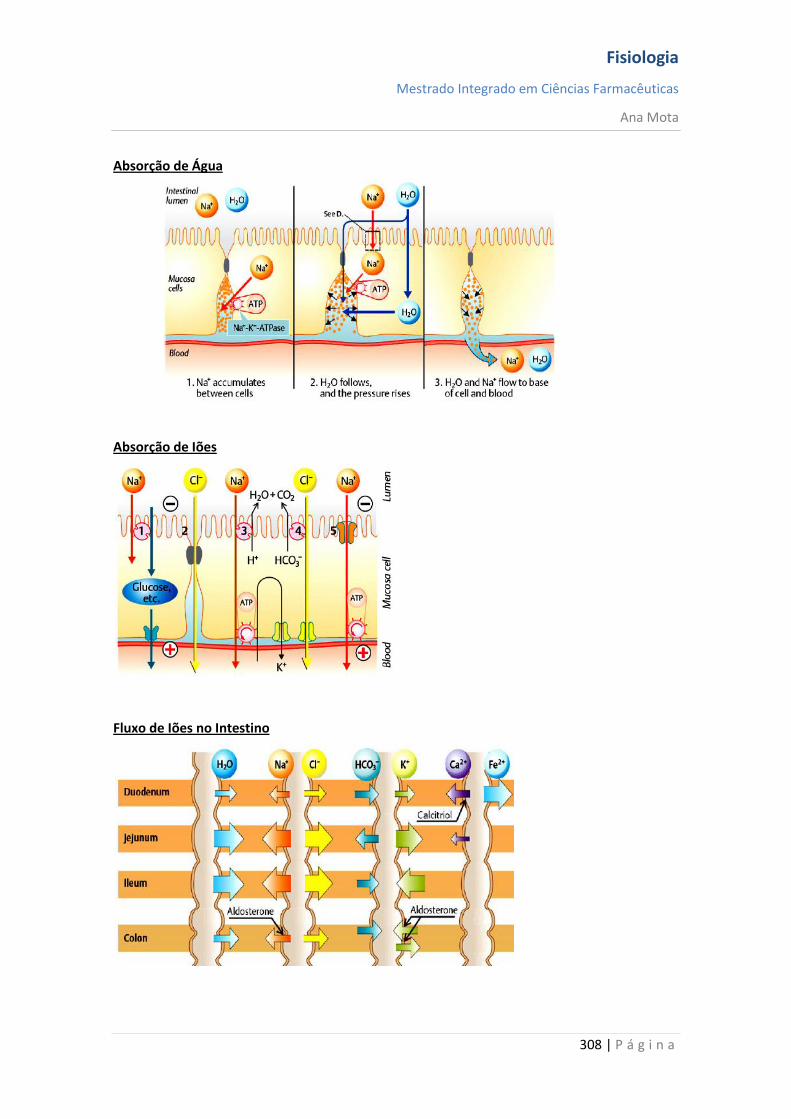
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
308 | P á g i n a
Absorção de Água
Absorção de Iões
Fluxo de Iões no Intestino
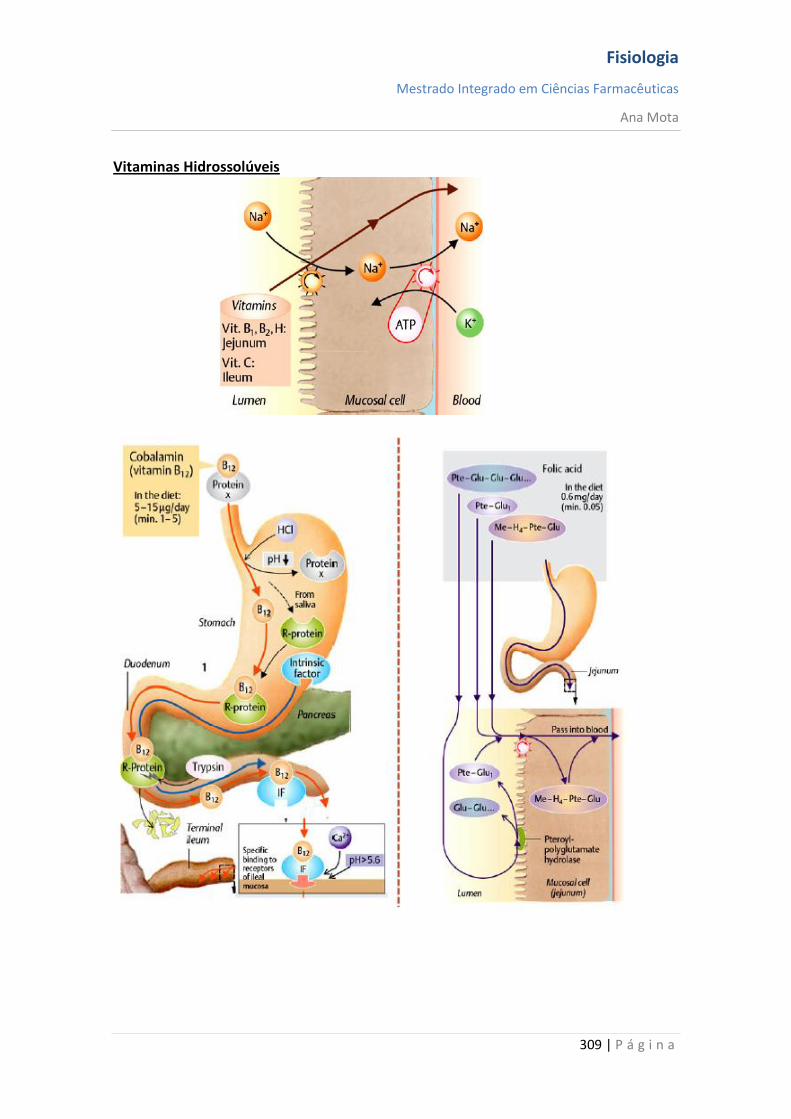
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
309 | P á g i n a
Vitaminas Hidrossolúveis
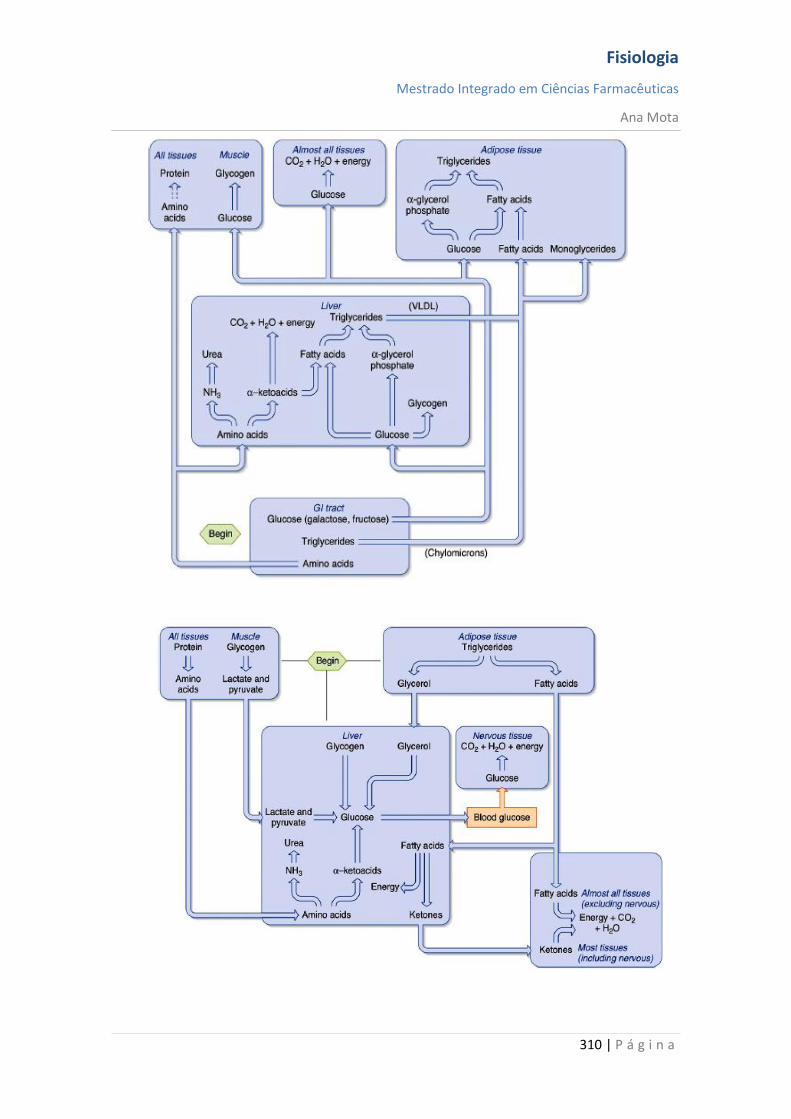
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
310 | P á g i n a
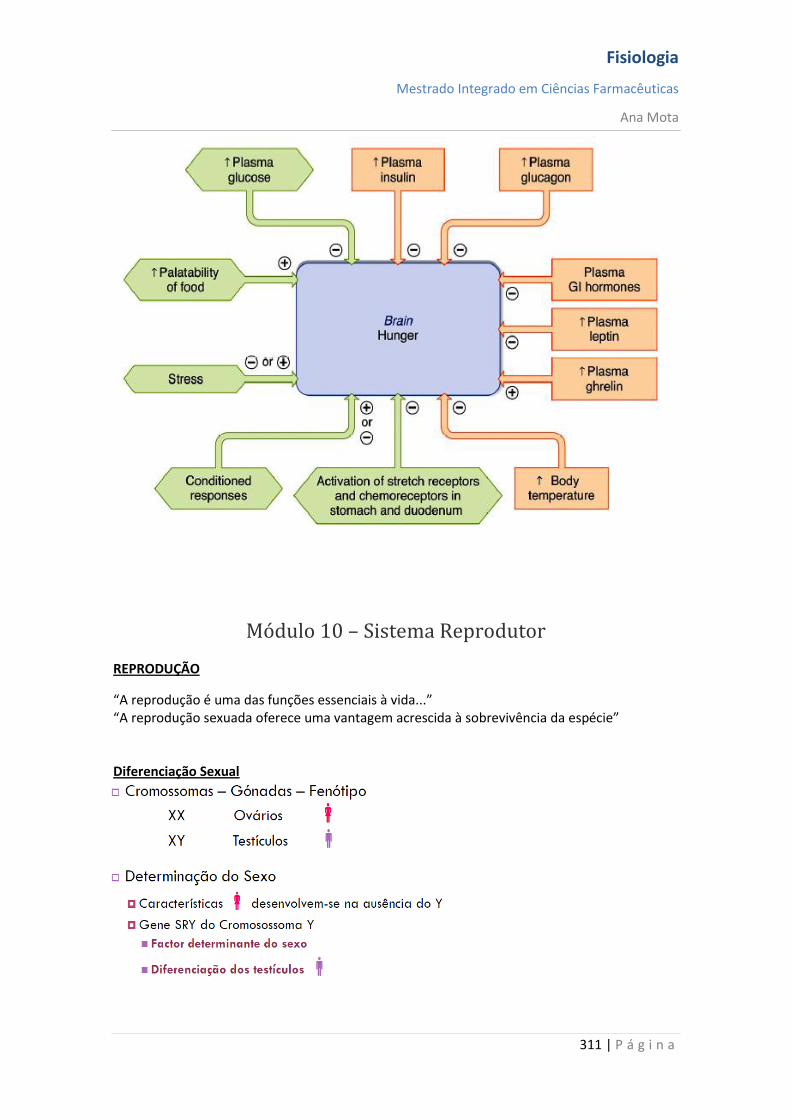
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
311 | P á g i n a
Módulo 10 – Sistema Reprodutor
REPRODUÇÃO
“A reprodução é uma das funções essenciais à vida...” “A reprodução sexuada oferece uma vantagem acrescida à sobrevivência da espécie”
Diferenciação Sexual

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
312 | P á g i n a
Hormonas Sexuais
Os testículos secretam diversas hormonas sexuais masculinos, colectivamente denominados androgénios, incluindo testosterona, di-hidrotestosterona e androstenediona. Todavia, a testosterona é muito mais abundante do que as demais hormonas, de modo que ela pode ser considerada como a hormona testicular fundamental, embora, grande parte, se não a maioria, é convertida na hormona mais activa, di-hidrotestosterona, nos tecidos-alvo.
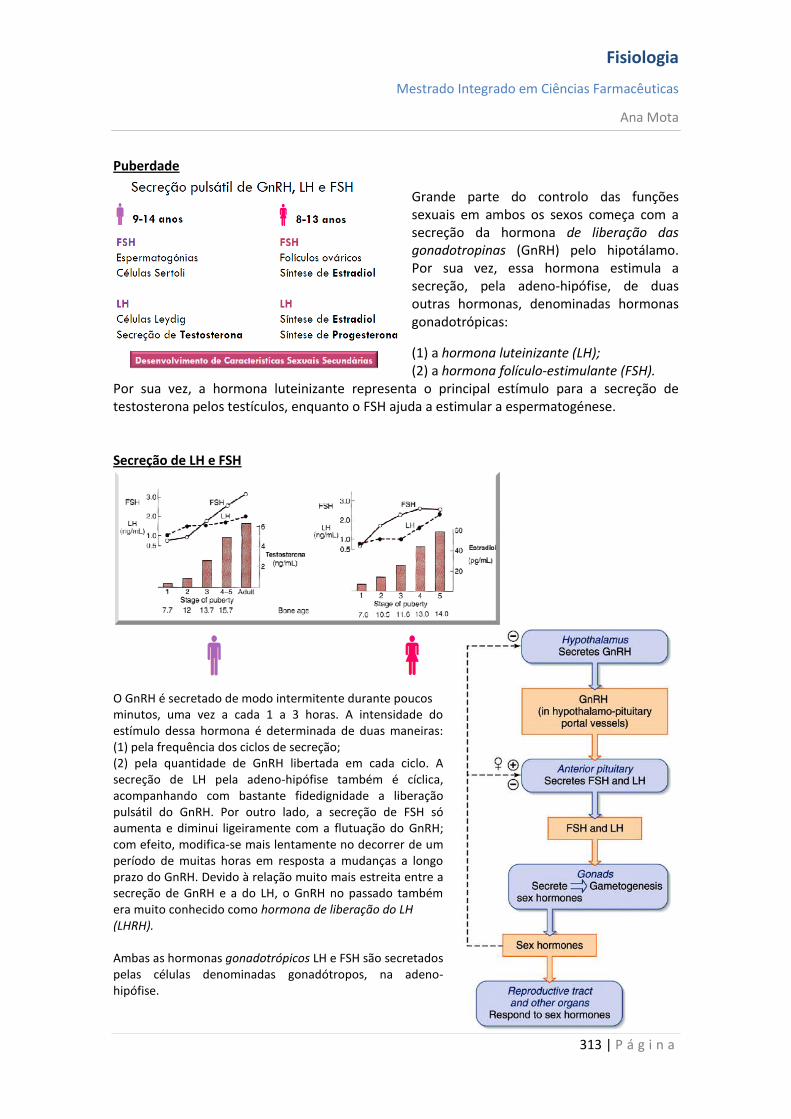
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
313 | P á g i n a
Puberdade
Grande parte do controlo das funções sexuais em ambos os sexos começa com a secreção da hormona de liberação das gonadotropinas (GnRH) pelo hipotálamo. Por sua vez, essa hormona estimula a secreção, pela adeno-hipófise, de duas outras hormonas, denominadas hormonas gonadotrópicas:
(1) a hormona luteinizante (LH); (2) a hormona folículo-estimulante (FSH).
Por sua vez, a hormona luteinizante representa o principal estímulo para a secreção de testosterona pelos testículos, enquanto o FSH ajuda a estimular a espermatogénese.
Secreção de LH e FSH
O GnRH é secretado de modo intermitente durante poucos minutos, uma vez a cada 1 a 3 horas. A intensidade do estímulo dessa hormona é determinada de duas maneiras: (1) pela frequência dos ciclos de secreção; (2) pela quantidade de GnRH libertada em cada ciclo. A secreção de LH pela adeno-hipófise também é cíclica, acompanhando com bastante fidedignidade a liberação pulsátil do GnRH. Por outro lado, a secreção de FSH só aumenta e diminui ligeiramente com a flutuação do GnRH; com efeito, modifica-se mais lentamente no decorrer de um período de muitas horas em resposta a mudanças a longo prazo do GnRH. Devido à relação muito mais estreita entre a secreção de GnRH e a do LH, o GnRH no passado também era muito conhecido como hormona de liberação do LH (LHRH). Ambas as hormonas gonadotrópicos LH e FSH são secretados pelas células denominadas gonadótropos, na adeno-hipófise.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
314 | P á g i n a
Na ausência total do GnRH do hipotálamo, os gonadótropos quase não secretam LH e FSH. O LH e o FSH são glicoproteínas; todavia, a quantidade de hidratos de carbono ligado à proteína nas moléculas varia de modo considerável em diferentes condições, o que pode modificar as potências de sua actividade. Tanto o LH quanto o FSH exercem seus efeitos sobre os tecidos-alvo dos testículos ao activar o sistema de segundo mensageiro de monofosfato de adenosina cíclico (AMPc), que, por sua vez, activa sistemas enzimáticos específicos nas respectivas células-alvo.
Características Sexuais 2as
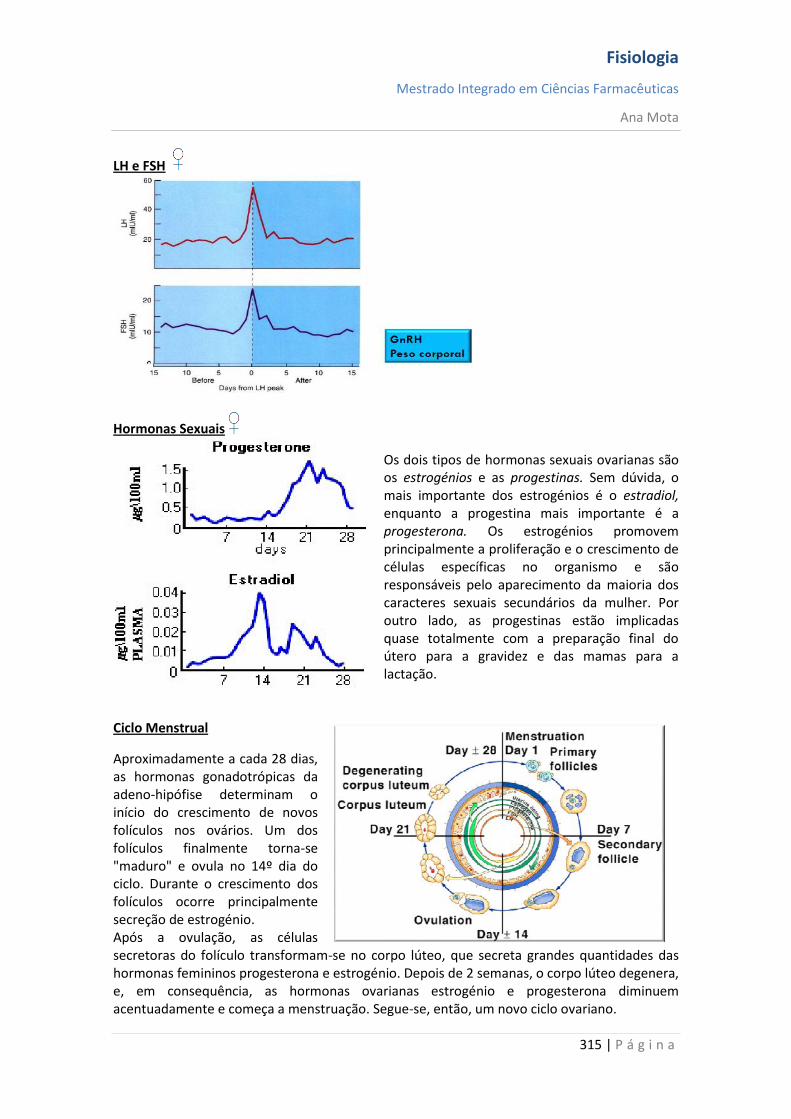
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
315 | P á g i n a
LH e FSH
Hormonas Sexuais
Os dois tipos de hormonas sexuais ovarianas são os estrogénios e as progestinas. Sem dúvida, o mais importante dos estrogénios é o estradiol, enquanto a progestina mais importante é a progesterona. Os estrogénios promovem principalmente a proliferação e o crescimento de células específicas no organismo e são responsáveis pelo aparecimento da maioria dos caracteres sexuais secundários da mulher. Por outro lado, as progestinas estão implicadas quase totalmente com a preparação final do útero para a gravidez e das mamas para a lactação.
Ciclo Menstrual
Aproximadamente a cada 28 dias, as hormonas gonadotrópicas da adeno-hipófise determinam o início do crescimento de novos folículos nos ovários. Um dos folículos finalmente torna-se "maduro" e ovula no 14º dia do ciclo. Durante o crescimento dos folículos ocorre principalmente secreção de estrogénio. Após a ovulação, as células secretoras do folículo transformam-se no corpo lúteo, que secreta grandes quantidades das hormonas femininos progesterona e estrogénio. Depois de 2 semanas, o corpo lúteo degenera, e, em consequência, as hormonas ovarianas estrogénio e progesterona diminuem acentuadamente e começa a menstruação. Segue-se, então, um novo ciclo ovariano.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
316 | P á g i n a
Oogénese
Entre os 40 e 50 anos de idade, os ciclos sexuais geralmente tornam-se irregulares e a ovulação deixa de ocorrer durante muitos desses ciclos. Depois de poucos meses e alguns anos, os ciclos cessam por completo. Esse período, durante o qual cessam os ciclos e as hormonas sexuais femininos diminuem rapidamente até quase zero, é denominado menopausa. A causa da menopausa é a "exaustão" dos ovários. Durante toda a vida reprodutiva da mulher, cerca de 400 folículos primordiais desenvolvem-se em folículos vesiculares e ovulam, enquanto literalmente centenas de milhares de óvulos degeneram. Com cerca de 45 anos de idade, poucos folículos primordiais ainda são estimulados pelo FSH e LH, e a produção de estrogénios pelos ovários diminui, à medida que o número de folículos primordiais se aproxima de zero. Quando a produção de estrogénios cai abaixo de um nível crítico, os estrogénios não mais conseguem inibir a produção de FSH e de LH; tampouco são capazes de ocasionar um surto ovulatório de LH e de FSH para produzir os ciclos ovulatórios. Na verdade, o FSH e o LH (principalmente o primeiro) são, então, produzidos em quantidades grandes e contínuas. Os estrogénios são produzidos em quantidades suberíticas durante curto período de tempo após a menopausa; todavia, no decorrer de alguns anos, à medida que os últimos folículos primordiais tornam-se atrésicos, a produção de estrogénios pelos ovários cai e atinge quase zero. Na época da menopausa, a mulher precisa reajustar sua vida, passando de uma vida fisiologicamente estimulada pela produção de estrogénios e progesterona para uma fase destituída dessas hormonas. Com frequência, a perda dos estrogénios provoca alterações fisiológicas acentuadas na função do organismo. Esses sintomas são de magnitude suficiente para exigir tratamento em cerca de 15% das mulheres. Se o aconselhamento não tiver êxito, a administração diária de um estrogénios em pequenas quantidades irá reverter os sintomas, e, ao reduzir gradualmente a dose, a mulher pós-menopáusica poderá evitar a ocorrência de sintomas graves. Infelizmente, esse tipo de tratamento pode prolongar os sintomas.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
317 | P á g i n a
Testosterona
Em geral, a testosterona é responsável pelas características peculiares do corpo masculino. Mesmo durante a vida fetal, os testículos são estimulados peta gonadotropina coriónica proveniente da placenta, produzindo quantidades moderadas de testosterona durante todo o desenvolvimento fetal e durante três ou mais semanas após o nascimento. Depois desse período, não há praticamente qualquer produção de testosterona durante a meninice até aproximadamente a idade de 10 a 13 anos. A partir aí, a produção de testosterona aumenta
rapidamente pelo estímulo das hormonas gonadotrópicas da adeno-hipófise no início da puberdade e persiste durante a maior parte da vida, diminuindo rapidamente depois dos 50 anos de idade, atingindo, aos 80 anos, um nível que corresponde a 20 a 50% do valor máximo.
SISTEMA REPRODUTOR
Espermatogénese e transporte de esperma;
Erecção e ejaculação de sémen.
O testículo é composto de até 900 tubos seminíferos enovelados, cada um com mais de meio metro de comprimento em média, onde são formados os espermatozóides. A seguir, o esperma desemboca no epidídimo, outro tubo enovelado com aproximadamente 6 metros de comprimento.
O epidídimo leva ao canal deferente, que se alarga para formar a ampola do canal deferente, imediatamente antes de sua penetração na glândula prostática. As vesículas seminais, cada uma localizada num lado da próstata, esvaziam-se na extremidade prostática da ampola, e o conteúdo da ampola e da vesícula seminal passam para o ducto ejaculador, que segue através do corpo da glândula prostática para desaguar na uretra interna. Os ductos prostáticos esvaziam-se, por sua vez, no ducto ejaculador. Por fim, a uretra é a última ligação entre o testículo e o exterior. A uretra recebe muco derivado de grande número de pequenas glândulas uretrais localizadas em toda sua extensão, bem como das glândulas bulbouretrais bilaterais (glândulas de Cowper), situadas próximo à origem da uretra.
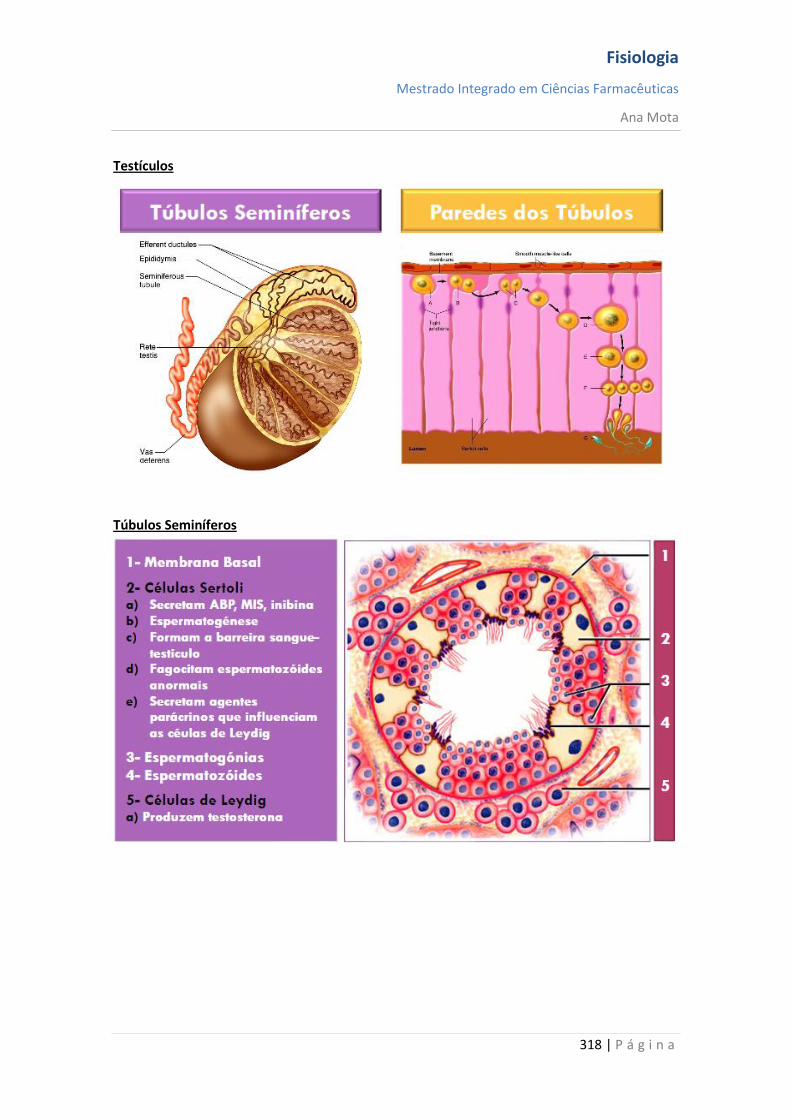
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
318 | P á g i n a
Testículos
Túbulos Seminíferos

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
319 | P á g i n a
Espermatogénese
1 ciclo = 65-74 Dias
Durante um período de 24 dias, em média, cada espermatogónia que atravessa a barreira na camada de células de Sertoli modifica-se progressivamente e aumenta, formando um espermatócito primário muito grande. Ao cabo de 24 dias, cada um desses espermatócitos primários divide-se para formar dois espermatócitos secundários. Todavia, essa divisão não é normal. Na verdade, é denominada primeira divisão meiótica. Na fase inicial dessa divisão, todo o DNA dos 46 cromossomas é replicado. Nesse processo, cada um dos 46 cromossomas transformam-se em dois cromátideos, que permanecem ligados entre si nos centrómeros, e cada um dos dois cromátideos tem os genes duplicados daquele
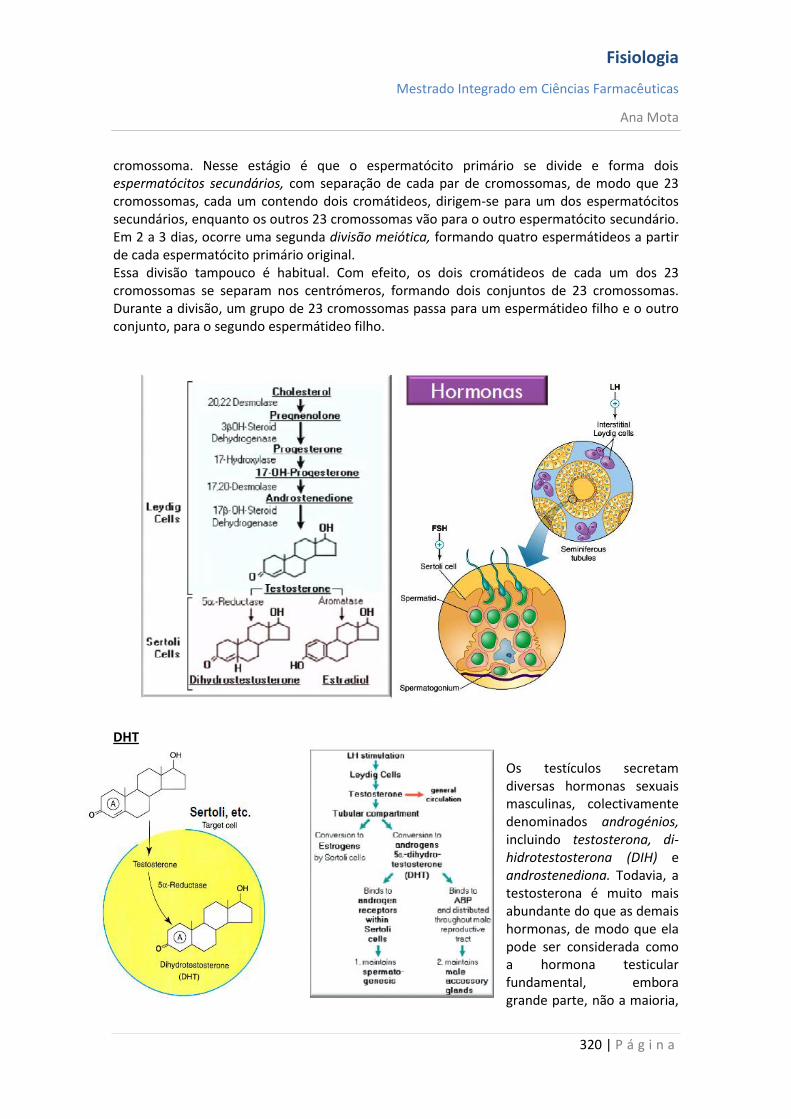
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
320 | P á g i n a
cromossoma. Nesse estágio é que o espermatócito primário se divide e forma dois espermatócitos secundários, com separação de cada par de cromossomas, de modo que 23 cromossomas, cada um contendo dois cromátideos, dirigem-se para um dos espermatócitos secundários, enquanto os outros 23 cromossomas vão para o outro espermatócito secundário. Em 2 a 3 dias, ocorre uma segunda divisão meiótica, formando quatro espermátideos a partir de cada espermatócito primário original. Essa divisão tampouco é habitual. Com efeito, os dois cromátideos de cada um dos 23 cromossomas se separam nos centrómeros, formando dois conjuntos de 23 cromossomas. Durante a divisão, um grupo de 23 cromossomas passa para um espermátideo filho e o outro conjunto, para o segundo espermátideo filho.
DHT
Os testículos secretam diversas hormonas sexuais masculinas, colectivamente denominados androgénios, incluindo testosterona, di-hidrotestosterona (DIH) e androstenediona. Todavia, a testosterona é muito mais abundante do que as demais hormonas, de modo que ela pode ser considerada como a hormona testicular fundamental, embora grande parte, não a maioria,
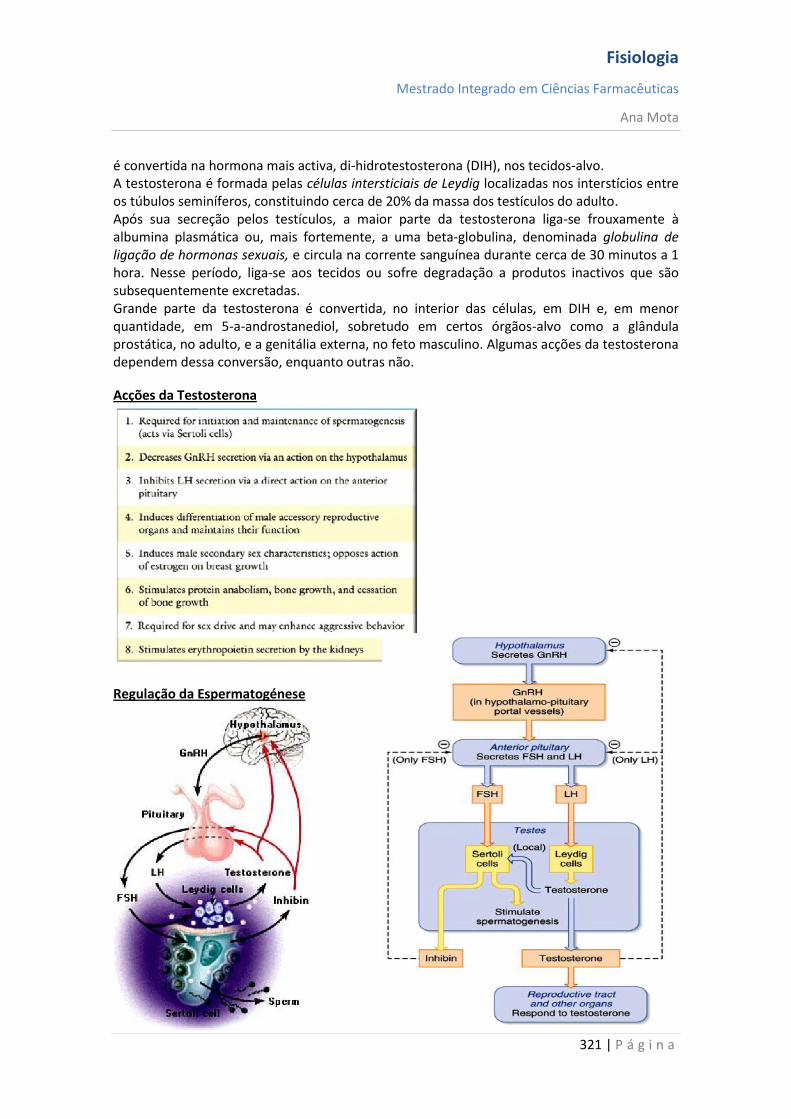
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
321 | P á g i n a
é convertida na hormona mais activa, di-hidrotestosterona (DIH), nos tecidos-alvo. A testosterona é formada pelas células intersticiais de Leydig localizadas nos interstícios entre os túbulos seminíferos, constituindo cerca de 20% da massa dos testículos do adulto. Após sua secreção pelos testículos, a maior parte da testosterona liga-se frouxamente à albumina plasmática ou, mais fortemente, a uma beta-globulina, denominada globulina de ligação de hormonas sexuais, e circula na corrente sanguínea durante cerca de 30 minutos a 1 hora. Nesse período, liga-se aos tecidos ou sofre degradação a produtos inactivos que são subsequentemente excretadas. Grande parte da testosterona é convertida, no interior das células, em DIH e, em menor quantidade, em 5-a-androstanediol, sobretudo em certos órgãos-alvo como a glândula prostática, no adulto, e a genitália externa, no feto masculino. Algumas acções da testosterona dependem dessa conversão, enquanto outras não.
Acções da Testosterona
Regulação da Espermatogénese

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
322 | P á g i n a
A testosterona secretada pelos testículos em resposta ao LH possui o efeito recíproco de impedir a secreção adeno-hipofisária de LH. Exerce esse efeito de duas maneiras: 1. A maior parte da inibição resulta do efeito directo da testosterona sobre o hipotálamo, no sentido de diminuir a secreção de GnRH. Por sua vez, a secreção diminuída de GnRH determina a redução correspondente na secreção adeno-hipofisária de LH e FSH, e a diminuição do LH reduz a secreção de testosterona pelos testículos. Por conseguinte, toda vez que a secreção de testosterona for muito grande, esse efeito automático de feedback negativo, ao operar através do hipotálamo e da adeno-hipófise, reduz a secreção de testosterona até seu nível funcional normal. Por outro lado, a presença de testosterona em quantidades muito pequenas induz o hipotálamo a secretar grandes quantidades de GnRH, com elevação correspondente na secreção adeno-hipofisária de LH e FSH e aumento da secreção de testosterona pelos testículos; 2. É provável que a testosterona também tenha um fraco efeito de feedback negativo actuando directamente sobre a adenohipófise no sentido de diminuir a secreção de LH. Consequentemente, acredita-se que também ocorra um pequeno grau de regulação da ligação de testosterona por esse mecanismo. O FSH liga-se a receptores específicos para essa hormona fixados às células de Sertoli nos túbulos seminíferos. Essa ligação induz o crescimento dessas células e a secreção de várias substâncias espermatogénicas por elas. Simultaneamente, a testosterona que se difunde nos túbulos a partir das células de Leydig nos espaços intersticiais também exerce forte efeito trópico sobre a espermatogénese. Para iniciar a espermatogénese, é necessária a presença de FSH e de testosterona, embora, uma vez ocorrida a estimulação inicial, a testosterona por si só possa manter a espermatogénese por longo tempo.
Erecção e Ejaculação

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
323 | P á g i n a
Sémen
Fertilização
SISTEMA REPRODUTOR
Feminino
Oogénse e Ciclo Menstrual.
Regulação do Ciclo
Menstrual. Fertilização.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
324 | P á g i n a
Os principais órgãos do aparelho reprodutor feminino no ser humano, incluindo, entre os mais importantes, os ovários, as trompas de Falópio, o útero e a vagina. A reprodução começa com o desenvolvimento dos óvulos nos ovários. No meio de cada ciclo sexual mensal, apenas um óvulo é expelido de um folículo ovariano, na cavidade abdominal, a seguir, esse óvulo passa através de uma das trompas de Falópio e penetra no útero - caso seja fertilizado por um espermatozóide, implanta-se no útero, onde se transforma em feto, placenta e membranas fetais.
Ciclo Menstrual
Aproximadamente 2 dias antes do final do ciclo mensal, a secreção das hormonas ovarianas, estrogénios e progesterona diminui acentuadamente e atinge baixos níveis, seguindo-se, então, a menstruação. A menstruação é causada pela súbita redução dos estrogénios a da progesterona, em particular desta última, ao final do ciclo ovariano mensal. O primeiro efeito observado consiste numa redução da estimulação das células endometriais por essas duas hormonas, seguida rapidamente pela involução do próprio endométrio até cerca de 65% de sua espessura anterior. Durante as 24 horas que precedem o início da menstruação, os vasos sanguíneos sinuosos que irrigam a camada mucosa do endométrio tornam-se vasoespásticos, presumivelmente devido a algum efeito da involução, como a liberação de material vasoconstritor— possivelmente um dos tipos vasoconstritores de prostaglandinas presentes em quantidades abundantes nessa fase. O vasoespasmo e a perda do estímulo hormonal provocam a necrose inicial do endométrio, em particular dos vasos sanguíneos situados no estrato vascular. Em consequência, o sangue exsuda para a camada vascular do endométrio, e as áreas hemorrágicas aumentam no decorrer de um período de 24 a 36 horas. Gradualmente, as camadas externas necróticas do endométrio descolam-se do útero no local das hemorragias, até que, aproximadamente 48 horas após o início da menstruação, todas as camadas superficiais do endométrio já sofreram descamação. A massa do tecido descamado e o sangue na cavidade uterina, possivelmente em associação com os efeitos contrateis das prostaglandinas, desencadeiam as contracções uterinas que expelem seu conteúdo. Durante a menstruação normal, ocorre perda aproximada de 40 ml de sangue e 35 ml de líquido seroso. Esse líquido menstrual normalmente não coagula, devido à liberação de fibrinolisina juntamente com o material endometrial necrótico. Todavia, se houver sangramento excessivo da superfície uterina, a quantidade de fibrinolisina pode não ser suficiente para impedir a coagulação. A presença de coágulos durante a menstruação constitui-se, em geral, numa prova clínica de patologia uterina. Dentro de 4 a 7 dias após o início da menstruação, a perda de sangue cessa, visto que, nessa época, o endométrio já está totalmente reepitelizado.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
325 | P á g i n a
Ovários e Folículos
Ao nascimento, cada óvulo, na criança do sexo feminino, é circundado por camada única de células da granulosa, e o óvulo, com sua bainha de células da granulosa, é denominado folículo primordial, conforme ilustrado na figura. Durante toda a infância, acredita-se que as células da granulosa forneçam a nutrição necessária para o óvulo, além de secretar um factor inibidora maturação dos ovócitos, que mantém o óvulo em seu estado primordial, detendo-o na profase da divisão meiótica por todo esse período de tempo. A seguir, após a puberdade, quando o FSH e o LH da adeno-hipófise começam a ser secretados em grandes quantidades, todo o ovário, juntamente com alguns de seus folículos, começa a crescer. A primeira fase do crescimento folicular consiste em aumento moderado do próprio óvulo, cujo diâmetro aumenta por duas a três vezes. Segue-se, então, o crescimento de camadas adicionais de células da granulosa, e, nesse estágio, o folículo passa a ser conhecido como folículo primário. Parte do processo de desenvolvimento até esse estágio pode ocorrer na ausência de FSH e de LH, mas o desenvolvimento posterior não é possível sem a presença dessas duas hormonas. Oogénese
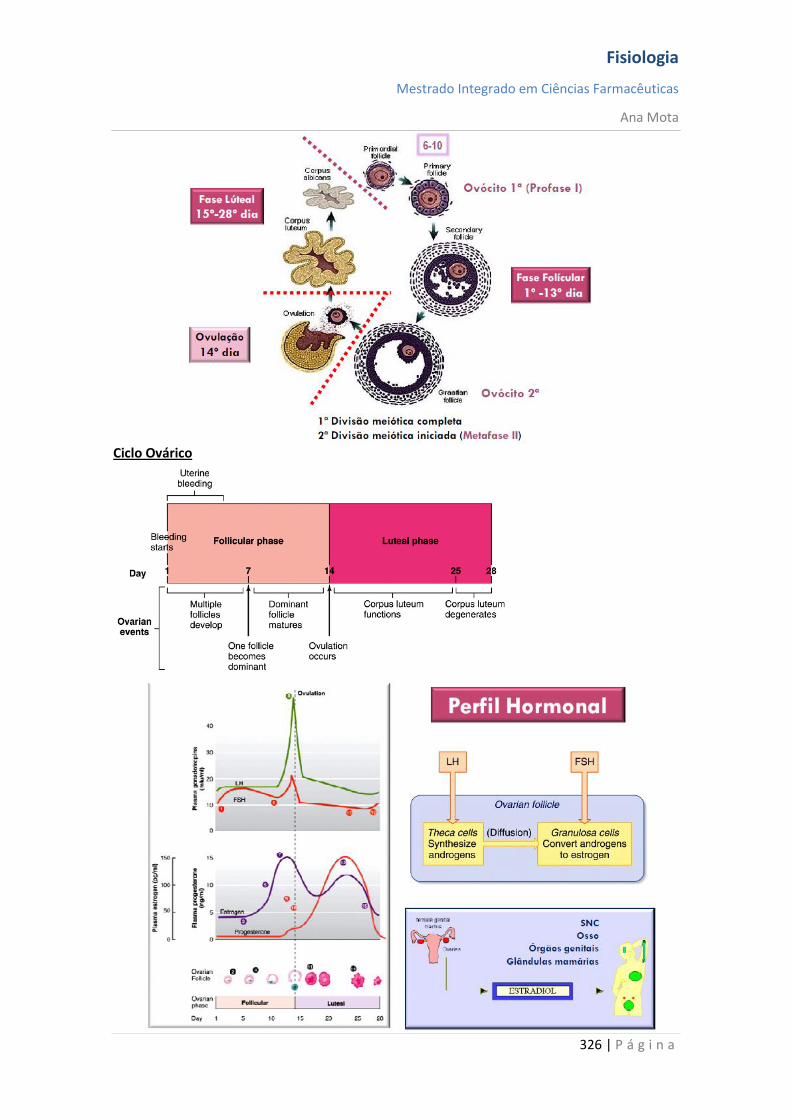
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
326 | P á g i n a
Ciclo Ovárico
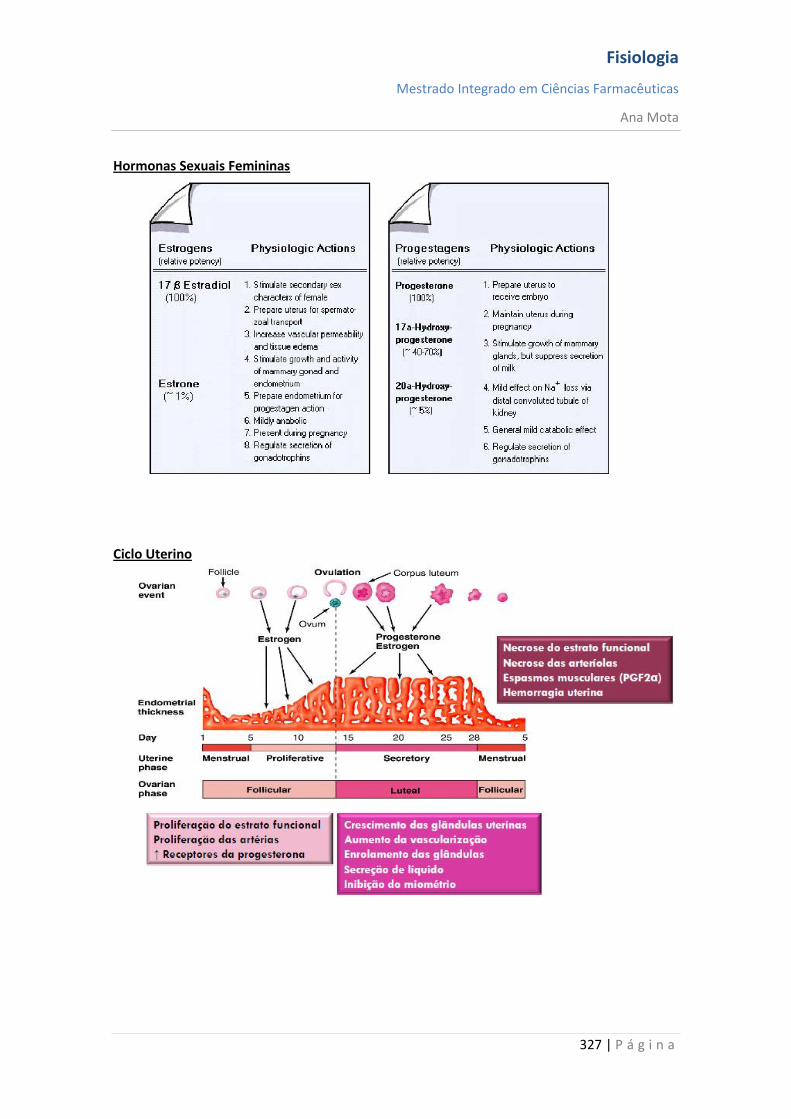
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
327 | P á g i n a
Hormonas Sexuais Femininas
Ciclo Uterino
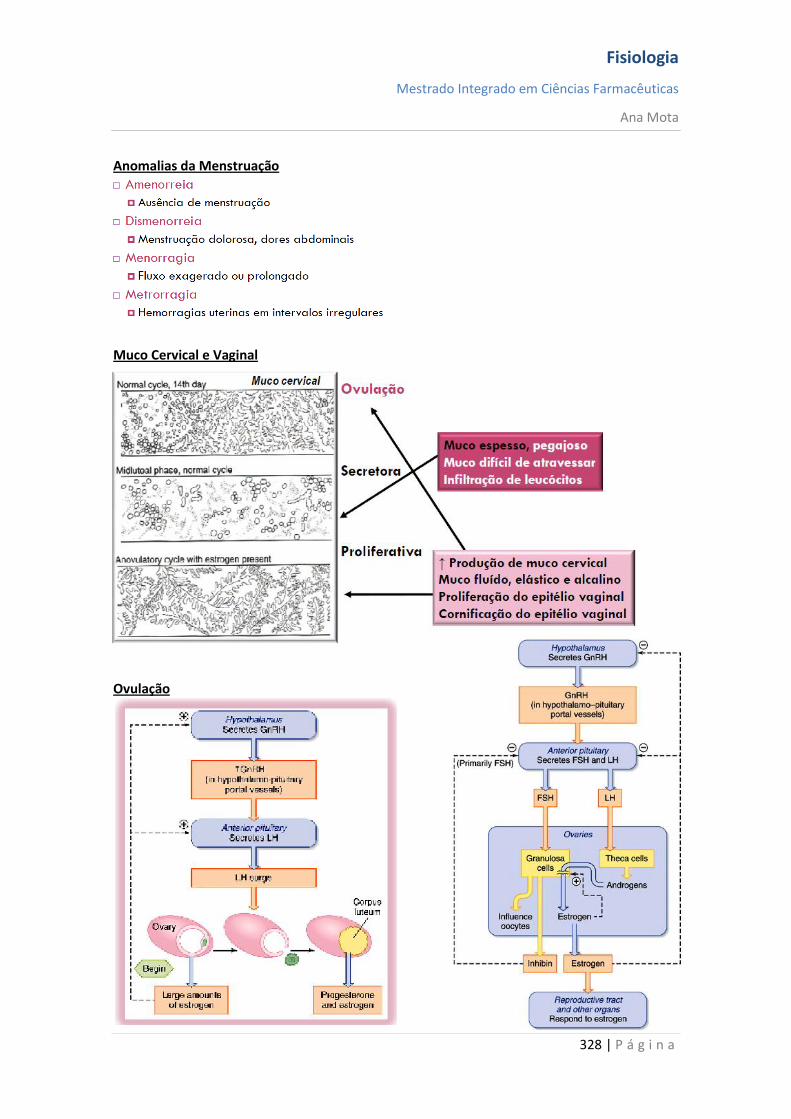
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
328 | P á g i n a
Anomalias da Menstruação
Muco Cervical e Vaginal
Ovulação
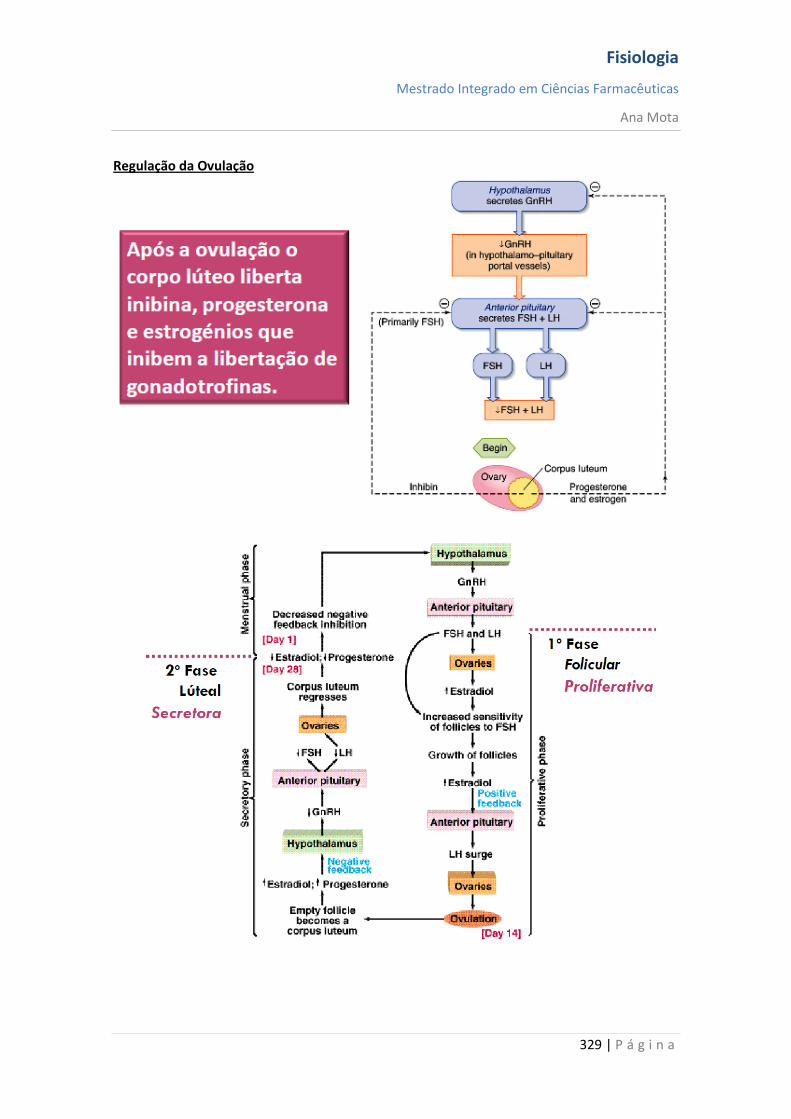
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
329 | P á g i n a
Regulação da Ovulação
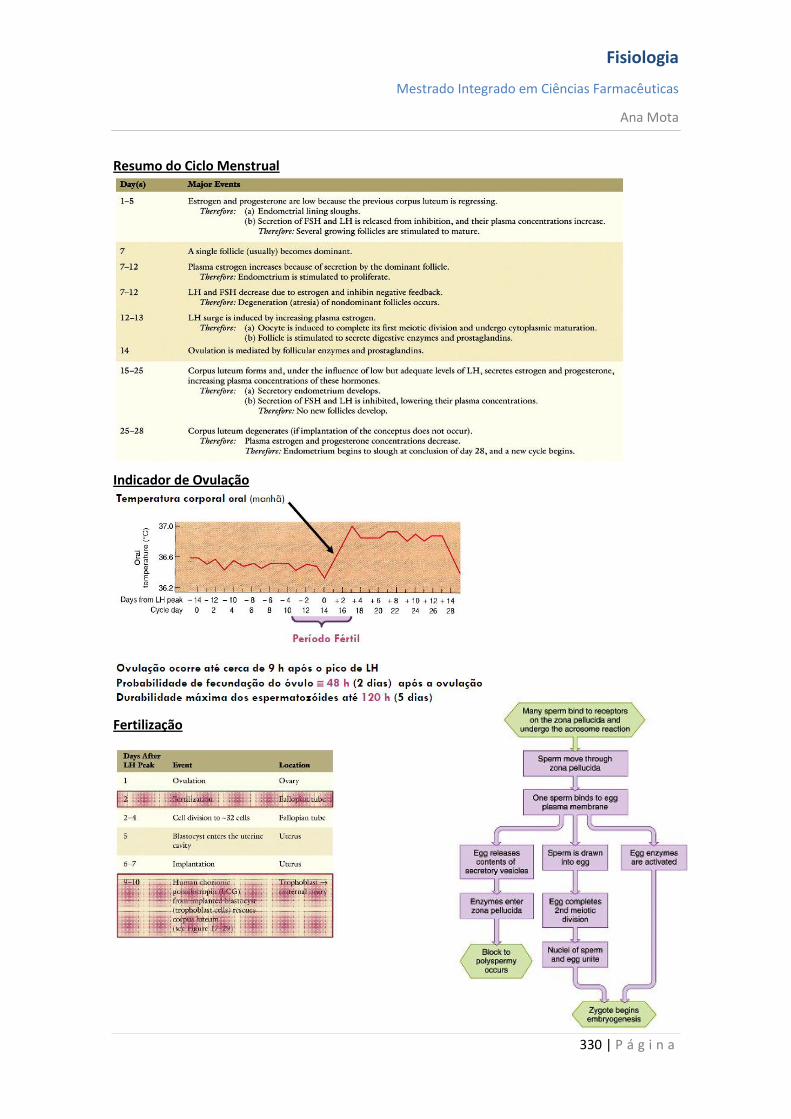
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
330 | P á g i n a
Resumo do Ciclo Menstrual
Indicador de Ovulação
Fertilização
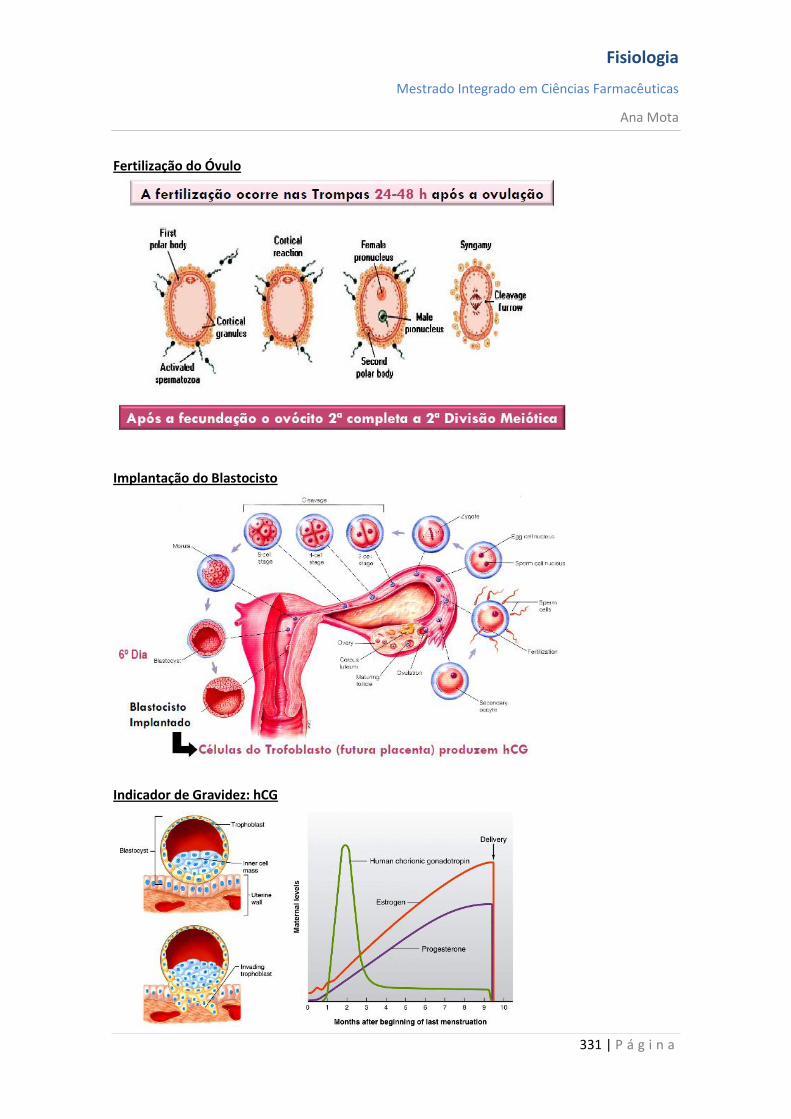
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
331 | P á g i n a
Fertilização do Óvulo
Implantação do Blastocisto
Indicador de Gravidez: hCG
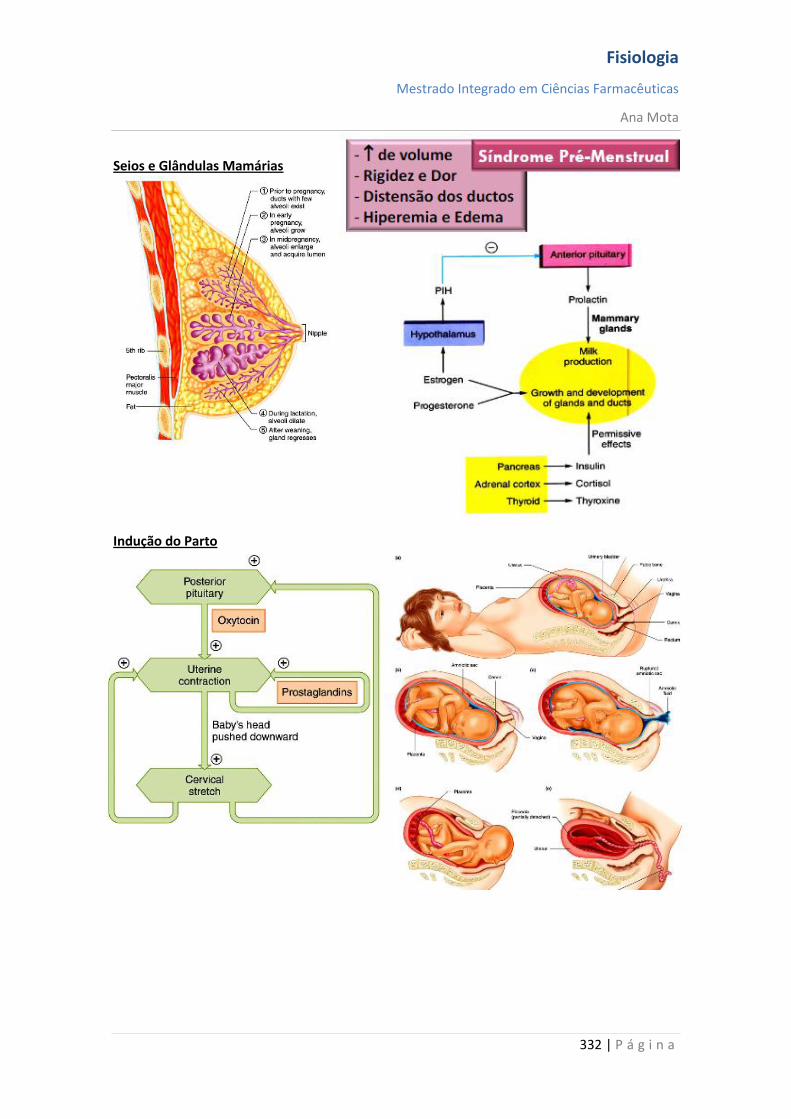
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
332 | P á g i n a
Seios e Glândulas Mamárias
Indução do Parto
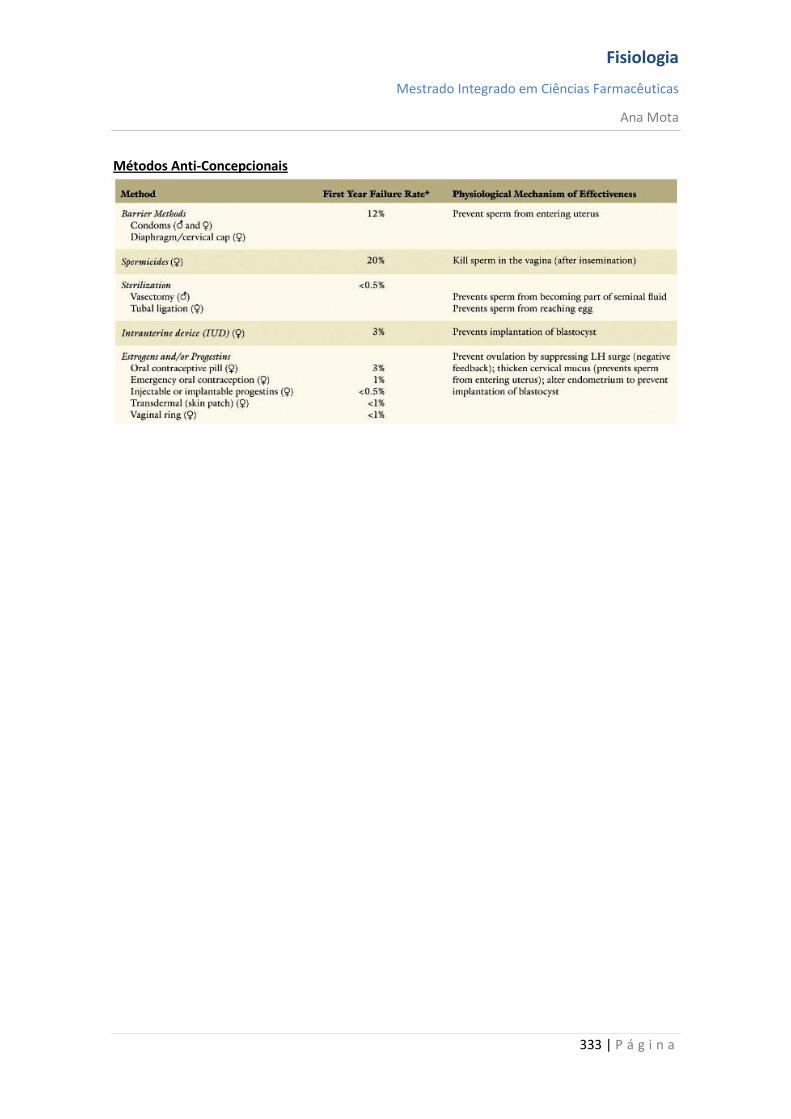
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
333 | P á g i n a
Métodos Anti-Concepcionais
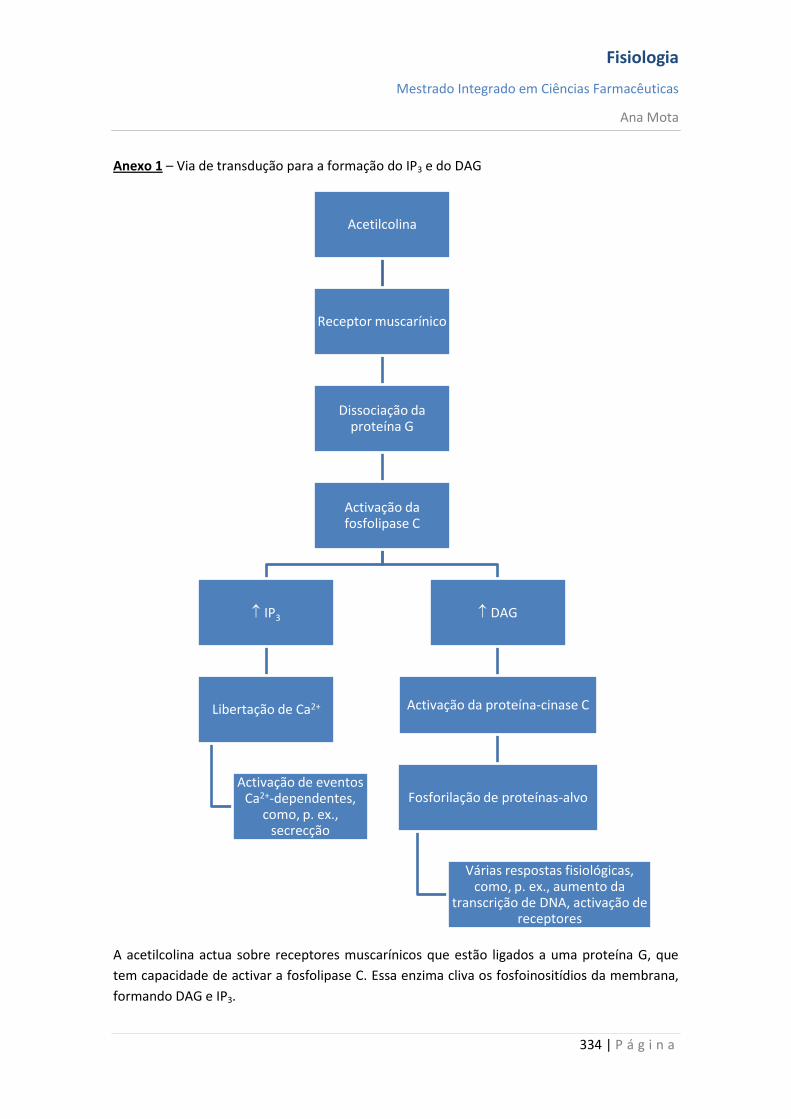
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
334 | P á g i n a
Anexo 1 – Via de transdução para a formação do IP3 e do DAG
A acetilcolina actua sobre receptores muscarínicos que estão ligados a uma proteína G, que
tem capacidade de activar a fosfolipase C. Essa enzima cliva os fosfoinositídios da membrana,
formando DAG e IP3.
Acetilcolina
Receptor muscarínico
Dissociação da proteína G
Activação da fosfolipase C
IP3
Libertação de Ca2+
Activação de eventos Ca2+-dependentes,
como, p. ex., secrecção
DAG
Activação da proteína-cinase C
Fosforilação de proteínas-alvo
Várias respostas fisiológicas, como, p. ex., aumento da
transcrição de DNA, activação de receptores

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
335 | P á g i n a
Anexo 2 – Vias que levam à síntese de prostaglandinas e de leucotrienos, a partir dos
fosfolípidicos da membrana
Anexo 3 - Um feedback positivo abre os canais de sódio Primeiro, enquanto a membrana da fibra nervosa permanece sem sofrer qualquer perturbação, nenhum potencial de acção ocorre na fibra nervosa normal. Contudo, se algum factor produzir uma elevação inicial suficiente do potencial de membrana, a partir do valor de -90 mV, em direcção ao zero, essa elevação da voltagem irá fazer com que muitos canais de sódio dependentes da voltagem comecem a abrir-se. Isso permite o influxo rápido de iões sódio, o que produz um maior aumento do potencial de membrana e abertura de um número maior de canais de sódio dependentes da voltagem e resultando num influxo de iões sódio para o interior da fibra. Obviamente, esse processo é um ciclo vicioso de feedback positivo que, caso esse feedback seja suficientemente intenso, irá prosseguir até que todos os canais de sódio dependentes da voltagem fiquem activos (abertos). Em seguida, dentro de fracção de milissegundo, a elevação do potencial de membrana produz o início da inactivação dos canais de sódio, além da abertura dos canais de potássio, e o potencial de acção logo chega ao fim.
Fosfolipídico da membrana
Ácido araquidónico
Ciclooxigenase
Prostaglandinas, tromboxanos, p.
ex., PGE2
Lipoxigenase
Leucotrienos, p. ex., LTC4, LTB4
Fosfolipases A, C e D
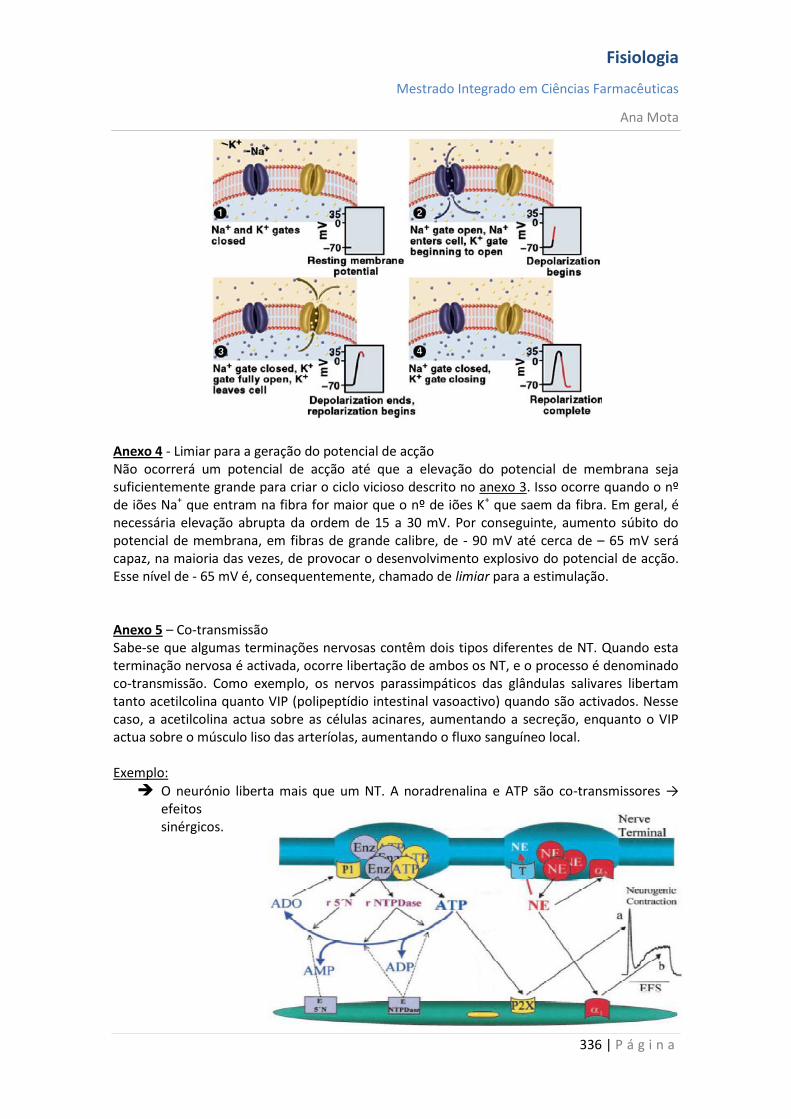
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
336 | P á g i n a
Anexo 4 - Limiar para a geração do potencial de acção Não ocorrerá um potencial de acção até que a elevação do potencial de membrana seja suficientemente grande para criar o ciclo vicioso descrito no anexo 3. Isso ocorre quando o nº de iões Na+ que entram na fibra for maior que o nº de iões K+ que saem da fibra. Em geral, é necessária elevação abrupta da ordem de 15 a 30 mV. Por conseguinte, aumento súbito do potencial de membrana, em fibras de grande calibre, de - 90 mV até cerca de – 65 mV será capaz, na maioria das vezes, de provocar o desenvolvimento explosivo do potencial de acção. Esse nível de - 65 mV é, consequentemente, chamado de limiar para a estimulação. Anexo 5 – Co-transmissão Sabe-se que algumas terminações nervosas contêm dois tipos diferentes de NT. Quando esta terminação nervosa é activada, ocorre libertação de ambos os NT, e o processo é denominado co-transmissão. Como exemplo, os nervos parassimpáticos das glândulas salivares libertam tanto acetilcolina quanto VIP (polipeptídio intestinal vasoactivo) quando são activados. Nesse caso, a acetilcolina actua sobre as células acinares, aumentando a secreção, enquanto o VIP actua sobre o músculo liso das arteríolas, aumentando o fluxo sanguíneo local. Exemplo: O neurónio liberta mais que um NT. A noradrenalina e ATP são co-transmissores →
efeitos sinérgicos.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
337 | P á g i n a
Anexo 6 – NT e neuromodulador NT: empacotado em vesículas e é libertado em fendas Neuromodulador: modifica a actividade dos NT, neurónio e actividade sináptica; não é um mensageiro!
Anexo 7 – Astrogliose reactiva As células da micróglia produzem mensageiros que alertam os astrócito da existência de uma lesão. Estes proliferam e limitam a zona da lesão, impedindo o seu alastramento. Os astrócitos produzem mensageiros inflamatórios; As neutrofinas são importantes na protecção e regeneração dos neurónios.
Astrogliose moderada
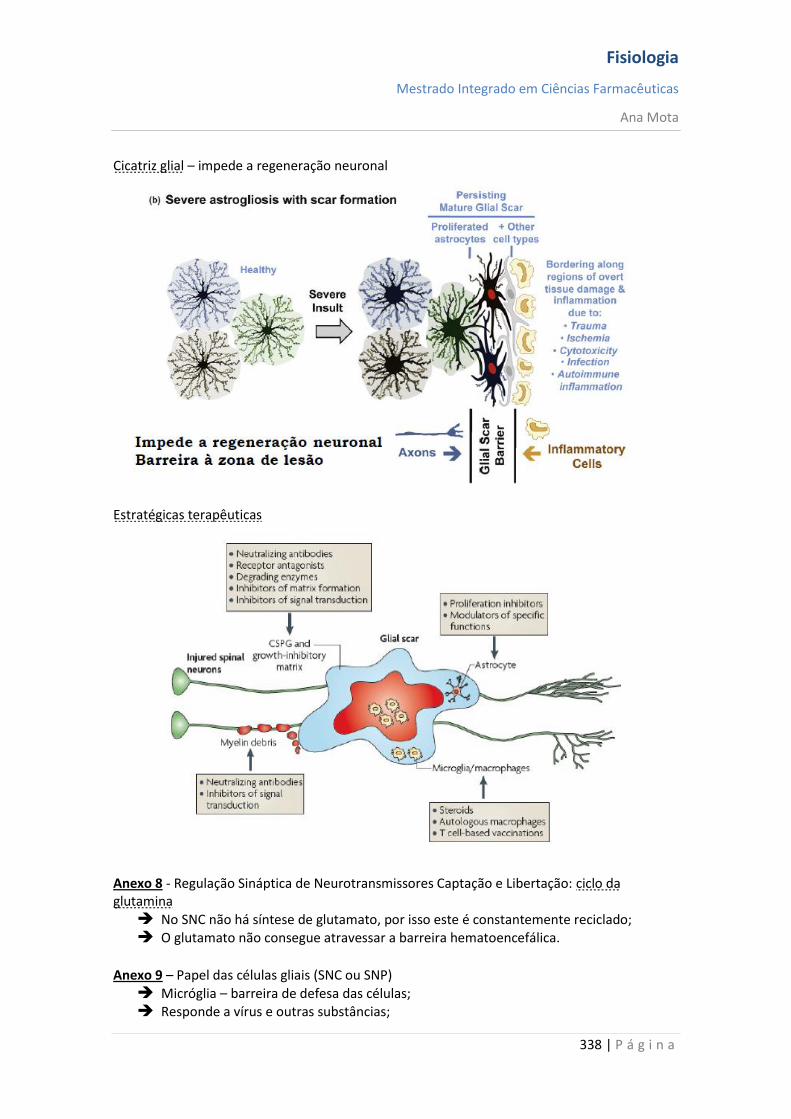
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
338 | P á g i n a
Cicatriz glial – impede a regeneração neuronal
Estratégicas terapêuticas
Anexo 8 - Regulação Sináptica de Neurotransmissores Captação e Libertação: ciclo da glutamina No SNC não há síntese de glutamato, por isso este é constantemente reciclado; O glutamato não consegue atravessar a barreira hematoencefálica.
Anexo 9 – Papel das células gliais (SNC ou SNP) Micróglia – barreira de defesa das células; Responde a vírus e outras substâncias;

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
339 | P á g i n a
Subjacente a receptores nociceptivos: substância P, glutamato. Quando existe SP ou glutamato no meio – produção de ATP, prostanglandinas e outras
substâncias → modificação da transmissão da informação nociceptiva; Glutamato:
- AMPA – ionotrópico que provoca despolarização da membrana em que o 2º neurónio desencadeia um PA; - NMDA – que, normalmente, está bloqueada devido a uma determinada concentração de magnésio (estímulos de fraca intensidade não bloqueiam, mas estas células são activadas por aminoácidos excitatórios (glutamato)). O grau de despolarização é muito elevado e suficiente para a sua activação. Ocorre um aumento da actividade do 2º neurónio → de transmissão → maior percepção e sensibilidade da dor. Anexo 10 – Contracção muscular O PA nos túbulos T induz os receptores DHPR, o que permite a libertação de Ca 2+ do Retículo Sarcoplasmático para o sarcoplasma.
DHPR é um elemento acoplador, ou seja, uma vez sensibilizado pela variação da voltagem da membrana (potencial de acção), activa fisicamente o RyR1 (Ryanodine receptor). Rianodina um receptor é uma proteína que, em humanos, é codificada pelo RYR1 gene. Este gene codifica um receptor de rianodina encontrada no músculo-esquelético. As funções da proteína codificada como um canal de libertação de cálcio no retículo sarcoplasmático, mas também serve para conectar o retículo sarcoplasmático.
Anexo 11 – Cardiomiócito
Fibra muscular cardíaca.
Estrutura:
Apresenta estrutura diferente da Fibra muscular lisa e da Fibra muscular esquelética, devido às particularidades do funcionamento do coração.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
340 | P á g i n a
Funcionamento:
Apresenta potencial de automatismo, ou seja, de ter contracções musculares espontâneas e cíclicas;
Isto ocorre por movimentos de iões através da membrana celular; O ciclo de movimentos iónicos em uma célula excitável, como é o cardiomiócito, é
chamado de Potencial de acção.
Irrigação de sangue da célula:
A célula tem de ser sempre bem irrigada por sangue, pois, se houver a falta de irrigação de sangue no cardiomiócito ira resultar em um enfarte do miocárdio (músculo responsável pelas rigorosas contracções involuntárias do coração).
Anexo 12 – Pressões pulmonares
À medida que a pressão intrapleural diminui, o volume alveolar aumenta, a pressão alveolar baixa para valores inferiores ao da pressão atmosférica e o ar flui para os pulmões. Á medida que o ar entra para os pulmões, a pressão alveolar aumenta e iguala a pressão atmosférica, no final da inspiração.
Pelo contrário, à medida que a pressão intrapleural aumenta, o volume alveolar diminui, a pressão alveolar sobe para valores superiores à pressão atmosférica e o ar sai dos pulmões. O ar ao sair dos pulmões provoca uma diminuição da pressão alveolar que se iguala à pressão atmosférica no final da expiração.
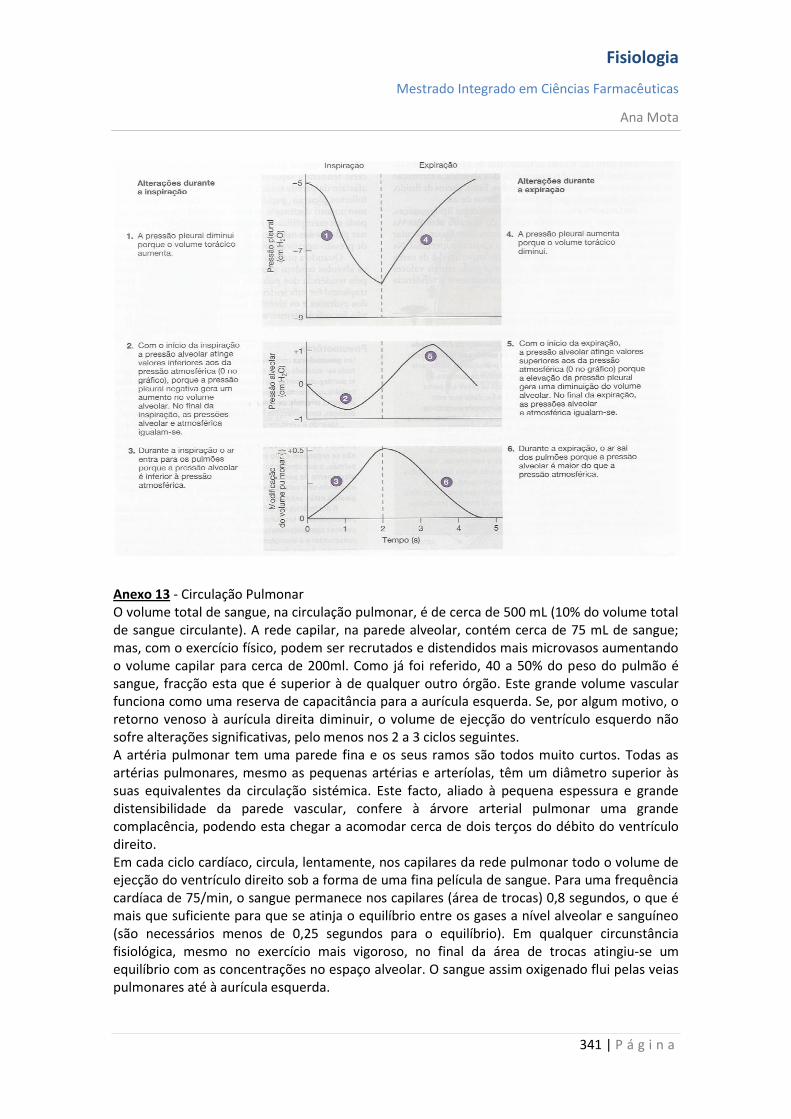
Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
341 | P á g i n a
Anexo 13 - Circulação Pulmonar O volume total de sangue, na circulação pulmonar, é de cerca de 500 mL (10% do volume total de sangue circulante). A rede capilar, na parede alveolar, contém cerca de 75 mL de sangue; mas, com o exercício físico, podem ser recrutados e distendidos mais microvasos aumentando o volume capilar para cerca de 200ml. Como já foi referido, 40 a 50% do peso do pulmão é sangue, fracção esta que é superior à de qualquer outro órgão. Este grande volume vascular funciona como uma reserva de capacitância para a aurícula esquerda. Se, por algum motivo, o retorno venoso à aurícula direita diminuir, o volume de ejecção do ventrículo esquerdo não sofre alterações significativas, pelo menos nos 2 a 3 ciclos seguintes. A artéria pulmonar tem uma parede fina e os seus ramos são todos muito curtos. Todas as artérias pulmonares, mesmo as pequenas artérias e arteríolas, têm um diâmetro superior às suas equivalentes da circulação sistémica. Este facto, aliado à pequena espessura e grande distensibilidade da parede vascular, confere à árvore arterial pulmonar uma grande complacência, podendo esta chegar a acomodar cerca de dois terços do débito do ventrículo direito. Em cada ciclo cardíaco, circula, lentamente, nos capilares da rede pulmonar todo o volume de ejecção do ventrículo direito sob a forma de uma fina película de sangue. Para uma frequência cardíaca de 75/min, o sangue permanece nos capilares (área de trocas) 0,8 segundos, o que é mais que suficiente para que se atinja o equilíbrio entre os gases a nível alveolar e sanguíneo (são necessários menos de 0,25 segundos para o equilíbrio). Em qualquer circunstância fisiológica, mesmo no exercício mais vigoroso, no final da área de trocas atingiu-se um equilíbrio com as concentrações no espaço alveolar. O sangue assim oxigenado flui pelas veias pulmonares até à aurícula esquerda.

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
342 | P á g i n a
Há um padrão anatómico de ramificação vascular arterial, acompanhando a dos bronquíolos, que contribui para o acoplamento ventilação-difusão. As veias, contudo, passam nos septos interlobulares. O grande número de vias circulatórias em paralelo diminui a resistência ao fluxo sanguíneo. A vasculatura pulmonar é muito inervada pelo simpático. No entanto, ao contrário do que acontece na circulação sistémica, a regulação central não é preponderante. A regulação depende predominantemente de influências metabólicas. O sangue também chega aos pulmões através de várias artérias brônquicas (circulação sistémica), uma parcela de 1 a 2% do débito cardíaco. Este sangue arterial é oxigenado, por oposição ao que provém das artérias pulmonares. Permite suprir as necessidades do tecido conjuntivo de suporte, glândulas e paredes das vias aéreas e vasos, com excepção da unidade respiratória terminal, que recebe nutrientes pela circulação pulmonar. Também aquece e humidifica o ar. Regressa à aurícula esquerda pelas veias pulmonares, sendo portanto o retorno ao coração esquerdo 1 a 2% superior ao do direito, o que se repercute nos débitos ventriculares. A circulação pulmonar sofre variações de pressão hidrostática em função da gravidade, que se traduzem por uma diferença de 25cm H2O entre as extremidades superior e inferior do pulmão (1cm H2O /cm altura). A nível arterial, esta acção não é muito significativa; mas, a nível venoso, as pressões nos vértices pulmonares caem para níveis inferiores aos da pressão atmosférica e alveolar (nalgumas fases do ciclo respiratório), o que leva ao colapso venoso e à limitação do fluxo de sangue nessa zona. É possível dividir o pulmão em zonas funcionais de fluxo sanguíneo, que dependem das pressões relativas dos vasos pulmonares em relação à pressão alveolar (vide infra).
Anexo 14

Fisiologia
Mestrado Integrado em Ciências Farmacêuticas
Ana Mota
343 | P á g i n a
Referências e agradecimentos: GUYTON, A; John E. Hall; Tratado de Fisiologia Médica; 10ª edição; Guanabara Koogan;
Rio de Janeiro; 2002; Aulas teóricas Fisiologia 2010/2011; Apontamentos laboratoriais FMUP; Sebenta “Fisiologia 2007”; Apontamentos da Tânia =D .