scale-dependent patterns of variability of a grazing parrotfish (leptoscarus vaigiensis) in a...
TRANSCRIPT

ORIGINAL PAPER
Scale-dependent patterns of variability of a grazing parrotfish(Leptoscarus vaigiensis) in a tropical seagrass-dominated seascape
Martin Gullstrom • Charlotte Berkstrom •
Marcus C. Ohman • Maria Bodin • Mattis Dahlberg
Received: 20 August 2010 / Accepted: 1 March 2011 / Published online: 18 March 2011
� Springer-Verlag 2011
Abstract Although herbivorous fish form critical linkages
between primary producers and higher trophic levels, the
major factors regulating their spatial structure in seagrass
systems remain poorly understood. The present study
examined the parrotfish Leptoscarus vaigiensis in seagrass
meadows of a tropical embayment in the western Indian
Ocean. Stomach content analysis and direct field observa-
tions showed that L. vaigiensis is an efficient grazer, feeding
almost exclusively on seagrass leaves. Seagrass shoot den-
sity was highly correlated to all density variables (total,
juvenile and subadult) and juvenile biomass of L. vaigiensis,
while subadult biomass was predicted by distance to neigh-
bouring coral habitat. Moreover, density and biomass of
predatory fish (piscivores) were predicted by seagrass can-
opy height and the distribution patterns of predators followed
those of L. vaigiensis. Hence, factors at local (seagrass
structural complexity and feeding mode) and landscape scale
levels (seascape context and distribution of piscivores) likely
mutually structure herbivorous fish communities. The
findings underscore the importance of incorporating multi-
ple scale-dependent factors when managing coastal seagrass
ecosystems and their associated key species.
Introduction
Key species play a critical role in determining community
patterns in time and space. Herbivores may encompass
such a role as they form an important functional linkage
between primary producers and predatory consumers of
natural food webs (Valentine and Duffy 2006; Moksnes
et al. 2008). They can trigger cascades both downwards
and upwards in the food-web system, hence functioning as
key conduits for the transfer of energy influencing com-
munity structure and ecosystem processes. Herbivores have
received much attention in terrestrial ecosystems (e.g.
Huntly 1991; Crawley 1997; Bardgett and Wardle 2003) as
well as in some aquatic ecosystems such as coral reefs (e.g.
Mumby et al. 2006; Hughes et al. 2007; Burkepile and Hay
2008); however, remarkably few studies have focused
explicitly on herbivorous fish in seagrass systems.
In the tropical shallow-water seascape (sensu Ogden
1988), and in seagrass biotopes in particular, grazing par-
rotfish can be the dominating feeding category (Lugendo
et al. 2005; Gullstrom et al. 2008), even though other ver-
tebrate herbivores (e.g. waterfowl, turtles and sirenians)
may in some areas locally affect seagrass biomass through
intense grazing (Valentine and Heck 1999). The key func-
tion of parrotfish has been confirmed in a number of studies
(e.g. Kirsch et al. 2002; Alcoverro and Mariani 2004;
Goecker et al. 2005) and can significantly contribute to
ecosystem dynamics and ecological stability (Valentine and
Duffy 2006; Unsworth et al. 2007a). Together with grazing
invertebrates, such as sea urchins and gastropods, seagrass-
Communicated by D. Goulet.
M. Gullstrom (&) � C. Berkstrom
Department of Systems Ecology, Stockholm University,
SE-106 91 Stockholm, Sweden
e-mail: [email protected]
M. Gullstrom � M. C. Ohman � M. Dahlberg
Department of Zoology, Stockholm University,
SE-106 91 Stockholm, Sweden
M. Gullstrom
Department of Marine Ecology - Kristineberg,
University of Gothenburg, SE-451 78 Fiskebackskil, Sweden
M. Bodin
Department of Marine Ecology - Tjarno,
University of Gothenburg, SE-452 96 Stromstad, Sweden
123
Mar Biol (2011) 158:1483–1495
DOI 10.1007/s00227-011-1665-z

grazing fish occasionally control seagrass productivity and
shoot density (Valentine and Heck 1999). In fact, seagrass-
grazing fish alone can potentially consume significant
quantities of seagrass production (Kirsch et al. 2002). As
typical for natural communities, the abundance and distri-
bution of grazing seagrass fish depend on competitive biotic
and abiotic factors (Duffy 2006), which operate across a
wide range of spatial and temporal scales (Levin 1992).
There is however a poor understanding of how interacting
scale-dependent processes in the tropical seascape are
structuring herbivorous fish community patterns.
Seagrass meadows have a multifunctional role within
the coastal seascape, e.g. offering habitat, nursery ground,
food and refuges for numerous fish and invertebrate species
(Jackson et al. 2001; Connolly and Hindell 2006; Nagel-
kerken 2009) as well as influencing biogeochemical and
physical processes (Larkum et al. 2006). Although their
function as nurseries is debatable (Beck et al. 2001; Heck
et al. 2003), seagrass systems have long been known to
provide juvenile fish with food and shelter (e.g. Edgar and
Shaw 1995; Lugendo et al. 2006; Unsworth et al. 2007b),
and they typically harbour greater densities and biomasses
of fish than adjacent unvegetated benthic habitats (Pollard
1984; Bell and Pollard 1989). Furthermore, seagrass
meadows of different species composition often support
distinct assemblages of animals even when meadows are
located near one another (Middleton et al. 1984; Blaber
et al. 1992; Gullstrom et al. 2002; Hyndes et al. 2003).
A number of factors may influence habitat usage by
seagrass-associated fish. Within the tropical seascape,
commonly configured as a mangrove–seagrass–coral reef
continuum (Ogden 1988), the abundance and distribution
of fish are affected by factors at different scales (Pittman
et al. 2004; Dorenbosch et al. 2007; Gullstrom et al. 2008),
even though the relative importance of these factors are not
yet understood. At finer scales, the structural complexity of
seagrass meadows has been suggested to be of influential
importance (Heck and Orth 1980; Bell and Westoby 1986a,
b; Connolly 1994a; Hyndes et al. 2003; Gullstrom et al.
2008). Heck and Orth (1980) suggested that changes in
physical complexity of seagrasses (e.g. shoot density or
canopy height) alter predator foraging efficiency, which in
turn affect abundances and distribution patterns of preda-
tory fish and their prey. This theory has been identified as
the predation hypothesis and might positively affect the
abundance of prey organisms like herbivorous fish, espe-
cially during juvenile life stages. In contrast, Bell and
Westoby (1986a, b) suggested that a high number of
juvenile fish in denser seagrass habitats is primarily a result
of settlement patterns and habitat preference, rather than
reduced predation pressure. They do, however, acknowl-
edge that habitat preference, once larvae have settled, may
have been selected by predation pressure, and hence
predation becomes the ultimate cause of patterns in abun-
dance and distribution. Connolly (1994a, b), on the other
hand, argued that prey availability in seagrass systems is
the crucial agent determining fish numbers and habitat use.
Considering that habitat patches within the shallow
tropical seascape are linked through energy exchange by
fish movement, it is assumed that not only fine-scale
characteristics (habitat complexity, depth, etc.) but also
broad-scale factors such as surrounding seascape configu-
ration (Pittman et al. 2007), proximity to adjacent habitats
(Gullstrom et al. 2008), patch size (Bohnsack et al. 1994)
and spatial arrangement of habitat patches (Grober-Duns-
more et al. 2007) may determine the structure of seagrass-
associated fish communities. The degree of influence of
scale likely depends on contemporary settings in terms of
species-specific home ranges, frequency of daily migration
and the extent to which fish species accomplish ontogenetic
habitat shifts.
The focal point of this study was to examine how the
spatial distribution of the herbivorous parrotfish Leptosc-
arus vaigiensis is related to various ecological and physical
components within a tropical seagrass-dominated seascape
mosaic. We hypothesised that spatial patterns of variability
of L. vaigiensis were determined by factors at scales
ranging from local seagrass characteristics to landscape
structure. Explicitly, we assessed whether density, biomass
and distribution patterns of L. vaigiensis of different life
stages (size classes) were structured by: (1) feeding mode
(food choice and consumption rate), (2) predator presence
(abundance and distribution of potential predators), (3)
seagrass structural complexity (shoot density, shoot bio-
mass and canopy height), (4) local physical water condi-
tions (water temperature, salinity and depth) and/or (5)
landscape configuration (distance to mangrove and coral-
reef habitats).
Materials and methods
Study area
The present study was carried out in Chwaka Bay (6�6–130S, 39� 24–310E, Fig. 1), a shallow embayment (mean
depth: 3.2 m) situated on the east coast of Zanzibar Island
(Unguja), Tanzania. The bay is semi-enclosed, fringed by
an extensive mangrove forest in the south and protected
from intense wave action (from the open Indian Ocean) by
heterogeneous patch reefs at the bay entrance in the north.
A network of channels with water currents predominantly
forced in a north–south direction is widely spread
throughout the bay. The tidal regime is semi-diurnal and
asymmetric with stronger currents at ebb compared with
flood. At high spring tide, the water body of Chwaka
1484 Mar Biol (2011) 158:1483–1495
123

covers an area of 50 km2 while only 20 km2 is covered by
water during low spring tide (Tobisson et al. 1998). The
mean tidal range is 3.2 m at spring tide and 0.9 m at neap
tide (Cederlof et al. 1995).
The bay bottom primarily consists of biogenic deposits
and erosion products derived from fossil limestone terraces
(Tobisson et al. 1998). Seagrasses, interspersed with vari-
ous macroalgae, grow as either large monospecific or
mixed-species meadows in tidal and subtidal areas
(Gullstrom et al. 2006). Eleven species of seagrass are
present, of which Enhalus acoroides, Thalassia hemprichii,
Cymodocea rotundata, C. serrulata and Thalassodendron
ciliatum are the most dominant. Intertidal flats in the
western, central and south-eastern parts of the bay are
predominantly covered by T. hemprichii, Cymodocea spp.
and the calcareous algae Halimeda spp., while the subtidal
central area is mainly composed of E. acoroides and/or
T. ciliatum. Channels are characterised by a mix of
macrophyte assemblages (seagrass and macroalgae) and
unvegetated bottom.
For this study, eight sampling sites were chosen (Fig. 1).
These sites comprised meadows dominated by E. acoroides
(E1, E2 and E3), meadows dominated by T. hemprichii
(T1, T2 and T3), a mixed meadow of E. acoroides, Thal-
assia hemprichii, Thalassodendron ciliatum and sand (M),
and an adjacent unvegetated area (U). The sites dominated
by E. acoroides and T. hemprichii were chosen to represent
large homogenous seagrass meadows. Site U was excluded
from regression analyses due to its lack of seagrass.
Leptoscarus vaigiensis
The seagrass parrotfish L. vaigiensis, also known as the
marbled, blue-spotted or slender parrotfish, is a member of
the family Scaridae and the only known gonochorist
within this fish family (Robertson et al. 1982). This
herbivorous parrotfish is widespread (Nakamura and Sano
2004; Lugendo et al. 2005; Gullstrom et al. 2008) and of
importance in artisanal fisheries for subsistence use (Gell
and Whittington 2002; Davies et al. 2009). It has a par-
ticularly wide distribution with a geographical range from
South Africa to the northern part of the Red Sea and
eastward to Japan and New Zealand, being the only scarid
extending into subtropical regions (Froese and Pauly
2009). Leptoscarus vaigiensis has been found to associate
with drifting algae in parts of its life cycle (Ohta and
Tachihara 2004), but is mostly found in seagrass meadows
and sometimes on hard substrates with a heavy cover of
macroalgae (Randall et al. 1997). It reaches a maximum
size of 35 cm (Froese and Pauly 2009) and has been
recorded to feed predominantly on seagrass leaves
(Almeida et al. 1999; Gell and Whittington 2002; Naka-
mura et al. 2003).
Potential predators
In terms of fish feeders, and according to Froese and Pauly
(2009), Gullstrom et al. (2008) and morphological char-
acteristics, we classified 22 species as potential predators of
L. vaigiensis within the study area. For each predatory
species, we also considered whether there are any onto-
genetic changes in feeding preferences (following Froese
and Pauly 2009).
Data collection
Field surveys were carried out during two periods, the first
extending from November 2002 to January 2003 and the
second one from January to February 2004. During the first
field period (2002/2003), we sampled fish in daytime at
high tide (±2 h) over four consecutive spring tide periods
(except for U, which was only sampled three times due to
Unguja
0 10
Km
N
UngujaChwaka
Bay
6° S
39° 30' E
39° 30' E
6° S
A f r i c a
E3E2E1T3
U
T2
T1
M
Coral reef
MangroveMangrove
10 km
Michamvipeninsula
Fig. 1 Map of study sites in
Chwaka Bay, Zanzibar Island
(Unguja), Tanzania. T1, T2 and
T3 represent sites dominated by
Thalassia hemprichii; E1, E2
and E3 represent sites
dominated by Enhalusacoroides; M represents a site
mixed with E. acoroides,
T. hemprichii, Thalassodendronciliatum and bare sediment; and
U represents an unvegetated
area
Mar Biol (2011) 158:1483–1495 1485
123

logistical weather constraints) using a beam trawl, with an
opening of 2 m in width and 0.6 m in height, and an
attached net with an unstretched mesh size of 6 mm and a
cod end of 1 mm. The sampling was carried out in a
200 9 150 m grid in each of the eight field sites (E1, E2,
E3, T1, T2, T3, M and U). Six semi-randomly selected
200-m transect hauls (replicates) were conducted on each
sampling occasion at every site. All fish were sorted, frozen
and stored on ice for further analysis. In the laboratory,
individuals were identified to the lowest taxonomic level
possible (following Smith and Heemstra 1991), counted,
measured for total length (TL) to the nearest millimetre,
and wet-weighed to the nearest 0.01 g. Based on species0
maximum length (Froese and Pauly 2009), all fishes were
separated into different life stages (size classes) following
Nagelkerken and van der Velde (2002). Accordingly,
juveniles represent individuals of less than one-third of the
species0 maximum length, subadults one-third to two-thirds
of the species0 maximum length and adults more than two-
thirds of the species0 maximum length.
In the survey 2002/2003, we also measured seagrass
structural complexity in terms of shoot density, shoot
(above-ground) biomass and canopy height. These mea-
surements were performed within 20 randomly selected
100-m2 spots in each seagrass habitat site as used for the
sampling of fish. At each of the 20 spots in all seagrass
habitat sites, the density of seagrass shoots was quantified
in two randomly placed 0.0625-m2 quadratic frames. The
seagrass was collected and subsequently analysed in the
laboratory for shoot biomass (dry weight) after being dried
at 80�C during 72 h. To measure canopy height, 8 frames
were placed randomly within each 100-m2 spot and in each
frame 16 seagrass plants were measured according to
Duarte and Kirkman (2001). The three seagrass structural
complexity variables were here used as predictors in
regression analyses, while they were previously analysed in
more detail (see Gullstrom et al. 2008).
During the second field survey (2004), a handful of
seagrass shoots was collected haphazardly at five spots in
four randomly selected locations (25 9 25 m in size) at
each seagrass habitat site dominated by E. acoroides or
T. hemprichii in order to measure and compare the amount
of epiphytes on seagrass leaves of different meadows. After
been carefully rinsed and scraped from epiphytes, all sea-
grass leaves were dried at 80�C during 72 h. Subsequently,
seagrass and epiphytes were weighed (dry and wet)
separately.
To attain physical environmental conditions, water
temperature, salinity and depth were recorded near the top
of the seagrass canopy at the start and end positions of each
fish sampling haul performed. The mean of the two mea-
surements for each environmental variable was used as
predictor data in the regression analyses (see ‘‘Statistical
analyses’’). An additional variable estimated was geo-
graphical position within the embayment, here represented
as the proximity from the central part of each seagrass
locality to the nearest part of mangrove and coral-reef
habitats. Hence, in order to relate fish density and biomass
to a site position, the estimated distances (attained from a
Landsat satellite image) were used as predictor variables in
the regression analyses.
During the field survey 2004, gut content analysis was
carried out for 15 specimens of L. vaigiensis caught in the
meadows dominated by E. acoroides using the same beam
trawl as mentioned above. Furthermore, in situ fish bite
rates on seagrass leaves were recorded during daytime at
high tide (±2 h) in two seagrass meadows dominated by
E. acoroides (E2 and E3) using SCUBA. The reason for
choosing E. acoroides-dominated meadows for the bite rate
study was because the abundance of L. vaigiensis in this
type of meadow has been shown to be significantly greater
than in meadows dominated by T. hemprichii (see Gulls-
trom et al. 2008). Foraging and grazing mode for individual
L. vaigiensis (n = 37) were observed until a change in
behaviour (due to disturbance from the diver) was noticed
(which ranged from 1 to 12 min for different specimens).
Fork length of each fish was estimated to the nearest
centimetre.
Statistical analyses
A nested analysis of variance (ANOVA) was used to
compare epiphyte biomass on seagrass leaves. Meadow
type (2 levels) was a fixed factor, while Site (3 levels) was
a random factor nested within Meadow type. The mixed
(M) and vegetation-free (U) sites were both excluded from
the analysis of epiphyte data. Variability in density and
biomass of L. vaigiensis and its potential predators was
analysed using a mixed-model statistical design with
Meadow type (2 levels, fixed) and Occasion (4 levels,
random) treated as orthogonal factors, and Site (3 levels)
as a random factor nested within Meadow type. Prior to
all analyses, the assumption to meet homogeneity of vari-
ances was checked by Levene’s (1960) test and data were
square-root-transformed. Since all fish data sets showed
heteroscedasticity despite transformation, we did not use
traditional ANOVAs but permutational multivariate anal-
ysis of variance (PERMANOVA, Anderson 2001). In
contrast to traditional ANOVA, significance assessment in
PERMANOVA is carried out using a permutation proce-
dure, rather than relying on nominated p-values assuming
normality (Anderson 2005). Hence, in this study, PER-
MANOVA tests comprised a more proper alternative to
the mixed-model ANOVAs. Euclidean distance was used
as resemblance measure, and all data were square-root-
transformed (with 999 permutations).
1486 Mar Biol (2011) 158:1483–1495
123
Stepwise multiple linear regression analysis was used to
explore the relative importance of various continuous
predictor variables, i.e. seagrass structural complexity
(shoot density, shoot biomass and canopy height), local
physical water conditions (temperature, salinity and water
depth) and proximity from sampled seagrass sites to
neighbouring habitats (mangrove and coral reef) on density
and biomass of L. vaigiensis (juveniles and subadults
separately) and its potential predators, respectively. Prior to
the analysis, the predictor variables were checked for col-
linearity and the probability criteria to enter or remove
predictor variables were specified to 0.05 and 0.1,
respectively.
Results
Comparison of seagrass meadow types
Thalassia hemprichii meadows were generally less dense
(mean shoot density: 768–135 shoots per m2) and shorter
(mean canopy height: 9–21 cm) compared with E. acoro-
ides meadows (mean shoot density: 274–439 shoots per m2;
mean canopy height: 28–49 cm), while the mean shoot
biomass was comparable in the two meadow types (ranging
among sites from 62 to 105 g DW per m2). The mixed
meadow comprised a structural complexity level similar to
meadows dominated by E. acoroides (mean shoot density:
423 shoots per m2; mean canopy height: 34 cm; mean
shoot biomass: 95 g DW per m2). A more detailed com-
parison between the two seagrass habitats can be found in
Gullstrom et al. (2008).
Food preference and feeding mode
A combination of stomach content analysis and direct field
observations showed that L. vaigiensis is an efficient sea-
grass grazer. The analyses of food items in guts from fifteen
L. vaigiensis specimens (mainly juveniles, but also a few
subadults) revealed that more than 95% of the gut contents
consisted of seagrass plant material. In addition, the field
observations of grazing mode support that seagrass was
virtually exclusively the food source of L. vaigiensis, as
intact bites of seagrass leaves were taken and no scraping of
epiphytes was seen. Seagrass items were constantly con-
sumed with an estimated feeding rate of seven bites per
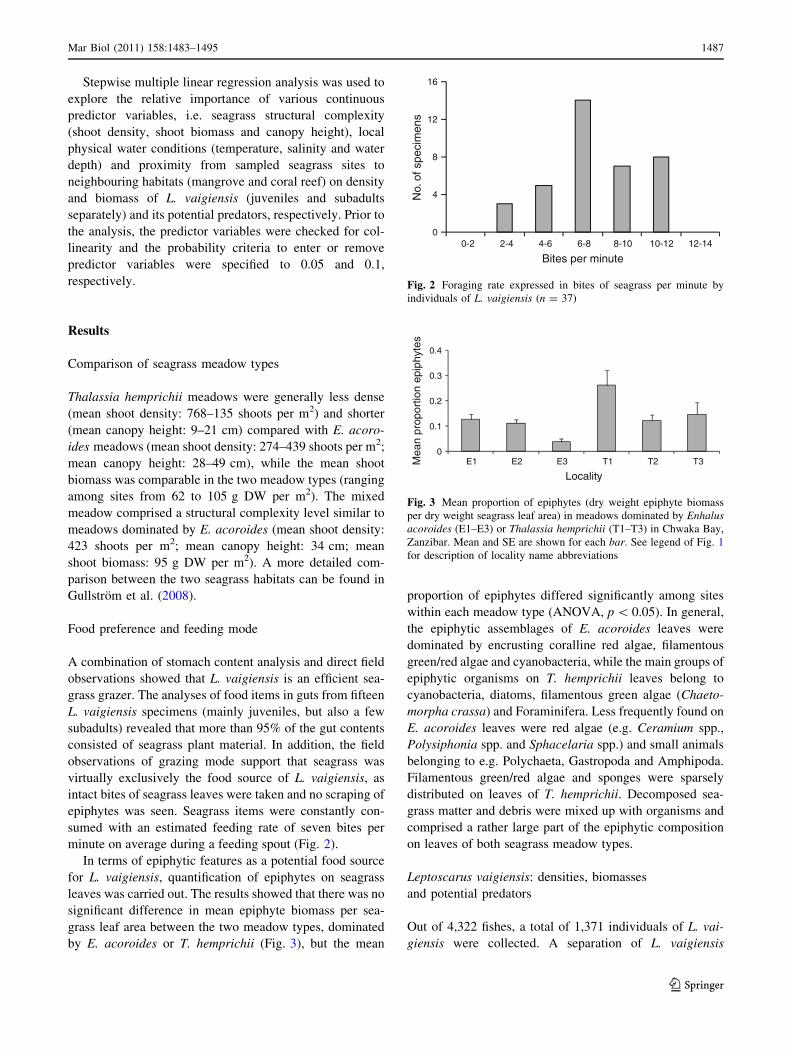
minute on average during a feeding spout (Fig. 2).
In terms of epiphytic features as a potential food source
for L. vaigiensis, quantification of epiphytes on seagrass
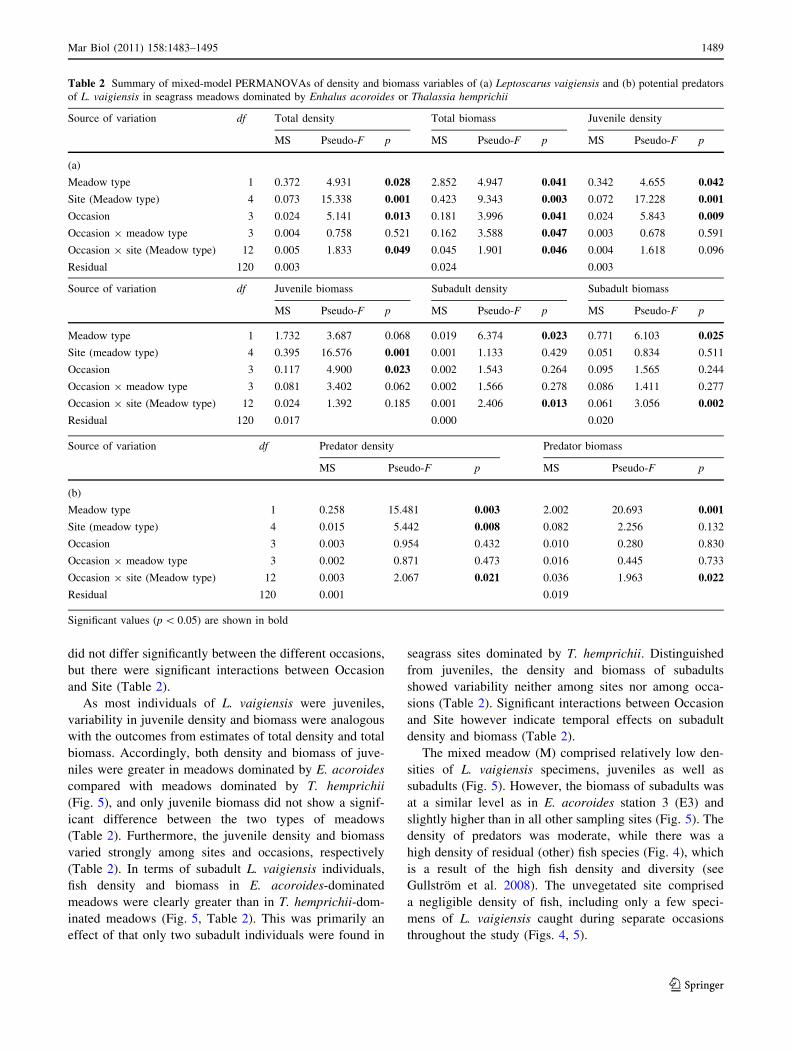
leaves was carried out. The results showed that there was no
significant difference in mean epiphyte biomass per sea-
grass leaf area between the two meadow types, dominated
by E. acoroides or T. hemprichii (Fig. 3), but the mean
proportion of epiphytes differed significantly among sites
within each meadow type (ANOVA, p \ 0.05). In general,
the epiphytic assemblages of E. acoroides leaves were
dominated by encrusting coralline red algae, filamentous
green/red algae and cyanobacteria, while the main groups of
epiphytic organisms on T. hemprichii leaves belong to
cyanobacteria, diatoms, filamentous green algae (Chaeto-
morpha crassa) and Foraminifera. Less frequently found on
E. acoroides leaves were red algae (e.g. Ceramium spp.,
Polysiphonia spp. and Sphacelaria spp.) and small animals
belonging to e.g. Polychaeta, Gastropoda and Amphipoda.
Filamentous green/red algae and sponges were sparsely
distributed on leaves of T. hemprichii. Decomposed sea-
grass matter and debris were mixed up with organisms and
comprised a rather large part of the epiphytic composition
on leaves of both seagrass meadow types.
Leptoscarus vaigiensis: densities, biomasses
and potential predators
Out of 4,322 fishes, a total of 1,371 individuals of L. vai-
giensis were collected. A separation of L. vaigiensis
0
4
8
12
16
0-2 2-4 4-6 6-8 8-10 10-12 12-14
No.
of s
peci
men
s
Bites per minute
Fig. 2 Foraging rate expressed in bites of seagrass per minute by
individuals of L. vaigiensis (n = 37)
0
0.1
0.2
0.3
0.4
E1 E2 E3 T1 T2 T3Mea
n pr
opor
tion
epip
hyte
s
Locality
Fig. 3 Mean proportion of epiphytes (dry weight epiphyte biomass
per dry weight seagrass leaf area) in meadows dominated by Enhalusacoroides (E1–E3) or Thalassia hemprichii (T1–T3) in Chwaka Bay,
Zanzibar. Mean and SE are shown for each bar. See legend of Fig. 1
for description of locality name abbreviations
Mar Biol (2011) 158:1483–1495 1487
123

specimens into life stages showed that 1,301 were juveniles
and 70 were subadults, while no adult fishes were observed.
In total, seven herbivorous species were identified
(Ctenochaetus striatus, Calotomus spinidens, Hipposcarus
harid, L. vaigiensis, Scarus ghobban, S. psittacus and
Siganus sutor). Leptoscarus vaigiensis was strikingly the
most abundant herbivorous fish species and accounted for
68% of all herbivorous specimens (Fig. 4). Twenty-two
species were classified as potential predators to L. vaigi-
ensis (Table 1), with the majority being preferentially
associated with coral and seagrass habitats (Fig. 4).
A comparison of fish densities of the different study
sites indicates similar distribution patterns for L. vaigiensis
and its potential predators. In general, the densities of fish
were greater in E. acoroides-dominated meadows com-
pared with meadows dominated by T. hemprichii (Fig. 4,
Table 2). Total density and total biomass estimates of
L. vaigiensis showed strong variations among sites as well
as among occasions (Table 2), indicating both spatial and
temporal variability. While density of predatory fish also
demonstrated among-site variability, predatory biomass did
not (Table 2). In addition, predatory density and biomass
Table 1 List of potential
predator fish species of
Leptoscarus vaigiensis
Fish are distinguished (based on
Gullstrom et al. 2008, Froese
and Pauly 2009 and this study)
into the habitat preference
groups G (generalists),
M/S (mangrove–seagrass-
associated fish), C/S (coral–
seagrass-associated fish),
S (seagrass residents) and
U (fish exclusively associated
with unvegetated habitat)
Family Species Habitat preference
Aulostomidae (trumpetfishes) Aulostomus chinensis C/S
Balistidae (triggerfishes) Rhinecanthus aculeatus C/S
Dasyatidae (stingrays) Dasyatis kuhlii M/S
Haemulidae (grunts) Plectorhincus gaterinus C/S
Holocentridae (squirrelfishes, soldierfishes) Neoniphon sammara G
Labridae (wrasses) Cheilinus trilobatus C/S
C. undulatus C/S
Cheilio inermis C/S
Oxycheilinus digramma C/S
Lethrinidae (emperors or scavengers) Lethrinus lentjan G
L. variegatus S
Lutjanidae (snappers) Lutjanus fluviflamma G
Muraenidae (moray eels) Echidna nebulosa C/S
Ostraciidae (boxfishes) Ostracion cubicus C/S
Scorpaenidae (scorpionfishes or rockfishes) Scorpaena scrofa C/S
Serranidae (sea basses: groupers, fairy basslets) Epinephelus caeruleopunctatus G
Grammistes sexlineatus C/S
Sphyraenidae (barracudas) Sphyraena sp.
Synanceiidae (stonefishes) Synanceia verrucosa C/S
Syngnathidae (pipefishes, seahorses) Syngnathoides biaculeatus S
Synodontidae (lizardfishes) Saurida gracilis G
Terapontidae (grunters or tigerperches) Pelates quadrilineatus M/S
0
10
20
30
40
50
60
70
80
90
100
E1 E2 E3 T1 T2 T3 M U
OthersPredatorsHerbivores
Spe
cim
ens
1000
m-2
Other fish
Other predators
Coral-seagrass-associated predators
Other herbivores
Leptoscarus
E1 E2 E3 T1 T2 T3 M U E1 E2 E3 T1 T2 T3 M U
Fig. 4 Mean density and
proportion of fish in all
sampling localities. See legend
of Fig. 1 for description of
locality abbreviations
1488 Mar Biol (2011) 158:1483–1495
123
did not differ significantly between the different occasions,
but there were significant interactions between Occasion
and Site (Table 2).
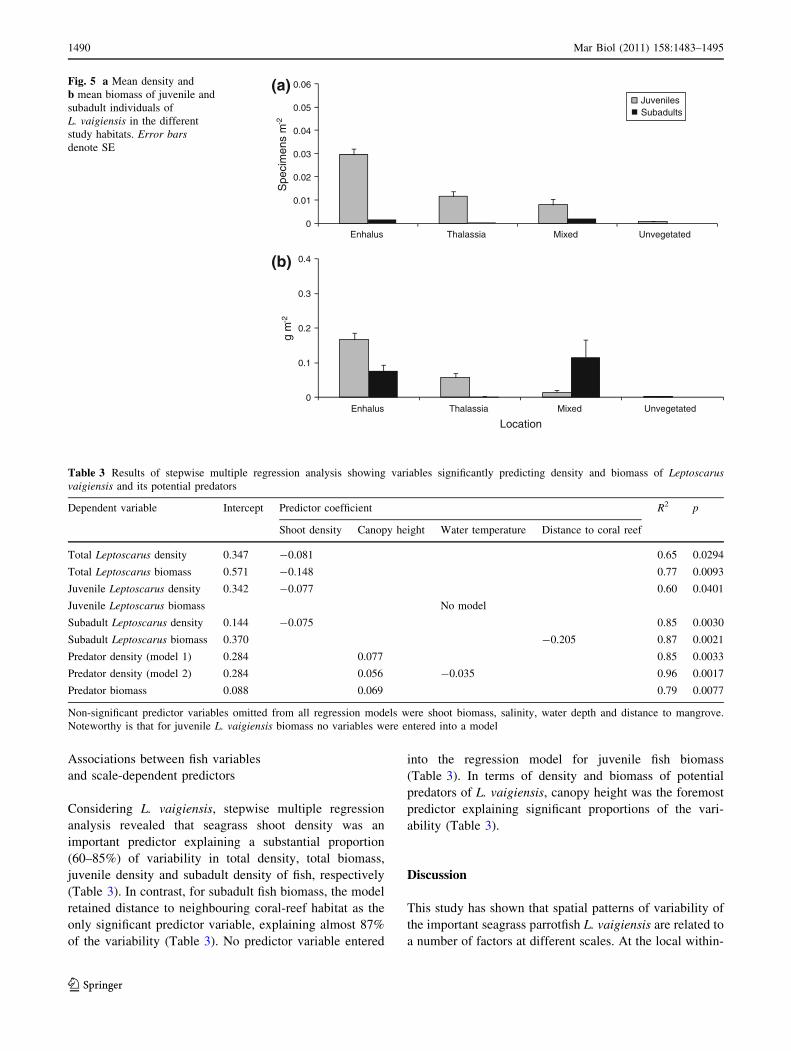
As most individuals of L. vaigiensis were juveniles,
variability in juvenile density and biomass were analogous
with the outcomes from estimates of total density and total
biomass. Accordingly, both density and biomass of juve-
niles were greater in meadows dominated by E. acoroides
compared with meadows dominated by T. hemprichii
(Fig. 5), and only juvenile biomass did not show a signif-
icant difference between the two types of meadows
(Table 2). Furthermore, the juvenile density and biomass
varied strongly among sites and occasions, respectively
(Table 2). In terms of subadult L. vaigiensis individuals,
fish density and biomass in E. acoroides-dominated
meadows were clearly greater than in T. hemprichii-dom-
inated meadows (Fig. 5, Table 2). This was primarily an
effect of that only two subadult individuals were found in
seagrass sites dominated by T. hemprichii. Distinguished
from juveniles, the density and biomass of subadults
showed variability neither among sites nor among occa-
sions (Table 2). Significant interactions between Occasion
and Site however indicate temporal effects on subadult
density and biomass (Table 2).
The mixed meadow (M) comprised relatively low den-
sities of L. vaigiensis specimens, juveniles as well as
subadults (Fig. 5). However, the biomass of subadults was
at a similar level as in E. acoroides station 3 (E3) and
slightly higher than in all other sampling sites (Fig. 5). The
density of predators was moderate, while there was a
high density of residual (other) fish species (Fig. 4), which
is a result of the high fish density and diversity (see
Gullstrom et al. 2008). The unvegetated site comprised
a negligible density of fish, including only a few speci-
mens of L. vaigiensis caught during separate occasions
throughout the study (Figs. 4, 5).
Table 2 Summary of mixed-model PERMANOVAs of density and biomass variables of (a) Leptoscarus vaigiensis and (b) potential predators
of L. vaigiensis in seagrass meadows dominated by Enhalus acoroides or Thalassia hemprichii
Source of variation df Total density Total biomass Juvenile density
MS Pseudo-F p MS Pseudo-F p MS Pseudo-F p
(a)
Meadow type 1 0.372 4.931 0.028 2.852 4.947 0.041 0.342 4.655 0.042
Site (Meadow type) 4 0.073 15.338 0.001 0.423 9.343 0.003 0.072 17.228 0.001
Occasion 3 0.024 5.141 0.013 0.181 3.996 0.041 0.024 5.843 0.009
Occasion 9 meadow type 3 0.004 0.758 0.521 0.162 3.588 0.047 0.003 0.678 0.591
Occasion 9 site (Meadow type) 12 0.005 1.833 0.049 0.045 1.901 0.046 0.004 1.618 0.096
Residual 120 0.003 0.024 0.003
Source of variation df Juvenile biomass Subadult density Subadult biomass
MS Pseudo-F p MS Pseudo-F p MS Pseudo-F p
Meadow type 1 1.732 3.687 0.068 0.019 6.374 0.023 0.771 6.103 0.025
Site (meadow type) 4 0.395 16.576 0.001 0.001 1.133 0.429 0.051 0.834 0.511
Occasion 3 0.117 4.900 0.023 0.002 1.543 0.264 0.095 1.565 0.244
Occasion 9 meadow type 3 0.081 3.402 0.062 0.002 1.566 0.278 0.086 1.411 0.277
Occasion 9 site (Meadow type) 12 0.024 1.392 0.185 0.001 2.406 0.013 0.061 3.056 0.002
Residual 120 0.017 0.000 0.020
Source of variation df Predator density Predator biomass
MS Pseudo-F p MS Pseudo-F p
(b)
Meadow type 1 0.258 15.481 0.003 2.002 20.693 0.001
Site (meadow type) 4 0.015 5.442 0.008 0.082 2.256 0.132
Occasion 3 0.003 0.954 0.432 0.010 0.280 0.830
Occasion 9 meadow type 3 0.002 0.871 0.473 0.016 0.445 0.733
Occasion 9 site (Meadow type) 12 0.003 2.067 0.021 0.036 1.963 0.022
Residual 120 0.001 0.019
Significant values (p \ 0.05) are shown in bold
Mar Biol (2011) 158:1483–1495 1489
123
Associations between fish variables
and scale-dependent predictors
Considering L. vaigiensis, stepwise multiple regression
analysis revealed that seagrass shoot density was an
important predictor explaining a substantial proportion
(60–85%) of variability in total density, total biomass,
juvenile density and subadult density of fish, respectively
(Table 3). In contrast, for subadult fish biomass, the model
retained distance to neighbouring coral-reef habitat as the
only significant predictor variable, explaining almost 87%
of the variability (Table 3). No predictor variable entered
into the regression model for juvenile fish biomass
(Table 3). In terms of density and biomass of potential
predators of L. vaigiensis, canopy height was the foremost
predictor explaining significant proportions of the vari-
ability (Table 3).
Discussion
This study has shown that spatial patterns of variability of
the important seagrass parrotfish L. vaigiensis are related to
a number of factors at different scales. At the local within-
0
0.01
0.02
0.03
0.04
0.05
0.06
Enhalus Thalassia Mixed Unvegetated
Spe
cim
ens
m-2
(a)JuvenilesSubadults
0
0.1
0.2
0.3
0.4
Enhalus Thalassia Mixed Unvegetated
g m
-2
Location
(b)
Fig. 5 a Mean density and
b mean biomass of juvenile and
subadult individuals of
L. vaigiensis in the different
study habitats. Error barsdenote SE
Table 3 Results of stepwise multiple regression analysis showing variables significantly predicting density and biomass of Leptoscarusvaigiensis and its potential predators
Dependent variable Intercept Predictor coefficient R2 p
Shoot density Canopy height Water temperature Distance to coral reef
Total Leptoscarus density 0.347 -0.081 0.65 0.0294
Total Leptoscarus biomass 0.571 -0.148 0.77 0.0093
Juvenile Leptoscarus density 0.342 -0.077 0.60 0.0401
Juvenile Leptoscarus biomass No model
Subadult Leptoscarus density 0.144 -0.075 0.85 0.0030
Subadult Leptoscarus biomass 0.370 -0.205 0.87 0.0021
Predator density (model 1) 0.284 0.077 0.85 0.0033
Predator density (model 2) 0.284 0.056 -0.035 0.96 0.0017
Predator biomass 0.088 0.069 0.79 0.0077
Non-significant predictor variables omitted from all regression models were shoot biomass, salinity, water depth and distance to mangrove.
Noteworthy is that for juvenile L. vaigiensis biomass no variables were entered into a model
1490 Mar Biol (2011) 158:1483–1495
123

meadow scale, seagrass structural complexity and feeding
mode had great influences on the spatial distribution and
abundance of L. vaigiensis, while at the broader landscape
scale, observed patterns highlight the importance of pred-
ator density and seascape structure in determining the
composition of this herbivorous parrotfish. These findings
emphasise the need to consider multiple factors at different
scales to better understand distribution patterns of coastal
fish (cf. Pittman et al. 2004; Gullstrom et al. 2008).
Leptoscarus vaigiensis is a herbivore and feed directly
on seagrass leaves, leaving behind distinct semi-circular
bite marks (McClanahan et al. 1994). In situ observations
of feeding mode in the present study confirmed this feeding
behaviour and showed that grazing occurs constantly and
with a high frequency during active feeding hours of the
day. Furthermore, based on stomach content analysis, we
found that L. vaigiensis preferentially feed on seagrass
leaves. Accordingly, this study confirms the key function of
L. vaigiensis being an efficient seagrass grazer, potentially
comparable to other dominant grazers of the region (e.g.
Alcoverro and Mariani 2004; Eklof et al. 2008). The results
correspond with other studies conducted on parrotfish
foraging in seagrass habitats. For instance, in the Carib-
bean, the bucktooth parrotfish Sparisoma radians was
found to feed predominantly on seagrasses and at a similar
consumption rate to L. vaigiensis in the present study
(Lobel and Ogden 1981; Goecker et al. 2005).
Herbivores can obtain part (or most) of their nutritional
needs in the form of nitrogen from epiphytes (e.g. Tomas
et al. 2005), and hence, it has been suggested that grazing
on seagrasses is determined by nitrogen availability
assimilated by their associated leaf-growing epiphytes
(Valentine and Duffy 2006). In contrast to epiphytes, sea-
grass tissue contains poor nutritional value (e.g. Bjorndal
1980) and is rather unattractive as a food source. This is
because seagrasses have high C/N ratio (Duarte 1990) as
well as high cellulose content, which may be difficult to
digest (Lawrence 1975). Epiphytised seagrass leaves might
therefore be a critical factor explaining herbivore–seagrass
interactions (Tomas et al. 2005). In this study, the pro-
portion of epiphytes in the two seagrass meadow types,
dominated by E. acoroides or T. hemprichii, did not sig-
nificantly differ from each other, and since the density of L.
vaigiensis clearly differed between the two meadow types,
epiphytes might be of inferior importance for the distri-
bution of L. vaigiensis. In addition, the epiphyte commu-
nity composition showed an overlap between meadow
types, which also with respect to nutritional value suggests
a minor impact on the distribution of fish. It should though
be acknowledged that the epiphyte proportion varied
among sites, and hence the effect of epiphytes on the
choice of food by L. vaigiensis cannot be entirely ignored.
Differences in seagrass nutrient concentration might
also influence grazing patterns and in turn the spatial
distribution of herbivorous fish. Duarte (1990) showed
that the content of carbon and nitrogen in seagrass leaves
varies between different species; however, E. acoroides
and T. hemprichii did not differ in terms of their C/N
ratios. With respect to nutritional preferences by grazing
parrotfish, contradictory findings from the literature
indicate great variability. For instance, in the Caribbean,
Goecker et al. (2005) demonstrated that S. radians
selected seagrasses with the highest nitrogen content,
while in the northern Florida Keys, Kirsch et al. (2002)
found no relationship between C/N ratio and grazing
intensity by parrotfish. Nevertheless, T. hemprichii has
been observed to be eaten more frequently than
E. acoroides by herbivorous fish in Kenya (Alcoverro
and Mariani 2004), and furthermore, Dahlgren (2006)
showed a preference for T. hemprichii over E. acoroides
by L. vaigiensis in Tanzania. This suggests that L. vai-
giensis is more common in seagrass meadows dominated
by T. hemprichii, if distribution patterns follow those of
preferred food sources. In our study, however, both
densities and biomasses of fish were significantly higher
in seagrass meadows dominated by E. acoroides com-
pared with those dominated by T. hemprichii, and instead
predator avoidance by hiding in a structurally compatible
seagrass habitat (E. acoroides) may be in priority prior
to the choice of food. It has been suggested that certain
foraging species will balance the predation risk with
resource availability in order to maximise net energy
gain and growth (Orth et al. 1984; Werner and Hall
1988; Warfe and Barmuta 2004). This trade-off between
foraging success and safety has been successfully examined
in different shallow-water assemblages (Werner et al. 1983;
Werner and Hall 1988; Gotceitas 1990; Bologna and Heck
1999; Jormalainen et al. 2001; Salita et al. 2003) and might
well exist for L. vaigiensis. It might be that L. vaigiensis
actually prefers to feed on T. hemprichii, but spends most of
the time in E. acoroides due to the ability of getting pro-
tection from predators. A recent study examining patterns
of grazing bite marks within the edge zone of meadows
composed of T. hemprichii and E. acoroides indicates that
this theory may be accurate (Gullstrom et al. unpublished
data).
It is likely that the spaces between shoots in meadows
with relatively low shoot density (such as our study
meadows comprised of E. acoroides or large-spaced
T. hemprichii) match the sizes of juvenile and subadult
fishes, but restrict larger predators. Bartholomew et al.
(2000) reasoned that inter-structural spaces between sea-
grass shoots that are smaller than a predator’s width would
impede the predator’s movement, and hence provide
Mar Biol (2011) 158:1483–1495 1491
123

sufficient protection in predator–prey encounters. Further-
more, Heck and Orth (2006) highlighted that predators’
foraging from above the seagrass canopy might be influ-
enced by existing seagrass structural complexity. They
suggested that the height of the leaves and the degree to
which they overlap could have an impact on the ability of
down-looking predators to detect prey. In the case of
L. vaigiensis, it may be that the height of E. acoroides
leaves disallow the accessibility of predators and make
L. vaigiensis difficult to detect. Moreover, L. vaigiensis is
well camouflaged with its cryptic coloration and marbled
pattern, a strategy also noted for other seagrass fishes,
including pipefishes and seahorses (Heck and Orth 2006).
In this study, we found a strong negative effect of seagrass
shoot density on the variability of both juvenile and sub-
adult densities of L. vaigiensis, further emphasising the
importance of seagrass habitat complexity in determining
fish distribution patterns. This suggests that L. vaigiensis
prefers meadows comprised of the longer and less dense
E. acoroides to the shorter and denser T. hemprichii.
Among-site differences within each meadow type might
have further strengthened the findings, as the structural
complexity of the highly diverse seagrass meadows in the
studied embayment (Chwaka Bay) vary in a remarkable
manner (long plants intermingling with short plants)
compared with meadows of other regions (Gullstrom et al.
2008). In terms of potential predators of L. vaigiensis,
strong positive relationships were found between seagrass
canopy height and the density and biomass of predatory
fish, respectively. Hence, L. vaigiensis and its potential
predators seem to utilise similar seagrass habitats, although
their co-occurrence is likely of a complex nature.
The ability of seagrass leaves and rhizomes to protect
from predation is one of the major tenets of the seagrass
nursery role hypothesis (Heck and Orth 2006). Body size,
and hence age, may influence an animal’s ability to avoid
predators and/or to harvest resources from the habitat
itself (Werner and Hall 1988). Because these abilities
change with body size during ontogeny, an ontogenetic
shift in habitat or resource use may occur. Numerous
studies have shown such an age-related habitat shift for
tropical fish species moving between mangroves, seagrass
beds and coral reefs (e.g. Nagelkerken et al. 2001, 2002).
By comparing fish assemblage compositions in meadows
dominated by Zostera capricorni and Posidonia australis,
Middleton et al. (1984) found that smaller members of
certain fish families were more common in the short-
leaved Zostera meadows, while that larger individuals of
corresponding families preferred the longer-leaved Posi-
donia meadows. In our study, juveniles were found in
all sites, with the highest densities and biomasses in
the long-leaved E. acoroides meadows. In contrast, sub-
adults were not found in the short-leaved T. hemprichii
meadows (with the exception of two individuals) reach-
ing their density and biomass peaks in the mixed seagrass
meadow, which is located in the slightly deeper area of
the embayment. Adult specimens of L. vaigiensis were
neither found in meadows dominated by T. hemprichii
nor in those dominated by E. acoroides, but have been
observed to occupy e.g. the edges of seagrass habitats in
the slightly deeper water (M. Gullstrom, pers. obs.).
These observations suggest a possible migration of
L. vaigiensis specimens into deeper and more suitable
habitats with age. Furthermore, subadult biomass was
strongly correlated with the distance to neighbouring
coral-reef habitat. Our results seem to conform to Pron-
ker (2002), who found that juveniles of L. vaigiensis
exhibit an ontogenetic shift from seagrass meadows at
the interior parts of Chwaka Bay towards seagrass
meadows near the bay mouth (i.e. near the coral-reef
area). In addition, the highest densities of juveniles were
found in the interior seagrass meadows, while the highest
densities of subadults and adults were found in seagrass
meadows closer to the bay entrance. As most potential
predators of L. vaigiensis are reef-associated, the prox-
imity of coral reefs to seagrass habitats may have an
effect on juvenile L. vaigiensis distribution.
In summary, we have shown that the herbivorous sea-
grass parrotfish L. vaigiensis is an efficient seagrass grazer,
suggesting its key role in a seagrass-dominated environ-
ment like Chwaka Bay. Besides the influence of feeding
mode, our results suggest that the spatial distribution of
seagrass-grazing fish like L. vaigiensis might be influenced
by contemporary density of predators, seagrass structural
complexity and landscape configuration. We conclude that
multiple scale-dependent factors are adequately responsi-
ble for spatial patterns of variability of grazing seagrass
fish assemblages, all from local-scale within-meadow
characteristics to landscape-scale factors. Further research
is encouraged to improve the understanding of spatial
variability of grazing fish communities and to facilitate the
efficacy of management and conservation applications of
coastal seagrass systems.
Acknowledgments We wish to thank Mcha Manzi for assistance
throughout all field periods, and Mohammed Nur Mohammed, Saleh
Yahya, Lisa Adelskold, Maria Asplund, Maricela de la Torre Castro
and Johan Eklof for assistance during part of the field work. We are
also grateful to Kristina Halling, Nils Kautsky, Jerker Lokrantz, and
Per Nilsson for useful comments on earlier versions of the manu-
script. Furthermore, thanks to Mats Lindegarth for statistical advice as
well as Mats Bjork for helping with identification of epiphytes. The
Institute of Marine Sciences (University of Dar es Salaam) at Zan-
zibar provided research facilities and we appreciate the support given
by Narriman Jiddawi. The study was supported by the Swedish
International Development cooperation Agency (Sida) through the
Sida/SAREC Bilateral Marine Science Programme between Sweden
and Tanzania as well as the Sida/SAREC-funded project ‘Changes in
tropical seagrass beds induced by eutrophication’.
1492 Mar Biol (2011) 158:1483–1495
123

References
Alcoverro T, Mariani S (2004) Patterns of fish and sea urchin grazing
on tropical Indo-Pacific seagrass beds. Ecography 27:361–365
Almeida AJ, Marques A, Saldanha L (1999) Some aspects of the
biology of three fish species from the seagrass beds at Inhaca
Island, Mozambique. Cybium 23:369–376
Anderson MJ (2001) A new method for non-parametric multivariate
analysis of variance in ecology. Aust Ecol 26:32–46
Anderson MJ (2005) PERMANOVA: a FORTRAN computer
program for permutational multivariate analysis of variance.
Department of Statistics, University of Auckland, New Zealand
Bardgett RD, Wardle DA (2003) Herbivore-mediated linkages
between aboveground and belowground communities. Ecology
84:2258–2268
Bartholomew A, Diaz RJ, Cicchetti G (2000) New dimensionless
indices of structural habitat complexity: predicted and actual
effects on a predator’s foraging success. Mar Ecol Prog Ser
206:45–58
Beck MW, Heck KL, Able KW, Childers DL, Eggleston DB,
Gillanders BM, Halpern B, Hays CG, Hoshino K, Minello TJ,
Orth RJ, Sheridan PF, Weinstein MR (2001) The identification,
conservation, and management of estuarine and marine nurseries
for fish and invertebrates. Bioscience 51:633–641
Bell JD, Pollard DA (1989) Ecology of fish assemblages and fisheries
associated with seagrasses. In: Shepherd SA (ed) Biology of
seagrasses–a treatise on the biology of seagrasses with special
reference to the Australian region. Elsevier, Amsterdam,
pp 565–609
Bell JD, Westoby M (1986a) Abundance of macrofauna in dense
seagrass is due to habitat preference, not predation. Oecologia
68:205–209
Bell JD, Westoby M (1986b) Importance of local changes in leaf
height and density to fish and decapods associated with
seagrasses. J Exp Mar Biol Ecol 104:249–274
Bjorndal KA (1980) Nutrition and grazing behaviour of the green
turtle Chelonia mydas. Mar Biol 56:147–154
Blaber SJM, Brewer DT, Salini JP, Kerr JD, Conacher C (1992)
Species composition and biomass of fishes in tropical seagrasses
at Groote Eylandt, Northern Australia. Estuar Coast Shelf Sci
35:605–620
Bohnsack JA, Harper DE, McClellan DB, Hulsbeck M (1994) Effects
of reef size on colonization and assemblage structure of fishes at
artificial reefs off southeastern Florida, USA. Bull Mar Sci
55:796–823
Bologna PAX, Heck KL (1999) Differential predation and growth
rates of bay scallops within a seagrass habitat. J Exp Mar Biol
Ecol 239:299–314
Burkepile DE, Hay ME (2008) Herbivore species richness and
feeding complementarity affect community structure and func-
tion on a coral reef. Proc Natl Acad Sci USA 105:16201–16206
Cederlof U, Rydberg L, Mgendi M, Mwaipopo O (1995) Tidal
exchange in a warm tropical lagoon: Chwaka Bay, Zanzibar.
Ambio 24:458–464
Connolly RM (1994a) Removal of seagrass canopy: effects on small
fish and their prey. J Exp Mar Biol Ecol 184:99–110
Connolly RM (1994b) The role of seagrass as preferred habitat for
juvenile Sillaginodes punctata (Cuv. & Val.) (Sillaginidae,
Pisces): habitat selection or feeding? J Exp Mar Biol Ecol
180:39–47
Connolly RM, Hindell JS (2006) Review of nekton patterns and
ecological processes in seagrass landscapes. Estuar Coast Shelf
Sci 68:433–444
Crawley MJ (1997) Plant–herbivore dynamics. In: Crawley MJ (ed)
Plant ecology. Blackwell Science, Oxford, pp 401–474
Dahlgren A (2006) Consumption of seagrass by herbivore parrotfish
in major fishing grounds of Chwaka Bay, Zanzibar. Master’s
Thesis, University of Gothenburg, Sweden
Davies TE, Beanjara N, Tregenza T (2009) A socio-economic
perspective on gear-based management in an artisanal fishery in
south-west Madagascar. Fish Manage Ecol 16:279–289
Dorenbosch M, Verberk WCEP, Nagelkerken I, van der Velde G
(2007) Influence of habitat configuration on connectivity
between fish assemblages of Caribbean seagrass beds, man-
groves and coral reefs. Mar Ecol Prog Ser 334:103–116
Duarte CM (1990) Seagrass nutrient content. Mar Ecol Prog Ser
67:201–207
Duarte CM, Kirkman H (2001) Methods for the measurement of
seagrass abundance and depth distribution. In: Coles RG (ed)
Global seagrass research methods. Elsevier, Amsterdam,
pp 141–153
Duffy JE (2006) Biodiversity and the functioning of seagrass
ecosystems. Mar Ecol Prog Ser 311:233–250
Edgar GJ, Shaw C (1995) The production and trophic ecology of
shallow-water fish assemblages in southern Australia 1. Species
richness, size-structure and production of fishes in Western Port,
Victoria. J Exp Mar Biol Ecol 194:53–81
Eklof JS, Gullstrom M, e Castro M, Muthiga N, Uku J, Muthiga N,
Lyimo T, Bandeira SO (2008) Sea urchin overgrazing of
seagrasses: a review of causes, consequences and management.
Estuar Coast Shelf Sci 79:569–580
Froese R, Pauly D (eds) (2009) FishBase. http://www.fishbase.org
Gell FR, Whittington MW (2002) Diversity of fishes in seagrass beds
in the Quirimba Archipelago, northern Mozambique. Mar
Freshw Res 53:115–121
Goecker ME, Heck KL, Valentine JF (2005) Effects of nitrogen
concentrations in turtlegrass Thalassia testudinum on consump-
tion by the bucktooth parrotfish Sparisoma radians. Mar Ecol
Prog Ser 286:239–248
Gotceitas V (1990) Plant stem density as a cue in patch choice by
foraging juvenile bluegill sunfish. Environ Biol Fish 29:227–232
Grober-Dunsmore R, Frazer TK, Lindberg WJ, Beets J (2007) Reef
fish and habitat relationships in a Caribbean seascape: the
importance of reef context. Coral Reefs 26:201–216
Gullstrom M, de la Torre-Castro M, Bandeira SO, Bjork M, Dahlberg
M, Kautsky N, Ronnback P, Ohman MC (2002) Seagrass
ecosystems in the Western Indian Ocean. Ambio 31:588–596
Gullstrom M, Lunden B, Bodin M, Kangwe J, Ohman MC, Mtolera
SP, Bjork M (2006) Assessment of changes in the seagrass-
dominated submerged vegetation of tropical Chwaka Bay
(Zanzibar) using satellite remote sensing. Estuar Coast Shelf
Sci 67:399–408
Gullstrom M, Bodin M, Nilsson PG, Ohman MC (2008) Structural
complexity and landscape configuration as determinants of tropical
fish assemblage composition. Mar Ecol Prog Ser 363:241–255
Heck KL Jr, Orth RJ (1980) Seagrass habitats: the roles of habitat
complexity, competition and predation in structuring associated
fish and motile macroinvertebrate assemblages. In: Kennedy VS
(ed) Estuarine perspectives. Academic Press, New York,
pp 449–464
Heck KL Jr, Orth RJ (2006) Predation in seagrass beds. In: Larkum
AWD, Orth RJ, Duarte CM (eds) Seagrasses: biology, ecology
and conservation. Springer, Netherlands, pp 537–550
Heck KL Jr, Hays G, Orth RJ (2003) Critical evaluation of the nursery
role hypothesis for seagrass meadows. Mar Ecol Prog Ser
253:123–136
Hughes TP, Rodrigues MJ, Bellwood DR, Ceccarelli D, Hoegh-
Guldberg O, McCook L, Moltschaniwskyj N, Pratchett MS,
Steneck RS, Willis B (2007) Phase shifts, herbivory, and the
resilience of coral reefs to climate change. Curr Biol 17:360–365
Mar Biol (2011) 158:1483–1495 1493
123

Huntly N (1991) Herbivores and the dynamics of communities and
ecosystems. Annu Rev Ecol Syst 22:477–503
Hyndes GA, Kendrick AJ, MacArthur LD, Stewart E (2003)
Differences in the species- and size-composition of fish assem-
blages in three distinct seagrass habitats with differing plant and
meadow structure. Mar Biol 142:1195–1206
Jackson EL, Rowden AA, Attrill MJ, Bossey SJ, Jones MB (2001)
The importance of seagrass beds as a habitat for fishery species.
Oceanogr Mar Biol Annu Rev 39:269–303
Jormalainen V, Honkanen T, Heikkila N (2001) Feeding preference
and performance of a marine isopod on seaweed hosts: cost of
habitat specialization. Mar Ecol Prog Ser 220:219–230
Kirsch KD, Valentine JF, Heck KL Jr (2002) Parrotfish grazing on
turtlegrass Thalassia testudinum: evidence for the importance of
seagrass consumption in food web dynamics of the Florida Keys
National Marine Sanctuary. Mar Ecol Prog Ser 227:71–85
Larkum AWD, Orth RJ, Duarte CM (eds) (2006) Seagrasses: biology,
ecology and conservation. Springer, Netherlands
Lawrence J (1975) On the relationship between marine plants and sea
urchins. Oceanogr Mar Biol Annu Rev 13:213–286
Levin SA (1992) The problem of pattern and scale in ecology.
Ecology 73:1943–1967
Lobel PS, Ogden JC (1981) Foraging by the herbivorous parrotfish
Sparisoma radians. Mar Biol 64:173–183
Lugendo BR, Pronker A, Cornelissen I, de Groene A, Nagelkerken I,
Dorenbosch M, van der Velde G, Mgaya YD (2005) Habitat
utilisation by juveniles of commercially important fish species in
a marine embayment of Zanzibar, Tanzania. Aquat Living
Resour 18:149–158
Lugendo BR, Nagelkerken I, van der Velde G, Mgaya YD (2006)
The importance of mangroves, mud and sand flats, and
seagrass beds as feeding areas for juvenile fishes in Chwaka
Bay, Zanzibar: gut content and stable isotope analyses. J Fish
Biol 69:1639–1661
McClanahan TR, Nugues M, Mwachireya S (1994) Fish and sea
urchin herbivory and competition in Kenyan coral reef lagoons:
the role of reef management. J Exp Mar Biol Ecol 184:237–254
Middleton MJ, Bell JD, Burchmore DA, Pollard DA, Pease BC
(1984) Structural differences in the fish communities of Zosteracapricorni and Posidonia australis seagrass meadows in Botany
Bay, New South Wales. Aquat Bot 18:89–109
Moksnes P-O, Gullstrom M, Tryman K, Baden S (2008) Trophic
cascades in a temperate seagrass community. Oikos 117:763–777
Mumby PJ, Dahlgren CP, Harborne AR, Kappel CV, Micheli F,
Brumbaugh DR, Holmes KE, Mendes JM, Broad K, Sanchirico
JN, Buch K, Box S, Stoffle RW, Gill AB (2006) Fishing, trophic
cascades, and the process of grazing on coral reefs. Science
311:98–101
Nagelkerken I (2009) Evaluation of nursery function of mangroves
and seagrass beds for tropical decapods and reef fishes: patterns
and underlying mechanisms. In: Nagelkerken I (ed) Ecological
connectivity among tropical coastal ecosystems. Springer, Lon-
don, pp 357–399
Nagelkerken I, van der Velde G (2002) Do non-estuarine mangroves
harbour higher densities of juvenile fish than adjacent shallow-
water and coral reef habitats in Curacao (Netherlands Antilles)?
Mar Ecol Prog Ser 245:191–204
Nagelkerken I, Kleijnen S, Klop T, van den Brand RACJ, Cocheret de
la Moriniere E, van der Velde G (2001) Dependence of
Caribbean reef fishes on mangroves and seagrass beds as nursery
habitats: a comparison of fish faunas between bays with and
without mangroves/seagrass beds. Mar Ecol Prog Ser
214:225–235
Nagelkerken I, Roberts CM, van der Velde G, Dorenbosch M, van
Riel MC, Cocheret de la Moriniere E, Nienhuis PH (2002) How
important are mangroves and seagrass beds for coral-reef fish?
The nursery hypothesis tested on an island scale. Mar Ecol Prog
Ser 244:299–305
Nakamura Y, Sano M (2004) Comparison between community
structures of fishes in Enhalus acoroides- and Thalassiahemprichii-dominated seagrass beds on fringing coral reefs in
the Ryukyu Islands, Japan. Ichthyol Res 51:38–45
Nakamura Y, Horinouchi M, Nakai T, Sano M (2003) Food habits of
fishes in a seagrass bed on a fringing coral reef at Iriomote
Island, southern Japan. Ichthyol Res 50:15–22
Ogden JC (1988) The influence of adjacent systems on the structure
and function of coral reefs. Proc 6th Int Coral Reef Symp,
Australia, 1:123–129
Ohta I, Tachihara K (2004) Larval development and food habits of the
marbled parrotfish, Leptoscarus vaigiensis, associated with
drifting algae. Ichthyol Res 51:63–69
Orth RJ, Heck KL Jr, Van Montfrans J (1984) Faunal communities in
seagrass beds: a review of the influence of plant structure and
prey characteristics on predator-prey relationships. Estuaries
7:339–350
Pittman SJ, McAlpine CA, Pittman KM (2004) Linking fish and
prawns to their environment: a hierarchical landscape approach.
Mar Ecol Prog Ser 283:233–254
Pittman SJ, Caldow C, Hile SD, Monaco ME (2007) Using seascape
types to explain the spatial patterns of fish in the mangroves of
SW Puerto Rico. Mar Ecol Prog Ser 348:273–284
Pollard DA (1984) A review of ecological studies on seagrass-fish
communities, with particular reference to recent studies in
Australia. Aquat Bot 18:3–42
Pronker A (2002) Utilization of seagrass, mangrove and other bay
habitats by commercially important fish species in Chwaka Bay,
Zanzibar, Tanzania. Master’s Thesis, University of Nijmegen,
Netherlands
Randall JE, Allen GR, Steene RC (1997) The complete divers0 and
fishermen0s guide to fishes of the great barrier reef and coral sea.
Crawford House Publishing Pty Ltd, Bathurst
Robertson DR, Reinboth R, Bruce RW (1982) Gonochorism,
protogynous sex-change and spawning in three sparisonmatinine
parrotfishes from the western Indian Ocean. Bull Mar Sci
32:868–879
Salita JT, Ekau W, Saint-Paul U (2003) Field evidence on the
influence of seagrass landscapes on fish abundance in Bolinao,
northern Philippines. Mar Ecol Prog Ser 247:183–195
Smith MM, Heemstra PC (1991) Smith0s sea fishes. Southern Book
Publishers, Johannesburg
Tobisson E, Andersson J, Ngazi Z, Rydberg L, Cederlof U (1998)
Tides, monsoons and seabed: local knowledge and practice in
Chwaka Bay, Zanzibar. Ambio 27:677–685
Tomas F, Turon X, Romero J (2005) Effects of herbivores on a
Posidonia oceanic seagrass meadow: importance of epiphytes.
Mar Ecol Prog Ser 287:115–125
Unsworth RKF, Wylie E, Smith DJ, Bell JJ (2007a) Diel trophic
structuring of seagrass bed fish assemblages in the Wakatobi
Marine National Park, Indonesia. Estuar Coast Shelf Sci
72:81–88
Unsworth RKF, Taylor JD, Powell A, Bell JJ, Smith DJ (2007b) The
contribution of scarid herbivory to seagrass ecosystem dynamics
in the Indo-Pacific. Estuar Coast Shelf Sci 74:53–62
Valentine JF, Duffy JE (2006) The central role of grazing in seagrass
ecology. In: Larkum AWD, Orth RJ, Duarte CM (eds)
Seagrasses: biology, ecology and conservation. Springer, Neth-
erlands, pp 463–501
Valentine JF, Heck KL Jr (1999) Seagrass herbivory: evidence for the
continued grazing of marine grasses. Mar Ecol Prog Ser
176:291–302
1494 Mar Biol (2011) 158:1483–1495
123

Warfe DM, Barmuta LA (2004) Habitat structural complexity
mediates the foraging success of multiple predator species.
Behav Ecol 141:171–178
Werner EE, Hall DJ (1988) Ontogenetic habitat shifts in bluegill: the
foraging rate-predation risk trade-off. Ecology 69:1352–1366
Werner EE, Gilliam JF, Hall DJ, Mittelbach GG (1983) An
experimental test of the effects of predation risk on habitat use
in fish. Ecology 64:1540–1548
Mar Biol (2011) 158:1483–1495 1495
123