salts and amino acids as stabilising agents for reconstituted collagen fibres
TRANSCRIPT

BIOCHIMICA ET BIOPHYSICA ACTA 553
BBA 45 O85
SALTS AND AMINO ACIDS AS STABILISING AGENTS
FOR RECONSTITUTED COLLAGEN FIBRES
J. K. CANDLISH AND G. R. TRISTRAM
Department of Physiology and Biochemistry, St Salvator's College, University of St. Andrews, St. Andrews (Great Britain)
(Received January I7th, 1964)
SUMMARY
Collagen fibres have been reconstituted by warming neutralised collagen solutions in the presence of various relevant substances, and their ease of dispersion in KI and urea subsequently determined. Of the substances which appeared to inhibit resolubili- sation of the fibres, a particular study was made of salts and amino acids. Anion and cation series were obtained showing the relative effect of ions, which is probably a consequence of their salting-out potency. All the amino acids tested had a similar, weakly inhibiting effect on fibre redispersion. Kinetic studies showed that the amino acids stabilise the soluble collagen against precipitation but that the fibres once formed are less easily dispersed than controls. The inhibition of dispersion by serine is maximal at pH 6.5 and it is postulated that under these conditions the charge properties of the two ampholytes are summated in such a way that non-specific cross-linkage during fibre formation is suppressed, allowing the formation of specific cross-links. I t cannot be assumed that similar effects operate in vivo, but it is obvious that very simple connective-tissue components may quite markedly influence the properties of collagen fibres.
INTRODUCTION
It has been pointed out by SCHMITT 1 that, following "descriptive" and then "molec- ular" phases of collagen study, we are now entering a phase concerned above all with the bioregulative factors governing the control of collagen fibrogenesis. The present study is intended as a contribution to this last phase. The basis of the experiments lay in the long known fact that acid extracts of skin, when neutralised or precipitated with the correct ionic strength of salt, tend to form fibres with the native 7oo-A longitudinal periodicity observable in the electron microscope s. It was found, howev- er3, * that collagen could be extracted directly from tissues with dilute neutral salt solutions and that such extracts, when warmed to body temperature, quickly formed rigid gels made up of fibres of the native type. It was postulated that this spontaneous fibre formation is analogous to the precipitation of a soluble collagen--isotope experiments 5 implicated the neutral salt-soluble fraction of skin--shed from the fibroblast surface. Later studies have tended to confirm that the formation of collagen fibres is an extracellular process 6-8.
Biochim. Biophys. Acta, 88 (1964) 553-563

554 J.K. CANDLISH, G. R. TRISTRAM
In this study, collagen fibres were formed by warming neutral solutions of it in the presence of various substances which were thought to be of interest in the context of fibrogenesis. Other workers have already studied 9-11 the effects of environment on the rate of fibre formation, but here, on the other hand, we have studied the properties of the fibres once produced, a convenient parameter being the ease of dispersion of the fibres in urea and KI. Redispersion studies will give an index of stability and resistance to solubilisation, these being obviously important for a connective-tissue component situated outwith the protective cell membrane.
Urea is a well-known protein denaturant which at room temperature begins to disrupt the collagen structure at 3 M (ref. 12), whereas KI has a threshold of 0.5 M and solubilises the collagen at lower concentrations without denaturation 13.
GROSS 14 showed that if thermally prepared collagen gels are cooled within a limiting time, then they become less opaque due to resolubilisation of part of the reconstituted fibres. The rate and extent of opacity decrease on cooling was used here also as an index of the resistance of fibres to dispersion.
The physicochemical mechanisms underlying collagen fibrogenesis have been studied by many workers 15-1s. A preliminary report of this work has been published TM.
EXPERIMENTAL Collagen
The acetic acid-soluble fraction of calf skin was prepared as previously 2°. This preparation gave 0.46 % ash, 17.89 % nitrogen, and 13. 9 % hydroxyproline (both of these on an ash-free basis). Hydroxyproline was assayed by the LEACH zl modification
f the method of NEUMANN AND LOGAN. O
Stock collagen solutions
The collagen after exhaustive dialysis against ion-free water, that is, in the form of a salt-free gel, was suspended in a measuring cylinder with an equal volume of twice the desired final concentration of buffer. After homogenisation in a Waring blendor, the mixture was stirred for 2-3 days, then filtered through a thick mat of cotton wool. This has been found to be effective for the removal of all particulate matter. The pH and protein concentration of the filtrate was then checked and if necessary adjusted to the desired values. Usually the stock solution was made up in buffer at pH 5-5; it could be stored thus for about I month at 7 °.
Fibre dispersion studies
Immediately before the start of an experiment some of the stock solution was taken to pH 7.4 with 0.2 N NaOH (at this point aliquots of the solution were taken for protein assay or the amount of alkali added could be measured). The collagen concen- tration was kept as near as possible to 0.2 mg/ml and 4-ml portions of this were added to a series of test tubes containing I ml of the desired concentration of material under study or I ml water as control. The test tubes were then placed in a water bath at 37 ° and left for 15 h. Thereafter the tubes were shaken, causing synaeresis and the pro- duction of protein threads. Aliquots of urea or KI solution were then added to give the desired concentration of dispersant, usually I M KI or 2.5 M urea. The tubes were then shaken mechanically at 37 ° and the time required for complete disappearance of each
Biochim. Biophys. Acta, 88 (1964) 553-563

EFFECT OF SALTS AND AMINO ACIDS ON COLLAGEN 555
of the fibre aliquots noted. This could be done simply and reproducibly by rotating the tubes gently in a strong beam of light.
Fraction of the collagen precipitated Aliquots of the neutralised collagen stock solution were added to tubes containing
the desired materials as before, incubated at 37 ° for 15 h and shaken to cause syn- aeresis. The insoluble fractions were then strained off on mats of cotton wool and the supernatant estimated for nitrogen by Nesslerisation, with subsequent use of the con- version factor of 17.89 % nitrogen.
Kinetic studies
The rate of thermal reconstitution of collagen was studied by following changes in the absorbancy of neutralised collagen in i-cm cells in the Unicam S.P. 500 spectro- photometer at 400 m#. A plate perforated by a pinhole was placed in front of the photocell shutter, as described by WOOD n. The constant-temperature attachment for the spectrophotometer was used in conjunction with an external bath to give the desired temperature in the cells.
Protein estimation
The collagen content of solutions was routinely measured by the microKjeldahl method, using the conversion factor of 17.89 % nitrogen.
Chemicals
These were all used as purchased from reliable dealers, without further purifica- tion. Buffer materials were reagent grade, KI and urea A.R. grade.
RESULTS
Table I gives qualitative observations regarding the ease of dispersion of collagen fibres reconstituted in the presence of various other materials, the collagen being in acetate buffer (I o.16) or in phosphate buffer (I o.I3), and the results being expressed relative to water as control. Conditions were specifically designed (to give relatively short blank dispersion times) such that conserving, i.e. solubilisation inhibiting, effects were most easily manifested, but where a substance had a marked solubilisation-promoting effect, this was noted. The results obtained appeared to be qualitatively similar when either acetate or phosphate buffer was used. I t was found, however, that phosphate being a relatively strong conserving agent in its own right (cf. Fig. 2) tended to mask or swamp the effect of more weakly conserving substances. The quantitative data then were all obtained using acetate buffer. The choice of buffer is important. WOOD n found that the rate of precipitation in phosphate was greater at pH 6.1 and pH 8 than at pH 7, whereas BENSUSAN AND HOYT 1° using Tris buffer, found that an optimum pH of 8 was obtained. Acetate buffer as used here gave maximum rate of about pH 7.8 (Fig. 7 b) and so is similar in this respect to Tris buffer. Acetate buffer was used in preference here since the presence of nitrogen renders protein assay more difficult. Acetic acid appears to solubilise collagen by binding in the unionised form to the protein chain; this may not occur, however, in the pH rahge of fibre'formation (5-9) and may be one of the factors preventing fibre formation at lower pH 2.,
Biochim. Biophys. Acta, 88 (1964) 553-563

556 j.K. C A N D L I S H , G. R. T R I S T R A M
T A B L E I
QUALITATIVE EFFECT OF SUBSTANCES ON THE RESISTANCE TO DISPERSION BY UREA AND K [ OF THERMALLY RECONSTITUTED COLLAGEN
O, n o c h a n g e as c o m p a r e d t o c o n t r o l ; + , i n c r e a s e d r e s i s t a n c e a n d - - , d e c r e a s e d r e s i s t a n c e , a s c o m p a r e d t o c o n t r o l .
Substance Concentration range in Resistance to dispersion as fibre-forming solution compared to control
B o v i n e s e r u m a l b u m i n 8 - 2 5 % of c o l l a g e n o C h o n d r o i t i n s u l p h a t e 8 20 % of c o l l a g e n o H y a l u r o n i c a c i d 8 20 % of c o l l a g e n o H e p a r i n 8 - 2 0 % of c o l l a g e n i n h i b i t s g e l l i n g G l y , A s p , L y s , Ser , P r o ~o -5 lO -3 M + G l u c o s e 1o 5 - 1 o -3 M o N - A c e t y l g l u c o s a m i n e Io 4 - 1 o - 3 M o N a F , N a ~ H P O 4, N a 2 S O 4. 0 . 0 5 - 0 . 2 M + S o d i u m c i t r a t e , s o d i u m a c e t a t e , NaC1 0 . 0 5 - 0 . 2 M + N a B r , L i B r o . o 5 - o . 2 M - - F o r m a l d e h y d e 1-IO ~o + R i b o f l a v i n 5 ' - p h o s p h a t e I - i o m g % + L- a n d D - a s c o r b a t e Io 6 - 1 o - 4 M + F u m a r i c a c i d l o 6 - 1 o 4 M o His tamine 1o 4 i o ~ M - - C o r t i s o n e a c e t a t e o. i - I m g % o D e o x y c o r t i c o n e a c e t a t e o . I i r a g % o C r e a t i n i n e lO -4 i o 2 M - - 5 - H y d r o x y t r y p t a m i n e c r e a t i n i n e
s u l p h a t e Io 4 - 1 o - 3 M o
Series for the effectiveness of cat ions (as sulphates) and anions (as sodium salts) for prevent ing dispersion were obtained (Figs. I and 2) : Ca 2+ > M@+ > K + > NH4+ > Li+; S04 ~- > HP042- > F - > citrate m acetate > C1- > Br .
Li + was placed after NHa+in the series due to its sharp decrease in act iv i ty above o .I M. H e a v y - m e t a l cat ions such as Zn 2+ or Cu 2+ tended to inhibit fibre formation
altogether.
6 0 CO2+ K + &
• ~, 5oF-If A \
\
"O 0.1 0 2 0.3 Molarity of cot:ion
Fig. I. Time for complete dispersion of col lagen fibre al iquots b y i .o ~ K I as a function of the con- centrat ion of cat ions in the fibre-forming solution. B lanks (water replacing salt solution) sub- tracted. Prote in concentrat ion, o.13 mg/ml . Solvent , acetate buffer (I o.16, p H 7.4}. Other con-
dit ions as g iven in EXPERIMENTAL.
Biochim. Biophys. Acta, 88 (1964) 5 5 3 - 5 6 3
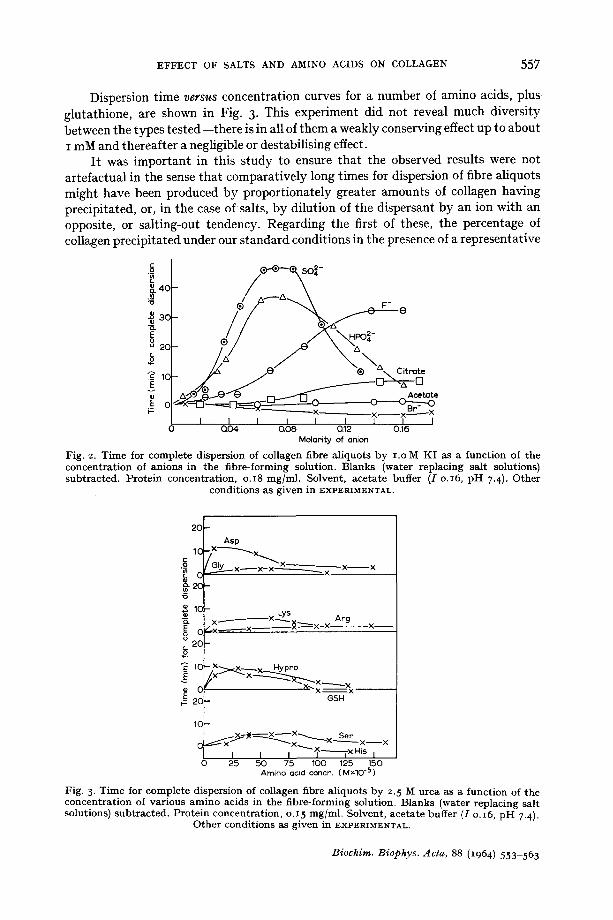
EFFECT OF SALTS AND AMINO ACIDS ON COLLAGEN 5 5 7
Dispersion time v e r s u s concentration curves for a number of amino acids, plus glutathione, are shown in Fig. 3. This experiment did not reveal much diversity between the types tested--there is in all of them a weakly conserving effect up to about I mM and thereafter a negligible or destabilising effect.
It was important in this study to ensure that the observed results were not artefactual in the sense that comparatively long times for dispersion of fibre aliquots might have been produced by proportionately greater amounts of collagen having precipitated, or, in the case of salts, by dilution of the dispersant by an ion with an opposite, or salting-out tendency. Regarding the first of these, the percentage of collagen precipitated under our standard conditions in the presence of a representative
~. 4C
u 2¢
~_ ~o E
~ o ! i-=
o--O~o,~-
I.D/ " ~ ~ e e
J / // /
Q04 Q08 Q12 0.16 Molority of onion
Fig. 2. T ime for comple te dispersion of collagen fibre a l iquots b y i .o M K I as a func t ion of the concen t ra t ion of an ions in the f ibre-forming solut ion. B lanks (water replacing salt solutions) subtracted. Pro te in concent ra t ion , o.18 mg/ml . Solvent , acetate buffer ([ o.16, p H 7.4). Other
condi t ions as g iven in EXPERIMENTAL.
23 t Asp
o= I/^. ~ x -
i "~ ICF . Lys ~- ix - - - - ' - - ' - ' " ' ~ - " ~ } ~ x - X o °E 2C C ~ x ' - - - - - - x ' ~
1C - X ' - - ~ X ~ x Hypro E f x ~ x ~
C x x - - ~x 2C GSH
Arg
~ X x
X ~
lC-- .~____~x~-~-J~x~ Set
~X.....~ X ~ x - - x I I I ~--~---T x His I
0 25 50 75 100 125 150 Amino ocid concn. (M*IO -5)
Fig. 3. T ime for comple te dispers ion of collagen fibre a l iquots b y 2. 5 M urea as a func t ion of the concen t r a t ion of va r ious a m i n o acids in t he f ibre- forming solut ion. B lanks (water replacing salt solutions) sub t rac ted . P ro te in concen t ra t ion , o.I 5 mg/ml . Solvent , acetate buffer (I o. 16, p H 7-4).
Other condit ions as g iven in EXPERIMENTAL.
Biochim. Biophys. Acta, 88 (1964) 553-563
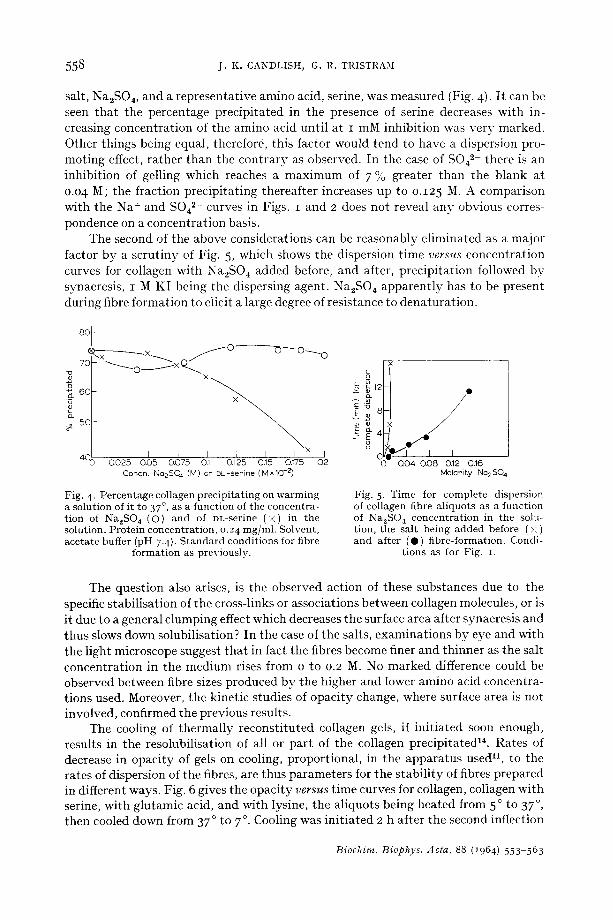
558 j . K . CANDLISH, G. R. TRISTRAM
salt, Na2SO 4, and a representative amino acid, serine, was measured (Fig. 4). I t can be seen that the percentage precipitated in the presence of serine decreases with in- creasing concentration of the amino acid until at I mM inhibition was very marked. Other things being equal, therefore, this factor would tend to have a dispersion pro- moting effect, rather than the contrary as observed. In the case of SO42- there is an inhibition of gelling which reaches a maximum of 7 % greater than the blank at o.o4 M; the fraction precipitating thereafter increases up to o.125 M. A comparison with the Na + and S042- curves in Figs. I and 2 does not reveal any obvious corres- pondence on a concentration basis.
The second of the above considerations can be reasonably eliminated as a major factor by a scrutiny of Fig. 5, which shows the dispersion time v e r s u s concentration curves for collagen with Na2SO ~ added before, and after, precipitation followed by synaeresis, I M KI being the dispersing agent. Na2SO ~ apparently has to be present during fibre formation to elicit a large degree of resistance to denaturation.
8o I i ) ~ x ~ ~ y O -
7C ~ o ~ X ~ --0---~ 0 ~ . _ 0
o 6C
0
o_ 5C
4C I I I I x 0025 0.05 0.075 0.1 0.125 0.15 0.175
Concn. No2SO 4 (M) o r DL-ser ine ( M x l O -2)
I Q2
Fig. 4. Pe rcen tage collagen p rec ip i t a t ing on w a r m i n g a solut ion of it to 37 °, as a func t ion of the concen t ra - t ion of Na2SO 4 (O) a n d of DL-serine ( × ) in the solut ion. Pro te in concen t ra t ion , o. 24 mg/ml . Solvent , ace ta te buffer (pH 7.4). S t a n d a r d condi t ions for fibre
fo rma t ion as previously .
L ~ ~12
8
E 4 o
o o
x I
/ / o
I~10 I I I Q04 0.08 0.12 0.16
Molarity Na2SO 4
Fig. 5 - T i m e for comple te dispers ion of collagen fibre a l iquots as a func t ion of Na2SO t concen t r a t ion in the solu-- t ion, the sal t being added before ( x ) and af ter (O) f ibre-format ion. Condi-
t ions as for Fig. i.
The question also arises, is the observed action of these substances due to the specific stabilisation of the cross-links or associations between collagen molecules, or is it due to a general clumping effect which decreases the surface area after synaeresis and thus slows down solubilisation? In the case of the salts, examinations by eye and with the light microscope suggest that in fact the fibres become finer and thinner as the salt concentration in the medium rises from o to 0.2 M. No marked difference could be observed between fibre sizes produced by the higher and lower amino acid concentra- tions used. Moreover, the kinetic studies of opacity change, where surface area is not involved, confirmed the previous results.
The cooling of thermally reconstituted collagen gels, if initiated soon enough, results in the resolubilisation of all or part of the collagen precipitated 14. Rates of decrease in opacity of gels on cooling, proportional, in the apparatus u s e d 11, to the rates of dispersion of the fibres, are thus parameters for tile stability of fibres prepared in different ways. Fig. 6 gives the opacity v e r s u s time curves for collagen, collagen with serine, with glutamic acid, and with lysine, the aliquots being heated from 5 o to 37 °, then cooled down from 37 ° to 7 °. Cooling was initiated 2 h after the second inflection
Biochim. Biophys. Acta, 88 (1964) 553 563
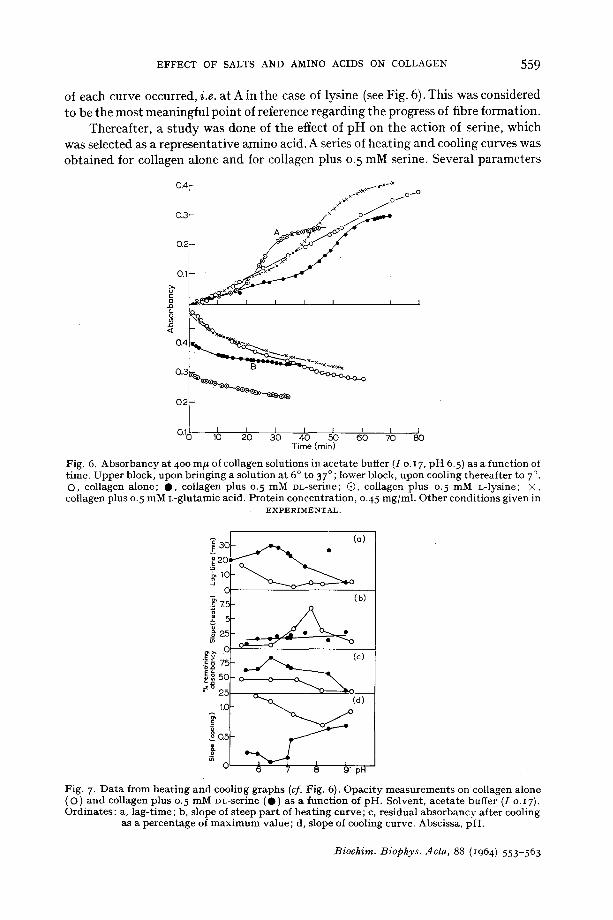
EFFECT OF SALTS AND AMINO ACIDS ON COLLAGEN 5 5 9
of each curve occurred, i.e. at A in the case of lysine (see Fig. 6). This was considered to be the most meaningful point of reference regarding the progress of fibre formation.
Thereafter, a s tudy was done of the effect of pH on the action of serine, which was selected as a representative amino acid. A series of heating and cooling curves was obtained for collagen alone and for collagen plus 0. 5 mM serine. Several parameters
0.4 ~ :,.-× -:~ x~tx~X ~o~O
0.3 /× o '''/~"
O.f~ ~ ~
~ ® ' ~ I [ I I I I
~ -
~ ' ~ L ~ " -~22~&~"~ ×-2
O.1o ,0 4o oo Time (rain)
Fig. 6. A b s o r b a n c y a t 400 m # of collagen solut ions in ace ta te buffer (I o. 17, p H 6.5) as a func t ion of t ime. Uppe r block, upon b r ing ing a solut ion a t 6 ° to 37 °; lower block, upon cooling thereaf te r to 7 °. O, collagen alone; 0 , col lagen p lus 0. 5 mM DL-serine; @, collagen p lus 0. 5 m M L-lysine; × , collagen p lus 0. 5 m M L-glu-tamic acid. P ro te in concen t ra t ion , o.45 mg/ml . O the r condi t ions given in
E X P E R I M E N T A L .
•
~20~
5
~ 0
.-: g 75
~o 2~ 1.(1
(b)
4o (c)
0 0 0 ~ _ _
~ (d)
pN
Fig. 7. D a t a f rom h e a t i n g a n d cooling g raphs (cf. Fig. 6). Opac i ty m e a s u r e m e n t s on collagen a lone (O) and collagen p lus 0. 5 m M DL-serine (O) as a func t ion of pH. Solvent , ace ta te buffer (I o.I7). Ord ina tes : a, l ag- t ime; b, s lope of s teep pa r t of h e a t i n g curve ; c, res idual a b s o r b a n c y af te r cooling
as a pe rcen tage of m a x i m u m value ; d, slope of cooling curve. Abscissa, pH.
Biochim. Biophys. Acta, 88 (1964) 553 563

560 j. K. CANDLISH, G. R. TRISTRAM
were then measured, and these have been plotted in Fig. 7 against pH. They are (a) the lag period or nucleation period as defined by WOOD 32 ; (b) the slope of the steep growth phase 3~ after the first inflection ; (c) the fractional remaining opacity when cooling had produced its maximum effect and which is proportional to the number of collagen fibres remaining intact ; and (d) the slope of the linear part of the cooling curve which was invariably found, for example, at B in Fig. 6. Gelling could not in this system be induced below pH 5.4 or above pH 9.3.
DISCUSSION General
From Table I it can be seen that, in acetate and phosphate buffers, and relative to water as control, many substances tend to make fibres resistant to dispersion. Of these, perhaps the only predictable ones are formaldehyde, which is a well-known tanning agent, and riboflavin 5'-phosphate, which would be expected to have a cross- linking effect due to its quinoid character 13. The other conserving agents can be divided into three groups: (a) some salts, (b) amino acids, (c) ascorbic acid and related enols. Group (c) has been dealt with in a separate detailed study ~'4, and this report deals particularly with salts and amino acids.
It is noticeable that none of the polysaccharides tested appeared to influence the stability of collagen fibres (cf. GROSS14). KEECH 25 has already noted that heparin inhi- bits the clotting of collagen.
Egect of salts The ion series noted in the RESULTS section are essentially the pair of Hofmeister
series as given in standard texts 2e. The Hofmeister series governs the potency of ions as regards interaction with colloids, for example, swelling, salting-out, and viscosity. I t is a consequence of the different degree of hydration of ions; the more intense field of the smaller ions leads to a greater hydration than larger ions of the same charge. The effect of the ions then, in this study, should be directly proportional to a power of the charge and inversely proportional to a power of the ionic radius. Taking as an arbitrary measurement of effectiveness for the inhibition of fibre redispersion the upward slopes of the graphs in Figs. I and 2, and plotting these against the charge squared divided by the ionic radius squared, graphs are obtained which show a roughly linear correla- tion. Although this relation has not been shown to be perfect, it is obvious that it is a salting-out phenomenon we have been here observing. Increasing the ionic strength of, for example, F-, as opposed to CI-, means that collagen during coagulation loses more of its envelope of water and thus charged groups can achieve a more intimate fit, while there is more chance of hydrophobic sites associating, and, the activity of water being lowered, there will be a greater tendency for protein side chains to form hydrogen bonds with other side chains rather than with chains of water molecules. From Figs. I and 2 it is apparent that there is an optimal amount of water removable for fibre stability. Though there is no evidence to point to specific effects of ions on fibrogenesis in vivo, it is possible, at least, that salting-out phenomena may not be merely labora- tory artifices. Is it fortuitous that bone contains large quantities of phosphate, citrate and fluoride27? Skin also contains large quantities of citrate ~s. Moreover, the high con- centration of calcium in aged skin ~ may be one of the factors causing its stability and insolubility, due to the abstraction of water from the fibres.
Biochim. Biophys. Acta, 88 (1964) 553-563
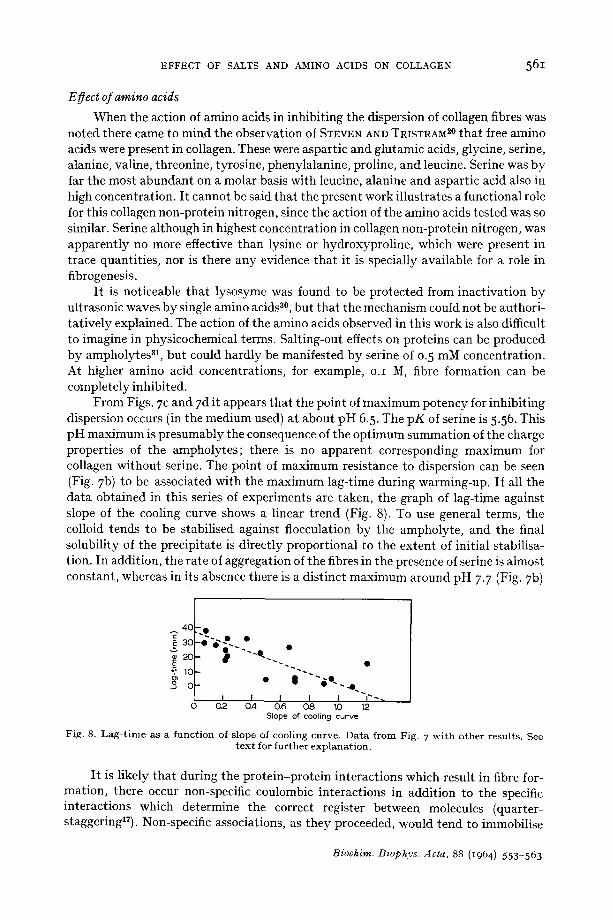
EFFECT OF SALTS AND AMINO ACIDS ON COLLAGEN 561
EBect of amino acids
When the action of amino acids in inhibiting the dispersion of collagen fibres was noted there came to mind the observation of STEVEN AND TRISTRAM 2° that free amino acids were present in collagen. These were aspartic and glutamic acids, glycine, serine, alanine, valine, threonine, tyrosine, phenylalanine, proline, and leucine. Serine was by far the most abundant on a molar basis with leucine, alanine and aspartic acid also in high concentration. It cannot be said that the present work illustrates a functional role for this collagen non-protein nitrogen, since the action of the amino acids tested was so similar. Serine although in highest concentration in collagen non-protein nitrogen, was apparently no more effective than lysine or hydroxyproline, which were present in trace quantities, nor is there any evidence that it is specially available for a role in fibrogenesis.
I t is noticeable that lysosyme was found to be protected from inactivation by ultrasonic waves by single amino acids a°, but that the mechanism could not be authori- tatively explained. The action of the amino acids observed in this work is also difficult to imagine in physic•chemical terms. Salting-out effects on proteins can be produced by ampholytes al, but could hardly be manifested by serine of 0.5 mM concentration. At higher amino acid concentrations, for example, o.I M, fibre formation can be completely inhibited.
From Figs. 7 c and 7 d it appears that the point of maximum potency for inhibiting dispersion occurs (in the medium used) at about pH 6.5. The pK of serine is 5.56. This pH maximum is presumably the consequence of the optimum summation of the charge properties of the ampholytes; there is no apparent corresponding maximum for collagen without serine. The point of maximum resistance to dispersion can be seen (Fig. 7 b) to be associated with the maximum lag-time during warming-up. If all the data obtained in this series of experiments are taken, the graph of lag-time against slope of the cooling curve shows a linear trend (Fig. 8). To use general terms, the colloid tends to be stabilised against flocculation by the ampholyte, and the final solubility of the precipitate is directly proportional to the extent of initial stabilisa- tion. In addition, the rate of aggregation of the fibres in the presence of serine is almost constant, whereas in its absence there is a distinct maximum around pH 7-7 (Fig. 7b)
4C - ~
3c . . , "~ 2£ - •
• e 0• .o.
0 Q2 0.4 O6 0.8 10 t2 Slope of cooling curve
F ig . 8. L a g - t i m e as a f u n c t i o n of s lope of coo l i ng cu rve . D a t a f r o m Fig . 7 w i t h o t h e r r e su l t s . See t e x t for f u r t h e r e x p l a n a t i o n .
It is likely that during the protein-protein interactions which result in fibre for- mation, there occur non-specific coulombic interactions in addition to the specific interactions which determine the correct register between molecules (quarter- staggeringlY). Non-specific associations, as they proceeded, would tend to immobilise
Biochim. Biophys. Acta, 88 (1964) 5 5 3 - 5 6 3

5 6 2 J . K . CANDLISH, G. R. TRISTRAM
the collagen molecules and prevent the formation of the more specific interactions. Serine, however, may act by stabilising the collagen against non-specific associations, allowing the orderly formation of nuclei during the lag-period and subsequent fast aggregation to produce the fibres. Compare the control curve (collagen alone) in Fig. 7, having an almost non-existent lag-period and a long, slow increase in opacity with the curves for the amino acids which have distinct and lengthy lag-periods and distinct steep aggregation phases. The specific register between the chains having proceeded in the presence of the ampholyte, the resultant fibres disperse more slowly than ones with a high proportion of non-specific associations.
I t is notable that evidence has been gained 18 that the formation of ionic bonds between arginyl and acidic residues constitutes the mechanism of fibre formation in collagen, the lysyl residues being merely sites for general strengthening of coulombic interactions rather than sites for specific interactions to determine the structure of the fibres.
The fact that a smaller proportion of collagen finally precipitates in the presence of the ampholyte is indicative that either the ampholyte has a tendency to combine with specific sites in addition to non-specific ones and so further stabilise against flocculation, or that inhibition of non-specific associations in part of the collagen renders that part unable to flocculate. This latter would of course imply heterogeneity of the collagen, but it has already been shown aa that collagen is heterogeneous with respect to the fibre-forming properties of its components.
SCHMITT 1 has stated that "it is difficult to imagine that the sole control of such a major portion of body protein would be left to the biosynthetic centres on the proto- tropocollagen-forming cells, and perhaps to removal by tissue collagenases, all inter- vening stages being dependent on specificities of molecular interaction and the spon- taneous crystallisation of tropocollagen into acid-insoluble fibres involving no dynamic processes or enzyme-regulated systems". It is surely too early to make such a state- ment categorically. The present work has shown that not only the rate of collagen fibrogenesis, but its extent, and the final properties of the fibres, can be quite exten- sively modified by very simple substances known to be present in connective tissues.
ACKNOWLEDGEMENTS
We are indebted to the Scottish Hospital Endowments Research Trust for financial support ; we are grateful to Dr. I. WORRALL for discussions.
R E F E R E N C E S
1 F. O. SCHMITT, Leder, 14 (1963) 21. 2 F. O. SCHMITT, C. E . HALL AND iV[. A. JAKUS, J. Cellular Comp. Physiol., 20 (1942) i i . 3 j . GRoss , J . H. HIGHERGER AND F. O. SCHMITT, Proc. Natl. Acid. Sci. U.S., 41 (1953) i . 4 D. S. JACKSON AND J. H. FESSLER, Nature, 176 (1955) 69. 5 R. D. HARKNESS, A. M. MARKO, H. IV[. MUIR AND A. NEUBERGER, Biochem. J., 56 (1954) 558. 6 I~. l:{. PORTER AND G. D. PAPPAS, J. Biophys. Biochem. Cytol., 5 (1959) 153. 7 M. D. SCHOENBERG, A. HINMAN AND ~ . D. MOORE, Lab. [riPest., 9 (196o) 305 • 8 K. KAJIKAWA, Abstr. Japan. Med., I, (1961) 1488. 9 j . GROSS AND D. KIRK, J. Biol. Chem., 233 (1958) 355-
10 H. B. BENSUSAN AND B. L. HOYT, J. Am. Chem. Soc., 80 (1958) 719 . 11 G. C. WOOD, Biochem. J., 76 (196o) 598. 12 F. S. STEVEN AND G. t{. TRISTRAM, Biochem. J., 85 (1962) 207.
Biochim. Biophys. Acta, 88 (1964) 5 5 3 - 5 6 3

EFFECT OF SALTS AND AMINO ACIDS ON COLLAGEN 563
13 j . CANDLISH, Biochim. Biophys. Acta, 74 (1963) 275. la j . GROSS, J. Exptl. Med., lO8 (1958) 215. 15 j . T. RANDALL, F. BOOTH, R. E. BURGE, S. F. JACKSON AND F. C. KELLY, Syrup. Soc. Exptl.
Biol., 9 (1955) 127. 16 K. KUHN, W. GRASSMANN AND U. HOFMANN, Naturwissenschaften, 47 (196o) 258. 17 A. J. HODGE AND F. O. SCHMITT, Proc. Natl.Acad. Sci. U.S., 146 (196o) 186. is H. B. BENSUSAN, V. R. MUMAW AND A. W. SCANU, Biochemistry. i (1962) 215. 19 j . K. CANDLISH AND G. R. TRISTRAM, Biochem. J., 89 (1963) 79 P. 20 F. S. STEVEN AND G. R. TRISTRAM, Biochem. J., 83 (1962) 240. 21 A. A. LEACH, Biochem. J., 74 (196o) 61. 22 G. R. TRISTRAM, unpub l i shed resul ts . 23 K. H. GOSTAVSON, The Chemistry and Reactivity of Collagen, Academic Press, New York, 1956,
p. 279. 22 j . K. CANDLISH AND G. R. TRISTRAM, Biochim. Biophys. Acta, 78 (1963) 289. 2~ M. K. KEECH, J. Biophys. Biochem. Cytol., 9 (1961) 193. 26 H. B. BULL, PhysicalBiochemistry, Wiley, New York, 1951, p. 85. 27 W. D. ARMSTRONG AND L. SINGER, in G. WOLSTENHOLME AND G. O'CONNOR, Bone Structure and
Metabolism, Churchil l , London , 1956, p. lO 3. 28 H. E. FIRSCHEIN AND J. BELL, J . Biol. Chem., 236 (1961) 22. 29 A. I. LANSING, Science, lO6 (1947) 187. 3o F. M. DIETRICH, Nature, 195 (1962) 146. 31 j . T. EDSALL AND J. WYMANN, Biophysical Chemistry, Vol. i , Academic Press, New York, 1958,
p. 278. 22 G. C. WOOD, Biochem. J., 76 (196o) 605. 32 j . H. FESSLER, Biochem. J., 76 (196o) 452.
Biochim. Biophys. Acta, 88 (1964) 553-563