revised – may 24, 2000
TRANSCRIPT

1
REVISED – MAY 24, 2000
Evidence of an unusually long operator for the Fur (ferric uptake) repressor in the
aerobactin promoter of Escherichia coli
by
Lucía Escolar1, José Pérez-Martín and Víctor de Lorenzo*
Department of Microbial Biotechnology, Centro Nacional de Biotecnología CSIC, Campus de
Cantoblanco, 28049 Madrid, Spain
Running Title : The Fur operator in the aerobactin promoter
___________________________________________________________________________
1Current address: Institut für Genetik. Biozentrum, Weinbergweg, 22 06120 Halle (Saale)
Germany
___________________________________________________________________________
*Corresponding author : Víctor de Lorenzo
Department of Microbial Biotechnology
Centro Nacional de Biotecnología-CSIC
Campus de Cantoblanco, 28049 Madrid, Spain
Tel +34 91-585 4536, Fax +34 91-585 4506
E-mail: [email protected]
JBC Papers in Press. Published on May 31, 2000 as Manuscript M002839200 by guest on A
pril 10, 2018http://w
ww
.jbc.org/D
ownloaded from

2
SUMMARY
Production of the siderophore aerobactin in Escherichia coli is transcriptionally
metalloregulated through the iron-dependent binding of the Fur (ferric uptake
regulator) to a large region (>100 bp) within the cognate promoter in the pColV-K30
plasmid. We show in this article that such an unusually long operator results from the
specific addition of degenerate repeats 5'NATA/TAT3´ and not from a fortuitous
occupation of the DNA adjacent to the primary binding sites by an excess of the
repressor. Furthermore, the protection pattern revealed by DNase I and hydroxyl
radical footprinting reflected a side-by-side oligomerisation of the protein along an
extended DNA stretch. This type of DNA-protein interactions is more alike those
observed in some eukaryotic factors and nucleoid-associated proteins than typical of
specific prokaryotic regulators.
___________________________________________________________________________
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
INTRODUCTION
Prokaryotic transcriptional regulators bind DNA in order to repress or activate
expression of specific genes or groups of genes (1). Although the sequences
recognised can be extremely diverse, most regulatory proteins naturally bind discrete
target sites within the bacterial genome. However, some regulators (typically the
nucleoid-associated proteins, 2) are also known to bind somewhat degenerated
sequences or structural motifs, thus spreading DNA-protein interactions along
extended nucleotide sequences. This feature is shared with a variety of eukaryotic
regulators, typically those containing Zn-fingers such as the TFIIIA (3). In this
respect, the Fur protein of Escherichia coli displays both, properties found in specific
transcriptional factors and in more global regulators. Fur is the product of the fur
(ferric uptake regulation) gene (4, 5, 6, 7), which controls transcription of iron-
dependent promoters in many prokaryotes. This regulator is a Zn-containing, Fe2+-
binding protein (8) which inhibits transcription of distinct genes implicated in the
response to iron starvation when the metal is in excess in the medium (9, 10, 11, 12).
But, in addition, Fur appears to play an important role also in a variety of cell
functions unrelated to iron acquisition, such as the production of several virulence
determinants (13), the defence against oxygen radicals (14, 15), the acid shock
response (16), chemotaxis (17), metabolic pathways and others (18, 19, 20, 21, 22, 23).
The interaction of the Fur protein-Fe2+ complex with its operators has been
characterised with diverse techniques in several promoters of E. coli and other genera
(14, 24, 25, 26, 27, 28, 29, 30). These studies have revealed that every Fe-dependent
promoter contains a target DNA sequence with different degrees of similarity to a
palindromic 5'GATAATGATAATCATTATC3', 19 bp consensus box (26, 31, 32). More
recently, we have reinterpreted such a consensus as the combination of 3 repeats of
the simpler motif 5'NATA/TAT3´ (33), in which the thymines would be the bases
determining the type of contact of the Fur protein with such a minimal unit of
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
interaction. The corollary of this interpretation is that extended sites for Fur binding
could be naturally or artificially assembled by simply adding multiple adjacent
5'NATA/TAT3´ hexamers to a minimum of 3 repeats. This is a very attractive
possibility, since it would permit the generation of repertoires of binding sites of
varying entensions and affinities which would allow Fur to act in some promoters as
a very specific regulator and in others as a more general co-regulator (12). While this
notion has been substantiated using synthetic DNA sequences consisting of
synthesized 5'NATA/TAT3´ hexamers (33), it is unclear whether long sequence-
dependent Fur operators are operative in natural iron-regulated promoters. The
promoter of the operon responsible for the biosynthesis of the aerobactin siderophore
(referred hereafter as Paer) is particularly interesting in this respect (34, 26, 32, 33).
Unlike other promoters controlled by Fur in which the operator involves a clear-cut
target sequence (24, 27, 28, 29, 13), Paer is bound by the repressor to 3 distinct extents
depending on the concentration of the protein (Fig. 1; see below). While Fur binding
to the adjacent sites named I and II can be justified by their similarity to the
consensus, the massive protection of the further upstream sequences (the so called
polymerization region, Fig. 1) is intriguing, since it does not contain clear Fur
consensus boxes. Such an extensive occupation of the promoter by the repressor
spreading over 100 bp has been revealed not only by DNase I and hydroxyl radical
footprinting (32, see below) but also visualized directly through electron
microcoscopy (35). Other iron-regulated promoters appear to undergo such an ample
occupation as well (25, 36), so it might be a genuine phenomenon and not an
unspecific protection caused by an excess of the protein.
In this work, we show that the so far unaccounted binding of the Fur protein to the 5'
upstream region of the aerobactin promoter is due to the functionality of a long
operator composed of 9 adjacent 5'NATA/TAT3' hexamers. This operator, which is
entirely sequence-dependent, becomes effective only following the occupation of the
other two sites. These results support the notion that the binding of Fur to DNA is
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
mediated by the recognition of hexameric repeats and that an increasing number of
adjacent repeats allow a co-operative binding of the repressor mediated by lateral
protein-protein interactions. Furthermore, we argue that this type of interaction, that
has features reminiscent of some transcription factors (37) endows the protein with
the ability to behave both as a very specific repressor and as a more general regulator.
EXPERIMENTAL PROCEDURES
General procedures- The Fur protein used in all the assays was purified to
homogenity following the metallo-affinity purification protocol of Wee et al. (7).
According to (8), such purification protocol yields a Fur protein containing 1 atom of
Zn per repressor monomer, whose DNA-binding ability is responsive to Mn2+ in our
assays system (see below). Protein Fur concentrations indicated through this work
refer to the protein monomer. DNA techniques were run according to published
protocols (38).
DNA templates for footprinting assays- The organisation of the DNA fragments used in
footprinting assays is shown in Fig. 2. The fragment wt1 is a 368 bp EcoRI-PvuII
segment from plasmid pUC-LE15 which contains the region spanning positions -128
to +32 of the aerobactin promoter region (using as a reference the transcription start
site of the main promoter P1) as an EcoRI-BamHI plus a vector-born unrelated BamHI-
PvuII extension of 208 bp. The strategy for creating the promoter termed ∆50 is
sketched in Fig. 2A as well. Primers were devised for amplification of the sequence -
50 to +32 (thus excluding the P2 promoter) and the upstream extension region. This
fragment was recloned in pUC19 using the EcoRI and BamHI sites present in the
amplified fragments (BamHI already present and EcoRI entered with the rightwards
primer). In order to get a template of identical size to wt1 for the footprint assay, a
primer was engineered which contained a terminal NcoI site located at exactly the
same distance that the EcoRI site of the wt1 promoter (see Fig. 2B). Such a segment
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
was then entered at the single HincII site of the previous pUC19 derivative. This new
plasmid contained an insert in the vector that spans the new promoter construct
∆50. The fragment generated after restriction with NcoI-PvuII allowed a base-wise
comparison of its footprint with the wild-type fragment because the end-sites, EcoRI
or NcoI, were located exactly at the same point.
For the second series of templates shown in Fig. 2B, modified variants of the
aerobactin promoter with increasing distance between the Fur box I (Fig. 1) and the
downstream protected region were constructed as follows. In the EcoRI-BamHI insert
of plasmid pUC-LE15 a novel ClaI restriction site was introduced by site-directed
mutagenesis at the boundary betwen Fur boxes I and II (Fig. 2B) with the method of
Kunkel (39). Digestion of the resulting construct with ClaI, filling-in of the cohesive
ends and religation originated a novel NruI as well as +2 bp insertion between the
boxes. The same ClaI-digested plasmid was ligated to the linker 5'CGACCATGGT3',
which entered a novel NcoI site as well as a +10 insertion. Finally, NcoI digestion of
the resulting construct, filling-in the cohesive ends and religation generated the +14
bp along with a new NsiI site. The mutated segments were cloned back to pUC19
and used as the source of the end-labelled restriction fragments employed in the
footprinting assays. To this end, they were excised from these pUC19 derivatives as
EcoRI-PvuII or NcoI-PvuII segments (for labelling of the bottom strand) and purified
by electrophoresis on non-denaturing 5 % polyacrylamide gels. The overhanging
ends of the restriction fragments were then filled-in with [α32P] dATP and Klenow
polymerase, after which they were further purified from non incorporated
nucleotides on small Sephadex G-25 columns.
Footprinting with DNase I and hydroxyl radicals- DNA-protein interactions were
probed with DNase I as described in references 32 and 33. Samples were
preincubated for five minutes at 37º C with the amounts of the Fur protein indicated
in each case. Each tube was then added with 2.5 ng of DNase I and further incubated
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
for two minutes. Reactions were stopped, nucleic acids precipitated, dried and
directly resuspended in 7 M urea (with tracking dyes). Samples were loaded on 7-10
% polyacrylamyde sequencing gels with 7 M urea. A+G reactions (40) with the same
labeled DNA fragments or sequencing size markers were loaded onto the gels
together with the treated samples. Footprinting of DNA with hydroxyl radicals
generated in situ with Fe/EDTA/ascorbate were carried out (32) on Fur-DNA
mixtures prepared and preincubated in the same conditions than before.
RESULTS AND DISCUSSION
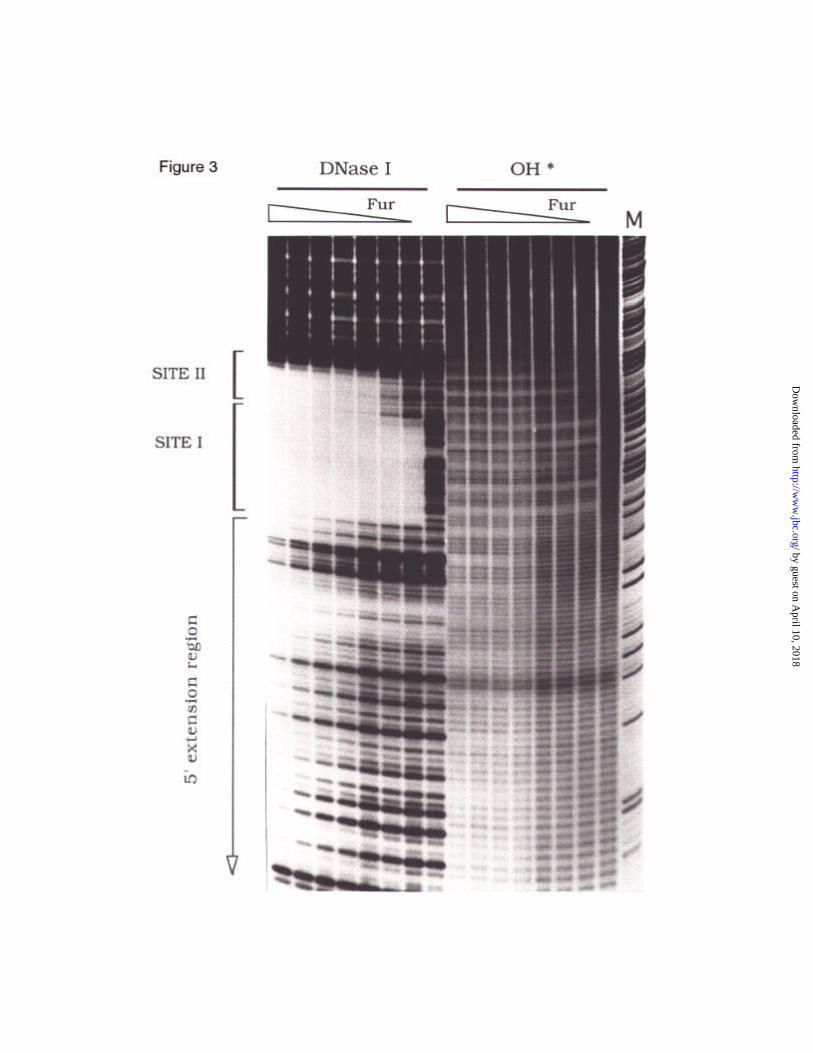
Visualisation of a continuous pattern of Fur-DNA interactions through the areobactin
promoter region- In order to match faithfully the extensions of each binding site for the
Fur protein along the Paer promoter with the specific bases involved in protein-DNA
contacts we made the experiment shown in Fig. 3. In it, we compared directly the
protections caused by increasing concentration of Fur-Mn2+ to either DNase I nicking
or hydroxyl radical cleavage of sugar-phosphate bonds (Mn was used instead of Fe2+
due its superior stability under the aerobic conditions of the experiment; 11, 12, 26).
The reference DNase I footprint to the left of the gel shown in Fig. 3 revealed the
position of each of the known 3 regions sequentially protected by growing repressor
concentrations. This includes first a 31 bp sequence (site I) spanning the -35 hexamer
of the P1 promoter, an additional 18-19 bp downstream protection (site II)
overlapping the -10 box, and the less defined further upstream region protected
towards 5´ (26). Although weaker than those of sites I and II, this last protection
involves exactly 60 bp, so that the addition of all sites covered by the protein at the
higher protein concentration comes to 110 bp. When the same protein-DNA contacts
are inspected in strict parallel with hydroxyl radical footprinting, some salient
features become apparent. First, that the distinction between the 3 binding regions
revealed by DNase becomes less clear-cut. At the higher Fur concentration, the whole
of the 110 bp DNA sequence displays a continuous and repetitive pattern of two
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
protected residues/four non-protected bases. The frame of such a regular pattern is
shifted only once and by one base at the very boundary between the DNase I site I
and the upstream protected region, to then resume the previous 2 protected/4
nonprotected mould through the further upstream region. Furthermore, the data of
Fig. 3 suggest that the occupation of such an upstream region does not commence
until the site II is fully bound by the repressor. Since we could not distinguish
discrete binding sites within the 110 bp region, but rather a continuum of repetitive
interactions, we wondered whether the entire DNA stretch actually functions as a
natural, extended operator of the type predicted by the re-interpretation of the Fur
consensus sequences presented before (33).
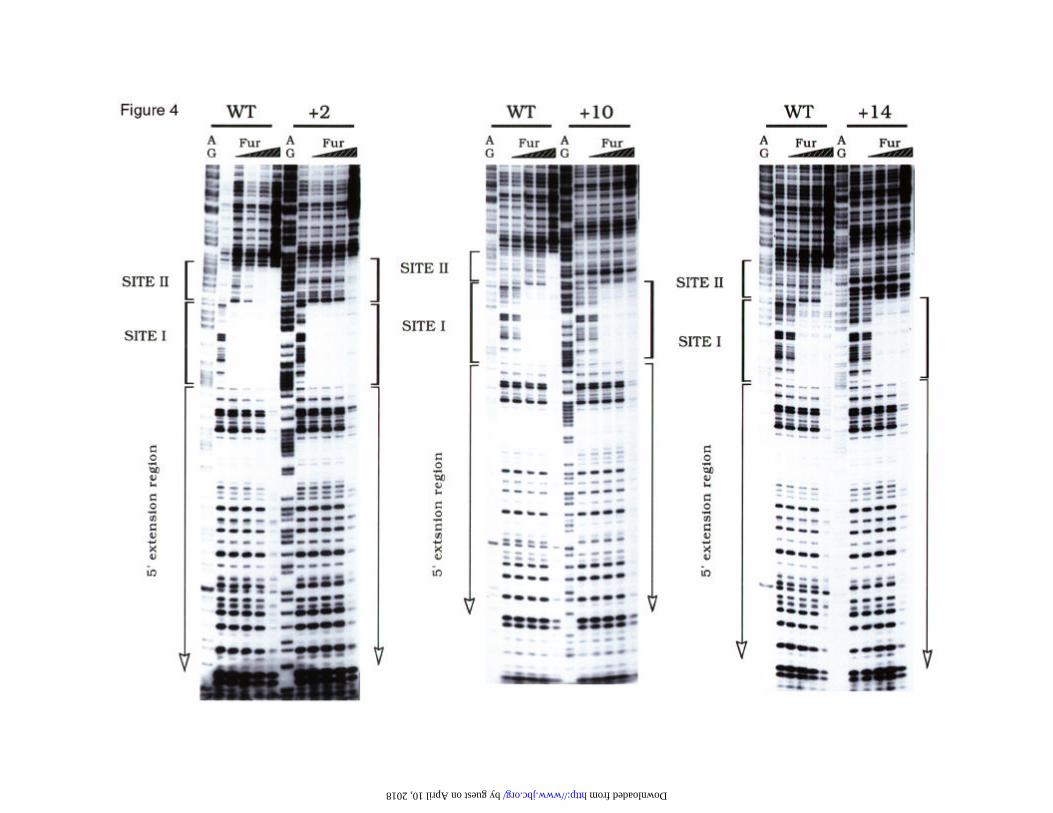
The primary Fur binding site in the Paer promoter nucleates the occupation of the adjacent
downstream sequence- Since the 50 bp sequence of the Paer promoter spanning DNase
sites I and II interacts invariably with the Fur protein with the 6-bp periodicity
discussed above, we first addressed whether such sites are independent (as they
should be by the 19 bp consensus criterium) or site II is a sequence-dependent
extension of site I. Such secondary sites are protected in most E. coli Fur-regulated
promoters, although the sequences can be very variable (24, 28, 29, 14). To address
this issue, we engineered a ClaI site next to the Fur box of site I (primary binding site
of the protein). This site was employed to insert extra bases that changed the relative
orientation of the downstream site II by 2, 10 or 14 bp (Fig. 2B). The expected result
of such insertions was to either offset moderately the two target sequences (+2), or to
separate them but keeping the phase of the DNA helix (+10) or to entirely offset and
separate sites I and II (+14). The resulting promoter variants were then footprinted
with DNase I in the presence of growing concentrations of Fur-Mn2+, with the results
shown in Fig. 4.
The effect of +2 bp insertion (Fig. 4) between the sites was relatively minor, since it
entered only a small change in the extension and strength of the occupation of site II.
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
The first 31 bp (site I) were protected to the same extension and the same protein
concentration than the wild type promoter. However, occupation of site II (which
was displaced further downstream: upwards in Fig. 4) required a significantly higher
Fur-Mn2+ level. Interestingly, the protection of the 5' extension region occurred at the
very same protein concentration than in the wild type promoter, thus suggesting that
such an extension is entirely independent of the presence of protein bound to site II.
Although these results indicated that occupation of site II is co-operative with that of
site I, they do not rule by themselves that both sites are indeed independent. This
issue, however, was unequivocally ascertained by the results of the +10 and +14
promoters. Regardless of the maintenance of the DNA helix phase (+10) or its full
disruption (+14), the increased distance between sites I and II resulted in the inability
of the downstream site to bind any protein. In both cases, the protection of site I was
in all comparable to the wild type Paer promoter. These data favour the notion that
site II is an extension of site I rather than a separate target sequence. As in the case of
the +2 template, the separation of the sites by longer insertions did not affect at all the
upstream 5' extensions, which were detected to the same extent and apparent
intensity than in the wild type promoter.
Extensive binding of Fur to the DNA adjacent to the primary binding site in the aerobactin
promoter is sequence-specific- The results above gave a preliminary hint on whether the
lateral enlargements of the protection caused by Fur on most iron-regulated
promoters of E. coli (24, 28, 29, 14), is sequence-specific or they just reflect an
artifactual occupation caused by a high protein concentration in vitro. This is a
reasonable doubt, since such prolongation not always matches the 19 bp consensus
Fur box (24, 28, 29, 14). The data of Fig. 4 show that not any sequence adjacent to site I
within the aerobactin promoter is suitable to become protected by a high
concentration of the repressor. Furthermore, extensions require a certain frame and
distance in respect to the primary site. However, the secondary site does include a
sequence stretch similar to the 19 bp consensus, thereby suggesting that frame,
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
distance and specific sequence are all necessary for the enlargement towards the site
II.
This is, however, less clear at the third protected region, where the extension
phenomenon is far more dramatic that in site II (Fig. 3). In this case, the upstream
protection observed spans 60 additional base pairs. Although such a protection has a
clear directionality and defined boundaries, the sequence involved does not show
any significant homology with the reference 19 bp Fur consensus box (26). It is thus
conceivable that such a massive protection is not specific and therefore irrelevant to
understand the metalloregulation of the promoter. To ascertain this question, we
simply prepared a new DNA template (Fig. 2A) in which we faithfully replaced the
upstream DNA by an unrelated sequence. The substitution was such that a DNA
fragment of identical size than that bearing the wild type Paer promoter could be
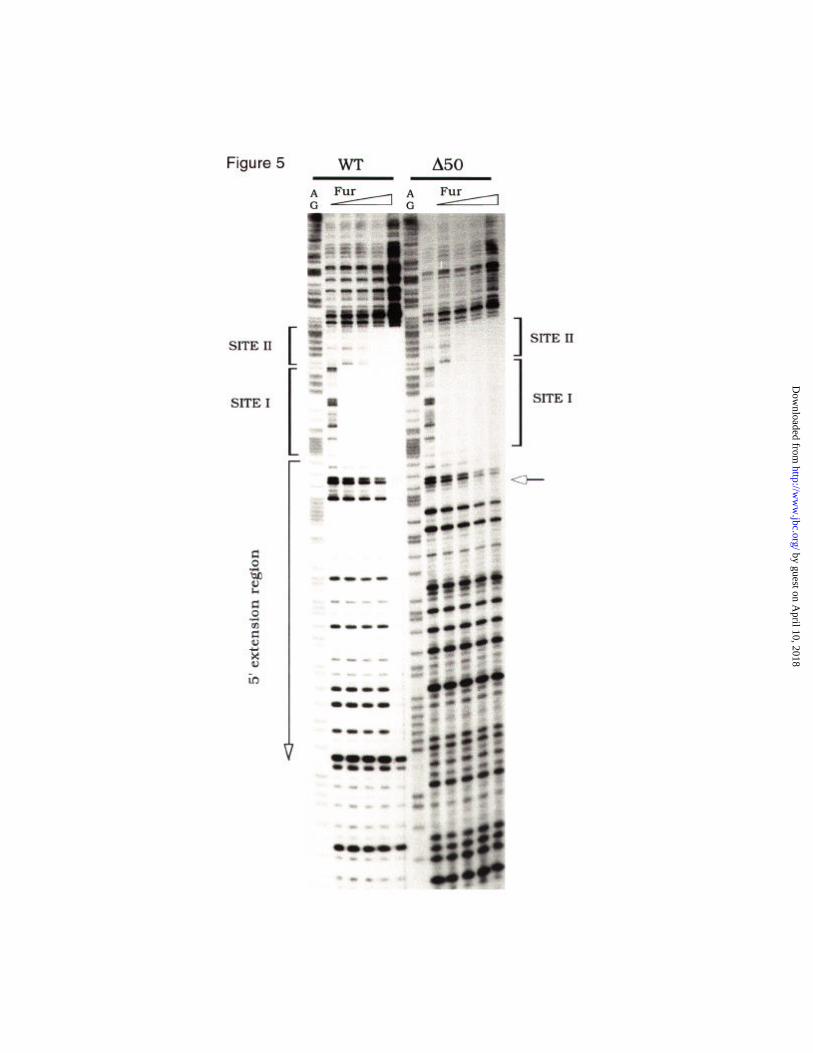
examined in parallel in DNase I footprinting assays. The results shown in Fig. 5
indicated that the unrelated sequence failed to bear any visible extension of the
footprint, even at the higher protein concentrations. Furthermore, the 5' boundary of
the protection was located exactly in the point were the heterologous sequence
started (marked with an arrow in Fig. 5). We thus conclude that the binding of Fur to
the third region is indeed sequence-dependent. We argue below that this cannot be
explained with the generally accepted 19 bp consensus model, but it is perfectly
compatible with the notion that a shorter 5'NATA/TAT3´ motif is the basic unit of Fur
binding.
Reinterpretation of the Fur operator within the aerobactin promoter- The data presented in
this work support the hypothesis (33) that Fur binding sites do not follow the
standard palindromic organisation of target sequences for regulators in prokaryotic
promoters (1). Instead, Fur operators of different extensions can be formed by
addition, in any orientation, of a minimum of 3 NATA/TAT hexamers. While this was
shown to be true for artificially assembled NATA/TAT multimers (33), this report
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
shows that this is the case also for a natural promoter such as that of the aerobactin
operon, whose extended binding sites for the repressor cannot be easily explained
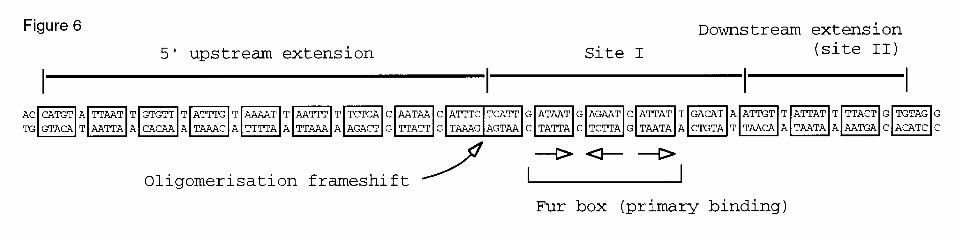
with the generally accepted 19 bp consensus model. Fig. 6 shows a reinterpretation of
the pattern of Fur-DNA interactions in the promoter as the result of a single, enlarged
operator that is formed by additions of up to 18 boxes whose frame give a maximum
match to the reference ATA/TAT pentamer. Such boxes are separated in all cases by
one intervening extra base. The one exception is at the boundary between the site I
and the protected upstream region, which lacks such an additional base, a fact that is
faithfully reflected in the hydroxyl radical footprint of the region (Fig. 3). It seems
that either the deletion of 1 base or the addition of two bases between boxes (as in the
artificial promoter +2, see Fig. 4) flaws the co-operative occupation of adjacent
hexamers, but does not inhibit it. In fact, it is revealing that such a naturally existing
deletion between site I and the upstream extension is required to frame maximally
the further upstream sequence to the reference NATA/TAT motif. But how does this
hypothesis equate the actual data?
The sequence that is protected by the lowest concentrations of Fur-Mn2+ includes 31
bp and, according to the hydroxyl radical footprinting of Fig. 3, consists of a whole of
five adjacent hexamers, three of them with a nearly perfect match to NATA/TAT.
The side repeats contain less conserved T residues and thus their occupation requires
a higher repressor concentration, what establishes the pause in the protection that is
clearly revealed by DNase I footprint (Fig. 3) and which defines site II. Such second
site would include three additional repeats. This extension certainly requires protein-
protein interactions with the repressor already bound to site I, in order to compensate
the divergence in the sequence. In fact, some hexamers have only a limited match
with the consensus. Thus, the downstream sequence may not bind by itself to the Fur-
Mn2+ complex, but it does in the context of the whole promoter. Finally, the long
upstream extension can also be sorted out as an array of adjacent Fur-binding
hexamers frame shifted by one base in respect to the sequence of boxes included in
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
sites I and II. While such a shift may explain the lower affinity, the hydroxyl radical
data of Fig. 3 shows that the shift resettles the pattern of protein-DNA interactions to
the maximum match with the NATA/TAT array. It thus appears that the sub optimal
alignment with the primary sites and the considerable sequence divergence of the
upstream region is balanced by a higher number of boxes which, as a result, produce
an unusually long operator.
Conclusion- Although not to the same dramatic extent than the aerobactin system,
many if not all iron-regulated promoters of E. coli (24, 36, 28, 29, 22, 14) contain Fur
target sequences that spread beyond the core iron box. No natural Fur binding sites
have been found to give less than a 31 bp footprint with DNase I, although the
minimal operator is only 19 bp. It thus looks likely that such adjacent sequences are
not casual but are indeed arrayed in a configuration of various 6 bp repeats with a
potential to interact specifically with the Fur protein as a whole. Extended sites might
tolerate a degree of divergence in the sequences involved, which could be
compensated by the higher overall affinity. These additional contacts might
strengthen the overall binding of the DNA segment to the regulator and do explain
why the protection is not limited to the consensus Fur box. The 6 bp box criteria
accounts for the variability and extension of the sequences protected by Fur in most
iron-regulated promoters and is also compatible with the relatively high amount of
Fur molecules (approx. 5000) found inside the cell (41, 16). The published DNase I
footprinting assays on several promoters (24, 28, 29, 14) can be consistently
reinterpreted as arrays of hexameric sequences akin to those of the aerobactin
promoter, in which the key T residues are conserved to various degrees. This mode of
Fur-DNA interaction, in which new Fur molecules must necessarily bind adjacent
hexamers through side-by-side oligomerisation, explains the gradual physiological
response observed in Fe2+-responsive systems, since it would make possible an entire
range of repression levels of iron-controlled promoters (12). The affinity for specific
promoters would vary depending on the number of repeats present on each operator
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
and the conservation of their sequences, thus generating a hierarchy of transcriptional
responses depending on small changes in the iron status of the cell. Such an ability of
Fur to control promoters through extensive DNA-protein interactions makes this
protein to be mechanistically closer to general regulators than to specific
transcriptional factors. In fact, since Fur is a Zn-containing protein (8) it is curious
that the type of DNA-protein interactions reported here have certain reminiscence to
the occupation of adjacent DNA sites by individual Zn fingers within eukaryotic
transcription factors such as TFIIIA (3, 37).
Acknowledgement- This work was supported by Contracts BIO4-CT97-2040 and QLRT-1999-00041 of the EU
and by Grant BIO98-0808 of the Comisión Interministerial de Ciencia y Tecnología. L. E.. was the recipient of a
Fellowship of Fundación Ramón Areces.
REFERENCES
1. Ptashne, M. (1992). A genetic switch. Cell Press and Blackwell Scientific Publications.
Cambridge, MA.
2. Azam, T.A. and Ishihama, A. (1999) Proc. Natl. Acad. Sci. USA 274, 33105-33113.
3. Wuttke, D. S., Foster, M. P., Case, D. A., Gottesfeld, J. M., Wright, P. E. (1997) J.
Mol. Biol. 273, 183-206
4. Bagg, A. and J. B. Neilands (1985) J. Bacteriol. 161, 450-453.
5. Hantke, K. (1984) Mol. Gen. Genet. 197, 337-341.
6. Saito, I., Wormald, M. R. and R. J. P. Williams (1991) Eur. J. Biochem. 197, 29-38.
7. Wee, S., Neilands, J. B., Bittner, M. L., Hemming, B. C., Haymore, B. L. and R.
Seetharam (1988) Biol. Metals 1, 62-68.
8. Althaus, E. W., Outten, C. E., Olson, K. E., Cao, H., and O'Halloran, T. V. (1999)
Biochemistry 38, 6559-6569
9. Bagg, A. and J. B. Neilands (1987) Microbiol. Rev. 51, 509-518.
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
10. Earhart, C. F. (1996) In Escherichia coli and Salmonella, Cellular and Molecular
Biology, pp 1075-1090. 2nd Ed. ASM Press, Washington, D. C.
11. Escolar, L., de Lorenzo, V. and Pérez-Martín, J. (1997) Mol. Microbiol. 26, 799-808.
12. Escolar, L., Pérez-Martín, J. and de Lorenzo, V. (1999) J. Bacteriol. 181, 6223-6229.
13. Litwin, M. and Calderwood, S. (1993)Clin. Microbiol. Rev. 6, 137-149.
14. Tardat, B. and Touati, D. (1993) Mol. Microbiol. 9, 53-63.
15. Zheng, M., Doan, B., Schneider, T. D. and Storz, G. (1999) J. Bacteriol. 181, 4639-
4643.
16. Hall, H. K. and J. W. Foster (1996) J. Bacteriol. 178, 5683-5691.
17. Karjalainen, T. K., Evans, D. G., Evans, D. J., Graham Jr., D. Y. and Lee, C.H.
(1991) Microb. Pathog. 11, 317-323.
18. Hantke, K. (1987) Mol. Gen. Genet. 210, 135-139.
19. Makemson, J. C. and Hastings, J. (1982) Curr. Microbiol. 7, 181-186.
20. McCarter, L. and Silverman, M. (1989) J. Bacteriol. 171, 731-736.
21. Ochsner, U.A. and Vasil, M. L. (1996). Proc. Natl. Acad. Sci. USA. 93, 4409-4414.
22. Stojiljkovic, I., Bäumer, A. J. and Hantke, K. (1994) J. Mol. Biol. 236, 531-545.
23. Tsolis, R.M., Bäumler, A.J., Stojiljkovic, I. and Heffron, F. (1995) J. Bacteriol. 177,
4628-4637.
24. Brickman, T. J., Ozenberger, B. A. and McIntosh, M. A. (1990) J. Mol. Biol. 212,
669-682.
25. Chai, S., Welch, T. J. and Crosa, J.H. (1998) J. Biol. Chem. 273, 33841-33847.
26. de Lorenzo, V., Wee, S., Herrero, M. and Neilands, J. B. (1987) J. Bacteriol. 169,
2624-2630.
27. Desai, P. J., Angerer, A. and Genco, C.A. (1996) J. Bacteriol. 178, 5020-5023.
28. Griggs, D. W. and Konisky, J. (1989) J. Bacteriol. 171, 1048-1054.
29. Hunt, M. D., Pettis, G. S. and McIntosh, M.A. (1994) J. Bacteriol. 176, 3944-3955.
30. Watnick, P.I., Butterton, J. R. and Calderwood, S. B. (1998) Gene 209, 65-70.
31. Calderwood, S. and Mekalanos, J. J. (1988). J. Bacteriol. 170, 1015-1017.
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15
32. de Lorenzo, V., Giovannini, F., Herrero, M. and Neilands, J.B. (1988). J. Mol. Biol.
203, 875-884.
33. Escolar, L., Pérez-Martín, J. and de Lorenzo, V. (1998) J. Mol. Biol. 283, 537-547.
34. Bindereif, A. and Neilands, J. B. (1985) J. Bacteriol. 162, 1039-1046.
35. Le Cam, E., Fréchon, D., Barray, M., Fourcade, A. and Delain, E. (1994) Proc. Natl.
Acad. Sci. USA 91, 11816-11820.
36. Fréchon, D. and Le Cam, E. (1994) Biochem. Biophys. Res. Commun. 201, 346-355.
37. Pizzi, S., Dieci, G., Frigeri, P., Piccoli, G., Stocchi, V. and Ottonello, S. (1999) J.
Biol. Chem. 274, 2539-4838.
38. Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecular cloning : A laboratory
manual. (Cold Spring Harbor Laboratory. Cold Spring Harbor, NY).
39. Kunkel, T. A., Roberts, J. D. and Zakour, R.A. (1987) Meth. Enzymol. 154, 367-382.
40. Maxam, A. M. and Gilbert, W. (1980). Meth. Enzymol. 65, 499-560.
41. Watnick, P. I., Eto, T., Takahashi, H. and Calderwood, S.B. (1997) J. Bacteriol. 179,
243-247.
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
LEGENDS TO THE FIGURES
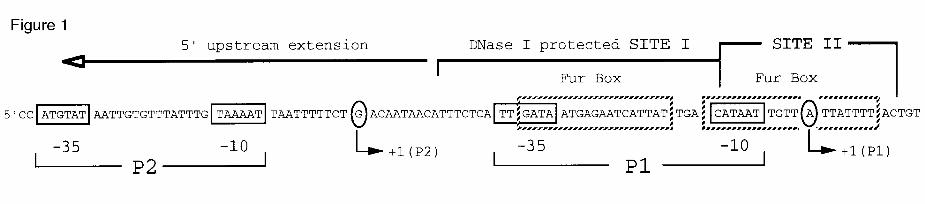
Fig. 1. Organisation of the aerobactin promoter region. The overall arrangement of
functional elements within the DNA segment placed at 5' in respect to the aerobactin
gene cluster is shown. The promoter region includes two -10/-35 hexamers, that
define promoters P1 (proximal) and P2 (distal). The primary target DNA sequences
for the Fur protein (sites I and II) and the upstream extension are pointed as defined
by DNase I footprinting (32 and Fig. 2), with an indication of the two segments with a
maximal coincidence with the 19 bp consensus Fur binding sequences
(5'GATAATGATAATCATTATC3', Fur boxes). The transcription start sites of each of
the promoters is indicated as well.
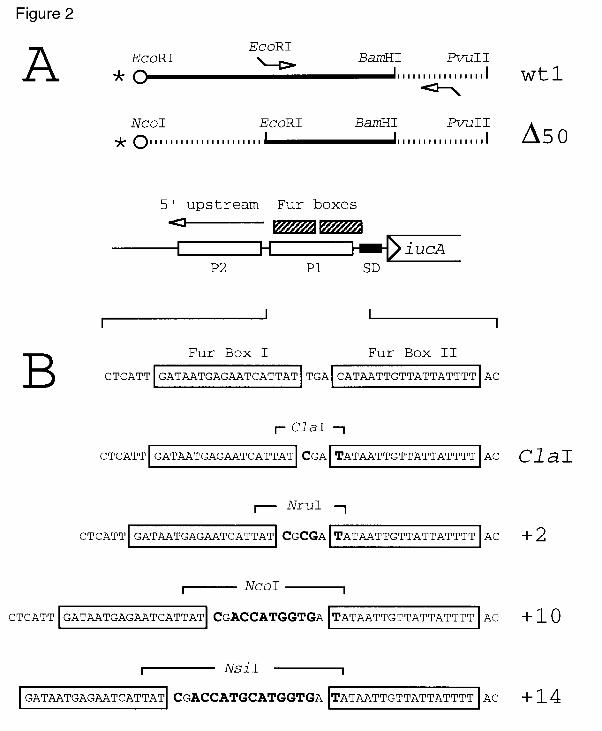
Fig. 2. Paer promoter variants used as templates for DNA footprinting analysis.
(A) The fragment wt1 is a 368 bp EcoRI-PvuII segment from plasmid pUC-LE15
spanning positions -128 to +32 of the aerobactin promoter region as an EcoRI-BamHI
plus a vector-born unrelated BamHI-PvuII extension. The promoter variant termed
∆50 was created by amplifying the sequence -50 to +32 as an EcoRI and BamHI
fragment, combining it with an NcoI-EcoRI extension of identical size of that of the
wt1 segment and cloning the whole in pUC19 (see text for explanation). The NcoI-
PvuII segment present in the resulting plasmid has its NcoI end located at exactly the
same distance to the Fur boxes than the EcoRI site of the wt1 fragment, thus allowing
a faithful compararison of its footprint with the wild-type fragment. The relative
position of each fragment in respect to the functional motifs of the aerobactin
promoter (led by the iucA gene) are indicated below, as well as the location of the
radioactive label (*) in the DNA fragments assayed. (B) Modified variants of the
aerobactin promoter with increasing distance between the Fur boxes I and II. The
boundary between the two boxes was entered with a novel ClaI site, which was
further employed for addition of 2, 10 or 14 bp (new bases in bold) as explained in the
Experimental procedures section. The mutated segments were cloned back to pUC19
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17
and used as the source of the end-labelled EcoRI-PvuII restriction fragments
employed in the footprints.
Fig. 3. Comparison of the DNase I and hydroxyl radical footprinting caused by the
Fur-Mn2+ complex on the wt aerobactin promoter. The wt1 DNA fragment (Fig.
2A), end-labelled at its EcoRI end was preincubated for 5 min at 37º C with increasing
amounts of the Fur protein (monomer): 0, 15, 35, 70, 150, 200, 250 y 350 nM. After
treatment with DNase I or hydroxyl radicals, the mixtures were processed as
previously described (32). Size markers from a sequencing reaction were loaded to
the right lane to identify the extent of the footprinted sequences. The boundaries of
the three regions sequentially protected by Fur are indicated.
Fig. 4. DNase I footprinting analysis of Fur-Mn2+ on Paer variants with increasing
distances between Fur boxes. The wild type promoter was compared in each gel
with its derivatives added with 2, 10 or 14 bases (see sketches in Fig. 2B). The
introduction of the ClaI site did not affect Fur binding (not shown). The end-labelled
restriction fragments were preincubated for 5 min at 37º C with Fur protein
(monomer) concentrations of 30, 60, 120 and 240 nM. After DNase I treatment the
reactions were processed as described in (26). A+G reactions (40) made on the same
fragments were loaded in parallel with each sample. The limits of the three sequential
regions protected by Fur are indicated.
Fig. 5. The extended binding of Fur-Mn2+ to the upstream promoter region is
sequence-dependent. The gel compares the ability of Fur-Mn2+ to bind to the wt
aerobactin promoter and to an equivalent variant (∆50, Fig. 2A) in which the region
upstream of the Fur site I has been replaced by an unrelated sequence of the same
size. Both wt1 and ∆50 fragments were preincubated for 5 min at 37º C with
increasing amounts of the Fur protein (monomer): 30, 60, 120 and 240 nM and
processed as before (32). A+G reactions (40) were carried out with the same labelled
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

18
DNA fragment and loaded onto the gels together with the treated samples. The start
of the substitution is indicated with an arrow to the right of the Figure. Note normal
occupation of sites I and II, but total lack of upstream extension in the ∆50 template.
Fig. 6. Reinterpretation of the Fur-protected sites in the aerobactin promoter. The
Figure shows the array of NATA/TAT hexamers at the Paer promoter region that
account for the data presented in this work. The boundaries of the primary and
secondary operators defined with DNase I footprint (sites I and sites II, respectively),
as well as the extension towards adjacent upstream sequences (e.g., the
polimerisation region), are indicated. The sequences are boxed in hexamers on the
basis of maximal similarity to the proposed minimal unit of interaction, which is
coincident with the pattern found in the OH radical assays (Fig. 3). Note the unique
frameshift of the array at the boundary between site I and the upstream extension.
The location and orientation of the 3 hexamers that determine the primary binding of
the repressor to the whole promoter and nucleates the subsequent upstream and
downstream extensions are pointed (for a discussion on the orientation of the
hexamers, see reference 33.
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Lucia Escolar, Jose Perez-Martin and Victor de Lorenzoaerobactin promoter of Escherichia coli
Evidence of an unusually long operator for the Fur (ferric uptake) repressor in the
published online May 31, 2000J. Biol. Chem.
10.1074/jbc.M002839200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from