receptortyrosinekinaseror2mediateswnt5a-induced ... ·...
TRANSCRIPT

Receptor Tyrosine Kinase Ror2 Mediates Wnt5a-inducedPolarized Cell Migration by Activating c-Jun N-terminalKinase via Actin-binding Protein Filamin A*□S
Received for publication, March 25, 2008, and in revised form, July 23, 2008 Published, JBC Papers in Press, July 30, 2008, DOI 10.1074/jbc.M802325200
Akira Nomachi‡1, Michiru Nishita‡2, Daisuke Inaba‡, Masahiro Enomoto‡§, Mayumi Hamasaki‡,and Yasuhiro Minami‡3
From the Departments of ‡Physiology and Cell Biology and §Pediatrics, Graduate School of Medicine, Kobe University, 7-5-1,Kusunoki-cho, Chuo-ku, Kobe 650-0017, Japan
The receptor tyrosine kinase Ror2 has recently been shown toact as an alternative receptor or coreceptor for Wnt5a and tomediate Wnt5a-induced migration of cultured cells. However,little is known about the molecular mechanism underlying thismigratory process. Here we show by wound-healing assays thatRor2plays critical roles inWnt5a-induced cellmigrationby reg-ulating formation of lamellipodia and reorientation of microtu-bule-organizing center (MTOC). Wnt5a stimulation inducesactivation of the c-JunN-terminal kinase JNKat thewound edgein a Ror2-dependent manner, and inhibiting JNK activity abro-gatesWnt5a-induced lamellipodia formation andMTOC reori-entation. Additionally, the association of Ror2 with the actin-binding protein filamin A is required for Wnt5a-induced JNKactivation and polarized cell migration. We further show thatWnt5a-induced JNK activation andMTOC reorientation can besuppressed by inhibiting PKC�. Taken together, our findingsindicate that Wnt5a/Ror2 activates JNK, through a processinvolving filamin A and PKC�, to regulate polarized cellmigration.
Ror2 belongs to the Ror family of evolutionally conservedreceptor tyrosine kinases (1) and acts as an alternative or core-ceptor for Wnt5a, a representative noncanonical Wnt protein(2–4). Duringmouse development, Ror2 plays essential roles indevelopmentalmorphogenesis (5, 6) and is expressed in variouscell types that display extensive migratory activities, includingneural crest-derived cells and mesenchymal cells (7). Loss- orgain-of-function analyses inmice,Xenopus laevis andCaenorh-abditis elegans reveal that, like Wnt5a, Ror2 and/or Ror2orthologs are required for convergent extension movements
and for polarity andmigration of several cell types during devel-opment (4–6, 8, 9).It has been proposed that Ror2 mediatesWnt5a signaling by
activating the Wnt-c-Jun N-terminal kinase (JNK)4 pathway,which regulates convergent extension movements in Xenopusgastrulation, and/or inhibiting the �-catenin-TCF pathway (2,4, 10). Wnt5a stimulation is known to promote cell migration(11–13), which Ror2 seems to mediate through its associationwith filamin A (FLNa) (3). However, it remains largelyunknown how Ror2 and FLNa function in Wnt5a-induced cellmigration and whether JNK is involved in this process.Polarized cell migration is essential for development and
wound-healing and requires rearrangements of microtubule(MT) and actin cytoskeletons (14, 15). In directionally migrat-ing cells in wounded monolayers of cultured cells (e.g. fibro-blasts) following external stimuli (e.g. epidermal growth factorand lysophosphatidic acid), MT arrays and actin filamentsbecome polarized facing to thewound edge. In such cells, selec-tively stabilized MTs (post-translationally detyrosinated tubu-lins or Glu tubulins) orient toward the leading edge and MTorganizing center (MTOC) is reoriented to lie between thenucleus and the leading edge (16). At the leading edge, actincytoskeletons are also reorganized to form lamellipodia, gener-ating driving force for polarized migration (17). On the otherhand, it has been established that JNK is involved in wound-healing in Drosophila (18, 19). Erk and p38 as well as JNK havealso been implicated in cell migration during wound closure offibroblastic and/or epithelial cells (20–24). Although JNK isactivated following Wnt5a stimulation of cultured cells (4, 25),its role in polarized cell migration remain elusive. Furthermore,it has recently been reported that Wnt5a-induced signalingpathway seems to cooperate with Par/aPKC pathway to medi-ate polarized reorganization of the microtubule cytoskeletonduring a wound response (26), but it is unclear how JNK caninteract functionally with components of the Par/aPKC path-way.We show by using in vitro wound-healing assays that Ror2
mediates polarized cell migration during Wnt5a-induced
* This work was supported in part by a Grant-in-Aid for Scientific Research inPriority Areas, a Grant-in-Aid for Scientific Research (B), and a Grant-in-Aidfor Young Scientists (B) from the Ministry of Education, Culture, Sports,Science, and Technology, Japan, the Uehara Memorial Foundation, theNaito Foundation, the Hyogo Science and Technology Association, andthe Astellas Foundation for Research on Metabolic Disorders. The costs ofpublication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked “advertisement” inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Fig. S1.
1 A research fellow of the Japan Society for the Promotion of Sciences.2 To whom correspondence may be addressed. Tel.: 81-78-382-5561; Fax:
81-78-382-5579; E-mail: [email protected] To whom correspondence may be addressed. Tel.: 81-78-382-5560; Fax:
81-78-382-5579; E-mail: [email protected].
4 The abbreviations used are: JNK, c-Jun N-terminal kinase; MTOC, microtu-bule-organizing center; FLNa, filamin A; GFP, green fluorescent protein;DMEM, Dulbecco’s modified Eagle’s medium; FBS, fetal bovine serum; WT,wild type; siRNA, short interference RNA; PKC, protein kinase C; MAP, mito-gen-activated protein; FLNa, filamin A; DAPI, 4�,6-diamidino-2-phenylin-dole; BrdU, bromodeoxyuridine.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 41, pp. 27973–27981, October 10, 2008© 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
OCTOBER 10, 2008 • VOLUME 283 • NUMBER 41 JOURNAL OF BIOLOGICAL CHEMISTRY 27973
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

wound closure by regulating lamellipodia formation andMTOC reorientation in migrating cells at the wound edge. Infact, these cellular events can be impaired by suppressedexpression of Ror2. Furthermore, JNK is activated at thewoundedge, in a Ror2-dependent manner, following Wnt5a stimula-tion, and inhibiting JNK activity abrogates Wnt5a-inducedlamellipodia formation and MTOC reorientation. We alsoshow that FLNa and its interaction with Ror2 are required forWnt5a-induced JNK activation and polarized cell migration.Moreover, we show that Wnt5a-induced JNK activation andMTOC reorientation can be suppressed by inhibiting PKC�,suggesting a possible functional link betweenWnt5a/Ror2/JNKandPar/aPKCpathways. Collectively, these results provide newinsights into the mechanism by which Wnt5a/Ror2 regulatesJNK activity, through a process involving FLNa and PKC�, forpolarized cell migration.
EXPERIMENTAL PROCEDURES
Plasmids, Antibodies, and Reagents—The cDNAs for Myc-FLNa and its �20mutant, isolated by restriction enzyme diges-tion from pMYC-C1 (3), were cloned into the retroviral vectorpMXs-puro (a gift from T. Kitamura). An anti-Ror2 antibodywas prepared as described (27). Antibodies against phospho-JNK (Thr-183/Tyr-185, Cell Signaling), JNK (FL, Santa CruzBiotechnology), phospho-p38 (Thr-180/Tyr-132, Cell Signal-ing), p38 (Cell Signaling), �-tubulin (Ab-1, Calbiochem), �-ac-tin (Sigma), Glu-tubulin (AB3201, Chemicon), �-tubulin(GTU-88, Sigma), phospho-c-Jun (KM-1, Santa Cruz Biotech-nology), phospho-ERK (Thr-202/Tyr-204, Cell Signaling), ERK(K-23, Santa Cruz Biotechnology), FLNa (FLMN01, AbCam),PKC� (C-20, Santa Cruz Biotechnology), bromodeoxyuridine(BrdU) (3D4, BD Biosciences), GFP (JL-8, Clontech), and Myc(A-14, Santa Cruz Biotechnology) were purchased commer-cially. SP600125, JNK inhibitor I (cell-permeable peptide con-sisting of HIV-TAT48–57 and 20-amino acid JNK bindingdomain of JIP1with two proline residues as spacer), and controlpeptide for JNK inhibitor I (cell-permeable peptide consistingof HIV-TAT48–57 with two proline residues) were purchasedfromCalbiochem. Anisomycin, actinomycin D, and BrdUwerefrom Sigma. Mouse-purified Wnt5a was obtained from R&DSystems. Cell-permeable pseudo-substrates for PKC�/�(Biomol), PKC�, PKC�, and PKC� (BIOSOURCE) were used.TheRor2 siRNA (1) (28),FLNa siRNA (29), control, nonspecificsiRNA (GUACCGCACGUCAUUCGUAUC) were purchasedfrom RNAi Co., Ltd. The Ror2 siRNA (2) (AAGACGAA-GUGGCAGAAGGAUGGGA) and RNAi Negative ControlDuplexes (Medium GC Duplex 2) were purchased fromInvitrogen. Conditionedmedia (CM)were harvested from con-fluent monolayers ofWnt5a/L or neo/L cells (30) that had beencultured in Dulbecco modified Eagle’s medium (DMEM) con-taining 5% fetal bovine serum (FBS). The CM were diluted inFBS-free DMEM (2:5�1:10) prior to use for cytoskeletal andcell motility analyses.Cell Culture, Retroviral Infection, Transfection, and Wound-
ing—NIH3T3 and SaOS2 cells were cultured in DMEM con-taining 10% FBS.M2 andA7 cells weremaintained as described(31). Packaging cell line Plat-A, provided by T. Kitamura, wasmaintained inDMEMcontaining 10% FBS, 1�g/ml puromycin
(Sigma), and 10 �g/ml blasticidin (Sigma). Plat-A cells weretransfected with retroviral plasmids using FuGENE 6 (Roche).After 48 h, viral supernatant was collected and filtered through0.45-�m size pore. M2 cells were infected with the viral super-natant in the presence of 4 �g/ml polybrene (Sigma) for 24 h.After infection, M2 cells were selected with 1 �g/ml puromy-cin, and single cell cloneswere isolated and screened for expres-sion of Myc-FLNa (WT or �20) by immunoblot analysis. Twoindependent stable clones for each construct (clones 10 and 2forWTand 3 and 7 for�20) were used. For siRNA transfection,we used GeneSilencer siRNA Transfection Reagent (GeneTherapy Systems) for NIH3T3 cells and Lipofectamine 2000(Invitrogen) for SaOS2 cells. For wound-healing assays, cellswere plated on fibronectin-coated coverslips in 12-well plates,grown to become confluent for 24 h, and scratched with apipette tip once across the coverslip. The wounded cells werewashed twice with phosphate-buffered saline and then incu-bated in the presence or absence of CM or purifiedWnt5a. Forimmunoblot analyses, confluent cells on fibronectin-coated 35mm dishes were scratched eight times across the dish, andimmediately after wounding, cultured media were replacedwith CM. The rate of wound closure was calculated by relativemigrating distance between the wound edges.Immunoblotting, Cell Staining, and Image Analysis—Cells
were lysed in lysis buffer (50 mM Tris-HCl (pH 7.4), 150 mMNaCl, 0.5% Nonidet P-40, 5 mM EDTA, 50 mM NaF, 1 mMNa3VO4, 1 mM phenylmethylsulfonyl fluoride, 10 �g/ml leu-peptin, and 10 �g/ml aprotinin). Whole cell lysates were sub-jected to immunoblot analysis as described (27). For BrdUlabeling, cells were incubatedwith 1�MBrdUduring the courseof the wound-healing assay and stained with anti-BrdU anti-body according to the manufacturer’s instruction. For phalloi-din staining, cells were fixed with 3.7% paraformaldehyde inphosphate-buffered saline, treated with 0.2% Triton X-100 inphosphate-buffered saline, and stained with rhodamine-phal-loidin (Invitrogen) or Alexa Fluor 488-phalloidin (Cambrex BioScience). For immunostaining, cells were fixed at �20 °C inmethanol. Fixed cells were stained with the respective antibod-ies as described (3). Nuclei were counterstained with 4�,6-dia-midino-2-phenylindole (DAPI). Fluorescent images wereobtained using a laser scanning confocal imaging system(LSM510, Carl Zeiss MicroImaging, Inc.) and processed usingPhotoshop CS (Adobe).
RESULTS AND DISCUSSION
Wnt5a Promotes Polarized Cell Migration in a Ror2-depend-entManner—When exposed to external stimuli that induce cellmigration, cells become polarized by reorganizing microtubuleand actin cytoskeletons, resulting in reorientation of MTOCand formation of an F-actin-rich membrane protrusion towardthe direction of cell movement. Because Wnt5a is one of thestimuli that induces cell migration in various cell types, includ-ing fibroblasts (3, 11–13), we examined whether or not Wnt5acould inducemicrotubule and actin rearrangements to regulatecell polarity by performing in vitro wound-healing assays. Tothis end, confluent monolayers of NIH3T3 cells, which expressRor2 endogenously, were treated with either control (neo) con-ditioned medium (CM) or Wnt5a CM. Because serum lipid,
Role of Ror2 in Wnt5a-induced Polarized Cell Migration
27974 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 41 • OCTOBER 10, 2008
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
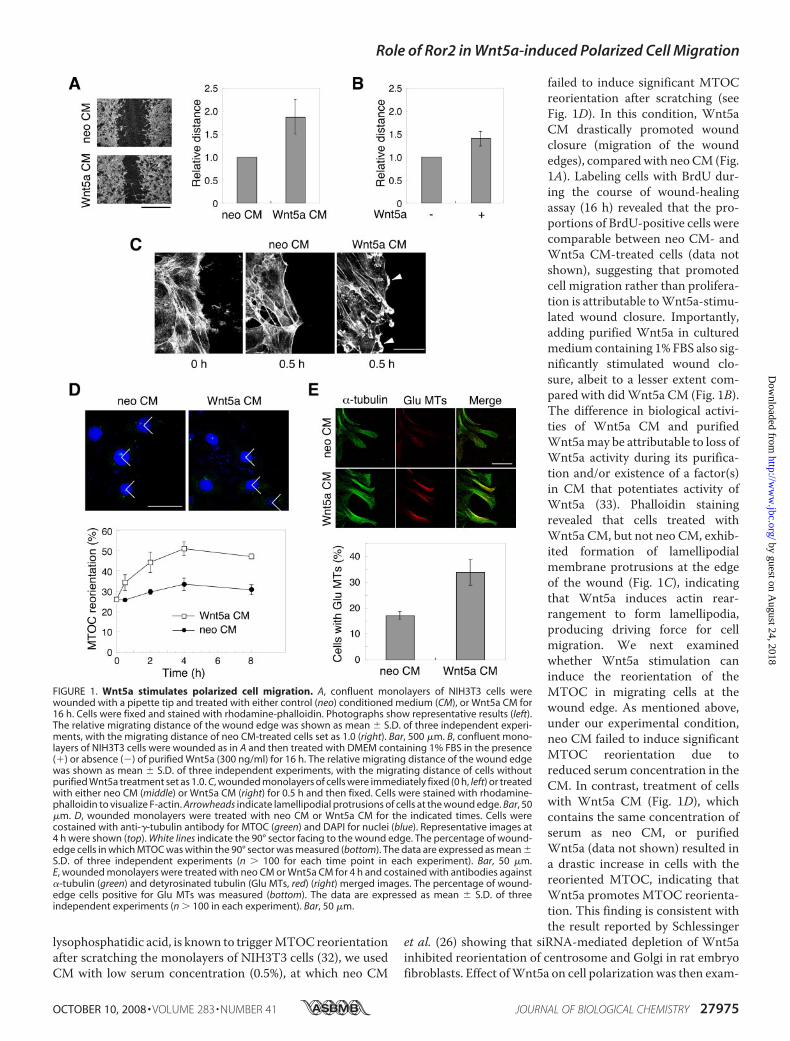
lysophosphatidic acid, is known to triggerMTOC reorientationafter scratching the monolayers of NIH3T3 cells (32), we usedCM with low serum concentration (0.5%), at which neo CM
failed to induce significant MTOCreorientation after scratching (seeFig. 1D). In this condition, Wnt5aCM drastically promoted woundclosure (migration of the woundedges), comparedwith neoCM (Fig.1A). Labeling cells with BrdU dur-ing the course of wound-healingassay (16 h) revealed that the pro-portions of BrdU-positive cells werecomparable between neo CM- andWnt5a CM-treated cells (data notshown), suggesting that promotedcell migration rather than prolifera-tion is attributable toWnt5a-stimu-lated wound closure. Importantly,adding purified Wnt5a in culturedmedium containing 1% FBS also sig-nificantly stimulated wound clo-sure, albeit to a lesser extent com-pared with didWnt5a CM (Fig. 1B).The difference in biological activi-ties of Wnt5a CM and purifiedWnt5amay be attributable to loss ofWnt5a activity during its purifica-tion and/or existence of a factor(s)in CM that potentiates activity ofWnt5a (33). Phalloidin stainingrevealed that cells treated withWnt5a CM, but not neo CM, exhib-ited formation of lamellipodialmembrane protrusions at the edgeof the wound (Fig. 1C), indicatingthat Wnt5a induces actin rear-rangement to form lamellipodia,producing driving force for cellmigration. We next examinedwhether Wnt5a stimulation caninduce the reorientation of theMTOC in migrating cells at thewound edge. As mentioned above,under our experimental condition,neo CM failed to induce significantMTOC reorientation due toreduced serum concentration in theCM. In contrast, treatment of cellswith Wnt5a CM (Fig. 1D), whichcontains the same concentration ofserum as neo CM, or purifiedWnt5a (data not shown) resulted ina drastic increase in cells with thereoriented MTOC, indicating thatWnt5a promotes MTOC reorienta-tion. This finding is consistent withthe result reported by Schlessinger
et al. (26) showing that siRNA-mediated depletion of Wnt5ainhibited reorientation of centrosome and Golgi in rat embryofibroblasts. Effect ofWnt5a on cell polarizationwas then exam-
FIGURE 1. Wnt5a stimulates polarized cell migration. A, confluent monolayers of NIH3T3 cells werewounded with a pipette tip and treated with either control (neo) conditioned medium (CM), or Wnt5a CM for16 h. Cells were fixed and stained with rhodamine-phalloidin. Photographs show representative results (left).The relative migrating distance of the wound edge was shown as mean � S.D. of three independent experi-ments, with the migrating distance of neo CM-treated cells set as 1.0 (right). Bar, 500 �m. B, confluent mono-layers of NIH3T3 cells were wounded as in A and then treated with DMEM containing 1% FBS in the presence(�) or absence (�) of purified Wnt5a (300 ng/ml) for 16 h. The relative migrating distance of the wound edgewas shown as mean � S.D. of three independent experiments, with the migrating distance of cells withoutpurified Wnt5a treatment set as 1.0. C, wounded monolayers of cells were immediately fixed (0 h, left) or treatedwith either neo CM (middle) or Wnt5a CM (right) for 0.5 h and then fixed. Cells were stained with rhodamine-phalloidin to visualize F-actin. Arrowheads indicate lamellipodial protrusions of cells at the wound edge. Bar, 50�m. D, wounded monolayers were treated with neo CM or Wnt5a CM for the indicated times. Cells werecostained with anti-�-tubulin antibody for MTOC (green) and DAPI for nuclei (blue). Representative images at4 h were shown (top). White lines indicate the 90° sector facing to the wound edge. The percentage of wound-edge cells in which MTOC was within the 90° sector was measured (bottom). The data are expressed as mean �S.D. of three independent experiments (n � 100 for each time point in each experiment). Bar, 50 �m.E, wounded monolayers were treated with neo CM or Wnt5a CM for 4 h and costained with antibodies against�-tubulin (green) and detyrosinated tubulin (Glu MTs, red) (right) merged images. The percentage of wound-edge cells positive for Glu MTs was measured (bottom). The data are expressed as mean � S.D. of threeindependent experiments (n � 100 in each experiment). Bar, 50 �m.
Role of Ror2 in Wnt5a-induced Polarized Cell Migration
OCTOBER 10, 2008 • VOLUME 283 • NUMBER 41 JOURNAL OF BIOLOGICAL CHEMISTRY 27975
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
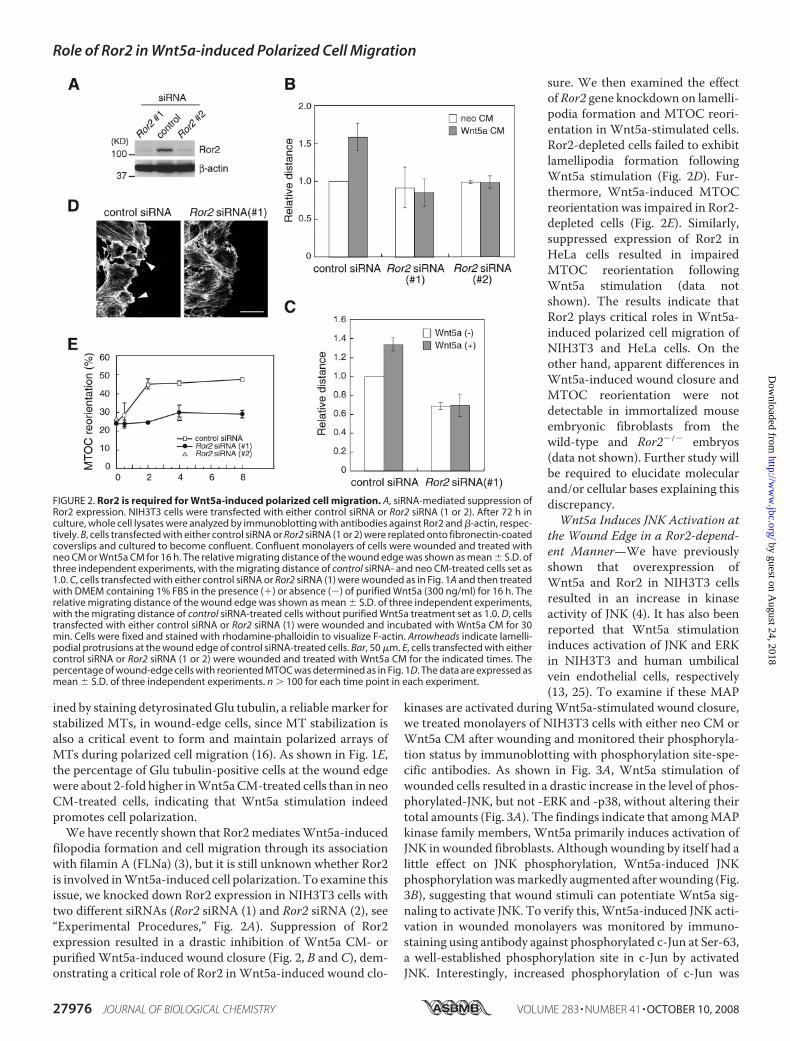
ined by staining detyrosinatedGlu tubulin, a reliablemarker forstabilized MTs, in wound-edge cells, since MT stabilization isalso a critical event to form and maintain polarized arrays ofMTs during polarized cell migration (16). As shown in Fig. 1E,the percentage of Glu tubulin-positive cells at the wound edgewere about 2-fold higher inWnt5aCM-treated cells than in neoCM-treated cells, indicating that Wnt5a stimulation indeedpromotes cell polarization.We have recently shown that Ror2mediatesWnt5a-induced
filopodia formation and cell migration through its associationwith filamin A (FLNa) (3), but it is still unknown whether Ror2is involved inWnt5a-induced cell polarization. To examine thisissue, we knocked down Ror2 expression in NIH3T3 cells withtwo different siRNAs (Ror2 siRNA (1) and Ror2 siRNA (2), see“Experimental Procedures,” Fig. 2A). Suppression of Ror2expression resulted in a drastic inhibition of Wnt5a CM- orpurifiedWnt5a-induced wound closure (Fig. 2, B and C), dem-onstrating a critical role of Ror2 inWnt5a-induced wound clo-
sure. We then examined the effectof Ror2 gene knockdown on lamelli-podia formation and MTOC reori-entation in Wnt5a-stimulated cells.Ror2-depleted cells failed to exhibitlamellipodia formation followingWnt5a stimulation (Fig. 2D). Fur-thermore, Wnt5a-induced MTOCreorientation was impaired in Ror2-depleted cells (Fig. 2E). Similarly,suppressed expression of Ror2 inHeLa cells resulted in impairedMTOC reorientation followingWnt5a stimulation (data notshown). The results indicate thatRor2 plays critical roles in Wnt5a-induced polarized cell migration ofNIH3T3 and HeLa cells. On theother hand, apparent differences inWnt5a-induced wound closure andMTOC reorientation were notdetectable in immortalized mouseembryonic fibroblasts from thewild-type and Ror2�/� embryos(data not shown). Further study willbe required to elucidate molecularand/or cellular bases explaining thisdiscrepancy.Wnt5a Induces JNK Activation at
the Wound Edge in a Ror2-depend-ent Manner—We have previouslyshown that overexpression ofWnt5a and Ror2 in NIH3T3 cellsresulted in an increase in kinaseactivity of JNK (4). It has also beenreported that Wnt5a stimulationinduces activation of JNK and ERKin NIH3T3 and human umbilicalvein endothelial cells, respectively(13, 25). To examine if these MAP
kinases are activated duringWnt5a-stimulated wound closure,we treated monolayers of NIH3T3 cells with either neo CM orWnt5a CM after wounding and monitored their phosphoryla-tion status by immunoblotting with phosphorylation site-spe-cific antibodies. As shown in Fig. 3A, Wnt5a stimulation ofwounded cells resulted in a drastic increase in the level of phos-phorylated-JNK, but not -ERK and -p38, without altering theirtotal amounts (Fig. 3A). The findings indicate that amongMAPkinase family members, Wnt5a primarily induces activation ofJNK in wounded fibroblasts. Although wounding by itself had alittle effect on JNK phosphorylation, Wnt5a-induced JNKphosphorylationwasmarkedly augmented afterwounding (Fig.3B), suggesting that wound stimuli can potentiate Wnt5a sig-naling to activate JNK. To verify this,Wnt5a-induced JNK acti-vation in wounded monolayers was monitored by immuno-staining using antibody against phosphorylated c-Jun at Ser-63,a well-established phosphorylation site in c-Jun by activatedJNK. Interestingly, increased phosphorylation of c-Jun was
FIGURE 2. Ror2 is required for Wnt5a-induced polarized cell migration. A, siRNA-mediated suppression ofRor2 expression. NIH3T3 cells were transfected with either control siRNA or Ror2 siRNA (1 or 2). After 72 h inculture, whole cell lysates were analyzed by immunoblotting with antibodies against Ror2 and �-actin, respec-tively. B, cells transfected with either control siRNA or Ror2 siRNA (1 or 2) were replated onto fibronectin-coatedcoverslips and cultured to become confluent. Confluent monolayers of cells were wounded and treated withneo CM or Wnt5a CM for 16 h. The relative migrating distance of the wound edge was shown as mean � S.D. ofthree independent experiments, with the migrating distance of control siRNA- and neo CM-treated cells set as1.0. C, cells transfected with either control siRNA or Ror2 siRNA (1) were wounded as in Fig. 1A and then treatedwith DMEM containing 1% FBS in the presence (�) or absence (�) of purified Wnt5a (300 ng/ml) for 16 h. Therelative migrating distance of the wound edge was shown as mean � S.D. of three independent experiments,with the migrating distance of control siRNA-treated cells without purified Wnt5a treatment set as 1.0. D, cellstransfected with either control siRNA or Ror2 siRNA (1) were wounded and incubated with Wnt5a CM for 30min. Cells were fixed and stained with rhodamine-phalloidin to visualize F-actin. Arrowheads indicate lamelli-podial protrusions at the wound edge of control siRNA-treated cells. Bar, 50 �m. E, cells transfected with eithercontrol siRNA or Ror2 siRNA (1 or 2) were wounded and treated with Wnt5a CM for the indicated times. Thepercentage of wound-edge cells with reoriented MTOC was determined as in Fig. 1D. The data are expressed asmean � S.D. of three independent experiments. n � 100 for each time point in each experiment.
Role of Ror2 in Wnt5a-induced Polarized Cell Migration
27976 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 41 • OCTOBER 10, 2008
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
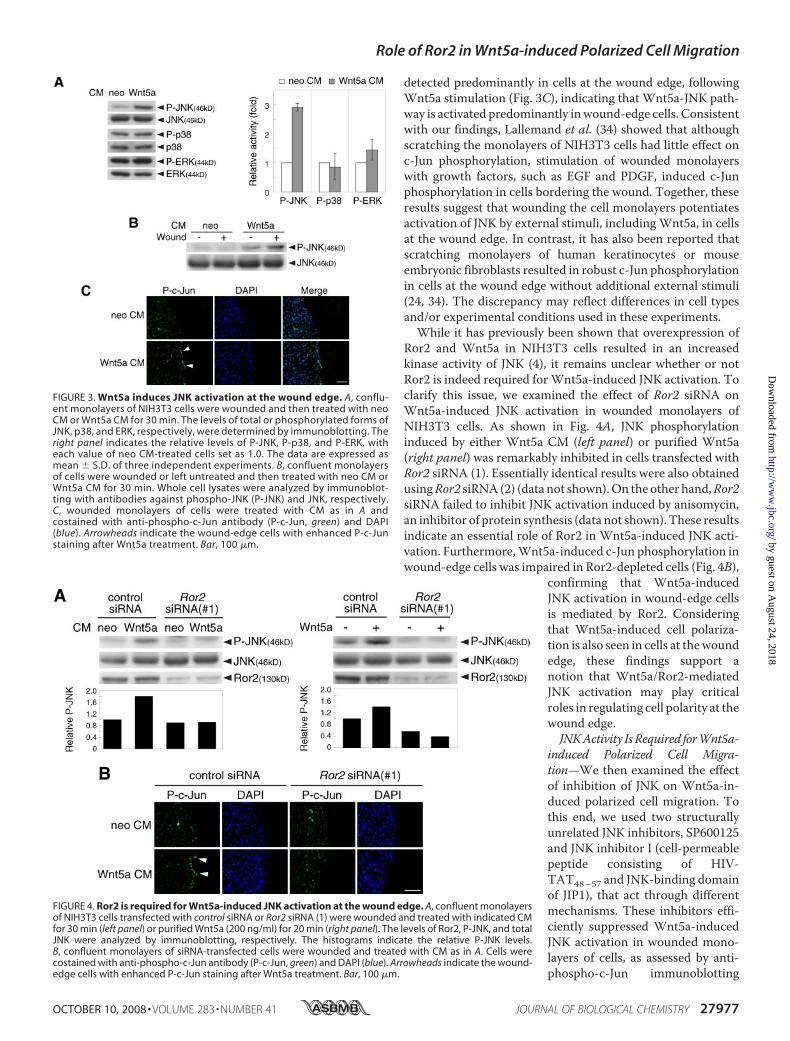
detected predominantly in cells at the wound edge, followingWnt5a stimulation (Fig. 3C), indicating that Wnt5a-JNK path-way is activated predominantly inwound-edge cells. Consistentwith our findings, Lallemand et al. (34) showed that althoughscratching the monolayers of NIH3T3 cells had little effect onc-Jun phosphorylation, stimulation of wounded monolayerswith growth factors, such as EGF and PDGF, induced c-Junphosphorylation in cells bordering the wound. Together, theseresults suggest that wounding the cell monolayers potentiatesactivation of JNK by external stimuli, includingWnt5a, in cellsat the wound edge. In contrast, it has also been reported thatscratching monolayers of human keratinocytes or mouseembryonic fibroblasts resulted in robust c-Jun phosphorylationin cells at the wound edge without additional external stimuli(24, 34). The discrepancy may reflect differences in cell typesand/or experimental conditions used in these experiments.While it has previously been shown that overexpression of
Ror2 and Wnt5a in NIH3T3 cells resulted in an increasedkinase activity of JNK (4), it remains unclear whether or notRor2 is indeed required forWnt5a-induced JNK activation. Toclarify this issue, we examined the effect of Ror2 siRNA onWnt5a-induced JNK activation in wounded monolayers ofNIH3T3 cells. As shown in Fig. 4A, JNK phosphorylationinduced by either Wnt5a CM (left panel) or purified Wnt5a(right panel) was remarkably inhibited in cells transfected withRor2 siRNA (1). Essentially identical results were also obtainedusingRor2 siRNA (2) (data not shown).On the other hand,Ror2siRNA failed to inhibit JNK activation induced by anisomycin,an inhibitor of protein synthesis (data not shown). These resultsindicate an essential role of Ror2 in Wnt5a-induced JNK acti-vation. Furthermore,Wnt5a-induced c-Jun phosphorylation inwound-edge cells was impaired in Ror2-depleted cells (Fig. 4B),
confirming that Wnt5a-inducedJNK activation in wound-edge cellsis mediated by Ror2. Consideringthat Wnt5a-induced cell polariza-tion is also seen in cells at thewoundedge, these findings support anotion that Wnt5a/Ror2-mediatedJNK activation may play criticalroles in regulating cell polarity at thewound edge.JNKActivity IsRequired forWnt5a-
induced Polarized Cell Migra-tion—We then examined the effectof inhibition of JNK on Wnt5a-in-duced polarized cell migration. Tothis end, we used two structurallyunrelated JNK inhibitors, SP600125and JNK inhibitor I (cell-permeablepeptide consisting of HIV-TAT48–57 and JNK-binding domainof JIP1), that act through differentmechanisms. These inhibitors effi-ciently suppressed Wnt5a-inducedJNK activation in wounded mono-layers of cells, as assessed by anti-phospho-c-Jun immunoblotting
FIGURE 3. Wnt5a induces JNK activation at the wound edge. A, conflu-ent monolayers of NIH3T3 cells were wounded and then treated with neoCM or Wnt5a CM for 30 min. The levels of total or phosphorylated forms ofJNK, p38, and ERK, respectively, were determined by immunoblotting. Theright panel indicates the relative levels of P-JNK, P-p38, and P-ERK, witheach value of neo CM-treated cells set as 1.0. The data are expressed asmean � S.D. of three independent experiments. B, confluent monolayersof cells were wounded or left untreated and then treated with neo CM orWnt5a CM for 30 min. Whole cell lysates were analyzed by immunoblot-ting with antibodies against phospho-JNK (P-JNK) and JNK, respectively.C, wounded monolayers of cells were treated with CM as in A andcostained with anti-phospho-c-Jun antibody (P-c-Jun, green) and DAPI(blue). Arrowheads indicate the wound-edge cells with enhanced P-c-Junstaining after Wnt5a treatment. Bar, 100 �m.
FIGURE 4. Ror2 is required for Wnt5a-induced JNK activation at the wound edge. A, confluent monolayersof NIH3T3 cells transfected with control siRNA or Ror2 siRNA (1) were wounded and treated with indicated CMfor 30 min (left panel) or purified Wnt5a (200 ng/ml) for 20 min (right panel). The levels of Ror2, P-JNK, and totalJNK were analyzed by immunoblotting, respectively. The histograms indicate the relative P-JNK levels.B, confluent monolayers of siRNA-transfected cells were wounded and treated with CM as in A. Cells werecostained with anti-phospho-c-Jun antibody (P-c-Jun, green) and DAPI (blue). Arrowheads indicate the wound-edge cells with enhanced P-c-Jun staining after Wnt5a treatment. Bar, 100 �m.
Role of Ror2 in Wnt5a-induced Polarized Cell Migration
OCTOBER 10, 2008 • VOLUME 283 • NUMBER 41 JOURNAL OF BIOLOGICAL CHEMISTRY 27977
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
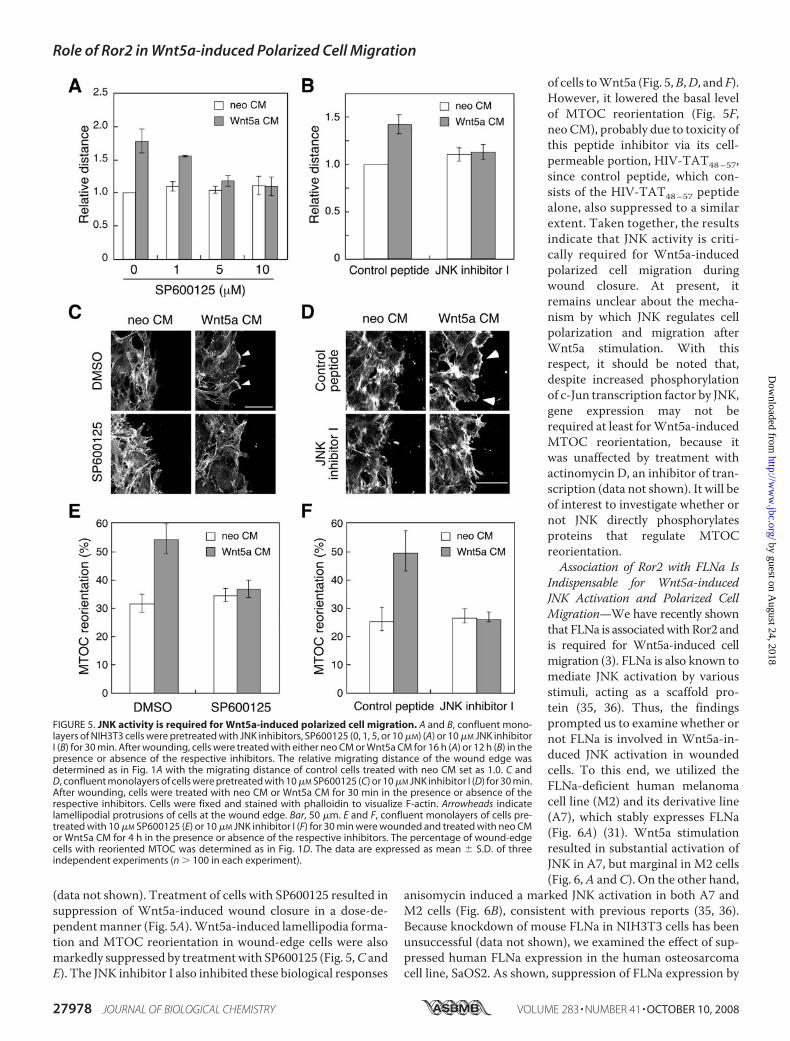
(data not shown). Treatment of cells with SP600125 resulted insuppression of Wnt5a-induced wound closure in a dose-de-pendentmanner (Fig. 5A).Wnt5a-induced lamellipodia forma-tion and MTOC reorientation in wound-edge cells were alsomarkedly suppressed by treatmentwith SP600125 (Fig. 5,C andE). The JNK inhibitor I also inhibited these biological responses
of cells toWnt5a (Fig. 5,B,D, andF).However, it lowered the basal levelof MTOC reorientation (Fig. 5F,neo CM), probably due to toxicity ofthis peptide inhibitor via its cell-permeable portion, HIV-TAT48–57,since control peptide, which con-sists of the HIV-TAT48–57 peptidealone, also suppressed to a similarextent. Taken together, the resultsindicate that JNK activity is criti-cally required for Wnt5a-inducedpolarized cell migration duringwound closure. At present, itremains unclear about the mecha-nism by which JNK regulates cellpolarization and migration afterWnt5a stimulation. With thisrespect, it should be noted that,despite increased phosphorylationof c-Jun transcription factor by JNK,gene expression may not berequired at least forWnt5a-inducedMTOC reorientation, because itwas unaffected by treatment withactinomycin D, an inhibitor of tran-scription (data not shown). It will beof interest to investigate whether ornot JNK directly phosphorylatesproteins that regulate MTOCreorientation.Association of Ror2 with FLNa Is
Indispensable for Wnt5a-inducedJNK Activation and Polarized CellMigration—We have recently shownthat FLNa is associatedwithRor2 andis required for Wnt5a-induced cellmigration (3). FLNa is also known tomediate JNK activation by variousstimuli, acting as a scaffold pro-tein (35, 36). Thus, the findingsprompted us to examine whether ornot FLNa is involved in Wnt5a-in-duced JNK activation in woundedcells. To this end, we utilized theFLNa-deficient human melanomacell line (M2) and its derivative line(A7), which stably expresses FLNa(Fig. 6A) (31). Wnt5a stimulationresulted in substantial activation ofJNK in A7, but marginal in M2 cells(Fig. 6,A andC). On the other hand,
anisomycin induced a marked JNK activation in both A7 andM2 cells (Fig. 6B), consistent with previous reports (35, 36).Because knockdown of mouse FLNa in NIH3T3 cells has beenunsuccessful (data not shown), we examined the effect of sup-pressed human FLNa expression in the human osteosarcomacell line, SaOS2. As shown, suppression of FLNa expression by
FIGURE 5. JNK activity is required for Wnt5a-induced polarized cell migration. A and B, confluent mono-layers of NIH3T3 cells were pretreated with JNK inhibitors, SP600125 (0, 1, 5, or 10 �M) (A) or 10 �M JNK inhibitorI (B) for 30 min. After wounding, cells were treated with either neo CM or Wnt5a CM for 16 h (A) or 12 h (B) in thepresence or absence of the respective inhibitors. The relative migrating distance of the wound edge wasdetermined as in Fig. 1A with the migrating distance of control cells treated with neo CM set as 1.0. C andD, confluent monolayers of cells were pretreated with 10 �M SP600125 (C) or 10 �M JNK inhibitor I (D) for 30 min.After wounding, cells were treated with neo CM or Wnt5a CM for 30 min in the presence or absence of therespective inhibitors. Cells were fixed and stained with phalloidin to visualize F-actin. Arrowheads indicatelamellipodial protrusions of cells at the wound edge. Bar, 50 �m. E and F, confluent monolayers of cells pre-treated with 10 �M SP600125 (E) or 10 �M JNK inhibitor I (F) for 30 min were wounded and treated with neo CMor Wnt5a CM for 4 h in the presence or absence of the respective inhibitors. The percentage of wound-edgecells with reoriented MTOC was determined as in Fig. 1D. The data are expressed as mean � S.D. of threeindependent experiments (n � 100 in each experiment).
Role of Ror2 in Wnt5a-induced Polarized Cell Migration
27978 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 41 • OCTOBER 10, 2008
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
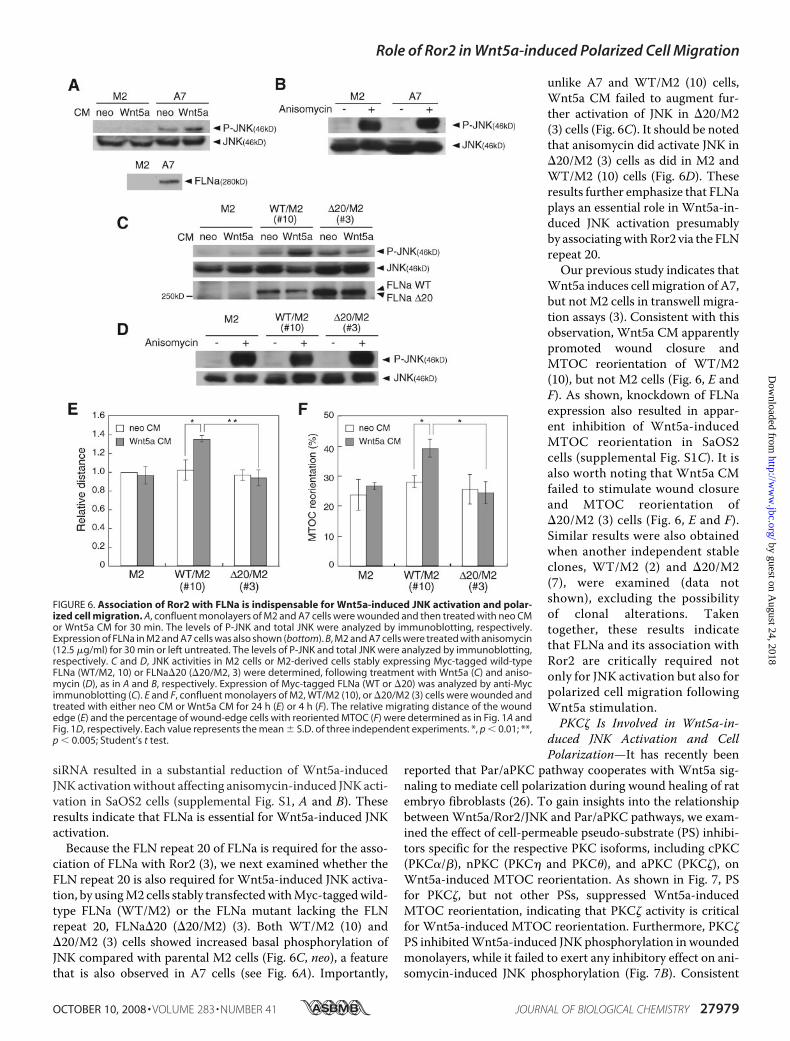
siRNA resulted in a substantial reduction of Wnt5a-inducedJNK activationwithout affecting anisomycin-induced JNK acti-vation in SaOS2 cells (supplemental Fig. S1, A and B). Theseresults indicate that FLNa is essential for Wnt5a-induced JNKactivation.Because the FLN repeat 20 of FLNa is required for the asso-
ciation of FLNa with Ror2 (3), we next examined whether theFLN repeat 20 is also required for Wnt5a-induced JNK activa-tion, by usingM2 cells stably transfectedwithMyc-taggedwild-type FLNa (WT/M2) or the FLNa mutant lacking the FLNrepeat 20, FLNa�20 (�20/M2) (3). Both WT/M2 (10) and�20/M2 (3) cells showed increased basal phosphorylation ofJNK compared with parental M2 cells (Fig. 6C, neo), a featurethat is also observed in A7 cells (see Fig. 6A). Importantly,
unlike A7 and WT/M2 (10) cells,Wnt5a CM failed to augment fur-ther activation of JNK in �20/M2(3) cells (Fig. 6C). It should be notedthat anisomycin did activate JNK in�20/M2 (3) cells as did in M2 andWT/M2 (10) cells (Fig. 6D). Theseresults further emphasize that FLNaplays an essential role in Wnt5a-in-duced JNK activation presumablyby associatingwithRor2 via the FLNrepeat 20.Our previous study indicates that
Wnt5a induces cell migration of A7,but not M2 cells in transwell migra-tion assays (3). Consistent with thisobservation, Wnt5a CM apparentlypromoted wound closure andMTOC reorientation of WT/M2(10), but not M2 cells (Fig. 6, E andF). As shown, knockdown of FLNaexpression also resulted in appar-ent inhibition of Wnt5a-inducedMTOC reorientation in SaOS2cells (supplemental Fig. S1C). It isalso worth noting that Wnt5a CMfailed to stimulate wound closureand MTOC reorientation of�20/M2 (3) cells (Fig. 6, E and F).Similar results were also obtainedwhen another independent stableclones, WT/M2 (2) and �20/M2(7), were examined (data notshown), excluding the possibilityof clonal alterations. Takentogether, these results indicatethat FLNa and its association withRor2 are critically required notonly for JNK activation but also forpolarized cell migration followingWnt5a stimulation.PKC� Is Involved in Wnt5a-in-
duced JNK Activation and CellPolarization—It has recently been
reported that Par/aPKC pathway cooperates with Wnt5a sig-naling to mediate cell polarization during wound healing of ratembryo fibroblasts (26). To gain insights into the relationshipbetween Wnt5a/Ror2/JNK and Par/aPKC pathways, we exam-ined the effect of cell-permeable pseudo-substrate (PS) inhibi-tors specific for the respective PKC isoforms, including cPKC(PKC�/�), nPKC (PKC� and PKC�), and aPKC (PKC�), onWnt5a-induced MTOC reorientation. As shown in Fig. 7, PSfor PKC�, but not other PSs, suppressed Wnt5a-inducedMTOC reorientation, indicating that PKC� activity is criticalfor Wnt5a-induced MTOC reorientation. Furthermore, PKC�PS inhibitedWnt5a-induced JNK phosphorylation in woundedmonolayers, while it failed to exert any inhibitory effect on ani-somycin-induced JNK phosphorylation (Fig. 7B). Consistent
FIGURE 6. Association of Ror2 with FLNa is indispensable for Wnt5a-induced JNK activation and polar-ized cell migration. A, confluent monolayers of M2 and A7 cells were wounded and then treated with neo CMor Wnt5a CM for 30 min. The levels of P-JNK and total JNK were analyzed by immunoblotting, respectively.Expression of FLNa in M2 and A7 cells was also shown (bottom). B, M2 and A7 cells were treated with anisomycin(12.5 �g/ml) for 30 min or left untreated. The levels of P-JNK and total JNK were analyzed by immunoblotting,respectively. C and D, JNK activities in M2 cells or M2-derived cells stably expressing Myc-tagged wild-typeFLNa (WT/M2, 10) or FLNa�20 (�20/M2, 3) were determined, following treatment with Wnt5a (C) and aniso-mycin (D), as in A and B, respectively. Expression of Myc-tagged FLNa (WT or �20) was analyzed by anti-Mycimmunoblotting (C). E and F, confluent monolayers of M2, WT/M2 (10), or �20/M2 (3) cells were wounded andtreated with either neo CM or Wnt5a CM for 24 h (E) or 4 h (F). The relative migrating distance of the woundedge (E) and the percentage of wound-edge cells with reoriented MTOC (F) were determined as in Fig. 1A andFig. 1D, respectively. Each value represents the mean � S.D. of three independent experiments. *, p � 0.01; **,p � 0.005; Student’s t test.
Role of Ror2 in Wnt5a-induced Polarized Cell Migration
OCTOBER 10, 2008 • VOLUME 283 • NUMBER 41 JOURNAL OF BIOLOGICAL CHEMISTRY 27979
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
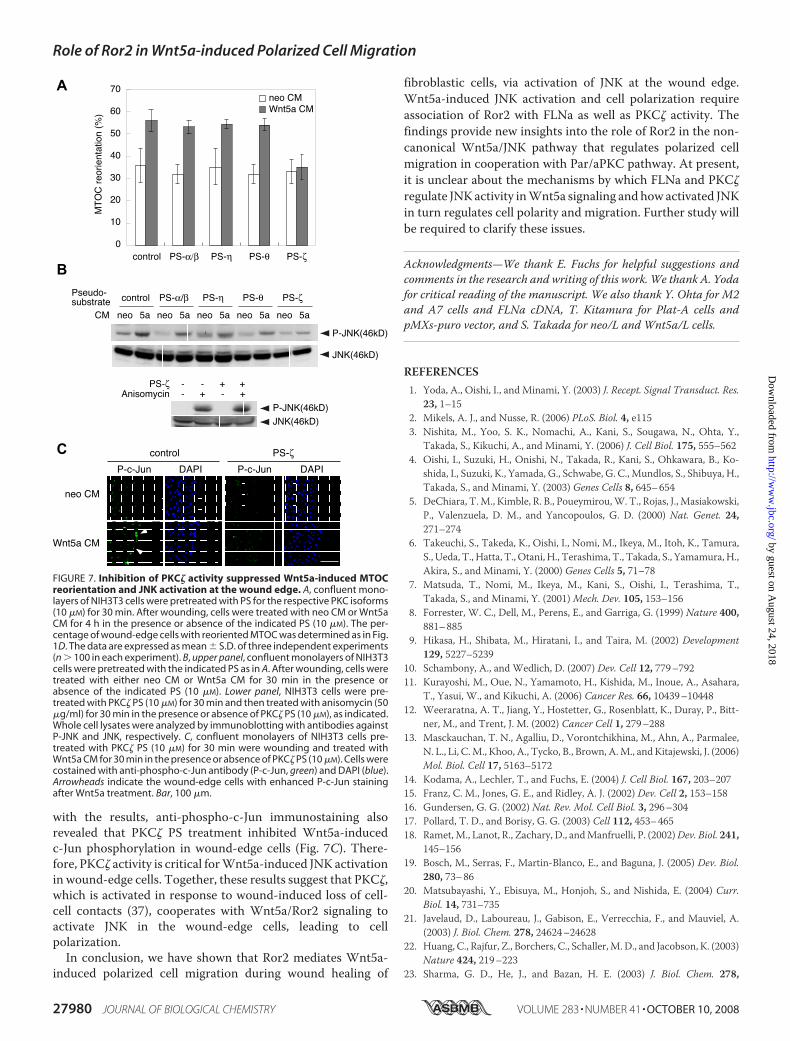
with the results, anti-phospho-c-Jun immunostaining alsorevealed that PKC� PS treatment inhibited Wnt5a-inducedc-Jun phosphorylation in wound-edge cells (Fig. 7C). There-fore, PKC� activity is critical forWnt5a-induced JNK activationin wound-edge cells. Together, these results suggest that PKC�,which is activated in response to wound-induced loss of cell-cell contacts (37), cooperates with Wnt5a/Ror2 signaling toactivate JNK in the wound-edge cells, leading to cellpolarization.In conclusion, we have shown that Ror2 mediates Wnt5a-
induced polarized cell migration during wound healing of
fibroblastic cells, via activation of JNK at the wound edge.Wnt5a-induced JNK activation and cell polarization requireassociation of Ror2 with FLNa as well as PKC� activity. Thefindings provide new insights into the role of Ror2 in the non-canonical Wnt5a/JNK pathway that regulates polarized cellmigration in cooperation with Par/aPKC pathway. At present,it is unclear about the mechanisms by which FLNa and PKC�
regulate JNKactivity inWnt5a signaling andhowactivated JNKin turn regulates cell polarity and migration. Further study willbe required to clarify these issues.
Acknowledgments—We thank E. Fuchs for helpful suggestions andcomments in the research and writing of this work. We thank A. Yodafor critical reading of the manuscript. We also thank Y. Ohta for M2and A7 cells and FLNa cDNA, T. Kitamura for Plat-A cells andpMXs-puro vector, and S. Takada for neo/L and Wnt5a/L cells.
REFERENCES1. Yoda, A., Oishi, I., and Minami, Y. (2003) J. Recept. Signal Transduct. Res.
23, 1–152. Mikels, A. J., and Nusse, R. (2006) PLoS. Biol. 4, e1153. Nishita, M., Yoo, S. K., Nomachi, A., Kani, S., Sougawa, N., Ohta, Y.,
Takada, S., Kikuchi, A., and Minami, Y. (2006) J. Cell Biol. 175, 555–5624. Oishi, I., Suzuki, H., Onishi, N., Takada, R., Kani, S., Ohkawara, B., Ko-
shida, I., Suzuki, K., Yamada, G., Schwabe, G. C., Mundlos, S., Shibuya, H.,Takada, S., and Minami, Y. (2003) Genes Cells 8, 645–654
5. DeChiara, T.M., Kimble, R. B., Poueymirou,W. T., Rojas, J., Masiakowski,P., Valenzuela, D. M., and Yancopoulos, G. D. (2000) Nat. Genet. 24,271–274
6. Takeuchi, S., Takeda, K., Oishi, I., Nomi, M., Ikeya, M., Itoh, K., Tamura,S., Ueda, T., Hatta, T., Otani, H., Terashima, T., Takada, S., Yamamura,H.,Akira, S., and Minami, Y. (2000) Genes Cells 5, 71–78
7. Matsuda, T., Nomi, M., Ikeya, M., Kani, S., Oishi, I., Terashima, T.,Takada, S., and Minami, Y. (2001)Mech. Dev. 105, 153–156
8. Forrester, W. C., Dell, M., Perens, E., and Garriga, G. (1999) Nature 400,881–885
9. Hikasa, H., Shibata, M., Hiratani, I., and Taira, M. (2002) Development129, 5227–5239
10. Schambony, A., and Wedlich, D. (2007) Dev. Cell 12, 779–79211. Kurayoshi, M., Oue, N., Yamamoto, H., Kishida, M., Inoue, A., Asahara,
T., Yasui, W., and Kikuchi, A. (2006) Cancer Res. 66, 10439–1044812. Weeraratna, A. T., Jiang, Y., Hostetter, G., Rosenblatt, K., Duray, P., Bitt-
ner, M., and Trent, J. M. (2002) Cancer Cell 1, 279–28813. Masckauchan, T. N., Agalliu, D., Vorontchikhina, M., Ahn, A., Parmalee,
N. L., Li, C.M., Khoo, A., Tycko, B., Brown, A.M., and Kitajewski, J. (2006)Mol. Biol. Cell 17, 5163–5172
14. Kodama, A., Lechler, T., and Fuchs, E. (2004) J. Cell Biol. 167, 203–20715. Franz, C. M., Jones, G. E., and Ridley, A. J. (2002) Dev. Cell 2, 153–15816. Gundersen, G. G. (2002) Nat. Rev. Mol. Cell Biol. 3, 296–30417. Pollard, T. D., and Borisy, G. G. (2003) Cell 112, 453–46518. Ramet,M., Lanot, R., Zachary, D., andManfruelli, P. (2002)Dev. Biol. 241,
145–15619. Bosch, M., Serras, F., Martin-Blanco, E., and Baguna, J. (2005) Dev. Biol.
280, 73–8620. Matsubayashi, Y., Ebisuya, M., Honjoh, S., and Nishida, E. (2004) Curr.
Biol. 14, 731–73521. Javelaud, D., Laboureau, J., Gabison, E., Verrecchia, F., and Mauviel, A.
(2003) J. Biol. Chem. 278, 24624–2462822. Huang, C., Rajfur, Z., Borchers, C., Schaller,M.D., and Jacobson, K. (2003)
Nature 424, 219–22323. Sharma, G. D., He, J., and Bazan, H. E. (2003) J. Biol. Chem. 278,
neo 5a
P-JNK(46kD)
JNK(46kD)
PS-ζPS-α/β PS-η PS-θ
CM
Pseudo-substrate
neo 5a neo 5a neo 5a neo 5a
control
Anisomycin - + - +
P-JNK(46kD)
JNK(46kD)
-PS-ζ - + +
neo CMWnt5a CM
MT
OC
reo
rient
atio
n (%
)70
60
50
40
30
20
10
0PS-α/β PS-η PS-θ PS-ζcontrol
A
C
B
DAPIP-c-Ju APIP-c-Jun
neo CM
Wnt5a CM
control PS-ζ
FIGURE 7. Inhibition of PKC� activity suppressed Wnt5a-induced MTOCreorientation and JNK activation at the wound edge. A, confluent mono-layers of NIH3T3 cells were pretreated with PS for the respective PKC isoforms(10 �M) for 30 min. After wounding, cells were treated with neo CM or Wnt5aCM for 4 h in the presence or absence of the indicated PS (10 �M). The per-centage of wound-edge cells with reoriented MTOC was determined as in Fig.1D. The data are expressed as mean � S.D. of three independent experiments(n � 100 in each experiment). B, upper panel, confluent monolayers of NIH3T3cells were pretreated with the indicated PS as in A. After wounding, cells weretreated with either neo CM or Wnt5a CM for 30 min in the presence orabsence of the indicated PS (10 �M). Lower panel, NIH3T3 cells were pre-treated with PKC� PS (10 �M) for 30 min and then treated with anisomycin (50�g/ml) for 30 min in the presence or absence of PKC� PS (10 �M), as indicated.Whole cell lysates were analyzed by immunoblotting with antibodies againstP-JNK and JNK, respectively. C, confluent monolayers of NIH3T3 cells pre-treated with PKC� PS (10 �M) for 30 min were wounding and treated withWnt5a CM for 30 min in the presence or absence of PKC� PS (10 �M). Cells werecostained with anti-phospho-c-Jun antibody (P-c-Jun, green) and DAPI (blue).Arrowheads indicate the wound-edge cells with enhanced P-c-Jun stainingafter Wnt5a treatment. Bar, 100 �m.
Role of Ror2 in Wnt5a-induced Polarized Cell Migration
27980 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 41 • OCTOBER 10, 2008
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21989–2199724. Jaeschke, A., Karasarides,M., Ventura, J. J., Ehrhardt, A., Zhang, C., Flavell,
R. A., Shokat, K. M., and Davis, R. J. (2006)Mol. Cell 23, 899–91125. Yamanaka, H.,Moriguchi, T.,Masuyama, N., Kusakabe,M., Hanafusa, H.,
Takada, R., Takada, S., and Nishida, E. (2002) EMBO Rep. 3, 69–7526. Schlessinger, K., McManus, E. J., and Hall, A. (2007) J. Cell Biol. 178,
355–36127. Kani, S., Oishi, I., Yamamoto, H., Yoda, A., Suzuki, H., Nomachi, A., Io-
zumi, K., Nishita, M., Kikuchi, A., Takumi, T., and Minami, Y. (2004)J. Biol. Chem. 279, 50102–50109
28. Paganoni, S., and Ferreira, A. (2005) J. Cell Sci. 118, 433–44629. Mammoto, A., Huang, S., and Ingber, D. E. (2007) J. Cell Sci. 120, 456–46730. Takada, R., Hijikata, H., Kondoh, H., and Takada, S. (2005)Genes Cells 10,
919–928
31. Cunningham, C. C., Gorlin, J. B., Kwiatkowski, D. J., Hartwig, J. H., Jan-mey, P. A., Byers, H. R., and Stossel, T. P. (1992) Science 255, 325–327
32. Palazzo, A. F., Joseph, H. L., Chen, Y. J., Dujardin, D. L., Alberts, A. S.,Pfister, K. K., Vallee, R. B., and Gundersen, G. G. (2001) Curr. Biol. 11,1536–1541
33. Yamamoto, S., Nishimura, O., Misaki, K., Nishita, M., Minami, Y., Yone-mura, S., Tarui, H., and Sasaki, H. (2008) Dev. Cell 15, 23–36
34. Lallemand, D., Ham, J., Garbay, S., Bakiri, L., Traincard, F., Jeannequin, O.,Pfarr, C. M., and Yaniv, M. (1998) EMBO J. 17, 5615–5626
35. Leonardi, A., Ellinger-Ziegelbauer, H., Franzoso, G., Brown, K., andSiebenlist, U. (2000) J. Biol. Chem. 275, 271–278
36. Marti, A., Luo, Z., Cunningham, C., Ohta, Y., Hartwig, J., Stossel, T. P.,Kyriakis, J. M., and Avruch, J. (1997) J. Biol. Chem. 272, 2620–2628
37. Etienne-Manneville, S., and Hall, A. (2001) Cell 106, 489–498
Role of Ror2 in Wnt5a-induced Polarized Cell Migration
OCTOBER 10, 2008 • VOLUME 283 • NUMBER 41 JOURNAL OF BIOLOGICAL CHEMISTRY 27981
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Hamasaki and Yasuhiro MinamiAkira Nomachi, Michiru Nishita, Daisuke Inaba, Masahiro Enomoto, Mayumi
by Activating c-Jun N-terminal Kinase via Actin-binding Protein Filamin AReceptor Tyrosine Kinase Ror2 Mediates Wnt5a-induced Polarized Cell Migration
doi: 10.1074/jbc.M802325200 originally published online July 30, 20082008, 283:27973-27981.J. Biol. Chem.
10.1074/jbc.M802325200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2008/07/31/M802325200.DC1
http://www.jbc.org/content/283/41/27973.full.html#ref-list-1
This article cites 37 references, 15 of which can be accessed free at
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from