immune mechanisms of allergen specific sublingual immunotherapy
DESCRIPTION
ÂTRANSCRIPT

Review article
Immune mechanisms of allergen-specific sublingual
immunotherapy
Allergen-specific immunotherapy has been used inhumans for almost a century with the aim to redirectinappropriate immune responses in atopic patients (1, 2).It has proven to be efficacious to treat type I allergies to avariety of allergens (3–8). While the parenteral (subcuta-neous) route of immunization is still a reference, localroutes (e.g. intranasal, oral) have been considered as analternative with mixed results, both in terms of efficacyand tolerance (9). Of note, a specific form of oraltolerance induction, i.e. sublingual immunotherapy(SLIT) is raising a lot of interest as a noninvasiveprocedure, as it has been shown to be efficacious,provided that high doses of allergen (i.e. 50–100-foldthe subcutaneous dose) are administered (10–25). In arecent meta-analysis, encompassing 22 clinical studiesevaluating SLIT in 979 patients with allergic rhinitis tohouse dust mite, pollens (from grass, parietaria, olive,ragweed, cupressus) and cat dander, it was concluded thatSLIT significantly reduces both symptoms and medica-tion requirements (26). Importantly, it is now widelyadmitted that SLIT is much safer than subcutaneousimmunotherapy (SCIT), with no evidence of anaphylacticshock recorded after more than 500 million doses admin-istered to humans (12, 17, 27, 28). Whereas SLIT has
been successfully used to treat allergic patients, ourunderstanding of the immunological mechanismsinvolved has been limited. We review herein recentscientific advances, which provide some clues on effec-tor/regulatory immune mechanisms elicited during suc-cessful allergen-specific immunotherapy in general, andSLIT in particular. Based on such improved biologicalfoundations, we comment on arising opportunities todesign second-generation sublingual allergy vaccinesrelying upon well-characterized recombinant allergens,capable of controlling T-cell polarization following vac-cine-mediated and subsequent natural exposure to theallergen.
Immunomodulation during allergen-specific immunotherapy
As summarized in Fig. 1, allergen-specific immunothera-py, whether it be SCIT or SLIT, is known to reduce bothimmediate as well as late-phase allergen-induced symp-toms, by acting both on humoral, as well as on cellularimmune mechanisms involved in allergic inflammation (4,29–33). Schematically, three categories of immunologicalchanges are induced by active immunotherapy, encompas-
Sublingual immunotherapy has been shown in some clinical studies to modulateallergen-specific antibody responses [with a decrease in the immunoglobulin E/immunoglobulin G4 (IgE/IgG4) ratio] and to reduce the recruitment and acti-vation of proinflammatory cells in target mucosa. Whereas a central paradigmfor successful immunotherapy has been to reorient the pattern of allergen-spe-cific T-cell responses in atopic patients from a T helper (Th)2 to Th1 profile,there is currently a growing interest in eliciting regulatory T cells, capable ofdownregulating both Th1 and Th2 responses through the production of inter-leukin (IL)-10 and/or transforming growth factor (TGF)-b. We discuss hereinimmune mechanisms involved during allergen-specific sublingual immuno-therapy (SLIT), in comparison with subcutaneous immunotherapy. DuringSLIT, the allergen is captured within the oral mucosa by Langerhans-likedendritic cells expressing high-affinity IgE receptors, producing IL-10 and TGF-b, and upregulating indoleamine dioxygenase (IDO), suggesting that such cellsare prone to induce tolerance. The oral mucosa contains limited number ofproinflammatory cells, such as mast cells, thereby explaining the well-establishedsafety profile of SLIT. In this context, second-generation vaccines based onrecombinant allergens in a native conformation formulated with adjuvants aredesigned to target Langerhans-like cells in the sublingual mucosa, with the aimto induce allergen-specific regulatory T cells. Importantly, such recombinantvaccines should facilitate the identification of biological markers of SLIT efficacyin humans.
P. Moingeon1, T. Batard1, R. Fadel1,F. Frati2, J. Sieber3, L. Van Overtvelt11Stallerg�nes, Antony, France; 2Stallergenes ItaliaS.r.l, Milan, Italy; 3Stallergenes GmbH & Co. KGGermany, Kamp-Lintfort, Germany
Key words: allergy vaccine; Langerhans cells;regulatory T lymphocyte; sublingual immunotherapy.
Philippe MoingeonResearch and DevelopmentStallergenes6 rue Alexis de Tocqueville92160 AntonyFrance
Accepted for publication 5 September 2005
Allergy 2006: 61: 151–165 Copyright � Blackwell Munksgaard 2006
ALLERGY
DOI: 10.1111/j.1398-9995.2006.01002.x
151

sing (i) modulation of allergen-specific antibody responses;(ii) reduction in recruitment and activation of proinflam-matory cells; and (iii) changes in the pattern of allergen-specific T-cell responses (Table 1). Although such biologi-cal changes are usually better documentedwith SCIT, thereare, in this regard, no clear-cut qualitative differencesbetween SCIT and SLIT, suggesting that immune mech-anisms at play are similar (Table 1).With respect to allergen-specific antibody responses,
SCIT often induces an initial increase in seric immuno-globulin E (IgE) levels, prior to a subsequent downregu-lation in the following months (34–39). In patients allergicto grass pollen, SCIT prevents the seasonal rise in IgEantibodies associated with natural exposure to allergens(37, 39). Moreover, successful SCIT protocols resulting inclinical improvement of patients often elicit allergen-specific IgG responses (mostly IgG1 and IgG4) and in afew reported cases, IgA responses (Table 1) (5, 40–44).Similarly, SLIT was shown to increase allergen-specificIgG4 levels comparedwith placebo (10, 11, 13),with amore
limited impact on specific IgE responses (18). A decrease inthe IgE/IgG4 ratio has been observed in a number of SLITstudies (11, 24, 45), with some exceptions (46). A meta-analysis of six SLIT studies with detailed analysis ofantibody responses concluded on a consistent increase inallergen-specific IgG4 levels (26). Such changes in the IgE/IgG4 ratio were found to correlate with a decrease in thelate-phase skin reaction to the allergen andwith the overallclinical efficacy of the vaccine in some studies (23, 24) butnot all (13, 18). In a recent phase I/II trial with grass pollentablets, SLIT was shown to elicit allergen-specific sericIgAs in a dose-dependent fashion (47) and a smallupregulation of IgA responses was also observed whenSLIT was used in house dust mite allergic patients (45).Altogether, allergen-specific IgG (and IgA) antibodiesinduced by immunotherapy are thought to contribute tothe positive clinical response through distinct and nonexclusive mechanisms: (i) these antibodies can competewith IgEs for binding to the allergen, thereby preventingboth basophil or mastocyte degranulation (38–40), as well
Figure 1. Humoral and cellular immune mechanisms involved in type I allergy and immunotherapy: an integrated view. Allergen-specific immunotherapy deals with the cause of type I allergies, by inducing long-term reorientation of an existing and inappropriateimmune response (associated with Th2 cytokine production, high IgE levels, activation of mast cells, basophils and eosinophils leadingto release of proinflammatory mediators such as histamine, eicosanoids and proteases). CD4+ T helper cells (e.g. Th1, Th2 or Tregulatory) stimulated following allergen exposure or desensitization are critical in controlling these various components of theimmune system through the production of distinct cytokines. IL-4 and IL-13 induce IgE class switching, IL-5 supports eosinophilrecruitment and activation, and IL-13 increases mucus production. Regulatory T cells inhibit both Th1 and Th2 responses through theproduction of IL-10 and TGFb. While atopic patients exhibit allergen-specific Th2 CD4+ cells, healthy people rather mount T regresponses when exposed to allergens. Thus, the purpose of allergen-specific immunotherapy is to restore tolerance by shifting T-cellresponses from Th2 to T Reg.
Moingeon et al.
152

Table1.
Modifications
ofimmunological
parametersduringallergen-specific
immunotherapy
(SCITandSLIT)inhumans
Parameters
Allergensused
Comments
Reference
Allergen-specific
antibody
responses(IgE,IgG1,IgG4,
IgA)
Venoms,pollens
(ragw
eed,birch,
grasses,parietaria,olive),dustmites
Used
as:
–naturalbiologicale
xtracts,allergoids
recombinant
protein(Bet
v1)
associated
with
anadjuvant
(e.g.
CaPO
4,Alum
,MPL)for
SCIT
–naturalbiologicale
xtracts,and
allergoids
withoutanyadjuvant
for
SLIT
Subcutaneous
immunotherapy
Immunoglobulin
isotypes:S
CITinducesan
initial
increase,followed
byasubsequent
decrease
ofspecificIgEs,aswellasan
increase
inallergen-specific
IgG1
andIgG4.
AnriseinIgG4/IgEratio
hasbeen
documentedwith
hypoallergenicformsof
recombinant
Betv1(peptidefra
gmentsandtrimer).Insomestudies,butnotall,
changesinaffinity
ofIgG4
antibodiesfortheallergen
duringimmunotherapy
have
been
show
n.AmonophosphoryllipidA(MPL)adjuvantedvaccinereducesseasonally
boostedIgEresponsesandinducesstrong
IgG1
andIgG4
responsesagainst
grass-pollenallergens.SCITelicits
allergen-specific
IgAresponsesdetectable
intheblood
5,34–40,
44,83
Functionalcharacterizationof
antibodies:IgG4
exhibitsomeblocking
activity.Insomebut
notallstudies,a
corre
lationwas
foundbetweenIgG(mostly
IgG4)concentrations
and
clinicaloutcom
es.IgG4antibodiesreduce
allergen
T-cellresponsesby
inhibiting
bindingof
allergen-IgEcomplexes
toantigen
presentingcells
40–44,48,4
9
Sublingualimmunotherapy
Immunoglobulin
isotypes:S
LITagainsthousedustmite
orgrasspollenincreasesspecific
IgEs
andIgG4,inassociationwith
animprovem
entof
therespiratoryfunction.
Several
studieswith
SLITreportinductionof
IgG4
responses,withoutanyincrease
ordecrease
ofIgEs
(i.e.ƒ
ofIgG4/IgEratio).Forexam
ple,SLITwith
aParietaria
judaicaextra
ctinchildrenreducesrhinitissymptom
sandboth
skin
andconjunctivalallergen
reactivity
inparallelw
ithincreasedspecificIgG4
seric
antibodies.In
contrast,preseasonalrush
SLITagainstP.judaicarhinoconjunctivitisor
SLITinasthmaticchildrensensitizedto
miteshadno
detectable
impact
onspecificIgEor
IgG4
levels,d
espite
improvem
ent
ofsymptom
anddrug
scores.A
phaseI/IIstudy
conductedwith
grasspollen(Phleum
pratense)tabletsdocumentedadose
dependentinductionof
allergen
specificIgG4
andIgAresponses.Ameta-analysisconductedon
sixSLITstudiesconcludedthat
the
combinedstandardizedmeandiffe
rences
(SMD)
betweentre
ated
patientsandcontrolsfor
changesinallergen-specific
IgEwas
0.22
(P¼
0.06),whereas
SMDwas
0.6(P<0.00001)
forallergen-specific
IgG4
10,11,14,23,24,26,
45,47
Recruitmentandactivationof
proinflammatorycells
(mast
cells,eosinophilsandbasophils)
Pollens
(ragw
eed,grasses,parietaria),
dustmites
Used
as:
–naturalbiologicale
xtracts,allergoids
associated
with
anadjuvant
(e.g.
CaPO
4,Alum
)for
SCIT
–naturalbiologicale
xtracts,allergoids
recombinant
protein(Phlp1,Betv1)
withoutanyadjuvant
forSLIT
Subcutaneous
immunotherapy
Impact
oncellrecruitment:SCITreducestherecruitmentinto
theskin
andmucosa(nose,lung)o
finflammatorycells
includingmastcells,eosinophils,b
asophils,n
eutro
phils
50–60
Impact
onproinflammatorymediators:a
decrease
inseric
eotaxinwas
foundto
beassociated
with
successfulIT.S
CITdecreasesthereleaseof
inflammatorymediators(e.g.h
istamine,
kinins,
PGD2)b
ybasophils
andmastcells
Sublingualimmunotherapy
Impact
oncellrecruitmentandactivation:Rush
SLITto
Parietaria
inducesareductionin
neutrophils
andeosinophils
recruitmentas
wella
sICAM
-1expression
aftereyeor
nasalchallenge,
incorre
lationwith
areductionin
both
symptom
anddrug
intake
scores.S
ublingual
challengewith
increasing
dosesof
recombinant
Phlp
1or
Betv1show
slim
itedreactogenicitywith
mastcells
63,65,66
Impact
onproinflammatorymediators:S
LITinchildrenwith
grasspollenallergydecreasesnasal
eosinophilcationicproteinlevelsandpreventsincrease
intryptasefollowingallergen-specific
challenge.In
childrenallergicto
D.pteronyssinus,SLITreducesseric
ECPandprolactin
levels
(thelatte
rlikelyassociated
with
areducedactivationof
Tlymphocytes).Along-te
rmimmunological
effect
ofSLITisto
reduce
residualsignsof
persistent
inflammation(decreaseinsolubleICAM
-1,in
mucosa,which
corre
lateswith
adecrease
inmetacholineresponsiveness)
61–65
Immune mechanisms of SLIT
153

Table1.
Continued
Parameters
Allergensused
Comments
Reference
T-cellresponsesandcytokine
patte
rns
Venoms/PLA2,dustmites,pollens
(ragw
eed/Am
ba1,grasses),cat
dander/Feld1
Used
as:
–naturalbiologicalextracts,
allergoids,p
urified
naturalallergen
(Amba1)or
peptides
associated
with
anadjuvant
(e.g.C
aPO4,A
lum,C
pGs)
forSC
IT–naturalbiologicalextracts,
withoutanyadjuvant
forSLIT
Subcutaneous
immunotherapy
Impacton
theTh1/Th2ratio:S
CITdownregulates
allergen-specific
Th2responses(bothin
termsof
cytokine
productionandproliferation)with
orwithoutstimulationof
allergen-
specificTh1cells.R
apidinvitro
allergen-inducedTh-2
lymphocyteapoptosishasbeen
observed
inonestudy.Increase
insignalinglymphocyticactivatingmolecules
(SLAM)
gene
expression
duringpollenIT.Insomestudies,Th2responsesareincreasedduringthe
titrationphase,
andtheTh2/Th1ratio
isonlysignificantlydecreasedduringthemaintenance
phase.Modificationof
Th2/Th1ratio
corre
lateswith
clinicalimprovem
ent,decrease
inlate-phase
skinreactions
andallergen-specific
IgElevels.A
ssociationbetweenallergens
(e.g.A
mba1)
andoligoCpGimmunostim
ulatorysequences,used
asamixor
asconjugate
vaccines
was
foundto
decrease
Th2andto
enhances
Th1responses
53,66–80,8
2,112
Modificationof
localT-cellresponses:d
ecreaseof
Th2cytokines(IL-4/IL-5/IL-3)a
ndincrease
ofTh1cytokines(IFNc,
IL-12),atlocallevels(e.g.intheskinfollowingintra
dermalgrasspollen
challenge).D
ecreaseinrecruitmentintarget
organs
ofTh2CD
4+Tcells
expressing
theCXCR1
receptor
81,112
Impacton
regulatory
Tcells:Inductionof
CD4+,C
D25+
Tregulatory
lymphocytes
andTr1IL-10
producingcells,intheblood.
Anincrease
inIL-10mRN
Aandproteinisdetected
inskinbiopsies
from
allergen
challenged
subjectsfollowingwaspvenomimmunotherapy.S
omeof
theseregulatory
Tcells
exhibitaCD
62L)
CCR5
+profile
suggestingacapacityto
migrate
toperipheraltissues.Early
orsustainedincrease
ofregulatory
cytokines(IL-10andTGF-b,respectively)isproposed
tocorre
late
with
successfulimmunotherapy.
83,84,90,112,148
Studiesconductedwith
peptides:studies
conductedwith
peptides
(Feld1,PLAA2)dem
onstrate
askew
ingof
theTh1/Th2ratio
towards
theformer,inductionof
SOCS
2andof
IL-10-producingcells.
Injections
ofsm
all(16
to17-mer)Feld1peptides
inhibitthecutaneouslate-phase
reactionto
whole
catdander,a
swella
sallergen-stim
ulated
productionby
PBMCs
ofboth
Th1(IFN-c)and
Th2(IL-4,
IL-13)cytokines,concom
itant
with
anincrease
inIL-10production
4,79,82
Sublingualimmunotherapy
Impacton
theTh1/Th2ratio:inadultswith
seasonalrhinoconjunctivitis,SLITwith
agrasspollen
extra
cthadno
detectableeffecton
sublingual
infiltra
tionby
CD3+
Tcells
orlocalproductionof
IL-12.Nostimulationof
T-cellproliferationor
cytokine
productioninchildrenallergicto
grass
pollen,
despite
apositiverescue
onmedicationuse.
Adecrease
inseric
IL-13andT-cell
proliferativeresponses(associatedwith
aneffect
onsymptom
s)hasbeen
observed
withoutany
detectable
change
inTh1/Th2ratio,inchildrentre
ated
with
D.pteronyssinusextra
cts
23,46,86
Impacton
regulatory
Tcells:Inapreliminaryreport,
inductionof
circulatingIL-10producing
peripheralT
cells
inhousedustmite
allergicpatients
87
Moingeon et al.
154

as allergen capture and presentation to T lymphocytes byFceRI+ and CD23+ antigen-presenting cells (APCs) (48,49); and (ii) such antibodies may act as blocking antibodiesby engaging low-affinity Fc receptors for immunoglobulins(e.g. FccRII) expressed by B lymphocytes, basophils, ormast cells. As FccRII receptors contain immunoreceptortyrosine-based inhibitorymotifs (ITIM), they transduce, asa consequence, negative signals preventing cellular activa-tion and release of soluble proinflammatory mediatorsfollowing co-aggregation with FCeRI receptors (42, 43).In a number of studies, SCIT was shown to inhibit both
the recruitment and activation in mucosa of proinflamma-tory cells involved in the allergic reaction (50–60). Forexample, successful SCIT has been associated with adecrease in the recruitment of mast cells, basophils andeosinophils in the skin, nose, eye and bronchial mucosa,following provocation or natural exposure to allergens(Table 1). Similarly, SLIT prevented the recruitment ofeosinophils in the eye or in the nose after allergen challenge(61–65). SLIT with grass pollen extracts was shown todecrease local or systemic levels of eosinophil cationicprotein (ECP), without any increase in tryptase (61, 62).Similarly, rush SLIT to parietaria reduced the recruitmentof neutrophils and eosinophils to the nasal mucosa as wellas the expression of the intercellular adhesion molecule-1(ICAM-1) (66).In the context of an emerging integrated picture of the
physiology of immune responses, the aforementionedchanges in immune parameters are assumed to be a directconsequence of an impact of immunotherapy on CD4+ T-cell responses (Fig. 1). It is well known that allergicpatients usually mount strong allergen-specific CD4+
T-cell responses of the T helper (Th)2 type, characterizedby the secretion of high amounts of interleukin (IL)-4, IL-5 and IL-13 cytokines (30, 37, 67). In this regard, a centralgoal for immunotherapy has been to reorient allergen-specific T-cell responses in atopic patients from a Th2 toTh1 profile [the latter being rather associated with theproduction of interferon (IFN)-c and IL-12 cytokines](53, 68–80). A number of studies have indeed correlatedsuccessful SCIT with the induction of Th1 responses and/or the decrease of Th2 cytokine production (thus, it is theTh1/Th2 balance which appears critical). Interestingly, ina recent SCIT study, the Th2/Th1 ratio was found toincrease during the titration phase, prior to decreasingduring the maintenance phase, in parallel with late-phaseskin reactivity and allergen-specific IgE levels (70). Inseveral SCIT studies with grass pollen, a switch from aTh2 to a Th1 profile was not consistently observed at asystemic level, but was rather detected locally (i.e. withinthe nasal mucosa or the skin) (53, 69, 81). This observa-tion emphasizes the importance of documenting immunechanges not only in peripheral blood, but also locally intarget organs. More recently, it has been shown that SCITinduces a new subset of CD4+ Th cells, called regulatoryT cells, which exhibit a capacity to downregulate both Th1and Th2 responses through the production of IL-10 and/
or transforming growth factor (TGF)-b (see below) (50,82–85). There is less evidence on the impact of SLIT onT-cell responses. In several studies conducted in childrenor adults with seasonal allergic rhinoconjunctivitis tograss pollen, no significant effect of SLIT on T-cellfunctions (i.e. cytokine production, proliferation) wasobserved (23, 46). SLIT does not induce any detectablechanges in the numbers of dendritic cells (DCs) nor Tlymphocytes in the epithelium or lamina propria of theoral mucosa (14). Immunization through the sublingualroute was nevertheless shown in other studies to decreasethe production of the Th2 cytokine IL-13 and theproliferation of PBMCs from patients allergic to housedust mite (86, 87). As of today, there is still no firmevidence that SLIT can induce regulatory T cells. Apreliminary study suggests that SLIT increases IL-10production in peripheral blood mononuclear cells(PBMCs) from house dust mite (HDM) allergic patientsfollowing in vitro stimulation with Dermatophagoidesfarinae antigens, but also with recall antigens (e.g.Candidaalbicans) or PHA, when compared with untreated allergicpatients (88). That some IL-10-secreting T cells are notallergen-specific raises the possibility of a bystanderimmunosuppressive effect of SLIT. Of note, high-doseSLIT regimens with ovalbumin in mice induces ova-specific T cells producing TGF-b in the spleen of sensitizedanimals (L. Van Overtvelt, P. Moingeon, unpublishedresults). Further studies are needed to document unam-biguously regulatory T cells induction during SLIT.
Regulatory T cells and allergy vaccines
Although both anergy and T-cell depletion are known tocontribute to the establishment of peripheral toleranceagainst environmental antigens, it is now broadly admit-ted that antigen-specific T-cell populations with suppres-sive/regulatory function play a key role in controllingimmune responses to both self- and nonself-antigens (89–95). These cells, termed regulatory T cells, are heteroge-neous, and include both: (i) naturally occurringCD4+CD25+ T cells and (ii) cells induced in theperiphery following antigen exposure (e.g. Tr1 cells,Th3 cells, and CD8+ regulatory T cells). These varioussubsets of regulatory T cells can be distinguished on thebasis of their surface markers and the pattern of cytokinesthey produce (Table 2). Antigen/allergen presenting DCsplay a critical role in the induction of T-cell-mediatedtolerance, in that immature DCs in the absence ofproinflammatory signals and possibly subsets of special-ized DCs can both support the differentiation of regula-tory T cells (96, 97). Regulatory T cells are usually anergicwith a low spontaneous proliferation rate, and are highlydependent on exogenous IL-2. They can downregulateboth Th1 and Th2 immune responses against viruses,bacteria, parasites and allergens (89–92), either by directcell–cell contact (involving PD1, membrane-bound
Immune mechanisms of SLIT
155

TGF-b or CTLA4 molecules) or through the productionof immunosuppressive cytokines such as TGF-b (Th3cells), or IL-10 (Tr1 cells) (Table 2) (90–92, 98, 99).Naturally occurring CD4+CD25+ regulatory T cells
represent 5–10% of peripheral CD4+ T cells in healthymice and humans. They are mainly thymus-derived, butcan also be induced in the periphery following sustainedantigen or TGF-b stimulation of CD4+CD25)-naive Tcells (99). These cells express the forkhead/winged helixtranscription factor Fox p3, which appears to play a keyrole in supporting their regulatory function (100).Type 1 regulatory (Tr1) T cells have a low-proliferative
capacity, produce high levels of IL-10 and TGF-b andlow levels of IL-2 and IL-4. Tr1 cells can be generated invitro by stimulating naive CD4+ T cells in the presence ofIL-10, with/without IFN-a (101), vitamin D3 plus dex-amethasone (102) or a combination of anti-CD46 plusanti-CD3 antibodies (103). CD4+CD25+ regulatory Tcells expressing the a4b7 integrin can also convert naiveCD4+ T cells into Tr1 cells (104).T helper type 3 (Th3) cells which produce TGF-b, IL-4
and IL-10 are induced following oral administration ofthe antigen (105). In vitro, CD4+CD25+ regulatory Tcells expressing the a4b1 integrin have been shown toconvert naive CD4+ T cells into Th3 cells (104).As of today, there is a growing evidence supporting the
role of regulatory T cells in controlling the developmentof asthma and allergic disease in a variety of models
(Fig. 2), although it is not clear yet which of the variousregulatory T cell subsets are the most important in thisregard (85, 92, 94). A revised version of the hygienehypothesis proposes that a limited exposure to infectiouspathogens during infancy, most particularly telluricmycobacteria and parasites, may prevent the establish-ment of not only a Th1, but also a T reg repertoire,thereby explaining in part the observed increase inprevalence of allergies in developed countries (106).Several studies document an association between atopyand a deficit in T reg functions. For example, childrenborn with a dysfunctional Fox p3 gene exhibit a deficit inCD4+CD25+ regulatory T cells and develop severeautoimmune diseases often associated with eczema,elevated IgE levels, eosinophilia and food allergy [thepolyendocrinopathy, enteropathy, and X-linked inherit-ance (IPEX) syndrome] (107). Moreover, for at leastsome atopic subjects with active disease, the suppressiveactivity of CD4+CD25+ regulatory T cells is significantlydecreased in vitro when compared with nonatopic indi-viduals, potentially explaining the loss of toleranceagainst allergens (108). In patients allergic to birchpollen, a functional deficit has also been observed inCD4+CD25+ regulatory T cells, which appears to peakduring the pollen season, and impacts the capacity ofsuch cells to inhibit Th2, but not Th1 responses (109).Moreover, DCs from children with allergic rhinitis can beimpaired in their capacity to produce IL-10 (110).
Table 2. Characteristics of naturally occurring and induced CD4+ and CD8+ regulatory T cells
Naturally occurring TR cellsCD4+CD25+
Peripherally induced TR cellsTr1 and CD4+CD25+/)
Peripherally induced TR cellsCD4+ Th3 CD8+ T Reg
Surface markersCD25 +++ ) to ++ ++ +GITR ++ ) ?ICOS ++ ++ ?CTLA-4 +++ + ++ )Other markers CD103, LAG3, CD122, neurophilin1 CD122, T1-ST2Foxp3 ++ ) (+/) after activation) ?CD45 RB low + + + +CD45 RO + + +
Cytokine secretedIL-10 +/) ++++ +/) ++TGF-b +/) ++ ++++ +/)IL-4 ) +/) +/)IL-5 ) ++ ?IL-2 ) +/) )IFN-c +/) ++ +/)
Cell contact (CTLA-4, PD1)IL-10, TGF-b (in vitro)
IL-10, TGF-bCell contact (CTLA-4, PD1) TGF-b IL-10, TGF-b
Suppressive mechanism(s)Differentiation/inductionfactor(s)
Immature or tolerogenicDCs
TGF-b
Immature or tolerogenicDCs
IL-10, IFN-aIL-4, anti-CD3/antiCD46, VitD3/dexamethasone, a4b7+TR
Immature or tolerogenicDCs
TGF-b, IL-4, IL-10, a4b1+TR
Immature or tolerogenicDCs?
Induction by allergen-specificimmunotherapy
++ (SCIT) +++ (SCIT, possibly SLIT) +++ (oral IT), likely SLIT -swallow Unknown
Moingeon et al.
156

Interestingly, allergen-specific IL-10-secreting Tr1 cellsare highly represented in healthy individuals in compar-ison with allergen-specific IL-4-secreting Th2 cells, sug-gesting that regulatory T cells are predominant duringnatural immune responses to environmental allergens innonatopic donors (111). Natural Tr1 responses are alsoinvolved in establishing phospholipase A2 tolerance inbeekeepers exposed to multiple stings (112, 113). Incontrast, this pattern of T-cell responses is reversed inallergic individuals with a heavy skewing towards Th2relative to T reg responses (111, 114). A functional role ofregulatory T cells has been demonstrated in vivo in mice,on the basis of adoptive transfer experiments showingthat allergen-specific CD4+CD25+ cells or Tr1 clonesinhibit allergen-induced Th2 responses and IgE produc-tion, as well as airway eosinophilia (90–92).Regulatory T lymphocytes can control an established
allergic response via distinct mechanisms (Fig. 2): IL-10and TGF-b decrease IgE production and enhance IgG4and IgA production, respectively. Both cytokines lowerthe release of proinflammatory mediators by downregu-lating IgE-dependent activation of basophils and mast
cells and by decreasing survival and activation of eosin-ophils. IL-10 and TGF-b also inhibit the production ofTh2 cytokines such as IL-4 and IL-5 (50, 90, 113). Inaddition, regulatory T cells exhibit a direct inhibitoryeffect on Th1 and Th2 T cells, through cell–cell contact,or by decreasing the antigen presenting function of DCs.
Recently, several successful immunotherapy studiesconducted through the subcutaneous route in patientswith either grass pollen, house dust mite or venom allergyhave shown an induction of various types of regulatory Tcells including CD4+CD25+ regulatory T cells (84, 90,115) or IL-10-producing CD4+CD25) Fox p3-Tr1 cells(82, 83). Importantly, the suppressive activity of regula-tory T cells induced in the course of immunotherapyappears to be allergen specific (83). Collectively, thesestudies suggest the importance of stimulating allergen-specific T regs during immunotherapy. They also provethat it is feasible to elicit regulatory T cells from a pool ofCD4+CD25)-naive T-cell progenitors in allergic patients,some of which may present a deficit in their T regfunction and repertoire. Early IL-10 production (withindays) and sustained TGF-b levels (over a year after
Figure 2. Regulatory T-cell dynamics in allergy. Regulatory T cells producing IL-10 and/or TGF-b are induced not only in atopicpatients by successful immunotherapy, but also during natural allergen exposure in healthy people. As per the hygiene hypothesis,limited exposure to bacteria and parasites in developed countries may result in a poor establishment of a T reg repertoire duringchildhood, thereby contributing to an increase in the frequency of allergies. Certain atopic patients have a deficit in their T regfunction, although several SCIT studies have shown that it is possible to induce allergen-specific regulatory T cells in atopic patients.Regulatory T cells can control and regulate all effector mechanisms activated during allergy and Th2 responses through the productionof IL-10/TGF-b and/or cell–cell contact. IL-10 is a potent suppressor of total and allergen-specific IgEs, whereas it induces anantibody isotype switch towards IgG4. TGF-b also decreases IgE production and induces immunoglobulin isotype switch towardsIgA. IL-10 and TGF-b act directly or indirectly on human airways to decrease both mucus production and airway hyper-reactivity.
Immune mechanisms of SLIT
157
immunization) have been proposed as potential biologicalcorrelates for successful immunotherapy (90).
Oral mucosa and immune responses
SLIT and induction of peripheral tolerance
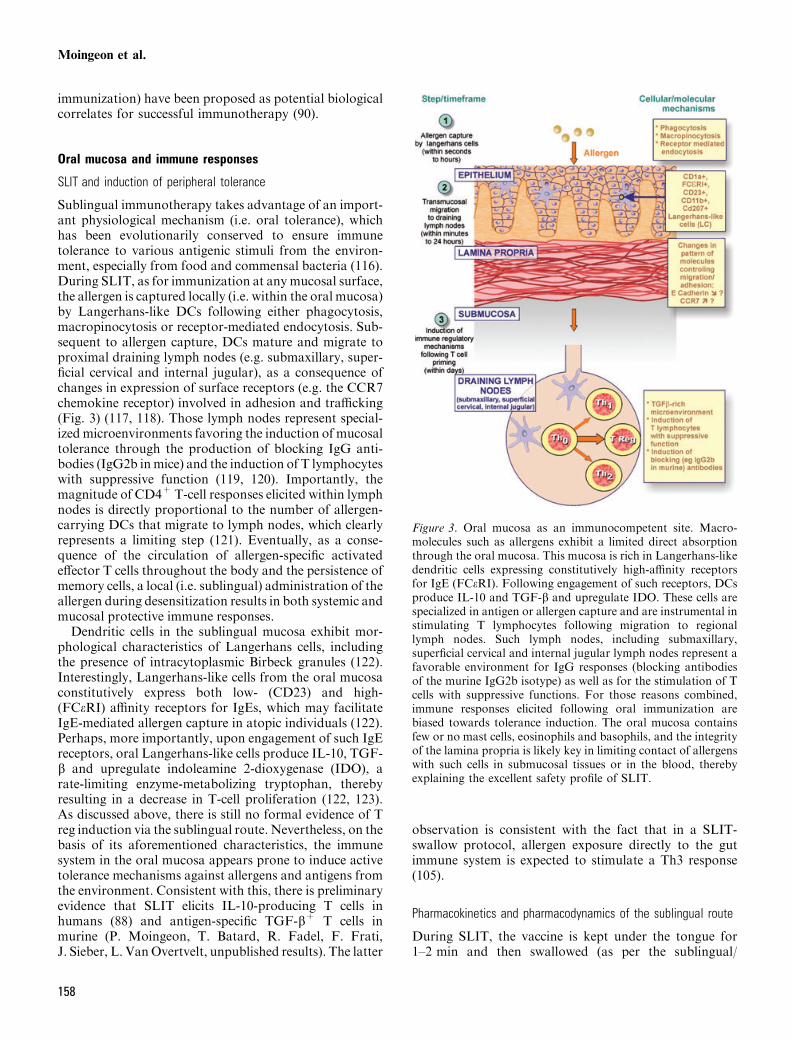
Sublingual immunotherapy takes advantage of an import-ant physiological mechanism (i.e. oral tolerance), whichhas been evolutionarily conserved to ensure immunetolerance to various antigenic stimuli from the environ-ment, especially from food and commensal bacteria (116).During SLIT, as for immunization at any mucosal surface,the allergen is captured locally (i.e. within the oral mucosa)by Langerhans-like DCs following either phagocytosis,macropinocytosis or receptor-mediated endocytosis. Sub-sequent to allergen capture, DCs mature and migrate toproximal draining lymph nodes (e.g. submaxillary, super-ficial cervical and internal jugular), as a consequence ofchanges in expression of surface receptors (e.g. the CCR7chemokine receptor) involved in adhesion and trafficking(Fig. 3) (117, 118). Those lymph nodes represent special-ized microenvironments favoring the induction of mucosaltolerance through the production of blocking IgG anti-bodies (IgG2b inmice) and the induction of T lymphocyteswith suppressive function (119, 120). Importantly, themagnitude of CD4+T-cell responses elicited within lymphnodes is directly proportional to the number of allergen-carrying DCs that migrate to lymph nodes, which clearlyrepresents a limiting step (121). Eventually, as a conse-quence of the circulation of allergen-specific activatedeffector T cells throughout the body and the persistence ofmemory cells, a local (i.e. sublingual) administration of theallergen during desensitization results in both systemic andmucosal protective immune responses.Dendritic cells in the sublingual mucosa exhibit mor-
phological characteristics of Langerhans cells, includingthe presence of intracytoplasmic Birbeck granules (122).Interestingly, Langerhans-like cells from the oral mucosaconstitutively express both low- (CD23) and high-(FCeRI) affinity receptors for IgEs, which may facilitateIgE-mediated allergen capture in atopic individuals (122).Perhaps, more importantly, upon engagement of such IgEreceptors, oral Langerhans-like cells produce IL-10, TGF-b and upregulate indoleamine 2-dioxygenase (IDO), arate-limiting enzyme-metabolizing tryptophan, therebyresulting in a decrease in T-cell proliferation (122, 123).As discussed above, there is still no formal evidence of Treg induction via the sublingual route. Nevertheless, on thebasis of its aforementioned characteristics, the immunesystem in the oral mucosa appears prone to induce activetolerance mechanisms against allergens and antigens fromthe environment. Consistent with this, there is preliminaryevidence that SLIT elicits IL-10-producing T cells inhumans (88) and antigen-specific TGF-b+ T cells inmurine (P. Moingeon, T. Batard, R. Fadel, F. Frati,J. Sieber, L. VanOvertvelt, unpublished results). The latter
observation is consistent with the fact that in a SLIT-swallow protocol, allergen exposure directly to the gutimmune system is expected to stimulate a Th3 response(105).
Pharmacokinetics and pharmacodynamics of the sublingual route
During SLIT, the vaccine is kept under the tongue for1–2 min and then swallowed (as per the sublingual/
Figure 3. Oral mucosa as an immunocompetent site. Macro-molecules such as allergens exhibit a limited direct absorptionthrough the oral mucosa. This mucosa is rich in Langerhans-likedendritic cells expressing constitutively high-affinity receptorsfor IgE (FCeRI). Following engagement of such receptors, DCsproduce IL-10 and TGF-b and upregulate IDO. These cells arespecialized in antigen or allergen capture and are instrumental instimulating T lymphocytes following migration to regionallymph nodes. Such lymph nodes, including submaxillary,superficial cervical and internal jugular lymph nodes represent afavorable environment for IgG responses (blocking antibodiesof the murine IgG2b isotype) as well as for the stimulation of Tcells with suppressive functions. For those reasons combined,immune responses elicited following oral immunization arebiased towards tolerance induction. The oral mucosa containsfew or no mast cells, eosinophils and basophils, and the integrityof the lamina propria is likely key in limiting contact of allergenswith such cells in submucosal tissues or in the blood, therebyexplaining the excellent safety profile of SLIT.
Moingeon et al.
158

swallow procedure). When the vaccine is immediatelyswallowed, clinical efficacy is substantially decreased(22, 124). On the basis of human studies demonstratingthat a limited therapeutic effect is observed when thevaccine is spat out, it has been proposed that sublingual-swallow immunotherapy is more efficient than sublin-gual-spit because it enhances the duration of contact ofthe allergen with the oral mucosa (124). Whereasimmunomodulation is initiated within minutes followingcontact with the oral mucosa, allergens absorbed in thegastrointestinal tract also probably contribute to theefficacy of SLIT.The tissue under the tongue is highly vascularized, with
blood vessels draining directly into the jugular vein. As aconsequence, small synthetic drugs administered sublin-gually are quickly absorbed and enter the bloodstreamwithout passing through the intestine and the liver (125,126). For example, the sublingual route is commonly usedin humans to administer the vasodilator nitroglycerin(glyceryl trinitrate), as a treatment for angina pectoris,leading to a plasmatic peak within 5 min with an overallbioavailability of approximately 70% (125). Similarly,sublingual administration of the opioid analgesic bupr-enorphine yields a bioavailability of up to 55% throughthe sublingual route, in comparison with 15% obtainedafter direct ingestion (126). In contrast with theseobservations made with small synthetic molecules, bio-distribution studies in humans demonstrated that there isno significant direct absorption of peptides or proteins –including allergens – through the sublingual mucosa(127–130). Following administration of the radiolabeledParietaria judaica pollen allergen (Par j 1), either as anorosoluble tablet or a solution kept in the mouth for1–1.5 or 20–30 min, respectively, no direct absorptioninto the blood was detected (129, 130). Only afterswallowing and contact with the gut mucosa, the radio-labeled Par j 1 allergen got distributed rapidly in theblood, with a plasmatic peak detectable within 2 h (129).Such biodistribution studies also suggested that signifi-cant amounts (e.g. 20% of the administered dose) of theallergen can persist for 2 h on the sublingual mucosalsurface, even though patients were allowed to rinse theirmouth extensively (129, 130). Consistent with this obser-vation, a limited proteolytic activity against allergens isfound in saliva, whereas in contrast exposure to gastro-intestinal fluids results in complete allergen degradation(131–133).As for SCIT, conventional SLIT protocols rely upon a
build-up phase (with a gradual increase in the dosingduring 4–6 weeks) and a maintenance phase (withadministration of the maximum dose one to three timesa week over several years). In the course of suchimmunization schemes, it has been proposed that bothdownregulation of proinflammory cells and upregulationof blocking IgG antibodies as well as IL-10 productionoccur quickly (i.e. within days) following immunotherapy(16). In contrast, a detectable impact on Th1 and Th2
responses and adaptive immunity is rather thought tooccur within months (16). Whereas immunological chan-ges can be detected shortly following immunotherapy, arecent study has suggested that SLIT is efficacious onsymptoms and medication after at least 1 year of therapy,with maximum benefit being observed only after 2 years(134). It should be emphasized, however, that several rushand ultrarush protocols for SLIT have demonstrated thatit can induce a decrease in skin reactivity and a readilydetectable clinical benefit within weeks or even days (16,66, 135). Whereas little is known on the duration ofSLIT-induced immunomodulation, several independentstudies have suggested that SLIT in children withrhinoconjunctivitis to either grass pollen or HDM,prevents asthma as well as new sensitizations (20, 136,137). Given that in some of these studies, a follow-up ofup to 10 years after the initiation of SLIT was performed,it is likely that immune memory mechanisms are estab-lished following SLIT.
Why high doses for SLIT?
It is well established that SLIT requires more allergen(at least 50–100 times) than SCIT to reach the samelevel of efficacy (15). One explanation for this is thatcurrent immunotherapy protocols rely upon an adjuvant(e.g. calcium phosphate or aluminum hydroxide) forsubcutaneous but not sublingual administration.Although it has been suggested that reaching a thresholdcumulative dose of allergen is important for a successfulSLIT, it is also possible that current vaccines do nottarget immune cells efficiently, most particularly DCs.Thus, high-dose regimens likely facilitate capture ofsufficient amounts of allergens by sentinel DCs withinthe oral mucosa, which clearly represents a critical stepto induce a strong and long-lasting T-cell response (121).Interestingly, numerous studies suggest that low or highdoses of antigens exhibit a radically distinct effect on Tlymphocyte stimulation, with low doses inducing Th2responses, whereas higher doses of antigen ratherstimulating Th1 CD4+ T cells (138, 139). Moreover,exposure to high doses of allergen during immunothera-py may promote the trafficking of Th1 over Th2 cells bymodulating surface expression of adhesion moleculesand chemokine receptors selectively on those cells (140).The influence of antigen dosage on regulatory T cells ispresently unclear: initial studies suggested that smalldoses of antigen administered orally were efficient ateliciting IL-10+ TGF-b+ Th3 cells in the gut (105).However, regulatory T cells are poorly reactive withantigen in vitro, raising the hypothesis that a strongantigenic stimulation may be necessary to induce pro-liferation and activation of such T cells. Importantly,high doses of antigen will provide a strong initial burstof T lymphocyte stimulation, leading to the establish-ment of T-cell-mediated immune memory and long-termtolerance (141).
Immune mechanisms of SLIT
159

SLIT safety profile
On the basis of extensive clinical experience, it is nowfirmlyestablished that SLIT is very well tolerated both in adultsand young children (22, 26, 27). Side effects are usuallylocal, encompassing oral itching, swelling, and irritation,and occur usually within minutes, thus, likely implying anIgE-mediated reaction. As inflammatory markers such astryptase orECPare not increased locally during SLIT, IgE-mediated reactions occurring in the mouth followingsublingual exposure to the vaccine are usually ratherlimited (62). Allergen-challenge experiments performed inhumans with recombinant allergens (e.g. rPhl p 1, rBet v 1)have confirmed that the sublingual mucosa is at least 10times less reactive than the nasal mucosa or the skin (142).This is likely explained by the fact that only limitednumbers of proinflammatory cells (such as mast cells) arepresent in the oral mucosa. Thus, in contrast to SCIT,allergens exposed to the oral mucosa are not in directcontact with basophils from the blood or mast cells intissues.While SLIT efficacy clearly correlates with allergen
dosage, high-dose regimens do not appear to enhancedramatically the frequency of systemic or local adverseevents, in contrast to SCIT (28). Importantly, high dosesof allergens have been shown in vitro to be less efficientthan low doses in inducing the release of proinflamma-tory mediators by mast cells (143). In a phase Idose-finding study with sublingual grass pollen tablets,high-dose regimens were very well tolerated provided thata short-dose escalation was conducted prior to reachingthe maintenance dose (144).
Implications for the development of second-generationsublingual vaccines
Based on our current understanding of immune mecha-nisms underlying allergen-specific immune responses,optimizing vaccines to control the type of T lymphocytes,and thus the pattern of cytokines induced during naturalexposure to the allergen or vaccination, is critical. Second-generation vaccines could possibly be more efficacious ifspecifically designed to elicit a strong allergen-specificregulatory T-cell response without exacerbation of the Th2allergic response, thereby limiting adverse reactions. Forthis, the sublingual route appearsmostly appropriate giventhat FCeRI+ IL-10/TGF-b-producing sentinel DCs pre-sent in the oral mucosa are likely to be prone to inducetolerance, as opposed to immunostimulation.While the first generation of sublingual vaccines
currently used is based on natural biological extracts,new vaccines are being developed which rely uponselected recombinant allergens (145, 146). There is, as oftoday, considerable interest in developing hypoallergenicversions of target allergens in which IgE-binding epitopes
have been knocked down by side-directed mutagenesis ordisruption of the three-dimensional structure of themolecule (145, 146). Such vaccines exhibit a reducedallergenic activity while retaining a capacity to elicitblocking IgG antibodies and T-cell recognition. Whilethis approach looks promising for vaccines administeredparenterally, a more appropriate strategy for SLIT israther to rely upon recombinant allergens presented in themost native conformation (147), to allow IgE-mediatedtargeting and capture of allergens by FCeRI+ DCs in theoral mucosa of atopic patients.
Strategies based on biological or synthetic adjuvantsand formulations to improve allergen presentation to oralLangerhans cells should be investigated to enhance SLITefficacy, reduce allergen dosing, and simplify immuniza-tion schemes. T reg adjuvants specially designed for thesublingual route may (i) target the allergen toLangerhans-like cells; (ii) trigger the production of IL-10 and/or TGF-b by antigen-presenting cells whileenhancing their capacity to migrate to local lymph nodesto ensure T cell priming; and (iii) facilitate the polariza-tion of T-cell responses towards regulatory T cells byengaging surface receptors such as ICOS, CD46 orspecific Toll-like receptors (e.g. TLR4).
Among many advantages, such molecularly definedvaccines will facilitate the identification of immunologicalcorrelates of clinical efficacy. Such biological surrogatemarkers would be extremely useful to ease the develop-ment of future vaccines, most particularly with the long-term goal of making vaccines tailored to patient-specificallergen sensitization patterns determined on the basis ofcomponent-resolved diagnostic. Importantly, whereas inthe past the immunological follow-up of immunotherapytrials focused on humoral (IgE/IgG4) responses, toolsand methods should rather be developed to monitor indetail allergen-specific CD4+ T-cell responses [e.g. usinghuman leukocyte antigen (HLA) class II-peptide solubletetramers, quantitative Elispot measurement of Th1/Th2/T reg cytokine-producing T cells, real-time PCR analysisof early events in T-cell polarization, etc.].
Altogether a better understanding of immune mecha-nisms involved in allergen-specific immunotherapy, com-bined with the power of molecular engineering andinnovative antigen delivery systems should offer theopportunity to rationally design second-generationrecombinant vaccines specifically tailored for the sublin-gual route. The development of such second-generationvaccines should help to confirm (or not) current workinghypotheses, to improve vaccine efficacy, and to evaluatesimpler administration schemes.
Acknowledgments
The authors wish to thank Danielle Michel for excellent secretarialassistance and Patrick Buchoux for help in preparing the graphics.
Moingeon et al.
160

References
1. Noon L. Prophylactic inoculationagainst hay fever. Lancet 1911;2:1572–1573.
2. Freeman J. Further observations on thetreatment of hay fever by hypodermicinoculations of pollen vaccine. Lancet1911;2:814–817.
3. Bousquet J, Lockey R, Malling HJ.World Health Organization positionpaper. Allergen immunotherapy: thera-peutical vaccines for allergic diseases.Allergy 1998;53(Suppl.):20–27.
4. Larche M. Specific immunotherapy.Brit Med Bull 2000;56:1019–1036.
5. Gehlhar K, Schlaak M, Becker W, BufeA. Monitoring allergen immunotherapyof pollen-allergic patients: the ratio ofallergen-specific IgG4 to IgG1 corre-lates with clinical outcome. Clin ExpAllergy 1999;29 (4):497–506.
6. Michils A, Mairesse M, Ledent C,Gossart B, Baldassare S, Duchateau J.Modified antigenic reactivity of anti-phospholipase A IgG antibodies inpatient allergic to bee venom: conver-sion with immunotherapy and relationto subclass expression. J Allergy ClinImmunol 1998;102:118–126.
7. Pichler CE, Helbling A, Pichler WJ.Three years of specific immunotherapywith house-dust-mite extracts in pa-tients with rhinitis and asthma: signifi-cant improvement of allergen-specificparameters and of non-specific bron-chial hyperreactivity. Allergy2001;56:301–306.
8. Moller C, Dreborg S, Ferdousi HA,Halken S, Host A, Jacobsen L et al.Pollen immunotherapy reduces thedevelopment of asthma in children withseasonal rhinoconjunctivitis (the PAT-Study). J Allergy Clin Immunol2002;109:251–256.
9. Canonica GW, Passalacqua G. Nonin-jection routes for immunotherapy.J Allergy Clin Immunol 2003;111:437–448.
10. Bousquet J, Scheinmann P, GuinnepainMT, Perrin-Fayolle M, Sauvaget J,Tonnel AB et al. Sublingual-swallowimmunotherapy (SLIT) in patients withasthma due to house-dust mites: adouble-blind, placebo-controlled study.Allergy 1999;54:249–260.
11. La Rosa M, Ranno C, Andre C, CaratF, Tosca MA, Canonica GW. Double-blind placebo-controlled evaluation ofsublingual-swallow immunotherapywith standardized Parietaria judaicaextract in children with allergic rhino-conjunctivitis. J Allergy Clin Immunol1999;104:425–432.
12. Wilson D, Torres Lima M, Durham S.Sublingual immunotherapy for allergicrhinitis. Cochrane Database Syst Rev2003;2:CD002893.
13. Clavel R, Bousquet J, Andre C. Clinicalefficacy of sublingual-swallowimmunotherapy: a double-blind, pla-cebo-controlled trial of a standardizedfive-grass-pollen extract in rhinitis.Allergy 1998;53:493–498.
14. Vourdas D, Syrigou E, Potamianou P,Carat F, Batard T, Andre C et al.Double-blind placebo controlled evalu-ation of sublingual immunotherapywith standardized olive pollen extract inpediatric patients with allergic rhino-conjunctivitis and mild asthma due toolive pollen sensitization. Allergy1998;53:662–672.
15. Bousquet J, Van Cauwenberge P,Khaltaev N. Aria workshop group,World Health Organization. Allergicrhinitis and its impact on asthma.J Allergy Clin Immunol2001;108(Suppl.):S147–S334.
16. Bousquet J. Sublingual immunothera-py: from proven prevention to putativerapid relief of allergic symptoms.Allergy 2005;60:1–3.
17. Frew AJ, Smith HE. Sublingual immu-notherapy. J Allergy Clin Immunol2001;107:441–444.
18. Pajno GB, Morabito L, Barberio G.Clinical and immunologic effects oflong-term sublingual immunotherapy inasthmatic children sensitized to mites: adouble-blind, placebo-controlled study.Allergy 2000;55:842–849.
19. Canonica GW, Compalati E, FumagalliF, Passalacqua G. Sublingual and oralimmunotherapy. Immunol Allergy ClinNorth Am 2004;24:685–704.
20. Marogna MM, Spadolini I, Massolo A,Canonica GW, Passalacqua G. Rand-omized controlled open study of sub-lingual immunotherapy for respiratoryallergy in real-life: clinical efficacy andmore. Allergy 2004;59:1205–1210.
21. Pajno GB, Vita D, Parmiani S, CaminitiL, La Grutta S, Barberio G. Impact ofsublingual immunotherapy on seasonalasthma and skin reactivity in childrenallergic to Parietaria pollen treated withinhaled fluticasone propionate. ClinExp Allergy 2003;33:1641–1647.
22. Passalacqua G, Guerra L, Pasquali M,Lombardi C, Canonica GW. Efficacyand safety of sublingual immunothera-py. Ann Allergy Asthma Immunol2004;93:3–12.
23. Torres Lima M, Wilson D, Pitkin L,Roberts A, Nouri-Aria K, Jacobson Met al. Grass pollen sublingual immuno-therapy for seasonal rhinoconjunctivi-tis: a randomized controlled trial. ClinExp Allergy 2002;32:507–514.
24. Troise C, Voltoni S, Canessa A, PecoraS, Negrini AC. Sublingual immuno-therapy in Parietaria pollen-inducedrhinitis: A double-blind study. J InvestAllergol Clin Immunol 1995;5:25–30.
25. Andre C, Perrin-Fayolle M, GrosclaudeM, Couturier P, Basset D, Cornillon Jet al. A double-blind placebo-controlledevaluation of sublingual immunothera-py with a standardized ragweed extractin patients with seasonal rhinitis. IntArch Allergy Immunol 2003;131:111–118.
26. Wilson DR, Torres Lima M, DurhamSR. Sublingual immunotherapy forallergic rhinitis: systematic review andmeta-analysis. Allergy 2005;60:4–12.
27. Agostinis F, Tellarini L, Canonica GW,Falagiani P, Passalacqua G. Safety ofsublingual immunotherapy with amonomeric allergoid in very youngchildren. Allergy 2005;60:133–138.
28. Grosclaude M, Bouillot P, Alt R,Leynadier F, Scheinmann P, Rufin Pet al. Safety of various dosage regimensduring induction of sublingual immu-notherapy. Int Arch Allergy Immunol2002;129:248–253.
29. Akdis CA, Blaser K. Mechanisms ofallergen-specific immunotherapy.Allergy 2000;55:522–530.
30. Durham SR, Till SJ. Immunologicchanges associated with allergen immu-notherapy. J Allergy Clin Immunol1998;102:157–164.
31. Till SJ, Francis JN, Nouri-Aria K,Durham SR. Mechanisms of immuno-therapy. J Allergy Clin Immunol2004;113:1025–1034.
32. Ebner C. Systemic immune response tospecific immunotherapy. Clin ExpAllergy 1998;28:781–783.
33. Garcia NM, Lynch NR, Di Prisco C.Lopez RI. Nonspecific changes inimmunotherapy with house dust ex-tract. J Invest Allergol Clin Immunol1995;5:8–24.
34. Lichtenstein LM, Ishizaka K, NormanPS, Sobotka AK, Hill BM. IgE anti-body measurements in ragweed hay fe-ver. Relationship to clinical severity andthe results of immunotherapy. J ClinInvest 1973;52:472–482.
161
Immune mechanisms of SLIT

35. Pierson-Mullany LK, Jackola D,Blumenthal M, Rosenberg A. Alteredallergen binding capacities of Amb a1-specific IgE and IgG4 from ragweed-sensitive patients receiving immuno-therapy. Ann Allergy Asthma Immunol2000;84:241–243.
36. Djurup R, Malling HJ. High IgG4antibody level is associated with failureof immunotherapy with inhalant aller-gens. Clin Allergy 1987;17:459–68.
37. Hakansson L, Heinrich C, Rak S,Venge P. Activation of B-lymphocytesduring pollen season. Effect of immu-notherapy. Clin Exp Allergy1998;28:791–798.
38. Garcia BE, Sanz ML, Gato JJ,Fernandez J, Oehling A. IgG4 blockingeffect on the release of antigen-specifichistamine. J Investig Allergol ClinImmunol 1993;3:26–33.
39. Mothes N, Heinzkill M, DrachenbergKJ, Sperr WR, Krauth MT, Majlesi Yet al. Allergen-specific immunotherapywith a monophosphoryl lipid A-adju-vanced vaccine: reduced seasonnalyboosted immunoglobulin E productionand inhibition of basophil histaminerelease by therapy-induced blockingantibodies. Clin Exp Allergy2003;33:1198–1208.
40. Niederberger V, Horak F, Vrtala S,Spitzauer S, Krauth MT, Valent P et al.Vaccination with genetically engineeredallergens prevents progression of aller-gic disease. Proc Natl Acad Sci USA2004;101(Suppl. 2):14677–14682.
41. Svenson M, Jacobi HH, Bodtger U,Poulsen LK, Rieneck K, Bendtzen K.Vaccination for birch pollen allergy.Induction of affinity-matured or block-ing IgG antibodies does not account forthe reduced binding of IgE to Bet v 1.Mol Immunol 2003;39:603–612.
42. Wachholz PA, Durham SR. Inductionof �blocking� IgG antibodies during im-munotherapy. Clin Exp Allergy2003;33:1171–1174.
43. Flicker S, Valenta R. Renaissance of theblocking antibody concept in type Iallergy. Int Arch Allergy Immunol2003;132:13–24.
44. Jakobsen CG, Bodiger U, Poulsen LK,Rogeen EL. Vaccination for birch pollenallergy: comparison of the affinities ofspecific immunoglobulins E, G1, and G4measured by surface plasmon resonance.Clin Exp Allergy 2005;36:193–198.
45. Bahceciler N, Arikan C, Taylor A,Akdis M, Blaser K, Barlan I et al. Im-pact of sublingual immunotherapy onspecific antibody levels in asthmaticchildren allergic to house dust mite.Intern Arch Immunol Allergy2005;136:287–294.
46. Rolinck-Werninghaus C, Kopp M,Liebke C, Lange J, Wahn U,Niggemann B. Lack of detectablealterations in immune responses duringsublingual immunotherapy in childrenwith seasonal allergic rhinoconjunctivi-tis to grass pollen. Int Arch AllergyImmunol 2005;136:134–141.
47. Malling HJ, Lund L, Ipsen H, PoulsenLK. Safety and immunological changesduring tablet based specific immuno-therapy. J Allergy Clin Immunol2005;115:S161 (abstract).
48. Van Neerven RJ, Wikborg T, Lund G,Jacobsen B, Brinch-Nielsen A, Arnved Jet al. Blocking antibodies induced byspecific allergy vaccination prevent theactivation of CD4+ T cells by inhibitingserum-IgE-facilitated allergen presenta-tion. J Immunol 1999;163:2944–2952.
49. Van Neerven RJ, Arvidsson M, IpsenH, Sparholt SH, Rak S, Wurtzen PA. Adouble-blind, placebo-controlled birchallergy vaccination study: inhibition ofCD23-mediated serum-immunoglobulinE-facilitated allergen presentation. ClinExp Allergy 2004;34:420–428.
50. Akdis C, Blaser K, Akdis M. Genes oftolerance. Allergy 2004;59:897–913.
51. Iliopoulos O, Proud D, Adkinson NFJr, Creticos PS, Norman PS, Kagey-Sobotka A et al. Effects of immuno-therapy on the early, late, andrechallenge nasal reaction to provoca-tion with allergen: changes in inflam-matory mediators and cells. J AllergyClin Immunol 1991;87:855–866.
52. Hakansson L, Heinrich C, Rak S,Venge P. Priming of eosinophil adhe-sion in patients with birch pollen allergyduring pollen season: effect of immu-notherapy. J Allergy Clin Immunol1997;99:551–562.
53. Durham SR, Ying S, Varney VA,Jacobson MR, Sudderick RM, MackayIS et al. Grass pollen immunotherapyinhibits allergen-induced infiltration ofCD4+ T lymphocytes and eosinophilsin the nasal mucosa and increases thenumber of cells expressing messengerRNA for interferon-c. J Allergy ClinImmunol 1996;97:1356–1365.
54. Durham SR, Varney VA, Gaga M,Jacobson MR, Varga EM, Frew AJet al. Grass pollen immunotherapydecreases the number of mast cells in theskin. Clin Exp Allergy 1999;29:1490–1496.
55. Monteseirin J, Bonilla I, Camacho J,Conde J, Sobrino F. Elevated secretionof myeloperoxidase by neutrophils fromasthmatic patients: the effect of immu-notherapy. J Allergy Clin Immunol2001;107:623–626.
56. Polosa R, Li Gotti F, Mangano G,Paolino G, Mastruzzo C, Vancheri Cet al. Effect of immunotherapy onasthma progression, BHR and sputumeosinophils in allergic rhinitis. Allergy2004;59:1224–1228.
57. Wilson DR, Nouri-Aria KT, WalkerSM, Pajno GB, O’Brien F, JacobsonMR et al. Grass pollen immunotherapy:symptomatic improvement correlateswith reductions in eosinophils and IL-5mRNA expression in the nasal mucosaduring the pollen season. J Allergy ClinImmunol 2001;107:971–976.
58. Wilson DR, Irani AM, Walker SM,Jacobson MR, Mackay IS, SchwartzLB et al. Grass pollen immunotherapyinhibits seasonal increases in basophilsand eosinophils in the nasal epithelium.Clin Exp Allergy 2001;31:1705–1713.
59. Furin MJ, Norman PS, Creticos PS,Proud D, Kagey-Sobotka A,Lichtenstein LM et al. Immunotherapydecreases antigen-induced eosinophilcell migration into the nasal cavity.J Allergy Clin Immunol 1991;88:27–32.
60. Polosa R, Li Gotti F, Mangano G,Mastruzzo C, Pistorio MP, Crimi N.Monitoring of seasonal variability inbronchial hyper-responsiveness andsputum cell counts in non-asthmaticsubjects with rhinitis and effect of spe-cific immunotherapy. Clin Exp Allergy2003;33:873–881.
61. Fanta C, Bohle B, Hirt W, Siemann U,Horak F, Kraft D et al. Systemicimmunological changes induced byadministration of grass pollen allergensvia the oral mucosa during sublingualimmunotherapy. Int Arch AllergyImmunol 1999;120:218–224.
62. Marcucci F, Sensi L, Frati F, SennaGE, Caninica GW, Parmiani S et al.Sublingual tryptase and ECP in childrentreated with grass pollen sublingualimmunotherapy (SLIT): safety andimmunologic implications. Allergy2001;56:1091–1095.
63. Passalacqua G, Albano M, FregoneseL, Riccio A, Pronzato C, Mela GS et al.Randomised controlled trial of localallergoid immunotherapy on allergicinflammation in mite-induced rhino-conjunctivitis. Lancet 1998;351:629–632.
64. Marcucci F, Frati F, Bernardini R,Novembre E, Barbato A, Pecora S.Effects on inflammation parameters of adouble blind, placebo controlled one-year course of SLIT in children mono-sensitized to mites. Allergy2003;58:657–662.
162
Moingeon et al.

65. Silvestri M, Spallarossa D, Battistini Eet al. Changes in inflammatory andclinical parameters and in bronchialhyperreactivity in asthmatic childrensensitized to house dust mites followingsublingual immunotherapy. J InvestAllergol Clin Immunol 2002;12:52–59.
66. Passalacqua G, Albano M, Riccio A,Fregonese L, Puccinelli P, Parmiani Set al. Clinical and immunologic effectsof a rush sublingual immunotherapy toParietaria species: a double-blind, pla-cebo-controlled trial. J Allergy ClinImmunol 1999;104:964–968.
67. El Biaze M, Boniface S, Koscher V,Mamessier E, Dupuy P, Milhe F et al. Tcell activation, from atopy to asthma:more a paradox than a paradigm.Allergy 2003;58:844–853.
68. Mavroleon D. Restoration of cytokineimbalance by immunotherapy. Clin ExpAllergy 1998;28:917–920.
69. Hamid QA, Schotman E, JacobsonMR, Walker SM, Durham SR. Increa-ses in IL-12 messenger RNA+ cellsaccompany inhibition of allergen-in-duced late skin responses after success-ful grass pollen immunotherapy.JAllergyClin Immunol 1997;99:254–260.
70. Benjaponpitak S, Oro A, Maguire P,Marinkovich V, DeKruyff H, UmetsuDT. The kinetics of change in cytokineproduction by CD4+ T cells duringconventional allergen immunotherapy.J Allergy Clin Immunol 1999;103:468–475.
71. Ebner C, Siemann U, Bohle B, WillheimM, Wiedermann U, Schenk S et al.Immunological changes during specificimmunotherapy of grass pollen allergy:reduced lymphoproliferative responsesto allergen and shift from Th2 to Th1 inT-cell clones specific for Phl p 1, a majorgrass pollen allergen. Clin Exp Allergy1997;27:1007–1015.
72. Secrist H, Chelen CJ, Wen Y, MarshallJD, Umetsu DT. Allergen immuno-therapy decreases IL-4 production inCD4+ T cells from allergic individuals.J Exp Med 1993;178:2123–2130.
73. Laaksonen K, Junikka M, Lahesmaa R,Terho EO, Savolainen J. In vitro aller-gen-induced mRNA expression of sign-aling lymphocytic activation moleculeby PBMC of patients with allergic rhi-nitis is increased during specific pollenimmunotherapy. J Allergy Clin Immu-nol 2003;112:1171–1177.
74. Meissner N, Kochs S, Coutelle J,Kussebi F, Baumgarten C, LowensteinH et al. Modified T-cell activation pat-tern during specific immunotherapy(SIT) in cat-allergic patients. Clin ExpAllergy 1999;29:618–625.
75. Majori M, Caminati A, Corradi M,Brianti E, Scarpa S, Pesci A. T-cellcytokine pattern at three time pointsduring specific immunotherapy formite-sensitive asthma. Clin Exp Allergy2000;30:341–347.
76. Gabrielsson S, Soderlund A, Paulie S,van der Pouw Kraan TC, Troye-Blomberg M, Rak S. Specific immuno-therapy prevents increased levels ofallergen-specific IL-4 and IL-13-produ-cing cells during pollen season. Allergy2001;56:293–300.
77. Guerra F, Carracedo J, Solana-Lara R,Sanchez-Guijo P, Ramirez R. Th2lymphocytes from atopic patients trea-ted with immunotherapy undergo rapidapoptosis after culture with specificallergens. J Allergy Clin Immunol2001;107:647–653.
78. Faith A, Richards DF, Verhoef A,Lamb JR, Lee TH, Hawrylowicz CM.Impaired secretion of interleukin-4 andinterleukin-13 by allergen-specific Tcells correlates with defective nuclearexpression of NF-AT2 and jun B: rele-vance to immunotherapy. Clin ExpAllergy 2003;33:1209–1215.
79. Oldfield WL, Larche M, Kay AB. Effectof T-cell peptides derived from Fel d 1on allergic reactions and cytokine pro-duction in patients sensitive to cats: arandomised controlled trial. Lancet2002;360:47–53.
80. O’Brien RM, Byron KA, Varigos GA,Thomas WR. House dust mite immu-notherapy results in a decrease in Der p2-specific IFN-c and IL-4 expression bycirculating T lymphocytes. Clin ExpAllergy 1997;27:46–51.
81. Wachholz PA, Nouri-Aria K, WilsonDR, Walker SM, Verhoef A, Till SJet al. Grass pollen immunotherapy forhayfever is associated with increases inlocal nasal but not peripheral Th1:Th2cytokine ratios. Immunology2002;105:56–62.
82. Muller U, Akdis CA, Fricker M, AkdisM, Blesken T, Bettens F et al. Success-ful immunotherapy with T-cell epitopepeptides of bee venom phospholipaseA2 induces specific T-cell anergy in pa-tients allergic to bee venom. J AllergyClin Immunol 1998;101:747–754.
83. Jutel M, Akdis M, Budak F, Aebischer-Casaulta C, Wrzyszcz M, Blaser K et al.IL-10 and TGF-b cooperate in the reg-ulatory T cell response to mucosalallergens in normal immunity and spe-cific immunotherapy. Eur J Immunol2003;33:1205–1214.
84. Gardner L, Thien F, Douglass J,Rolland J, O’Hehir R. Induction of Tregulatory cells by standardized housedust mite immunotherapy – an increasein CD4+CD25+ interleukin 10+ Tcells expressing peripheral tissue traf-ficking markers. Clin Exp Allergy2004;34:1209–1219.
85. Taylor A, Verhagen J, Akdis CA, AkdisM. T regulatory cells in allergy andhealth: a question of allergen specificityand balance. Int Arch Allergy Immunol2004;135:73–82.
86. Ippoliti F, De Santis W, Volterrani A,Lenti L, Canitano N, Lucarelli S et al.Immunomodulation during sublingualtherapy in allergic children. PediatrAllergy Immunol 2003;14:216–221.
87. Fenoglio D, Puppo F, Cirillo I,Vizzaccaro A, Ferrera A, Tosca MAet al. Sublingual specific immunothera-py reduces PBMC proliferations. EurAnn Allergy Clin Immunol2005;37:147–151.
88. Ciprandi G, Fenoglio D, Cirillo I,Vizzaccaro A, Ferrero A, Tosca MAet al. Sublingual HDM-specific immu-notherapy induces IL10 production:Preliminary report. Ann Allergy Asth-ma Immunol 2005;95:38–44.
89. Sakaguchi S, Sakaguchi N, Asano M,Itoh M, Toda M. Immunologic self-tolerance maintained by activated Tcells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a singlemechanism of self-tolerance causes var-ious autoimmune diseases. J Immunol1995;155:1151–1164.
90. Blaser K, Akdis C. Interleukin 10, Tregulatory cells and specific allergytreatment. Clin Exp Allergy2004;34:328–331.
91. Groux H, O’Garra A, Bigler M,Rouleau M, Antonenko S, de Vries JEet al. A CD4+ T-cell subset inhibitsantigen-specific T-cell responses andprevents colitis. Nature 1997;389:737–742.
92. Umetsu DT, Akbari O, DeKruyff RH.Regulatory T cells control the develop-ment of allergic disease and asthma.J Allergy Clin Immunol 2003;112:480–487.
93. Jonuleit H, Schmitt E. The regulatory Tcell family: distinct subsets and theirinterrelations. J Immunol 2003;171:6323–6327.
94. Hawrylowicz CM, O’Garra A. Potentialrole of interleukin-10 secreting regula-tory T cells in allergy and asthma.Nature Rev Immunol 2005;5:271–283.
163
Immune mechanisms of SLIT

95. Vieira PL, Christensen JR, Minaee S,O’Neill EJ, Barrat FJ, Boonstra A et al.IL-10-secreting regulatory T cells do notexpress Foxp3 but have comparableregulatory function to naturally occur-ring CD4+CD25+ regulatory T cells.J Immunol 2004;172:5986–5993.
96. Akbari O, DeKruyff RH, Umetsu DT.Pulmonary dendritic cells producingIL-10 mediate tolerance induced byrespiratory exposure to antigen. NatImmunol 2001;2:725–731.
97. Wakkach A, Fournier N, Brun V,Breittmayer JP, Cottrez F, Groux H.Characterization of dendritic cells thatinduce tolerance and T regulatory 1 celldifferentiation in vivo. Immunity2003;18:605–617.
98. Akdis CA, Blesken T, Akdis M,Wuthrich B, Blaser K. Role of inter-leukin 10 in specific immunotherapy.J Clin Invest 1998;102:98–106.
99. Yamagiwa S, Gray JD, Hashimoto S,Horwitz DA. A role for TGF-beta inthe generation and expansion ofCD4+CD25+ regulatory T cells fromhuman peripheral blood. J Immunol2001;166:7282–7289.
100. Fontenot JD, Gavin MA, RudenskyAY. Foxp3 programs the developmentand function of CD4+CD25+ regula-tory T cells. Nat Immunol 2003;4:330–336.
101. Levings MK, Sangregorio R, GalbiatiF, Squadrone S, de Waal Malefyt R,Roncarolo MG. IFN-alpha and IL-10induce the differentiation of human type1 T regulatory cells. J Immunol2001;166:5530–5539.
102. Barrat FJ, Cua DJ, Boonstra A,Richards DF, Crain C, Savelkoul HFet al. In vitro generation of interleukin10-producing regulatory CD4(+) Tcells is induced by immunosuppressivedrugs and inhibited by T helper type 1(Th1)- and Th2-inducing cytokines.J Exp Med 2002;195:603–616.
103. Kemper C, Chan AC, Green JM, BrettKA, Murphy KM, Atkinson JP. Acti-vation of human CD4+ cells with CD3and CD46 induces a T-regulatory cell 1phenotype. Nature 2003;421:388–392.
104. Stassen M, Fondel S, Bopp T, RichterC, Muller C, Kubach J et al. HumanCD25+ regulatory T cells: two subsetsdefined by the integrins alpha 4 beta 7or alpha 4 beta 1 confer distinct sup-pressive properties upon CD4+ Thelper cells. Eur J Immunol2004;34:1303–1311.
105. Weiner HL. Induction and mechanismof action of transforming growth factor-beta-secreting Th3 regulatory cells.Immunol Rev 2001;182:207–214.
106. Yazdanbakhsh M, Kremsner PG, VanRee R. Allergy, parasites and the hy-giene hypothesis. Science 2002;296:490–494.
107. Gambineri E, Torgerson T, Ochs H.Immune dysregulation, polyendocrin-opathy, enteropathy, and X-linkedinheritance (IPEX), a syndrome of sys-temic autoimmunity caused by muta-tions of Foxp3, a critical regulator of Tcell homeostasis. Curr Opin Rhumatol2003;15:430–435.
108. Ling EM, Smith T, Nguyen XD,Pridgeon C, Dallman M, Arbery Jet al. Relation of CD4+CD25+
regulatory T-cell suppression ofallergen-driven T-cell activation toatopic status and expression of allergicdisease. Lancet 2004;363:608–615.
109. Grindebacke H, Wing K, AnderssonAC, Suri-Payer E, Rak S, Rudin A.Defective suppression of Th2 cytokinesby CD4+CD25+ regulatory T cells inbirch allergics during birch pollen sea-son. Clin Exp Allergy 2004;34:1364–1372.
110. Gentile D, Schreiber R, Howe-Adams J,Trecki J, Patel A, Angelini B et al.Diminished dendritic cell interleukin 10production in atopic children. Ann Al-lergy Asthma Immunol 2004;92:538–544.
111. Akdis M, Verhagen J, Taylor A,Karamloo F, Karagiannidis C, CrameriR et al. Immune responses in healthyand allergic individuals are character-ized by a fine balance between allergen-specific T regulatory 1 and T helper 2cells. J Exp Med 2004;199:1567–1575.
112. Nasser S, Ying S, Meng Q, Kay AB,Ewan PW. Interleukin 10 levels increasein cutaneous biopsies of patientsundergoing wasp venom immunothera-py. Eur J Immunol 2001;31:3704–3713.
113. Akdis C, Joss A, Akdis M, Blaser K.Mechanisms of IL10 induced T cellinactivation in allergic inflammationand normal response to allergens. IntArch Allergy Immunol 2001;124:180–182.
114. Bullens DM, Van Den KC, Dilissen E,Kasran A, Ceuppens JL. Allergen-spe-cific T cells from birch-pollen-allergicpatients and healthy controls differ in Thelper 2 cytokine and in interleukin-10production. Clin Exp Allergy2004;34:879–887.
115. Nouri-Aria KT, Wachholz PA, FrancisJN, Jacobson MR, Walker SM,Wilcock LK et al. Grass pollen immu-notherapy induces mucosal and periph-eral IL-10 responses and blocking IgGactivity. J Immunol 2004;172:3252–3259.
116. Faria AM, Weimer HL. Oral tolerance:mechanisms and therapeutic applica-tions. Adv Immunol 1999;73:153–264.
117. Noirey N, Rougier N, Andre C, SchmittD, Vincent C. Langerhans like dendriticcells generated from cord blood pro-genitors internalize pollen allergens bymacropinocytosis, and part of mole-cules are processed and can activateautologous naive T lymphocytes. J Al-lergy Clin Immunol 2000;105:1194–1201.
118. Van Wilsem EJ, Breve J, Savelkoul H,Claessen A, Scheper RJ, Kraal G. Oraltolerance is determined at the level ofdraining lymph nodes. Immunobiology1995;194:403–414.
119. Aoyama-Kondo T, Yoshida T, TsobeK, Nakayama A, Asai J, Oka T et al.Characterization of antibody responsesof local lymph nodes to antigen givenunder the oral submucosa. Immunobi-ology 1992;184:372–383.
120. Van Helvoort JM, Samsom J, ChantryD, Jansen W, Schadee-Eestermans I,Thepen T et al. Preferential expressionof IgG2b in nose draining cervicallymph nodes and its putative role inmucosal tolerance induction. Allergy2004;59:1211–1218.
121. Martin-Fotecha A, Sebastiani S,Hopken U, Uguccioni M, Lipp M,Lanzavecchia A et al. Regulation ofdendritic cell migration to the draininglymph node: impact on T lymphocytetraffic and priming. J Exp Med2003;198:615–621.
122. Allam JP, Novak N, Fuchs C, Asen S,Berge S, Appel T et al. Characterizationof dendritic cells from human oralmucosa: a new Langerhans cell typewith high constitutive FC epsilon RIexpression. J Allergy Clin Immunol2003;112:141–148.
123. von Bubnoff D, Fimmers R, BogdanovM, Matz H, Koch S, Bieber T.Asymptomatic atopy is associated withincreased indoleamine 2,3-dioxygenaseactivity and interleukin 10 productionduring seasonal allergen exposure. ClinExp Allergy 2004;34:1056–1063.
124. Passalacqua G, Villa G, Altrinetti V,Falagiani P, Canonica GW, Mariani Get al. Sublingual swallow or spit?Allergy 2001;56:578–582.
125. Noonan PK, Benet LZ. Incomplete anddelayed bioavailability of sublingualnitroglycerin. Am J Cardiol1985;55:184–187.
126. Cleary JF. Pharmacokinetic and phar-macodynamic issues in the treatment ofbreakthrough pain. Semin Oncol1997;24(Suppl. 16):S13-S19.
164
Moingeon et al.

127. Senel S, Kremer M, Nagy K, Squier C.Delivery of bioactive peptides and pro-teins across oral (buccal) mucosa. CurrPharm Biotechnol 2001;2:175–186.
128. Falagiani P, Mistrello G. Pharmacoki-netics of allergens after local adminis-tration. Allergy 1997;52:17–21.
129. Bagnasco M, Mariani G, PassalacquaG, Motta C, Bartolomei M, Falagiani Pet al. Absorption and distribution kin-etics of the major Parietaria judaicaallergen (Par j 1) administered by non-injectable routes in healthy humanbeings. J Allergy Clin Immunol1997;100:122–129.
130. Bagnasco M, Passalacqua G, Villa G,Augeri C, Flamigni G, Borini E et al.Pharmacokinetics of an allergen and amonomeric allergoid for oromucosalimmunotherapy in allergic volunteers.Clin Exp Allergy 2001;31:54–60.
131. Walker GF, Langoth N, Bernkop-Schnurch A. Peptidase activity on thesurface of the porcine buccal mucosa.Int J Pharm 2002;233:141–147.
132. Igea JM, Cuevas M, Lazaro M, QuirceS, Cuesta J. Susceptibility of a grass-pollen oral immunotherapy extract tothe saliva and gastric fluid digestiveprocess. Allergol Immunopathol(Madr) 1994;22:55–59.
133. Einarsson R, Renck B, Taudorf E. Invitro studies of degradation of birch andtimothy pollen allergen preparations byhuman duodenal juice. Allergy1988;43:469–472.
134. Smith H, White P, Annila I, Poole J,Andre C, Frew A. Randomized con-trolled trial of high-dose sublingual im-munotherapy to treat seasonal allergicrhinitis. J Allergy Clin Immunol2004;114:831–837.
135. Voltolini S, Modena P, Minale P,Bignardi D, Troise C, Puccinelli P et al.Sublingual immunotherapy in tree pol-len allergy. Double-blind, placebo-con-trolled study with a biologicallystandardised extract of three pollens(alder, birch and hazel) administered bya rush schedule. Allergol Immunopa-thol (Madr) 2001;29:103–110.
136. Di Rienzo V, Marcucci F, Puccinelli P,Parmiani S, Frati F, Sensi L. Longlasting effect of sublingual immuno-therapy in children with asthma due tohouse dust mites. A 10 year prospectivestudy. Clin Exp Allergy 2003;33:206–210.
137. Novembre E, Galli E, Landi F,Caffarelli C, Pifferi M, De Marco Eet al. Coseasonal sublingual immuno-therapy reduces the development ofasthma in children with allergic rhino-conjunctivitis. J Allergy Clin Immunol2004;114:851–857.
138. Taams LS, van Eden W, Wauben MH.Dose dependent induction of distinctphenotypes: multiple level of T-cellanergy. J Immunol 1999;162:1974–1981.
139. Hosken NA, Shibuya K, Heath A,Murphy K, O’Garra A. The effect ofantigen dose on CD4+ T helper cellphenotype development in a T cellreceptor a/b transgene model. J ExpMed 1995;182:1579–1584.
140. Gardner L, Spyroglou L, O’Hehir R,Rolland J. Increased allergen concen-tration enhances IFN-c production byallergic donor T cells expressing a per-ipheral tissue trafficking phenotype.Allergy 2004;59:1308–1317.
141. Sprent J, Surch CD. Generation andmaintenance of memory T cells. CurrOpin Immunol 2001;13:248–254.
142. Marcucci F, Sensi L, Di Cara G, GidaroG, Incorvaia C, Frati F. Sublingualreactivity to rBet v1 and r Phl p1 inpatient with oral allergy syndrome. Int JPharmacol (in press).
143. Gonzalez-Espinosa C, Odom S, OliveraA, Hobson JP, Martinez ME, Oliveira-Dos-Santos A et al. Preferential signal-ling and induction of allergy-promotinglymphokines upon weak stimulation ofthe high affinity IgE receptor on mastcells. J Exp Med 2003;11:1453–1465.
144. Holst-Larsen T, Poulsen LK, Melac M,Combebias A, Andre C, Malling H-J. Aphase I-dose finding study to investigatethe safety and tolerability of grass pol-len sublingual immunotherapy given inincremental or in constant dosing regi-mens. J Allergy Clin Immunol2005;S1:529 (abstract).
145. Valenta R. The future of antigen-speci-fic immunotherapy of allergy. NatureRev Immunol 2002;2:446–453.
146. Mahler V, Vrtala S, Kuss O, DiepgenTL, Suck R, Cromwell O et al. Vaccinesfor birch pollen allergy based on gen-etically engineered hypoallergenicderivatives of the major birch pollenallergen, Bet v1. Clin Exp Allergy2004;34:115–122.
147. Batard T, Didierlaurent A, Chabre H,Mothes N, Bussieres L, Bohle B et al.Characterization of recombinant wild-type Bet v 1 as a candidate vaccineagainst birch pollen allergy. Int ArchAllergy Immunol 2005;136:239–249.
148. Francis JN, Till SJ, Durham SR.Induction of IL-10+ CD4+CD25+ Tcells by grass pollen immunotherapy.J Allergy Clin Immunol 2003;111:1255–1261.
165
Immune mechanisms of SLIT