algal colonies
TRANSCRIPT

Plant Physiol. (1988) 87, 603-6080032-0889/88/87/0603/06/$0 1.00/0
Establishment of Control Parameters for in Situ, AutomatedScreening of Sustained Hydrogen Photoproduction by IndividualAlgal Colonies'
Received for publication December 7, 1987 and in revised form January 27, 1988
D. A. GRAVES2, M. E. REEVES, AND E. GREENBAUM*Chemical Technology Division, Oak Ridge National Laboratory,3 Oak Ridge, Tennessee 37831
ABSTRACT
An apparatus was constructed which allowed automated screening ofindividual microalgal colonies for sustained ability to photoevolve H2during anaerobic photosynthesis. The main components of this apparatuswere a microcomputer, a He-Ne laser mounted on a computer-controlledX-Y translation stage, a flow-through chamber which contained an agarplate of colonies, and a H2 detector which interfaced with the microcom-puter for data collection. The system was capable of detecting a minimumproduction rate of 1 nanomole of H2 per hour per colony and provided anefficient means of screening relatively large numbers of algal colonies.Examination of the effect of the spacing of colonies on the agar plate,light intensity, stability of colonies within a screening period, colony age,chlorophyll content, and colony size on H2 yield indicated that, underoptimum conditions, yields from genetically uniform colonies varied byno more than a factor of 2 in their H2-producing ability. Therefore,colonies of algae whose H2 yields lie outside this intrinsic twofoldvariability can be identified and selected as natural variants or mutants.A description of the construction and of the apparatus is presented, andthe experimental results used to establish the control parameters forChlamydomonas reinhardtii colonies are discussed.
reported (2, 4, 6, 12, 17, 20, 21), they are incapable of measuringsustained steady state rates of H2 production. Since a period of3 h is required for adaptation of light-driven H2 evolution and 1to 2 h is required for H2 photoevolution to approach steady state(plus additional time for reestablishment of baseline conditions)in the case of liquid culture, only one clone can be screened permeasuring apparatus per day. To circumvent this lengthy andtedious process, an apparatus was constructed and interfaced toa microcomputer which allowed single colonies on an agar platecontaining multiple colonies to be screened sequentially for theirH2-producing ability. This apparatus, particularly the H2-sensingcomponent (10, 13), is not hindered by the inherent limitationsof other H2 detection methods. It can be used to examinerelatively large numbers of algal colonies for inherent variationin their steady state H2-producing ability, to evaluate putativemutants that are defective or enhanced in H2-producing ability,and to compare H2 photoevolution from cells growing on solidversus liquid medium.
MATERIALS AND METHODS
Hydrogen evolution by microalgae represents a unique variantof photosynthesis in which the energy-rich photoproduct is mo-lecular H2, rather than reduced CO2. Since the pioneering dis-covery of Gaffron and Rubin (8), several genera of microalgaehave been shown to be capable of photoevolving H2 underanaerobic conditions (3, 4, 23). The recent awareness of soma-clonal variation in clones of higher plants (7) and the findingthat repeated cycles of anaerobic photosynthesis and regrowth ofChlamydomonas reinhardtii improved both H2 yield and toler-ance to anaerobiosis (18, 22) prompted interest in inherentvariability in H2 photoevolution among individual colonies ofmicroalgae.
Prior techniques for analyzing individual members of an algalpopulation required growth of each clone in liquid and analysisof the clones individually. Although amperometric electrode andmanometric techniques for measuring H2 have previously been
'Research supported by the Gas Research Institute under contract5083-260-0880 and by the Division ofChemical Sciences, Office of BasicEnergy Sciences, United States Department of Energy.
2 Research Associate, Botany Department, University of Tennessee,Knoxville, TN 37996.
'Operated by Martin Marietta Energy Systems, Inc., under contractDE-AC05-840R2 1400 with the United States Department of Energy.
Chlamydomonas reinhardtii (University ofTexas Culture Col-lection, strain 90 [mt-]) was grown on minimal medium, pH 6.8(18), under photoautotrophic conditions at about 12 W m-2 and250 C with 14 h/10 h light-dark cycles. Liquid cultures werediluted and plated on minimal agar plates to give 10 to 30colonies per plate. The Chl content of single colonies was deter-mined by excising a colony from the plate along with a smallpiece of agar and placing it into a 1.5-ml microcentrifuge tubewith 0.5 ml of methanol. The colony was usually white after 1 hof dark incubation at room temperature in methanol. Chl ex-traction from postscreened colonies was more difficult but wasachieved by heating the extraction mixture to 65° C for 5 min.Chl was quantitated using the extinction coefficients of Mac-Kinney (15, 18). Colony diameter was determined by using anocular micrometer and a dissecting microscope. The algae wereroutinely checked for contamination by plating dilutions ofliquid cultures onto tryptic soy agar.One 100 x 15 mm Petri dish was housed in a two-piece glass
chamber. The top of the chamber was recessed to enclose thedish and grooved around the perimeter to accept an 0-ring. Inletand outlet ports for gas exchange were built into the top. Thebottom was a flat piece of glass. An agar plate was inverted andplaced on the bottom, and the top was positioned over the plateand sealed with an 0-ring. The entire assembly was sealed andheld in place with three bolts that passed through the top of thesupport frame, the glass plate, and a clamp on the top (Fig. 1,top, bottom). The chamber was connected to a regulated Hesupply and vented through the H2 sensor. The He was humidifiedby bubbling it through a cell containing doubly distilled H20
603

Plant Physiol. Vol. 87, 1988
FIG. 1. Assembled sample chamber con-taining an inverted plate of algal colonies.Top, An overhead view of the assembledchamber showing the clamp used to securethe chamber and the position of the plateof colonies. The small points of light nearthe laser beam are photographic artifacts.Bottom, A lateral view showing the supportframe, the sample chamber, and the frontsurface mirror connected to the laser forreflecting the beam to a colony.
604 GRAVFES ET AL.

AUTOMATED COLONY SCREENING
before being passed through the chamber. He flow was heldconstant at 25 ml min-'.
Colonies were individually irradiated using a He-Ne laser(model 80-2HA, Coherent, Palo Alto, CA [633-nm wave-length]). The laser beam was positioned over a colony using aX-Y translation stage (Micro Controle) adapted to carry thelaser. As the H2 analyzer was sensitive to moisture and pressurechanges, moisture traps consisting of 1 3x molecular sieves (LindeCorp.) immersed in ice were installed upstream from the ana-lyzer. The copper tubing connecting the various pieces was driedwith compressed air before each experiment because water wouldcondense and build up at the unions and form bubbles thatcould cause detectable pressure changes. H2 evolution was quan-titated using a calibration curve generated by applying knownamounts of current to an in-line electrolysis cell (10) whichcontained 1 mM NaOH and platinum electrodes. All connectionswere made using Swagelok connectors, glass and brass 0-ringfittings, copper tubing, Viton 0-rings, and spring clamps withlocking nuts. The gas sensor was a Snoopy Ethylene Detector(Bio-gas Detector Corp., Okemos, MI) modified for continuousgas flow measurements. This technology, employing a Figaro812 gas sensing SnO2 semiconductor, has been previously appliedfor the detection ofcombustible gases in continuous flow systems(10, 13, 18) and gas chromatographic configurations (13, 16).The gas sensor/calibration cell assembly used for measuring thephotoproduced H2 was similar to that described earlier (9, 10).Data collection and laser positioning were automatically per-
formed using a Hewlett-Packard 85 microcomputer via an IEEE-488 interface. A Klinger programmable stepper motor controllerwas driven by the computer to position the laser on and offselected colonies at predetermined time intervals. The X-Y co-ordinates for each colony to be screened were entered into thecomputer by manually positioning the laser beam on the colony;the coordinates were transmitted from the stepper motor con-troller to the computer over the IEEE-488 interface. The analogoutput from the H2 sensor was displayed on a Keithley model197 multimeter and collected over the IEEE-488 bus. Data wererecorded and stored on tape cassettes and also recorded directlyfrom the multimeter's analog output on a chart recorder. (SeeFigure 2 for a schematic representation of the apparatus.)
After a plate of colonies had been placed in the chamber andthe system sealed, the colonies were subjected to 4 h of darknessat room temperature for adaptation. During this time, calibration
data were collected by supplying current steps to the electrolysiscell and recording the sensor response for each step. Calibrationswere typically run with 9, 7, 5, and 3 gamp for 45 min at eachcurrent. Following dark adaptation, single colonies were screenedwith the laser beam positioned on the colony for a minimum of1 h. After the light-on period, the laser was moved offthe colonyto a small piece of black felt for light off. Baseline conditionswere reestablished during the light-offtime (minimum dark time,1 h). The H2 yield of single colonies was quantitated using thecalibration data generated by electrolyzing water. Software waswritten to plot the raw data, calculate the rates of H2 evolution,and determine the integrated yield of H2 during each light-onperiod. This software, written in BASIC language, is availableupon request.
RESULTS
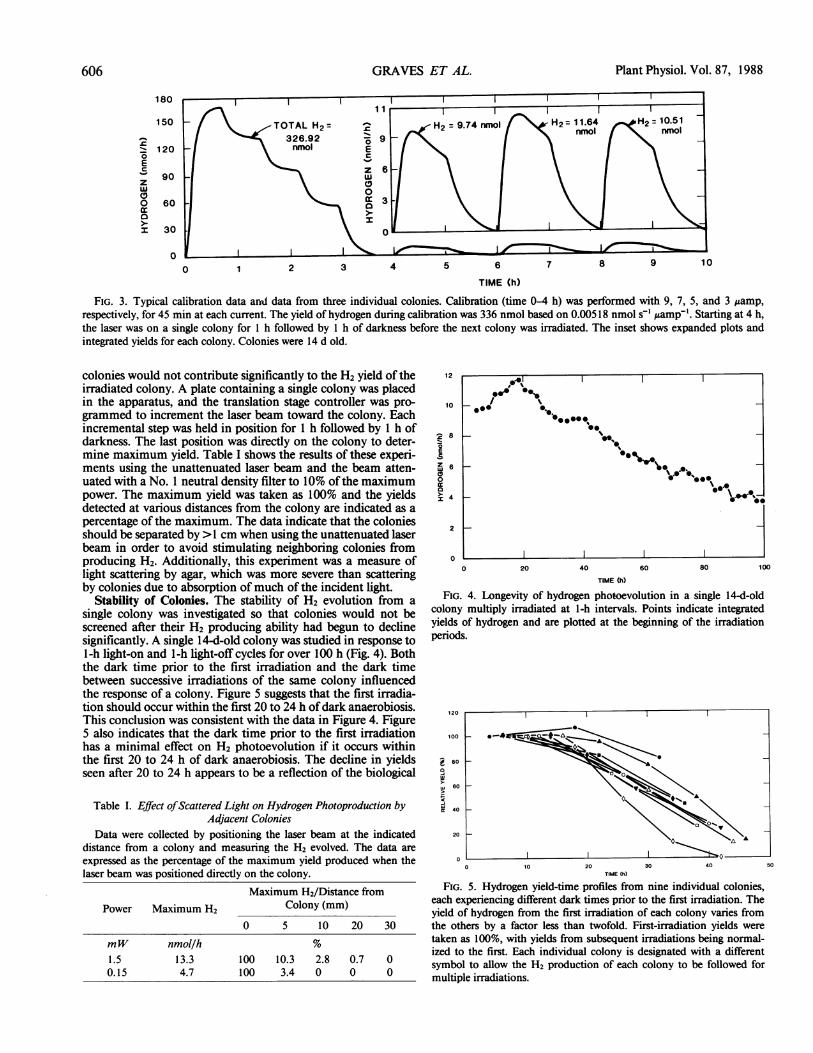
H2 photoevolution from individual algal colonies was easilymeasured using the described apparatus. Photoevolution rates ofless than 1 nmol H2 h-' were measurable with the gas analyzer.Figure 3 shows typical calibration data and the H2 productionby three individual colonies. The integrated experimental yieldofH2 during calibration is in close agreement with the theoreticalyield of 336 nmol calculated using 0.00518 nmol H2 s-' gamp'.Integrated yields from the calibration data were within 5% ofthetheoretical yield, indicating the accuracy ofthis analysis. Anothersignificant feature of the measurement system reflected in thedata shown in Figure 3 is the low noise level. The shape of theH2 evolution profiles accentuates the difference between thesemeasurements and those made with amperometric electrodetechniques. The ability to examine H2 evolution for extendedtime periods gives sustained photohydrogen evolution ratesrather than transient rates determined by short-term experimentsusing amperometric electrodes.
Several factors were examined which could affect real orapparent H2 photoevolution from colonies. The following factorswere considered in analyzing and interpreting the data: the age,Chl content, and size of colonies; light intensity; stability of thecolonies under anaerobiosis; the effect of scattered light onadjacent colonies; and the survivability and cloning of screenedalgal colonies.
Scattered Light. The effect of scattered light on stimulatingH2 evolution from adjacent colonies was examined to determinethe minimum separation between colonies so that neighboring
H2DETECTOR
REACTION CHAMBER
FIG. 2. Schematic diagram ofthe experimental apparatus.
FLOW REGULATOR
_ DIRECTION OFGAS FLOW
LASER BEAM.
ELECTROLYSISCELL
X-Y TRANSLATION STAGE H-P 85
605
GRAVES ET AL.606
180
150.-l
- 1200EcZ 90z
o 60
I 30
Plant Physiol. Vol. 87, 1988
00 1 2 3 4 5 6 7 8 9 10
TIME (h)
FIG. 3. Typical calibration data and data from three individual colonies. Calibration (time 0-4 h) was performed with 9, 7, 5, and 3 vamp,respectively, for 45 min at each current. The yield of hydrogen during calibration was 336 nmol based on 0.00518 nmol s-' ,amp-'. Starting at 4 h,the laser was on a single colony for 1 h followed by 1 h of darkness before the next colony was irradiated. The inset shows expanded plots andintegrated yields for each colony. Colonies were 14 d old.
colonies would not contribute significantly to the H2 yield of theirradiated colony. A plate containing a single colony was placedin the apparatus, and the translation stage controller was pro-grammed to increment the laser beam toward the colony. Eachincremental step was held in position for 1 h followed by 1 h ofdarkness. The last position was directly on the colony to deter-mine maximum yield. Table I shows the results of these experi-ments using the unattenuated laser beam and the beam atten-uated with a No. 1 neutral density filter to 10% ofthe maximumpower. The maximum yield was taken as 100% and the yieldsdetected at various distances from the colony are indicated as apercentage of the maximum. The data indicate that the coloniesshould be separated by >1 cm when using the unattenuated laserbeam in order to avoid stimulating neighboring colonies fromproducing H2. Additionally, this experiment was a measure oflight scattering by agar, which was more severe than scatteringby colonies due to absorption of much of the incident light.
Stability of Colonies. The stability of H2 evolution from asingle colony was investigated so that colonies would not bescreened after their H2 producing ability had begun to declinesignificantly. A single 14-d-old colony was studied in response to1-h light-on and 1-h light-off cycles for over 100 h (Fig. 4). Boththe dark time prior to the first irradiation and the dark timebetween successive irradiations of the same colony influencedthe response of a colony. Figure 5 suggests that the first irradia-tion should occur within the first 20 to 24 h ofdark anaerobiosis.This conclusion was consistent with the data in Figure 4. Figure5 also indicates that the dark time prior to the first irradiationhas a minimal effect on H2 photoevolution if it occurs withinthe first 20 to 24 h of dark anaerobiosis. The decline in yieldsseen after 20 to 24 h appears to be a reflection of the biological
Table I. Effect ofScattered Light on Hydrogen Photoproduction byAdjacent Colonies
Data were collected by positioning the laser beam at the indicateddistance from a colony and measuring the H2 evolved. The data areexpressed as the percentage of the maximum yield produced when thelaser beam was positioned directly on the colony.
Maximum H2/Distance fromPower Maximum H2 Colony (mm)
0 5 10 20 30
mW nmol/h %1.5 13.3 100 10.3 2.8 0.7 00.15 4.7 100 3.4 0 0 0
12
10 /
.020 4 0.60... 81W
'00* %~~028 8%0~~~~~~~~~~~~~@~0~~~~~~~~~~
S
0~~~~~~~~~~~~~~~
0 20 40 60 80 1OC
TIME (h)
FIG. 4. Longevity of hydrogen photoevolution in a single 14-d-oldcolony multiply irradiated at 1-h intervals. Points indicate integratedyields of hydrogen and are plotted at the beginning of the irradiationperiods.
120
8 A -~~~~~~~
80-
40-
20-~Y20~~~~~~~~~~~~~~~~0 0~~~~~~~~0A
0 10 20 30 40 5TIME (h)
FIG. 5. Hydrogen yield-time profiles from nine individual colonies,each experiencing different dark times prior to the first irradiation. Theyield of hydrogen from the first irradiation of each colony varies fromthe others by a factor less than twofold. First-irradiation yields weretaken as 100%, with yields from subsequent irradiations being normal-ized to the first. Each individual colony is designated with a differentsymbol to allow the H2 production of each colony to be followed formultiple irradiations.
0
so
AUTOMATED COLONY SCREENING
stability of the colonies rather than a consequence of the darktime prior to the first irradiation.Colony Age, Chl Content, and Size. The age of a colony,
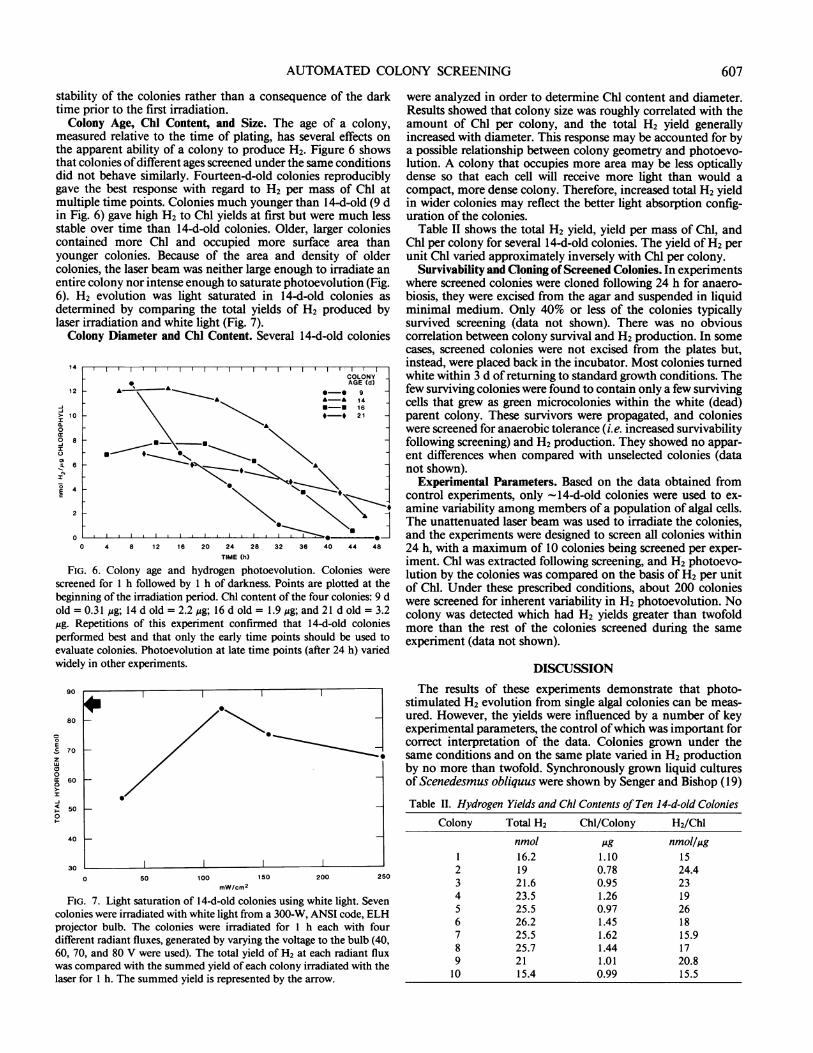
measured relative to the time of plating, has several effects onthe apparent ability of a colony to produce H2. Figure 6 showsthat colonies ofdifferent ages screened under the same conditionsdid not behave similarly. Fourteen-d-old colonies reproduciblygave the best response with regard to H2 per mass of Chl atmultiple time points. Colonies much younger than 14-d-old (9 din Fig. 6) gave high H2 to Chl yields at first but were much lessstable over time than 14-d-old colonies. Older, larger coloniescontained more Chl and occupied more surface area thanyounger colonies. Because of the area and density of oldercolonies, the laser beam was neither large enough to irradiate anentire colony nor intense enough to saturate photoevolution (Fig.6). H2 evolution was light saturated in 14-d-old colonies asdetermined by comparing the total yields of H2 produced bylaser irradiation and white light (Fig. 7).Colony Diameter and Chl Content. Several 144-old colonies
14 l lCOLONYAGE (d)
12 -0~~~~~A-A14
.-1 16
>- 10 21
0
8
E (h)
2
0 4 8 12 16 20 24 28 32 36 40 44 48
TIME (h)
FIG. 6. Colony age and hydrogen photoevolution. Colonies werescreened for 1 h followed by 1 h of darkness. Points are plotted at thebeginning of the irradiation period. Chl content of the four colonies: 9 dold = 0.31 Mg; 14 d old = 2.2 ,g; 16 d old = 1.9 ,ug; and 21 d old = 3.2Mg. Repetitions of this experiment confirmed that 14-d-old coloniesperformed best and that only the early time points should be used toevaluate colonies. Photoevolution at late time points (after 24 h) variedwidely in other experiments.
90
80
'5 70z /CD0
=: 60_ X_60
50 -
0
40-
30
0 50 100 150 200 250mW/cm2
FIG. 7. Light saturation of 14-d-old colonies using white light. Sevencolonies were irradiated with white light from a 300-W, ANSI code, ELHprojector bulb. The colonies were irradiated for 1 h each with fourdifferent radiant fluxes, generated by varying the voltage to the bulb (40,60, 70, and 80 V were used). The total yield of H2 at each radiant fluxwas compared with the summed yield of each colony irradiated with thelaser for 1 h. The summed yield is represented by the arrow.
were analyzed in order to determine Chl content and diameter.Results showed that colony size was roughly correlated with theamount of Chl per colony, and the total H2 yield generallyincreased with diameter. This response may be accounted for bya possible relationship between colony geometry and photoevo-lution. A colony that occupies more area may be less opticallydense so that each cell will receive more light than would acompact, more dense colony. Therefore, increased total H2 yieldin wider colonies may reflect the better light absorption config-uration of the colonies.
Table II shows the total H2 yield, yield per mass of Chl, andChl per colony for several 14-d-old colonies. The yield of H2 perunit Chl varied approximately inversely with Chl per colony.
Survivability and Cloning ofScreened Colonies. In experimentswhere screened colonies were cloned following 24 h for anaero-biosis, they were excised from the agar and suspended in liquidminimal medium. Only 40% or less of the colonies typicallysurvived screening (data not shown). There was no obviouscorrelation between colony survival and H2 production. In somecases, screened colonies were not excised from the plates but,instead, were placed back in the incubator. Most colonies turnedwhite within 3 d of returning to standard growth conditions. Thefew surviving colonies were found to contain only a few survivingcells that grew as green microcolonies within the white (dead)parent colony. These survivors were propagated, and colonieswere screened for anaerobic tolerance (i.e. increased survivabilityfollowing screening) and H2 production. They showed no appar-ent differences when compared with unselected colonies (datanot shown).
Experimental Parameters. Based on the data obtained fromcontrol experiments, only -14-d-old colonies were used to ex-amine variability among members of a population of algal cells.The unattenuated laser beam was used to irradiate the colonies,and the experiments were designed to screen all colonies within24 h, with a maximum of 10 colonies being screened per exper-iment. Chl was extracted following screening, and H2 photoevo-lution by the colonies was compared on the basis of H2 per unitof Chl. Under these prescribed conditions, about 200 colonieswere screened for inherent variability in H2 photoevolution. Nocolony was detected which had H2 yields greater than twofoldmore than the rest of the colonies screened during the sameexperiment (data not shown).
DISCUSSIONThe results of these experiments demonstrate that photo-
stimulated H2 evolution from single algal colonies can be meas-ured. However, the yields were influenced by a number of keyexperimental parameters, the control ofwhich was important forcorrect interpretation of the data. Colonies grown under thesame conditions and on the same plate varied in H2 productionby no more than twofold. Synchronously grown liquid culturesofScenedesmus obliquus were shown by Senger and Bishop (19)
Table II. Hydrogen Yields and Chl Contents of Ten 14-d-old ColoniesColony Total H2 Chl/Colony H2/Chl
nmol agg nmoltlg1 16.2 1.10 152 19 0.78 24.43 21.6 0.95 234 23.5 1.26 195 25.5 0.97 266 26.2 1.45 187 25.5 1.62 15.98 25.7 1.44 179 21 1.01 20.810 15.4 0.99 15.5
607

Plant Physiol. Vol. 87, 1988
to vary in H2 photoproduction during their life cycle. C. rein-hardtii growing as colonies did not display any life-cycle-depend-ent variation in H2 production when synchronous and asyn-chronous (grown in constant light) colonies were compared.Therefore, the value of twofold was taken as the normal rangeof variability for C. reinhardtii under our experimental condi-tions.The development of an apparatus with the sensitivity for
detecting photogenerated H2 from a single colony has a numberof pertinent applications. At a rate of 10 colonies per day, theapparatus can be used to screen natural populations of algae forH2 photoevolution. This approach for making putative identifi-cation of interesting new strains is much more efficient thanisolating pure cultures and screening them individually. Simi-larly, the apparatus can be used as a secondary screen to testputative mutants altered in their H2 producing ability and hy-drogenase. The simple addition of a wild-type colony to a plateof putative mutants will provide a powerful internal control forevaluating mutants. Since the general rule-of-thumb for algalmutants is that the phenotype should deviate from wildtype by5-fold to 10-fold (14), the inherent variability of 2-fold indicatedby this work will not interfere with mutant screening. Theapparatus can also be used for comparing the overall response ofalgal colonies growing on semisolid medium to liquid cultures.Low postscreening viability mandates a colony replicationscheme so that interesting colonies can be recovered.An important addition to the apparatus would be an 02 sensor
of sufficient sensitivity to detect photoevolved 02 (<10 nmol/h).A Hersch galvanic cell (1, 1 1) was installed in the system, butthe signal to noise ratio was too great to allow detection of 02evolved by a single 14-d-old colony. An 02 luminometer (5) wasanother option, but its use required that the helium flow fromthe reaction chamber be split into two streams, one for the H2analyzer and one for the luminometer. The gas analyzer issensitive to dimethyl sulfoxide, the solvent used in the lumi-nometer, and the luminometer cannot be placed downstreamfrom the analyzer because ofthe 02 flow that purges the analyzer.Splitting an already small signal was considered impractical, sothe luminometer was not tested.Even with the current inability to detect 02, the apparatus
described here is a powerful and quantitative tool which shouldfind pertinent applications in a number of areas involving an-aerobic photosynthesis and H2 photoevolution. It exceeds otherdetection methods in sensitivity and long-term stability and isuniquely suited for measuring steady state H2 photoevolution forextended periods of time. Combining point irradiation, using alaser with the H2 sensor, has produced a novel method forefficiently examining H2 photoevolution from individual clonesof an algal population.
Acknowledgments-The authors thank L. W. Jones and P. J. Hanson for their
comments and critical reading of the manuscript. They also thank M. A. Neal forsecretarial support.
LITERATURE CITED
1. BAKER WJ, JF COMBS, TL ZINN, AW WOTRING, RF WALL 1959 The glavaniccell oxygen analyzer. Ind Eng Chem 51: 727-730
2. BAMBERGER ES, D KING, DL ERBES, M GIBBS 1982 H2 and CO2 evolution byanaerobically adapted Chlamydomonas reinhardtii F-60. Plant Physiol 69:1268-1273
3. BISHOP NI, M FRICK, LW JONES 1977 Photohydrogen production in greenalgae: water serves as the primary substrate for hydrogen and oxygen pro-duction. In A Mitsui, S Miyachi, A San Pietro, S Tamura, eds, BiologicalSolar Energy Conversion. Academic Press, New York, pp 3-22
4. BRAND JJ 1983 Survey for Hydrogenase Activity in Algae. Final Report forSubcontract No. IB-1-9229-1 under Prime Contract No. EG-77-C-01-4042,Solar Energy Research Institute, Golden, CO
5. BURR A, D MAUZERALL 1968 The oxygen luminometer, an apparatus todetermine small amounts of oxygen, and application to photosynthesis.Biochem Biophys Acta 153: 614-624
6. BURRIS RH 1977 Energetics of Biological N2 Fixation. In Biological SolarEnergy Conversion. A Mitsui, S Miyachi, A San Pietro, S Tamura, eds,Academic Press, New York, pp 275-289
7. EVANS DA, WR SHARP, HP MEDINo-FILHO 1984 Somaclonal and gameto-clonal variation. Am J Bot 71: 759-744
8. GAFFRON H, J RUBIN 1942 Fermentative and photochemical production ofhydrogen in algae. J Gen Physiol 26: 219-240
9. GREENBAUM E 1981 Stoichiometry of hydrogen and oxygen photoproductionby chloroplasts, ferrodoxin, and hydrogenase. In M Moo-Young, CW Ro-binson, eds, Advances in Biotechnology, Vol II. Pergamon Press, Oxford, pp297-302
10. GREENBAUM E 1984 Biophotolysis of water: the light saturation curves. Pho-tobiochem Photobiophys 8: 323-332
11. HERSCH PA 1973 Couloximetry. Am Lab 5: 29-3912. JONES LW, NI BISHOP 1976 Simultaneous measurement of oxygen and hydro-
gen exchange from the blue-green alga anabaena. Plant Physiol 57: 659-66513. LAYZELL DB, GE WEAGLE, DT CAVIN 1984 A highly sensitive, flow through
H2 gas analyzer for use in nitrogen fixation studies. Plant Physiol 75: 582-585
14. LEVINE RP 1970 Preparation and properties of mutant strains of Chiamydo-monas reinhardtii. Methods Enzymol 23: 119-129
15. MACKINNEY G 1941 Absorption of light by chlorophyll solutions. J Biol Chem140: 315-322
16. MALLARD TM, CS MALLARD, HS HOLFIELD, TA LARUE 1977 Portable, gaschromatography for the acetylene reduction assay of nitrogenase. Anal Chem49:1275-1277
17. McBRIDE AC, S LEIN, RK TOGASAKI, A SAN PIETRO 1977 Mutational analysisof Chlamydomonas reinhardtii: application to biological solar energy con-version. In A Mitsui, S Miyachi, A San Pietro, S Tamura, eds, BiologicalSolar Energy Conversion. Academic Press, New York, pp 77-86
18. REEVES M, E GREENBAUM 1985 Long-term endurance and selection studies inhydrogen and oxygen photoproduction by Chiamydomonas reinhardtii.Enzyme Microb Technol 7: 169-174
19. SENGER H, NI BISHOP 1979 Observations on the photohydrogen producingactivity during the synchronous cell cycle of Scenedesmus obliquus. Planta145: 53-62
20. WANG RT 1980 Amperometric hydrogen electrode. Methods Enzymol 69:409-413
21. WANG R, FP HEALEY, J MYERS 1971 Amperometric measurement ofhydrogenevolution in Chlamydomonas. Plant Physiol 48: 108-110
22. WARD B, ME REEVES, E GREENBAUM 1985 Stress-selected Chlamydomonasreinhardtii for photoproduction of hydrogen. Biotechnol Bioeng Symp 15:501-507
23. WEAVER PF, S LIEN, M SEIBERT 1980 Photobiological production of hydrogen.Sol Energy 24: 3-45
608 GRAVES ET AL.