racjonalne stosowanie antybiotyków w weterynarii - researchgate
TRANSCRIPT
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/258341454
Oporność Salmonella i komensalnych Escherichia coli - skutek
stosowania antybiotyków czy epidemiologia zakażeń ?
Chapter · October 2013
CITATIONS
0READS
1,233
3 authors, including:
Some of the authors of this publication are also working on these related projects:
In simulacra studies on the reduction of azo-dyes and identification of their carcinogenic metabolites View project
European Network for Optimization of Veterinary Antimicrobial Treatment - CA18217 View project
Dariusz Wasyl
Państwowy Instytut Weterynaryjny
116 PUBLICATIONS 1,476 CITATIONS
SEE PROFILE
Magdalena Zając
Państwowy Instytut Weterynaryjny
47 PUBLICATIONS 622 CITATIONS
SEE PROFILE
All content following this page was uploaded by Dariusz Wasyl on 30 May 2014.
The user has requested enhancement of the downloaded file.

Racjonalne stosowanie antybiotyków
w weterynarii
monografia
pod redakcją naukową
dr. hab. Krzysztofa Niemczuka
dr Doroty Krasuckiej
Puławy, 2013

RADA WYDAWNICTW:
dr hab. Artur Rzeżutka - przewodniczący
prof. dr hab. Dariusz Bednarek
dr hab. Tomasz Cencek, prof. nadzw.
prof. dr hab. Krzysztof Kwiatek
prof. dr hab. Jacek Osek
prof. dr hab. Zygmunt Pejsak
dr hab. Mirosław Paweł Polak, prof. nadzw.
prof. dr hab. Michał Reichert
prof. dr hab. Jerzy Rola
prof. dr hab. Jan Franciszek Żmudziński
prof. dr hab. Jan Żmudzki
RECENZENCI:
prof. dr hab. Jacek Osek
prof. dr hab. Jan Żmudzki
REDAKTOR WYDANIA:
mgr Anetta Szymańska
SKŁAD I OPRACOWANIE GRAFICZNE:
dr hab. Krzysztof Niemczuk, prof. nadzw., dr Dorota Krasucka
©
Copyright by Państwowy Instytut Weterynaryjny
-Państwowy Instytut Badawczy
ISBN 978-83-89946-61-4
WYDAWCA:
Państwowy Instytut Weterynaryjny
-Państwowy Instytut Badawczy (PIWet-PIB)
DRUK I OPRAWA:
Zakład Planowania i Upowszechniania Badań PIWet-PIB
al. Partyzantów 57, 24-100 Puławy
Nakład: 225+25 egz.; Ark. wyd.: 8,89

Spis treści
Nadzór Inspekcji Weterynaryjnej nad produktami leczniczymi
weterynaryjnymi. Aktualne działania i problemy.
Janusz Związek, Ewa Maślikowska 5
Analiza stosowania leków przeciwdrobnoustrojowych
u trzody chlewnej, bydła i drobiu w Polsce w latach 2010 -2012.
Dorota Krasucka, Wojciech Cybulski, Agnieszka Klimowicz 25
Wybrane aspekty stosowania pasz leczniczych.
Monika Przeniosło-Siwczyńska, Krzysztof Kwiatek 41
Przeszłość, teraźniejszość i przyszłość antybiotyków stosowanych
w hodowli zwierząt w Unii Europejskiej (UE), w wybranych
państwach europejskich i na świecie – z punktu widzenia sektora
zdrowia zwierząt.
Peter Oostenbach 55
Zasady skutecznej i bezpiecznej antybiotykoterapii chorób
bakteryjnych drobiu.
Andrzej Koncicki 95
Racjonalne stosowanie antybiotyków u świń.
Zygmunt Pejsak, Marian Truszczyński 113
Przyczyny i skutki ograniczania stosowania antybiotyków u
zwierząt oraz alternatywne sposoby ich zastąpienia.
Marian Truszczyński, Zygmunt Pejsak 127

Probiotyki jako alternatywa antybiotykowych stymulatorów
wzrostu (ASW) w żywieniu zwierząt gospodarskich.
Jens Noesgaard Jørgensen, Bea Nielsen 147
Krajowa immunoprofilaktyka jako alternatywa dla
antybiotykoterapii chorób drobiu.
Dorota Krasucka, Katarzyna Kos, Agnieszka Klimowicz,
Ewa Łysiak, Aneta Woźniak., Małgorzata Lisiowska,
Grzegorz Tomczyk, Wojciech Cybulski 157
Zdrowie konsumenta wobec standardów antybiotykoterapii
weterynaryjnej.
Cezary Jacek Kowalski, Beata Łebkowska-Wieruszewska 171
Pozostałości antybiotyków w tkankach i produktach zwierzęcego
pochodzenia - realność zagrożeń.
Andrzej Posyniak, Jan Żmudzki, Anna Gajda, Małgorzata Gbylik,
Tomasz Błądek, Tomasz Śniegocki, Kamila Mitrowska,
Bogumił Biernacki, Marta Piątkowska, Maja Antczak, Jacek Osek,
Hanna Różańska 185
Oporność Salmonella i komensalnych Escherichia coli – skutek
stosowania antybiotyków czy epidemiologia zakażeń?
Dariusz Wasyl, Andrzej Hoszowski, Magdalena Zając 199
Metoda krążkowo – dyfuzyjna w weterynaryjnej diagnostyce
bakteriologiczne.
Dominika Borowska, Artur Jabłoński, Sylwia Zębek, Agnieszka
Nowak, Anna Grzesiak 237

199
Oporność Salmonella i komensalnych Escherichia coli
-skutek stosowania antybiotyków czy epidemiologia zakażeń?
Dariusz Wasyl, Andrzej Hoszowski, Magdalena Zając
Państwowy Instytut Weterynaryjny – Państwowy Instytut Badawczy,
w Puławach
Zjawisko oporności na substancje antybakteryjne od zawsze
intrygowało naukowców. Pytania, jak mikroorganizmy radzą sobie
z substancjami chemicznymi, w jaki sposób tworzą strategie umożliwiające
przetrwanie w środowisku zawierającym substancje antybiotyczne, jak są one
przekazywane pomiędzy komórkami i gatunkami mikroorganizmów oraz ich
nosicielami, wydają się obecnie bardziej złożone niż można było sądzić
w początkach ery antybiotykoterapii. Pomimo, że świat nauki dysponuje
zaawansowanymi metodami badawczymi, ciągle mamy do czynienia
z lukami w wiedzy na temat oporności i nie rozumiemy w pełni tego
zjawiska. Podstawowe ograniczenia wynikają ze złożoności mikrobiomu
występującego w różnych ekosystemach (Allen i wsp. 2013; Perchec-Merien
i Lewis, 2013) oraz faktu, że podejmowane badania mają charakter
fragmentaryczny – koncentrują się zwykle na wybranym zagadnieniu np.
danym rodzaju bakterii izolowanym z określonego źródła w określonym
czasie (Dahmen i wsp. 2012; Wasyl i wsp. 2012b). Również globalizacja
i związane z nią podróże człowieka oraz intensywna wymiana handlowa
utrudniają zrozumienie epidemiologii oporności na substancje antybakteryjne
(Davies i Davies, 2010; SVARM, 2012). Należy również wspomnieć,
że współczesne metody bakteriologiczne stwarzają możliwość wyhodowania
tylko części bakterii obecnych w mikrobiomie – oporność tych, których nie

200
da się uzyskać w warunkach in vitro stanowią obiekt analizy metagenomowej
(Simon i Daniel, 2011; Bhullar i wsp. 2012; Czekalski i wsp. 2012).
Oporne bakterie są źródłem poważnych zagrożeń zdrowotnych dla
ludzi i zwierząt (Morfin-Otero i wsp., 2012) oraz strat ekonomicznych
(Tadesse i wsp. 2012). Niebezpieczna dla człowieka oporność może być
generowana zarówno w obszarze zdrowia publicznego, jak i w łańcuchu
żywnościowym, skąd oporne bakterie mogą przedostawać się do człowieka
poprzez bezpośredni kontakt zwierząt i ludzi, lub pośrednio, wraz
z żywnością (EFSA, 2009; Davies i Davies, 2010).
Monitorowanie oporności patogenów bakteryjnych, takich jak np.
Salmonella, ma na celu odpowiednio wczesne reagowanie i przeciwdziałanie
zakażeniom groźnym dla zdrowia i życia (Morfin-Otero i wsp. 2012).
Wdrożenie programów zwalczania Salmonella u drobiu, wraz z rosnącą
świadomością społeczną, w znacznym stopniu ograniczyły zagrożenia
epidemiologiczne dla ludzi i zwierząt. Tym samym zmniejszyła się liczba
izolatów bakterii patogennych dostępnych do badań o charakterze
epidemiologicznym. Bakterie komensalne, takie jak Escherichia (E.) coli,
są w tej sytuacji istotną alternatywą (Tadesse i wsp. 2012; Allen i wsp. 2013).
Chociaż nie można wykluczyć wystąpienia fenotypów patogennych (Pitout,
2012), większość izolatów E. coli wchodzących w skład flory jelitowej
człowieka lub zwierząt może być uznawana za bakterie wskaźnikowe
(komensale) pozbawione cech chorobotwórczości, które można wykorzystać
w monitorowaniu oporności na substancje antybakteryjne (Kaesbohrer i wsp.
2012; Tadesse i wsp. 2012). Zaletą badania oporności komensali jest ich
powszechne występowanie i stosunkowo proste i wydajne metody ich izolacji
(Allen i wsp. 2013). Dodatkowo, w odróżnieniu od bakterii patogennych,
właściwie wyklucza się wpływ klonalnego szerzenia się zakażeń na wyniki
monitorowania oporności. Co więcej, możliwe wydaje się określenie

201
prawdopodobnych związków oporności bakterii komensalnych izolowanych
od zwierząt ze stosowaniem u nich przeciwbakteryjnych substancji czynnych.
Metody wykrywania oporności powinny być dostosowane do
przyjętego celu badania. Metody molekularne, charakteryzujące się
doskonałą czułością wykrywania oporności i identyfikacji odpowiedzialnych
za nią mechanizmów, są ciągle zbyt kosztowne i pracochłonne w rutynowych
badaniach o charakterze klinicznym (van der Bij i wsp. 2012). Samo też
wykrycie u danego izolatu bakteryjnego genu oporności nie musi oznaczać
jego ekspresji i być zgodne ze stwierdzaną fenotypowo opornością (Ozaki
i wsp. 2011; Wasyl i wsp. 2012b). Dlatego też, do monitorowania oporności
zalecane są klasyczne, wystandaryzowane metody bakteriologiczne (de Jong
i wsp. 2012; EFSA, 2012b; EFSA i ECDC, 2012; Tadesse i wsp. 2012).
Niezależnie od stosowanej metody oznaczania oporności kluczowe znaczenie
mają kryteria wykorzystywane przy interpretacji wyników. Kryteria kliniczne
tzw. „clinical breakpoints” są stosowane, aby ocenić prawdopodobieństwo
skutecznej antybiotykoterapii w konkretnym przypadku klinicznym (Morfin-
Otero i wsp. 2012; van der Bij i wsp. 2012). Podejście epidemiologiczne,
w którym wykorzystuje się tzw. „epidemiological cut-off values” wydaje się
bardziej właściwe w badaniach monitoringowych zarówno patogenów, jak
i bakterii komensalnych (Bronzwaer i wsp. 2008), gdyż daje możliwość nie
tylko wczesnego wykrycia zmian w oporności populacji bakterii,
ale i pojawienia się nieznanych dotąd mechanizmów oporności (EFSA,
2008). Ponadto, poprzez analizę ilościową wyników poprawia się możliwość
oceny tendencji dotyczących występowania oporności i jej zmian w czasie
(EFSA, 2012a; van der Bij i wsp. 2012).
Ranga naukowa problemu narastania oporności wśród bakterii, jej
związki ze stosowaniem antybiotyków oraz zagrożenia dla zdrowia
doprowadziły do intensyfikacji badań nad klinicznymi aspektami oporności

202
oraz monitorowania tego zjawiska zarówno u bakterii patogennych,
jak i komensali (Tadesse i wsp. 2012; Allen i wsp. 2013; Schroeter i wsp.
2013). W Unii Europejskiej podjęto próbę harmonizacji metod
monitorowania w celu większej porównywalności wyników uzyskiwanych
w różnych krajach (EFSA, 2012b; van der Bij i wsp. 2012). W związku
z tym, prowadzony obecnie monitoring obejmuje zarówno czynniki
zoonotyczne bezpośrednio zagrażające zdrowiu ludzi i zwierząt (Bronzwaer
i wsp. 2008), jak i bakterie komensalne, będące potencjalnym rezerwuarem
genów oporności i indykatorem stosowania substancji antybakteryjnych
u zwierząt (EFSA, 2008; EFSA i ECDC, 2012). Rekomendacje UE określają
szczegółowy schemat badań, metody i kryteria interpretacji wyników oraz
listę substancji antybakteryjnych, w stosunku do których powinien być
prowadzony monitoring (EFSA, 2008). W oparciu o te zalecenia w 2008 r.
w Zakładzie Mikrobiologii PIWet-PIB rozpoczęto monitorowanie oporności
Salmonella izolowanych od zwierząt, z pasz i żywności, a od 2009 r.
badaniami objęto również komensalne E. coli izolowane od zwierząt
rzeźnych na terenie całego kraju. Badania te są realizowane w ramach
działalności laboratorium referencyjnego (izolaty Salmonella uzyskane
w trakcie realizacji krajowych programów zwalczania Salmonella w stadach
niosek towarowych, brojlerów i indyków) oraz Programu Wieloletniego
(pozostałe Salmonella i E. coli).
Celem niniejszego opracowania jest przedstawienie wybranych
elementów prowadzonych badań, ukazujących zwłaszcza zmiany oporności
w czasie (EFSA, 2012a) oraz wniosków z prowadzonych badań
uzupełnionych analizami epidemiologicznymi.

203
Materiały i metody
Izolaty Salmonella: Monitoring oporności na substancje
antybakteryjne objął 3 795 szczepów Salmonella spośród ponad 11 500
izolatów zgromadzonych w Zakładzie Mikrobiologii PIWet-PIB w latach
2008 – 2012. Począwszy od 346 izolatów w 2008 r., 429 w 2009 r. ich liczba
wzrosła do około 1 000 Salmonella w kolejnych latach badań. Izolaty te
zostały uzyskane w ramach krajowych programów zwalczania Salmonella
w stadach hodowlanych kur (N=143), niosek towarowych (N=703),
brojlerów (N=769) i indyków (N=131) oraz innych badań różnych gatunków
zwierząt, w tym gadów (N=538), z żywności (N=514) i pasz (N=143).
Należały one do 241 serowarów, z których większość była reprezentowana
przez pojedyncze izolaty. Jednak siedem najczęściej notowanych serowarów
objęło aż 71% badanych izolatów: S. Enteritidis (N=1360), S. Infantis
(N=374), S. Typhimurium (N=330; w tym wariant jednofazowy),
S. Mbandaka (N=213), S. Newport (N=173), S. Virchow (N=133),
i S. Kentucky (N=111).
Izolaty E. coli: Monitoring oporności komensalnych E. coli objął
5 grup produkcyjnych zwierząt: brojlery, nioski towarowe pochodzące ze
stad likwidowanych po okresie nieśności, indyki, świnie i bydło. Obiektywną
ocenę poziomu i tendencji występujących w oporności na substancje
antybakteryjne osiągnięto poprzez losowe próbkobranie prowadzone przez
Inspekcję Weterynaryjną. Charakterystykę opisową populacji zwierząt
rzeźnych objętych pobieraniem próbek przedstawiono w tabeli 1. W styczniu
każdego roku zbierane były dane dotyczące liczby zwierząt poddanych
ubojowi w każdej rzeźni działającej na terenie kraju. Na podstawie tych
danych opracowywany był program pobierania próbek w ciągu kolejnych 12
miesięcy (luty – styczeń), który określał liczbę pobrań w wyznaczonych
zakładach. Liczba ta była proporcjonalna do przewidywanej w danym roku
204
wielkości uboju określonej grupy produkcyjnej zwierząt rzeźnych. Dla każdej
z 5 grup zwierząt objętych badaniami przewidziano 200 pobrań w ciągu roku.
Każda próbka obejmowała 3 wymazy z kloaki lub odbytu pobrane od
3 losowo wybranych zwierząt bezpośrednio po uboju. Próbki były pobierane
przez inspektorów weterynaryjnych przy pomocy jałowych wymazówek
z pożywką transportową i przesyłane bezpośrednio do laboratorium
PIWet-PIB. Dane opisujące próbkę, obejmujące datę, miejsce i źródło były
przesyłane elektronicznie do centralnej bazy danych zaprojektowanej
i zbudowanej wyłącznie na potrzeby realizacji tego zadania badawczego.
Badania laboratoryjne polegały na bezpośrednim posiewie próbki na agar
MacConkeya. Uzyskane kolonie wykazujące typową dla E. coli morfologię
identyfikowano biochemiczne, a następnie jeden izolat reprezentujący próbkę
był poddawany badaniu oporności na substancje antybakteryjne.
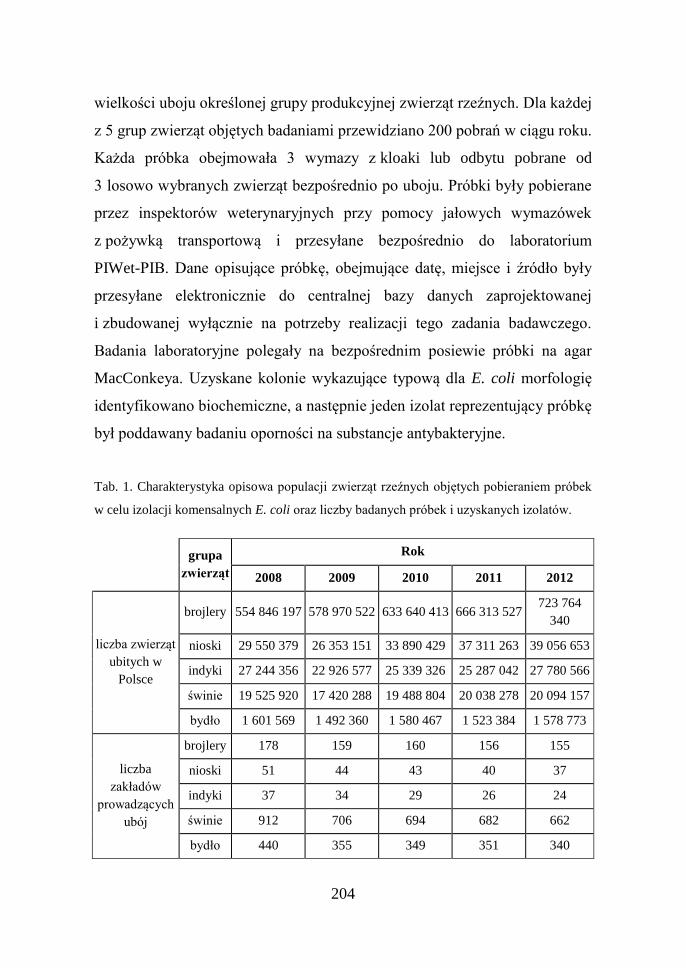
Tab. 1. Charakterystyka opisowa populacji zwierząt rzeźnych objętych pobieraniem próbek
w celu izolacji komensalnych E. coli oraz liczby badanych próbek i uzyskanych izolatów.
grupa
zwierząt
Rok
2008 2009 2010 2011 2012
liczba zwierząt
ubitych w
Polsce
brojlery 554 846 197 578 970 522 633 640 413 666 313 527 723 764
340
nioski 29 550 379 26 353 151 33 890 429 37 311 263 39 056 653
indyki 27 244 356 22 926 577 25 339 326 25 287 042 27 780 566
świnie 19 525 920 17 420 288 19 488 804 20 038 278 20 094 157
bydło 1 601 569 1 492 360 1 580 467 1 523 384 1 578 773
liczba
zakładów
prowadzących
ubój
brojlery 178 159 160 156 155
nioski 51 44 43 40 37
indyki 37 34 29 26 24
świnie 912 706 694 682 662
bydło 440 355 349 351 340
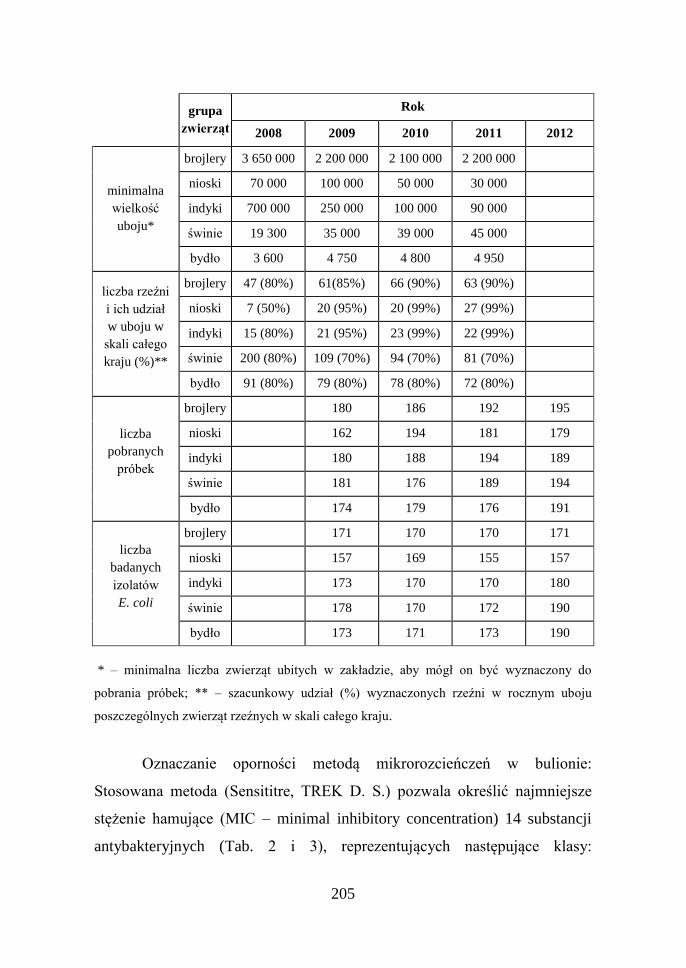
205
grupa
zwierząt
Rok
2008 2009 2010 2011 2012
minimalna
wielkość
uboju*
brojlery 3 650 000 2 200 000 2 100 000 2 200 000
nioski 70 000 100 000 50 000 30 000
indyki 700 000 250 000 100 000 90 000
świnie 19 300 35 000 39 000 45 000
bydło 3 600 4 750 4 800 4 950
liczba rzeźni
i ich udział
w uboju w
skali całego
kraju (%)**
brojlery 47 (80%) 61(85%) 66 (90%) 63 (90%)
nioski 7 (50%) 20 (95%) 20 (99%) 27 (99%)
indyki 15 (80%) 21 (95%) 23 (99%) 22 (99%)
świnie 200 (80%) 109 (70%) 94 (70%) 81 (70%)
bydło 91 (80%) 79 (80%) 78 (80%) 72 (80%)
liczba
pobranych
próbek
brojlery 180 186 192 195
nioski 162 194 181 179
indyki 180 188 194 189
świnie 181 176 189 194
bydło 174 179 176 191
liczba
badanych
izolatów
E. coli
brojlery 171 170 170 171
nioski 157 169 155 157
indyki 173 170 170 180
świnie 178 170 172 190
bydło 173 171 173 190
* – minimalna liczba zwierząt ubitych w zakładzie, aby mógł on być wyznaczony do
pobrania próbek; ** – szacunkowy udział (%) wyznaczonych rzeźni w rocznym uboju
poszczególnych zwierząt rzeźnych w skali całego kraju.
Oznaczanie oporności metodą mikrorozcieńczeń w bulionie:
Stosowana metoda (Sensititre, TREK D. S.) pozwala określić najmniejsze
stężenie hamujące (MIC – minimal inhibitory concentration) 14 substancji
antybakteryjnych (Tab. 2 i 3), reprezentujących następujące klasy:

206
betalaktamy, cefalosporyny, chinolony i fluorochinolony, fenikole,
aminoglikozydy, inhibitory przemian kwasu foliowego, tetracykliny
i polimyksyny. Uzyskane wartości MIC interpretowano zgodnie z kryteriami
epidemiologicznymi (EUCAST 2010, www.eucast.org). Wartość MIC
powyżej epidemiologicznej wartości odcięcia (ang. cut-off value) pozwala
zaliczyć izolat do kategorii NWT (ang. Non-Wild Type), która oznacza
obecność mechanizmu warunkującego oporność danego izolatu na badaną
substancję czynną. Kategoria NWT bywa też nazywana opornością
mikrobiologiczną (ang. microbiological resistance) dla odróżnienia od
oporności klinicznej (ang. clinical resistance) (EUCAST, 2000).
Analiza statystyczna: Częstość występowania oporności
mikrobiologicznej była wyrażana jako odsetek izolatów wykazujących
wartość MIC powyżej przyjętego kryterium interpretacji z zachowaniem 95%
przedziału ufności (95% CI). Trendy oporności w czasie w oparciu o wyniki
jakościowe (%WT i %NWT) analizowano w modelu regresji logistycznej,
a liniowa analiza regresji była wykorzystana do analizy przesunięć wartości
MIC (dane logarytmowane) (EFSA, 2012b). Wartość współczynnika
prawdopodobieństwa p 0,5 oznaczała istotną statystycznie zmianę, której
kierunek obrazowano na wykresach.
Metody molekularne: Wybrane mechanizmy oporności na chinolony
i cefalosporyny identyfikowano poprzez amplifikację i sekwencjonowanie
fragmentów genów (Veldman i wsp. 2011; Wasyl i wsp. 2012a; Wasyl
i wsp. 2012b). Do analizy pokrewieństwa izolatów wykorzystywano metodę
PFGE zgodną z protokołem PulseNet (Ribot i wsp. 2006).
Wyniki
Salmonella
Wśród badanych izolatów Salmonella stwierdzono występowanie
oporności mikrobiologicznej na każdą z badanych substancji antybak-
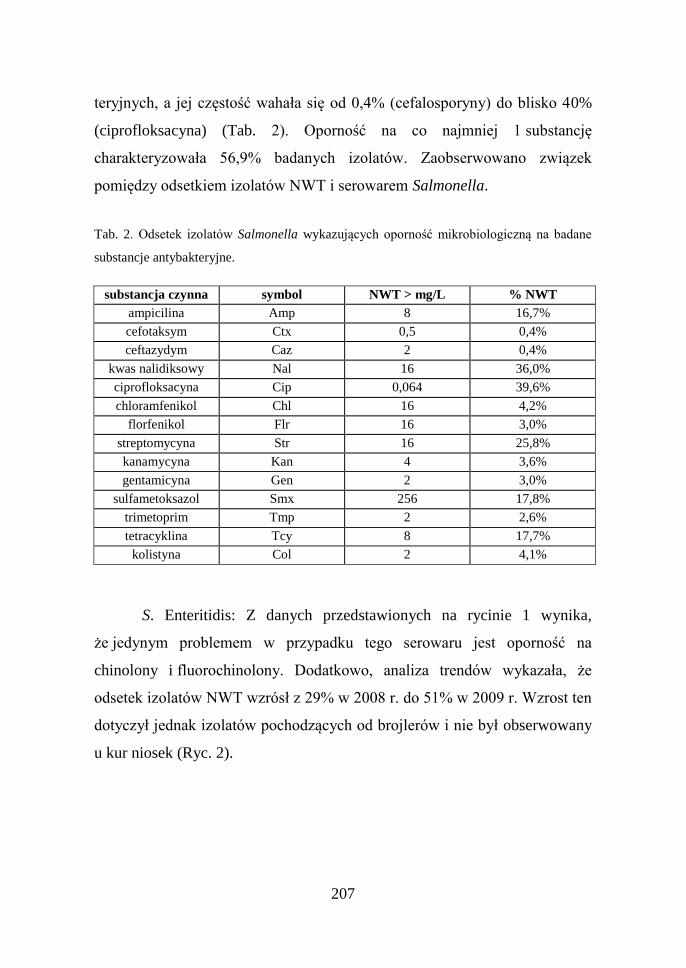
207
teryjnych, a jej częstość wahała się od 0,4% (cefalosporyny) do blisko 40%
(ciprofloksacyna) (Tab. 2). Oporność na co najmniej 1 substancję
charakteryzowała 56,9% badanych izolatów. Zaobserwowano związek
pomiędzy odsetkiem izolatów NWT i serowarem Salmonella.
Tab. 2. Odsetek izolatów Salmonella wykazujących oporność mikrobiologiczną na badane
substancje antybakteryjne.
substancja czynna symbol NWT > mg/L % NWT
ampicilina Amp 8 16,7%
cefotaksym Ctx 0,5 0,4%
ceftazydym Caz 2 0,4%
kwas nalidiksowy Nal 16 36,0%
ciprofloksacyna Cip 0,064 39,6%
chloramfenikol Chl 16 4,2%
florfenikol Flr 16 3,0%
streptomycyna Str 16 25,8%
kanamycyna Kan 4 3,6%
gentamicyna Gen 2 3,0%
sulfametoksazol Smx 256 17,8%
trimetoprim Tmp 2 2,6%
tetracyklina Tcy 8 17,7%
kolistyna Col 2 4,1%
S. Enteritidis: Z danych przedstawionych na rycinie 1 wynika,
że jedynym problemem w przypadku tego serowaru jest oporność na
chinolony i fluorochinolony. Dodatkowo, analiza trendów wykazała, że
odsetek izolatów NWT wzrósł z 29% w 2008 r. do 51% w 2009 r. Wzrost ten
dotyczył jednak izolatów pochodzących od brojlerów i nie był obserwowany
u kur niosek (Ryc. 2).
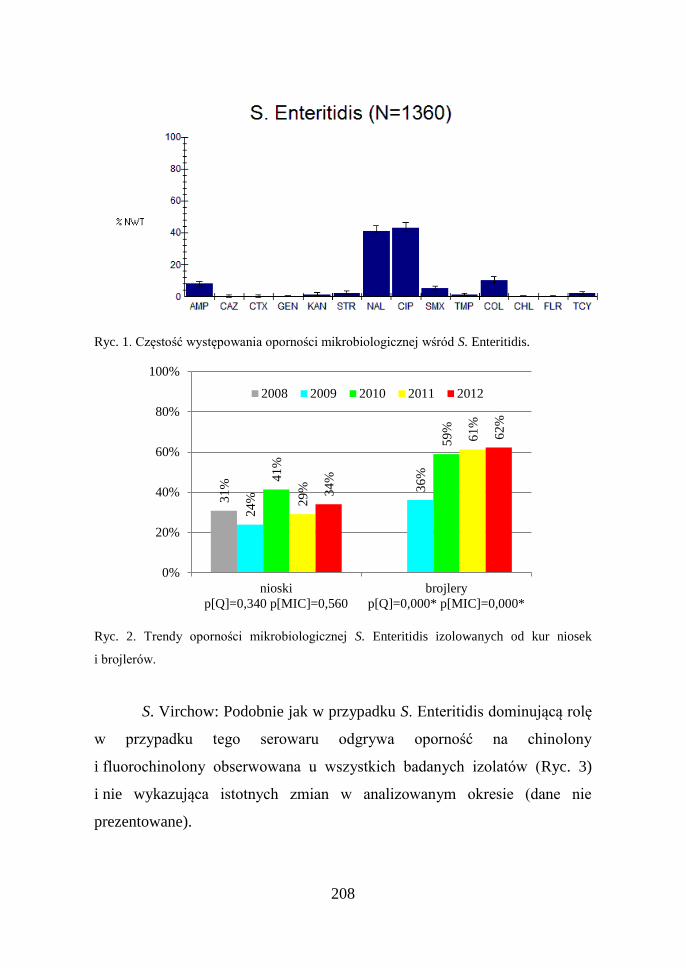
208
Ryc. 1. Częstość występowania oporności mikrobiologicznej wśród S. Enteritidis.
Ryc. 2. Trendy oporności mikrobiologicznej S. Enteritidis izolowanych od kur niosek
i brojlerów.
S. Virchow: Podobnie jak w przypadku S. Enteritidis dominującą rolę
w przypadku tego serowaru odgrywa oporność na chinolony
i fluorochinolony obserwowana u wszystkich badanych izolatów (Ryc. 3)
i nie wykazująca istotnych zmian w analizowanym okresie (dane nie
prezentowane).
31
%
24
% 3
6%
41
%
59
%
29
%
61
%
34
%
62
%
0%
20%
40%
60%
80%
100%
nioski
p[Q]=0,340 p[MIC]=0,560
brojlery
p[Q]=0,000* p[MIC]=0,000*
2008 2009 2010 2011 2012
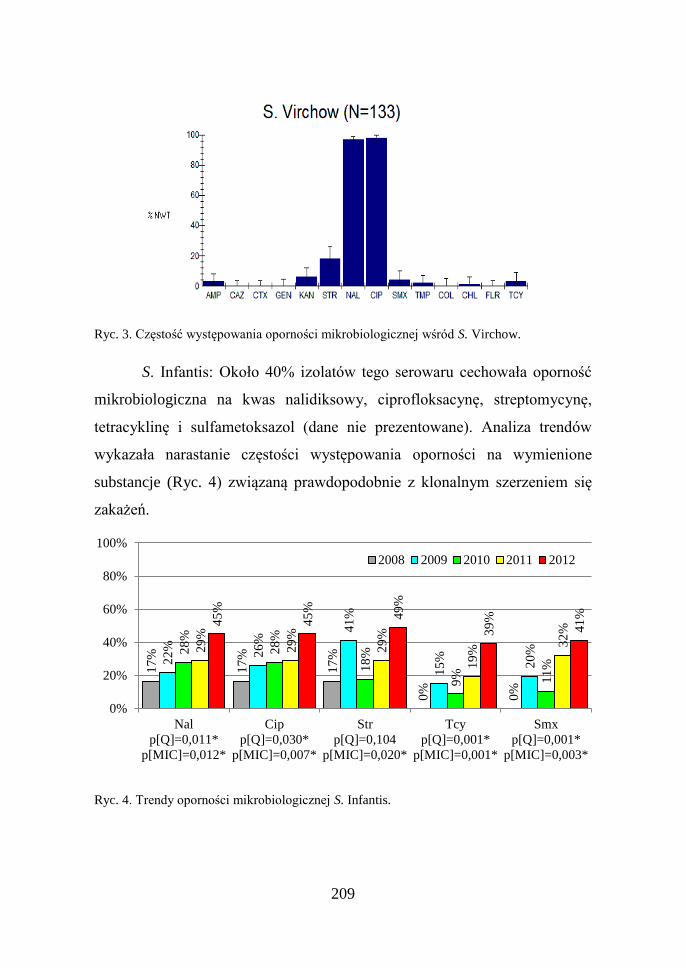
209
Ryc. 3. Częstość występowania oporności mikrobiologicznej wśród S. Virchow.
S. Infantis: Około 40% izolatów tego serowaru cechowała oporność
mikrobiologiczna na kwas nalidiksowy, ciprofloksacynę, streptomycynę,
tetracyklinę i sulfametoksazol (dane nie prezentowane). Analiza trendów
wykazała narastanie częstości występowania oporności na wymienione
substancje (Ryc. 4) związaną prawdopodobnie z klonalnym szerzeniem się
zakażeń.
Ryc. 4. Trendy oporności mikrobiologicznej S. Infantis.
17
%
17
%
17
%
0%
0%
22
%
26
% 4
1%
15
%
20
%
28
%
28
%
18
%
9%
11
%
29
%
29
%
29
%
19
% 32
% 45
%
45
%
49
%
39
%
41
%
0%
20%
40%
60%
80%
100%
Nal
p[Q]=0,011*
p[MIC]=0,012*
Cip
p[Q]=0,030*
p[MIC]=0,007*
Str
p[Q]=0,104
p[MIC]=0,020*
Tcy
p[Q]=0,001*
p[MIC]=0,001*
Smx
p[Q]=0,001*
p[MIC]=0,003*
2008 2009 2010 2011 2012
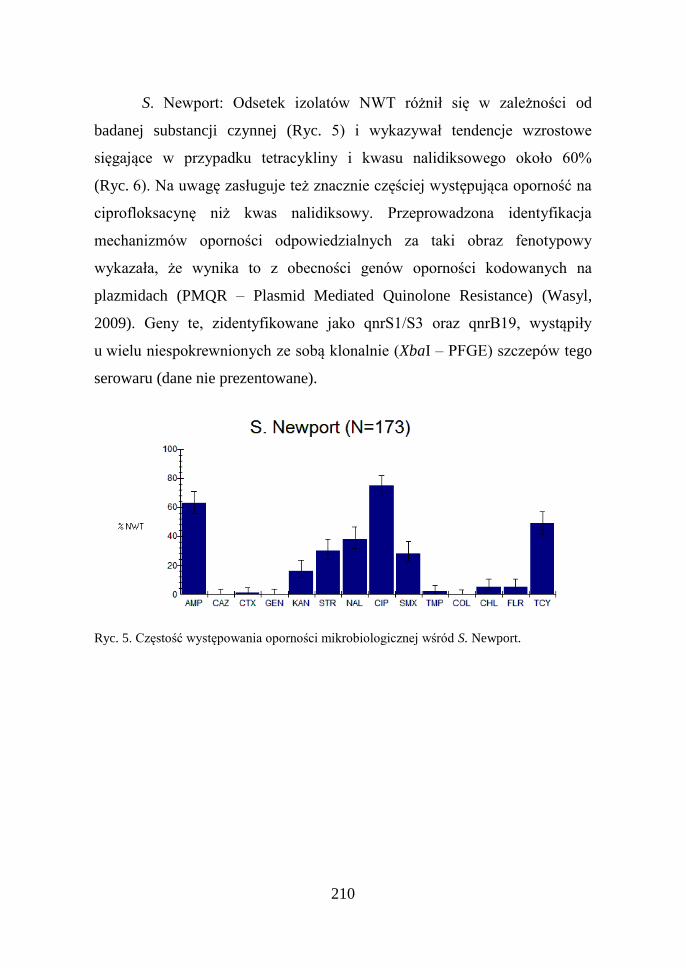
210
S. Newport: Odsetek izolatów NWT różnił się w zależności od
badanej substancji czynnej (Ryc. 5) i wykazywał tendencje wzrostowe
sięgające w przypadku tetracykliny i kwasu nalidiksowego około 60%
(Ryc. 6). Na uwagę zasługuje też znacznie częściej występująca oporność na
ciprofloksacynę niż kwas nalidiksowy. Przeprowadzona identyfikacja
mechanizmów oporności odpowiedzialnych za taki obraz fenotypowy
wykazała, że wynika to z obecności genów oporności kodowanych na
plazmidach (PMQR – Plasmid Mediated Quinolone Resistance) (Wasyl,
2009). Geny te, zidentyfikowane jako qnrS1/S3 oraz qnrB19, wystąpiły
u wielu niespokrewnionych ze sobą klonalnie (XbaI – PFGE) szczepów tego
serowaru (dane nie prezentowane).
Ryc. 5. Częstość występowania oporności mikrobiologicznej wśród S. Newport.
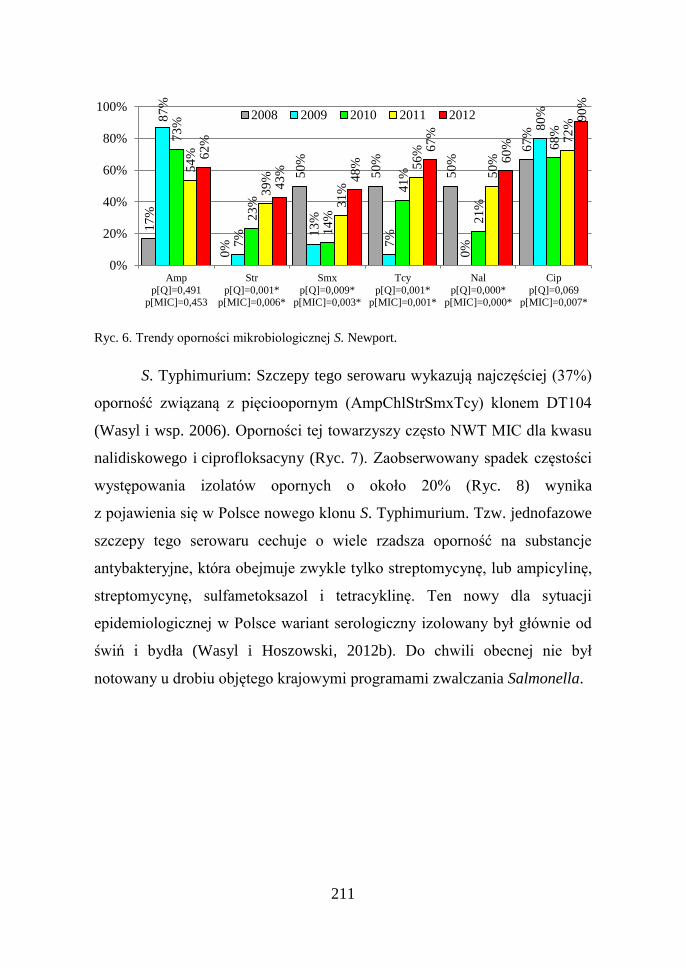
211
Ryc. 6. Trendy oporności mikrobiologicznej S. Newport.
S. Typhimurium: Szczepy tego serowaru wykazują najczęściej (37%)
oporność związaną z pięcioopornym (AmpChlStrSmxTcy) klonem DT104
(Wasyl i wsp. 2006). Oporności tej towarzyszy często NWT MIC dla kwasu
nalidiskowego i ciprofloksacyny (Ryc. 7). Zaobserwowany spadek częstości
występowania izolatów opornych o około 20% (Ryc. 8) wynika
z pojawienia się w Polsce nowego klonu S. Typhimurium. Tzw. jednofazowe
szczepy tego serowaru cechuje o wiele rzadsza oporność na substancje
antybakteryjne, która obejmuje zwykle tylko streptomycynę, lub ampicylinę,
streptomycynę, sulfametoksazol i tetracyklinę. Ten nowy dla sytuacji
epidemiologicznej w Polsce wariant serologiczny izolowany był głównie od
świń i bydła (Wasyl i Hoszowski, 2012b). Do chwili obecnej nie był
notowany u drobiu objętego krajowymi programami zwalczania Salmonella.
17
%
0%
50
%
50
%
50
% 6
7%
87
%
7%
13
%
7%
0%
80
%
73
%
23
%
14
%
41
%
21
%
68
%
54
%
39
%
31
%
56
%
50
%
72
%
62
%
43
%
48
% 6
7%
60
%
90
%
0%
20%
40%
60%
80%
100%
Amp
p[Q]=0,491p[MIC]=0,453
Str
p[Q]=0,001*p[MIC]=0,006*
Smx
p[Q]=0,009*p[MIC]=0,003*
Tcy
p[Q]=0,001*p[MIC]=0,001*
Nal
p[Q]=0,000*p[MIC]=0,000*
Cip
p[Q]=0,069p[MIC]=0,007*
2008 2009 2010 2011 2012
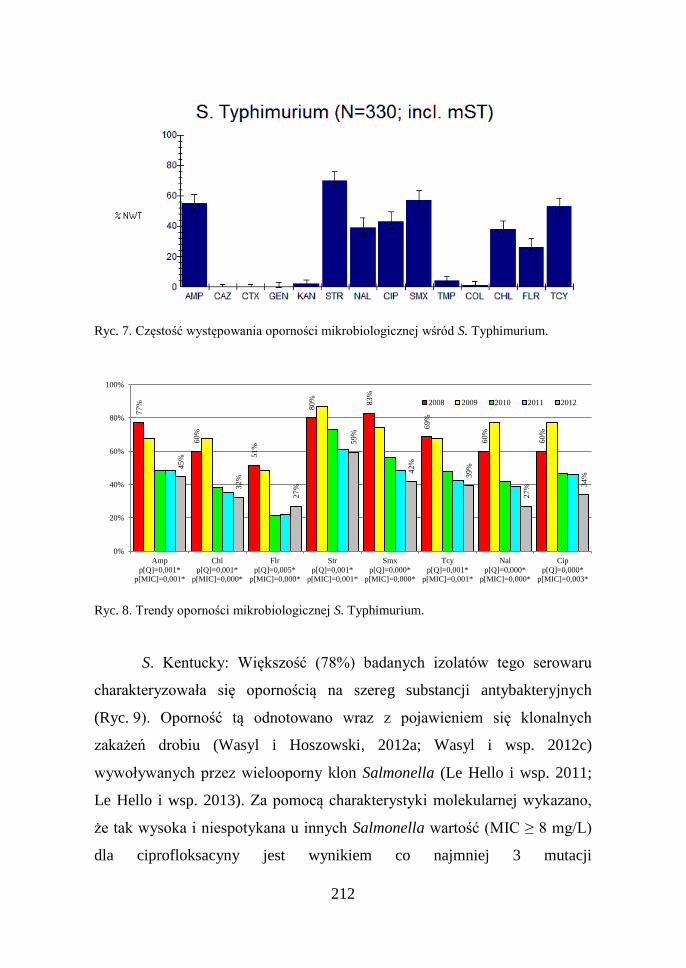
212
Ryc. 7. Częstość występowania oporności mikrobiologicznej wśród S. Typhimurium.
Ryc. 8. Trendy oporności mikrobiologicznej S. Typhimurium.
S. Kentucky: Większość (78%) badanych izolatów tego serowaru
charakteryzowała się opornością na szereg substancji antybakteryjnych
(Ryc. 9). Oporność tą odnotowano wraz z pojawieniem się klonalnych
zakażeń drobiu (Wasyl i Hoszowski, 2012a; Wasyl i wsp. 2012c)
wywoływanych przez wielooporny klon Salmonella (Le Hello i wsp. 2011;
Le Hello i wsp. 2013). Za pomocą charakterystyki molekularnej wykazano,
że tak wysoka i niespotykana u innych Salmonella wartość (MIC ≥ 8 mg/L)
dla ciprofloksacyny jest wynikiem co najmniej 3 mutacji
77%
60%
51%
80%
83%
69%
60%
60%
45%
32
%
27%
59%
42%
39%
27%
34%
0%
20%
40%
60%
80%
100%
Amp
p[Q]=0,001*
p[MIC]=0,001*
Chl
p[Q]=0,001*
p[MIC]=0,000*
Flr
p[Q]=0,005*
p[MIC]=0,000*
Str
p[Q]=0,001*
p[MIC]=0,001*
Smx
p[Q]=0,000*
p[MIC]=0,000*
Tcy
p[Q]=0,001*
p[MIC]=0,001*
Nal
p[Q]=0,000*
p[MIC]=0,000*
Cip
p[Q]=0,000*
p[MIC]=0,003*
2008 2009 2010 2011 2012
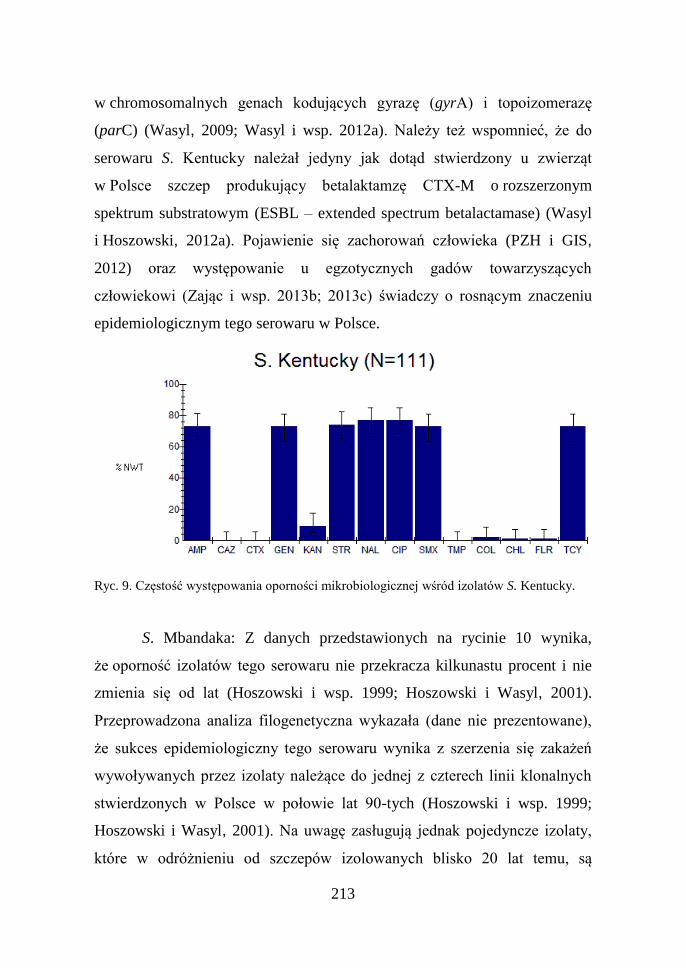
213
w chromosomalnych genach kodujących gyrazę (gyrA) i topoizomerazę
(parC) (Wasyl, 2009; Wasyl i wsp. 2012a). Należy też wspomnieć, że do
serowaru S. Kentucky należał jedyny jak dotąd stwierdzony u zwierząt
w Polsce szczep produkujący betalaktamzę CTX-M o rozszerzonym
spektrum substratowym (ESBL – extended spectrum betalactamase) (Wasyl
i Hoszowski, 2012a). Pojawienie się zachorowań człowieka (PZH i GIS,
2012) oraz występowanie u egzotycznych gadów towarzyszących
człowiekowi (Zając i wsp. 2013b; 2013c) świadczy o rosnącym znaczeniu
epidemiologicznym tego serowaru w Polsce.
Ryc. 9. Częstość występowania oporności mikrobiologicznej wśród izolatów S. Kentucky.
S. Mbandaka: Z danych przedstawionych na rycinie 10 wynika,
że oporność izolatów tego serowaru nie przekracza kilkunastu procent i nie
zmienia się od lat (Hoszowski i wsp. 1999; Hoszowski i Wasyl, 2001).
Przeprowadzona analiza filogenetyczna wykazała (dane nie prezentowane),
że sukces epidemiologiczny tego serowaru wynika z szerzenia się zakażeń
wywoływanych przez izolaty należące do jednej z czterech linii klonalnych
stwierdzonych w Polsce w połowie lat 90-tych (Hoszowski i wsp. 1999;
Hoszowski i Wasyl, 2001). Na uwagę zasługują jednak pojedyncze izolaty,
które w odróżnieniu od szczepów izolowanych blisko 20 lat temu, są
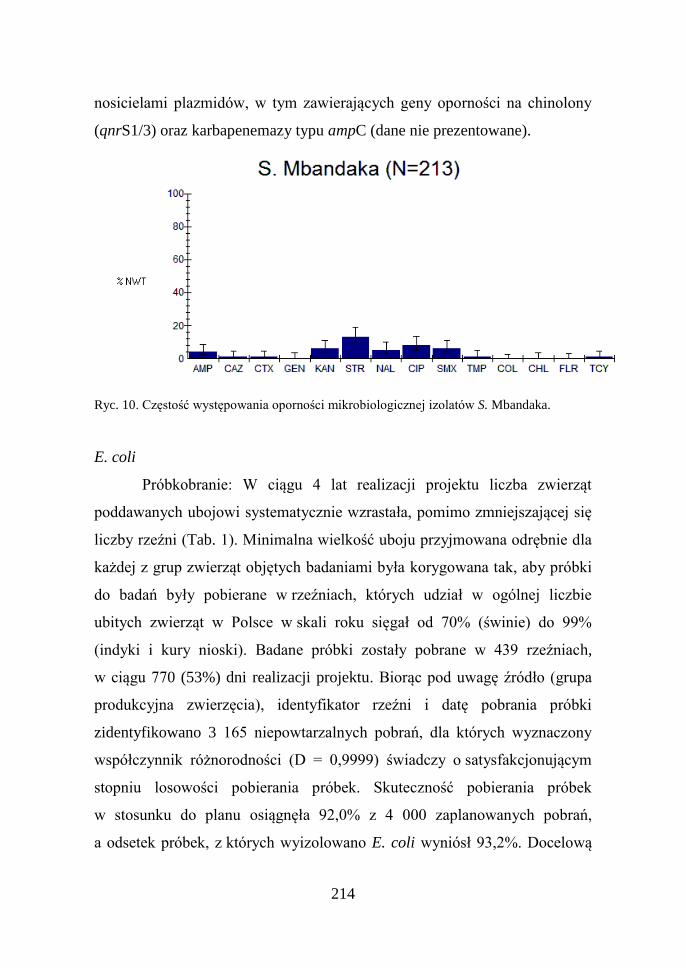
214
nosicielami plazmidów, w tym zawierających geny oporności na chinolony
(qnrS1/3) oraz karbapenemazy typu ampC (dane nie prezentowane).
Ryc. 10. Częstość występowania oporności mikrobiologicznej izolatów S. Mbandaka.
E. coli
Próbkobranie: W ciągu 4 lat realizacji projektu liczba zwierząt
poddawanych ubojowi systematycznie wzrastała, pomimo zmniejszającej się
liczby rzeźni (Tab. 1). Minimalna wielkość uboju przyjmowana odrębnie dla
każdej z grup zwierząt objętych badaniami była korygowana tak, aby próbki
do badań były pobierane w rzeźniach, których udział w ogólnej liczbie
ubitych zwierząt w Polsce w skali roku sięgał od 70% (świnie) do 99%
(indyki i kury nioski). Badane próbki zostały pobrane w 439 rzeźniach,
w ciągu 770 (53%) dni realizacji projektu. Biorąc pod uwagę źródło (grupa
produkcyjna zwierzęcia), identyfikator rzeźni i datę pobrania próbki
zidentyfikowano 3 165 niepowtarzalnych pobrań, dla których wyznaczony
współczynnik różnorodności (D = 0,9999) świadczy o satysfakcjonującym
stopniu losowości pobierania próbek. Skuteczność pobierania próbek
w stosunku do planu osiągnęła 92,0% z 4 000 zaplanowanych pobrań,
a odsetek próbek, z których wyizolowano E. coli wyniósł 93,2%. Docelową
215
liczbę 170 izolatów (EFSA, 2008) uzyskano corocznie z każdej grupy
zwierząt objętej badaniami, z wyjątkiem kur niosek. Interesujące,
że w przypadku kur niosek 70 z 638 (11,0%) próbek pochodziło od zwierząt
importowanych bezpośrednio przed ubojem z Holandii (N=21), Niemiec
(N=17), Słowacji (N=11) oraz Austrii, Belgii, Czech, Litwy i Łotwy.
W przypadku pozostałych grup zwierząt, próbki od zwierząt importowanych
zdarzały się incydentalnie ( 0,6%), a w przypadku bydła nie odnotowano
takiej sytuacji.
Oporność mikrobiologiczną stwierdzono w odniesieniu do każdej
z badanych substancji antybakteryjnych, a częstość jej występowania wahała
się od 0,9% (kolistyna) do 43,3% (tetracyklina) i zależała też od źródła
izolacji E. coli (Tab. 3). Zmiany częstości występowania oporności w czasie
odnotowano w przypadku 16 z 65 analizowanych kombinacji substancji
czynnej i źródła izolacji. W 9 przypadkach zmiana ta miała charakter
jakościowy, a w pozostałych dotyczyła przesunięcia wartości MIC nie
skutkujących zmianą częstości występowania kategorii NWT.
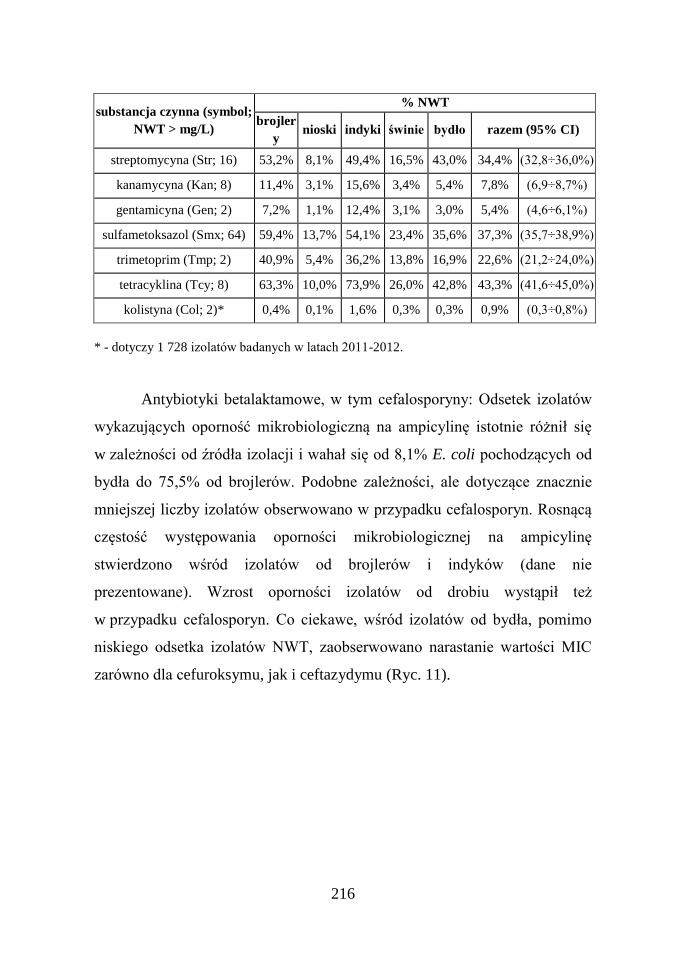
Tab. 3. Odsetek izolatów E. coli wykazujących oporność mikrobiologiczną na badane
substancje antybakteryjne.
substancja czynna (symbol;
NWT > mg/L)
% NWT
brojler
y nioski indyki świnie bydło razem (95% CI)
ampicilina (Amp; 8) 75,5% 34,3% 68,5% 26,1% 8,1% 42,3% (40,7÷44,0%)
cefotaksym (Ctx; 0,25) 8,7% 1,3% 3,8% 3,9% 1,4% 3,8% (3,1÷4,4%)
ceftazydym (Caz; 0,5) 8,1% 1,0% 3,0% 4,5% 1,4% 3,6% (2,9÷4,2%)
kwas nalidiksowy (Nal; 16) 74,0% 3,0% 53,8% 35,7% 6,9% 34,3% (32,7÷35,9%)
ciprofloksacyna (Cip; 0,064) 79,8% 3,3% 61,0% 42,6% 10,4% 39,0% (37,3÷40,6%)
chloramfenikol (Chl; 16) 17,3% 1,4% 20,3% 3,3% 7,7% 10,1% (9,1÷11,1%)
florfenikol (Flr, 16) 8,4% 0,7% 6,2% 0,9% 3,0% 3,8% (3,2÷4,5%)
216
substancja czynna (symbol;
NWT > mg/L)
% NWT
brojler
y nioski indyki świnie bydło razem (95% CI)
streptomycyna (Str; 16) 53,2% 8,1% 49,4% 16,5% 43,0% 34,4% (32,8÷36,0%)
kanamycyna (Kan; 8) 11,4% 3,1% 15,6% 3,4% 5,4% 7,8% (6,9÷8,7%)
gentamicyna (Gen; 2) 7,2% 1,1% 12,4% 3,1% 3,0% 5,4% (4,6÷6,1%)
sulfametoksazol (Smx; 64) 59,4% 13,7% 54,1% 23,4% 35,6% 37,3% (35,7÷38,9%)
trimetoprim (Tmp; 2) 40,9% 5,4% 36,2% 13,8% 16,9% 22,6% (21,2÷24,0%)
tetracyklina (Tcy; 8) 63,3% 10,0% 73,9% 26,0% 42,8% 43,3% (41,6÷45,0%)
kolistyna (Col; 2)* 0,4% 0,1% 1,6% 0,3% 0,3% 0,9% (0,3÷0,8%)
* - dotyczy 1 728 izolatów badanych w latach 2011-2012.
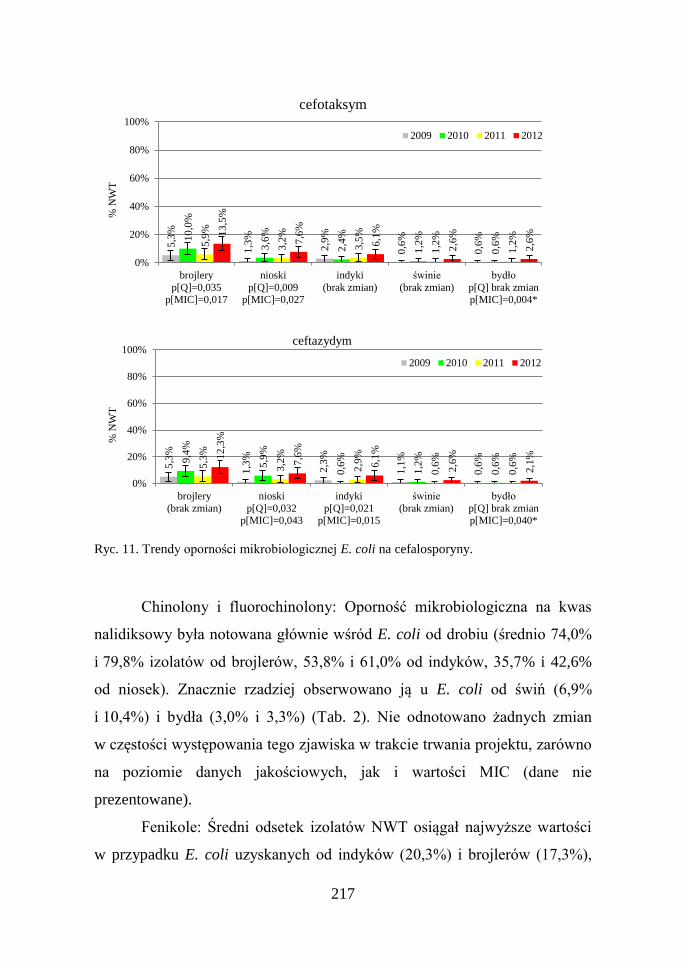
Antybiotyki betalaktamowe, w tym cefalosporyny: Odsetek izolatów
wykazujących oporność mikrobiologiczną na ampicylinę istotnie różnił się
w zależności od źródła izolacji i wahał się od 8,1% E. coli pochodzących od
bydła do 75,5% od brojlerów. Podobne zależności, ale dotyczące znacznie
mniejszej liczby izolatów obserwowano w przypadku cefalosporyn. Rosnącą
częstość występowania oporności mikrobiologicznej na ampicylinę
stwierdzono wśród izolatów od brojlerów i indyków (dane nie
prezentowane). Wzrost oporności izolatów od drobiu wystąpił też
w przypadku cefalosporyn. Co ciekawe, wśród izolatów od bydła, pomimo
niskiego odsetka izolatów NWT, zaobserwowano narastanie wartości MIC
zarówno dla cefuroksymu, jak i ceftazydymu (Ryc. 11).
217
Ryc. 11. Trendy oporności mikrobiologicznej E. coli na cefalosporyny.
Chinolony i fluorochinolony: Oporność mikrobiologiczna na kwas
nalidiksowy była notowana głównie wśród E. coli od drobiu (średnio 74,0%
i 79,8% izolatów od brojlerów, 53,8% i 61,0% od indyków, 35,7% i 42,6%
od niosek). Znacznie rzadziej obserwowano ją u E. coli od świń (6,9%
i 10,4%) i bydła (3,0% i 3,3%) (Tab. 2). Nie odnotowano żadnych zmian
w częstości występowania tego zjawiska w trakcie trwania projektu, zarówno
na poziomie danych jakościowych, jak i wartości MIC (dane nie
prezentowane).
Fenikole: Średni odsetek izolatów NWT osiągał najwyższe wartości
w przypadku E. coli uzyskanych od indyków (20,3%) i brojlerów (17,3%),
5,3
%
1,3
%
2,9
%
0,6
%
0,6
%
10
,0%
3,6
%
2,4
%
1,2
%
0,6
%
5,9
%
3,2
%
3,5
%
1,2
%
1,2
%
13
,5%
7,6
%
6,1
%
2,6
%
2,6
%
0%
20%
40%
60%
80%
100%
brojlery
p[Q]=0,035p[MIC]=0,017
nioski
p[Q]=0,009p[MIC]=0,027
indyki
(brak zmian)
świnie
(brak zmian)
bydło
p[Q] brak zmianp[MIC]=0,004*
% N
WT
cefotaksym
2009 2010 2011 2012
5,3
%
1,3
%
2,3
%
1,1
%
0,6
%
9,4
%
5,9
%
0,6
%
1,2
%
0,6
%
5,3
%
3,2
%
2,9
%
0,6
%
0,6
%
12
,3%
7,6
%
6,1
%
2,6
%
2,1
%
0%
20%
40%
60%
80%
100%
brojlery
(brak zmian)
nioski
p[Q]=0,032p[MIC]=0,043
indyki
p[Q]=0,021p[MIC]=0,015
świnie
(brak zmian)
bydło
p[Q] brak zmianp[MIC]=0,040*
% N
WT
ceftazydym
2009 2010 2011 2012
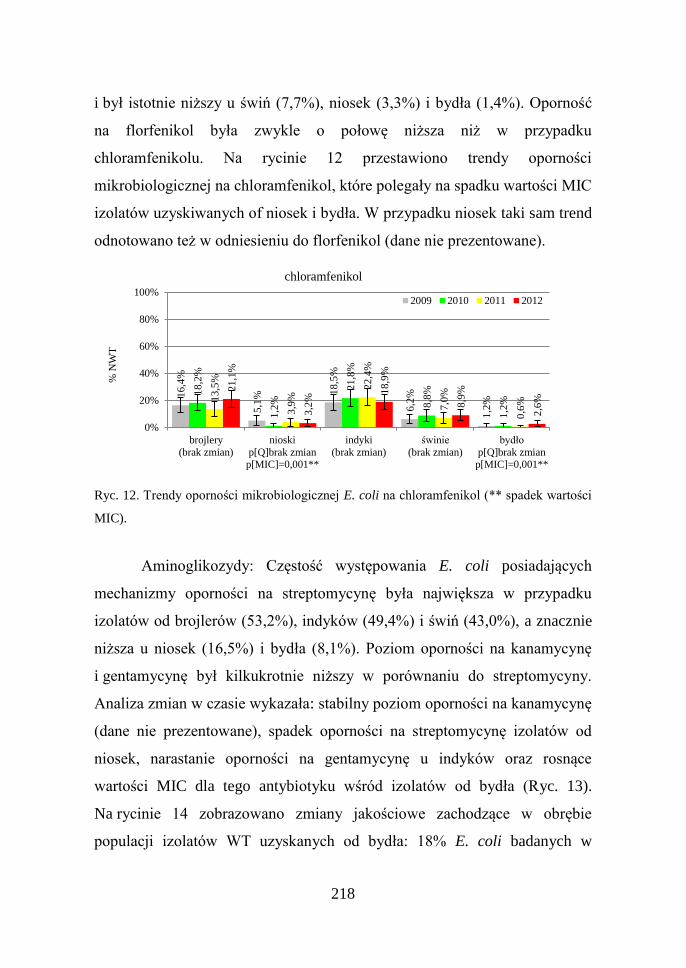
218
i był istotnie niższy u świń (7,7%), niosek (3,3%) i bydła (1,4%). Oporność
na florfenikol była zwykle o połowę niższa niż w przypadku
chloramfenikolu. Na rycinie 12 przestawiono trendy oporności
mikrobiologicznej na chloramfenikol, które polegały na spadku wartości MIC
izolatów uzyskiwanych of niosek i bydła. W przypadku niosek taki sam trend
odnotowano też w odniesieniu do florfenikol (dane nie prezentowane).
Ryc. 12. Trendy oporności mikrobiologicznej E. coli na chloramfenikol (** spadek wartości
MIC).
Aminoglikozydy: Częstość występowania E. coli posiadających
mechanizmy oporności na streptomycynę była największa w przypadku
izolatów od brojlerów (53,2%), indyków (49,4%) i świń (43,0%), a znacznie
niższa u niosek (16,5%) i bydła (8,1%). Poziom oporności na kanamycynę
i gentamycynę był kilkukrotnie niższy w porównaniu do streptomycyny.
Analiza zmian w czasie wykazała: stabilny poziom oporności na kanamycynę
(dane nie prezentowane), spadek oporności na streptomycynę izolatów od
niosek, narastanie oporności na gentamycynę u indyków oraz rosnące
wartości MIC dla tego antybiotyku wśród izolatów od bydła (Ryc. 13).
Na rycinie 14 zobrazowano zmiany jakościowe zachodzące w obrębie
populacji izolatów WT uzyskanych od bydła: 18% E. coli badanych w
16
,4%
5,1
% 18
,5%
6,2
%
1,2
% 1
8,2
%
1,2
%
21
,8%
8,8
%
1,2
%
13
,5%
3,9
% 2
2,4
%
7,0
%
0,6
%
21
,1%
3,2
% 18
,9%
8,9
%
2,6
%
0%
20%
40%
60%
80%
100%
brojlery
(brak zmian)
nioski
p[Q]brak zmianp[MIC]=0,001**
indyki
(brak zmian)
świnie
(brak zmian)
bydło
p[Q]brak zmianp[MIC]=0,001**
% N
WT
chloramfenikol
2009 2010 2011 2012
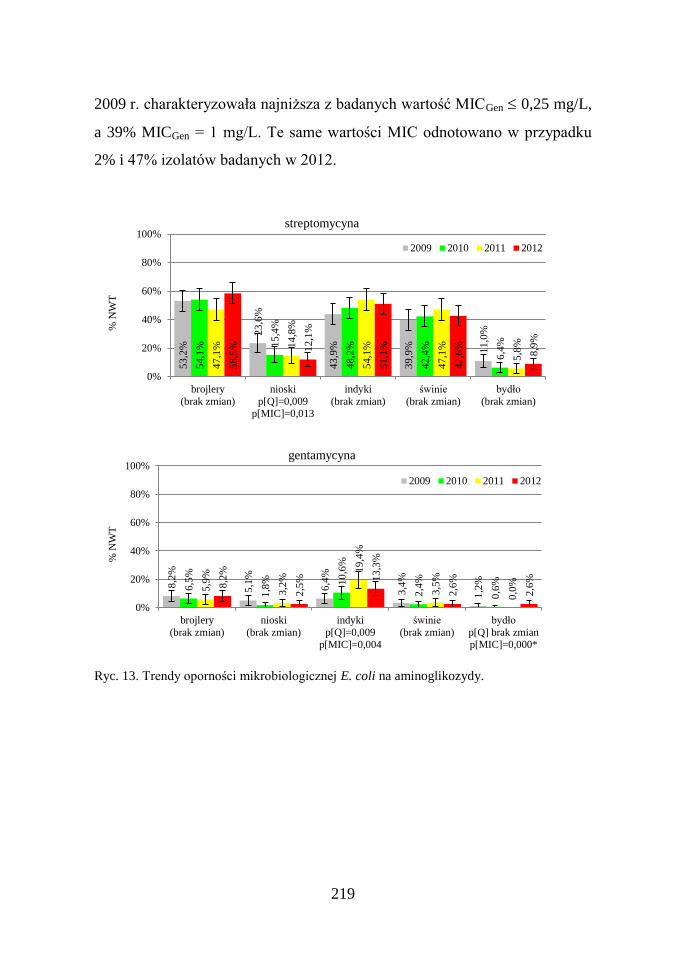
219
2009 r. charakteryzowała najniższa z badanych wartość MICGen 0,25 mg/L,
a 39% MICGen = 1 mg/L. Te same wartości MIC odnotowano w przypadku
2% i 47% izolatów badanych w 2012.
Ryc. 13. Trendy oporności mikrobiologicznej E. coli na aminoglikozydy.
53
,2%
23
,6%
43
,9%
39
,9%
11
,0%
54
,1%
15
,4%
48
,2%
42
,4%
6,4
%
47
,1%
14
,8%
54
,1%
47
,1%
5,8
%
58
,5%
12
,1%
51
,1%
42
,6%
8,9
%
0%
20%
40%
60%
80%
100%
brojlery
(brak zmian)
nioski
p[Q]=0,009p[MIC]=0,013
indyki
(brak zmian)
świnie
(brak zmian)
bydło
(brak zmian)
% N
WT
streptomycyna
2009 2010 2011 2012
8,2
%
5,1
%
6,4
%
3,4
%
1,2
%
6,5
%
1,8
%
10
,6%
2,4
%
0,6
%
5,9
%
3,2
% 1
9,4
%
3,5
%
0,0
%
8,2
%
2,5
%
13
,3%
2,6
%
2,6
%
0%
20%
40%
60%
80%
100%
brojlery
(brak zmian)
nioski
(brak zmian)
indyki
p[Q]=0,009p[MIC]=0,004
świnie
(brak zmian)
bydło
p[Q] brak zmianp[MIC]=0,000*
% N
WT
gentamycyna
2009 2010 2011 2012
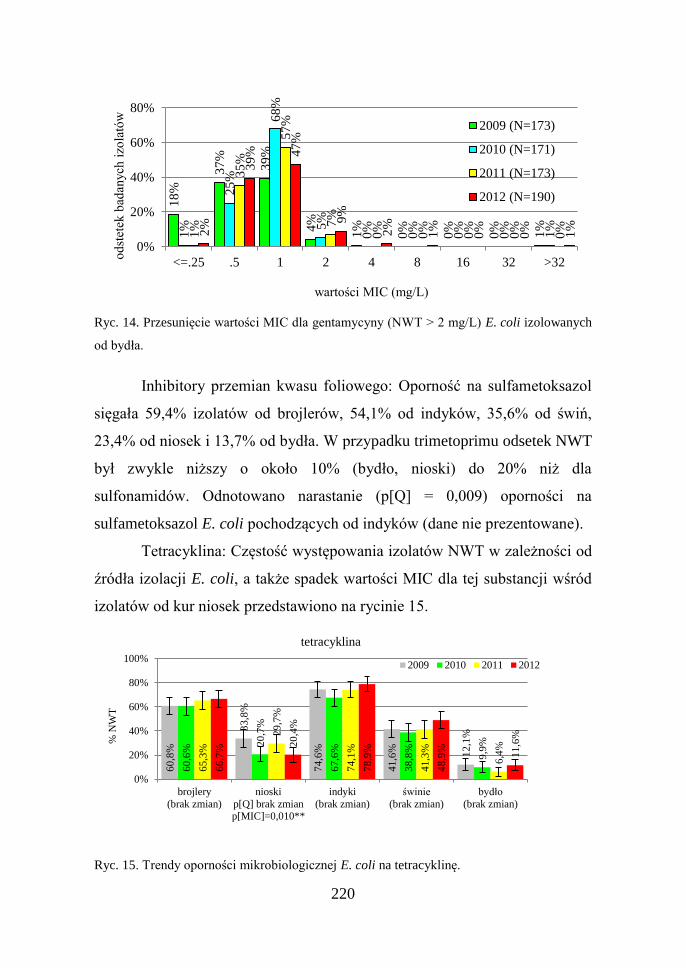
220
Ryc. 14. Przesunięcie wartości MIC dla gentamycyny (NWT > 2 mg/L) E. coli izolowanych
od bydła.
Inhibitory przemian kwasu foliowego: Oporność na sulfametoksazol
sięgała 59,4% izolatów od brojlerów, 54,1% od indyków, 35,6% od świń,
23,4% od niosek i 13,7% od bydła. W przypadku trimetoprimu odsetek NWT
był zwykle niższy o około 10% (bydło, nioski) do 20% niż dla
sulfonamidów. Odnotowano narastanie (p[Q] = 0,009) oporności na
sulfametoksazol E. coli pochodzących od indyków (dane nie prezentowane).
Tetracyklina: Częstość występowania izolatów NWT w zależności od
źródła izolacji E. coli, a także spadek wartości MIC dla tej substancji wśród
izolatów od kur niosek przedstawiono na rycinie 15.
Ryc. 15. Trendy oporności mikrobiologicznej E. coli na tetracyklinę.
18
%
37
%
39
%
4%
1%
0%
0%
0%
1%
1%
25
%
68
%
5%
0%
0%
0%
0%
1%
1%
35
%
57
%
7%
0%
0%
0%
0%
0%
2%
39
%
47
%
9%
2%
1%
0%
0%
1%
0%
20%
40%
60%
80%
<=.25 .5 1 2 4 8 16 32 >32
od
stet
ek b
adan
ych
izo
lató
w
wartości MIC (mg/L)
2009 (N=173)
2010 (N=171)
2011 (N=173)
2012 (N=190)
60
,8%
33
,8%
74
,6%
41
,6%
12
,1%
60
,6%
20
,7%
67
,6%
38
,8%
9,9
%
65
,3%
29
,7%
74
,1%
41
,3%
6,4
%
66
,7%
20
,4%
78
,9%
48
,9%
11
,6%
0%
20%
40%
60%
80%
100%
brojlery
(brak zmian)
nioski
p[Q] brak zmianp[MIC]=0,010**
indyki
(brak zmian)
świnie
(brak zmian)
bydło
(brak zmian)
% N
WT
tetracyklina
2009 2010 2011 2012

221
Kolistyna: Izolaty NWT obserwowano sporadycznie ( 0,4%),
najczęściej wśród E. coli pochodzących od indyków (1,6%). Substancja ta
została uwzględniona w prowadzonym monitoringu od roku 2011, dlatego
też nie było możliwe przeprowadzenie analizy trendów.
Dyskusja
Monitorowanie oporności na substancje antybakteryjne
W ostatnich latach wiele krajów podjęło systematyczne badania
oporności bakterii na substancje antybakteryjne (Bronzwaer i wsp. 2008;
Deckert i wsp. 2010; Ozaki i wsp. 2011; Morfin-Otero i wsp. 2012; Tadesse
i wsp. 2012). Prezentowane wyniki są efektem badań prowadzonych
w ramach monitoringu wdrożonego w krajach UE (EFSA, 2007; Bronzwaer
i wsp. 2008; EFSA, 2008). Jego niezaprzeczalną zaletą jest zharmonizowana
metodyka, która pozwala na bezpośrednie porównywanie pomiędzy izolatami
pochodzącymi z różnych źródeł, regionów geograficznych i okresów
(MARAN, 2012; Schroeter i wsp. 2013). Oznaczanie najmniejszego stężenia
hamującego stałego panelu substancji antybakteryjnych, daje możliwość
oceny trendów oporności w czasie. Analiza taka jest zwykle wykonywana
w odniesieniu do wyników jakościowych, otrzymywanych po interpretacji
MIC (Kronvall, 2010; Kaesbohrer i wsp. 2012; Morfin-Otero i wsp. 2012;
SVARM, 2012; Tadesse i wsp. 2012; van der Bij i wsp. 2012). Metoda ta
daje również możliwość analizy ilościowej, polegającej na bezpośrednim
porównywaniu częstości występowania bakterii charakteryzujących się
określoną wartością MIC (EFSA, 2012a). Prezentowane wyniki są jednym
z pierwszych opracowań tego typu (Lundin i wsp. 2008; Wasyl i wsp. 2013a;
Wasyl i wsp. 2013c).
Bardzo ważnym elementem monitoringu oporności jest przyjęcie
jednolitych kryteriów interpretacji wyników. Zastosowanie kryteriów
epidemiologicznych (epidemiological cut-off values), chociaż często

222
krytykowane przez klinicystów i przemysł farmaceutyczny (Schwarz i wsp.
2010; de Jong i wsp. 2012), daje możliwość wczesnego wykrycia oporności
występującej nawet na bardzo niskim poziomie (Lundin i wsp. 2008).
Potwierdzeniem tego jest stwierdzona w prezentowanych badaniach oporność
E. coli na cefalosporyny oraz jej trendy. Dodatkowym argumentem
przeciwko stosowaniu kryteriów klinicznych w badaniach monitoringowych
jest fakt, że izolaty pochodzą zwykle od klinicznie zdrowych zwierząt,
z żywności i pasz.
Możliwość wykrycia nowopojawiającej się oporności lub jej trendów
w czasie zależy od liczby izolatów objętych badaniami monitoringowymi
(Bronzwaer i wsp. 2008). Przyjęta zasada badania 170 izolatów okazała się
skuteczna w odniesieniu do E. coli i chociaż zastosowany program objął
zaledwie niewielki ułamek populacji ubijanych co roku zwierząt (Tab. 3),
to losowe i równomiernie pobierane, przez cały okres badań, próbki
pozwoliły ocenić skalę problemu oporności. Niekiedy jednak, tak jak to
miało miejsce w przypadku kur niosek, mogą występować trudności
z pozyskaniem próbek do badań. Ograniczona dostępność izolatów dotyczy
z reguły bakterii patogennych (EFSA, 2008). Prezentowane wyniki
oznaczania oporności Salmonella poddają w wątpliwość zalecany sposób
wyboru izolatów (EFSA, 2007; Bronzwaer i wsp. 2008), który powoduje,
że badania obejmują głównie najczęściej występujące serowary. Tym samym
serowary Salmonella o mniejszym znaczeniu epidemiologicznym, które
jednak mogą charakteryzować się opornością istotną z punktu widzenia
epidemiologii, mogą zostać pominięte w badaniach monitoringowych.
Przykładem takiej sytuacji był oporny na chinolony (pojedyncza mutacja
gyrA) szczep S. Stanley (Wasyl i wsp. 2013b), którego oporność została
zidentyfikowana w Polsce tylko dzięki dochodzeniu epidemiologicznemu
prowadzonemu

223
w innych krajach UE (ECDC, 2012). W przypadku Salmonella zdecydowanie
skuteczniejszym rozwiązaniem wydaje się uwzględnianie przynależności
serologicznej przy wyborze izolatów do oznaczania oporności. Pozwoliłoby
to uniknąć sytuacji zaobserwowanej w niniejszych badaniach, w których
blisko 40% badanych izolatów Salmonella należało do najczęściej
występującego serowaru Enteritidis. Sześć kolejnych serowarów było
reprezentowanych przez wystarczająco liczną grupę izolatów do
przeprowadzenia analizy trendów oporności, a 30% oznaczeń wykonanych
w ciągu 5-letniego cyklu badań dotyczyła pojedynczych szczepów
należących do 234 serowarów. W przypadku tych ostatnich dopiero wzrost
znaczenia epidemiologicznego pozwala wykryć i w pełni zidentyfikować
mechanizmy oporności na substancje antybakteryjne (Wasyl i wsp. 2012a;
Wasyl i Hoszowski, 2012a; Wasyl i wsp. 2012c; Wasyl i wsp. 2013b).
Oporność jako wynik presji selekcyjnej
Stosowanie substancji antybakteryjnych jest podstawowym
czynnikiem powodującym selekcję i rozprzestrzenianie się bakterii opornych.
Monitorowanie konsumpcji substancji antybakteryjnych u zwierząt rzeźnych,
realizowane równocześnie z monitorowaniem oporności izolowanych od nich
bakterii, pozwala na precyzyjne określenie tych związków (EMA, 2012;
Krasucka i wsp. 2012). Szczególnie przydatne w tym zakresie jest
wykorzystanie niepatogennych, indykatorowych E. coli występujących tak
u zwierząt rzeźnych, jak i wolnożyjących (de Jong i wsp. 2012; EFSA
i ECDC, 2012; SVARM, 2012; Allen i wsp. 2013). Prezentowane wyniki
oporności komensalnych E. coli odzwierciedlają preferencje stosowania
określonych substancji czynnych oraz różne praktyki stosowane w chowie
zwierząt. Dla przykładu, stwierdzono, że 79,9% E. coli pochodzących od
bydła nie wykazywała obecności jakichkolwiek mechanizmów oporności
(dane nie prezentowane). Analiza danych dotyczących zwierząt, od których

224
izolowano te bakterie wykazała, że było to głównie bydło dorosłe (średnia
wieku: 46 miesięcy, najstarsza sztuka: 214 miesięcy, incydentalne przypadki
uboju bydła w wieku poniżej 2 lat). Prawdopodobnie kierowano je do uboju
z powodu niesatysfakcjonującej mleczności, a nie podawano substancji
antybakteryjnych ze względu na wytwarzane przez te zwierzęta mleko.
Z kolei u brojlerów i indyków substancje te mogą być stosowane okresowo,
w celach metafilaktycznych w czasie tuczu przy zachowaniu zasad karencji
ich stosowania przed ubojem. W wyniku tak dużej presji selekcyjnej
zaledwie 5,1% E. coli od brojlerów i 11,3% od izolatów indyczych objętych
tymi badaniami należało do kategorii WT (dane nie prezentowane).
Kolejnym przykładem stosowania substancji antybakteryjnych, którego
efektem jest wzrost częstości występowania oporności mikrobiologicznej na
chinolony i fluorochinolony jest S. Enteritidis izolowane od brojlerów
(Ryc. 1). Odnotowany w tym samym czasie stabilny poziom tej oporności
wśród izolatów od kur może wynikać również z mniejszej presji związanej
ze stosowaniem substancji antybakteryjnych u ptaków w trakcie nieśności.
Różna częstość występowania oporności komensalnych E. coli
świadczy o preferencjach w stosowaniu substancji antybakteryjnych
u określonych gatunków zwierząt (EMA, 2012; Krasucka i wsp. 2012). Na
obraz oporności może też wpływać import zwierząt. Stwierdzenie
w obecnych badaniach takich próbek wskazuje, że międzynarodowy obrót
zwierząt może być czynnikiem szerzenia się bakterii opornych lub
determinantów oporności (Wasyl i wsp. 2012b).
Zakażenia klonalne a oporność na substancje bakteryjne
Charakterystyka wybranych izolatów E. coli przy użyciu
elektroforezy PFGE genomowego DNA wykazała, że izolaty wykazujące
podobną oporność nie są epidemiologicznie związane (dane nie
prezentowane). W przypadku Salmonella sytuacja jest odmienna, a wyniki

225
oznaczania oporności jednoznacznie świadczą o konieczności analizowania
oporności na substancje antybakteryjne w obrębie poszczególnych
serowarów. Ścisłe związki oporności z przynależnością serologiczną
Salmonella powodują, że każda zmiana sytuacji epidemiologicznej, taka jak
pojawienie się lub zanikanie jakiegoś serowaru na danym obszarze
geograficznym, czy też w określonym sektorze produkcji zwierzęcej, rzutuje
na poziom oporności uzyskiwanych izolatów Salmonella. W przypadku
bakterii patogennych ma to związek nie tyle ze stosowaniem substancji
antybakteryjnych, ale wynika z szerzenia zakażeń wywoływanych przez klon
bakteryjny charakteryzujący się określonym profilem oporności (Wasyl
i Hoszowski, 2012b; 2012a; Wasyl i wsp. 2013b). Obok wspomnianych już
S. Enteritidis i S. Stanley (SVARM, 2012), dotyczy to też właściwie
wszystkich izolatów S. Virchow (Ryc. 4) i S. Kentucky (Wasyl i Hoszowski,
2012a). Izolaty te charakteryzuje kodowana chromosomalnie oporność na
chinolony i fluorochinolony (Wasyl i wsp. 2013d), która szerzy się na drodze
pionowej (z komórki macierzystej na potomną), a nie horyzontalnej, poprzez
np. mechanizmy kodowane plazmidowo przekazywane pomiędzy komórkami
tej samej generacji bakterii. Wspomniany wcześniej wzrost oporności na tą
klasę antybiotyków odnotowany w przypadku brojlerów (Ryc. 1) jest
prawdopodobnie wynikiem selekcji i klonalnego rozprzestrzenienia się
szczepu w tym sektorze produkcji drobiarskiej.
Przedstawiona na rycinie 4 narastająca oporność S. Infantis na
5 badanych substancji (NalCipStrTcySmx) jest z kolei efektem
rozprzestrzeniania się szczepu o takim właśnie profilu oporności, który
stwierdzono u 33% badanych izolatów tego serowaru. Narastanie oporności
odnotowano również u S. Newport (Ryc. 6), jednakże w przypadku tego
serowaru oporność mikrobiologiczna na poszczególne substancje
antybakteryjne była rozproszona pomiędzy różnymi liniami klonalnymi (dane

226
nie prezentowane), występującymi w różnych źródłach. Ich wspólną cechą
było częste występowanie PMQR, głównie qnrS1/S3, ale też qnrB (Veldman
i wsp. 2011; Wasyl i wsp. 2013d). Fenotypowo obecność tych mechanizmów
oporności wyraża się zdecydowanie wyższym odsetkiem izolatów opornych
na ciprofloksacynę niż kwas nalidiksowy. Co ciekawe, aż 66% wszystkich
Salmonella, u których w niniejszych badaniach stwierdzano geny PMQR,
reprezentowało S. Newport. Wyjaśnienie przyczyn tego zjawiska jest
niezaprzeczalnie interesującym tematem przyszłych badań.
Spektakularny sukces epidemiologiczny, tak w Polsce jak i w kilku
innych krajach, odniósł S. Kentucky. Klon określany mianem ST198
ewoluował przez kilkadziesiąt lat w Afryce Północnej, a zakażenia człowieka
były związane głównie z wyjazdami turystycznymi (Le Hello i wsp. 2011).
W Polsce ten wielooporny szczep charakteryzujący się wysoką opornością na
ciprofloksacynę pojawił się w 2009 r. prawdopodobnie w związku
z importem jaj wylęgowych indyków (Wasyl i Hoszowski, 2012a; Wasyl
i wsp. 2012c). Podobną sytuację obserwuje się w Niemczech (Beutlich i wsp.
2012). We Francji natomiast odnotowano zakażenia S. Kentucky w stadach
indyków, gdzie źródłem zakażenia był człowiek (informacja dzięki
uprzejmości S. Granier, ANSES). Zwykle jednak, to zakażenia indyków są
przyczyną zanieczyszczenia mięsa indyczego, co stwarza zagrożenie dla
zdrowia publicznego (Beutlich i wsp. 2012; PZH i GIS, 2012). Co ciekawe,
przeprowadzone badania wykazały, że zakażeniu klonem ST198, ulegają
również utrzymywane w domach egzotyczne gady, takie jak węże i warany,
a kontakt z tymi zwierzętami może stać się alternatywną dla żywności drogą
zakażenia człowieka tym niebezpiecznym patogenem (Zając i wsp. 2013a).
W odróżnieniu od opisanych powyżej przykładów, w których zmiana
sytuacji epidemiologicznej w obrębie danego serowaru Salmonella
powodowała wzrost odsetka izolatów wykazujących oporność

227
mikrobiologiczną, w przypadku S. Typhimurium odnotowano spadek
częstości występowania tego zjawiska (Ryc. 8). Jest to również związane
z epidemiologią S. Typhimurium i pojawieniem się w Polsce tzw.
jednofazowego wariantu tego serowaru. Izolaty te charakteryzuje rzadziej
obserwowana oporność i występują one głównie u świń, bydła
i uzyskiwanym od tych zwierząt mięsie (Wasyl i Hoszowski, 2012b).
Przykład S. Mbandaka również wskazuje, że nabywanie oporności
bakterii nie jest konieczne do szerzenia się zarazka i narastania jego
znaczenia epidemiologicznego. Serowar ten pojawił się w Polsce w połowie
lat 90-tych wywołując głównie zakażenia drobiu wodnego (Hoszowski
i Wasyl, 2001). Obecnie należy do najczęściej występujących serowarów
zarówno u zwierząt, jak i ludzi (Hoszowski i wsp. 2012; PZH i GIS, 2012),
chociaż oporność pozostaje na niskim poziomie (Ryc. 10). Jednakże
pojedyncze izolaty S. Mbandaka wykazujące oporność na fluorochinolony
(qnrS1/S3) oraz cefalosporyny (cefalosporynazy typu ampC), której
determinanty są kodowane na plazmidach, dowodzą roli presji selekcyjnej
antybiotyków stosowanych w chowie zwierząt (dane nie prezentowane) i ich
roli w szerzeniu się oporności na antybiotyki. Należy dodać, że substancje te
są uznawane przez Światową Organizację Zdrowia za niezmiernie ważne
z punktu widzenia ochrony zdrowia człowieka (Collignon i wsp. 2009).
Podsumowanie
Prowadzony monitoring oraz analiza jego wyników na poziomie
jakościowym i ilościowym (EFSA, 2012a) daje możliwość oceny zmian
w czasie oporności Salmonella i komensalnych E. coli izolowanych od
zwierząt w aspekcie jakościowym i ilościowym. W przypadku Salmonella
zmiany w oporności powiązane są z sytuacją epidemiologiczną w obrębie
poszczególnych serowarów, a u komensalnych E. coli ze skalą stosowania
substancji antybakteryjnych u zwierząt. Nawet jeżeli częstość występowania

228
oporności wydaje się niska i stabilna, analiza dystrybucji wartości MIC może
wykazać przesunięcia w obrębie kategorii WT i NWT. Może to być
traktowane jako system wczesnego ostrzegania przed narastaniem oporności
(Lundin i wsp. 2008). Tendencje do narastania oporności na betalaktamy
(w tym cefalosporyn), aminoglikozydy i sulfonamidy odnotowano wyłącznie
wśród izolatów od drobiu. Wbrew ogólnie przyjętej koncepcji dotyczącej
powszechnego narastania oporności (Collignon i wsp. 2009),
w pojedynczych przypadkach notowano obniżanie się wartości MIC (np.
fenikole, tetracyklina u izolatów E. coli od kur niosek i bydła). Przedstawione
wyniki dowodzą skomplikowanego charakteru zjawiska oporności bakterii na
substancje antybakteryjne, którego zrozumienie wymaga zarówno badań
monitoringowych, identyfikacji i charakterystyki mechanizmów oporności
i zasad ich rozprzestrzeniania się, ale też całościowego podejścia do
problemu oporności jako naturalnego zjawiska występującego
w ekosystemie.
Piśmiennictwo:
Allen S.E., Janecko N., Pearl D.L., Boerlin P., Reid-Smith R.J., Jardine C.M.:
Comparison of Escherichia coli recovery and antimicrobial resistance
in cecal, colon, and fecal samples collected from wild house mice
(Mus musculus). J. Wildl. Dis. 2013, 49, 432-436.
Beutlich J., Guerra B., Schroeter A., Arvand M., Szabo I., Helmuth R.:
Highly ciprofloxacin resistant Salmonella enterica serovar Kentucky isolates
in turkey meat and a human patient. Berl. Munch. Tierarztl. Wochenschr.
2012, 125, 89-95.

229
Bhullar K., Waglechner N., Pawlowski A., Koteva K., Banks E.D., Johnston
M. D., Barton H. A., Wright G. D.: Antibiotic resistance is prevalent in an
isolated cave microbiome. PLoS ONE 2012, 7, e34953.
Bronzwaer S., Aarestrup F., Battisti A., Bengtsson B., Piriz Duran S.,
Emborg H.D., Kahlmeter G., Mevius D., Regula G., Sanders P., Teale C.,
Wasyl D., De Smet K., Torren Edo J., Tul. P., Deluyker H., Makela P.:
Harmonised monitoring of antimicrobial resistance in Salmonella and
Campylobacter isolates from food animals in the European Union. Clin.
Microbiol. Infect. 2008, 14, 522-533.
Collignon P., Powers J.H., Chiller T.M., Aidara-Kane A., Aarestrup F.M.:
World Health Organization ranking of antimicrobials according to their
importance in human medicine: A critical step for developing risk
management strategies for the use of antimicrobials in food production
animals. Clin. Infect. Dis. 2009, 49, 132-141.
Czekalski N., Berthold T., Caucci S., Egli A., Burgmann H.: Increased levels
of multiresistant bacteria and resistance genes after wastewater treatment and
their dissemination into Lake Geneva, Switzerland. Front. Microbiol. 2012,
3, 106.
Dahmen S., Haenni M., Madec J.Y.: IncI1/ST3 plasmids contribute to
thedissemination of the blaCTX-M-1 gene in Escherichia coli from several
animal species in France. J. Antimicrob. Chemother. 2012, 67, 3011-3012.
Davies J., Davies D.: Origins and evolution of antibiotic resistance.
Microbiol. Mol. Biol. Rev. 2010, 74, 417-433.
de Jong A., Thomas V., Simjee S., Godinho K., Schiessl B., Klein U., Butty
P., Valle M., Marion H., Shryock T.R.: Pan-European monitoring of

230
susceptibility to human-use antimicrobial agents in enteric bacteria isolated
from healthy food-producing animals. J. Antimicrob. Chemother. 2012, 67,
638-651.
Deckert A., Gow S., Rosengren L., Leger D., Avery B., Daignault D., Dutil
L., Reid-Smith R., Irwin R.: Canadian integrated program for antimicrobial
resistance surveillance (CIPARS) farm program: results from finisher pig
surveillance. Zoonoses Public. Health 2010, 57 (Suppl 1), 71-84.
ECDC: Joint ECDC/EFSA rapid risk assessment: Multi-country outbreak
of Salmonella Stanley infections. Update. ECDC, Stockholm. 2012.
EFSA: Report of the Task Force on Zoonoses Data Collection including
a proposal for a harmonized monitoring scheme of antimicrobial resistance
in Salmonella in fowl (Gallus gallus), turkeys, and pigs and Campylobacter
jejuni and C. coli in broilers. EFSA Journal 2007, 96, 1-46.
EFSA: Report from the Task Force on Zoonoses Data Collection including
guidance for harmonized monitoring and reporting of antimicrobial resistance
in commensal Escherichia coli and Enterococcus spp. from food animals.
EFSA Journal 2008, 141, 1-44.
EFSA: Joint opinion on antimicrobial resistance (AMR) focused on zoonotic
infections. Scientific opinion of the European Centre for Disease Prevention
and Control; Scientific opinion of the Panel on Biological Hazards; Opinion
of the Committee for Medicinal Products for Veterinary Use; Scientific
opinion of the Scientific Committee on Emerging and Newly Identified
Health Risks. EFSA Journal 2009, 7, 1372, 1-78.

231
EFSA: Technical specifications for the analysis and reporting of data on
antimicrobial resistance in the European Union Summary Report. EFSA
Journal 2012, 10, 2587, 1-53.
EFSA: Technical specifications on the harmonised monitoring and reporting
of antimicrobial resistance in Salmonella, Campylobacter and indicator
Escherichia coli and Enterococcus spp. bacteria transmitted through food.
EFSA Journal 2012, 10, 2742, 1-64.
EFSA, ECDC: The European Union Summary Report on antimicrobial
resistance in zoonotic and indicator bacteria from humans, animals and food
in 2010. EFSA Journal 2012, 10, 2598, 1-233.
EMA: Sales of veterinary antimicrobial agents in 19 EU/EEA countries
in 2010. ESVAC report. London, 2012
EUCAST: Terminology relating to methods for the determination
of susceptibility of bacteria to antimicrobial agents. Definitive Document
E. Def 1.2, 2000, 6 (9), 503-508.
Hoszowski A., Skarżyńska M., Wasyl D., Zając M., Lalak A., Samcik I.,
Wnuk D.: Serowary Salmonella występujące u zwierząt, w żywności
i paszach w Polsce w latach 2005-2010. Medycyna Weter. 2012, 68,
411-417.
Hoszowski A., Wasyl D.: Typing of Salmonella enterica subsp. Enterica
serovar Mbandaka isolates. Vet. Microbiol. 2001, 80, 139-148.
Hoszowski A., Wasyl D., Truszczyński M.: Epidemiological investigation of
Salmonella serovar Mbandaka strains isolated from animals, their feed and
food products in Poland during the years 1995 - 1997. Polish J. Vet. Sci.
1999, 2, 43-48.

232
Kaesbohrer A., Schroeter A., Tenhagen B.A., Alt K., Guerra B., Appel B.:
Emerging antimicrobial resistance in commensal Escherichia coli with public
health relevance. Zoonoses Public. Health 2012, 59 (Suppl 2), 158-165.
Krasucka D., Cybulski W., Klimowicz A.: Ocena stosowania substancji
przeciwdrobnoustrojowych u świń i bydła w Polsce na podstawie badań
sondażowych w 2010 roku. Medycyna Weter. 2012, 68, 106-109.
Kronvall G.: Antimicrobial resistance 1979-2009 at Karolinska hospital,
Sweden: normalized resistance interpretation during a 30-year follow-up on
Staphylococcus aureus and Escherichia coli resistance development. APMIS,
2010, 118, 621-639.
Le Hello S., Harrois D., Bouchrif B., Sontag L., Elhani D., Guibert V.,
Zerouali K., Weill F.-X.: Highly drug-resistant Salmonella enterica serotype
Kentucky ST198-X1: a microbiological study. Lancet Infect. Dis. 2013, 13,
672–679.
Le Hello S., Hendriksen R.S., Doublet B., Fisher I., Nielsen E.M., Whichard
J.M., Bouchrif B., Fashae K., Granier S.A., Jourdan-Da Silva N., Cloeckaert
A., Threlfall E.J., Angulo F.J., Aarestrup F.M., Wain J., Weill F.X.:
International spread of an epidemic population of Salmonella enterica
serotype Kentucky ST198 resistant to ciprofloxacin. J. Infect. Dis. 2011, 204,
675-684.
Lundin J.I., Dargatz D.A., Wagner B.A., Lombard J. E., Hill A.E., Ladely S.
R., Fedorka-Cray P.J.: Antimicrobial drug resistance of fecal Escherichia coli
and Salmonella spp. isolates from United States dairy cows. Foodborne
Pathog. Dis. 2008, 5, 7-19.

233
MARAN. Monitoring of antimicrobial resistance and antibiotic usage
in animals in the Netherlands. Central Veterinary Institute, Agricultural
Economics Research Institute, Wageningen, 2012.
Morfin-Otero R., Tinoco-Favila J.C., Sader H.S., Salcido-Gutierrez L.,
Perez-Gomez H.R., Gonzalez-Diaz E., Petersen L., Rodriguez-Noriega E.:
Resistance trends in gram-negative bacteria: surveillance results from two
Mexican hospitals, 2005-2010. BMC Res. Notes. 2012, 5, 277.
Ozaki H., Esaki H., Takemoto K., Ikeda A., Nakatani Y., Someya A.,
Hirayama N., Murase T.: Antimicrobial resistance in fecal Escherichia coli
isolated from growing chickens on commercial broiler farms. Vet. Microbiol.
2011, 150, 132-139.
Perchec-Merien A.M., Lewis G.D.: Naturalized Escherichia coli from New
Zealand wetland and stream environments. FEMS Microbiol. Ecol. 2013, 83,
494-503.
Pitout J.D.: Extraintestinal pathogenic Escherichia coli: a combination of
virulence with antibiotic resistance. Front. Microbiol. 2012, 3, 9.
PZH: Choroby Zakaźne i zatrucia w Polsce w 2011 r. Państwowy Zakład
Higieny, Główny Inspektor Sanitarny, Warszawa, 2012.
Ribot E.M., Fair M.A., Gautom R., Cameron D.N., Hunter S.B.,
Swaminathan B., Barrett T.J.: Standardization of pulsed-field gel
electrophoresis protocols for the subtyping of Escherichia coli O157:H7,
Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3,
59-67.
Schroeter A., Tenhagen B.A., Alt K., Fetsch A., Stingl K., Guerra B.,
Heckenbach K., Helmuth R., Beutlich J., Hensel A., Appel B., Käsbohrer A:

234
German antimicrobial resistance situation in the food chain – DARLink 2009.
BfR Wissenschaft, Berlin, 2013.
Schwarz S., Silley P., Simjee S., Woodford N., van Duijkeren E., Johnson
A.P., Gaastra W.: Assessing the antimicrobial susceptibility of bacteria
obtained from animals. Vet. Microbiol, 2010, 141, 1-4.
Simon C., Daniel R.: Metagenomic analyses: past and future trends. Appl.
Environ. Microbiol. 2011, 77, 1153-1161.
SVARM: Swedish veterinary antimicrobial resistance monitoring. Uppsala,
Sweden: The National Veterinary Institute, Uppsala, 2012.
Tadesse D.A., Zhao S., Tong E., Ayers S., Singh A., Bartholomew M.J.,
McDermott P.F.: Antimicrobial drug resistance in Escherichia coli from
humans and food animals, United States, 1950-2002. Emerg. Infect. Dis.
2012, 18, 741-749
van der Bij A.K., van Dijk K., Muilwijk J., Thijsen S.F., Notermans D.W.,
de Greeff S., van de Sande-Bruinsma N.: Clinical breakpoint changes and
their impact on surveillance of antimicrobial resistance in Escherichia coli
causing bacteraemia. Clin. Microbiol. Infect. 2012, 18, E466-472.
Veldman K., Cavaco L.M., Mevius D., Battisti A., Franco A., Botteldoorn
N., Bruneau M., Perrin-Guyomard A., Cerny T., de Frutos Escobar C.,
Guerra B., Schroeter A., Gutierrez M., Hopkins K., Myllyniemi A.L., Sunde
M., Wasyl D., Aarestrup F.M.: International collaborative study on the
occurrence of plasmid-mediated quinolone resistance in Salmonella enterica
and Escherichia coli isolated from animals, humans, food and the
environment in 13 European countries. J. Antimicrob. Chemother. 2011, 66,
1278-1286.

235
Wasyl D.: Mechanizmy oporności Salmonella na chinolony. Medycyna Wet.
2009, 65, 516-520.
Wasyl D., Domańska-Blicharz K., Hoszowski A.: Quinolone resistance
mechanisms in Salmonella Kentucky with high-level ciprofloxacin
resistance. Proceedings of 3rd ASM Conference on Antimicrobial Resistance
in Zoonotic Bacteria and Foodborne Pathogens in Animals, Humans and the
Environment. Aix-en-Provence, Francja, 26-29 June, 2012.
Wasyl D., Hasman H., Cavaco L.M., Aarestrup F.M.: Prevalence and
characterization of cephalosporin resistance in nonpathogenic Escherichia
coli from food-producing animals slaughtered in Poland. Microb. Drug.
Resist. 2012, 18, 79-82.
Wasyl D., Hoszowski A.: First isolation of ESBL-producing Salmonella and
emergence of multiresistant Salmonella Kentucky in turkey in Poland. Food.
Res. Int. 2012, 45, 958–961.
Wasyl D., Hoszowski A.: Occurrence and characterization of monophasic
Salmonella enterica serovar Typhimurium (1,4,[5],12:i:-) of non-human
origin in Poland. Foodborne Pathog. Dis. 2012, 9, 1037-1043.
Wasyl D., Hoszowski A., Domańska-Blicharz K.: Salmonella Kentucky
epidemics in turkey, Poland, 2009 – 2011. In 9th international Symposium on
turkey Diseases. H. M. Hafez, ed. World Veterinary Poultry Association,
Institute of Poultry Diseases, Free University Berlin, Berlin. 2012, pp.
128-134.
Wasyl D., Hoszowski A., Zając M., Szulowski K.: Antimicrobial resistance
in commensal Escherichia coli isolated from animals at slaughter. Front.
Microbiol. 2013, 4, 221.

236
Wasyl D., Sandvang D., Skov M.N., Baggesen D.L.: Epidemiological
characteristics of Salmonella Typhimurium isolated from animals and feed
in Poland. Epidemiol. Infect. 2006, 134, 179-185.
Wasyl D., Zając M., Domanska-Blicharz K., Hoszowski A.: “Ad hoc” study
on Salmonella Stanley outbreak strains. Proceedings of International
Symphosium on Salmonella and Salmonellosis. Saint Malo, Francja, 27-29
May, 2013.
Wasyl D., Zając M., Hoszowski A.: Antibiotic resistance trends
in Salmonella, 2008-2012. Proceedings of International Symphosium
on Salmonella and Salmonellosis. Saint Malo, Francja, 27-29 May, 2013.
Wasyl D., Zając M., Hoszowski A.: Prevalence and characterisation
of quinolone resistance mechanisms in Salmonella isolated in Poland.
Proceedings of International Symphosium on Salmonella and Salmonellosis.
Saint Malo, Francja, 27-29 May, 2013.
Zając M., Wasyl D., Hoszowski A., Le Hello S., Szulowski K.: Genetic
lineages of Salmonella enterica serovar Kentucky spreading in pet reptiles.
Vet. Microbiol. 2013, 166, 686–689.
Zając M., Wasyl D., Hoszowski A., Szulowski K.: Genotypic diversity
of Salmonella Kentucky isolated from reptiles, poultry food, feed and
environmental samples in Poland. Proceedings of International Symphosium
on Salmonella and Salmonellosis. Saint Malo, Francja, 27-29 May, 2013.
Zając M., Wasyl D., Hoszowski A., Szulowski K.: Public healh relevant
Salmonella serovards in reptiles. Proceedings of Med-Vet-Net Association
Internarnational Scientific Conference. Kgs. Lingby, Dania, 24-25 June,
2013.
View publication statsView publication stats