protein structure and function. chapter2. from structure to function
TRANSCRIPT
Protein Structure and Function

CHAPTER2. From Structure to Function

There are many levels of protein function Biochemist: biochemical role of an individual protein Cell biologist, geneticist: cellular roles judged by the
phenotype of its deletion or signal pathway Physiologist, developmental biologist: cellular or
organism level
Protein can have many functions on different levels ex) tubulin
4 fundamental biochemical functions:Binding, catalysis, switching and structural elements
2-0. Overview : The Structural Basis of Protein Function
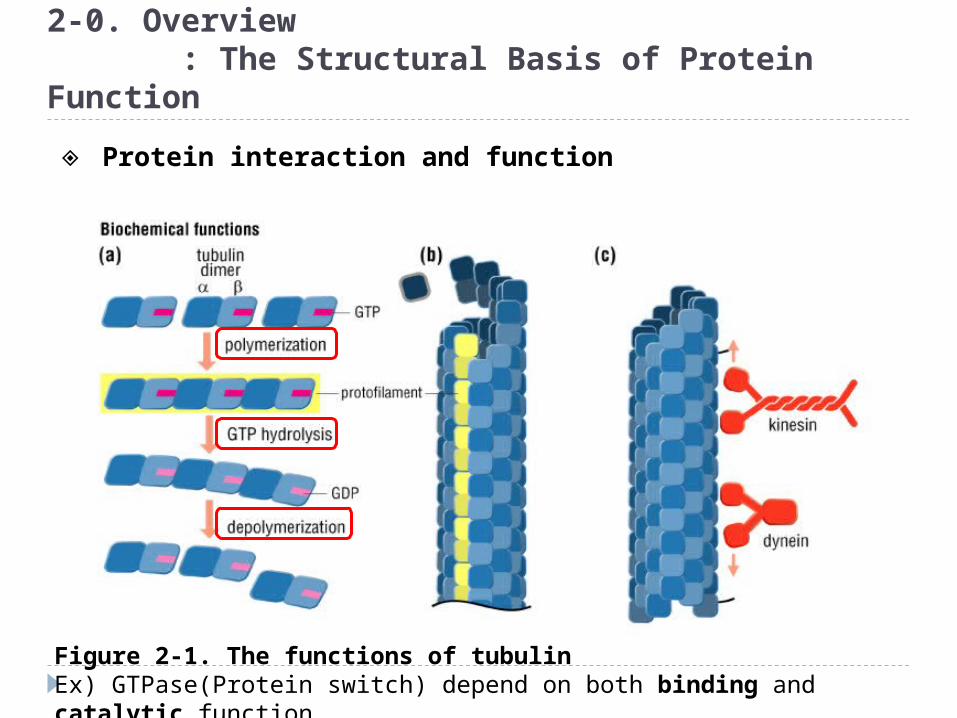
Figure 2-1. The functions of tubulinEx) GTPase(Protein switch) depend on both binding and catalytic function.
◈ Protein interaction and function

2-0. Overview : The Structural Basis of Protein Function
Figure 2-1. The functions of tubulin
Assemblies of microtubules, dynein and other microtubule-associated proteins form flagella that propel sperm
Microtubules and associated motor proteins form a network of “tracks” on which vesicles are moved around in cells. Taxol anti-cancer drug
Radial spoke

TAXOL Paclitaxel 은 Taxus brevifolia Nutt. (Taxaceae)( 미국 주목나무 -Western Yew) 의
주피 (bark) 에서 추출한 diterpenoid taxane 유도체로 천연 taxane ring 을 가진 alkaloid 이다 . (1) 이 taxane ring 과 ester side chain 이 cytotoxicity 작용을 가지는 것으로 알려졌다 .(1) Paclitaxel 은 수용성이 극히 낮아 제제화하기 위해 polyoxyethylated castor oil 과 absolute ethanol 의 혼합물인 Cremorphor® EL 이라는 용제를 사용하고 있다 . (3) 따라서 임상적으로 이러한 용제가 과량 투여되므로 심장독성과 과민반응이 발생하는 것으로 보고되고 있다 . Taxol 와 tubulin complex 구조
Mechanism of ActionPaclitaxel 은 1992 년 12 월에 난치성 난소암의 치료제로 FDA 의 승인을 받은 항암제로 세포 내 microtubule 의 assembly 를 증진시키고 disassembly 를 저해함으로써 항암효과를 나타내는 독특한 약물이다 .(2) Microtubule 이란 세포질 (cytoplasm) 에 있는 가는 tube 같은 구조로써 세포의 골격 형성과 motility 를 유지하며 유사분열 중기에서 방추사의 형성과 분열시 이동에 관여한다 .(2) Microtubule 은 tubulin dimer 라는 고유의 subunit 로 이루어졌고 이 tubulin 이 assembly 되어 microtubule 을 형성한다 . .(2) 형성된 microtubule 은 유사분열시 기능수행을 위해서는 disassembly 되어야 한다 . .(2) Paclitaxel 은 암세포의 microtubule 의 assembly 를 증진 시키고 일단 형성된 microtubule 을 안정화시켜 polymerization 상태로 남아있게 한다 .(2) Fuchs 과 Johnson 의 초기 발견에 의하면 , microtubule 의 disassembly 를 저해하여 유사분열에 필요한 방추사의 형성을 억제하므로 세포 주기상 암 세포가 G2 기와 M 기에 머무르게 되어 cytotoxic effect 를 가진다 . 이는 vincristine, vinblastine, colchicines, podophyllotoxin, maytansine 과 같은 항암제와 같은 작용이나 ,(1) paclitaxel 은 다른 항암제와 달리 microtubule 을 stabilize시키고 , depolymerization 을 억제시킨다 .(2) 또한 이 G2 기와 M 기 는 방사선에 매우 민감한 주기이므로 radiation therapy 와 병용할 시 cytotoxicity 가 증가하게 된다 .
2-1. Recognition, Complementarity and Active Sites
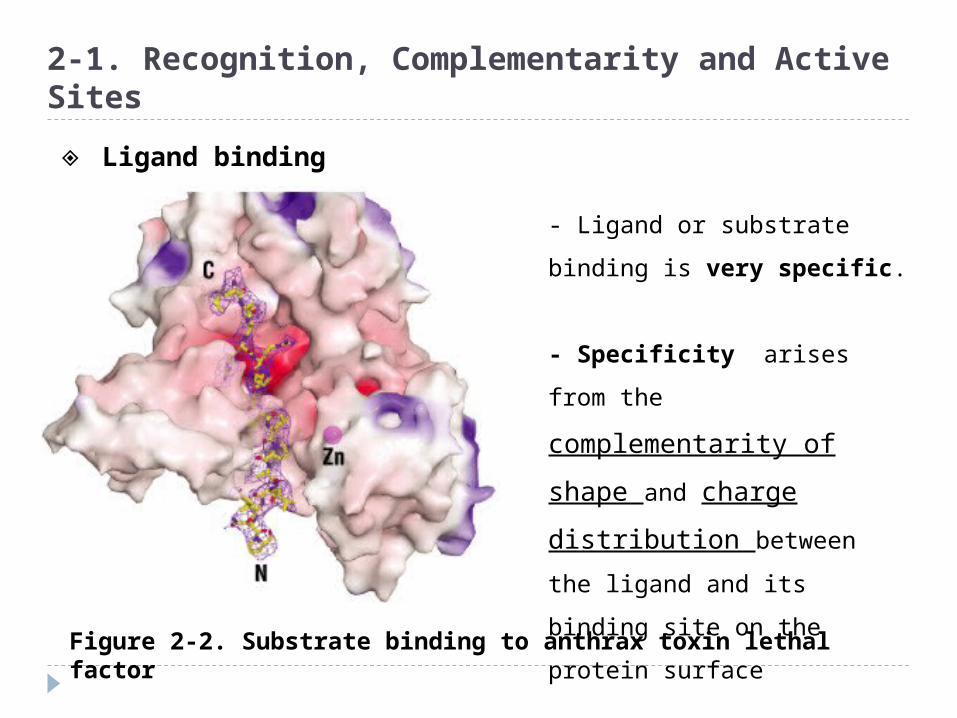
Figure 2-2. Substrate binding to anthrax toxin lethal factor
- Ligand or substrate binding
is very specific.
- Specificity arises from the
complementarity of shape
and charge distribution
between the ligand and its
binding site on the protein
surface
◈ Ligand binding
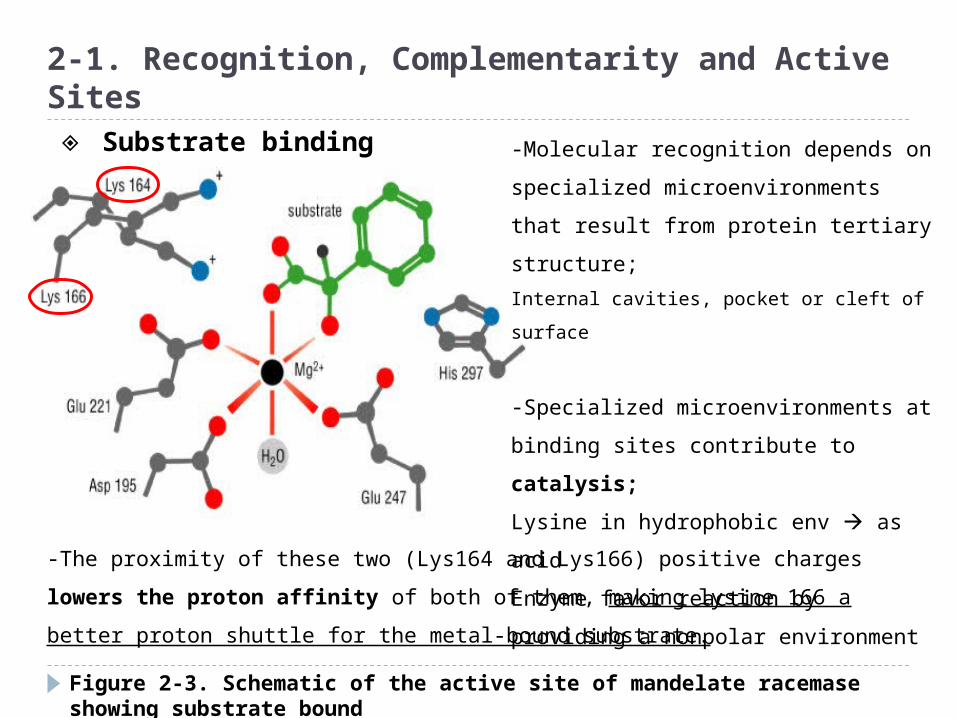
Figure 2-3. Schematic of the active site of mandelate racemase showing substrate bound
2-1. Recognition, Complementarity and Active Sites
-Molecular recognition depends on
specialized microenvironments that
result from protein tertiary structure;
Internal cavities, pocket or cleft of surface
-Specialized microenvironments at
binding sites contribute to catalysis;
Lysine in hydrophobic env as acid
Enzyme favor reaction by providing a
nonpolar environment
-The proximity of these two (Lys164 and Lys166) positive charges lowers the
proton affinity of both of them, making lysine 166 a better proton shuttle for the
metal-bound substrate.
◈ Substrate binding

Figure 2-4. Tight fit between a protein and its ligand
2-2. Flexibility and Protein Function
-The flexibility of tertiary structure
allows proteins to adapt to their
ligands;
Charge configuration, potential H-
bond complementarity
- Lack-and-key analogy :
implies rigidity of the protein(the
lock) and of the ligand(the key)
-Induced fit model : both protein
and the ligands are naturally flexible
◈ Ligand binding model

Figure 2-5. HIV protease, and enzyme from the virus that causes AIDS, bound to three different inhibitors
2-2. Flexibility and Protein Function
Haloperidol
Crixivan
Peptide analog of the natural substrate
Each inhibitor clearly has a quite different structure, yet all bind tightly to the active site(Because of protein structure flexibility)
◈ Protein structure flexibility
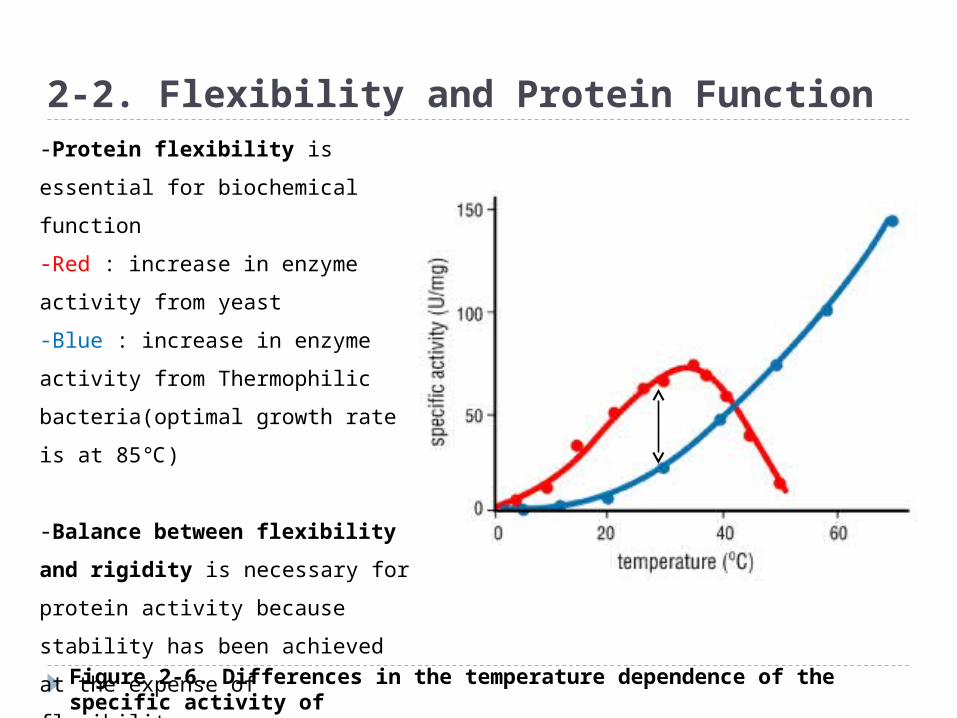
Figure 2-6. Differences in the temperature dependence of the specific activity of D-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) from two organisms
2-2. Flexibility and Protein Function-Protein flexibility is essential for
biochemical function
-Red : increase in enzyme activity
from yeast
-Blue : increase in enzyme activity
from Thermophilic bacteria(optimal
growth rate is at 85℃)
-Balance between flexibility
and rigidity is necessary for
protein activity because stability
has been achieved at the expense
of flexibility.
-Genetic engineering in
Enzyme?

Figure 2-7. Example of a large conformational change in adenylate kinase
2-2. Flexibility and Protein Function
Some proteins undergo very large shape changes when the correct ligand binds
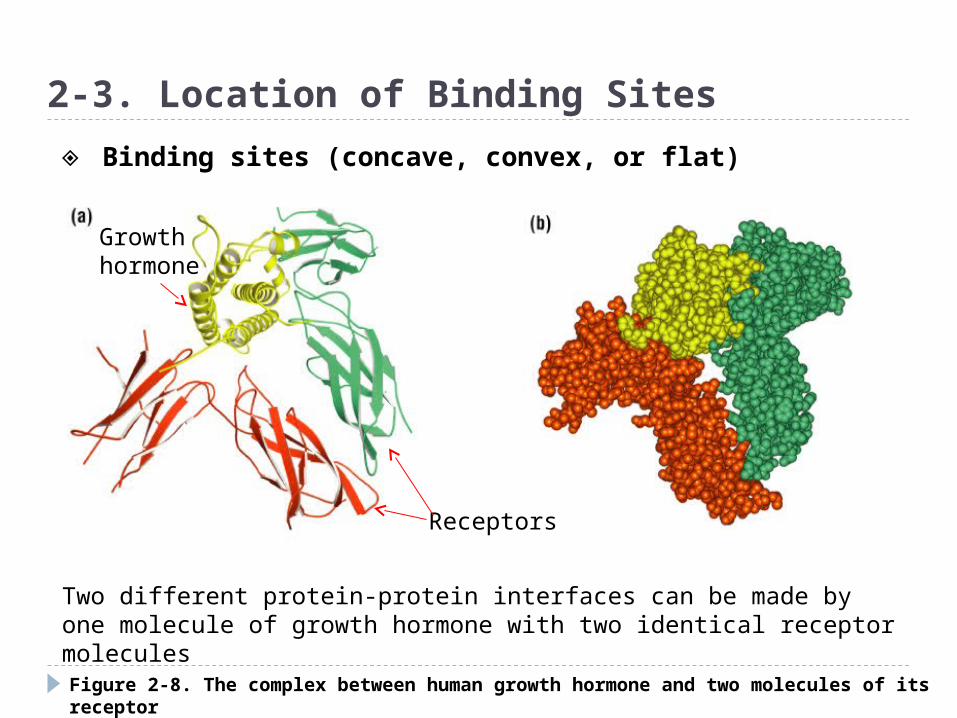
Figure 2-8. The complex between human growth hormone and two molecules of its receptor
2-3. Location of Binding Sites
Two different protein-protein interfaces can be made by one molecule of growth hormone with two identical receptor molecules
Growth hormone
Receptors
◈ Binding sites (concave, convex, or flat)

Figure 2-9. Two protein-DNA complexes
2-3. Location of Binding Sites
Many binding sites for RNA or DNA on proteins are protruding loops or alpha helices that fit into the major and the minor grooves of the nucleic acid
Zinc finger
Helix-turn-helix motif
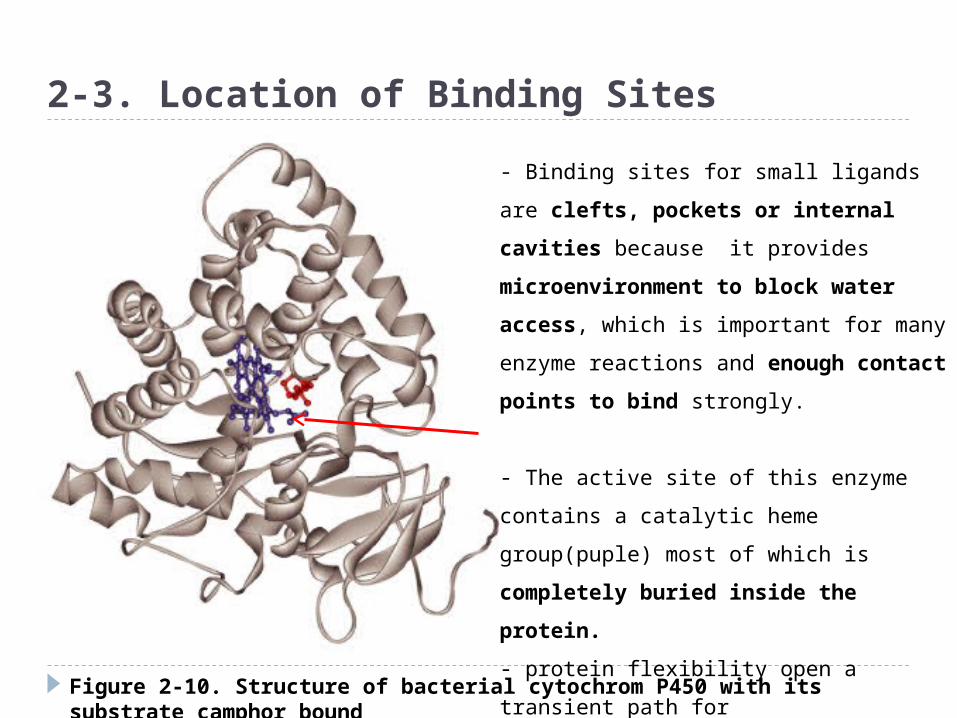
Figure 2-10. Structure of bacterial cytochrom P450 with its substrate camphor bound
2-3. Location of Binding Sites
- Binding sites for small ligands are clefts,
pockets or internal cavities because it
provides microenvironment to block
water access, which is important for
many enzyme reactions and enough
contact points to bind strongly.
- The active site of this enzyme contains a
catalytic heme group(puple) most of which
is completely buried inside the
protein.
- protein flexibility open a transient path
for
Penetration of the substrate
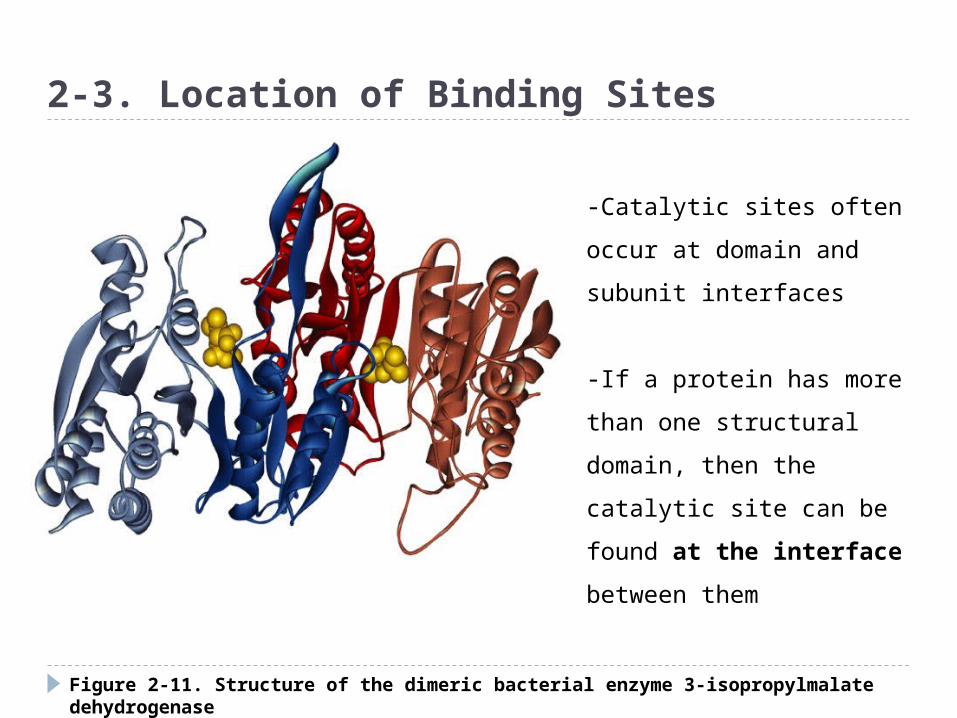
Figure 2-11. Structure of the dimeric bacterial enzyme 3-isopropylmalate dehydrogenase
2-3. Location of Binding Sites
-Catalytic sites often occur at
domain and subunit interfaces
-If a protein has more than
one structural domain, then
the catalytic site can be found
at the interface between
them

Figure 2-12. Surface view of the heme-binding pocket of cytochrome c6, with hydrophobic residues indicated in yellow
2-4. Nature of Binding Sites
-Binding sites generally have a
higher than average amount of
exposed hydrophobic surface
◈ Binding site characteristic
Heme
Large hydrophobic areas on the surface of a protein lead to self-associationAnd oligomerization. But binding sites for small molecules are usually too small and concave to allow the protein to oligomerize.
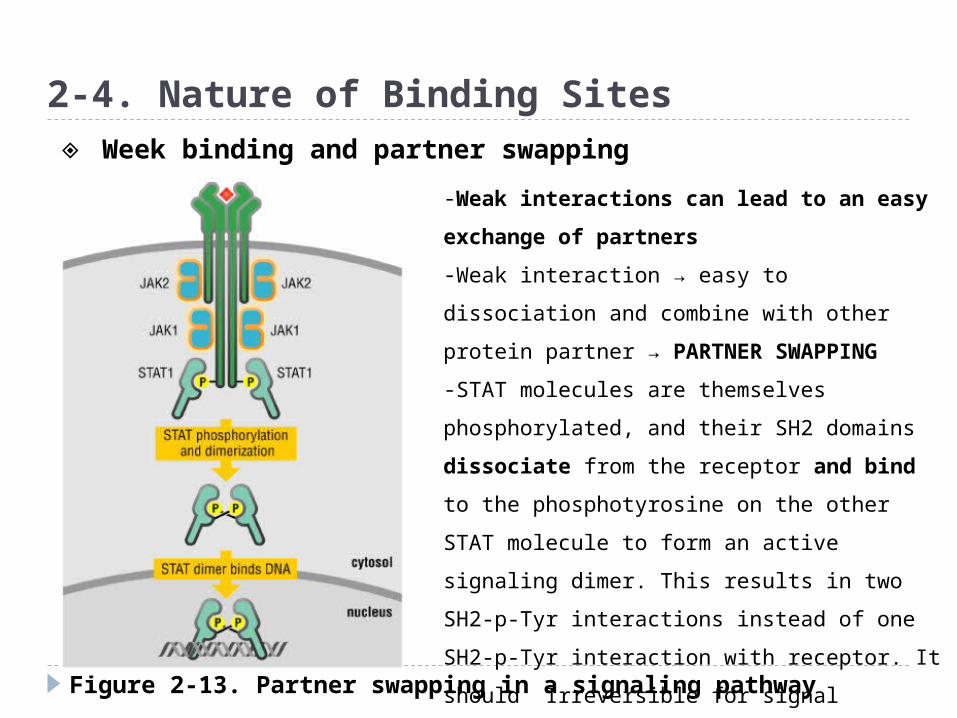
Figure 2-13. Partner swapping in a signaling pathway
2-4. Nature of Binding Sites
-Weak interactions can lead to an easy
exchange of partners
-Weak interaction → easy to dissociation and
combine with other protein partner →
PARTNER SWAPPING
-STAT molecules are themselves
phosphorylated, and their SH2 domains
dissociate from the receptor and bind to the
phosphotyrosine on the other STAT molecule
to form an active signaling dimer. This results
in two SH2-p-Tyr interactions instead of one
SH2-p-Tyr interaction with receptor. It should
Irreversible for signal transduction
◈ Week binding and partner swapping
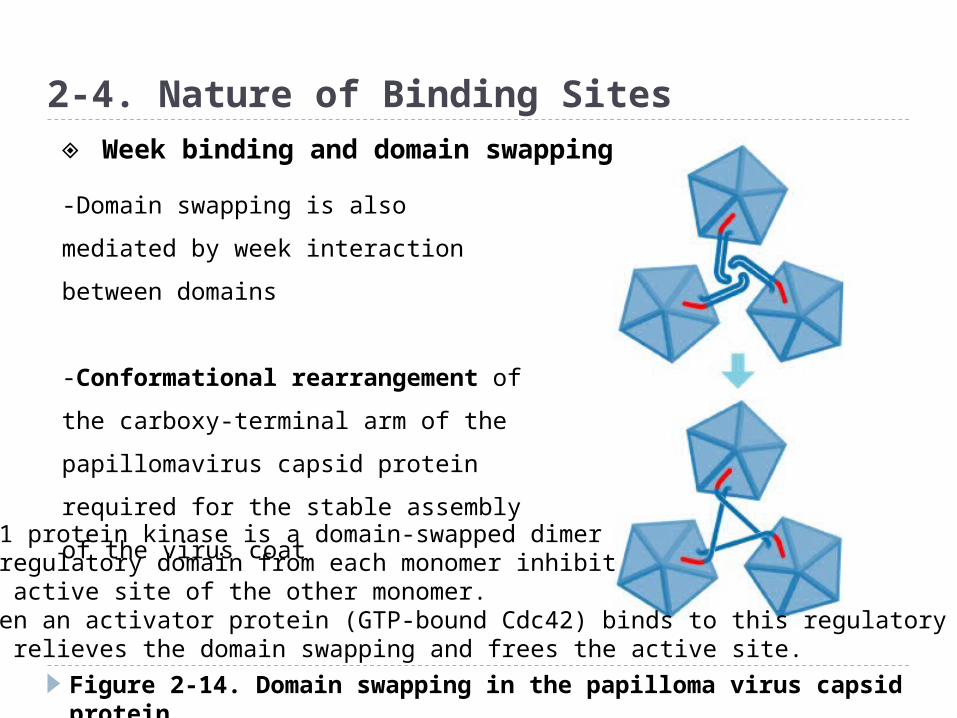
Figure 2-14. Domain swapping in the papilloma virus capsid protein
2-4. Nature of Binding Sites
-Domain swapping is also mediated by
week interaction between domains
-Conformational rearrangement of
the carboxy-terminal arm of the
papillomavirus capsid protein required
for the stable assembly of the virus
coat
◈ Week binding and domain swapping
PAK1 protein kinase is a domain-swapped dimer-A regulatory domain from each monomer inhibitthe active site of the other monomer.-When an activator protein (GTP-bound Cdc42) binds to this regulatory domain-It relieves the domain swapping and frees the active site.

Displacement of water also drives binding events for binding between ligand and protein, water
layer (hydrogen shell) must be disrupted or partially displaced.
Thermodynamics? Enthalphy? Between Waters and protein-
waters, in hydrophobic environment How about entrophy?
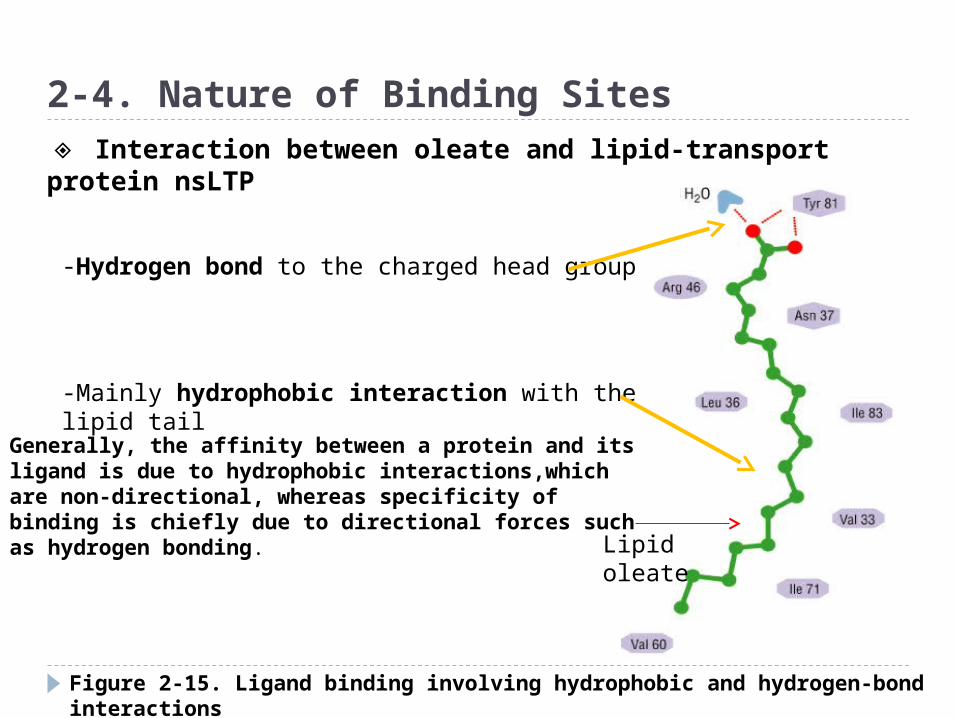
Figure 2-15. Ligand binding involving hydrophobic and hydrogen-bond interactions
2-4. Nature of Binding Sites
Lipid oleate
-Hydrogen bond to the charged head group
-Mainly hydrophobic interaction with the lipid tail
◈ Interaction between oleate and lipid-transport protein nsLTP
Generally, the affinity between a protein and its ligand is due to hydrophobic interactions,which are non-directional, whereas specificity of binding is chiefly due to directional forces such as hydrogen bonding.

Figure 2-16. Structure of the 50S (large) subunit of the bacterial ribosome
2-5. Functional Properties of Structural Proteins
-In some cases, structural
proteins are assisted by DNA
and RNA, lipid and carbohydrate
molecules.
-Ex). Ribosome: has over a
hundred different protein
components(for stabilization,
catalytic function)
-Dynamics in proteins;
muscle, actin filaments,
fibrinogen
Blue : protein Red & grey : RNA
◈ Proteins as frameworks, connectors and scaffolds

Figure 2-17. Structure of collagen
2-5. Functional Properties of Structural Proteins
Permanent structural proteins
-Silk, collagen, elastin, or keratin
-Collagen : fibrous component of tendons.
Collagen is a triple helix of three protein
chain made up repeating GlyXY
sequence(where X is often proline).
- Hydrophobic interactions, H-bond and cross-
linking of lysine residues (peptidyl aldehydes by
lysyl oxidase)
◈ Some structural proteins only form stable assemblies

Figure 2-18. The Ste5p scaffold in MAPK cascade
2-5. Functional Properties of Structural Proteins
-In the signal transduction,
specific protein recruitment is
required.
-Sometimes this recruitment
occurs by localizing components
to the cell membrane.
-But the other cases specific
structural proteins serve as
scaffold proteins
◈ Some structural proteins serve as scaffolds
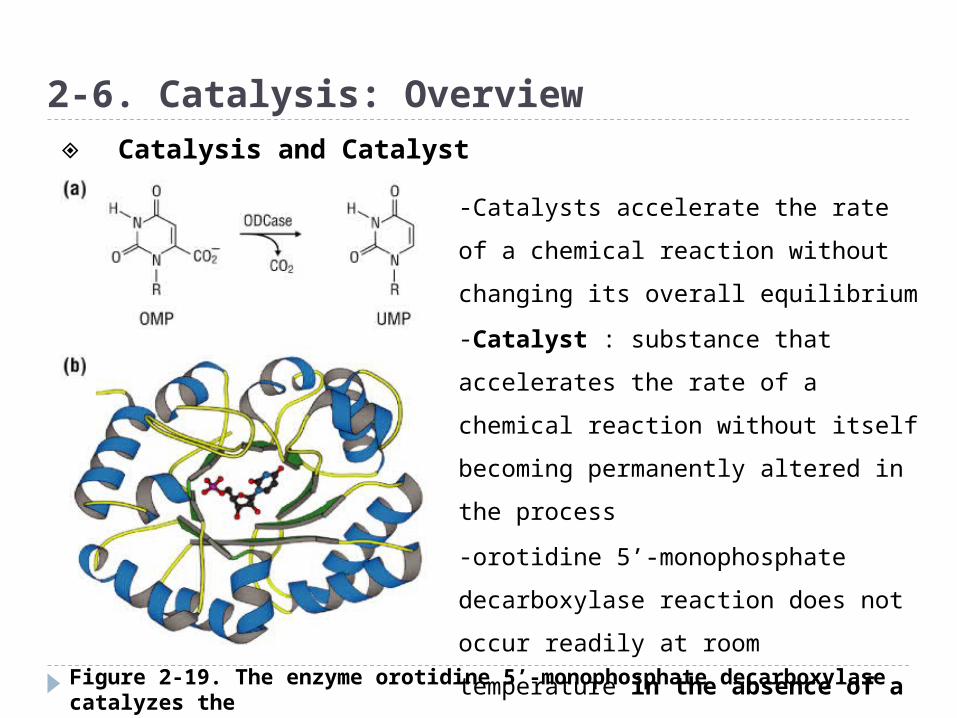
Figure 2-19. The enzyme orotidine 5’-monophosphate decarboxylase catalyzes the transformation of orotidine 5’-monophosphate to uridine 5’-monophosphate
2-6. Catalysis: Overview
-Catalysts accelerate the rate of a
chemical reaction without changing
its overall equilibrium
-Catalyst : substance that
accelerates the rate of a chemical
reaction without itself becoming
permanently altered in the process
-orotidine 5’-monophosphate
decarboxylase reaction does not
occur readily at room temperature in
the absence of a catalyst.
◈ Catalysis and Catalyst
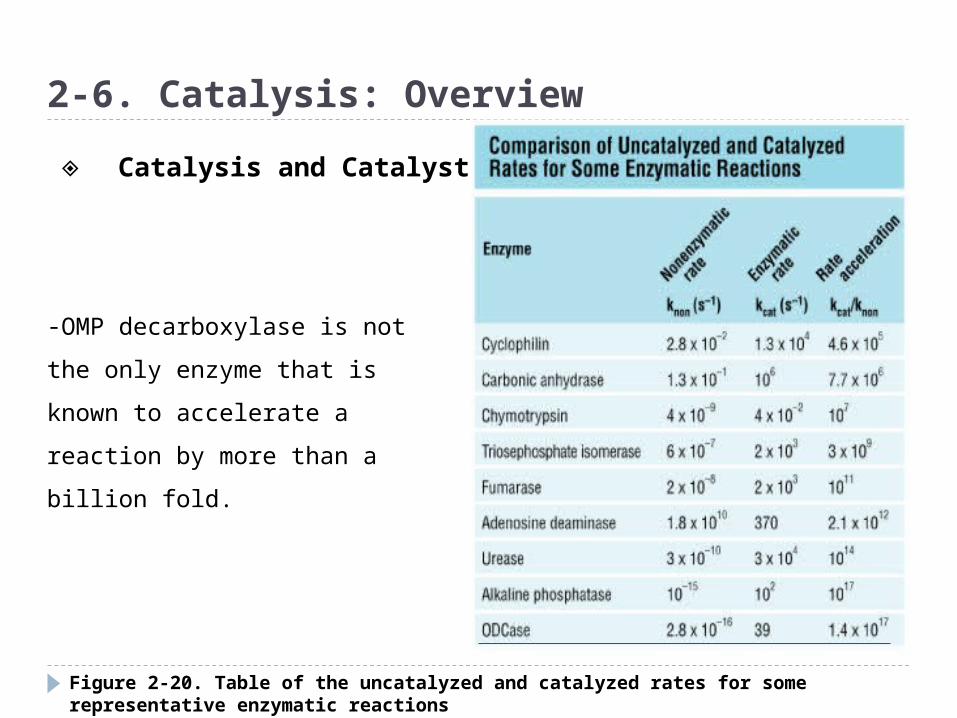
Figure 2-20. Table of the uncatalyzed and catalyzed rates for some representative enzymatic reactions
2-6. Catalysis: Overview
-OMP decarboxylase is not the
only enzyme that is known to
accelerate a reaction by more
than a billion fold.
◈ Catalysis and Catalyst
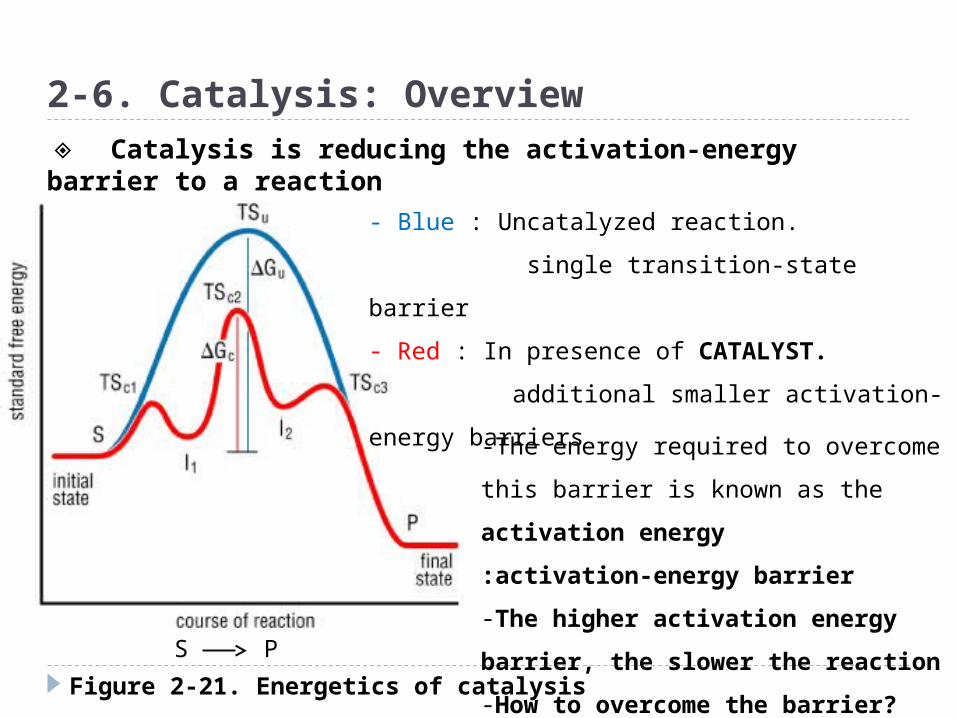
Figure 2-21. Energetics of catalysis
2-6. Catalysis: Overview
- Blue : Uncatalyzed reaction.
single transition-state barrier
- Red : In presence of CATALYST.
additional smaller activation-energy
barriers
S P
-The energy required to overcome this
barrier is known as the activation
energy
:activation-energy barrier
-The higher activation energy
barrier, the slower the reaction
-How to overcome the barrier?
◈ Catalysis is reducing the activation-energy barrier to a reaction

Figure 2-23. Schematic diagram showing some of the ways in which electrostatic interactions can influence the binding of a ligand to a protein
2-7. Active-Site Geometry
Negative substrate
Positive potential
•Reactive groups in enzyme active sites are optimally positioned to interact
With the substrate-primarily by van der Waals interactions and complementary H-bonds and electrostatic interactions
-Too tight substrate binding reduces efficiency as a catalyst. Why?
-Specific complex is necessary for productive collision; correct orientation
Which induces atomic orbitals can overlap to allow the appropriate bonds to be formed or broken.
-enzyme offer the time and place for second substrate to bind to enzyme.
Gated binding
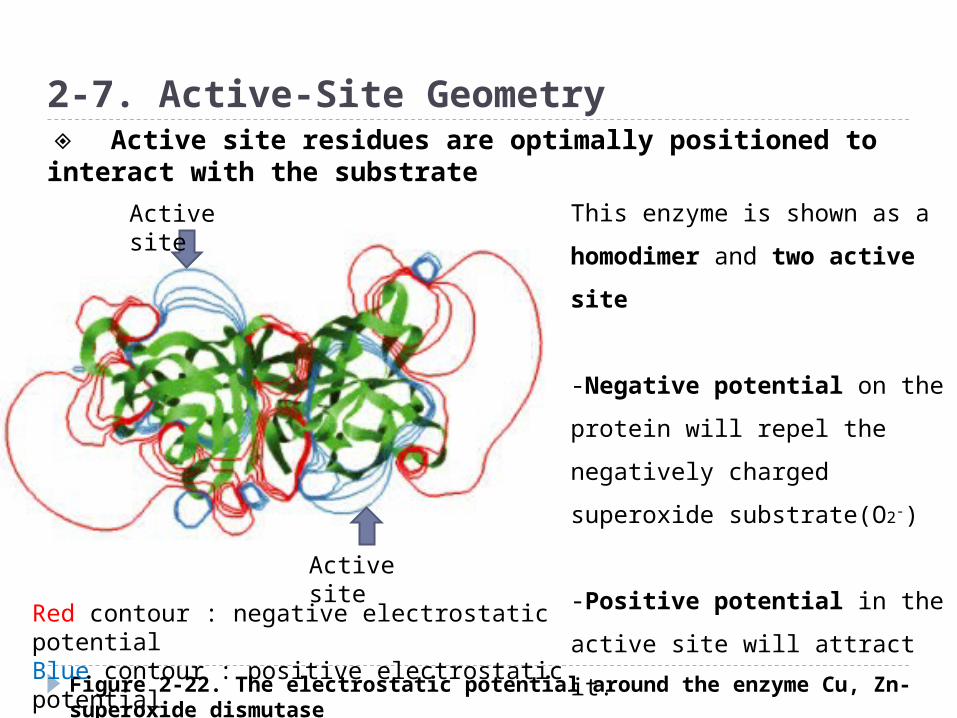
Figure 2-22. The electrostatic potential around the enzyme Cu, Zn-superoxide dismutase
2-7. Active-Site Geometry
Active site
Active site
Red contour : negative electrostatic potentialBlue contour : positive electrostatic potential
This enzyme is shown as a
homodimer and two active
site
-Negative potential on the
protein will repel the
negatively charged superoxide
substrate(O2-)
-Positive potential in the
active site will attract it.
◈ Active site residues are optimally positioned to interact with the substrate
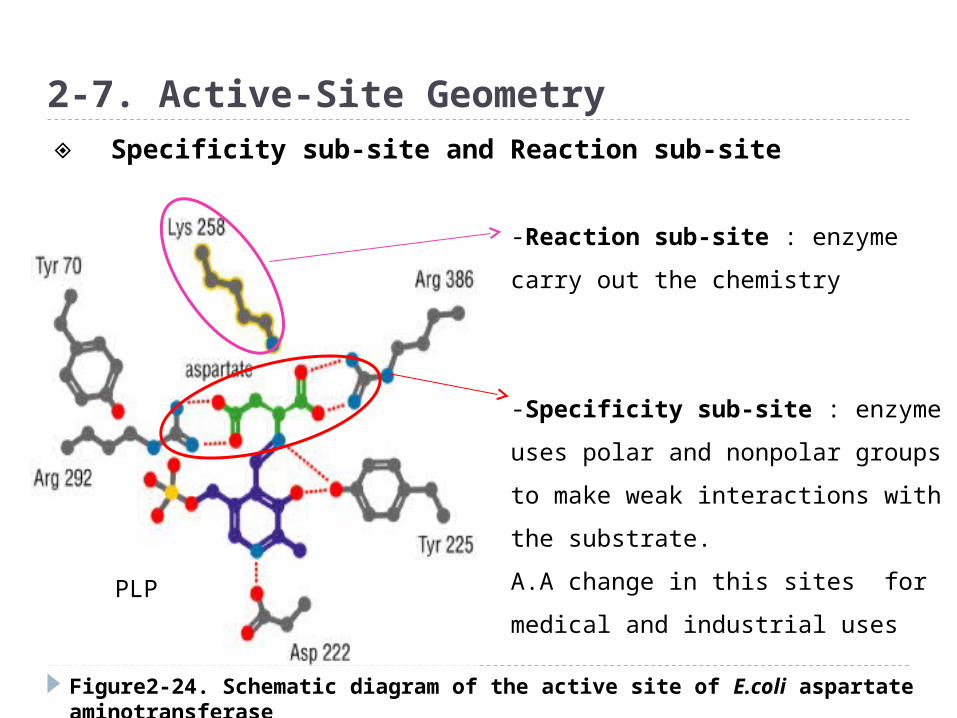
Figure2-24. Schematic diagram of the active site of E.coli aspartate aminotransferase
2-7. Active-Site Geometry
-Reaction sub-site : enzyme carry
out the chemistry
-Specificity sub-site : enzyme
uses polar and nonpolar groups to
make weak interactions with the
substrate.
A.A change in this sites for medical
and industrial uses
◈ Specificity sub-site and Reaction sub-site
PLP
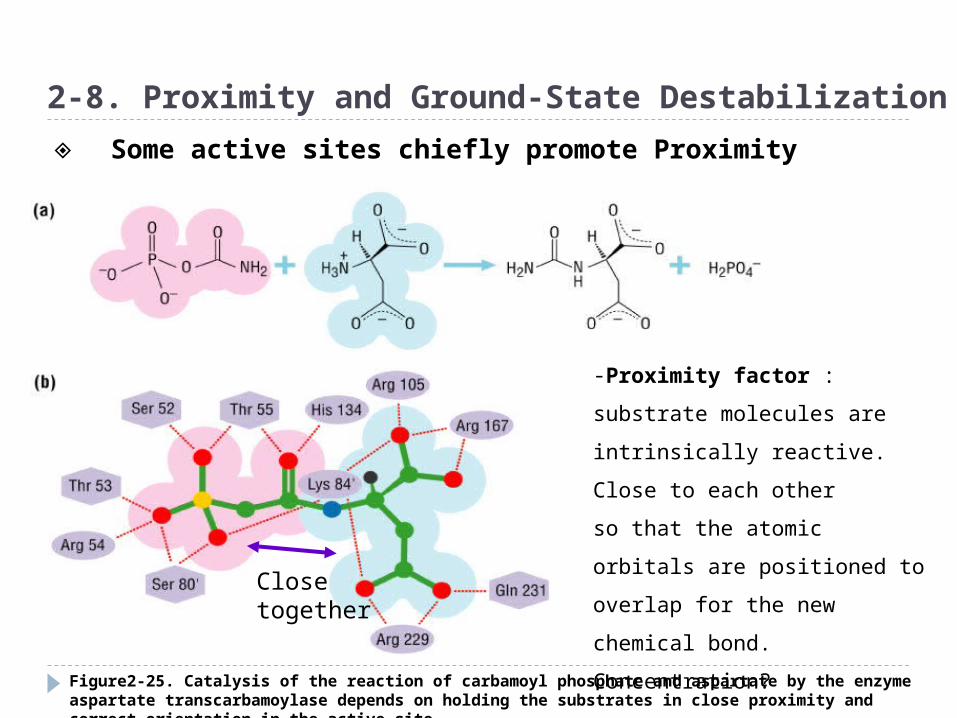
Figure2-25. Catalysis of the reaction of carbamoyl phosphate and aspartate by the enzyme aspartate transcarbamoylase depends on holding the substrates in close proximity and correct orientation in the active site
2-8. Proximity and Ground-State Destabilization
-Proximity factor : substrate
molecules are intrinsically
reactive. Close to each other
so that the atomic orbitals are
positioned to overlap for the
new chemical bond.
Concentration?Close together
◈ Some active sites chiefly promote Proximity
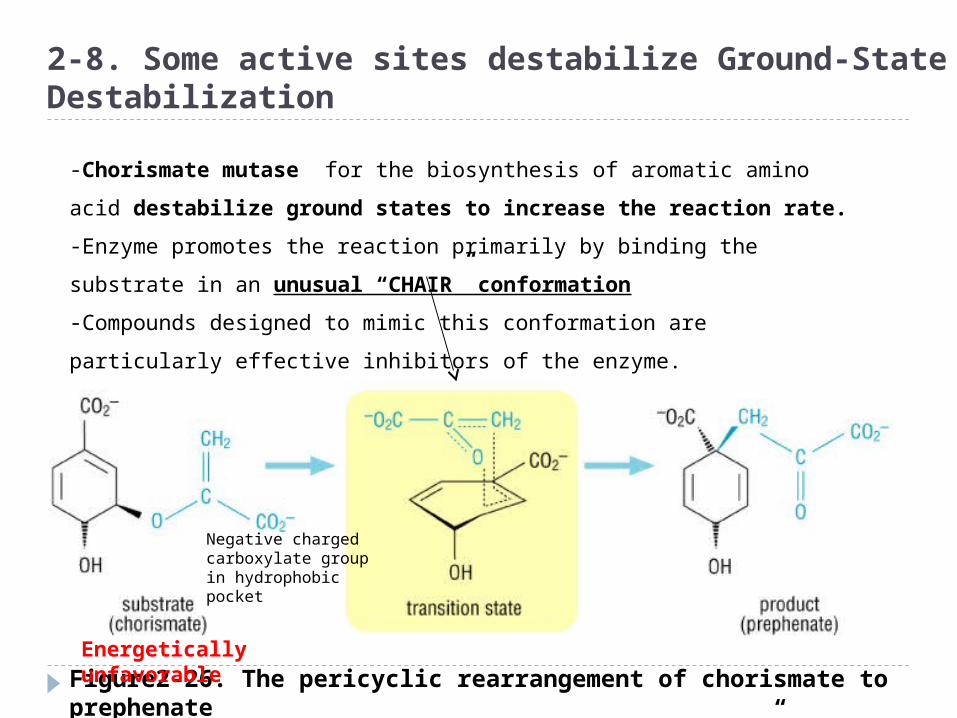
Figure2-26. The pericyclic rearrangement of chorismate to prephenate via the proposed “chair-like” transition state
2-8. Some active sites destabilize Ground-State Destabilization
-Chorismate mutase for the biosynthesis of aromatic amino acid
destabilize ground states to increase the reaction rate.
-Enzyme promotes the reaction primarily by binding the substrate in an
unusual “CHAIR” conformation
-Compounds designed to mimic this conformation are particularly
effective inhibitors of the enzyme.
Energetically unfavorable
Negative charged carboxylate group in hydrophobic pocket

Figure2-27. Effect of binding energy on enzyme catalysis
2-9. Stabilization of Transition States and Exclusion of Water
Substrate and product are of equal energy, with a large free-energy barrier between them.
ES : enzyme and substrate complexEP : enzyme and product complex
◈ Binding energyIf the transition state can be bound more tightlythan the substrate, activation energy will be reducedThe differential binding of enzyme for these two stateIs the driving force of reactions
a = new interactions with transition stateb = stabilization

Many active sites must protect their substrates from water, but must be accessible at the same time
When an active site is in its closed conformation, it is protected from water. But substrates, products?
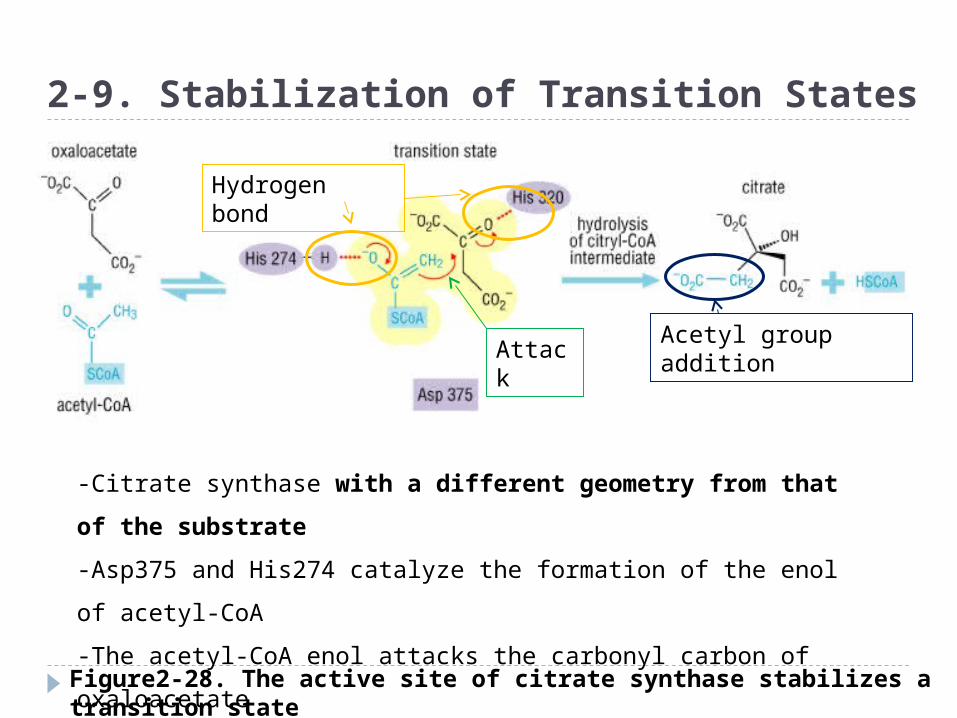
Figure2-28. The active site of citrate synthase stabilizes a transition state with a different geometry from that of the substrate
2-9. Stabilization of Transition States
-Citrate synthase with a different geometry from that of the
substrate
-Asp375 and His274 catalyze the formation of the enol of acetyl-
CoA
-The acetyl-CoA enol attacks the carbonyl carbon of oxaloacetate
-Addition of the elements of the acetyl group at this portion
Hydrogen bond
AttackAcetyl group addition

Figure2-29. Phosphoglycerate kinase (PGK) undergoes a conformational change in its active site after substrate binds
2-9. Stabilization of Transition States and Exclusion of Water
- If an enzyme active site dies not start out perfectly complementary to
the transition state, the enzyme undergo conformational changes
that increase that complementarity
Unliganded PGK Complex with its substrate
◈ Binding and conformational changes

Figure2-30. NAD-dependent lactate dehydrogenase has a mechanism for excluding water from the active site once substrates are bound
2-9. Stabilization of Transition States and Exclusion of Water
- Many active sites must protect their substrates from water,
but must be accessible at the same time
-Resting enzyme exist in an open state to which substrates can bind
readily
-Substrate binding trigger the conformational changes to the closed
form
-Opening and/or closing of the lid is the rate-determining step in the
reaction
Hydride ions (H-) are unstable in water, so the active site must be shield from bulk solvent. Why?

2-10. Redox Reactions
A mammalian cell produces over 10,000 different
proteins→ more than half are enzymes
→ thousands of different enzymes with thousands of different substrates
and products
→ 4 similar reaction
→ Oxidation/reduction
→ addition/elimination
→ hydrolysis
→ decarboxylatoin
◈ Enzyme chemical reactions
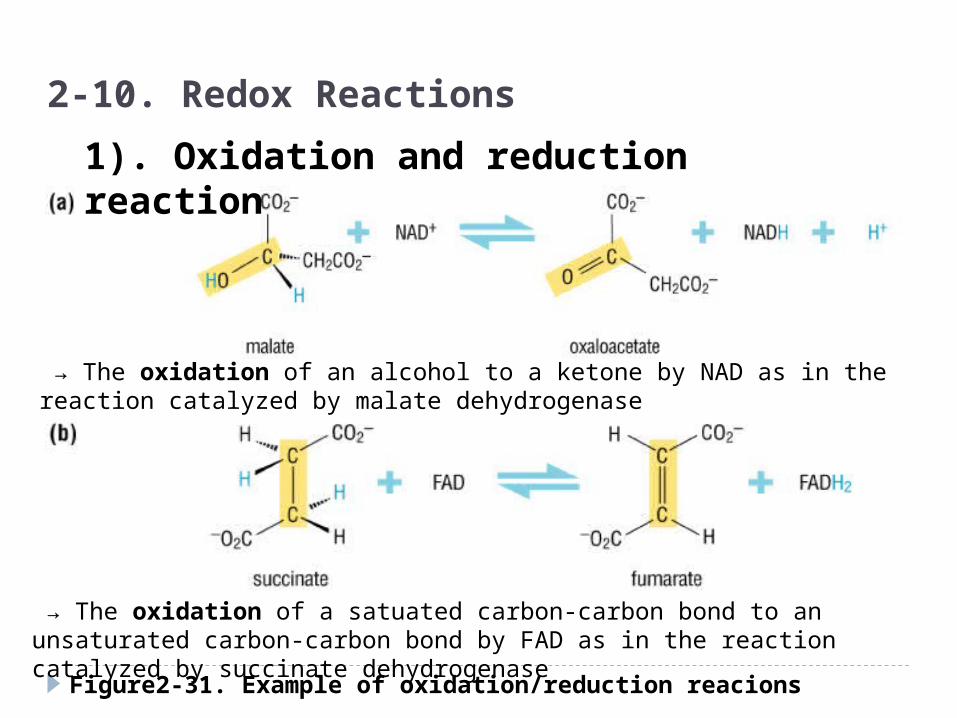
Figure2-31. Example of oxidation/reduction reacions
2-10. Redox Reactions
1). Oxidation and reduction reaction
→ The oxidation of an alcohol to a ketone by NAD as in the reaction catalyzed by malate dehydrogenase
→ The oxidation of a satuated carbon-carbon bond to an unsaturated carbon-carbon bond by FAD as in the reaction catalyzed by succinate dehydrogenase

Figure2-31. Example of oxidation/reduction reacions
2-10. Redox Reactions
1). Oxidation and reduction reaction
→ In the first step of the pathway for the conversion of cholesterol to pregnenolone

Figure2-32. Examples of addition/elimination reactions
2-11. Addition/Elimination, Hydrolysis and Decarboxylation
2). Addition and elimination reaction
→ Addition of water across the C=C of fumarate to create the HC-COH group of malate, a reaction catalyzed by fumarase. Only L-malate
→ Addition of acetate to the carbonyl carbon of oxaloacetate in the aldol condensation reaction catalyzed by citrate synthase
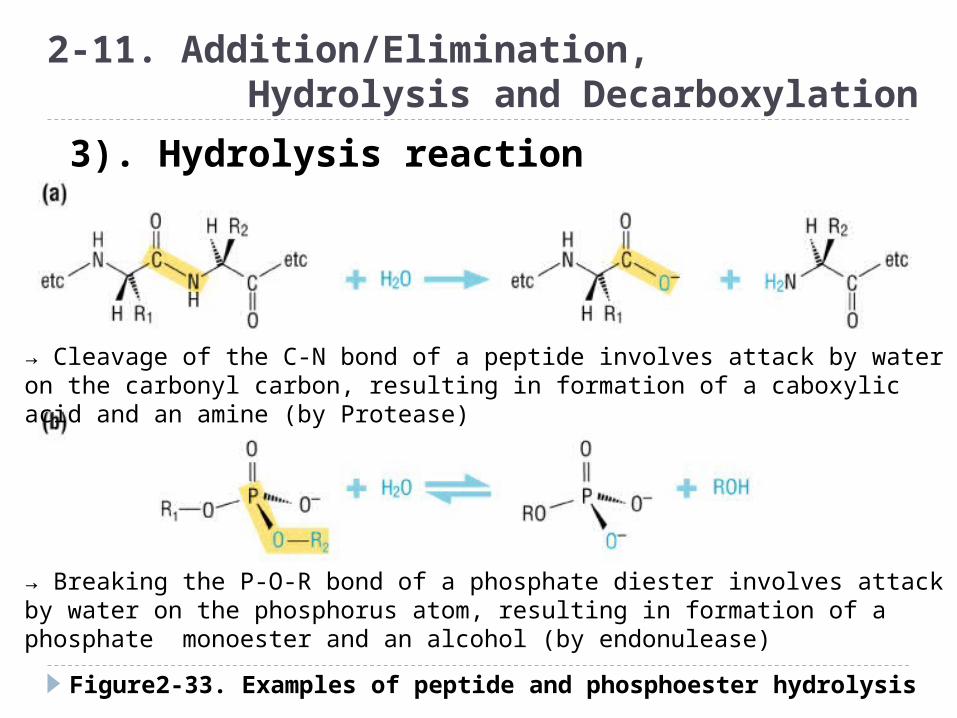
Figure2-33. Examples of peptide and phosphoester hydrolysis
2-11. Addition/Elimination, Hydrolysis and Decarboxylation
3). Hydrolysis reaction
→ Cleavage of the C-N bond of a peptide involves attack by water on the carbonyl carbon, resulting in formation of a caboxylic acid and an amine (by Protease)
→ Breaking the P-O-R bond of a phosphate diester involves attack by water on the phosphorus atom, resulting in formation of a phosphate monoester and an alcohol (by endonulease)
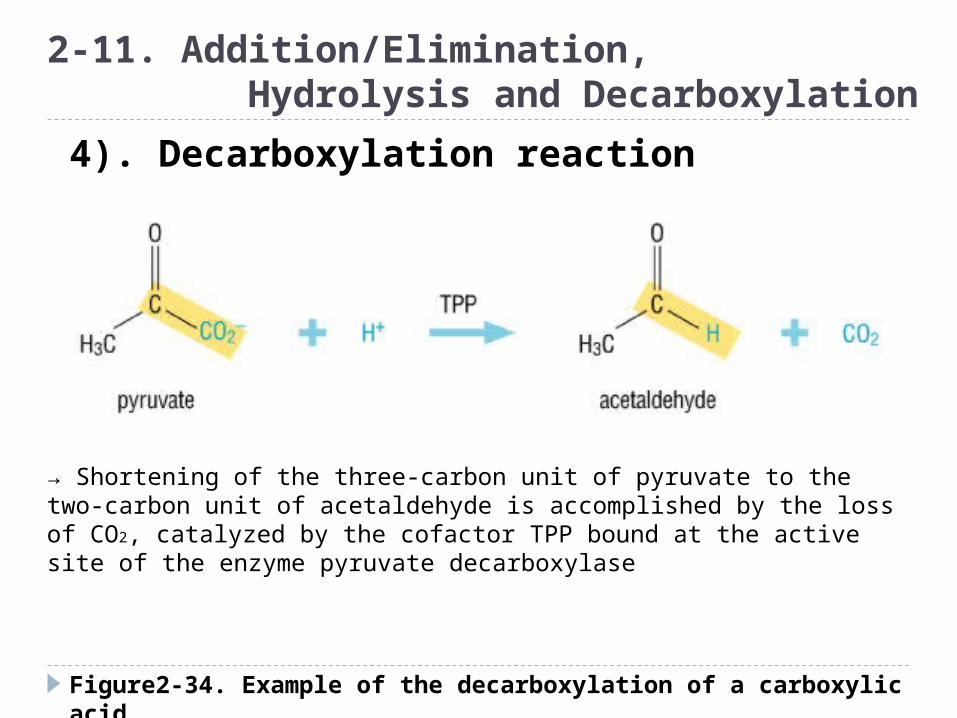
Figure2-34. Example of the decarboxylation of a carboxylic acid
2-11. Addition/Elimination, Hydrolysis and Decarboxylation
4). Decarboxylation reaction
→ Shortening of the three-carbon unit of pyruvate to the two-carbon unit of acetaldehyde is accomplished by the loss of CO2, catalyzed by the cofactor TPP bound at the active site of the enzyme pyruvate decarboxylase

Figure2-35. Table of pKa values for some common weak acids in biology
2-12. Active-Site Chemistry
- Acid : proton
donating group
-Base : proton
accepting group
-Acid-base
catalysis : proton
transferring catalysis
-pKa value : proton
affinity
◈ Acid-base catalysis

Figure2-36. Active site of lysozyme
2-12. Active-Site Chemistry
-Enzymes can increase the efficiency of acid-base reactions by
changing the intrinsic pKa values of the groups involved
-Two carboxylic acid side chins(Asp and Glu) are found in the active site of
lysozyme
-Glu 35 pKa = 4 in solution, But with no water around to accept a proton,
the carboxylic acid tend to hang on to its hydrogen. pKa=6
pKa=4 pKa=6

Figure2-37. Table of organic cofactors
2-13. Many active sites use cofactors to assist catalysis
-Not every biological reaction can be carried out efficiently using only
the chemical properties of the 20 naturally occurring amino acids.
Ex) Unpaired electron
-To overcome limitations, many enzyme active sites contain non-
amino-acid cofactors that allow specialized chemical functions
-Cofactors can be as small as metal ion / as large as heterocyclic
organometallic complex (Heme)
-Coenzyme : cofactors that are organic compounds and assist
catalysis
-Right cofactor binding to the right protein at the right
time!!
◈ Cofactor and coenzyme

Figure2-37. Table of organic cofactors
2-13. Cofactors
In humans most cofactors are derived from vitamins and minerals in the diet

Figure2-37. Table of organic cofactors
2-13. Cofactors
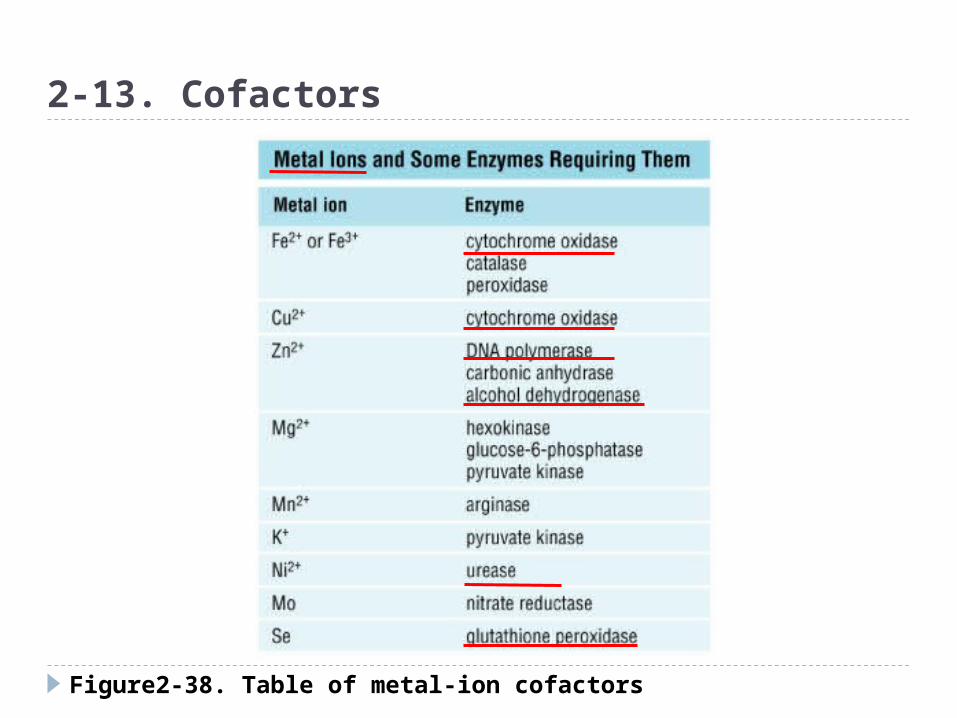
Figure2-38. Table of metal-ion cofactors
2-13. Cofactors

Figure2-39. The coenzyme lysine tyrosylquinone
2-13. Cofactors
-LTQ : unusaul coenzyme in
copper amine oxidase
-LTQ is synthesized by the
addition of a lysine side chain of
the enzyme itself
-Is essential for the proper cross-
linking of collagen and elastin
Lys side chain
Oxidized form of tyrosine
Organic cofactor can be synthesized by modification of amino acid side chain
-GFP (green fluorescent protein), chromophore is synthesized by the protein itself from the reaction of a tyrosine with neighboring serine and glycine.
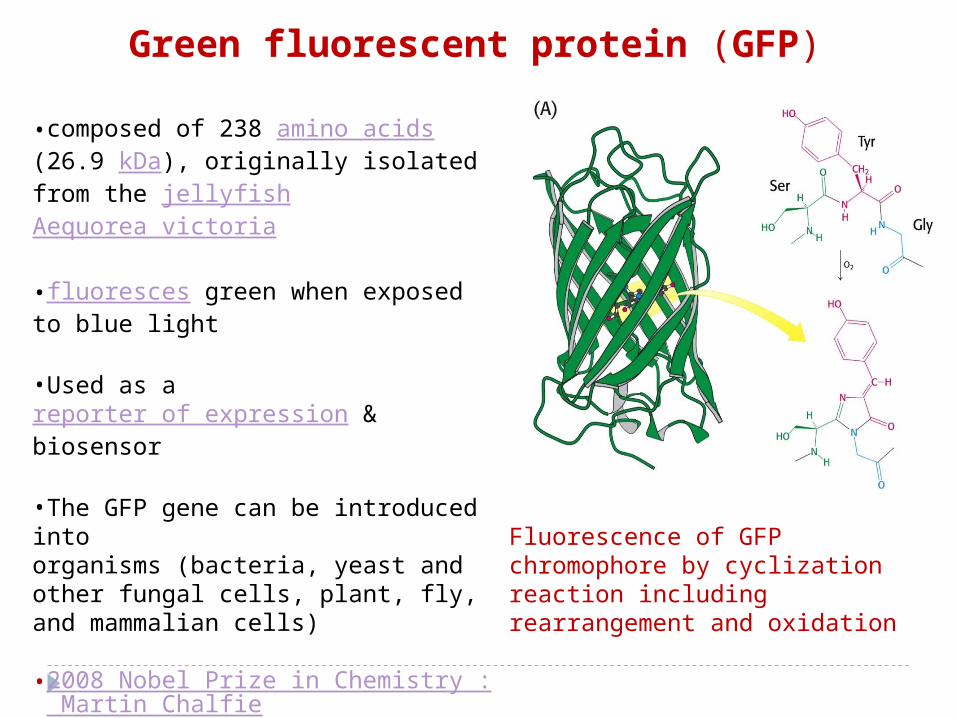
Fluorescence of GFP chromophore by cyclization reaction including rearrangement and oxidation
•composed of 238 amino acids (26.9 kDa), originally isolated from the jellyfish Aequorea victoria
•fluoresces green when exposed to blue light
•Used as a reporter of expression & biosensor
•The GFP gene can be introduced into organisms (bacteria, yeast and other fungal cells, plant, fly, and mammalian cells)
•2008 Nobel Prize in Chemistry : Martin Chalfie
, Osamu Shimomura and Roger Y. Tsien
•A typical beta barrel structure
Green fluorescent protein (GFP)

Figure2-40. The chemical steps in peptide hydrolysis catalyzed by the serine protease chymotrypsin
2-14. Multi-Step Reactions
To achieve high-energy demanding catalysis, the reactions need to break up into a number of steps, each of which has a lower-energy transition state instead of stabilization of transition state.
(a)Use the hydroxyl group of a
serine side chain and
nucleophile attack the
carbonyl carbon of the amide
bond
(Ser OH group was activated by
His and close to the substrate.
Asp-His-Ser is called catalytic
triad)
◈ Multi-step reaction
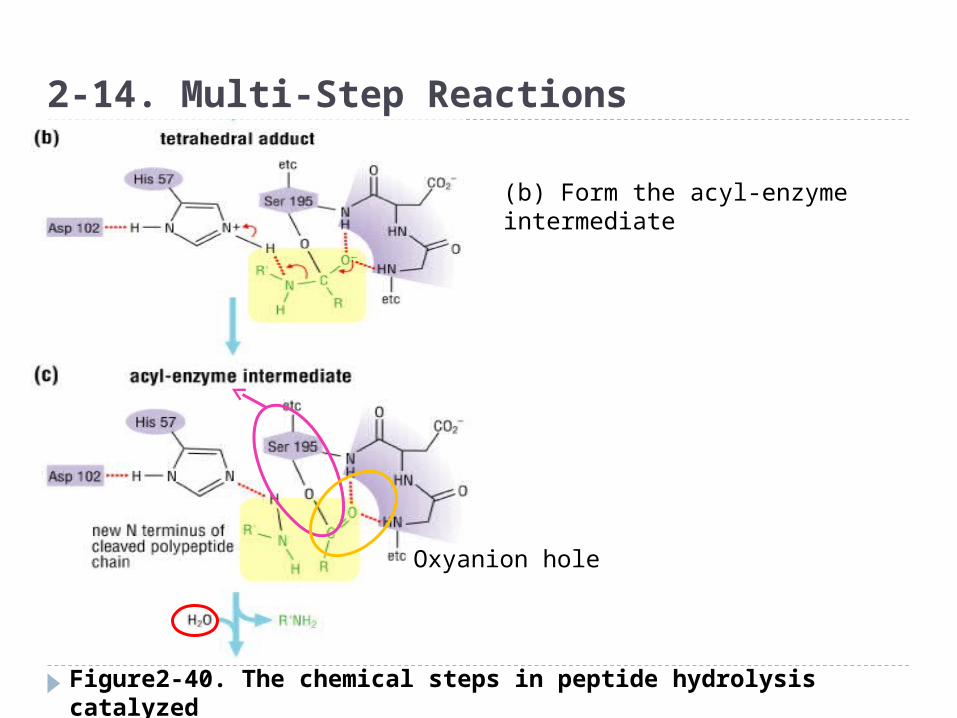
Figure2-40. The chemical steps in peptide hydrolysis catalyzed by the serine protease chymotrypsin
2-14. Multi-Step Reactions
Oxyanion hole
(b) Form the acyl-enzyme intermediate
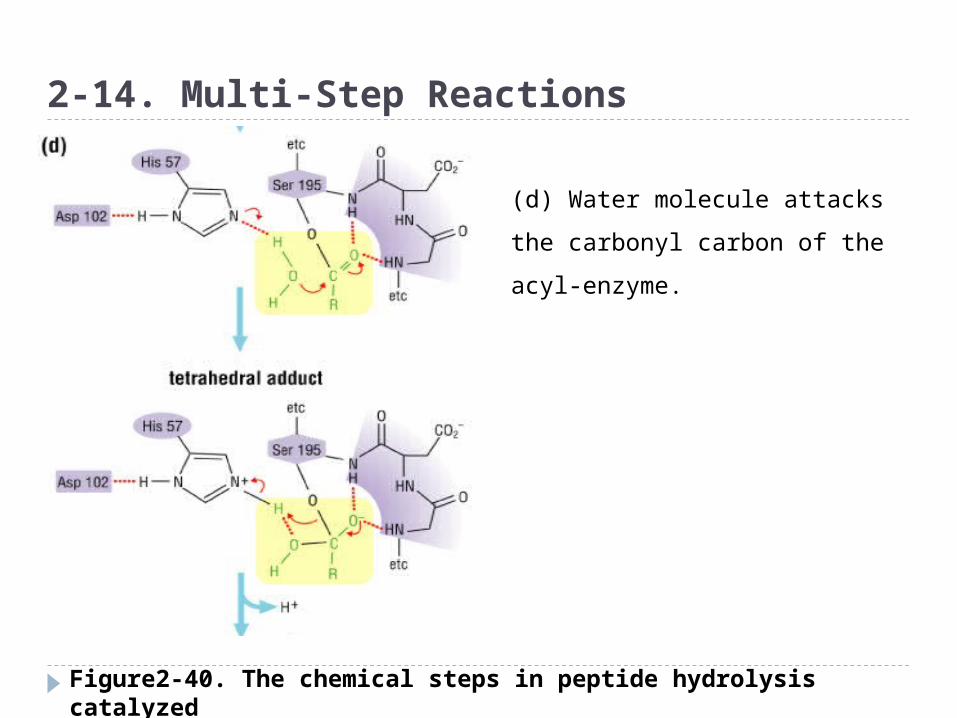
Figure2-40. The chemical steps in peptide hydrolysis catalyzed by the serine protease chymotrypsin
2-14. Multi-Step Reactions
(d) Water molecule attacks the
carbonyl carbon of the acyl-
enzyme.
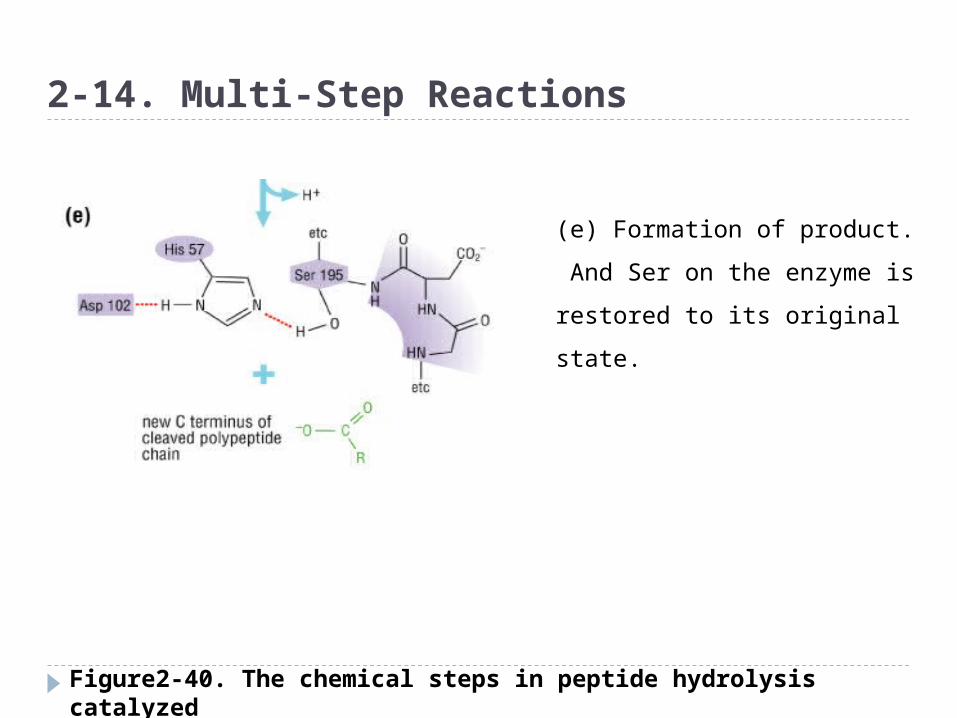
Figure2-40. The chemical steps in peptide hydrolysis catalyzed by the serine protease chymotrypsin
2-14. Multi-Step Reactions
(e) Formation of product.
And Ser on the enzyme is
restored to its original state.

2-15. Multifunctional Enzymes
- catalyze more than one chemical transformation- one or more active sites- Three classes 1) Two reaction take place consecutively at the
same active site 2) Two separate chemical reactions are
catalyzed by two distinct active sites in different domains
3) Two or more reactions are catalyzed by two or more distinct active sites which are connected by internal channels
◈ Bifunctional (or multifunctional) enzymes
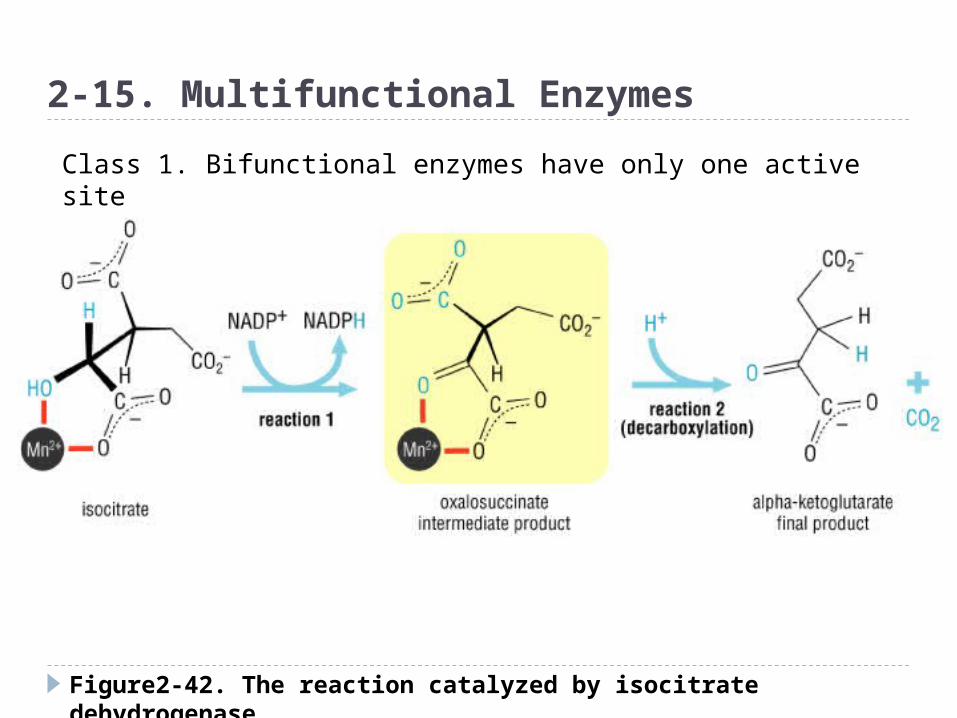
Figure2-42. The reaction catalyzed by isocitrate dehydrogenase
2-15. Multifunctional Enzymes
Class 1. Bifunctional enzymes have only one active site

Figure2-43. The bifunctional enzyme, AICAR transformylase-IMP cyclohydrolase (ATIC) is a single enzyme with two distinct active sites
2-15. Multifunctional EnzymesClass 2. Bifunctional enzymes contain two active site
Intertwined homodimer
transformylase cyclohydrolase
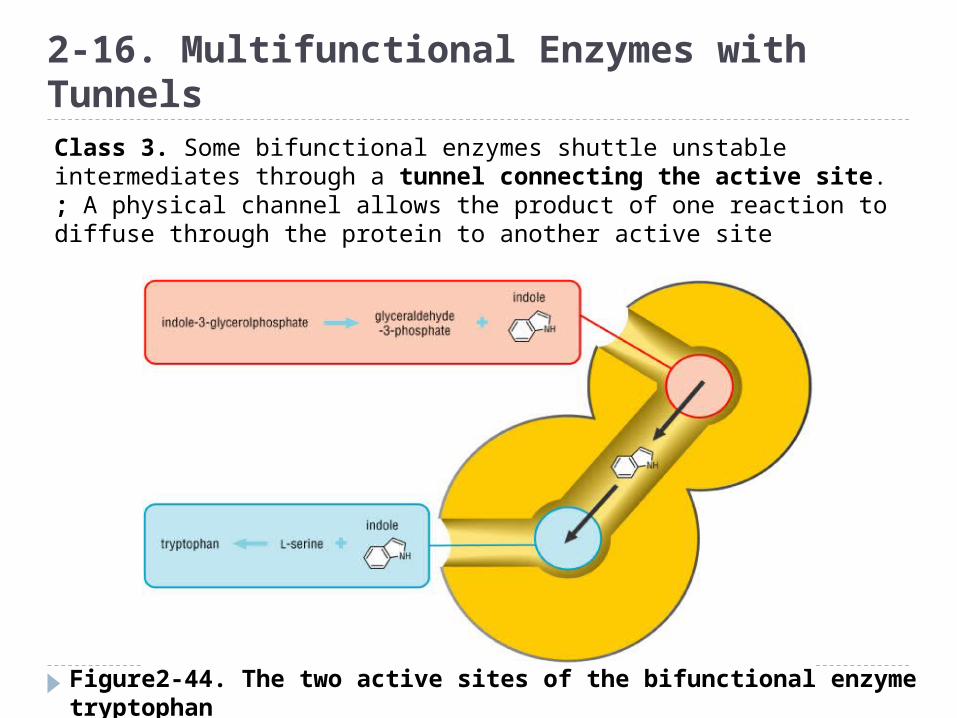
Figure2-44. The two active sites of the bifunctional enzyme tryptophan synthase are linked by an internal channel
2-16. Multifunctional Enzymes with TunnelsClass 3. Some bifunctional enzymes shuttle unstable intermediates through a tunnel connecting the active site.; A physical channel allows the product of one reaction to diffuse through the protein to another active site

Figure2-45. Three consecutive reactions are catalyzed by the three active sites of the enzyme carbamoyl phosphate synthetase
2-16. Multifunctional Enzymes with Tunnels
-Carbamoyl phosphate synthetase
-The single-chain protein has three
separate active sites connected by
two tunnels through the interior of the
protein
-The entire journey from first substrate to
final product covers a distance of nearly
100ÅGlutamine is hydrolyzed to ammoniaAmmonia + carboxyphosphate (ATP+HCO3-) carbamateCarbamate + ATP carbamoyl phosphate + ADP

Some enzymes have non-enzymatic functions Regulatory functions; transcription factors,
signaling proteins, essential cofactors in protein synthesis, cytokines or growth factors
Thymidylate synthase also functions as an RNA-binding proteins
- bind to mRNAs including transcripts of the p53 and the myc family genes and repress the translation of these genes
- Good target for anti-cancer drugs