protein adhesion, friction, unfolding, compaction
DESCRIPTION
Protein adhesion, friction, unfolding, compaction. D. Horinek, A. Alexander-Katz, A. Serr, Roland Netz, TU München. spider-silk peptide adhesion and friction at surfaces hydrophobic versus hydrophilic adhesion (all-atomistic MD simulations) shear-induced protein unfolding in blood - PowerPoint PPT PresentationTRANSCRIPT

Protein adhesion, friction, unfolding, compactionD. Horinek, A. Alexander-Katz, A. Serr,
Roland Netz, TU München
1) spider-silk peptide adhesion and friction at surfaces hydrophobic versus hydrophilic adhesion
(all-atomistic MD simulations)
2) shear-induced protein unfolding in blood fluctuation-induced hydrodynamic instabilities(hydrodynamic simulations, scaling arguments)
3) anomalous polymer sedimentation- conformational changes at high sedimentation rates
(compaction versus stretching)

Forces at Hydrophobic Interfaces
E. E. Meyer, K. J. Rosenberg, J. Israelachvili PNAS 2006, 103, 15739
Hydrophobic forces act between particles whose surfaces do not posess polar groups, regardless of the exact chemical composition.
Hydrophobic forces give rise to many different phenomena,
Short-ranged versus long-ranged
Theoretically, hydrophobic forces are not uniquely defined.
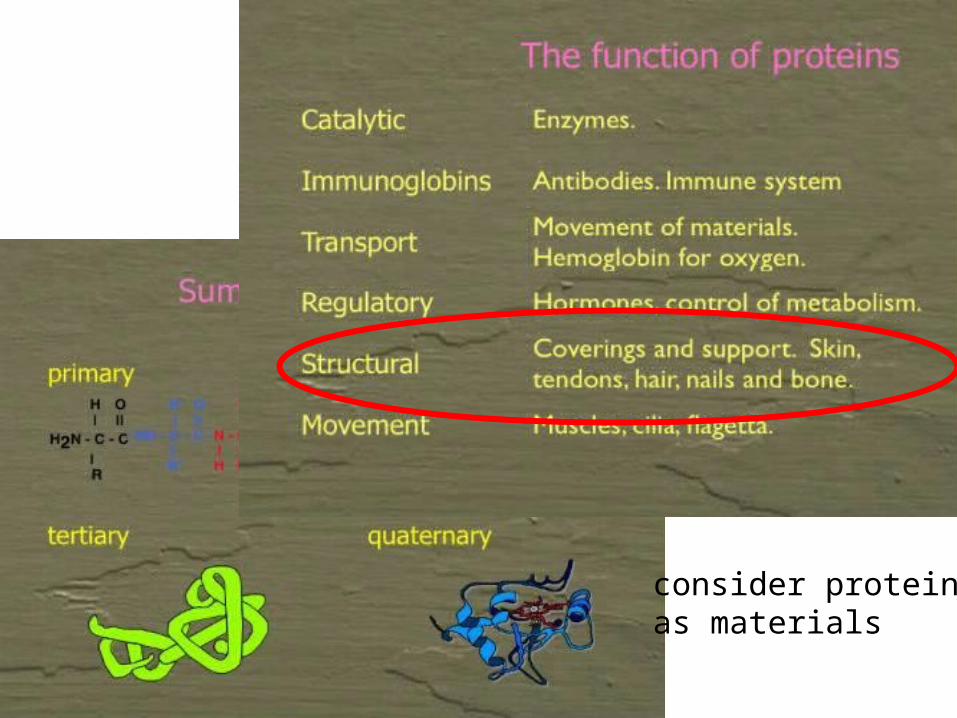
consider proteins as materials
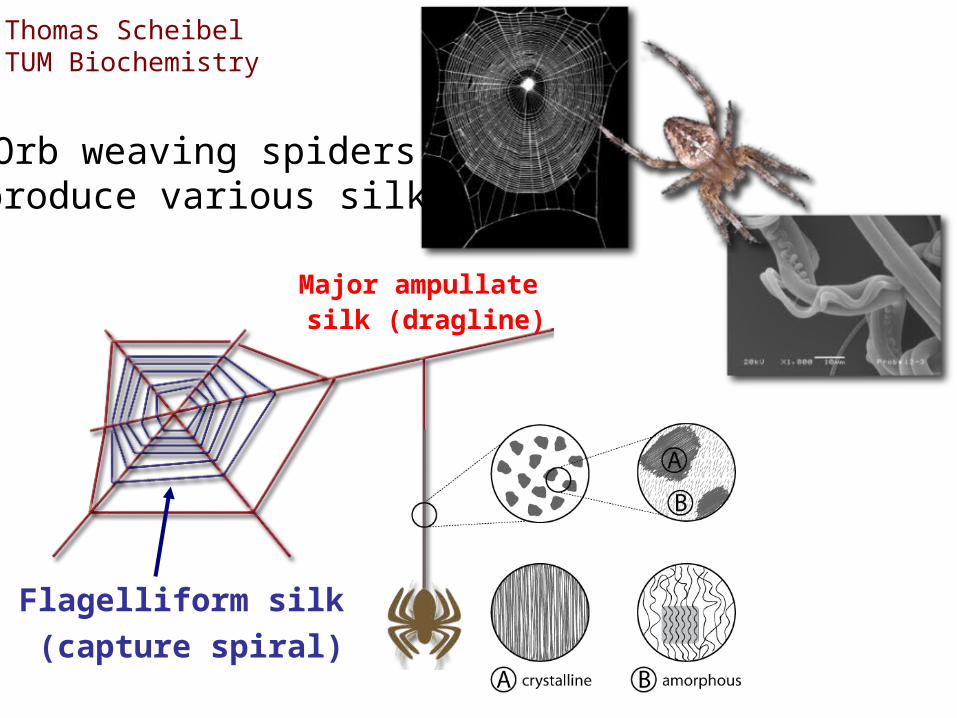
Orb weaving spiders produce various silks
Major ampullate silk (dragline)
Flagelliform silk
(capture spiral)
Thomas ScheibelTUM Biochemistry

Thomas ScheibelTUM Biochemistry
Andreas BauschTUM Biophysics
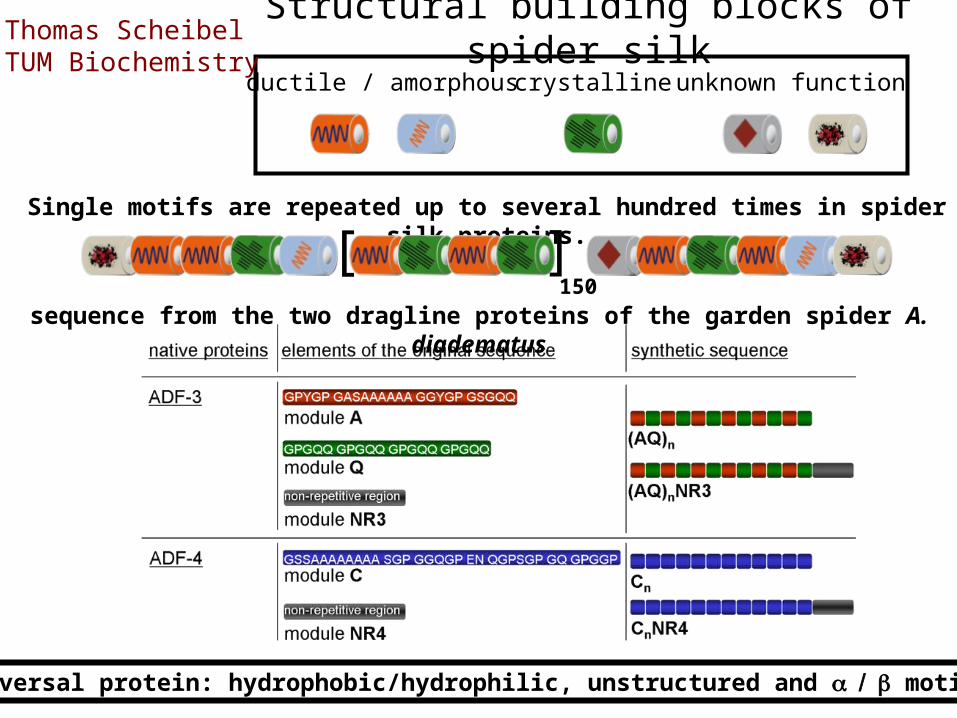
sequence from the two dragline proteins of the garden spider A. diadematus
Single motifs are repeated up to several hundred times in spider silk proteins.[ ]
150
Structural building blocks of spider silkductile / amorphouscrystallineunknown function
Thomas ScheibelTUM Biochemistry
Universal protein: hydrophobic/hydrophilic, unstructured and motifs
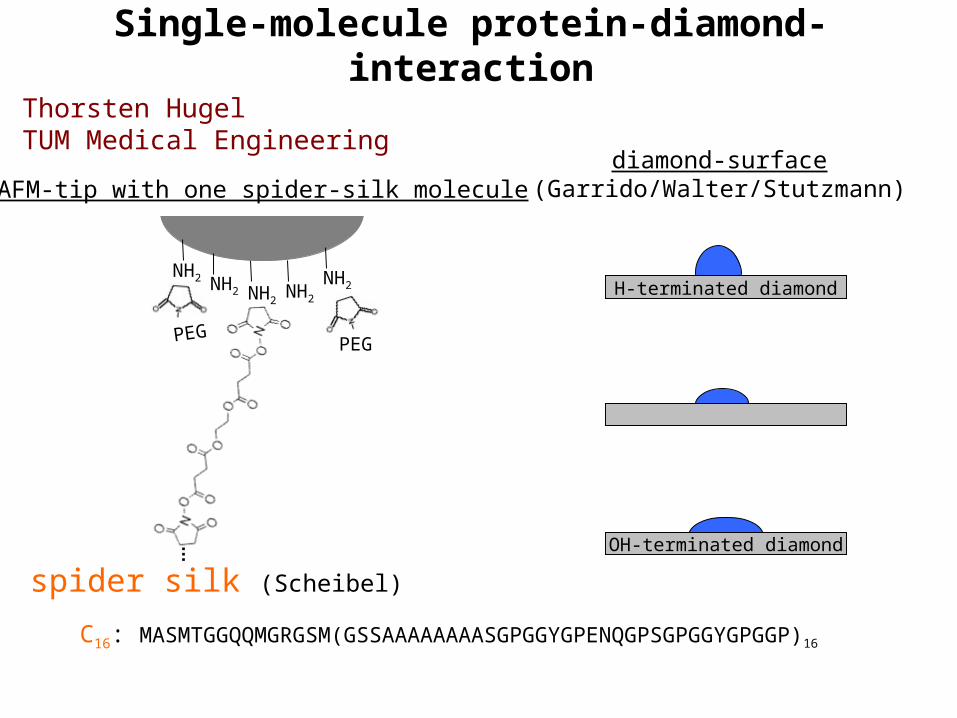
Single-molecule protein-diamond-interaction
PEG
NH2
spider silk (Scheibel)
AFM-tip with one spider-silk molecule
NH2
NH2NH2
NH2
PEG
diamond-surface(Garrido/Walter/Stutzmann)
H-terminated diamond
OH-terminated diamond
C16: MASMTGGQQMGRGSM(GSSAAAAAAAASGPGGYGPENQGPSGPGGYGPGGP)16
Thorsten HugelTUM Medical Engineering
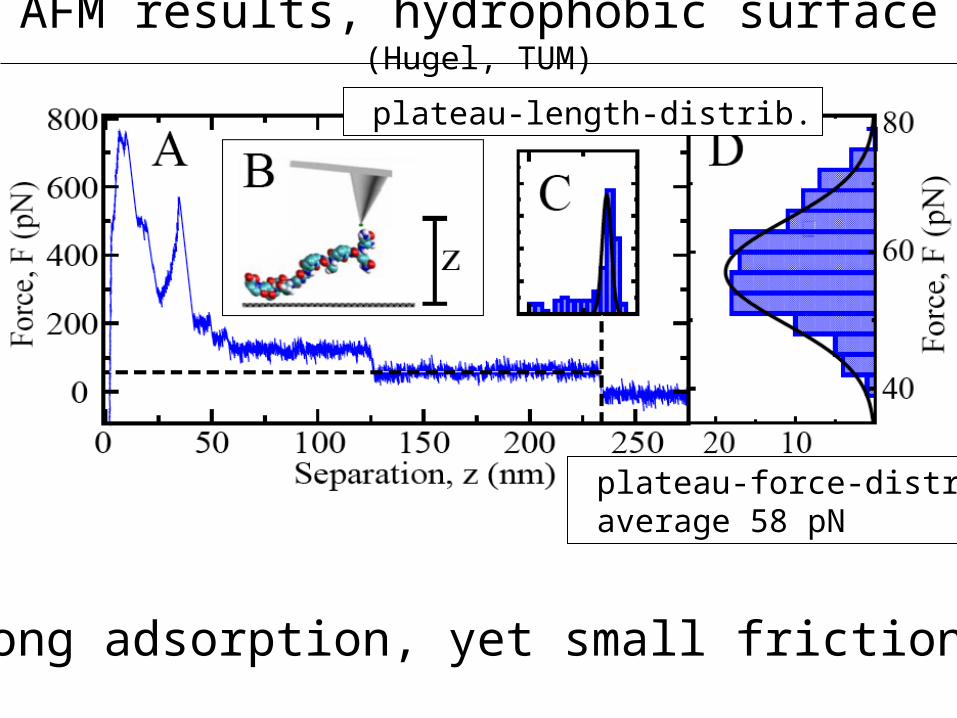
AFM results, hydrophobic surface (Hugel, TUM)
plateau-length-distrib.
plateau-force-distrib. average 58 pN
-strong adsorption, yet small friction

R
F
FT
if applied tangential force FT smallerthan rate-dependent frictional resistance, polymer sticks; --> angle self-adjustsSerr, Netz, EPL 73, 292 (2006)
FN
Vertical pulling at constant speed, low frictionVertical pulling at constant speed, high friction
Zur Anzeige wird der QuickTime™ Dekompressor „YUV420 codec“
benötigt.Zur Anzeige wird der QuickTime™ Dekompressor „YUV420 codec“
benötigt.

MD Simulations (Dominik Horinek)
For simulations, the spider silk C16 motif is cut in three pieces:
GSSAAAAAAAASGPGGYGPENQGPSGPGGYGPGGP GSSAAAAAAA fragment 1GPGGYGPENQGPSGPGGYGPGGP GSSAAAAAAAASGPGGYGPENQGPSG fragment 2 2GP GSSAAAAAAAASGPGGYGPENQGPSGPGGYGPGGP fragment 3
water (SPC)
peptide fragment (Gromos96)
surface (Gromos96)
H-terminated diamond
OH-terminated diamond
Alkane SAM
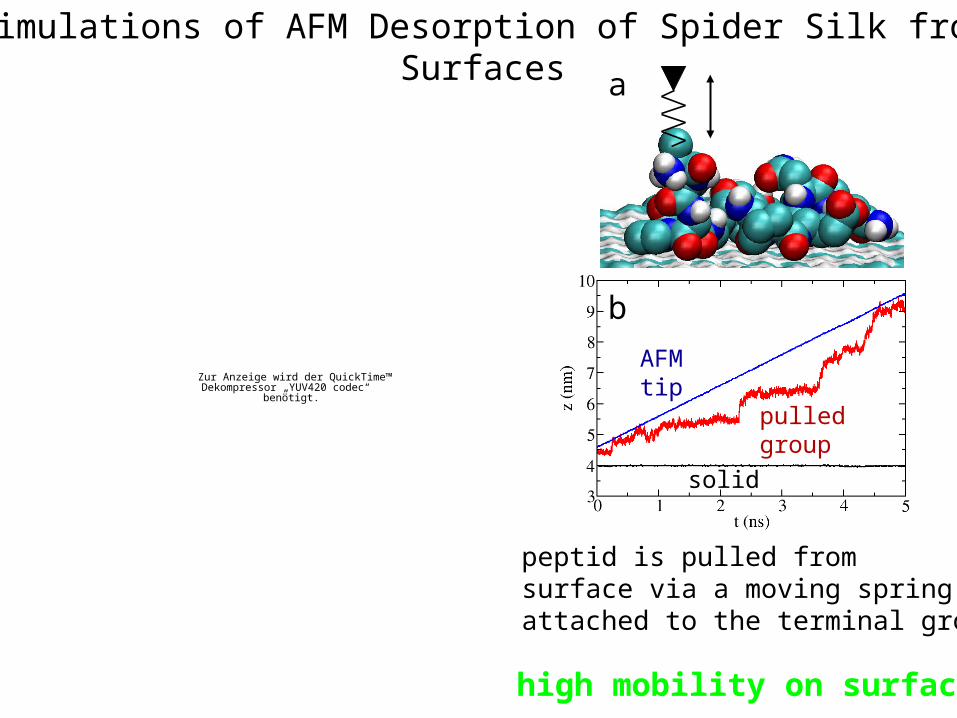
Simulations of AFM Desorption of Spider Silk from Surfaces
AFM tip
pulled group
solid
a
b
peptid is pulled fromsurface via a moving springattached to the terminal group
Zur Anzeige wird der QuickTime™ Dekompressor „YUV420 codec“
benötigt.
high mobility on surface!
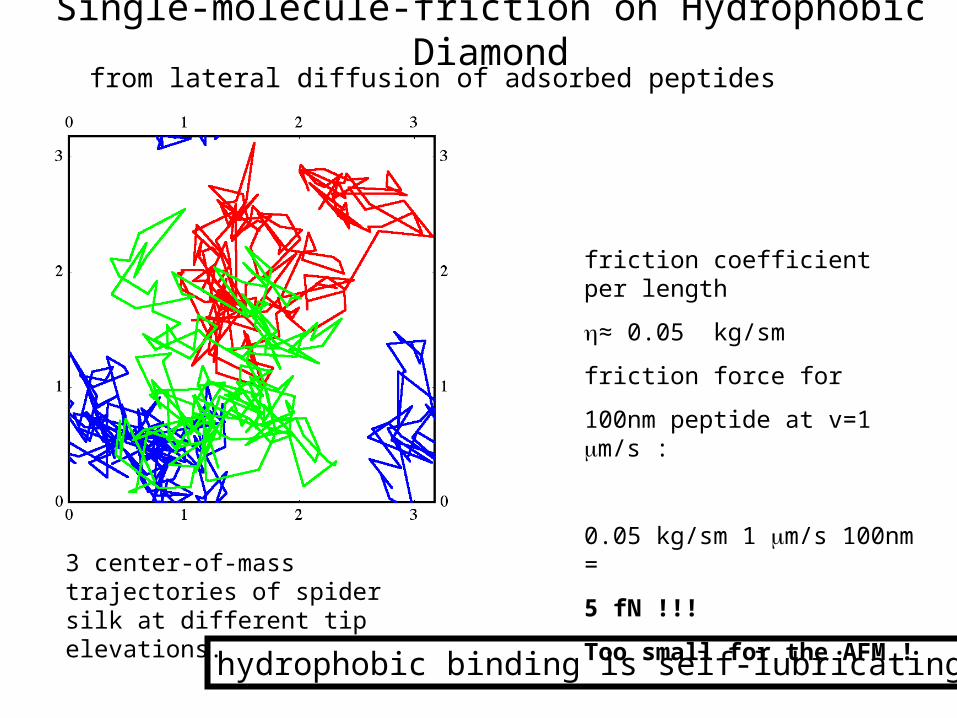
friction coefficient per length
≈ 0.05 kg/sm
friction force for
100nm peptide at v=1 m/s :
0.05 kg/sm 1 m/s 100nm =
5 fN !!!
Too small for the AFM !3 center-of-mass trajectories of spider silk at different tip elevations.
Single-molecule-friction on Hydrophobic Diamondfrom lateral diffusion of adsorbed peptides
hydrophobic binding is self-lubricating

10
0.1
a
b
c
d
Desorption Forces from MD at various pulling ratesHorinek, RRN, PNAS (2008)
1
Fdes = k (zspring -zAA)
pulling rate 10 m/s
pulling rate 1 m/s
pulling rate 0.1 m/s
hydrophobic surfaceaverage plateau force: 54 pN(experimental: 58 pN with NaCl)
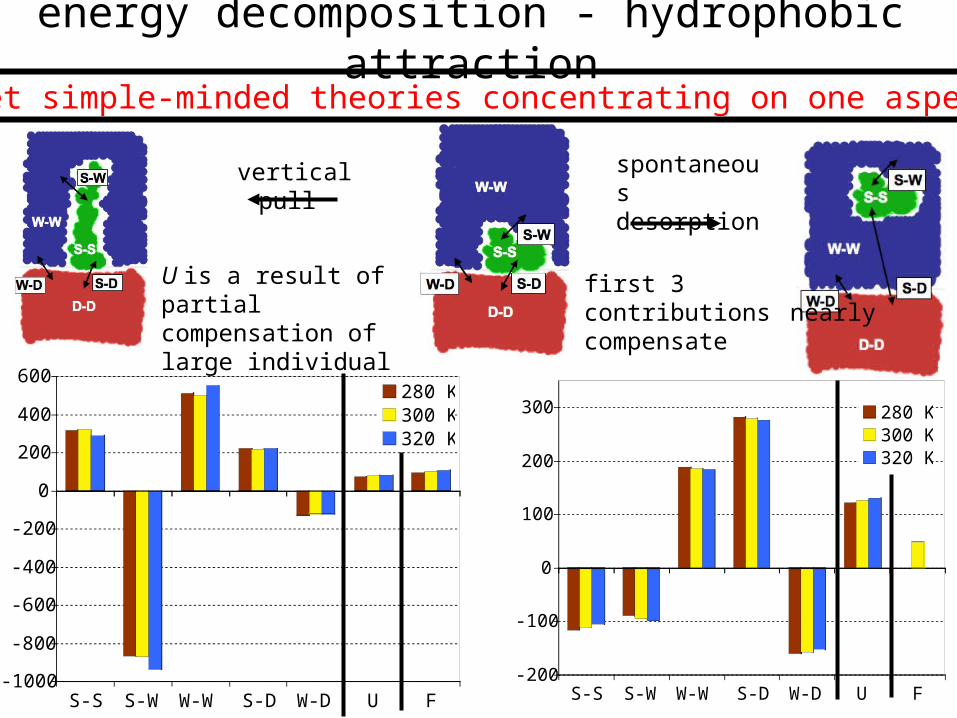
energy decomposition - hydrophobic attraction
vertical pull
U is a result of partial compensation of large individual energies
-1000
-800
-600
-400
-200
0
200
400
600
S-S S-W W-W S-D W-D U F
280 K300 K320 K
first 3 contributions nearly compensate
spontaneous desorption
-200
-100
0
100
200
300
S-S S-W W-W S-D W-D U F
280 K300 K320 K
forget simple-minded theories concentrating on one aspect !!

hydrophilic OH-terminated diamond
desorption (at most) doubled on hydrophilic surface
large friction due to breaking and reformation of hydrogen bonds !!
Pulling rate 0.1 m/sLarge hysteresis !
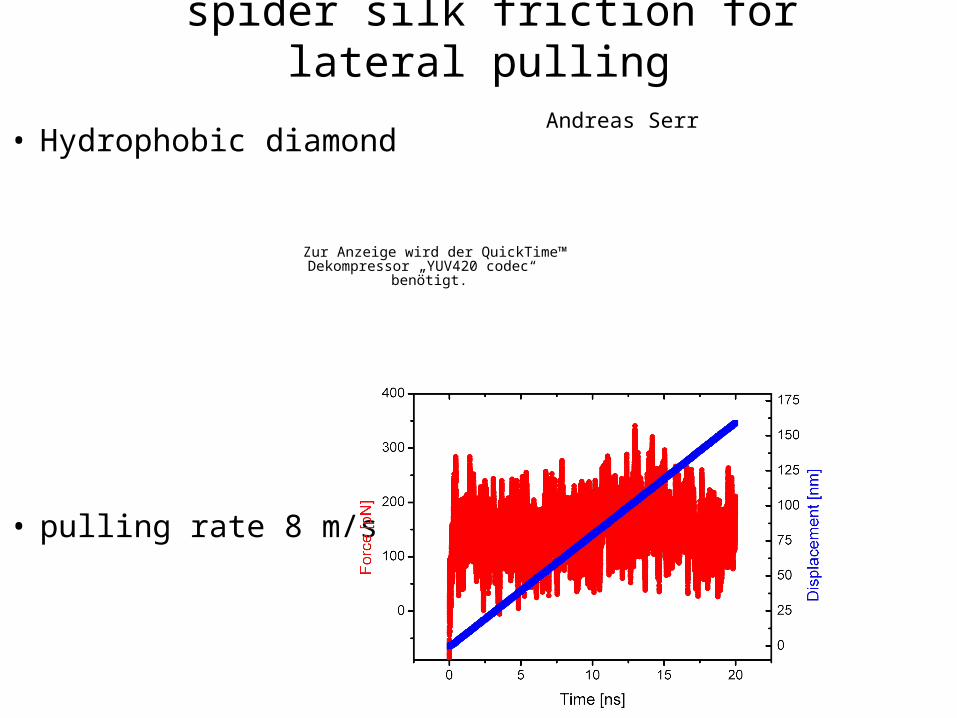
spider silk friction for lateral pullingAndreas Serr
• Hydrophobic diamond
• pulling rate 8 m/s
Zur Anzeige wird der QuickTime™ Dekompressor „YUV420 codec“
benötigt.
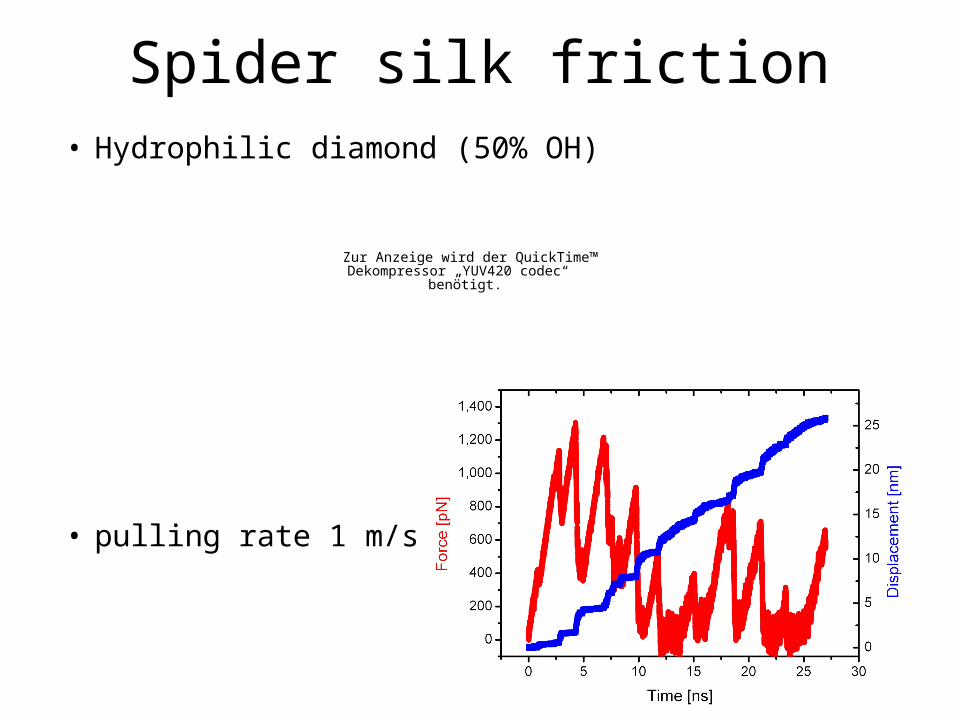
• Hydrophilic diamond (50% OH)
• pulling rate 1 m/s
Spider silk friction
Zur Anzeige wird der QuickTime™ Dekompressor „YUV420 codec“
benötigt.

Single-molecule peptide friction• mobility per monomer
hydrophilic diam
hydrophobic diam
bulk water (perfect match with exp.)
peptide glides on vacuum: „hydrophobic binding is self-lubricating“
30-fold friction increase for hydrophilic surface: driven diffusion in corrugated binding potential of 6kBT (Frenkel-Kontorova-Tomlinson)
simulation 0.1m/s -> 5pN
simulation 0.1m/s -> 200pN
experimental friction forces: o.k. agreement with exp. data

- adhesive proteins bind to BOTH hydrophilic and hydrophobic surfaces strongly (5 kBT per amino acid)
- nano-friction on hydrophobic/philic substrates is very different (effective adhesive properties depend on binding free energy AND surface friction ! gecko, scotch tape)
- in all cases, effective interaction involves direct interactions as well as water-ordering effects!
FN
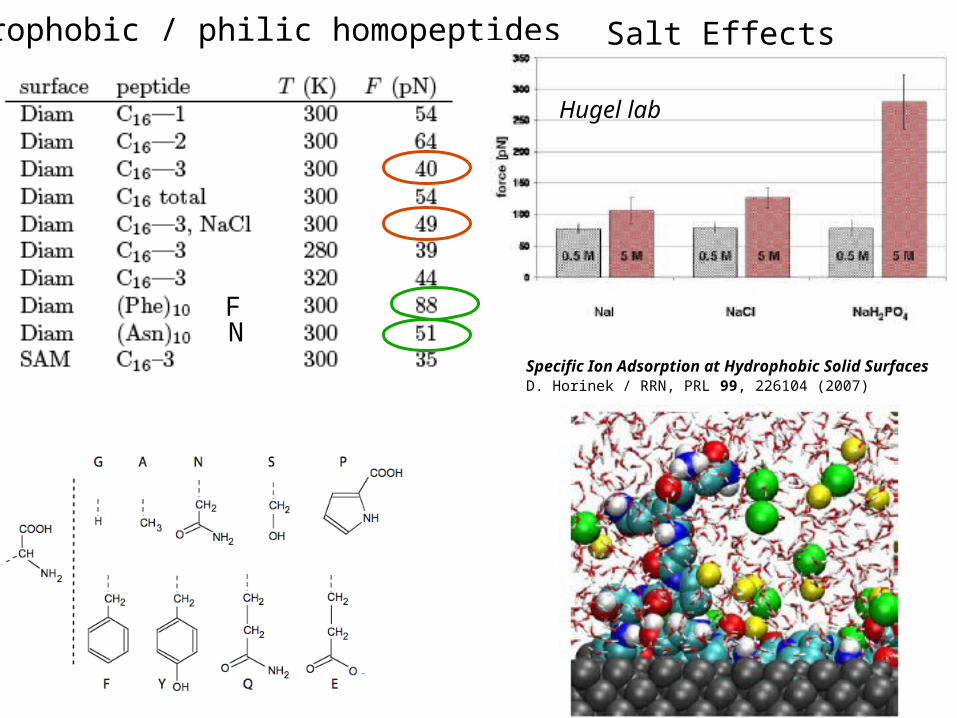
hydrophobic / philic homopeptides Salt Effects
Hugel lab
Specific Ion Adsorption at Hydrophobic Solid Surfaces D. Horinek / RRN, PRL 99, 226104 (2007)

pressure between 2 hydrophobic surfaces from Poisson-Boltzmann:
screenable contribution to hydrophobic attraction
1 mM salt: weak but long-ranged
100 mM salt: strong but short-ranged
DOES NOT YET EXPLAIN PEPTIDE ION SPECIFICITY!

blood functions:
- oxygene - transport (& Hemoglobin)- nutrient - transport (glucose, amino-acids, fat ....)- waste - transport (CO2l urea, lactatic acid ...)- immuno reactione ( lymphocytes, antibodies ...)- signal - transduction (hormons ...)- regulation of temperature and pH of body- coagulation, vascular repair

capillaries connect arteries and veins they are 5-10 microns thick and are lined by a single-cell-layer: the endothelium

action
since the endothelial layer is thin, it ruptures easily !
the von-Willebrand-factor (vWF) helps fixing capillaries

DockingTransport Fusion
von-Willebrand Faktor (globular !!!)
von-Willebrand Faktor(fibers !!!)Blood
IntracellularVesicels (packaged proteins)
vWf unfolds in shear

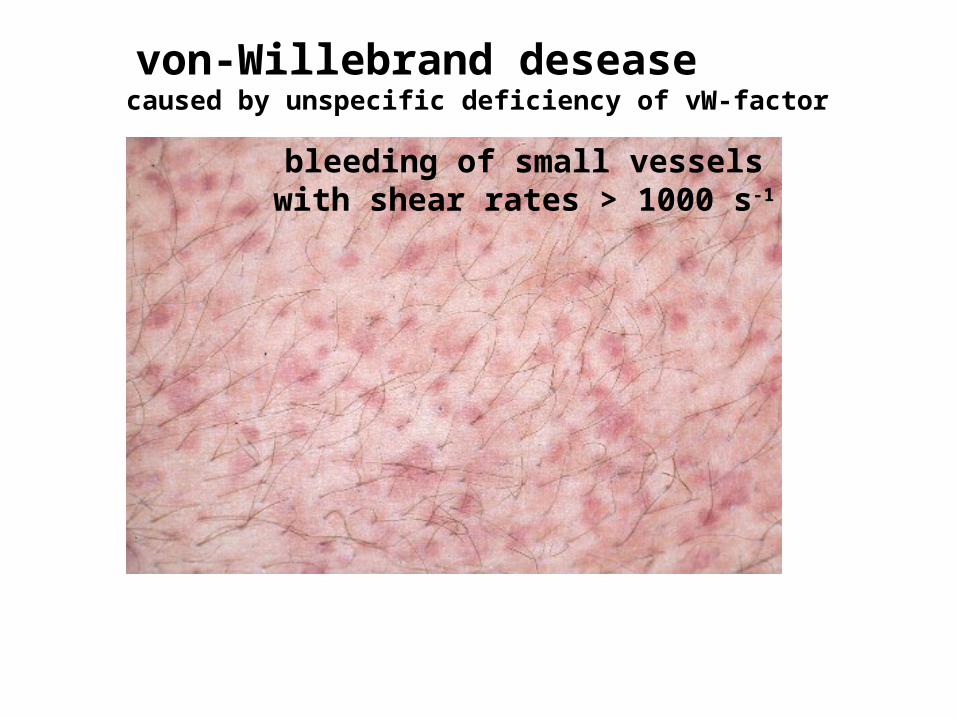
von-Willebrand desease caused by unspecific deficiency of vW-factor
bleeding of small vessels with shear rates > 1000 s-1


the vWf is the largest watersoluble protein in the body --- why ???
monomer(2500 aminoacids)
dimer
multimer (a few hundred units)
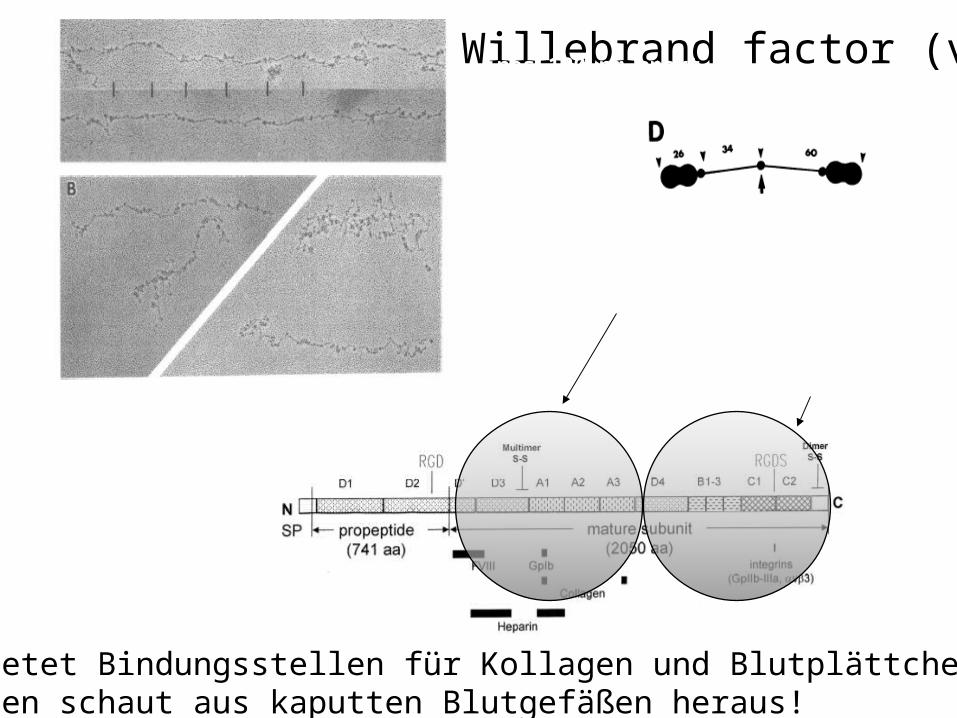
von-Willebrand factor (vWf)
Large globular structure~ 25x6.5 nm
Rod + central nodule~ 30 + 6 nm
Lines 120 nm apart
Fowler et al
vWf bietet Bindungsstellen für Kollagen und Blutplättchen,Kollagen schaut aus kaputten Blutgefäßen heraus!

Was stimuliert die Entfaltung des vWf ??
Hypothese: Scherfluss in kleinen Blutgefäßen bewirktEntfaltung des Proteins!
Hagen-Poiseuille Gesetz für Strömung im Rohr:Flüssigkeitsstrom geht wie R4
Strömungsgeschwindigkeit ist Null an der WandScherung verformt Proteine und Blutkörperchen
Experimentelle Untersuchung an künstlichen Blutgefäßen!
R

High Frequency Input
(Source of SAW)
Hydrophilic Channel
Hydrophobic Surface
Surface Acoustic Wave(Nanopump)
LiNbO3
(Piezoelectric)
Flow-chamber Chip - Wixforth&Schneider, Augsburg
200µm
1mm
40mm
V = 8µl

Real-time movie of stretched vWf above critical shear rate
Zur Anzeige wird der QuickTime™ Dekompressor „Cinepak“
benötigt.
collapsed stretched10 µm
BelowCriticalShearγc AboveCriticalShearγc
10µm
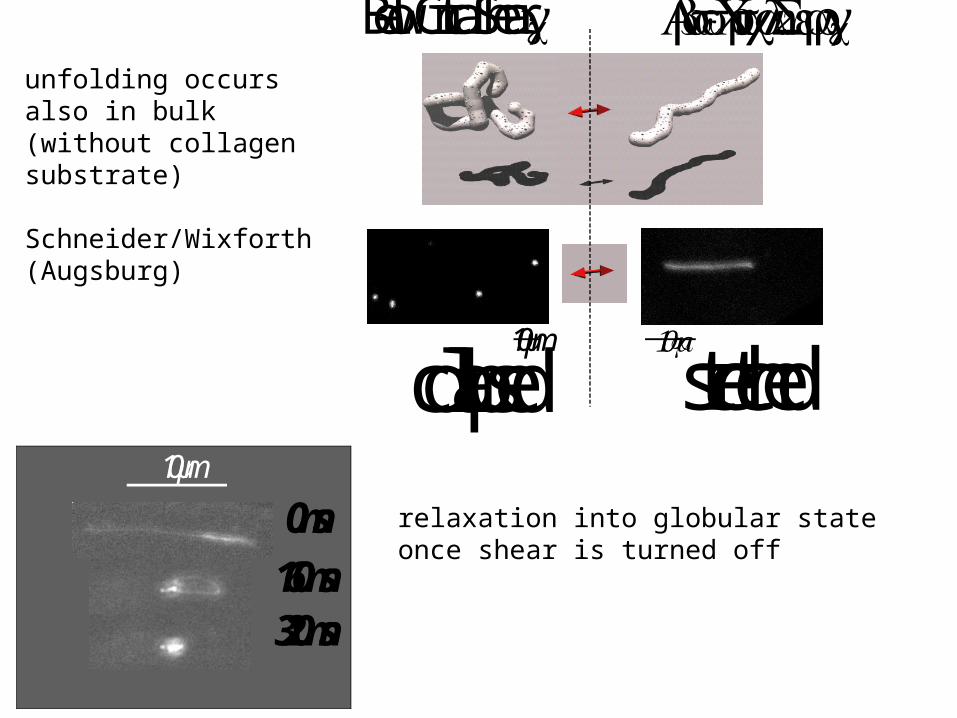
unfolding occursalso in bulk (without collagensubstrate)
Schneider/Wixforth(Augsburg)
0 ms
320 ms160 ms
10 µm
relaxation into globular stateonce shear is turned off
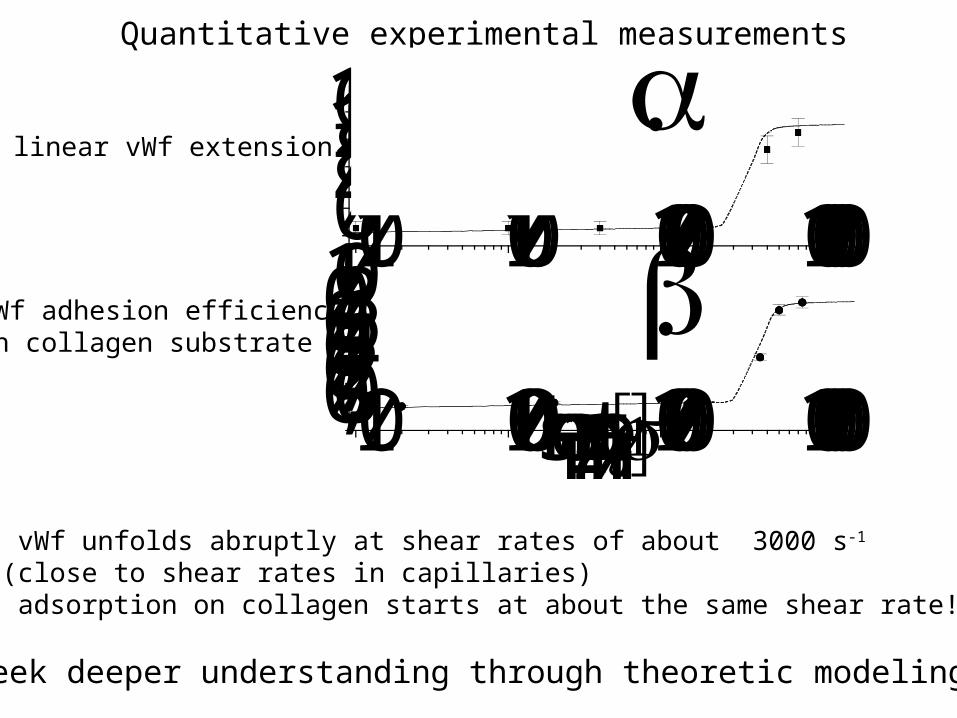
Fig. 410 100 1000 100000,00,20,40,60,81,010 100 1000 100000481216
shearrate γ[s-1]normalized rate ofadhesion
[end to end distance
µ ]mshearrateγ[s-1]
a.
b.
vWf unfolds abruptly at shear rates of about 3000 s-1
(close to shear rates in capillaries) adsorption on collagen starts at about the same shear rate!
Quantitative experimental measurements
linear vWf extension
vWf adhesion efficiency on collagen substrate
Seek deeper understanding through theoretic modeling !
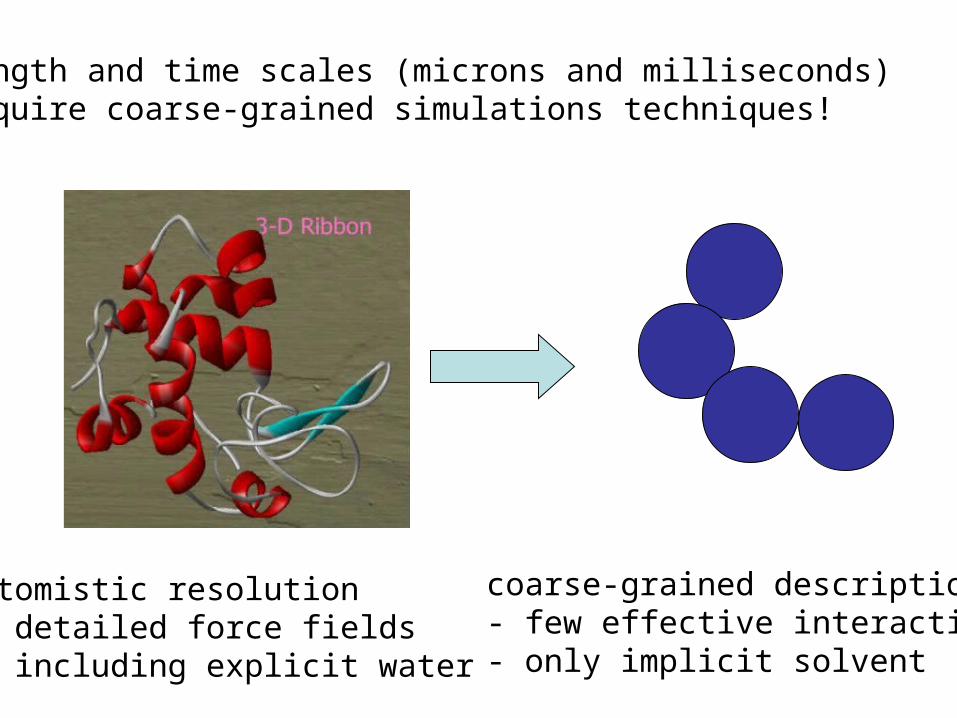
length and time scales (microns and milliseconds) require coarse-grained simulations techniques!
atomistic resolution - detailed force fields - including explicit water
coarse-grained description - few effective interactions - only implicit solvent

human bacterium sinking cylinder
H2O: = 0.001 Pa s; = 1000 kg/m3
v = 1 m/sl = 1 m
Re = 106 v ~ 10-7 m/sl = 1 Re = 10-7
v = 10-5 m/sl = 1 Re = 10-5
Hydrodynamics at low Reynolds numbers
Stationary Navier-Stokes equation
one obtains the creeping flow equation.
If the Reynolds number , ,

€
Hαβ (r) =1
8πηrδαβ +ˆ r α ˆ r β[ ]
(Oseen-Tensor)
flow-field due to point-force at origin:
€
uα(r)=Hαβ (r) f β1
for many particles the superposition principle is valid:
€
uα(r)= Hαβ (r −ri) fiβ
i
∑
invert to get forces for prescribedsolvent velocity distribution !!
Next: add thermal noise

Hydrodynamic Brownian simulation techniques
Random force
€
ξi(t)ξ j( ′ t ) =6t μ ij kBT δ(t− ′ t )
Mobility matrix:
€
t μ ij =Dij /kBT =μ0 δij +
t H (ri,rj)
€
μ0 = 6πRη( )−1
self mobility: hydrodyn. interact.
equivalent to Smoluchowski equation for particle distribut. W(rj,t) :
€
∂W∂t
=∂∂ri
Dij
∂W∂rj
−μij f jW⎡
⎣ ⎢ ⎤
⎦ ⎥ i, j
∑ with solution:
€
W ≅e−U /kBT
€
m r j(t)t μ ij +˙ r i(t) =
t μ ij f j(t)+ξi(t)Velocity of
i-th particle:
deterministic force
€
f j(t) =−∂U(t) /∂rj (t)+E
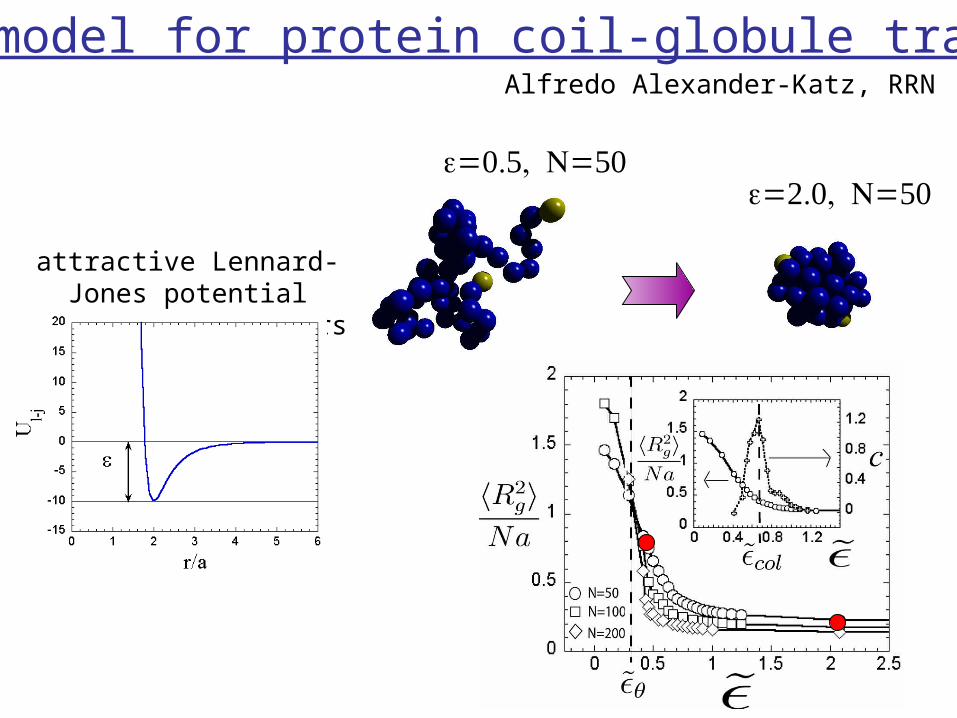
simple model for protein coil-globule transition
attractive Lennard-Jones potential between all monomers
0000
Alfredo Alexander-Katz, RRN

globule in shear flow, =2.5, γ=1.2 Alfredo Alexander-Katz, RRN
Zur Anzeige wird der QuickTime™ Dekompressor „YUV420 codec“
benötigt.
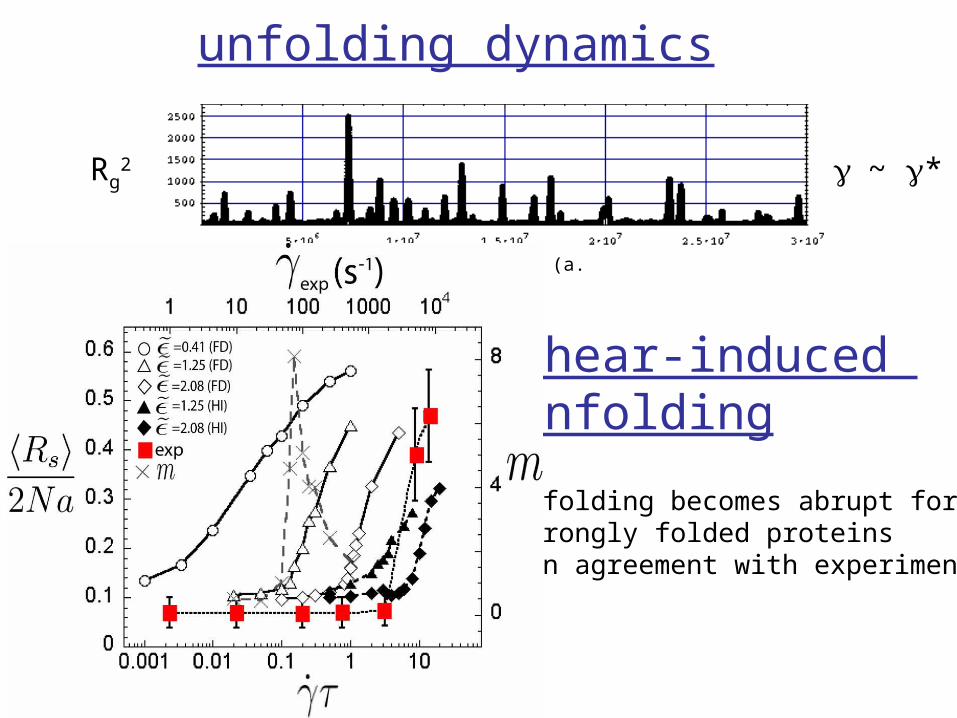
unfolding dynamics
Rg2
time (a. u.)
γ~ γ*
shear-induced unfolding
unfolding becomes abrupt for strongly folded proteins(in agreement with experiments)
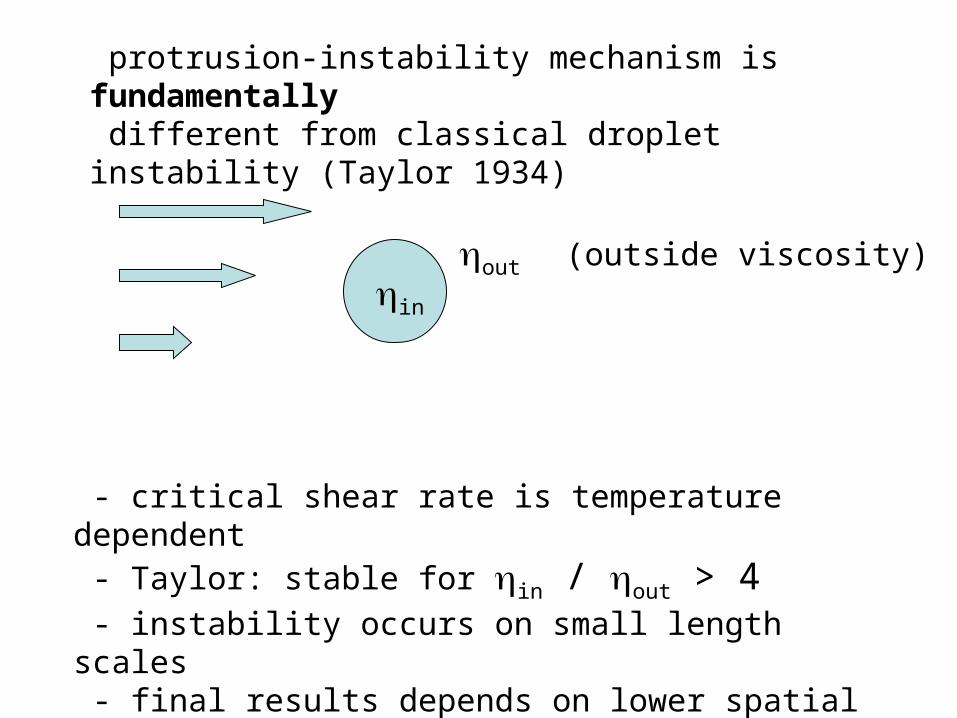
protrusion-instability mechanism is fundamentally different from classical droplet instability (Taylor 1934)
- critical shear rate is temperature dependent - Taylor: stable for in / out > 4 - instability occurs on small length scales - final results depends on lower spatial cutoff
out (outside viscosity)in

minimal model for shear-induced globule unfolding:“force balance on protrusions”
shear-force on protrusion -free draining (with slip)
€
fshear ≈ (γ•
R)(l /a)μ−1
from equipartition theorem lf=kBT--> „typical“ protrusion length
€
l ≈(kBT /Δε)1/α
€
f ≈ Δεl α −1cohesive force on protrusion(1sharp interface, diffuse interface)
--> typical cohesive force on protrusion fcoh
relative velocitysphere/solvent
# monomers
friction coefficient of one monomer
€
fshear ≈ γ•
l 3R−1a−1μ−1-hydrodynamic case (no slip)

critical protrusion length fcoh = fshear
free draining
hydrodynamic
€
γ*τ•
≈ Δε 2 /α a /R
€
γ*τ•
≈ Δε 4 /αR /a
€
τ =a2 /kBTμ

L: protein contour lengtha: protein monomer radius
€
γ*•
≈ L1/ 3(Δε /kBT)4 /α /a3
scaling of critical shear rate (with hydrodynamics) :
to unfold a protein with typical cohesion energy in a capillary vessel one needs huge monomers with a radius of 10 nm, close to vWf
=2kBT, N=100, γ=1000s-1, ----> a = 10nm !!enormously large
monomer size !!!
now connect to classical hydrodynamic instability theory (Taylor, Kelvin-Helmholtz) and assume protrusions are controlled by surface tension /a2 and 1
€
γ*•
≈ L1/ 3(σ /kBT)4 a5
instability at small length scales !!A. Alexander-Katz, RRN: PRL (2006), PNAS (2007) ……..

polymer separation in the ultracentrifuge:sedimentation anomaly at large driving fields
G: sedimentation force per monomerN: monomer number
velocity v = GN
mobility 1R 1N
velocity v ≈ G N1-
sedimentation rate S = v/G ≈ N1-
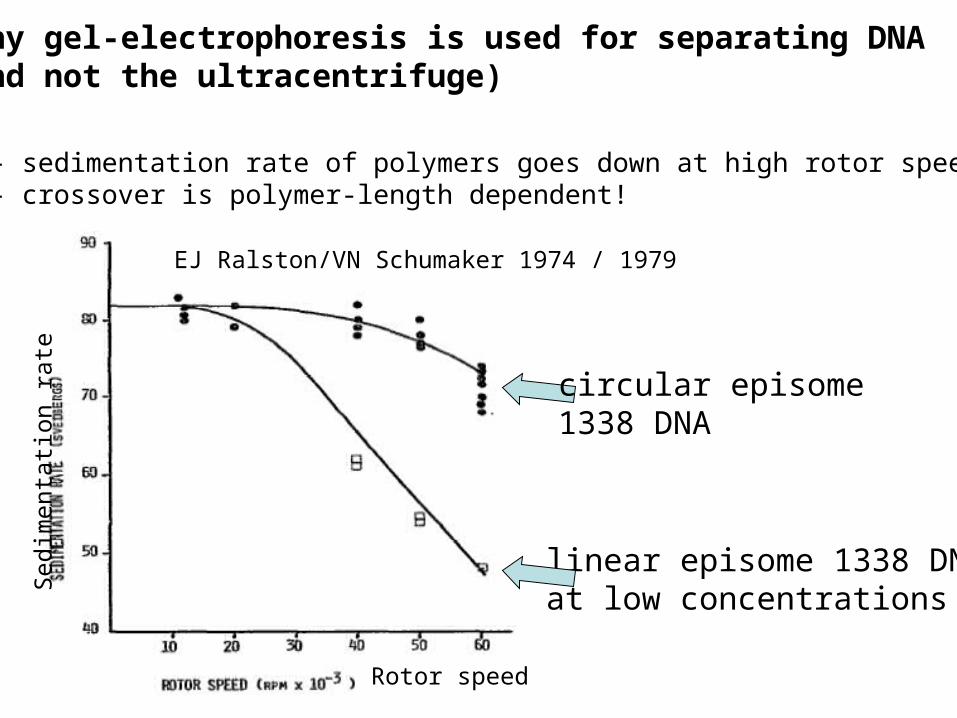
why gel-electrophoresis is used for separating DNA(and not the ultracentrifuge)
Sed
imen
tatio
n ra
te
Rotor speed
linear episome 1338 DNAat low concentrations
EJ Ralston/VN Schumaker 1974 / 1979
- sedimentation rate of polymers goes down at high rotor speeds- crossover is polymer-length dependent!
circular episome 1338 DNA
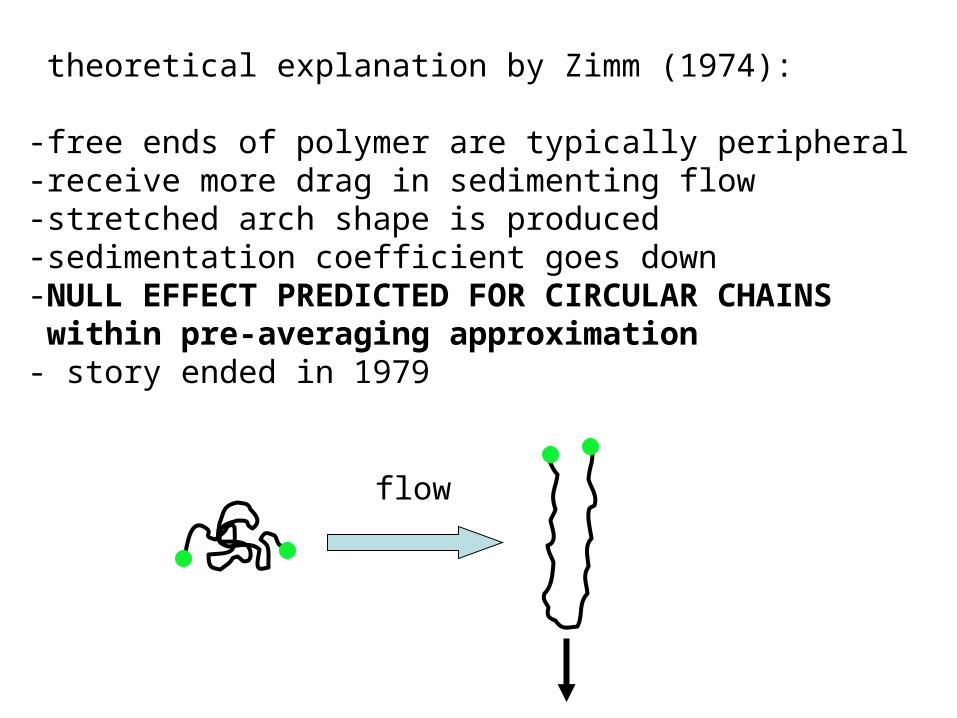
theoretical explanation by Zimm (1974):
-free ends of polymer are typically peripheral-receive more drag in sedimenting flow -stretched arch shape is produced-sedimentation coefficient goes down-NULL EFFECT PREDICTED FOR CIRCULAR CHAINS within pre-averaging approximation- story ended in 1979
flow

Crumpling and stretching of sedimenting flexible chain (Schlagberger, Netz, PRL 2007)
motion
Zur Anzeige wird der QuickTime™ Dekompressor „YUV420 codec“
benötigt.

short chains, small fields: hydrodynamic collapse of sedimenting polymers
radius
sedimentationforce
hydrodynamic drag -> internal recirculation with velocity -> recirculation time scale
€
v ≈GN /ηR
€
τ flow ≈ R /v ≈ ηR2 /GN
compare with coil relaxation time „scrambled/collapsed coil“ for τR>τflow or Ga/kBT >N-1-
€
τR ≈ ηR3 /kBT
v

stable stationary state: vh≈vt and thus Nh ≈ (ln N)3/2
(tail stretched by recirculation force, sedimentation reduced w.r.t. coil)
head velocity vh ≈ Nh2/3
tail velocity vt ≈ ln(Nt)
Long chains, large driving fields: tadpole structure(recirculation too weak to pull tail in …..)
sedimentation force
sedi
m.
rate
small fields, long chains: weak stretching perturbation analysis
S = v/G ≈ N1- [ 1 - c G2 N2+2 (same scaling as Zimm!)

tadpoles obtained with ring-polymersonly in full hydrodynamicsimulation !
full hydrodynamicsimulation
realistic theory needs to incorporate hydrodynamic interactions and entanglement effects (essential for compaction) !
hydrodynamic simulation using Zimm‘s preaveraging approximation

interfacial water structure
non-equilibriumeffects hydrodynamic effects
conformational fluctuations
protein adhesion / dynamics