probing excitation-contraction coupling in trachealis smooth muscle with the mycotoxin cyclopiazonic...
TRANSCRIPT

Clinical and Experimental Pharmacology and Physiology (1996) 23,733-737
SYMPOSIUM 7th Southeast Asian Western Pacific Regional Meeting of Pharmacologists
Pharmacology of Natural Products
PROBING EXCITATION-CONTRACTION COUPLING IN TRACHEALIS SMOOTH MUSCLE WITH THE MYCOTOXIN
CYCLOPIAZONIC ACID
Daniel Amoako, Yun Qian, Chiu-Yin Kwan and Jean-Pierre Bourreau Department of Physiology, The Univerjity of Hong Kong, Hong Kong
S U M M A R Y
1. Muscarinic stimulation-induced tonic contraction of air- way smooth muscle is independent of membrane potential. This contraction is not sensitive to inhibition by voltage- operated CaZ+ channel blockers or by K' channel openers.
2. Cyclopiazonic acid (CPA) inhibits Ca2+ loading of internal stores but does not affect maximal tonic contraction induced by acetylcholine (ACh) in steady state conditions.
3. After depletion of internal Ca2+ stores with CPA, ACh- induced tonic contraction becomes dependent upon values of membrane potential. The contraction is then sensitive to voltage-operated Ca2+ channel blockers and to K+ channel openers.
4. Treatment of trachealis muscle with CPA potentiates the Mz-mediated component of ACh stimulation, but this poten- tiation is not entirely responsible for the switch in excitation- contraction (E-C) coupling.
5. It is proposed that depletion of internal Ca2+ stores with CPA and promotion of Mz-stimulation can lead to a switch in E-C coupling in trachealis smooth muscle from pharmaco- to electromechanical mode, perhaps by targeting a plasma mem- brane K+ channel.
Key words: airway smooth muscle, cyclopiazonic acid, di- hydropyridines, excitation Contraction coupling, internal Ca2+ stores, muscarinic stimulation, potassium channel opener.
INTRODUCTION
Cyclopiazonic acid (CPA) is an indole tetramic acid metabolite produced by Aspergillus and Penicillium (Fig. la, insert) and this mycotoxin is toxic to muscle.' Cyclopiazonic acid is a potent inhibitor of Ca2+/ Mg2+-ATPase activity and Ca2+ uptake in skeletal, cardiac and smooth muscle sarcoplasmic reticulum (SR).24 Its lack of effect on Na+/ K+-ATPase, H+/ K+-ATPase,
Correspondence: Dr J-P Bourreau, Department of Physiology, Faculty of Medicine, The University of Hong Kong, 5 Sassoon Road, Hong Kong.
Presented at the 7th Southeast Asian Western Pacific Regional Meeting of Pharmacologists, Manila, Philippines, November 1995.
Received 2 February 1996; accepted 15 February 1996.
mitochondria1 F1-ATPase, plasmalemma Ca2+/ Mg2+-ATPase,5 L-type Ca2+ channels and CaZ+-activated K+ channels6 makes it a potentially valuable tool to study the functional role of internal Ca2+ stores in excitation-contraction (E-C) coupling in muscles. In smooth muscle, CPA completely inhibits oxalate stimulated 45Ca2+ uptake in microsomal fractions enriched in SR.4 Cyclopiazonic acid, however, is variably efficient in pre- venting the loading of agonist-recruitable internal Ca2+ stores in multicellular smooth muscle preparations (Fig. 1 b)' and, unlike thapsigargin (another SR Ca2+/ MgZ+-ATPase inhibitor), it also decreases passive Ca2+ efflux from SR,8 which may complicate interpretation of its effect in functional studies. Despite, or perhaps because of, these limitations, CPA can be a useful tool to unmask the processes of E-C coupling in smooth muscle, as illustrated later with airway smooth muscle.
In the respiratory system, activity of airway smooth muscle modulates resistance to air flow and abnormal smooth muscle reactivity to stimuli is a major component in obstructive lung diseases. Unlike in vascular smooth muscle, contraction of airways induced by agonist stimulation (muscarinic) is not sensitive to organic Ca2+ entry blockers, perhaps as a result of the pharmacomechanical mode of coupling (i.e. independent of changes in membrane potential) between muscarinic stimulation and muscle contraction.9 The coupling mode between the stimulation of muscarinic cholinoceptors and airway muscle contraction can change. We have observed that after inhibition of the SR Ca2+-pumps with CPA, muscarinic E-C coupling in airway smooth muscle was switched from a pharmacomech- anical to an electromechanical mode. Tension development due to muscarinic stimulation became sensitive to inhibition by L-type Ca2+-entry blockers and potassium channels openers.'O
Muscarinic stimulation and contraction in trachealis smooth muscle Airway smooth muscles contract tonically in response to muscarinic stimulation. This contraction is associated with a biphasic change in cytosolic free Ca2+ concentration and with a graded membrane depolarization that often oscillates. Acti- vation of M3-muscarinic cholinoceptor subtypes by ACh stimulates phospholipase C, resulting in the formation of intracellular second messengers, inositol trisphosphate ( I h ) and diacylglycerol (DAG), which act, respectively, to release

734 D Amoako et al.
Ca2+ from internal IPS-sensitive stores (responsible for a phasic increase in cytosolic free Ca2+ concentration) and to change the sensitivity of the contractile machinery to Ca2+.11,L2 Release of Ca2+ from IP3-sensitive internal stores is transient and occurs immediately after muscle stimulation. It cannot, therefore, be responsible for the sustained tension observed during prolonged ACh stimulation and, in experiments performed in medium deprived of Ca2+, ACh stimulation leads only to transient contraction, which can be accounted for by the release of Ca2+ from internal stores.13 Influx of extracellular Ca2+ is required to maintain tone during stimulation.
0.6 1 * AC7 I
0 60 120 180 240 300 360 +e------- 0.4
Ca2+ entry during muscarinic stimulation in airway smooth muscle The route of CaZ+ entry into the cytosol in airway smooth muscle during muscarinic stimulation is multiple. L-type voltage-operated Ca*+ channels (VOCC) have been identified in airway muscles together with non-specific cation channels (NSCC) that Ca2+ can permeate and receptor-operated Ca2+ channels (ROCC).14p l5 VOCC activate at - 30 to - 15 mV and maximum current is seen at positive potentials. Steady-state inactivation curves have predicted that the probability of channel open time would be very low at a membrane potential as high as - 20 mV, but the slow depolarization induced by ACh stimulation can, however, activate these channels, leading to Ca2+ entry into the cell.l4'l6 Muscarinic stimulation de- polarizes airway smooth muscle plasma membrane (PM) from approximately - 60 mV to approximately - 30 mV. Acetyl- choline evokes large inward currents that result from activation of non-specific cation conductances and chloride conduc- tances.17 Activation of these currents is controlled by internal agonist-releasable Ca2+ stores and compounds that release Ca2+ from internal stores (caffeine, ryanodine) elicit inward currents similar to those induced by acetylcholine stimulation, while depletion of the stores with thapsigargin or CPA inhibits these currents.18 The physiological significance of this membrane depolarization has puzzled physiologists for many years, as voltage-dependent Ca2+ entry into cells marginally contributes to muscarinic stimulation-induced contraction. Blockade of VOCC with organic Ca2' channel blockers has little or no effect on the development of airway smooth muscle tone.148l9 Repolarizing pulses that bring back membrane potential to - _ -
Time (s ) resting values during ACh stimulation have no effect on tension.l6 Potassium channel openers, which repolarize the membrane during ACh stimulation to levels below the resting membrane potential have also marginal effect on tonic tension.10 Thus, airway smooth muscle uses a mode of coupling between muscarinic stimulation and contraction that is independent of membrane potential changes (pharmacomechanical coupling) and this particularity may be responsible for the lack of any clinical effect of Ca2+ channel blockers in the management of
25
0
Fig. 1. (a) Effect of cyclopiazonic acid (CPA; insert) on changes in the free cytosolic Ca2+ concentration induced by acetylcholine (ACh, 3 pmol/L) stimulation in canine trachealis. Changes in cytosolic free Ca2+ concentration were monitored in thin strip preparations obtained from canine trachealis incubated for 2 h at 37OC with lOpmol/L Fura 2 AM. Changes in free cytosolic Ca2' concentration are represented by changes in the ratio of fluorescence intensity obtained during excitation at 340 and 380nm (340/380). Preparations were incubated for 30 min with 30 pmol/L CPA. The open circle (0) indicates recordings obtained after CPA incubation. (b) Effect of 30pmol/L CPA on contractions induced by 100 grnol/ L ACh stimulation in bovine trachealis, in Ca2+- free and in Ca*+-containing solutions. Note the lack of effect of CPA on the maximal tension obtained in the Caz+-containing solution. (D), Ca*+-free control response; (W), f CPA in Caz+-free solution; (m), control response in Caz+-containing (1.25 mmol/ L) solution; (a), f CPA in Ca*+-containing solution.
most obstructive lung diseases.
What is the role of L-type Ca2+ channel activation in airway muscle contraction during muscarinic stimulation? According to the 'superficial buffer barrier' model proposed by van Breemen>o CaZT originating from entry across PM is in- volved in SR loading, which ultimately determines the magni- tude of IP3-induced Ca2+ release. The 'superficial buffer barrier' model explains the dependence of force development on the rate of Ca2+ influx rather than its magnitude as the fenestrated superficial SR, separated from the cell PM by a narrow restricted cytoplasmic space, partially buffers enhanced Ca2+ entry by active uptake. According to this model, we have proposed that Ca2+ influx via VOCC was used to restore or maintain intracellular, agonist recruitable Ca2+ stores.13 This proposal is supported by experiments in canine trachealis showing that blockade of voltage-dependent Ca2+ channels with nifedipine results in a decrease in the Ca2' content of internal stores, while exposure to the VOCC activator, Bay K
CPA and airway
150 -- A ?
0 Y
8 u) 100 --
2
v
C 0
0 .- I
I
5 0 -
muscle contraction 735
8644, increases the Ca2+ content of internal stores.7,13.18 These findings indicate that a true dihydropyridine-sensitive Ca2+ channel (rather than a non-specific effect of these compounds) is implicated in the Ca2+ homeostasis of these stores.21
Relationship between internal Ca2+ store loading and tonic muscle contraction Depletion of internal Ca*+ stores with CPA, or with ryanodine, which increases Ca2+ leak from stores, does not significantly affect tonic contraction of airway muscle in steady state condition^,^, ‘3 indicating that tonic contraction is not directly dependent upon the loading status of internal Ca2+ stores. However, prolonged exposure to nifedipine, which prevents loading and decreases the content of internal Ca2+ stores, progressively decreases tonic contraction of the muscle repeti- tively stimulated with ACh.22 This slowly developing inhibitory effect of nifedipine, which is absent when internal stores are fully loaded, is a direct consequence of decreased internal Ca2+ stores content on the sensitivity of tonic contraction to in- hibition by nifedipine. In airway smooth muscle, the less loaded internal CaZ+ stores are the more potent nifedipine is at inhibiting tonic contraction induced by muscarinic stimu- lation.’~ 10 Given the well established pharmacomechanical coupling mode of this stimulation (i.e. its lack of membrane potential dependency), it was believed at first that the nif- edipine-sensitive influx of Ca2+ following internal Ca2+ store depression was due to activation of a second messenger-acti-
0.7 i I v 0.6
.- - 0.5 I A C h
s 0.4)-+,- +-I- l- W 0 60 120 180 240 300 360 420 480 C W 0
Cro
0.6
0.5 * ACh
0.4 i--
0.3 4 I 0 120 240 360 480 600
Time (s)
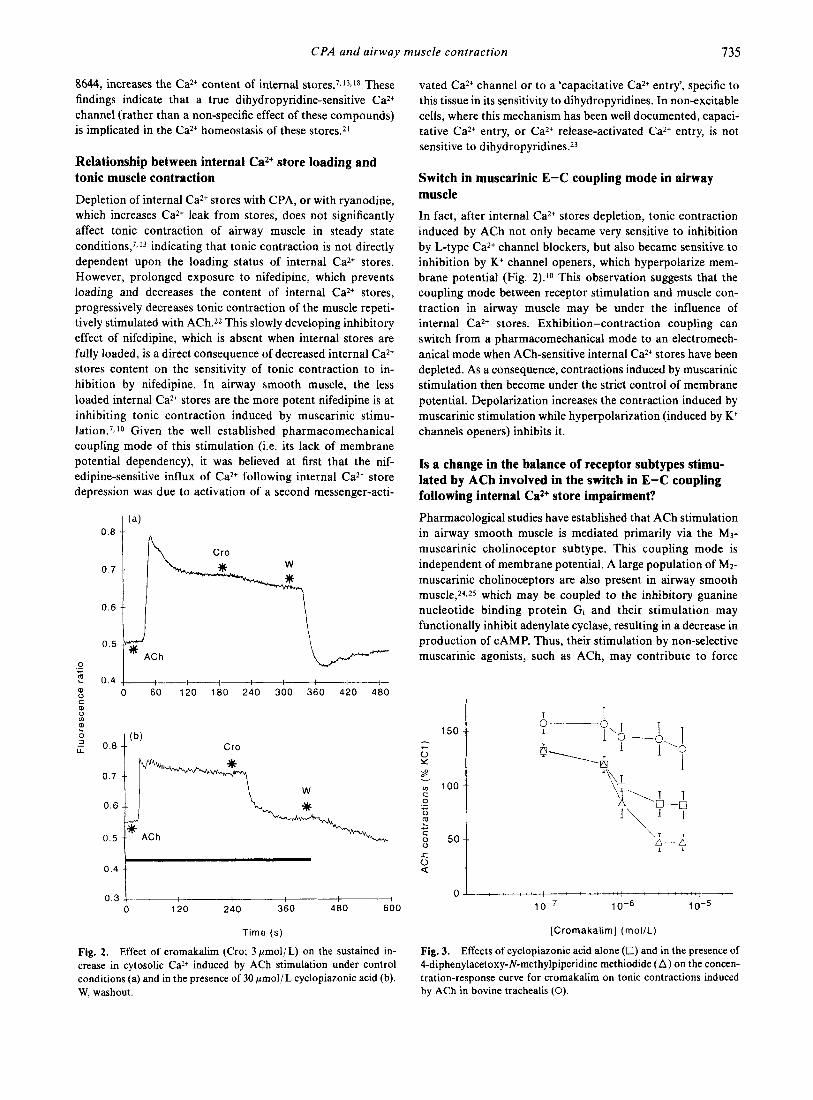
Fig. 2. Effect of cromakalim (Cro; 3 pmol/ L) on the sustained in- crease in cytosolic Ca2+ induced by ACh stimulation under control conditions (a) and in the presence of 30 pmol/L cyclopiazonic acid (b). W. washout.
vated Ca2+ channel or to a ‘capacitative Ca2+ entry’, specific to this tissue in its sensitivity to dihydropyridines. In non-excitable cells, where this mechanism has been well documented, capaci- tative Ca2+ entry, or Ca2+ release-activated Ca2+ entry, is not sensitive to dihydr~pyridines.~~
Switch in muscarinic E-C coupling mode in airway muscle In fact, after internal Ca2+ stores depletion, tonic contraction induced by ACh not only became very sensitive to inhibition by L-type Ca2+ channel blockers, but also became sensitive to inhibition by K+ channel openers, which hyperpolarize mem- brane potential (Fig. 2).10 This observation suggests that the coupling mode between receptor stimulation and muscle con- traction in airway muscle may be under the influence of internal Ca2+ stores. Exhibition-contraction coupling can switch from a pharmacomechanical mode to an electromech- anical mode when ACh-sensitive internal Ca2+ stores have been depleted. As a consequence, contractions induced by muscarinic stimulation then become under the strict control of membrane potential. Depolarization increases the contraction induced by muscarinic stimulation while hyperpolarization (induced by K+ channels openers) inhibits it.
Is a change in the balance of receptor subtypes stimu- lated by ACh involved in the switch in E-C coupling following. internal Ca2+ store impairment? Pharmacological studies have established that ACh stimulation in airway smooth muscle is mediated primarily via the M3- muscarinic cholinoceptor subtype. This coupling mode is independent of membrane potential. A large population of M2- muscarinic cholinoceptors are also present in airway smooth m u ~ c l e , ~ ~ . ~ ~ which may be coupled to the inhibitory guanine nucleotide binding protein Gi and their stimulation may functionally inhibit adenylate cyclase, resulting in a decrease in production of CAMP. Thus, their stimulation by non-selective muscarinic agonists, such as ACh, may contribute to force
8 0 Q
0 4 , : : : : : + : : : : : : : : ; : : : : . : : : : :
I 0-7 10-6 10-5
(Cromakalim] (mol /L)
Fig. 3. Effects of cyclopiazonic acid alone (0) and in the presence of 4-diphenylacetoxy-N-methylpiperidine methiodide (A) on the concen- tration-response curve for cromakalirn on tonic contractions induced by ACh in bovine trachealis (0).

736 D Arnoako et al.
development by decreasing the activity of Caz+-dependent K+- channels (channel activity is modulated by CAMP-dependent protein kinase). This would oppose activation of these channels by Ca2+ release from internal stores following agonist stimu- lation and contribute to agonist-induced depolarization leading to Ca2+ influx via activation of VOCC.26J7
We have tested whether decreasing the content of internal ACh-sensitive Ca2+ stores could unmask (or amplify) the M2 component of muscarinic stimulation, thus promoting an electromechanical mode of coupling. Under standard 'in vitro' conditions, M2-receptor antagonists have no significant effect on contractions induced by various muscarinic agonists. After depletion of internal stores with CPA, the potency of the Mj- receptor antagonist 4-DAMP to inhibit contractions of bovine trachealis induced by concentrations of ACh larger than 1 pmol /L decreased, while the potency of the M2-receptor antagonist gallamine increased.28 This amplification of the M2 component of muscarinic stimulation by ACh probably con- tributes to, but cannot fully account for, the switch in E-C coupling observed after CPA. Also, Fig. 3 shows that after CPA, E-C coupling can be switched further toward a n electromechanical mode by competition at the M3-receptor level. Thus, it is possible that the extent of phospholipase C activation following M3-receptor stimulation is a prime element controlling the E-C coupling mode. In this regard, Baba and coworkers29 have observed that in swine tracheal smooth muscle, inhibition of PIP2 hydrolysis following muscle stimu- lation with phorbol ester could induce a switch in E-C coupling from pharmaco- to electromechanical, similar to the switch we observed after CPA treatment.
CONCLUSION
Control of the E-C coupling mode in airway muscle during muscarinic stimulation is a complex process. The use of CPA has revealed that factors such as the status of internal Ca2+ stores and the stimulation of receptor subtypes can modulate the E-C coupling mode in trachealis muscle. The recognition of the microenvironment in smooth muscle and the role of large conductance, CaZ+-activated, outward rectifying, voltage- dependent K' channel activity in modulating vascular smooth muscle c0ntraction3~ suggests that these ionic channels, which are also present in airway may be the key element, targeted by these factors, responsible for determining the mode of E-C coupling in airways.
ACKNOWLEDGEMENTS
This work was supported by a grant from the Hong Kong Research Grant Council (grant 315/93M). The authors appreci- ate the skilful contribution of M r YW Wong in performing some of the experiments described.
REFERENCES
1 . Norred WP, Morrissey RE, Riley RT, Cole RJ, Dorner JW. Distribution, excretion and skeletal muscle effects of the mycotoxin ['4C]cyclopiazonic acid in rats. Food Chem. Toxicol. 1985; 23: 1069-76,
2. Goeger DE, Riley RT, Dorner JW, Cole RJ. Cyclopiazonic inhibition of Caz+-transport ATPase in rat skeletal muscle SR vesicles. Biochem. Pharmacol. 1988; 3 7 978-81.
3. Lahouratate P, Quiniou MJ, Leoty C. Effect of cyclopiazonic acid, an inhibitor of the CaZt-ATPase of sarcoplasmic reticulum, on CaZt transport, contraction and relaxation in cardiac muscle. Adv. Exp. Med. Bid. 1992; 311: 343-5.
4. Darby P, Kwan CY, Daniel EE. Use of calcium pump inhibitors in the study of calcium regulation in smooth muscle. Biol. Signals
5. Seidler NW, Jona I, Vegh M, Martonosi A. Cyclopiazonic acid is a specific inhibitor of the CaZ+-ATPase of sarcoplasmic reticulum. J. Biol. Chem. 1989; 264 17816-23.
6. Suzuki M, Muraki K, Imaizumi Y, Watanabe M. Cyclopiazonic acid, an inhibitor of the sarcoplasmic reticulum CaZ+-pump, reduces Caz+-dependent K+ currents in guinea pig smooth muscle. Br. J. Pharmacol. 1992; 107: 134-40.
7. Bourreau JP, Kwan CY, Daniel EE. Two distinct pathways for external Ca2+ to refill ACh-sensitive internal stores in canine airway smooth muscle. Am. J. Physiol. 1993; 265: C28-35.
8. Missiaen L, De-Smedt H, Droogmans G , Casteels R. 2,5-di-(tert- butyl)-1 ,4-Benzohydroquinone and cyclopiazonic acid decrease the Caz+ permeability of endoplasmic reticulum. Eur. J. Pharmacol.
9. Coburn RF, Baron CB. Coupling mechanisms in airway smooth muscle. Am. J. Physiol. 1990; 258: 11 19-33.
10. Bourreau J-P. Cross talk between plasma membrane and sarco- plasmic reticulum in canine airway smooth muscle. Biol. Signals
11. Chilvers ER, Barnes PJ, Nahorski SR. Characterization of agonist-stimulated incorporation of [3H]myo-inositol into inositol phospholipids and [3H]inositol phosphate formation in tracheal smooth muscle. Biochem. J. 1989; 262: 739-46.
12. Baron CB, Ring M, Coburn RF. Inositol lipids turnover and compartmentalization in canine trachealis smooth muscle. Am. J.
13. Bourreau J-P, Abela AP, Kwan CY, Daniel EE. Acetylcholine calcium stores refilling directly involves a dihydropyridine-sensitive channel in dog trachea. Am. J. Physiol. 1991; 261: C497-505.
14. Coburn RE Electromechanical coupling in canine trachealis muscle: Acetylcholine contraction. Am. J. Physiof. 1979; 236:
15. Murray RK, Kotlikoff MI. Receptor-activated calcium influx in human airway smooth muscle cells. J. Physiol. 1991; 435 123-44.
16. Daniel EE, Jury J, Serio R, Jager LP. Role of depolarization and calcium in contraction of canine trachealis from endogenous and exogenous acetylcholine. Can. J. Physiol. Pharmacol. 1991; 69:
17. Janssen L, Sims SM. Acetylcholine activates non-selective cation and chloride conductances in canine and guinea-pig tracheal myocytes. J. Physiol. 1992; 453: 197-218.
18. Janssen LJ, Sims SM. Emptying and refilling of CaZ+ stores in tracheal myocytes as indicated by ACh-evoked currents and contraction. Am. J. Physiol. 1993; 265: (377-86.
19. Coburn RF, Yamaguchi T. Membrane potential dependent and independent tension in canine tracheal smooth muscle. J. Phar- macol. Exp. Ther. 1977; 201: 276-84.
20. Chen Q , Cannel M, van Breemen C. The superficial buffer barrier in vascular smooth muscle. Can. J. Physiol. Pharmacol. 1992; 70:
21. Daniel EE, Bourreau J-P, Abela A, Jury J. The internal calcium store in airway muscle: Emptying, refilling and chloride. Biochem. Pharmacol. 1992; 43: 29-37.
22. Bourreau J-P, Kwan CY, Daniel EE. Excitation-contraction coupling in canine tracheal smooth muscle cells. In: Frank GB, Bianchi CP, ter Keurs HEDJ (eds). Excitation-contraction coupling in skeletal, cardiac and smooth muscle. Plenum Press, New York.
1993; 2 293-304.
1992; 227: 391-4.
1993; 2: 272-83.
Physiol. 1989; 256: C375-83.
C177-84.
518-25.
509-14.
1992; 375-7.

CPA and airway muscle contraction 737
23. Putney JW. Capacitative calcium entry revisited. Cell Calcium
24. Lucchesi E, Scheid CR, Romano FD. Ligand binding and G- protein coupling of muscarinic receptors in airway smooth muscle. Am. J. Physiol. 1990; 258: C730-8.
25. Misle AJ, Lippo De Becemberg I, Gonzales de Alfonzo R, Alfonzo MJ. Methocramine binding sites sensitive to alkylation on mus- carinic receptors from tracheal smooth muscle. Biochem. Phar- macol. 1992; 48: 191-5.
26. Hiramatsu T, Kume H, Kotlikoff MI, Takagi K. Role of calcium- activated potassium channels in the relaxation of tracheal smooth muscle by forskolin. Clin. Exp. Pharmacol. Physiol. 1994; 21:
1990; 11: 61 1-24.
361-15.
27. Kume H, Mikawa K, Takagi K, Kotlikoff MI. Role of G-proteins and KC. channels in the muscarinic and /3-adrenergic regulation of airway smooth muscle. Am. J. Physiol. 1995; 268: L221-9.
28. Amoako DK, Kwan CY, Bourreau JP. Switch in excitation-con- traction coupling in bovine trachealis: Due to a switch in muscarinic receptor subtypes stimulation? CAPS News Commun. 1995; 14: 51.
29. Baba K, Baron CB, Coburn R E Phorbol ester effects on coupling mechanisms during cholinergic contractions of swine tracheal smooth muscle. J. Physiol. 1989; 412 23-42.
30. Nelson MT, Cheng H, Rubart M, Santana LF, Bonev AD, Knot HJ, Lederer WJ. Relaxation of arterial smooth muscle by calcium sparks. Science 1995; 270 633-1.