printed symposium on replication of viral nucleic acids · activity by chloramphenicol suggests...
TRANSCRIPT

BACTERIOLOGICAL REVIEWS, June, 1966Copyright @ 1966 American Society for Microbiology
Symposium on Replication of Viral Nucleic AcidsII. Ribonucleic Acid Virus Replication'
LUCILLE SHAPIRO AND J. T. AUGUSTDepartment of Molecular Biology, Albert Einstein College of Medicine, Bronx, New York
INTRODUCTION................................................................ 279INDUCTION OF AN RNA VIRUS RNA POLYMERASE............................... 280PURIFICATION AND CHARACTERIZATION OF AN RNA VIRUS RNA POLYMERASE . ...... 281
Purification and Properties of the Enzyme...................................... 281Comparison Between RNA Virus RNA Polymerase and Host RNA Polymerase ...... 282RNA Product of the Reaction................................................. 282
POLYMERASE AcTIvITIEs ASSOCIATED WITH OTHER RNA VIRUSES ................... 284CONCLUSIONS................................................................ 285LITERATURE CITED............................................................ 285
INTRODUCTION
A cell infected with a ribonucleic acid (RNA)-containing virus has the unique capability ofproducing viral progeny from information en-coded in the RNA strand of the infecting par-ticle. The role of viral RNA in initiating its ownreplication is of major interest because of theinsight this special process may provide intomechanisms for transfer of genetic information.It is believed that RNA of the infecting virusserves both as messenger, to direct the synthesisof proteins required for phage multiplication,and as template, for replication of viral RNA.At present, two viral-specific synthetic functionsare recognized. The RNA genome must provideinformation for synthesis of coat protein, a
function demonstrated in vitro (40), and it hasbeen shown that a virus-specific enzyme, orseries of enzymes, is involved in the synthesis ofprogeny RNA (3, 4, 8, 25, 26, 58). Whereas thede novo synthesis of both protein and RNA arevital for these processes (12-18, 46), deoxy-ribonucleic acid (DNA) synthesis does not ap-pear necessary. Replication of RNA viruseswas not inhibited by mitomycin C, or fluori-nated pyrimidines (11, 49). Furthermore, tran-scription of bacterial DNA may not be requiredfor the replicative process, because a normalyield of the bacteriophage MS2 was obtainedfrom protoplasts treated with actinomycin (27).In these experiments, however, protoplasts wereprepared 5 min after infection, and actinomycinwas added 10 min later. Under these circum-stances, an early function requiring bacterial
1 A contribution to a symposium held at the An-nual Meeting of the American Society for Microbiol-ogy, Atlantic City, N.J., 29 April 1965, with JohnHolland as convener and Consultant Editor.
DNA could have escaped detection. There re-mains the possibility that some function in-volving bacterial DNA may influence viral RNAsynthesis. Evidence from several sources indi-cates that regulatory mechanisms similar tothose exerted on bacterial RNA synthesis affectRNA virus replication. The synthesis of bac-teriophage RNA appears to be controlled byamino acids in a manner similar to that of cellu-lar RNA and to be influenced by the RC locus,"RNA control gene," of the bacterial chromo-some (21). Phage RNA synthesis is also subjectto control by energy source "shift-down" (21).Another mechanism for the control of cellularRNA is the poorly understood process wherebyinfection with T-even bacteriophage inhibitsbacterial RNA synthesis (43). It was recentlyfound that infection by T-even phage inhibitsRNA phage replication as well (41; Leventhal,unpublished data). It is not known, however,whether this control operates at the level of hostDNA. Another observation was that infection ofultraviolet-treated bacteria by RNA phage re-sults in the accumulation of double-strandedviral RNA (41, 44). It was suggested that ahost-cell function which normally prevents theaccumulation of this double-stranded form wasdamaged by radiation (41). RNA phage replica-tion may not exhibit the autonomy of the T-evenphages which are capable of multiplying in bac-teria treated with ultraviolet light (2). From theseaccumulated data, it seems possible that the ul-timate expression of the message contained inthe entering viral RNA strand is not completelyindependent of some function involving bac-terial DNA.
Replication of viral RNA requires that theparental strand be used to specify the uniquesequence of ribonucleotides in progeny RNA.
279
Vol. 30, No. 2Printed in U.S.A.
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from

SHAPIRO AND AUGUST
RNA
U nATP
A nUTP
C nGTP
G 0CTP
Mg+ +
RNA
U rAMP
A UMP+
C GMP
G CMP]
PPi4n
FIG. 1. Predicted activity of RNA-dependent RNApolymerase.
One molecular mechanism for template-directednucleic acid synthesis involves specific hydrogenbonding between purine and pyrimidine bases asset forth in the Watson and Crick (54) modelof DNA structure. If the replication of single-stranded RNA viruses is also to involve hy-drogen-bonded base pairing, a nucleic acid ofcomplementary base composition must be syn-thesized. A possible alternative mechanism isthe direct synthesis of RNA identical to parentalRNA, eliminating the intervention of a comple-mentary polynucleotide (51). There have beenmany reports, however, of virus-specific RNAwith characteristics of a double-stranded struc-ture (7, 19, 30, 33, 39, 44, 56, 57). Parental virusRNA has been found in this form (18, 19, 32,53, 56), and double-stranded RNA was reportedto be infectious (1, 39, 45). There is also evidencethat replication of RNA in a reaction catalyzedby an RNA virus RNA polymerase occurs by atemplate mechanism utilizing specific hydrogenbonds between complementary base pairs (47,48). By analogy to the double-stranded DNAreplicative form of the bacteriophage cpX-174(50), it has been postulated that single-strandedRNA viruses require the formation of an inter-mediate for the formation of progeny RNA.Conclusive identification of the RNA directlyinvolved in replication of progeny viruses, aswell as a detailed description of secondarystructure, has not been presented, however.The series of events following RNA virus in-
fection logically suggest a requirement for RNA-mediated RNA replication and the synthesis ofat least one virus-specific RNA polymerase. Areaction utilizing RNA as template and ribo-nucleoside triphosphates as substrates is notknown to occur in vitro in normal uninfectedcells, whether animal, plant, or bacterial. Severalreports have indicated the presence of new RNApolymerase activity after RNA virus infection(3, 4, 8, 25, 26, 58). It is the purpose of this re-view to discuss the characteristics of this newenzyme(s) and to present data elucidating thebiochemical mechanism whereby parental single-
stranded RNA leads to the synthesis of manyidentical copies of RNA for progeny virus.
INDUCTION OF AN RNA VIRus RNA POLYMERAE
An RNA-dependent RNA polymerase couldbe predicted to act as outlined in Fig. 1. Thein vitro assay of such an enzymatic activity utilizesribonucleoside triphosphate substrates and iscarried out in the presence of RNA, Mg++,deoxyribonuclease, and phosphate buffer. Thedeoxyribonuclease is included to inhibit DNA-dependent RNA polymerase activity, and phos-phate buffer excludes polynucleotide phos-phorylase activity. Under these conditions, invitro ribonucleotide incorporation was foundto increase after infection with RNA viruses(Fig. 2). The inhibition of this RNA polymeraseactivity by chloramphenicol suggests that syn-thesis of a new protein is required for theexpression of this activity (36). New RNA poly-merase activity has been found after mengovirusinfection of L cells (8); bacteriophage MS2(25, 58), f2 (4), and Q3 (26) infection of Escher-ichia coli; and turnip yellow mosaic virus in-fection of Chinese cabbage leaves (3). It is gen-erally assumed that this new activity is that ofan RNA-dependent RNA polymerase inducedby virus infection. It is also possible, however,that the activity present in crude extracts, inwhole or in part, is that of the host RNA poly-merase utilizing a new type of RNA as template.
c
1, 0.8
E
0 0.6
a-
°0.CL0-00
.* 0.2iLa.CD
Time after infection (min)FIG. 2. Effect of RNA phage infection on ribonu-
cleotide incorporation. Alumina extracts were preparedfrom Escherichia coli collected before and after infec-tion with phage f2 at intervals as shown on the abscissa.The incorporation ofguanosine inonophosphate (GMP)was measured as described in Table 1.
280 BAcrERIOL. REv.
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RNA VIRUS REPLICATION
TABLE 1. Requirements for guanosinemonophosphate (GMP) incorporation*
GMP in-corooa
Systemt tora-(pumoles/
5 min)
Complete............................... 72.5Minus MgC2 ......................... 3.2Minus ribonucleic acid................ 6.6Minus CTP, UTP, and ATP........... 2.5Minus GTP, CTP, UTP, and ATP: plus
[C14] dGTP, dCTP, dTTP, and dATP. 1.0Minus GTP, CTP, UTP, and ATP: plus
[C'4] GDP, CDP, UDP, and ADP... 1.9Minus ribonucleic acid and deoxyribo-
nuclease; plus thymus deoxyribonu-cleic acid........................... 10.1
Minus deoxyribonuclease; plus ribo-nuclease (1.0,ug).................... 0.8
Minus deoxyribonuclease ...... ....... 71.6Plus actinomycin D (25,ug)........... 74.1
* The ribonucleic acid (RNA) virus RNA poly-merase assay measured the rate of incorporationof radioactivity from a labeled ribonucleosidetriphosphate into an acid-insoluble form (6).The standard reaction mixture (0.2 ml) contained25 ,moles of potassium phosphate buffer, pH7.0; 5,moles of,-mercaptoethanol; 2,moles ofMgCl2; 20 m,umoles of [C'4]-GTP (8.0 X 106 countsper min per umole); 50 m,umoles each of ATP,CTP, and UTP; 300 m,umoles of Escherichia coliR-RNA; 5 ,ug of pancreatic deoxyribonuclease;and 3 ,ug of protein of the enzyme fraction.
f Abbreviations: CTP, cytidine triphosphate;UTP, uridine triphosphate; ATP, adenosine tri-phosphate; GTP, guanosine triphosphate; dGTP,deoxyguanosine triphosphate; CDP, cytidinediphosphate; dTTP, deoxythymidine triphos-phate; dATP, deoxyadenosine triphosphate; GDP,guanosine diphosphate; UDP, uridine diphos-phate; ADP, adenosine diphosphate.
It is not known whether synthesis of progenyRNA is entirely the function of a new virus-in-duced enzyme(s). It is possible that, once a newenzyme catalyzes the synthesis of RNA comple-mentary to the parental strand, a replicative formserves as template for the host RNA polymerase.It has been reported that double-stranded RNAof reovirus serves as primer for the normallyDNA-dependent RNA polymerase, in vitro (24,35). Precise identification of the RNA-dependentpolymerase activity in crude extracts of virus-infected cells must await clarification of the bio-chemical details of viral RNA synthesis.
PURIFICATION AND CHARACTERIZATION OF ANRNA VIRUs RNA POLYMERASE
One approach to understanding the biochemi-cal mechanism of RNA virus multiplication is
to purify and characterize the virus-inducedpolymerase enzyme(s). Purification of one RNAvirus RNA polymerase was facilitated by use of amutant phage, f2 su-11. Bacteriophage su-11produces viable progeny only in bacteria con-taining a suppressor gene. In E. coli K38, astrain lacking a suppressor gene, su-11 does notproduce progeny, either viable or defective (36).However, the RNA virus RNA polymerase ac-tivity in extracts of K38 infected with su-11 isabout five times greater than normal, and an ex-cess of phage-specific RNA is produced.
Purification and Properties of the EnzymeThe RNA virus RNA polymerase, isolated
from E. coli K38 infected with f2 su-1 1, has beenpurified approximately 100-fold (6). RNA syn-thesis catalyzed by the partially purified enzymewas dependent on the presence of Mg++, RNA,and the four ribonucleoside triphosphates (Table1). The reaction did not occur when manganousions replaced magnesium ions or when DNA re-placed RNA; nor did deoxyribonucleotides orribonucleoside diphosphates replace the ribo-nucleoside triphosphates. Ribonucleoside tri-phosphate dependence was demonstrated evenwhen adenosine triphosphate (ATP) was thelabeled substrate, indicating the absence ofpolyriboadenylate polymerase activity (5). En-zymatic synthesis of homopolymers with anyof the ribonucleoside triphosphates as substratewas never observed. The reaction was completelyinsensitive to deoxyribonuclease, but was in-hibited by 0.1 gg/ml of ribonuclease. Actinomy-cin D, in concentrations as high as 100 ,g/ml,did not inhibit the reaction; by comparison,DNA-primed synthesis of RNA was inhibitedby 1 to 4 ,ug/ml of actinomycin D (23, 28).The purified enzyme was dependent upon the
addition of RNA for activity. RNA from manydifferent sources satisfied this requirement. Prepa-rations of bacterial soluble and ribosomal RNAand viral RNA were equally effective in stimu-lating the reaction catalyzed by the RNA virusRNA polymerase isolated from cells infectedwith f2 su-11 (Table 2). Synthetic polyribonu-cleotides, either homopolymers or copolymers,did not satisfy the RNA requirement for eithercomplementary or noncomplementary incor-poration of ribonucleotides. Changes in thesecondary structure of the synthetic polymerswere without effect; polyadenylic acid pluspolyuridylic acid and polyinosinic acid plus poly-cytidylic acid were ineffective as primers.
In the presence of a high concentration ofinorganic pyrophosphate, the RNA virus RNApolymerase catalyzed the incorporation of pyro-phosphate into ribonucleoside triphosphates
VOL. 30, 1966 281
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from

SHAPIRO AND AUGUST
TABLE 2. RNA requiremnent for guaniosiniemonophosphate (GMP) incorpora tionl*
G.MP in-RNA added corporation
(AAsmoles/5min)
None 8f2. .... 103TMV 1................................. 106Es/eieric/ia coli B ribosomal ........... 104Streptococculs ftiecalis ribosomal ........ 101Micrococelcs /ysodeikticuts ribosomal 108Clos tridilnti pasteurianuin ribosomal .. 107E. coli B soluble 101E. coli W soluble ...................... 105S. faecalis soluble .................... 105
* The standard assay procedure was used withC14-guanosine triphosphate (8.2 X 101 countsper min per ,umole) and 5 ,ug of enzyme protein ineach tube, except that E. coli R-RNA was omitted.Different RNA preparations were added as indi-cated, by use of 200 to 3Co m,umoles of each prepa-ration.
(6). This is in accord with an enzymatic mecha-nism utilizing nucleoside triphosphates as sub-strate (10, 22). Inorganic pyrophosphate (PP1)exchange required the presence of ribonucleosidetriphosphates, Mg++ and RNA.
Comparison Betueen RNA Virus RNA Pol4yneracseand Host RNA Polymnerase
Both the RNA virus RNA polymerase andthe DNA-dependent RNA polymerase catalyzethe synthesis of RNA. There are several obvioussimilarities in the details of the reactions, butthere are also significant differences (Table 3).The outstanding difference is in the nature ofthe template utilized in the reaction. The RNAvirus RNA polymerase will utilize RNA butnot DNA as template, whereas single-strandedRNA is a poor template in the normally DNA-
dependent RNA polymerase reaction (52). How-ever, the DNA-dependent enzyme will utilizeribohomopolymer templates very efficiently andwill also catalyze ribohomopolymer synthesis(20, 34, 42). In contrast, the RNA virus RNApolymerase appears to be more specific, for itwill utilize only natural RNA as template anddoes not catalyze homopolymer synthesis (6).Furthermore, the viral polymerase purified fromcells infected with f2 su-11 is insensitive to anti-serum which specifically inactivates the hostRNA polymerase (Shapiro and August, un-
published dlata). This characterization of thepartially purified RNA virus RNA polymeraseappears to establish the fact that at least one new
enzyme is involved in the replication of the RNAbacteriophage f2.
RNA Procict of'dhe Reaction
The accumulated evidence supports the con-
clusion that an intermediate or "replicative form"of RNA may be involved in the multiplicationof single-stranded RNA viruses (7, 19, 30, 33,37, 39, 44, 56). Thus, the enzyme(s) associatedwith the replication of vir-al RNA may be in-volved either in the formation of a replicativeform or in the use of the replicative form duringsynthesis of progeny RNA, or both. With theassumption that the formation of hydrogen-bonded base pairs occurs in these processes, bothsteps would require the synthesis of a comple-mentary copy of the RNA directing the reaction.Further elucidation of the role, in these processes,
of the viral RNA polymerase induced by f2
su-11 infection has been obtained (i) by analysisof the base composition of the RNA product, (ii)by studies of base analogue substitution, and(iii) by characterization of some of the physicalproperties of the RNA product of the reaction.The base composition of the RNA preduct was
determined by the technique of nearest neighbor
TABLE 3. Comi1parisonl ofRNA polymerase eiizymues
Characteristic of the reaction Xiral RNA-dependent DNA-dependentRNA p)olymerase R\A polymea.se
Substrates .......................... ATP, UTP, GTP, CTP* ATP, UTP, GTP, CTP*Metal ion ..........................mg Mn++ (Mg++)DNA requirement .......... No YesRNA requirement.................. Yes NoRibohomopolymer as template No YesRibohomopolymer synthesis No YesPPi exchange ...................... Yes YesActinomycin inhibition ............. No YesSensitive to RNA polymerase anti-serum .. No Yes
* See second footnote to Table 1.
282 BACTERIOL. REV.
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from
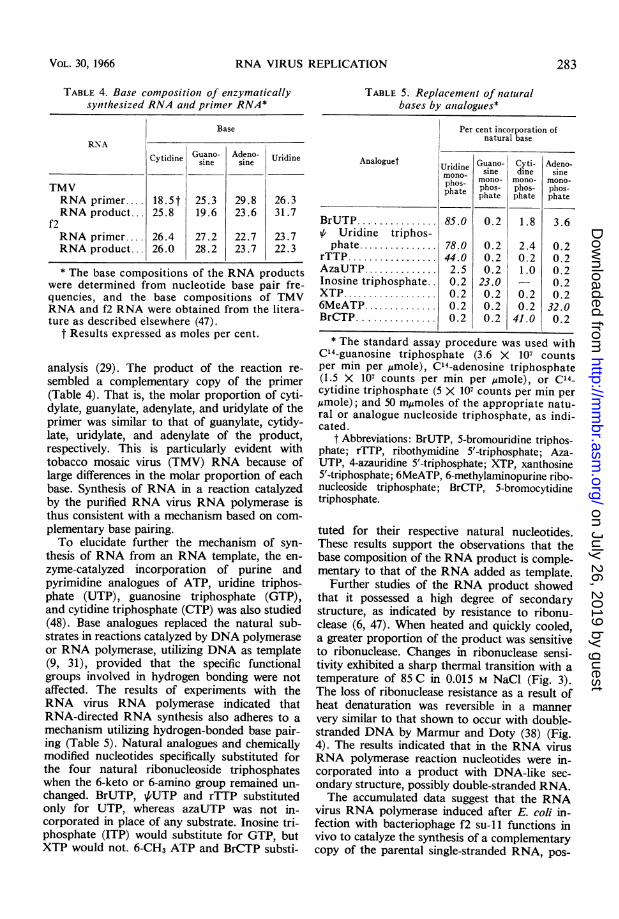
RNA VIRUS REPLICATION
TABLE 4. Base compositionz of enzymaticallysynthesized RNA antd primer RNA*
Base
RNACytidine Guano- Adeno- UridineCyiiesine sine
TMVRNA primer.... 18.5t 25.3 29.8 26.3RNA product... 25.8 19.6 23.6 31.7
f2RNA primer.... 26.4 27.2 22.7 23.7RNA product... 26.0 28.2 23.7 22.3
* The base compositions of the RNA productswere determined from nucleotide base pair fre-quencies, and the base compositions of TMVRNA and f2 RNA were obtained from the litera-ture as described elsewhere (47).
t Results expressed as moles per cent.
analysis (29). The product of the reaction re-sembled a complementary copy of the primer(Table 4). That is, the molar proportion of cyti-dylate, guanylate, adenylate, and uridylate of theprimer was similar to that of guanylate, cytidy-late, uridylate, and adenylate of the product,respectively. This is particularly evident withtobacco mosaic virus (TMV) RNA because oflarge differences in the molar proportion of eachbase. Synthesis of RNA in a reaction catalyzedby the purified RNA virus RNA polymerase isthus consistent with a mechanism based on com-plementary base pairing.To elucidate further the mechanism of syn-
thesis of RNA from an RNA template, the en-zyme-catalyzed incorporation of purine andpyrimidine analogues of ATP, uridine triphos-phate (UTP), guanosine triphosphate (GTP),and cytidine triphosphate (CTP) was also studied(48). Base analogues replaced the natural sub-strates in reactions catalyzed by DNA polymeraseor RNA polymerase, utilizing DNA as template(9, 31), provided that the specific functionalgroups involved in hydrogen bonding were notaffected. The results of experiments with theRNA virus RNA polymerase indicated thatRNA-directed RNA synthesis also adheres to amechanism utilizing hydrogen-bonded base pair-ing (Table 5). Natural analogues and chemicallymodified nucleotides specifically substituted forthe four natural ribonucleoside triphosphateswhen the 6-keto or 6-amino group remained un-changed. BrUTP, /6UTP and rTTP substitutedonly for UTP, whereas azaUTP was not in-corporated in place of any substrate. Inosine tri-phosphate (ITP) would substitute for GTP, butXTP would not. 6-CH3 ATP and BrCTP substi-
Analoguet
BrUTP...............VI Uridine triphos-phate...............
rTTP.................AzaUTP..............Inosine triphosphate..XTP..................6MeATP..............BrCTP...............
Per cent incorporation ofnatural base
Uridine Guano- Cyti- Adeno-monno- sine dine sine
phos-phate
85.0
78.044.02.50.20.20.20.2
mono-phos-phate
0.2
0.20.20.223.00.20.20.2
mono-phos-phate
1.8
2.40.21.0
0.20.241.0
mono-phos-phate
3.6
0.20.20.20.20.232.00.2
* The standard assay procedure was used withC'4-guanosine triphosphate (3.6 X 107 countsper min per umole), C14-adenosine triphosphate(1.5 X 107 counts per min per ,umole), or C14-cytidine triphosphate (5 X 107 counts per min per,umole); and 50 myrmoles of the appropriate natu-ral or analogue nucleoside triphosphate, as indi-cated.
t Abbreviations: BrUTP, 5-bromouridine triphos-phate; rTTP, ribothymidine 5'-triphosphate; Aza-UTP, 4-azauridine 5'-triphosphate; XTP, xanthosine5'-triphosphate; 6MeATP, 6-methylaminopurine ribo-nucleoside triphosphate; BrCTP, 5-bromocytidinetriphosphate.
tuted for their respective natural nucleotides.These results support the observations that thebase composition of the RNA product is comple-mentary to that of the RNA added as template.
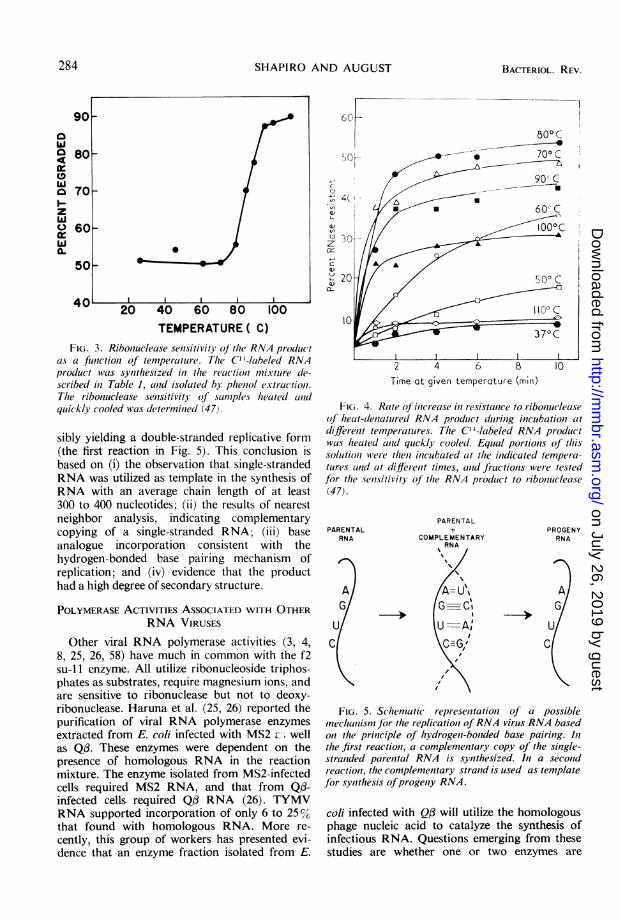
Further studies of the RNA product showedthat it possessed a high degree of secondarystructure, as indicated by resistance to ribonu-clease (6, 47). When heated and quickly cooled,a greater proportion of the product was sensitiveto ribonuclease. Changes in ribonuclease sensi-tivity exhibited a sharp thermal transition with atemperature of 85 C in 0.015 M NaCl (Fig. 3).The loss of ribonuclease resistance as a result ofheat denaturation was reversible in a mannervery similar to that shown to occur with double-stranded DNA by Marmur and Doty (38) (Fig.4). The results indicated that in the RNA virusRNA polymerase reaction nucleotides were in-corporated into a product with DNA-like sec-ondary structure, possibly double-stranded RNA.The accumulated data suggest that the RNA
virus RNA polymerase induced after E. coli in-
fection with bacteriophage f2 su-l functions invivo to catalyze the synthesis of a complementarycopy of the parental single-stranded RNA, pos-
TABLE 5. Replacement of niaturalbases by anIalogues*
-~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
VOL. 30, 1966 283
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from
SHAPIRO AND AUGUST
900w0
CD
0
z
(Lu
80k
70F
60k
50
20 40 60 80 100
TEMPERATURE( C)
FIG. 3. Riboniuclease sentsitivity of' the RNA produic t
as5 a funictioIn of temperature. The C11 labeled RNApr-oduct was synlthesized in the reaction mixture dte-scribed in Table 1, and isolated by phentol extractioni.Tlhe ribonuclease sensitivity of salflpleAs heated anldquickly cooled was determinied (47).
sibly yielding a double-stranded replicative form(the first reaction in Fig. 5). This conclusion isbased on (i) the observation that single-strandedRNA was utilized as template in the synthesis ofRNA with an average chain length of at least300 to 400 nucleotides; (ii) the results of nearestneighbor analysis, indicating complementarycopying of a single-stranded RNA; (iii) baseanalogue incorporation consistent with thehydrogen-bonded base pairing mechanism ofreplication; and (iv) evidence that the producthad a high degree of secondary structure.
POLYMERASE ACTIVITIEs ASSOCIATED WITH OTHERRNA VIRUSES
Other viral RNA polymerase activities (3, 4,8, 25, 26, 58) have much in common with the f2
su-11 enzyme. All utilize ribonucleoside triphos-phates as substrates, require magnesium ions, andare sensitive to ribonuclease but not to deoxy-ribonuclease. Haruna et al. (25, 26) reported thepurification of viral RNA polymerase enzymesextracted from E. coli infected with MS2 wellas Qj. These enzymes were dependent on thepresence of homologous RNA in the reactionmixture. The enzyme isolated from MS2-infectedcells required MS2 RNA, and that from Qf-infected cells required Q3 RNA (26). TYMVRNA supported incorporation of only 6 to 25%/othat found with homologous RNA. More re-
cently, this group of workers has presented evi-dence that an enzyme fraction isolated from E.
60
800 c
a)c
a)
c)zOf
ca1)ui
Time at given temperature (min)
FIG. 4. Rate oJ inlcri-ease in esistanice to ribonucleaseof heat-dtiac-tured RNA produict durinig inicubationi at
different temperatulres. The C'4-labeled RNA productwas heated anid quckly cooled. Equial portionis of thissolutioni were theln inicubated at the inidicated tempera-
tures acnd at difl rent times, anid fiactionis wer e testedfor the selsitivity of the RNA pr-oduct to riboniuclease(47).
PARENTALRNA
A
G
UlC
PROGENYRNA
A|G
UlCt
FIG. 5. Schematic representation of a possiblemechaiisni for the replicationt ofRNA virus RNA basedon the priniciple of hiydrogeni-bonided base pairinlg. In
the first reactioni, a complemenitary copy of the sinigle-stranided parenital RNA is synzthesized. In a seconidreactioll, the complementary stranid is used as templatefor synthesis ofprogeniy RNA.
coli infected with Qf3 will utilize the homologousphage nucleic acid to catalyze the synthesis ofinfectious RNA. Questions emerging from thesestudies are whether one or two enzymes are
.
284 BACTERIOL. REV.
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RNA VIRUS REPLICATION
present in the purified enzyme fraction andwhether a polynucleotide complementary to theviral RNA exists in the reaction mixture. Furtherelucidation of the enzymatic mechanism and theRNA products of the MS2 and Qj3 reactions willpermit a clearer analysis of the RNA requirementexhibited by these enzymes and of the viral poly-merase isolated from E. coli infected by f2 su-1 1.
Activity from another enzyme isolated fromMS2-infected E. coli has been studied by Weiss-mann and his co-workers (51, 55, 58). This en-zyme was purified in association with endogenousRNA and in this state appeared to catalyze thesynthesis of progeny RNA (57). It was proposedthat the enzyme, as isolated, utilized the replica-tive form as template and yielded progeny RNAvia a semiconservative mechanism of replication(55). It is as yet unknown whether this enzymehas properties in common with the other enzymesisolated from cells infected with f2 su-1 1, MS2,or Q,B.
CONCLUSIONS
The evidence presented favors the conceptthat replication of single-stranded viral RNAproceeds by the known mechanism of base pair-ing, requiring the synthesis of RNA with a basecomposition complementary to that of parentalRNA (Fig. 5). Once formed, the complement ofthe entering viral strand would act as templatefor the synthesis of progeny RNA by means,perhaps, of asymmetric semiconservative repli-cation. Whether a single virus-induced polymeraseor two different enzymes catalyze the synthesisof viral complement and progeny is a matter ofspeculation at this time. Although much has beenlearned about the replication of the small RNAviruses, it is apparent that there are many detailsof this mechanism that remain to be elucidated.
ACKNOWLEDGMENTSPortions of this work were supported by grants
from the National Institutes of Health (1-TI-GM-1191-02, GM-11936-03, and GM-11301-04) and theNational Science Foundation (GB-2073), and by theHealth Research Council of the City of New Yorkunder Contract No. 1-211.We wish to thank J. Marmur for his interest and
valuable contributions to the preparation of thismanuscript.
LITERATURE CITED1. AMMANN, J., H. DELIUS, AND P. H. HOFSCHNEI-
DER. 1964. Isolation and properties of an intactphage-specific replicative form of RNA phageM12. J. Mol. Biol. 10:557-561.
2. ANDERSON, T. F. 1948. The growth of T2 viruson ultraviolet-killed host cells. J. Bacteriol.56:403-410.
3. ASTIER-MANIFACIER, S., AND P. CORNUET. 1964.Replication de l'acide ribonucleique du virusde la mosaique jaune du Nanet. Etude d'uneRNA polymerase. Compt. Rend. 259:4401-4404.
4. AUGUST, J. T., S. COOPER, L. SHAPIRO, AND N.D. ZINDER. 1963. RNA phage-induced RNApolymerase. Cold Spring Harbor Symp. Quant.Biol. 28:95-97.
5. AUGUsr, J. T., P. J. ORTIZ, AND J. HURWITZ.1962. Ribonucleic acid-dependent ribonucleo-tide incorporation. J. Biol. Chem. 237:3786-3793.
6. AUGUST, J. T., L. SHAPIRO, AND L. EOYANG.1965. Replication of RNA viruses. I. Char-acterization of a viral RNA-dependent RNApolymerase. J. Mol. Biol. 11:257-271.
7. BALTIMORE, D., Y. BECKER, AND J. E. DARNELL.1964. Virus-specific double-stranded RNA inpoliovirus-infected cells. Science 143:1034-1036.
8. BALTIMORE, D., AND R. FRANKLIN. 1963. A newribonucleic acid polymerase appearing afterMengovirus infection of L-cells. J. Biol. Chem.238:3395-3400.
9. BESSMAN, M. J., I. R. LEHMAN, J. ADLER, S. ZIM-merman, E. S. Simms, AND A. KORNBERG.1958. Enzymatic synthesis of deoxyribonucleicacid. III. The incorporation of pyrimidine andpurine analogues into deoxyribonucleic acid.Proc. Natl. Acad. Sci. U.S. 44:633-640.
10. BESSMAN, M. J., I. R. LEHMAN, E. S. SIMMS, ANDA. KORNBERG. 1958. Enzymatic synthesis ofdeoxyribonucleic acid. II. General propertiesof the reaction. J. Biol. Chem. 233:171-177.
11. COOPER, S., AND N. D. ZINDER. 1962. The growthof an RNA bacteriophage: the role of DNAsynthesis. Virology 18:405-411.
12. COOPER, S., AND N. D. ZINDER. 1963. The growthof an RNA bacteriophage: the role of proteinsynthesis. Virology 20:605-612.
13. DARNELL, J. E., AND L. LEVINTOW. 1960. Polio-virus protein: source of amino acids and timecourse of synthesis. J. Biol. Chem. 235:74-77.
14. DAVIS, J. E., AND R. L. SINSHEIMER. 1963. Thereplication of bacteriophage MS2. I. Transferof parental nucleic a-id to progeny phage. J.Mol. Biol. 6:203-207.
15. DELIUS, H., AND P. H. HOFSCHNEIDER. 1964. Twoeffects of inhibition of protein synthesis on thereplication of M12 bacteriophage RNA. J.Mol. Biol. 10:554-556.
16. DoI, R. H., AND S. SPIEGELMAN. 1963. Conserva-tion of a viral RNA genome during replicationand translation. Proc. Natl. Acad. Sci. U.S.49:353-360.
17. ELLIS, D. B., AND W. PARANCHYCH. 1963. Syn-thesis of ribonucleic acid and protein in bac-teria infected with an RNA bacteriophage. J.Cellular Comp. Physiol. 62:207-213.
18. ERIKSON, R. L., M. L. FENWICK, AND R. M.FRANKLIN. 1964. Replication of bacteriophageRNA: studies on the fate of parental RNA. J.Mol. Biol. 10:519-529.
VOL. 30, 1966 285
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from

SHAPIRO AND AUGUST
19. ERIKSON, R. L., M. L. FENWICK, AND R. M.FRANKLIN. 1965. Replication of bacteriophageRNA: some properties of the parental-labeledreplicative intermediate. J. Mol. Biol. 13:399-406.
20. Fox, C. F., W. S. ROBINSON, R. HASELKORN, ANDS. B. WEISS. 1962. Enzymatic synthesis of ribo-nucleic acid. III. The ribonucleic acid-primedsynthesis of ribonucleic acid with Micrococcuslysodeikticus ribonucleic acid polymerase. J.Biol. Chem. 239:186-195.
21. FRIESEN, J. D. 1965. Control of bacteriophageRNA synthesis in E. coli. J. Mol. Biol. 13:220-233.
22. FURTH, J. J., J. HURWITZ, AND M. ANDERS. 1962.The role of deoxyribonucleic acid in ribonu-cleic acid synthesis. I. The purification andproperties of RNA polymerase. J. Biol. Chem.237:2611-2619.
23. GOLDBERG, 1. H., M. RABINOWITZ, AND E. REICH.1962. Basis of actinomycin action. I. DNAbinding and inhibition of RNA polymerasesynthetic reactions by actinomycin. Proc.Natl. Acad. Sci. U.S. 48:2094-2101.
24. GOMATOS, P. J., R. M. KRUG, AND I. TAMM. 1964.Enzymic synthesis of RNA with reovirus RNAas template. I. Characteristics of the reactioncatalyzed by the RNA polymerase fromEscherichia coli. J. Mol. Biol. 9:193-207.
25. HARUNA, I., K. Nozu, Y. OHTAKA, AND S. SPIE-GELMAN. 1963. An RNA "replicase" inducedby and selective for a viral RNA: isolationand properties. Proc. Natl. Acad. Sci. U.S.50:905-911.
26. HARUNA, I., AND S. SPIEGELMAN. 1965. Specifictemplate requirements of RNA replicases.Proc. Natl. Acad. Sci. U.S. 54:579-587.
27. HAYWOOD, A. M., AND R. L. SINSHEIMER. 1963.Inhibition of protein synthesis of E. coli proto-plasts by actinomycin D. J. Mol. Biol. 6:247-249.
28. HURW1TZ, J., J. J. FURTH, M. MALAMY, AND M.ALEXANDER. 1962. The role of deoxyribonu-cleic acid in ribonucleic acid synthesis. III.The inhibition of the enzymatic synthesisof ribonucleic acid and deoxyribonucleicacid by actinomycin D and proflavin. Proc.Natl. Acad. Sci. U.S. 48:1222-1230.
29. JOSSE, J., A. D. KAISER, AND A. KORNBERG. 1961.Enzymatic synthesis of deoxyribonucleic acid.VIII. Frequencies of nearest neighbor basesequences in deoxyribonucleic acid. J. Biol.Chem. 236:864-875.
30. KAERNER, H. C., AND H. HOFFMANN-BERLING.1964. Synthesis of double-stranded RNA inRNA-phage infected E. coli cells. Nature 202:1012-1013.
31. KAHAN, F., AND J. HURWITZ. 1962. The role ofdeoxyribonucleic acid in ribonucleic acid syn-thesis. IV. The incorporation of pyrimidineand pure analogues into ribonucleic acid. J.Biol. Chem. 237:3778-3785.
32. KELLY, R. B., J. L. GOULD, AND R. L. SINSHEI-
MER. 1965. The replication of bacteriophageMS2. IV. RNA components specifically associ-ated with infection. J. Mol. Biol. 11:562-575.
33. KELLY, R. B., AND R. L. SINSHEIMER. 1964. Anew RNA component in MS2-infected cells. J.Mol. Biol. 8:602-605.
34. KRAKOW, J. S., AND S. OCHOA. 1963. Ribonucleicacid polymerase of Azotobacter vinelandii. I.Priming by polyribonucleotides. Proc. Natl.Acad. Sci. U.S. 49:88-94.
35. KRUG, R. M., P. J. GOMATOS, AND I. TAMM. 1965.Enzymic synthesis of RNA with reovirus RNAas template. II. Nearest neighbor analysis ofthe products of the reaction catalyzed by theE. coli RNA polymerase. J. Mol. Biol. 12:872-880.
36. LODISH, H. E., S. COOPER, AND N. D. ZINDER.1964. Host-dependent mutants of bacteriophagef2. IV. On the biosynthesis of a viral RNApolymerase. Virology 24:60-70.
37. MANDEL, H. G., R. E. F. MATTHEWS, A. MATUS,AND R. K. RALPH. 1964. Replicative form ofplant viral RNA. Biochem. Biophys. Res.Commun. 16:604-609.
38. MARMUR, J., AND P. DOTY. 1961. Thermal re-naturation of deoxyribonucleic acid. J. Mol.Biol. 3:585-594.
39. MONTAGNIER, L., AND F. K. SANDERS. 1963.Replicative form of encephalomyocarditis virusribonucleic acid. Nature 199:664-667.
40. NATHANS, D., G. NOTANI, J. H. SCHWARTZ, ANDN. D. ZINDER. 1962. Biosynthesis of the coatprotein of coliphage f2 by E. coli extracts. Proc.Natl. Acad. Sci. U.S. 48:1424-1431.
41. NEUBAUER, Z., AND ZAVADA, V. 1965. The ca-pacity of the bacterial host for the reproductionof the RNA phage f2. Biochem. Biophys. Res.Commun. 20:1-3.
42. NIYoGI, S. K., AND A. STEVENS. 1965. Studies ofthe ribonucleic acid polymerase from E. coli.III. Studies with synthetic polyribonucleotidesas templates. J. Biol. Chem. 240:2587-2592.
43. NOMURA, M., K. MATSUBARA, K. OKAMOTO, R.FUJIMURA. 1962. Inhibition of host nucleic acidand protein synthesis by bacteriophage T4: itsrelationship to the physical and functional in-tegrity of host chromosomes. J. Mol. Biol.5:535-549.
44. NONOYAMA, M., AND Y. IKEDA. 1964. Ribonu-clease-resistant RNA found in cells of Escher-ichia coli infected with RNA phage. J. Mol.Biol. 9:763-771.
45. PONS, M. 1964. Infectious double-stranded polio-virus RNA. Virology 24:467-473.
46. SALZMAN, N. P., AND E. D. SEBRING. 1961. Thesource of poliovirus ribonucleic acid. Virology13:258-260.
47. SHAPIRO, L., AND J. T. AUGUST. 1965. Replicationof RNA viruses. II. The RNA product of areaction catalyzed by a viral RNA-dependentRNA polymerase. J. Mol. Biol. 11:272-284.
48. SHAPIRO, L., AND J. T. AUGUSr. 1965. Replicationof RNA viruses. III. Utilization of ribonucleo-tide analogues in the reaction catalyzed by an
286 BACTrERIOL. REV.
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from

VOL. 30, 1966 RNA VIRUS R
RNA virus RNA polymerase. J. Mol. Biol. 14:214-220.
49. SIMON, E. H. 1961. Evidence for the nonpartici-pation of DNA in viral RNA synthesis. Virol-ogy 13:105-118.
50. SINSHEIMER, R. L., B. STARMAN, C. NAGLER, ANDS. GUTHRIE. 1962. The process of infectionwith bacteriophage ox-174. I. Evidence for a"replicative form." J. Mol. Biol. 4:142-160.
51. SPIEGELMAN, S., I. HARUNA, I. B. HOLLAND, G.BEAUDREAU, AND D. MILLS. 1965. The synthesisof a self-propagating and infectious nucleicacid with a purified enzyme. Proc. Natl. Acad.Sci. U.S. 54:919-927.
52. STEVENS, A., AND J. HENRY. 1964. Studies of theribonucleic acid polymerase from E. coli. I.
Purification of the enzyme and studies of ribo-nucleic acid formation. J. Biol. Chem. 239:196-203.
53. VAN STYVENDAELE, B., AND W. FIERS. 1965. Puri-fication of the replicative form of the bacterio-phage MS2. Arch. Intern. Physiol. Biochim.73:536-537.
EPLICATION 287
54. WATSON, J. D., AND F. H. C. CRICK. 1953. Astructure for deoxyribose nucleic acid. Nature171:737-738.
55. WEISSMANN, C. 1965. Replication of viral RNA.VII. Further studies on the enzymatic repiica-tion of MS2 RNA. Proc. Natl. Acad. Sci. U.S.54:202-207.
56. WEISSMANN, C., P. BORST, R. H. BURDON, H. A.BILLETER, AND S. OCHOA. 1964. Replication ofviral RNA. III. Double-stranded replicativeform of MS2 phage RNA. Proc. Natl. Acad.Sci. U.S. 51:682-690.
57. WEISSMANN, C., P. BORST, R. H. BURDON, M. A.BILLETER, AND S. OCHOA. 1964. Replication ofviral RNA. IV. Properties of RNA synthetaseand enzymatic synthesis of MS2 phage RNA.Proc. Natl. Acad. Sci. U.S. 51:890-897.
58. WEISSMANN, C., L. SIMON, AND S. OCHOA. 1963.Induction by an RNA phage of an enzymecatalyzing incorporation of ribonucleotidesinto ribonucleic acid. Proc. Natl. Acad. Sci.U.S. 49:407-414.
on July 26, 2019 by guesthttp://m
mbr.asm
.org/D
ownloaded from