pontificia universidad catolica de valparaiso · i pontificia universidad catolica de valparaiso...
TRANSCRIPT

i
PONTIFICIA UNIVERSIDAD CATOLICA DE VALPARAISO FACULTAD DE RECURSOS NATURALES
ESCUELA CIENCIAS DEL MAR
Caracterización de los patrones de edad y crecimiento de juveniles de anchoveta (Engraulis ringens) de la zona norte de Chile
asociados a eventos El Niño y La Niña.
Proyecto para optar al titulo de Ingeniero Pesquero
por
Mackarena Francisca Gómez Beltrán
Valparaíso, 2013

ii
COMISIÓN DEL PROYECTO DE TITULO; Profesor Guía : Dr. Guido Plaza Pasten Profesor : Dr. Eleuterio Yáñez Rodríguez Investigador : M. Sc. Francisco Cerna Troncoso

iii

iv
AUTORIZACIÓN DE USO
Al presentar este proyecto como último requisito para la obtención del título de
Ingeniero Pesquero, autorizo a la Escuela de Ciencias del Mar de la Pontificia Universidad Católica de Valparaíso, para que disponga libremente de ella. Autorizo además reproducciones parciales o totales de este proyecto solo con fines académicos. Mackarena Francisca Gómez Beltrán

v
DEDICATORIA
A toda mi familia, en especial a mi abuela por sus años de dedicación, a mis padres y hermanos por su apoyo y cariño incondicional.

vi
AGRADECIMIENTOS
Quiero expresar mis más sinceros agradecimientos a la persona que con su esfuerzo y años de dedicación hicieron posible alcanzar esta tan anhelada meta, mi abuela Ana Gaete Canales, quién siempre se esmeró en conseguir que fuese “alguien en la vida” y hacerme entender que el único camino para conseguirlo era solo a través del estudio y la responsabilidad, gracias por la preocupación y su empeño en querer que fuese una profesional, gracias por todo.
Agradecer a mi madre Rosa Beltrán y padre Johnny Tapia quienes siempre a pesar de las condiciones adversas estuvieron ahí y me brindaron su apoyo siempre cuando los necesite.
A mi gran amiga Jenniffer Angulo con la que he compartido incontables momentos importantes en mi vida, a sus padres y hermanos entre ellos Tania Angulo quienes con su alegría y cariño me han permitido formar parte de su familia, a Alan Soto por sus tan sabios consejos, a Priscilla y Gina por incentivarme siempre a terminar con esta etapa. A Mario Neira por su incondicional apoyo y desinteresada ayuda en todo, por su energía y alegría que contagia y por su tan linda e inigualable compañía de este último tiempo.
A mi profesor guía Sr. Guido Plaza, por brindarme la oportunidad de poder realizar este último y tan importante proceso en su laboratorio, a mis compañeros de laboratorio Javier Leguá por su colaboración en la realización de este proyecto, y en especial a Alejandra Hernández y María de los Ángeles Cornejo por su excelente acogida al aposento de la sabiduría, por su ayuda y apoyo durante todo este proceso y por tan buenos momentos compartidos
A mis compañeros y amigos de universidad quienes de una u otra manera formaron parte en esta etapa, en especial a Francisca López Campos por su amistad incondicional durante todo este tiempo.
Y a todos los Profesores de la Escuela Ciencias del Mar de la PUCV quienes han contribuido de una u otra manera en mi formación como profesional, en especial al profesor Rodolfo Olivarí Q.E.P.D. quien con su especial forma de ser, su cercanía y disposición de ayudar siempre, han dejado un imborrable y lindo recuerdo.

vii
CONTENIDO
Pág PORTADA COMISIÓN DEL PROYECTO DE TITULO
ACTA DE REVISIÓN DE PROYECTO DE TITULO
AUTORIZACIÓN DE USO
DEDICATORIA
AGRADECIMIENTOS
CONTENIDO
INDICE DE TABLAS
INDICE DE FIGURAS
RESUMEN
ABSTRACT
1. INTRODUCCIÓN
2. OBJETIVOS
2.1 Objetivo general
2.2 Objetivos específicos
3. ANTECEDENTES
3.1 Otolimetría
3.2 Micro-incrementos primarios
3.2.1 El primordium
3.2.2 El primer anillo primario
3.2.3 Centro secundarios de crecimiento (CSC)
3.2.4 Estructuras discontinuas (ED)
3.3 Preparación de otolitos de peces juveniles
3.4 Retro-calculo
3.4.1 Método Proporcional
3.4.2 Método de regresión lineal
3.5 De la especie
i
ii
iii
iv
v
vi
ix
xi
xiv
xv
1
3
3
3
4
4
6
7
7
8
8
8
9
10
10
11

viii
3.6 De la pesquería
3.7 Estadios tempranos de peces teleósteos
3.8 Pesquerías y el ambiente
3.8.1 El Niño Oscilación del Sur (ENOS)
3.8.2 Efectos de “El Niño” en la pesquería de la anchoveta
3.8.3 La fase fría del ciclo ENOS “La Niña”
3.8.4 Ocurrencia Histórica del ENOS
4. METODOLOGIA
4.1 Aspectos Generales
4.2 Caracterización de la micro-estructura de otolitos
4.2.1 Aplicación del protocolo SGEM para preparación de otolitos
4.2.2 Proceso de pulido de otolito
4.2.3 Agrupación de imágenes
4.2.4 Lectura de los micro-incrementos primarios
4.3 Crecimiento y ambiente
4.3.1 Crecimiento de juveniles de anchoveta Engraulis ringens en
presencia de los eventos “Niño” y “Niña”
4.3.2 Procedimientos para la evaluación y comparación del
crecimiento
4.4 Análisis estadístico comparativo del crecimiento
4.4.1 Comparación de los modelos de crecimiento
4.4.2 Comparación grosor del micro-incremento
4.4.3 Comparación tasas de crecimiento retro-calculadas
5. RESULTADOS
5.1 Estadísticas Generales
5.2 Análisis de la microestructura de otolitos
5.2.1 Simetría entre pares de otolitos
5.2.2 Caracterización la micro-estructura de otolitos de anchoveta
juvenil.
5.2.3 Primordio
12
13
15
15
16
16
17
19
19
19
19
21
22
23
24
24
25
26
26
27
28
30
30
31
31
32
33

ix
5.2.4 Marca de eclosión y primera alimentación
5.2.5 Grosores micro-incrementos
5.3 Análisis para determinar diferencias en el crecimiento de juveniles de
anchoveta Engraulis ringens en presencia de los eventos Niño y Niña.
5.3.1 Relación talla-edad
5.3.2 Tasas de crecimiento diarias retro-calculadas
5.3.3 Grosor de los micro-incrementos
6. DISCUSIÓN
6.1 Micro-estructura de otolitos juveniles de anchoveta Engraulis ringens
6.2 Crecimiento en la etapa juvenil asociado a los Eventos el Niño y la Niña
7. CONCLUSIONES
8. REFERENCIAS
33
33
34
37
37
43
46
49
49
51
53
54

x
INDICE DE TABLAS
Pág Tabla 1. . Validación de los incrementos diarios en otolitos de especies de clupeiformes, Marcador químico utilizado oxytetraciclina. Extraído de Castellano y Vasconcellos (1995). Tabla 2. Métodos de tipo proporcional más empleados en el uso de las técnicas de retro-cálculo. Tabla 3. Zona de recolección de ejemplares juveniles de anchoveta en la zona Norte de Chile. Tabla 4. Diseño balanceado de análisis de varianza de medidas repetidas (ANDEVA-RM)) para determinar diferencias en los grosores entre cohortes de años con eventos El Niño. Tabla 5. Diseño balanceado de análisis de varianza de medidas repetidas (ANDEVA-RM)) para determinar diferencias en los grosores entre cohortes de años con eventos El Niño y La Niña Tabla 6. Diseño balanceado de análisis de varianza de medidas repetidas (ANDEVA-RM)) para determinar diferencias en el crecimiento entre cohortes de años con eventos El Niño y La Niña. Tabla 7. Otolitos de anchovetas juveniles analizados por año, área y mes para el análisis de micro-incrementos. Tabla 8 Análisis multivariado de medidas repetidas para determinar diferencias significativas en las lecturas realizadas por los lectores. Lambda de Wilks= estadístico de prueba multivariado, P> 0,05. Tabla 9. Marca de eclosión y de primera alimentación en juveniles de anchoveta asociados a ventos El Niño y La Niña. Tabla 10. Fluctuación, media y extensión de los grosores de los micro-incrementos por sección de otolitos juveniles de la zona Norte de Chile, asociados a eventos de El Niño y La Niña. Tabla 11. Rango de longitud y edad analizada en juveniles de anchoveta para el evento El Niño 1973 y 1982, y para el evento La Niña 2009-2010. Tabla 12. Parámetros derivados de los ajustes del modelo de crecimiento de Laird Gompertz para cada evento y año.
5
10
19
27
28
29
30
32
34
35
38
39

xi
Pág
Tabla 13. Parámetros derivados de los ajustes del modelo de crecimiento de Laird Gompertz para cada evento en la estación de verano. Tabla 14. Procedimiento de contraste para la curva de crecimiento del modelo Laird–Gompertz derivada del análisis estacional de verano para anchoveta de la zona norte. Subíndices (73, año 1973; 82, año 1982; 10, año 2010). Tabla 15. Contraste multivariado global entre los eventos El Niño y La Niña para las tasas de crecimiento retro-calculadas. Tabla 16. Contraste multivariado estacional entre los eventos El Niño y La Niña para las tasas de crecimiento retro-calculadas. Tabla 17. Contraste multivariado del análisis global de medidas repetidas entre los eventos El Niño (1973 y 1982) para el grosor de los micro-incrementos Tabla 18. Contraste multivariado del análisis global de medidas repetidas entre los eventos El Niño y La Niña para los grosores de los micro-incrementos. Tabla 19. Contraste multivariado para la estación de verano entre los eventos El Niño y La Niña para las tasas de crecimiento retro-calculadas.
41
43 44 45 46 47 47

xii
INDICE DE FIGURAS
Pág
Figura 1. A: Localizacion de los otolitos en el aparato vestibular (Secor et al., 1992) y B: aspecto general del laberinto auditivo derecho por la cara medial. Figura 2. Fotografías de microscopía óptica ilustrando rasgos de la micro-estructura de los otolitos de peces juveniles (1 &2) Sagittae de un juveniles de Sardinops melanosctitus (Plaza et al. com. pers.); (3) región del primordio de una otolito sagita del clupeido Etremeus teres (Plaza et al., 2005); (4 & 5) Secciones frontales y sagitales de un juvenil de un pez de roca (Sebastes inermis, Plaza et al., 2001). P= primordio; me = marca a la eclosión; pa = marca a la primera alimentación; CSC = centros secundarios de crecimiento. Figura 3. Relación lineal entre el crecimiento del pez y el crecimiento del otolito. Es el principio de dependencia lineal base para la utilización del método retro-cálculo (Chugunova, 1963) Figura 4. Cardumen de anchoveta de alta densidad Figura 5. Distribución de la anchoveta (Engraulis ringens) en Perú y norte de Chile. Figura 6. Ilustración del mecanismo “bigger-is-better”, donde individuos de una misma edad difieren en tamaño. Figura 7. Aumento de la TSM en las costas sudamericanas correspondiente a la llegada del fenómeno de El Niño Figura 8. Enfriamiento de la TSM en las costas sudamericanas, correspondiente a la llegada del fenómeno de La Niña
Figura 9. Anomalías de la TSM de la base datos de NOAA.
Figura 10. Sistema de almacenamiento de otolitos, denominado “cartones”
Figura 11. Ilustración método SGEM: preparación de otolitos para el análisis de micro-incrementos. A: Aplicación de cera termoplástica; B: resina de dos componentes en partes iguales; C: Mezcla de los componentes de resina epóxica; D y E: montaje del otolito (Fuente: Plaza et al. 2005). Figura 12. A: fotografía bajo luz trasmitida de un otolito sin pulir, no se diferencian los micro-incrementos diarios, con aumento 400X, B: fotografía de otolito pulido, donde pueden observarse claramente los anillos diarios, C: imagen de otolito sobre pulido, en donde se pierde completamente la resolución de sus micro-incrementos.
4
7
9
11
12
14
15
17
18
20
21
21

xiii
Pág
Figura 13. Imagen bajo luz trasmitida de la secuencia completa de los micro-incrementos desde el primordio hasta el borde del otolito de juvenil de anchoveta: edad 88 días, LT= 9,5 cm, Aumento: 400X.
Figura. 14. Agrupación de imágenes secuenciales de la micro-estructura de otolitos para la lectura de micro-incrementos primarios en juveniles de anchoveta. Figura 15. Conteo de anillos y medición de radios de un otolito juvenil de anchoveta a través del programa analizador de imágenes. Figura 16. Otolito de anchoveta juvenil de 11,5 cm, donde se observa claramente a través de su cara externa, el rostro y antirostro y las pequeñas irregularidades de sus bordes ventral y dorsal (Aumento 100X).
Figura 17. Imágenes bajo luz trasmitida agrupadas de la micro-estructura de otolitos para la lectura de micro-incrementos primarios en juveniles de anchoveta: A) edad 117 días, LT= 11,5 cm, aumento 400X, zona Arica, 1973, B: edad 116 días, LT= 11,5 cm, aumento 400X, Zona Mejillones, año 1982 y C: edad 104 días, LT= 11 cm, aumento 400X, zona Iquique, 2010) Figura 18. Fotografias bajo luz trasmitida del analisis micro-estructural de otolitos juveniles de anchoveta de la zona norte de Chile, donde se observa visiblemente el primordio en los tres ejemplares :A) edad 117 dias, LT = 11 cm, zona Arica, 1973; B) edad 134 dias, LT = 12 cm, zona Iquique,1982 y C) edad 116 dias, LT = 11,5 cm, zona Iquique. 2010; Pr; Primordio, Me; marca de eclosion, Mpa; marca primera alimentacion (Aumento 400X) Figura 19. Fotografia bajo luz trasmitida de la micro-estructura de un otolito juvenil de anchoveta; edad 119 dias, LT=11,5 cm, zona Mejillones, año 2010, Pr; primordio, Ad; anillo doble, Asd; anillo sub-diario. Figura 20. Fotografia bajo luz trasmitida de la lectura de un otolito juvenil de anchoveta, edad 116 dias, LT= 11,5 cm, zona Mejillones, año 1982, en la esquina inferior izquierda, se observa el grosor de los micro-incrementos a medida que aumenta el número de micro-incrementos del otolito. Figura 21. Grosor medio de micro-incrementos en otolitos juveniles de anchoveta de la zona Norte de Chile, para eventos El Niño y La Niña. Figura 22. Relación talla-edad de juveniles de anchoveta mediante ajuste del modelo de crecimiento de Laird - Gompertz ( ) realizado por cada evento El Niño 1973, 1982 y evento El Niña” 2009-2010 ajustando la tasa instantánea de crecimiento diario ( ). (a) corresponde al ajuste por evento global promedio de la longitud total (mm) derivada del retro-calculo desde el primer día de vida y (b) corresponde al ajuste por evento promedio de la longitud total (mm) derivada del retro-calculo desde el día 40 de vida.
22
22
23
31
32
33
35
36
36
40

xiv
Figura 23. Relación talla-edad de juveniles de anchoveta mediante ajuste estacional para la época de verano del modelo de crecimiento de Laird - Gompertz ( ) realizado por cada evento El Niño (1973 y 1982) y evento La Niña (2009-2010) ajustando la tasa instantánea de crecimiento diario ( ). (a) corresponde al ajuste por evento global promedio de la longitud total (mm) derivada del retro-calculo desde el primer día de vida.(b) corresponde al ajuste por evento promedio de la longitud total (mm) derivada del retro-calculo desde el día 40 de vida. Los parámetros del ajuste se indican en la Tabla 12 Figura 24.A: Análisis global de la tasa de crecimiento retro-calculada media anual de los micro-incrementos por evento. B: Análisis estacional de la tasa de crecimiento retro-calculada media anual para la época de verano por evento. La línea vertical muestra el punto de corte del análisis de medidas repetidas. Figura 25. A: Grosor medio anual de los micro-incrementos para el evento El Niño y La Niña usando un enfoque global. B: Grosor medio anual realizado para el verano de cada evento. La línea vertical muestra el punto de corte del análisis de medidas repetidas.
42
45
48

xv
RESUMEN
Estudios recientes han demostrado que el crecimiento y la sobrevivencia están directamente relacionados en los estadios tempranos de peces teleósteos. El estudio de las fases de vida temprana de los peces marinos resulta muy interesante y fundamental para entender la dinámica de las poblaciones de peces y por ende un mejor manejo y administración de los recursos pesqueros. En este contexto, en el presente estudio se llevó a cabo un análisis comparativo de la edad y crecimiento en juveniles de la anchoveta (Engraulis ringens) en la zona norte de Chile. La determinación de la edad fue realizada mediante la lectura de micro-incrementos diarios en otolitos sagitales, en ejemplares < 12 cm de LT y edades entre 60 y 178 días. Los juveniles fueron recolectados en cruceros de reclutamiento en la zona norte específicamente Arica, Iquique y Antofagasta, durante los años 1973 y 1982, asociados a evento El Niño y 2009-2010, con influencia de un evento La Niña. El grosor medio de los micro-incrementos fluctuó entre 4 y 13 μm, siendo distintivos a través de toda la micro-estructura de los otolitos. Las relaciones talla-edad se ajustaron a un modelo Laird-Gompertz, utilizando longitudes retro-calculadas, a través del método del intercepto biológico propuesto por Campana (1990). Las tasas de crecimiento globales al punto de inflexión fluctuaron entre 1,24 - 1,64 mmd-1, alcanzándose a edades entre 45 y 46 días. En el ajuste estacional, realizado para la época de verano, las tasas de crecimiento variaron entre 1,32 y 1,64 mmd-1 a edades de 44 y 45 días. Estas tasas de crecimiento elevadas se obtuvieron independientes de los eventos El Niño y La Niña, indicando que esta especie tiene una gran plasticidad durante sus estadios tempranos, maximizando su potencial de crecimiento en sus primeros meses de vida. Palabras claves: otolito, anchoveta, retro-cálculo, El Niño, La Niña.
ABSTRACT

xvi
Recent studies have shown that growth and survival are directly related in the early stages of teleost fish. In addition, the study of early life stages of marine fish is an very interesting and important issue to understand the dynamics of fish populations and contribute to enhance the management and conservation of fishery resources. In this context, the present study addressed to make a comparative analysis of age and growth in juveniles of the anchovy Engraulis ringens in northern Chile. Age determination was made by reading daily micro-increments in sagittal otoliths in specimens <12 cm total length and between 60 and 178 days. Juveniles were collected on a research campaign made to evaluate recruitment in northern Chile, during the years 1973 and 1982, associated with the "El Niño" event and 2009-2010, influenced by "La Niña" event. The average thickness of the micro-increments ranged between 4 and 13 microns, being distinctive throughout the microstructure of otoliths. The age-length relationships were fitted to a Gompertz-Laird model, using back-calculated lengths through the Biological Intercept method proposed by Campana (1990). Global growth rates at the inflection point ranged from 1.24 to 1.64 mmd-
1, reached at ages between 45 and 46 days. In a seasonal approach, made for the summer, growth rates ranged between 1.32 and 1.64 mmd-1 at ages 44 and 45 days. These high growth rates were obtained irrespective of El Niño and/or La Niña events. The findings suggest this species has a great plasticity in its early stages, maximizing their growth potential in their first months of life.
Keywords: Otolith, anchovy, back-calculation, El Niño, La Niña.

1
1. INTRODUCCIÓN
La edad y el crecimiento de los peces resultan indispensables en la dilucidación de la biología de las especies, para un adecuado manejo y ordenación pesquera (Leta & Keim, 1982). De los estudios de edad pueden ser determinados datos básicos, tales como: edad de primera madurez, éxito del reclutamiento, estructura de edad poblacional, cambios poblacionales causados por las tasas de explotación entre otros. (Morales-Nin, 1991). Por lo tanto, es importante conocer la edad en la que una especie alcanza la longitud adecuada para la pesca y su reclutamiento a la pesquería, así como sus parámetros de crecimiento y mortalidad que son fundamentales y constituyen la base de los modelos de dinámica de poblaciones, que son utilizados en la regulación y análisis de pesquerías (Tresierra & Culquichicon, 1993).
La edad de muchas especies marinas puede determinarse a partir de los anillos de crecimiento presentes en algunas estructuras duras, como escamas, otolitos, vértebras, espinas y radios (Araya & Cubillos, 2002). Los anillos de crecimiento estacionales en otolitos están formados por una zona translúcida y otra opaca, constituyendo entre ambas un anillo anual (denominado anulo), cuyo crecimiento registra la cronología histórica desde el nacimiento hasta la fecha de muerte del pez (Casselman, 1990). Alternativamente, los anillos de crecimiento diario (o micro-incrementos, Panella, 1971) son solamente visibles a través de un microscopio durante el primer año de vida del pez, debido a que su crecimiento es mayor durante dicho período. Numerosos estudios de edad y crecimiento, basados en la micro-estructura de otolitos, (Campana & Jones, 1992; Gorini et al., 2010; Jones, 1992; Morales-Nin, 1991; Quiñones & López, 2001; Chong et al., 2007) indican que los incrementos diarios presentes en estos, pueden ser usados para determinar la edad de larvas y juveniles de peces y así la duración del desove, desarrollo, eventos y estados tempranos de la historia de vida. La ventaja de utilizar otolitos a nivel diario es que son las únicas estructuras que registran consistentemente los eventos en los estadios tempranos de la vida, permaneciendo esta información inalterada a lo largo del tiempo (Campana & Nielson, 1985; Jones, 1992).
Una de las aplicaciones importantes derivada del análisis de los otolitos a nivel diario es la determinación precisa de las tasas de crecimiento durante la vida temprana en el mar. Luego una vez disponible la tasas de crecimiento, se puede estimar la tasa de mortalidad larval (Lough et al., 1982). Las tasas de mortalidad en estadios tempranos suelen ser muy elevadas en algunas especies pelágicas como la sardina y anchoveta, recursos de suma importancia económica para nuestro país. Aproximadamente el 99% de la progenie de estas especies muere y el 1% restante es el destinado a sostener la pesquería y a mantener la viabilidad de la especie. (Plaza & Cerna, 2011).

2
En Chile la anchoveta (Engraulis ringens) es uno de los recursos de mayor importancia económica, que junto con la sardina sostienen la pesquería pelágica de la zona norte (Arcos et al., 2004; Aliaga et al., 2001). La anchoveta se caracteriza por ser un recurso costero, de rápido crecimiento, ciclo de vida corto, posee elevadas tasas de mortalidad natural; su distribución espacial, biomasa y reclutamiento son fuertemente influenciados por factores ambientales. Por lo tanto, resulta indispensable tener conocimiento de la importancia que tienen las condiciones ambientales durante el primer período de crecimiento rápido, que no sólo determina cambios en la longitud media de los peces de una cohorte, sino también tiene incidencia directa sobre la actividad reproductiva venidera.
Existe una variada y abundante literatura bien fundamentada (Yánez et al., 1992;
Cañón, 1986; Martínez et al., 1985; Serra, 1983; Espino, 1999; Lombarte, 1990; Methot, 1983; Cubillos & Arcos, 2002; Cubillos et al., 1998) que apoya la tesis que las fluctuaciones en el ambiente físico tienen un notable impacto en la pesquería de la anchoveta en diferentes escalas de tiempo y espacio. Dentro de estas fluctuaciones El Niño y La Niña aparecen como los más relevantes a la hora de explicar los cambios en abundancia, vulnerabilidad y distribución de esta especie (Avaria et al., 2004). El aspecto más discutido ha sido el hecho de que la anchoveta se distribuye a mayor profundidad ante el predominio de aguas sub-superficiales cálidas asociadas a eventos El Niño, tornando a este recurso menos vulnerable a la pesquería de cerco Sosa (1999). No obstante, existe falta de información acerca de los efectos que producen los eventos de El Niño y La Niña en el crecimiento durante los estadios tempranos de esta especie, siendo de vital importancia tener conocimiento de lo que realmente sucede en esta fase en donde el recurso es considerablemente vulnerable.
La presente tesis propone evaluar el crecimiento durante los estadios tempranos de
la anchoveta (Engraulis ringens) bajo la influencia de eventos el Niño y Niña en la zona norte de Chile. Para este propósito se efectuó un estudio acucioso de la microestructura de otolitos de juveniles de esta especie, utilizando muestras históricas para ambos eventos.

3
2. OBJETIVOS
2.1 Objetivo general Caracterizar los patrones de edad y crecimiento de juveniles de anchoveta (Engraulis ringens) de la zona norte de Chile, asociados a eventos El Niño y la Niña. 2.2 Objetivos específicos
2.2.1. Caracterizar la micro-estructura de otolitos de anchoveta juveniles de la zona
estudio. 2.2.1. Reconstruir los patrones de crecimiento diario de juveniles de anchoveta,
utilizando técnicas de retro-cálculo, en presencia del evento Niño (1972-1973 y 1982-1983) y el evento Niña (2009-2010).
2.2.1. Comparar el crecimiento retro-calculado durante los eventos Niño y Niña
utilizando técnicas multivariadas.
4
3. ANTECEDENTES
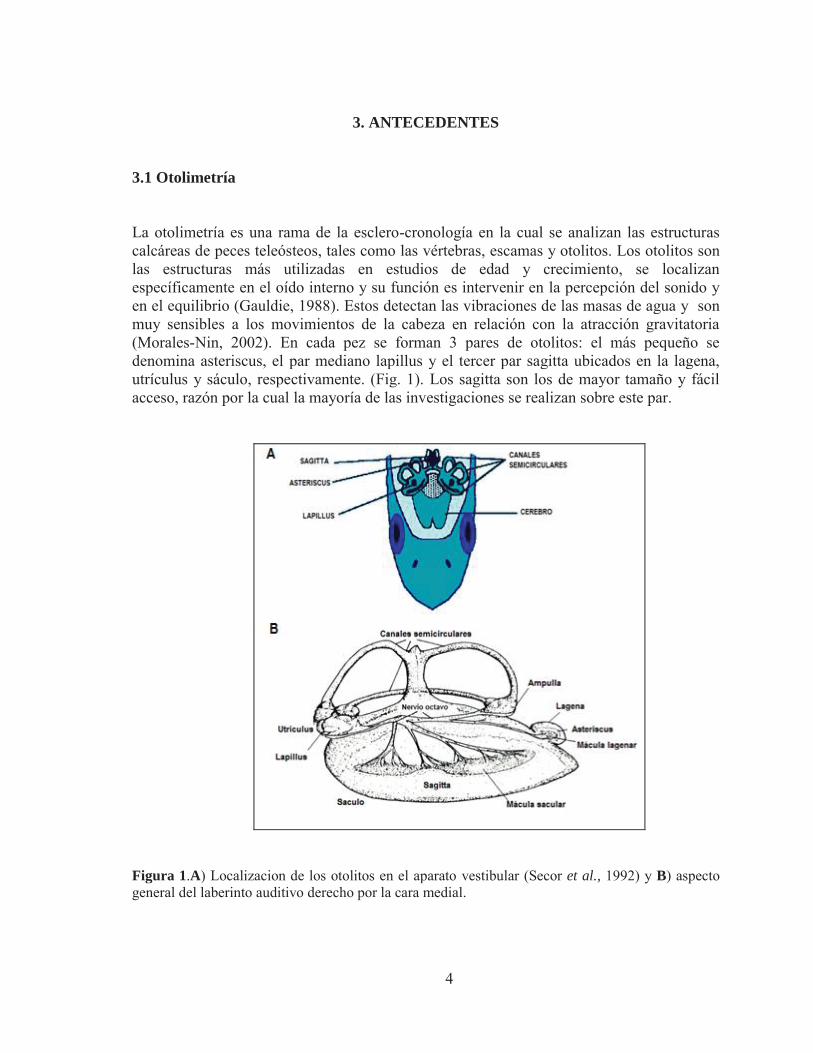
3.1 Otolimetría La otolimetría es una rama de la esclero-cronología en la cual se analizan las estructuras calcáreas de peces teleósteos, tales como las vértebras, escamas y otolitos. Los otolitos son las estructuras más utilizadas en estudios de edad y crecimiento, se localizan específicamente en el oído interno y su función es intervenir en la percepción del sonido y en el equilibrio (Gauldie, 1988). Estos detectan las vibraciones de las masas de agua y son muy sensibles a los movimientos de la cabeza en relación con la atracción gravitatoria (Morales-Nin, 2002). En cada pez se forman 3 pares de otolitos: el más pequeño se denomina asteriscus, el par mediano lapillus y el tercer par sagitta ubicados en la lagena, utrículus y sáculo, respectivamente. (Fig. 1). Los sagitta son los de mayor tamaño y fácil acceso, razón por la cual la mayoría de las investigaciones se realizan sobre este par.
Figura 1.A) Localizacion de los otolitos en el aparato vestibular (Secor et al., 1992) y B) aspecto general del laberinto auditivo derecho por la cara medial.

5
Los otolitos están conformados por una matriz proteica sobre la cual se depositan cristales de carbonato de calcio en forma de aragonito y de una fibra de colágeno-proteína similar a la otolina (Degens et al., 1969). El otolito crece por la deposición de material sobre la superficie, y según Pannella (1971) estos depósitos poseen una naturaleza diaria, lo que fue confirmado posteriormente por Brothers et al. (1976) y Jones (1986) en Engraulis morda y otras especies (Tabla 1). Este depósito cíclico se debe a los ritmos internos del metabolismo del calcio y de la síntesis de aminoácidos (Simkiss, 1974). El resultado de este proceso cíclico es la formación de los incrementos de crecimiento diario en el otolito, compuestos por una unidad continua o incremental y una unidad discontinua (Pannella, 1971, 1974; Dunkelberguer et al., 1980). La zona incremental se compone de micro-cristales como agujas de aragonito rodeadas por la matriz orgánica y la zona discontinua; esta compuesta principalmente por capas concéntricas de materia orgánica (Mugiya et al., 1981; Watabe et al., 1982; Morales-Nin, 1986 b)
Tabla 1. Validación de los incrementos diarios en otolitos de especies de clupeiformes, Marcador químico utilizado oxytetraciclina. Extraído de Castellano y Vasconcellos (1995).

6
La formación diaria de los incrementos de crecimiento depende de un ritmo circadiano endocrino que se sincroniza a edad temprana con el fotoperiodo u otros factores externos diarios (Tanaka et al., 1981; Radtke & Dean, 1982; Campana & Nielson, 1985)
El grosor de los incrementos y la densidad de los micro-cristales depende de la fase de crecimiento (Irie, 1960). Así, en los periodos de crecimiento rápido los incrementos son gruesos con unidades discontinuas bien desarrolladas y en los períodos de crecimiento lento los incrementos son más finos, compactos y continuos. Frecuentemente se encuentran dos o más subunidades en los incrementos, probablemente causadas por ritmos de alimentación, migraciones (Pannella, 1974; 1980) y cambios de la temperatura (Brothers, 1978; Campana, 1983; Campana & Neilson, 1982). 3.2 Micro-incrementos primarios
Los micro-incrementos primarios (MP) (Fig. 2, Fotos 1~4) son formados por la sucesiva deposición de una capa rica en carbonato de calcio y de una capa rica en proteína pero de baja concentración en carbonato de calcio. Ambas capas son denominadas zonas L y D (Fig. 2, Foto 2), apareciendo como una zona clara y oscura bajo luz trasmitida, respectivamente. Pannella (1971) fue quien descubrió estas estructuras y fue el primero en sugerir que su frecuencia de producción era diaria. El grosor de los MP puede fluctuar desde menos de 1 μm hasta los 15 μm o más. El grosor de la zona L es de alrededor de 1 μm, mientras que el grosor de la zona D puede ser más variable. Los micro-incrementos de la zona D son solamente visibles a través de un microscopio, principalmente durante el primer año de vida del pez, debido a que su crecimiento es mayor durante dicho período. No obstante, los MP se siguen formando durante la vida del pez, aunque su periodicidad diaria es difícil de evaluar debido a que su grosor disminuye notoriamente, cayendo más allá de la resolución de la microscopía óptica e incluso electrónica. Algunos incrementos sub-diarios pueden ocurrir, particularmente en aquellos períodos de crecimiento rápido. Estas estructuras son menos uniformes y regulares que los incrementos primarios, y por consiguiente pueden fácilmente ser distinguidos por un lector experimentado (Campana & Jones, 1992).
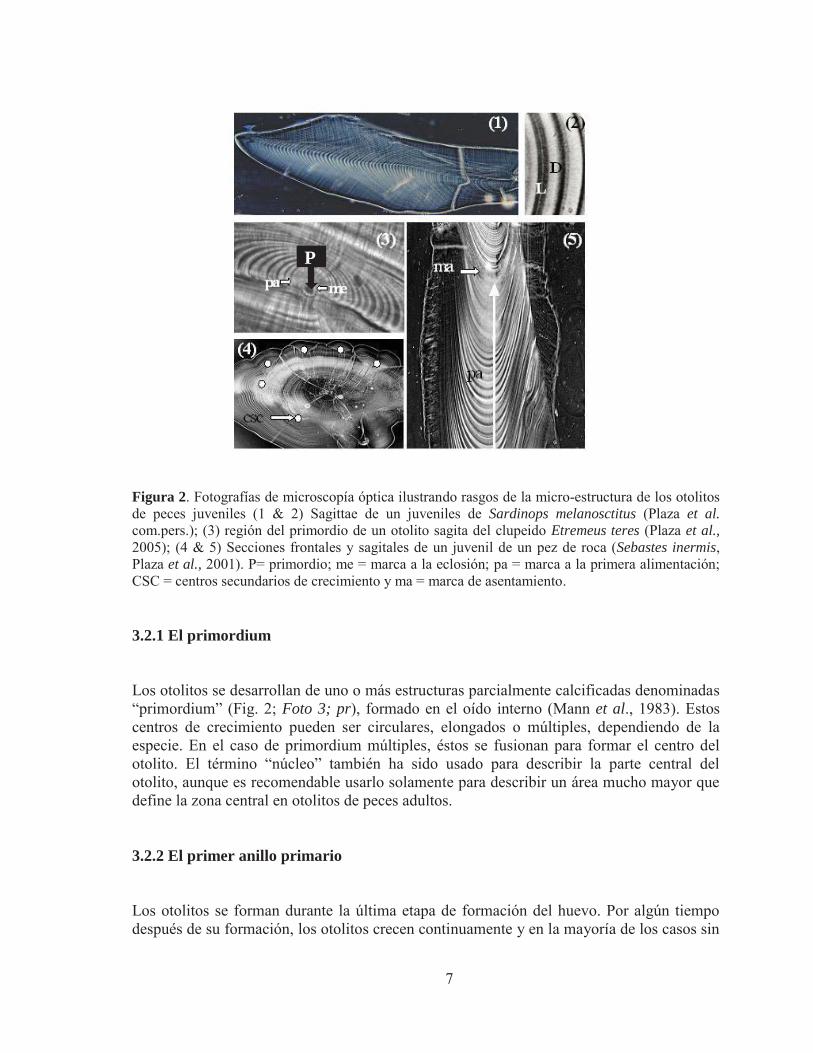
7
P
Figura 2. Fotografías de microscopía óptica ilustrando rasgos de la micro-estructura de los otolitos de peces juveniles (1 & 2) Sagittae de un juveniles de Sardinops melanosctitus (Plaza et al. com.pers.); (3) región del primordio de un otolito sagita del clupeido Etremeus teres (Plaza et al., 2005); (4 & 5) Secciones frontales y sagitales de un juvenil de un pez de roca (Sebastes inermis, Plaza et al., 2001). P= primordio; me = marca a la eclosión; pa = marca a la primera alimentación; CSC = centros secundarios de crecimiento y ma = marca de asentamiento. 3.2.1 El primordium Los otolitos se desarrollan de uno o más estructuras parcialmente calcificadas denominadas “primordium” (Fig. 2; Foto 3; pr), formado en el oído interno (Mann et al., 1983). Estos centros de crecimiento pueden ser circulares, elongados o múltiples, dependiendo de la especie. En el caso de primordium múltiples, éstos se fusionan para formar el centro del otolito. El término “núcleo” también ha sido usado para describir la parte central del otolito, aunque es recomendable usarlo solamente para describir un área mucho mayor que define la zona central en otolitos de peces adultos. 3.2.2 El primer anillo primario Los otolitos se forman durante la última etapa de formación del huevo. Por algún tiempo después de su formación, los otolitos crecen continuamente y en la mayoría de los casos sin

8
producir capas concéntricas u incrementos primarios. El comienzo de la formación del primer incremento primario varía de una especie a otra, pero en la mayoría de las especies

9
estudiadas, el primer incremento aparece prominente y bien definido (Fig. 2; Foto 3; pa). En algunas especies la formación del primer incremento coincide con la eclosión (Lagardere, 1989), (Fig.2 Foto 3, me=marca de eclosión), y en otros con la primera alimentación (Geffen, 1982), (Fig. 2; Foto 3; pa=marca de primera alimentación). En otras especies, estructuras concéntricas han sido observadas antes de la eclosión, aunque aún no está claro si estas estructuras son incrementos diarios o meramente artefactos visuales. 3.2.3 Centro secundarios de crecimiento (CSC) En algunas especies es posible apreciar centros secundarios de crecimiento análogos al primordium (Fig. 2; Foto 4; círculos blancos), del cual emanan nuevas estructuras concéntricas. Estos CSC son más comunes en el otolito más grande (sagita). Varias hipótesis han sido planteadas para explicar el origen de estos CSC, entre las cuales se pueden citar: cambio de hábitat (Sogard, 1991), cambios ontogenéticos en la dieta (Marks & Conover, 1993) y/o cambios fisiológicos asociados a la metamorfosis (Hare & Cowen, 1994). 3.2.4 Estructuras discontinuas (ED) Las ED son elementos sobresalientes e irregulares que ocurren a través de los incrementos primarios. Estas ED pueden interrumpir la sucesión de los incrementos primarios en forma cíclica u acíclica. Las ED pueden ser distinguidas bajo magnificaciones altas o bajas y generalmente requieren que el otolito sea previamente preparado. Las ED son típicas de todas las especies y probablemente son productos del estrés ambiental al que los individuos están sujetos. Algunas ED están relacionadas a eventos tales como asentamiento y/o metamorfosis y aparecen como drásticos cambios en el grosor del incremento (Fig. 2; Foto 5; ma= marca de asentamiento; pa= Incrementos primarios post-asentamiento) 3.3. Preparación de otolitos de peces juveniles Para observar la micro-estructura de los otolitos de peces juveniles es necesario obtener una lámina de alrededor de 50 μm de grosor en el plano sagital, frontal o transversal. Este proceso normalmente tiene que ser llevado a cabo manualmente, usando lijas de distinta graduación. El procedimiento básico consiste en embeber el otolito en resina epóxica, pulirlo hasta revelar el primordio y los MP y posteriormente montar la sección en un portaobjeto, para finalmente observarla a través de un microscopio óptico. Mayores detalles de estos procedimientos han sido descrito por Secor et al. (1992) y Plaza et al. (2005).

11
3.4. Retro-calculo El análisis de la micro-estructura de los otolitos permite, mediante los micro-incrementos, reconstruir los patrones de crecimiento a través de una técnica llamada retro-calculo (Francis, 1990). El análisis retrospectivo o retro-cálculo es una técnica que permite utilizar esa información para estimar dimensiones corporales en edades o etapas previas de la historia de vida. Específicamente se usan las dimensiones de las marcas de crecimiento de la estructura dura (otolito) (Fig. 3) junto a la longitud del cuerpo al momento de la captura, para estimar la longitud al tiempo de formación de cada una de estas marcas.
Figura 3. Relación lineal entre el crecimiento del pez y el crecimiento del otolito. Es el principio de dependencia lineal, base para la utilización del método retro-cálculo (Chugunova, 1963)
El retro-cálculo es una herramienta de gran utilidad, ya que se puede monitorear las tasas de crecimiento promedio de varias clases de edad en la población, identificando de esta forma años o estaciones de crecimiento lento o rápido (Smith, 1983).
Existen dos métodos que se pueden utilizar al aplicar retro-cálculo, el método proporcional y el método de regresión. Estos utilizan los parámetros de la ecuación que relaciona la longitud del pez y el tamaño del otolito al momento de la captura y asumen que existe una proporcionalidad entre el crecimiento somático y el crecimiento del otolito.

13
3.4.1 Método proporcional Este método incorpora información de cada individuo, lo que da cuenta de la variación individual. En esta aproximación se asume que existe una proporcionalidad entre el crecimiento somático y el crecimiento del otolito (Carlander 1981; Campana 1990). En la Tabla 2 se muestran los modelos proporcionales mas utilizados. Tabla 2. Métodos de tipo proporcional más empleados en el uso de las técnicas de retro-cálculo.
Método Dahl-Lea
Se basa en que el otolito crece proporcionalmente a la longitud del pez.
Método Frasser-
Lee
Se basa en el supuesto "El incremento de crecimiento del otolito es, en promedio, una proporción constante del incremento de crecimiento del pez"
Método Hile
Se basa en el supuesto que "El porcentaje o desviación relativa del tamaño de cualquier otolito, desde el tamaño teórico, es constante al momento de formación de todos los annuli".
Método intercepto
biológico
Emplea un intercepto determinado biológicamente mas que uno estimado estadísticamente. Considera el intercepto del comienzo de la proporcionalidad entre el tamaño del otolito y la longitud del pez.
3.4.2 Método de regresión lineal Este método utiliza la ecuación para estimar las longitudes pretéritas a cada edad directamente desde las marcas periódicas, sin preocuparse de la variación en el tamaño del otolito entre los peces.
Los métodos de retro-cálculo se basan en que existe una proporcionalidad entre el
crecimiento del otolito y el crecimiento del pez.(Campana, 1990; Araya & Cubillos, 2002; Carlander, 1981) Sin embargo, hay un número de especies en donde la relación longitud pez-otolito es no lineal (Wright et al., 1990) El retro-cálculo en estas especies es difícil; además cuando se retro-calcula desde una relación curvilínea hay un supuesto implícito, que el punto de inflexión de la curva ocurre al mismo tamaño pez - tamaño otolito en cada pez. En estos casos el retro-cálculo no se debería realizar, debido a la violación de este supuesto lo que entregaría resultados dudosos.

14
3.5 De la especie La anchoveta es una especie pelágica de hábitos costeros, aguas superficiales moderadamente frías, con rangos que oscilan entre 16° y 23°C en verano y de 14° a 18°C en invierno. Vive en cardúmenes altamente densos en áreas que pueden estar entre 36 km y 180 km de la costa (Fig. 4). Los cardúmenes de anchovetas se desplazan en aguas superficiales de hasta 50 metros de profundidad en el día y suben en la noche. Esta especie posee un crecimiento rápido, ciclo de vida corto con tres a cuatro años de longevidad, elevadas tasas de mortalidad natural (Aliaga et al., 2001) y marcadamente influenciada por factores ambientales (bióticos y abióticos) en todas las etapas de su ciclo vital (Yañez et al., 1990).
Figura 4. Cardumen de anchoveta de alta densidad
La anchoveta (Engraulis ringens), se distribuye desde Zorritos (4°30’S) al norte del Perú hasta la Isla Chiloé (42º 30’S) (Serra et al., 1979). En esta área de distribución se distinguen tres unidades de stock: norte-centro de Perú, sur de Perú-norte de Chile y centro-sur de Chile (Alheit & Ñiquen, 2004). Las unidades de stock del norte-centro de Perú (03°30’- 16°00’S) y del sur de Perú-norte de Chile (16°01’ – 24°00S), registran las mayores concentraciones (Fig. 5).

15
Figura 5. Distribución de la anchoveta (Engraulis ringens) en Perú y norte de Chile.
La actividad reproductiva de la anchoveta abarca casi todo el año; Sin embargo, existen dos periodos de mayor intensidad, el principal en invierno primavera (julio-septiembre) y el otro en el verano (febrero-marzo) (FAO, 1988). Con respecto a la talla de primera madurez sexual, se estima que se produce a los 12 cm (Cubillos & Arancibia, 1993)
Cubillos et al.(1999) señalan que la estrategia reproductiva de este recurso consiste
en desovar al término del invierno (agosto-septiembre) cuando las condiciones oceanográficas generan convergencia debido a la dominancia de vientos norte que permiten el transporte hacia la costa, lo que favorece la retención de huevos. Sin embargo en menos de un mes estas condiciones cambian a un sistema de surgencia, favoreciendo la producción de alimento y su concentración en zonas costeras, y la sobrevivencia de larvas y juveniles (Castro et al., 2000).
3.6 De la pesquería En Chile la anchoveta es uno de los recursos de mayor importancia para la actividad pesquera. A lo largo de las costas de nuestro país existen 2 unidades de pesquerías que explotan este recurso. Unidades que se encuentran en la zona norte (I y II Regiones) y en la zona centro-sur (V a X Regiones).

16
Esta especie se encuentra en régimen de plena explotación y anualmente se determina una cuota de captura por unidad de pesquería (I-II, III-IV y V-X región). Esta cuota está subdividida por pesquería industrial y artesanal, posteriormente se prorratea por zona y período. 3.7 Estadios tempranos de peces teleósteos El estudio de las fases de vida temprana de los peces marinos resulta interesante y fundamental para entender la dinámica de las poblaciones y por ende desarrollar un mejor manejo de los recursos pesqueros (Fiuman, 2002). Existe gran variabilidad en la supervivencia durante este periodo de desarrollo, la cual está relacionada con la variabilidad en el reclutamiento, siendo este último determinante de la fuerza de las clases anuales, y por lo tanto del tamaño de la población. Por esta razón existe un gran interés económico y científico por reconocer los factores y condiciones que influyen en el reclutamiento durante las fases de vida temprana (Plaza & Cerna, 2011).
Existen varias hipótesis propuestas para explicar las altas tasas de mortalidad en los estadios de vida temprana, las cuales se describen a continuación. La primera de ellas es la propuesta por Hjort (1914) y se le conoce con el nombre, de “periodo crítico” Esta se relaciona con el periodo después de la absorción del saco vitelino, donde las larvas no son capaces de encontrar alimento para su primera alimentación y perecen inevitablemente por inanición. Durante este periodo ocurre el 99% de mortalidad de la progenie. La siguiente hipótesis es denominada “match-mismatch” por Cushing (1975), quien propuso que muchos peces han adaptado sus ciclos reproductivos a los ciclos de producción planctónica, para garantizar la sobrevivencia de la progenie. Es decir, si el tiempo de alimentación no coincide con la estación de producción planctónica, se gatillan clases anuales muy débiles. De forma paralela Lasker (1975) propuso que la densidad de las presas en el plancton es crucial para la sobrevivencia larval. Esta hipótesis establece que los parches de presas planctónicas se desestabilizan por condiciones oceanográficas desfavorables, aumentando considerablemente la mortalidad larval. En caso contrario, las condiciones del ambiente calmas y estables permiten zonas favorables para la progenie, con alta concentración de alimento favoreciendo la sobrevivencia. Parrish (1976) propuso la teoría de la “dispersión larval”, causada por las corrientes que desplazan la progenie fuera de las zonas de alimentación, provocando la muerte de las larvas por inanición. Bailey & Houde (1989) propusieron la hipótesis de la “predación larval”, en la que establecen que las fluctuaciones de las clases de peces están directamente relacionadas con las variaciones en la abundancia de los predadores de los estadios tempranos de un especie determinada. Cury & Roy (1989) propusieron la “ventana ambiental optima”, relacionando directamente los reclutamientos y los factores ambientales en áreas de surgencia.

17
Estudios recientes han sugerido que existe una conexión entre el crecimiento y la mortalidad, la cual puede explicar la supervivencia de los estadios tempranos de peces teleósteos. Esta hipótesis sugiere que el crecimiento opera bajo la acción de cuatro mecanismos, los cuales no son necesariamente mutualmente excluyentes. El primero de estos sugiere que si la mortalidad decrece funcionalmente con el tamaño del pez, entonces aquellos individuos más grandes a cualquier edad definida, tendrían menor probabilidad de mortalidad que aquellos más pequeños (bigger-is-better mechanism; Leggett & DeBlois, 1994). (Fig. 6)
Figura 6. Ilustración del mecanismo “bigger-is-better”, donde individuos de una misma edad difieren en tamaño.
El segundo mecanismo establece que si la mortalidad es tamaño-dependiente, entonces las tasas de crecimiento mayores podrían reducir el tiempo en el cual los individuos experimentan elevadas tasas de mortalidad. En otras palabras, una cohorte que exhibe tasas de crecimientos lento, puede tener un período larvario prolongado y una mortalidad acumulativa muy alta. En contraste aquellos individuos que crecen rápidamente tienen mayor probabilidad de sobrevivir (“growth-rate” mechanism; Shepherd & Cushing, 1980).
El tercer mecanismo sugiere que aquellos individuos que hacen las transiciones ontogenéticas (e.g., saco vitelino a primera alimentación, larva a juvenil) a edades mas tempranas, tienen mayor probabilidad de sobrevivir y viceversa (“stage duration” mechanism"; Houde, 1997).
Finalmente, el mecanismo propuesto por Takasua et al. (2003; 2004) denominado
“crecimiento selectivo”, establece que las larvas que crecen mas rápidamente tienen mas

18
probabilidades de sobrevivir que aquellas de crecimiento lento. Sin embargo este proceso puede estar mediado por el efecto de la “predación”. Existen ocasiones en donde los predadores pueden preferir larvas de crecimiento lento y mal nutridas, como también casos en que para los predadores larvas más grandes y robustas sean más fáciles de detectar.

19
3.8 Pesquerías y ambiente 3.8.1 El Niño Oscilación del Sur (ENOS) El ENOS corresponde a un evento de interacción océano-atmosférico que se presenta en el océano Pacífico ecuatorial central (Maturana et al., 2004). Este Consiste en una oscilación entre una fase cálida y una fase fría, El Niño y La Niña respectivamente (McPhaden, 1999), denominada Oscilación del Sur (OS), Esta se manifiesta principalmente por el calentamiento o enfriamiento anormal de la temperatura superficial del mar (TSM).
La etapa inicial de ENOS (El Niño) comienza con el intercambio de masa de aire entre los océanos Indico y Pacífico que invierte la dirección de los vientos predominantes (alisios), los cuales se debilitan a gran escala (Maturana et al., 2004), Esto provoca transmisión de energía desde la atmosfera hacia el océano, desencadenando el calentamiento de las aguas que se desplazan a través de ondas Kelvin, las cuales arriban a las costas sudamericanas del Pacífico (Cazeneuve et al., 2000) (Fig. 7).
Figura 7. Aumento de la TSM en las costas sudamericanas correspondiente a la llegada del fenómeno de El Niño.
El calentamiento de las aguas al que se ha hecho referencia, está asociado con el aumento en la temperatura superficial del mar (TSM), el cual se manifiesta a través de anomalías positivas de la TSM las que se obtienen por la diferencia entre el valor observado y el valor histórico del lugar en el que se ha realizado la medición.
En esta situación inestable del sistema Océano-Atmósfera, El Niño puede ser
caracterizado por sus efectos principales. Los más importantes que se presentan en las costas occidentales de Sudamérica (lado este del Pacífico), son los siguientes:

20
Debilitamiento de los vientos alisios, Disminución de la presión atmosférica Aumento de la temperatura superficial del mar (TSM) y del aire. Profundización de la termoclina (zona que separa las aguas superficiales y
profundas). Cambios en la disponibilidad y distribución de los recursos marinos.
3.8.2 Efectos de El Niño en la pesquería de la anchoveta La variabilidad ambiental causada por el ENOS juega un papel crítico en la abundancia y distribución de los recursos pelágicos. Esto depende en gran medida de la duración del evento, la intensidad, la época de ocurrencia, y en especial de la composición faunística presente en el ecosistema al tiempo de su ocurrencia (Avaria et al., 2004).
Son varias las maneras en que El Niño puede afectar un recurso pesquero. La principal es el efecto sobre las surgencias normalmente son muy ricas en nutrientes. Debido a las masas de agua caliente estas se empobrecen y se produce una falla en la cadena trófica El mayor efecto que produce el evento de El Niño en la anchoveta (Engraulis ringens) se ve reflejado en los drásticos cambios de disponibilidad, accesibilidad y abundancia. Cuando la TSM se incrementa la anchoveta se repliega hacia la costa y se profundiza, observándose un comportamiento netamente demersal (Sosa, 1999). 3.8.3 La fase fría del ciclo ENOS La fase fría del ENOS (La Niña) corresponde a un evento de interacción océano-atmósfera que se caracteriza por la aparición de temperaturas oceánicas superficiales inusualmente bajas en el centro y este del Océano Pacífico ecuatorial (Fig. 8).
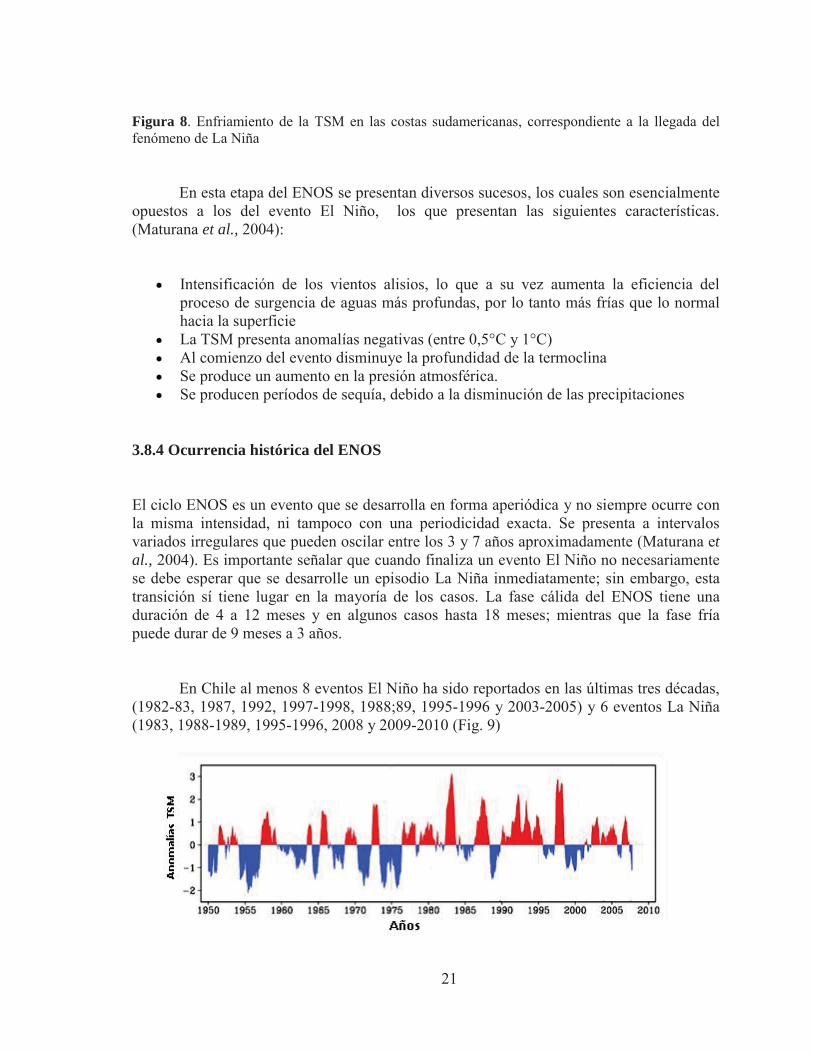
21
Figura 8. Enfriamiento de la TSM en las costas sudamericanas, correspondiente a la llegada del fenómeno de La Niña
En esta etapa del ENOS se presentan diversos sucesos, los cuales son esencialmente
opuestos a los del evento El Niño, los que presentan las siguientes características. (Maturana et al., 2004):
Intensificación de los vientos alisios, lo que a su vez aumenta la eficiencia del proceso de surgencia de aguas más profundas, por lo tanto más frías que lo normal hacia la superficie
La TSM presenta anomalías negativas (entre 0,5°C y 1°C) Al comienzo del evento disminuye la profundidad de la termoclina Se produce un aumento en la presión atmosférica. Se producen períodos de sequía, debido a la disminución de las precipitaciones
3.8.4 Ocurrencia histórica del ENOS El ciclo ENOS es un evento que se desarrolla en forma aperiódica y no siempre ocurre con la misma intensidad, ni tampoco con una periodicidad exacta. Se presenta a intervalos variados irregulares que pueden oscilar entre los 3 y 7 años aproximadamente (Maturana et al., 2004). Es importante señalar que cuando finaliza un evento El Niño no necesariamente se debe esperar que se desarrolle un episodio La Niña inmediatamente; sin embargo, esta transición sí tiene lugar en la mayoría de los casos. La fase cálida del ENOS tiene una duración de 4 a 12 meses y en algunos casos hasta 18 meses; mientras que la fase fría puede durar de 9 meses a 3 años.
En Chile al menos 8 eventos El Niño ha sido reportados en las últimas tres décadas,
(1982-83, 1987, 1992, 1997-1998, 1988;89, 1995-1996 y 2003-2005) y 6 eventos La Niña (1983, 1988-1989, 1995-1996, 2008 y 2009-2010 (Fig. 9)

22
Figura 9. Anomalías de la TSM estandarizadas de la base de datos de NOAA.

23
4. METODOLOGÍA
4.1 Aspectos generales El desarrollo y cumplimiento de los objetivos de este estudio se llevó a cabo bajo el marco del proyecto “Validación de formación de anillos primarios y macro-anillos de crecimiento en otolitos de anchoveta de la zona norte” ID: N° 4728-31 LP 11 SUBPESCA.
La información utilizada en este estudio fue otorgada por el Instituto de Fomento
pesquero (IFOP) que, puso a disposición datos y muestras históricas de otolitos correspondientes a los años 1972-1973 y 1982-1983, colectados en distintos períodos relacionados con el evento ENSO. Mientras que las muestras de los años 2009-2010 son provenientes del muestreo regular que realiza el IFOP en la pesquería de la zona norte 18°21’03’’S y 24°00'00’’S desde la costa hasta las 200 millas náuticas (Tabla 3). Tabla 3. Zona de recolección de ejemplares juveniles de anchoveta en la zona Norte de Chile
ZONA Límite Norte - Limite sur
Arica 18° 21' 03'' S - 19° 30' 00'' S Iquique 19° 30' 00'' S - 21° 25' 00'' S Antofagasta 21° 26' 00'' S - 24° 00' 00'' S
4.2 Caracterización de la micro-estructura de otolitos Para una adecuada caracterización de la micro-estructura de otolitos juveniles se requirió de secciones con una elevada resolución, permitiendo una clara identificación de los micro-incrementos diarios. Para este propósito se realizó un proceso de preparación de otolitos que se llevó a cabo utilizando el método SGEM propuesto por Plaza et al., (2005), el cual se describe a continuación:
4.2.1 Aplicación del protocolo SGEM para preparación de otolitos En primer lugar, fue necesario extraer los otolitos sagitales de su sistema de almacenamiento, el cual consistía en dispensadores llamados comúnmente “cartones”,

24
encontrándose cada par de otolito fijo a este cartón con resina epóxica. Los otolitos fueron cuidadosamente removidos por medio de acetona diluida al 10%. Posteriormente se extrajo el otolito izquierdo del par, el cual fue inmediatamente fijado mediante protocolo SGEM. Una vez finalizada la remoción y para evitar el desprendimiento o pérdida del otolito derecho, la estructura calcárea se almacenó en un nuevo “cartón” (Fig. 10).
Figura 10. Sistema de almacenamiento de otolitos, denominado “cartones”
Para el montaje de los otolitos, inicialmente se etiquetó el portaobjetos con su ID respectivo, código de zona, mes y año. Posteriormente se calentó a una temperatura de 90 a 150°C, y se frotó ligeramente una barra de cera termoplástica, la que al contacto con el portaobjetos caliente se derritió formando un delgado residuo, el que se fijó una vez enfriado. Se mezclaron en partes similares resina epóxica de dos componentes (Araldite 2011 A y B), procurando que la unión fuese lo más lenta y cuidadosa posible, para así evitar la formación de burbujas. Una vez mezclados homogéneamente los componentes, se tomó una pequeña porción, depositándola en el portaobjetos previamente preparado con la cera termoplástica.
El procedimiento básico del SGEM consiste en embeber el otolito en la resina. Cabe
mencionar que antes de depositar la resina en el portaobjetos ésta debe pasar por la llama de una lámpara de alcohol (mechero), para eliminar la mayor parte de las burbujas que quedaron al mezclar ambos componentes. Los otolitos pequeños fueron fácilmente embebidos en una gota de resina, mientras que los de mayor tamaño requirieron mayor cantidad. La precaución más importante es asegurar que los otolitos quedaran completamente cubiertos por la resina. Luego se deja a temperatura ambiente hasta que la mezcla se fija completamente (Fig. 11).

25
Figura 11. Ilustración método SGEM: para la preparación de otolitos para el análisis de micro-incrementos. A: aplicación de cera termoplástica; B: resina de dos componentes en partes iguales; C: mezcla de los componentes de resina epóxica; D y E: montaje del otolito (Fuente: Plaza et al., 2005). 4.2.2 Proceso de pulido de otolito Los otolitos fijados fueron pulidos manualmente con lijas de diferentes granos (diámetro: 1000 -1200 y 5 – 12 μm), en posición sagital por ambas caras, hasta obtener la mejor resolución del primordio en la cara cóncava, teniendo la precaución de no sobre-pulir los bordes y perder la resolución de estos en la cara convexa.
Los otolitos utilizados en este estudio no fueron suficientemente transparentes como para diferenciar los micro-incrementos diarios a simple vista (Fig.12A). El objetivo principal fue obtener un área claramente visible de los anillos (Fig.12 B), para su posterior lectura. Para esto se procedió a evaluar el proceso del pulido mediante la obtención de fotografías constante por medio del microscopio, con el propósito de evitar el sobre pulido y perder la resolución de los micro-incrementos. (Fig.12 C).
c
Figura 12. A: fotografía bajo luz trasmitida de un otolito sin pulir, donde no se diferencian los micro-incrementos diarios, con aumento de 400X. B: fotografía de otolito pulido, donde pueden

26
observarse claramente los anillos diarios. C: imagen de otolito sobre pulido, en donde se pierde completamente la resolución de sus micro-incrementos.
27
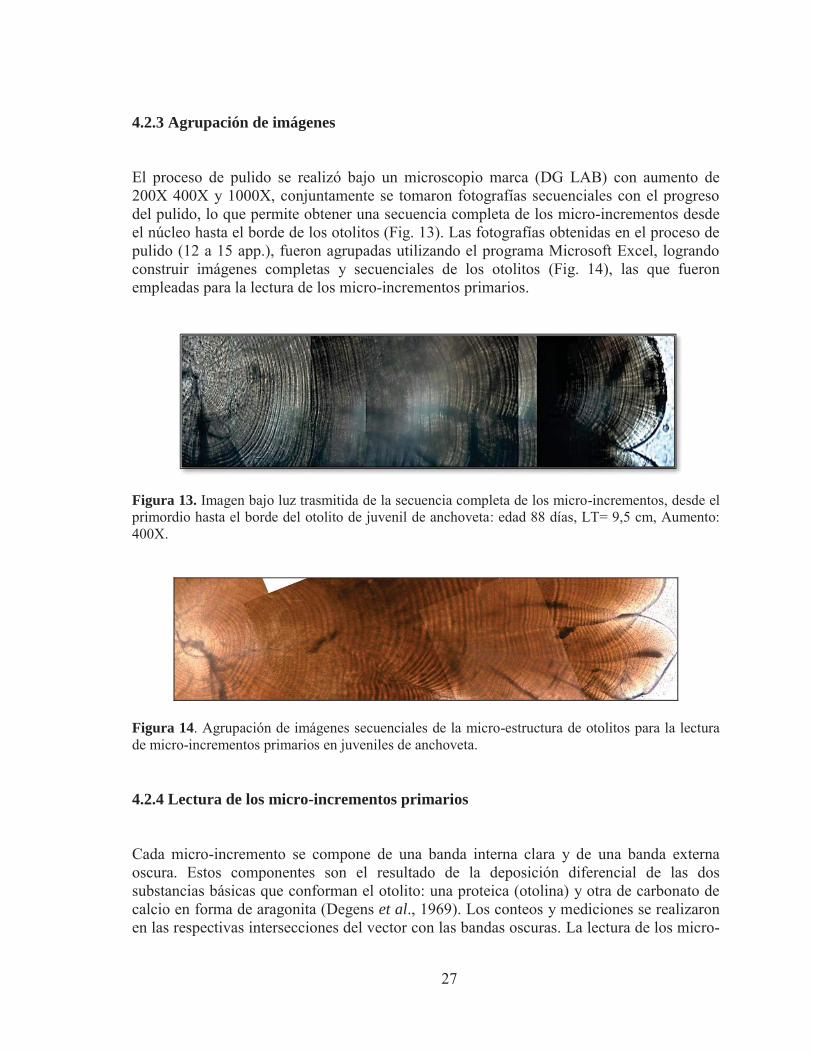
4.2.3 Agrupación de imágenes
El proceso de pulido se realizó bajo un microscopio marca (DG LAB) con aumento de 200X 400X y 1000X, conjuntamente se tomaron fotografías secuenciales con el progreso del pulido, lo que permite obtener una secuencia completa de los micro-incrementos desde el núcleo hasta el borde de los otolitos (Fig. 13). Las fotografías obtenidas en el proceso de pulido (12 a 15 app.), fueron agrupadas utilizando el programa Microsoft Excel, logrando construir imágenes completas y secuenciales de los otolitos (Fig. 14), las que fueron empleadas para la lectura de los micro-incrementos primarios.
Figura 13. Imagen bajo luz trasmitida de la secuencia completa de los micro-incrementos, desde el primordio hasta el borde del otolito de juvenil de anchoveta: edad 88 días, LT= 9,5 cm, Aumento: 400X.
Figura 14. Agrupación de imágenes secuenciales de la micro-estructura de otolitos para la lectura de micro-incrementos primarios en juveniles de anchoveta.
4.2.4 Lectura de los micro-incrementos primarios Cada micro-incremento se compone de una banda interna clara y de una banda externa oscura. Estos componentes son el resultado de la deposición diferencial de las dos substancias básicas que conforman el otolito: una proteica (otolina) y otra de carbonato de calcio en forma de aragonita (Degens et al., 1969). Los conteos y mediciones se realizaron en las respectivas intersecciones del vector con las bandas oscuras. La lectura de los micro-

28
incrementos primarios (MP) fue realizada por dos lectores, con el objetivo de establecer concordancia y exactitud en la identificación de los MP. Para esto se utilizaron imágenes digitalizadas de los otolitos con aumento de 200X y 400X.
Los micro-incrementos fueron medidos mediante la proyección de una línea recta,
en el sector donde se observó la mayor resolución posible, desde el núcleo hasta el borde del otolito, utilizando para la medición la herramienta “Caliper” del analizador de imágenes IMAGE-Pro.Plus (Fig. 15).
Figura 15. Conteo de anillos y medición de radios de un otolito juvenil de anchoveta a través del programa analizador de imágenes.
Para evaluar diferencias entre las lecturas de micro-incrementos diarios efectuada por los dos lectores se realizó un análisis multivariante de medidas repetidas (ANDEVA–RM), utilizando el programa estadístico SPSS. Para este análisis se utilizó como estadístico de prueba Lambda de Wilks (λ), el cual mide las desviaciones dentro de cada grupo respecto de las desviaciones totales, con la siguiente ecuación:
donde:
S.C.R : Suma de los cuadrados intra grupos S.C.T : Suma de los cuadrados totales

29
Hipótesis: Se rechaza la hipótesis nula H0 que expresa que no existen diferencias significativas entre los lectores si el valor p, asociado al resultado observado, es igual o menor que el nivel de significación establecido (0,05).

30
La fecha de nacimiento fue estimada restando la edad total observada (en días) a la fecha de captura, lo que permitió diferenciar las cohortes mes de nacimiento para efectuar las comparaciones. Cabe señalar que todas las mediciones realizadas se efectuaron en el otolito izquierdo, ya que investigaciones previas en la materia señalan que no existen diferencias significativas entre ambos, dejando el otolito derecho de respaldo en caso de ser requerido. (Carreño, 2010; Proyecto FIP 2009-17) 4.3 Crecimiento y ambiente 4.3.1 Crecimiento de juveniles de anchoveta en presencia de eventos El Niño y La Niña.
Para identificar diferencias significativas entre el crecimiento de juveniles menores o iguales a 12 cm entre eventos El Niño (1972-1973, 1982-1983) y La Niña (2009-2010) se desarrollaron tres procedimientos: el primero consistió en comparar la relación talla-edad a la fecha de captura para el rango de edad y longitud de los individuos muestreados, el segundo correspondió a una comparación de los grosores de los micro-incrementos diarios como proxy del crecimiento somático y el tercero tiene relación con la comparación de las tasas de crecimiento, las cuales fueron estimadas a través de técnicas de retro-cálculo.
El tamaño de muestra disponible para el presente estudio permitió realizar dos análisis uno global y otro estacional, los cuales se aplicaron en todos los procedimientos propuestos para evaluar y comparar el crecimiento de los ejemplares estudiados. Análisis Global: En este análisis se trabajó con todos los ejemplares nacidos durante cada uno de los eventos, siendo estos El Niño 1972-1973 y 1982-1983, y La Niña 2009-2010. Análisis Estacional: En el análisis estacional se agruparon todos los ejemplares nacidos en una misma estación dentro de cada uno de los eventos. Para este caso la estación de verano fue la que logró el mayor tamaño de muestra, por lo que se decidió trabajar sólo con esa cohorte.

31
4.3.2 Procedimiento para la evaluación y comparación del crecimiento Relación talla-edad En este procedimiento inicialmente se realizó un análisis global para obtener una proximidad del comportamiento de la relación talla-edad en cada evento. Para ello fue necesario reconstruir los patrones de crecimiento diario, objetivo que se llevó a cabo utilizando técnicas de retro-cálculo para estimar la longitud al tiempo de formación de cada una de las marcas presentes en los otolitos (Ricker, 1975). El método utilizado para tal propósito fue el del intercepto biológico, el cuál emplea un intercepto determinado biológicamente (Campana, 1990), que se define por la siguiente ecuación:
donde: Li : Longitud del pez al momento de formación del i-ésimo día
Lc : Longitud del pez al momento de la captura Ri : Tamaño del otolito al momento de formación del i-ésimo día Rc : Tamaño del otolito al momento de la captura
Rm : Tamaño del otolito al momento de la eclosión Lo : Longitud del pez al momento de la eclosión
La longitud al momento de la eclosión que se utilizó fue 2,5 mm, definido por Llanos-Rivera y Castro (2006) para la zona de Antofagasta (23°) a una temperatura de 15°C. Una vez que se logró reconstruir las longitudes previas, éstas fueron ajustadas a un modelo de crecimiento Laird-Gompertz de la siguiente manera:
donde:

32
L : Longitud asintótica α: Tasa de crecimiento instantánea X0: Edad al punto de inflexión
Con los parámetros entregados por el modelo se obtuvo la tasa de crecimiento instantánea (TICD) y la tasa de crecimiento absoluta al punto de inflexión (punto que marca el mayor crecimiento TCApi), las cuales se estimaron de la siguiente manera:
Los modelos se ajustaron siguiendo dos criterios, en el primero se modeló la
relación talla-edad desde la marca de eclosión hasta los primeros 130 días de vida y el segundo se modeló sólo la fracción juvenil a partir de los 40 días de vida (40-130). Esta doble aproximación fue necesaria debido que el retro-calculo depende de una relación lineal significativa entre el tamaño del otolito y el tamaño del pez, relación que sólo ha sido validada para la etapa juvenil en esta especie. En el caso de las estimaciones de las tasas de crecimiento modelando la relación talla-edad desde 0 a 130 días, se asumió que la relación entre el tamaño del otolito fue lineal desde larva a juvenil. 4.4 Análisis estadístico comparativo del crecimiento 4.4.1 Comparación de los modelos de crecimiento Los parámetros de crecimiento fueron estimados utilizando el método Dual Quasi-Newton, implementado en el módulo PROCNLMIXED del Programa Estadístico SAS Inc (2007). La bondad de ajuste de cada modelo fue evaluada examinando el cuadrado medio residual y el coeficiente de determinación (r2). Los parámetros L , y X0 fueron comparados usando el módulo "Contrast Statement” en PROCNLMIXED, en el cual el “Test-F” usa el método delta para aproximar matriz de varianza-covarianza. 4.4.2 Comparación grosor del micro-incremento Los grosores de los micro-incrementos de los ejemplares estudiados fueron calculados por medio de los radios de los micro-incrementos diarios, los que se obtuvieron a través de la

33
herramienta “Caliper” del analizador de imágenes IMAGE-Pro.Plus. Dicho cálculo se determinó mediante la siguiente formula:
donde:
Rd(i) : Radio micro-incremento a la edad i. Para este objetivo, inicialmente se compararon los grosores de los micro-incrementos de una misma cohorte mensual de los dos eventos El Niño 1973 y 1982 para determinar significativas entre un mismo evento, para luego comparar puntualmente los grosores de una cohorte mensual entre un evento El Niño y un evento La Niña. Para esto se realizó un análisis de varianza de medidas repetidas (ANDEVA-RM), siguiendo los procedimientos descritos por Chambers & Miller (1995). El análisis correspondió a un diseño mixto de dos factores inter-sujeto (evento-año) e intra-sujeto (grosor micro-incremento), con los individuos agrupados en filas y los grosores como factor de repetición. Para disminuir el elevado número de repeticiones el análisis se realizó a intervalos de 5 días; 5, 10, 15, 20, 25, 30, 35, 40……etc. La última repetición correspondió al juvenil de menor edad recolectado.
En las Tablas 4 y 5 se puede observar la demostración del diseño balanceado del
ANDEVA-RM para la comparación de los grosores entre eventos Niños y entre eventos Niño y Niña Tabla 4. Diseño balanceado de análisis de varianza de medidas repetidas (ANDEVA-RM)) para determinar diferencias en los grosores entre cohortes de años con eventos El Niño
Factores inter-sujeto Factores intra-sujetos: Grosor micro-incremento diario
Evento Año Casos Dia 5
Día 10
Dia 15
Dia 20
Día 25
Día 30
Día 35
Día 40
Día 45
Día 50
Día 55
Día 60
El Niño 1973 C1 x x x x x x x x x x X x C1 x x x x x x x x x x X x Cn x x x x x x x x x x X x
El Niño 1982 C1 x x x x x x x x x x X x C1 x x x x x x x x x x X x Cn x x x x x x x x x x X x
34
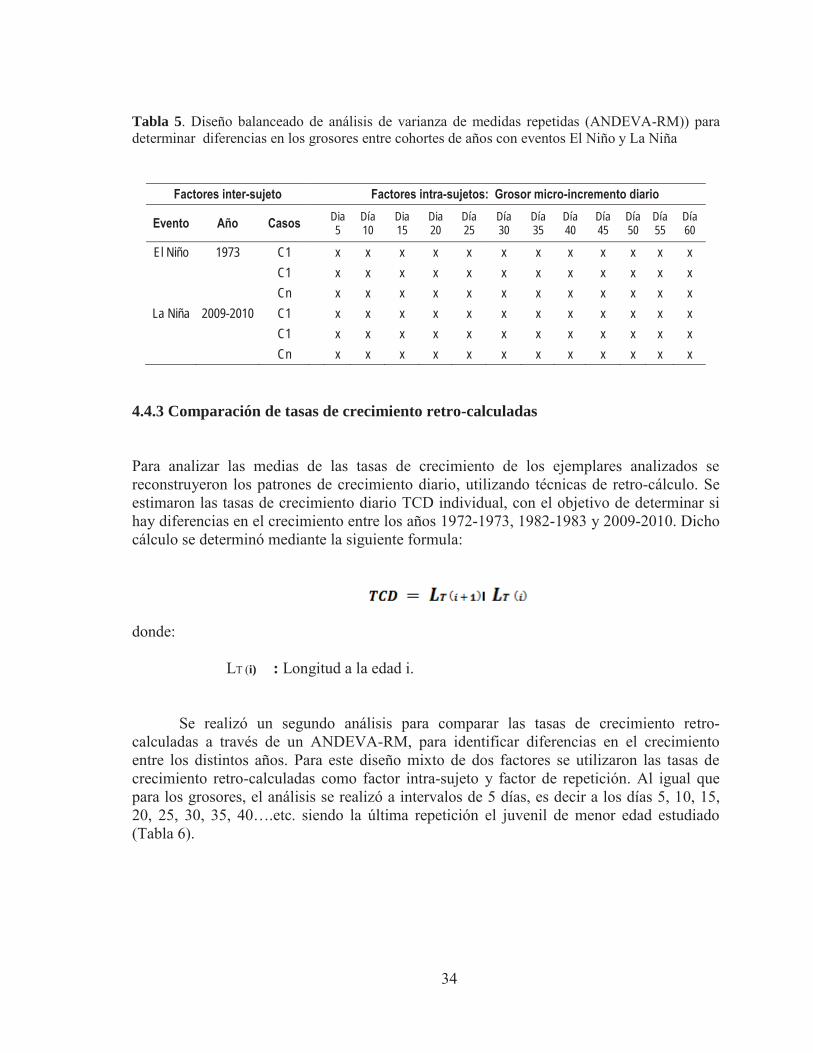
Tabla 5. Diseño balanceado de análisis de varianza de medidas repetidas (ANDEVA-RM)) para determinar diferencias en los grosores entre cohortes de años con eventos El Niño y La Niña
Factores inter-sujeto Factores intra-sujetos: Grosor micro-incremento diario
Evento Año Casos Dia 5
Día 10
Dia 15
Dia 20
Día 25
Día 30
Día 35
Día 40
Día 45
Día 50
Día 55
Día 60
El Niño 1973 C1 x x x x x x x x x x x x C1 x x x x x x x x x x x x Cn x x x x x x x x x x x x
La Niña 2009-2010 C1 x x x x x x x x x x x x C1 x x x x x x x x x x x x Cn x x x x x x x x x x x x
4.4.3 Comparación de tasas de crecimiento retro-calculadas Para analizar las medias de las tasas de crecimiento de los ejemplares analizados se reconstruyeron los patrones de crecimiento diario, utilizando técnicas de retro-cálculo. Se estimaron las tasas de crecimiento diario TCD individual, con el objetivo de determinar si hay diferencias en el crecimiento entre los años 1972-1973, 1982-1983 y 2009-2010. Dicho cálculo se determinó mediante la siguiente formula:
donde:
LT (i) : Longitud a la edad i.
Se realizó un segundo análisis para comparar las tasas de crecimiento retro-calculadas a través de un ANDEVA-RM, para identificar diferencias en el crecimiento entre los distintos años. Para este diseño mixto de dos factores se utilizaron las tasas de crecimiento retro-calculadas como factor intra-sujeto y factor de repetición. Al igual que para los grosores, el análisis se realizó a intervalos de 5 días, es decir a los días 5, 10, 15, 20, 25, 30, 35, 40….etc. siendo la última repetición el juvenil de menor edad estudiado (Tabla 6).
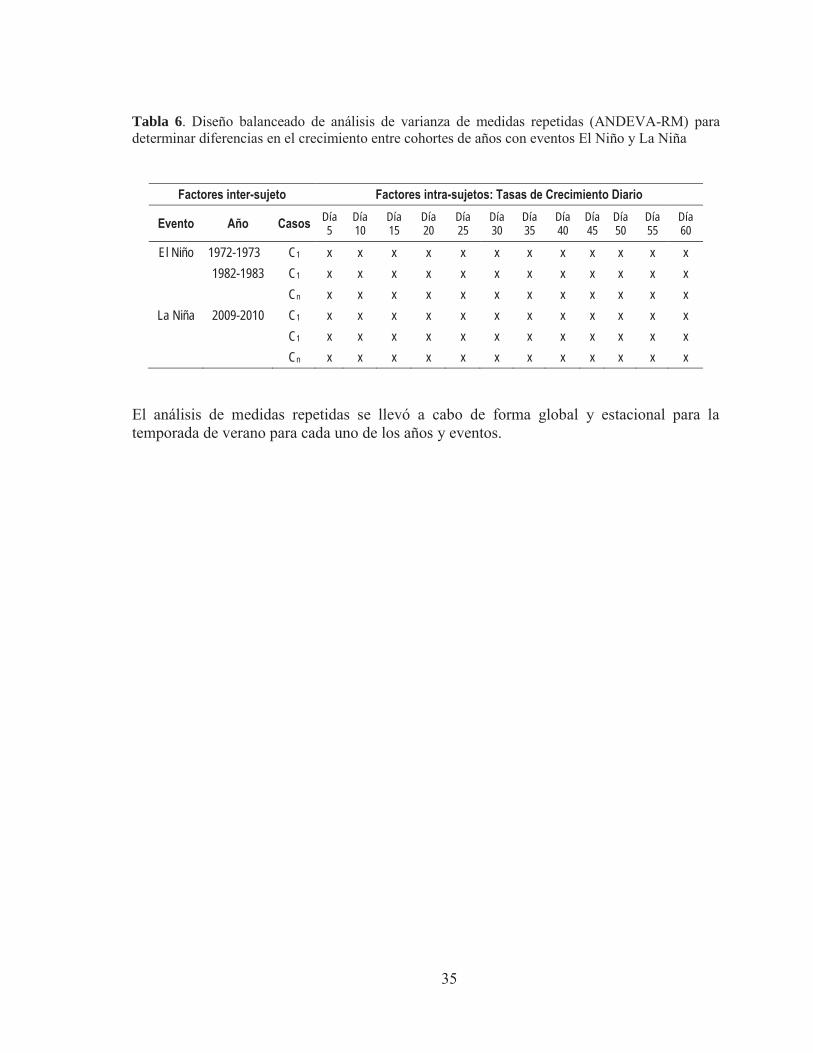
35
Tabla 6. Diseño balanceado de análisis de varianza de medidas repetidas (ANDEVA-RM) para determinar diferencias en el crecimiento entre cohortes de años con eventos El Niño y La Niña
Factores inter-sujeto Factores intra-sujetos: Tasas de Crecimiento Diario
Evento Año Casos Día 5
Día 10
Día 15
Día 20
Día 25
Día 30
Día 35
Día 40
Día 45
Día 50
Día 55
Día 60
El Niño 1972-1973 C1 x x x x x x x x x x x x 1982-1983 C1 x x x x x x x x x x x x Cn x x x x x x x x x x x x
La Niña 2009-2010 C1 x x x x x x x x x x x x C1 x x x x x x x x x x x x Cn x x x x x x x x x x x x
El análisis de medidas repetidas se llevó a cabo de forma global y estacional para la temporada de verano para cada uno de los años y eventos.

36
5. RESULTADOS
5.1 Estadísticas Generales
Los juveniles analizados en este estudio fueron 376 ejemplares, con tallas que
fluctuaron entre los 5,5 y 12 cm de longitud total y edades que variaron entre 60 y 178 días. Del total, 276 correspondieron al evento La Niña del periodo 2009-2010, 56 del evento El Niño de 1982 y 44 al evento El Niño de 1973. El detalle del tamaño de muestra, longitud media, edad media y área de estudio se presenta en la Tabla 7.
Tabla 7. Otolitos de anchovetas juveniles analizados por año, área y mes para el análisis de micro-incrementos.
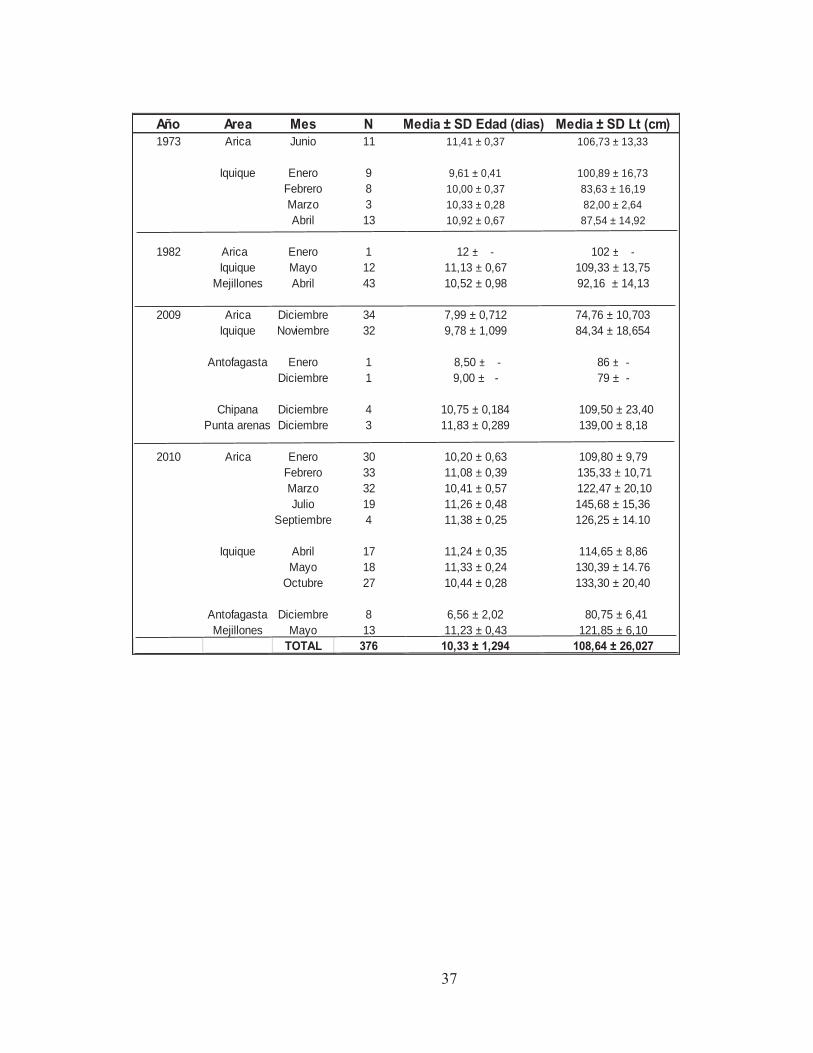
37
Año Area Mes N Media ± SD Edad (dias) Media ± SD Lt (cm)1973 Arica Junio 11 11,41 ± 0,37 106,73 ± 13,33
Iquique Enero 9 9,61 ± 0,41 100,89 ± 16,73Febrero 8 10,00 ± 0,37 83,63 ± 16,19Marzo 3 10,33 ± 0,28 82,00 ± 2,64Abril 13 10,92 ± 0,67 87,54 ± 14,92
1982 Arica Enero 1 12 ± - 102 ± -Iquique Mayo 12 11,13 ± 0,67 109,33 ± 13,75
Mejillones Abril 43 10,52 ± 0,98 92,16 ± 14,13
2009 Arica Diciembre 34 7,99 ± 0,712 74,76 ± 10,703Iquique Noviembre 32 9,78 ± 1,099 84,34 ± 18,654
Antofagasta Enero 1 8,50 ± - 86 ± -Diciembre 1 9,00 ± - 79 ± -
Chipana Diciembre 4 10,75 ± 0,184 109,50 ± 23,40Punta arenas Diciembre 3 11,83 ± 0,289 139,00 ± 8,18
2010 Arica Enero 30 10,20 ± 0,63 109,80 ± 9,79Febrero 33 11,08 ± 0,39 135,33 ± 10,71Marzo 32 10,41 ± 0,57 122,47 ± 20,10Julio 19 11,26 ± 0,48 145,68 ± 15,36
Septiembre 4 11,38 ± 0,25 126,25 ± 14.10
Iquique Abril 17 11,24 ± 0,35 114,65 ± 8,86Mayo 18 11,33 ± 0,24 130,39 ± 14.76
Octubre 27 10,44 ± 0,28 133,30 ± 20,40
Antofagasta Diciembre 8 6,56 ± 2,02 80,75 ± 6,41Mejillones Mayo 13 11,23 ± 0,43 121,85 ± 6,10
TOTAL 376 10,33 ± 1,294 108,64 ± 26,027
38
5.2 Análisis de la microestructura de otolitos
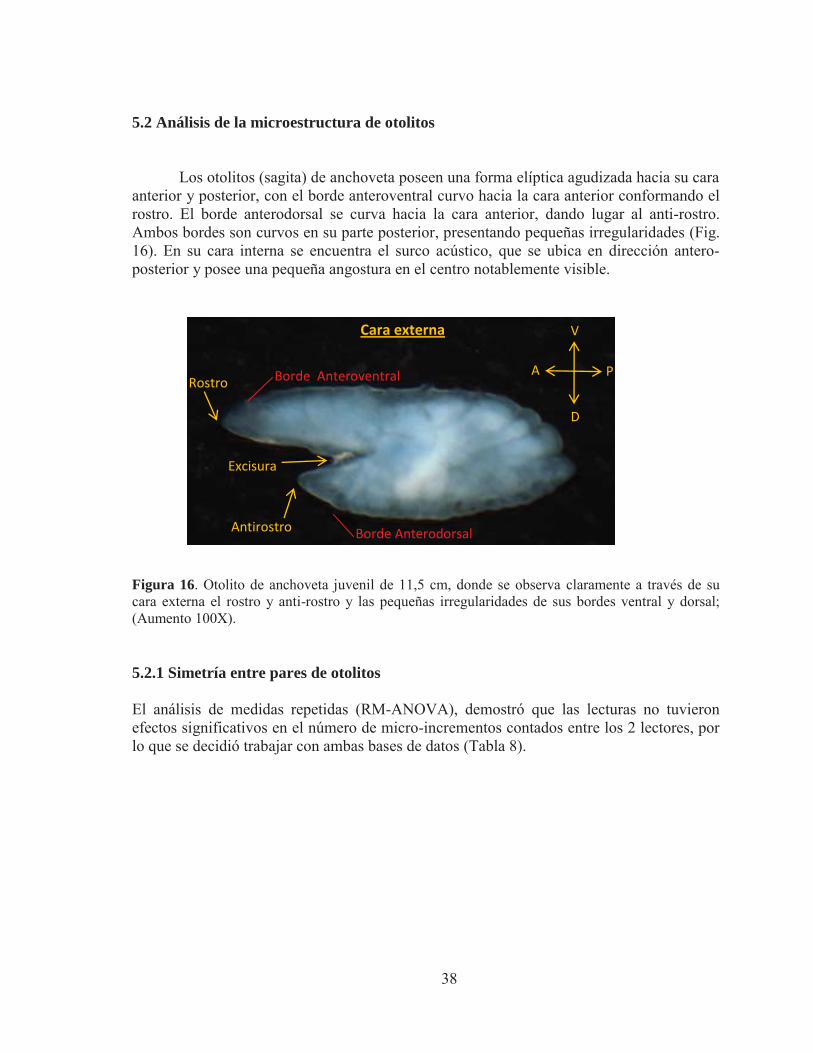
Los otolitos (sagita) de anchoveta poseen una forma elíptica agudizada hacia su cara
anterior y posterior, con el borde anteroventral curvo hacia la cara anterior conformando el rostro. El borde anterodorsal se curva hacia la cara anterior, dando lugar al anti-rostro. Ambos bordes son curvos en su parte posterior, presentando pequeñas irregularidades (Fig. 16). En su cara interna se encuentra el surco acústico, que se ubica en dirección antero-posterior y posee una pequeña angostura en el centro notablemente visible.
Figura 16. Otolito de anchoveta juvenil de 11,5 cm, donde se observa claramente a través de su cara externa el rostro y anti-rostro y las pequeñas irregularidades de sus bordes ventral y dorsal; (Aumento 100X).
5.2.1 Simetría entre pares de otolitos El análisis de medidas repetidas (RM-ANOVA), demostró que las lecturas no tuvieron efectos significativos en el número de micro-incrementos contados entre los 2 lectores, por lo que se decidió trabajar con ambas bases de datos (Tabla 8).
V
D
P A Rostro
Antirostro
Excisura
Cara externa
Borde Anteroventral
Borde Anterodorsal
39
Tabla 8. Análisis multivariado de medidas repetidas para determinar diferencias significativas en las lecturas realizadas por los lectores. Lambda de Wilks= estadístico de prueba multivariado, P> 0,05.
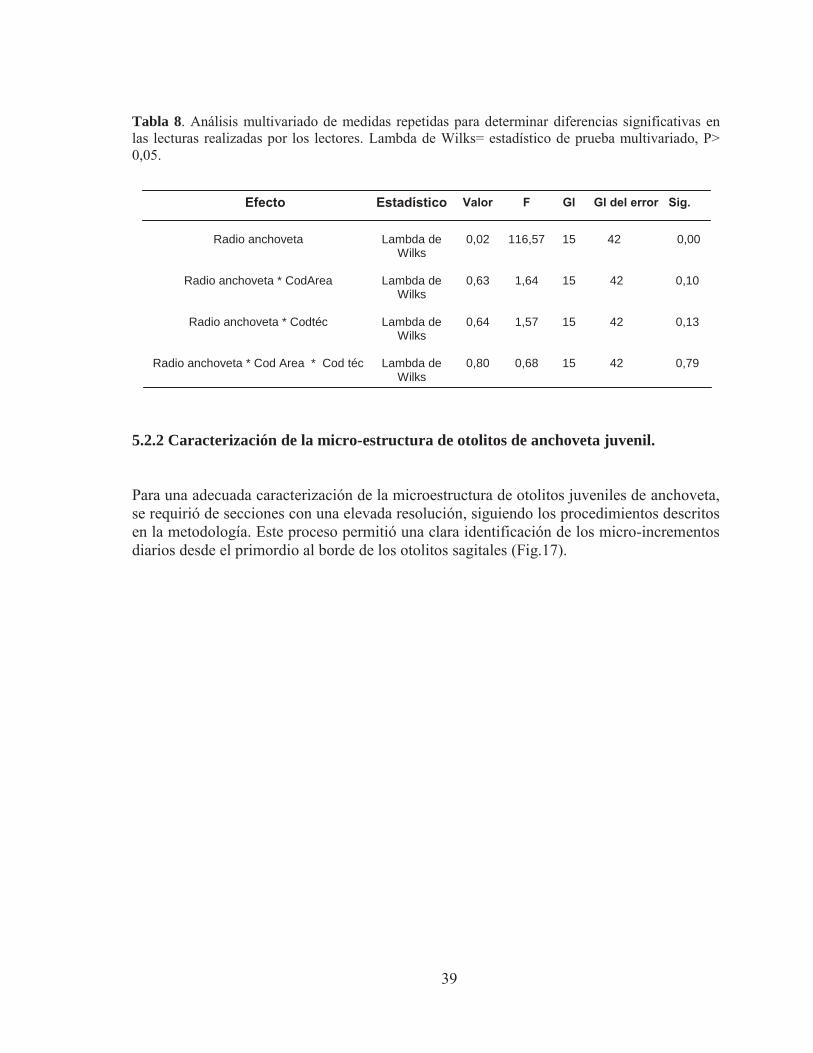
5.2.2 Caracterización de la micro-estructura de otolitos de anchoveta juvenil. Para una adecuada caracterización de la microestructura de otolitos juveniles de anchoveta, se requirió de secciones con una elevada resolución, siguiendo los procedimientos descritos en la metodología. Este proceso permitió una clara identificación de los micro-incrementos diarios desde el primordio al borde de los otolitos sagitales (Fig.17).
Efecto Estadístico Valor F Gl Gl del error Sig.
Radio anchoveta
Lambda de
Wilks 0,02 116,57 15 42 0,00
Radio anchoveta * CodArea
Lambda de
Wilks 0,63 1,64 15 42 0,10
Radio anchoveta * Codtéc
Lambda de
Wilks 0,64 1,57 15 42 0,13
Radio anchoveta * Cod Area * Cod téc
Lambda de
Wilks 0,80 0,68 15 42 0,79
40
Figura 17. Imágenes bajo luz trasmitida agrupadas de la micro-estructura de otolitos para la lectura de micro-incrementos primarios en juveniles de anchoveta: A) edad 117 días, LT= 11,5 cm, aumento 400X, zona Arica, 1973, B) edad 116 días, LT= 11,5 cm, aumento 400X, Zona Mejillones, 1982 y C) edad 104 días, LT= 11 cm, aumento 400X, zona Iquique, 2010) 5.2.3 Primordio
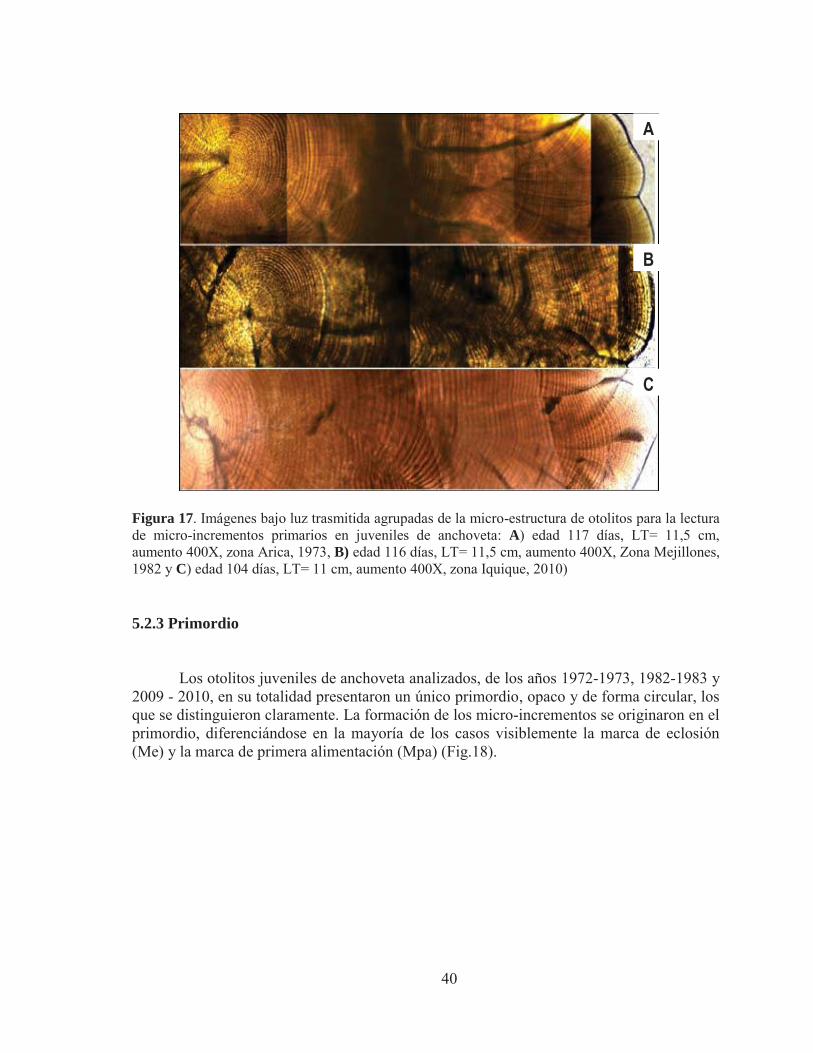
Los otolitos juveniles de anchoveta analizados, de los años 1972-1973, 1982-1983 y
2009 - 2010, en su totalidad presentaron un único primordio, opaco y de forma circular, los que se distinguieron claramente. La formación de los micro-incrementos se originaron en el primordio, diferenciándose en la mayoría de los casos visiblemente la marca de eclosión (Me) y la marca de primera alimentación (Mpa) (Fig.18).
A
B
C

41
A B C
PrMe
Me
Mpa
Figura 18 . Fotografias bajo luz trasmitida del analisis micro-estructural de otolitos juveniles de la zona norte de Chile, donde se observa visiblemente el primordio en los tres ejemplares: A) edad 117 dias, LT = 11 cm, zona Arica, 1973; B) edad 134 dias, LT = 12 cm, zona Iquique, 1982 y C) edad 116 dias, LT = 11,5 cm, zona Iquique, 2010; Pr: Primordio, Me: marca de eclosion y Mpa: marca primera alimentacion (Aumento 400X). 5.2.4 Marca de eclosión y primera alimentación La marca de eclosión en la mayoría de los otolitos estudiados se exhibió de manera muy definida y clara, encontrándose a una distancia media del primordio de 6,89 μm para ejemplares del evento El Niño 1972-1973. Para el evento El Niño 1982-1983 la media fue de 8,19 μm, mientras que para el periodo 2009-2010 perteneciente al evento La Niña la media correspondió a 6,43μm (Tabla 9).
En relación con la marca de primera alimentación, en general fue claramente visible. Sin embargo, en algunos casos se presentó de forma poco clara y más difusa en comparación con la marca de eclosión. La media para las muestras de los años 1972-1973 y 1982-1983 (eventos El Niño) fue de 17,1 μm y 19,6 μm respectivamente. Para el periodo 2009-2010 la media correspondió a 19,3 μm. En los tres eventos esta marca se presentó en promedio a los 3-4 dias de vida.
Tabla 9. Marca de eclosión y de primera alimentación en juveniles de anchoveta asociados a ventos El Niño y La Niña.
Evento / Año Media ± SD ME (μm) Media ± SD MPA (μ El Niño 1972-1973 6,89 ± 2,84 17,1 ± 4,94 El Niño 1982 8,19 ± 3,14 19,6 ± 5,09 La Niña 2009-2010 6,43 ± 0,76 19,3 ± 3,81

42
5.2.5 Grosor de microincrementos
En la mayoría de las muestras analizadas de los eventos El Niño 1972-1973 y 1982-1983 y La Niña 2009-2010, se presentó un patrón de formación similar en los grosores de los micro-incrementos primarios (MP). Por ejemplo, se observó un aumento progresivo en el grosor medio hasta llegar a un plateu a aproximadamete los dos primeros meses de vida, después del cual el grosor medio disminuyó notoriamente en la etapa juvenil tardía.
Los grosores de los micro-incrementos fueron variando en las distintas secciones del
otolito, denominadas para este caso A, B y C. Los incrementos de menor grosor o ancho mas pequeño se encontraron en una primera instancia cercanos al primordio (sección A), observándose nítidamente gracias a su forma concéntrica muy bien definida. En la sección B se nota un visible aumento en el grosor de los micro-incrementos definidos muy claramente. En esta zona se pudo constatar también la presencia de anillos sub-diarios y dobles, que presentaron una forma más irregular discontínua y poco clara. Llegando al borde del otolito (sección C) los grosores disminuyeron notablemente resultando un poco mas complicado en algunos casos distinguir los micro-incrementos (Fig. 19).
B C
C
Pr
A
A
B
Asd
Ad

43
Figura 19. Fotografia bajo luz trasmitida de la micro-estructura de un otolito juvenil de anchoveta: edad 119 dias, LT=11,5 cm, zona Mejillones, 2010; Pr; primordio, Ad; anillo doble y Asd; anillo sub-diario.
44
En la Tabla 10 se puede observar la media de los grosores de los micro-incrementos en las secciones A, B y C, y la media de los días en los que se extendieron estas secciones.
Tabla 10. Fluctuación, media y extensión de los grosores de los micro-incrementos por sección de otolitos juveniles de la zona Norte de Chile, asociados a eventos de El Niño y La Niña.
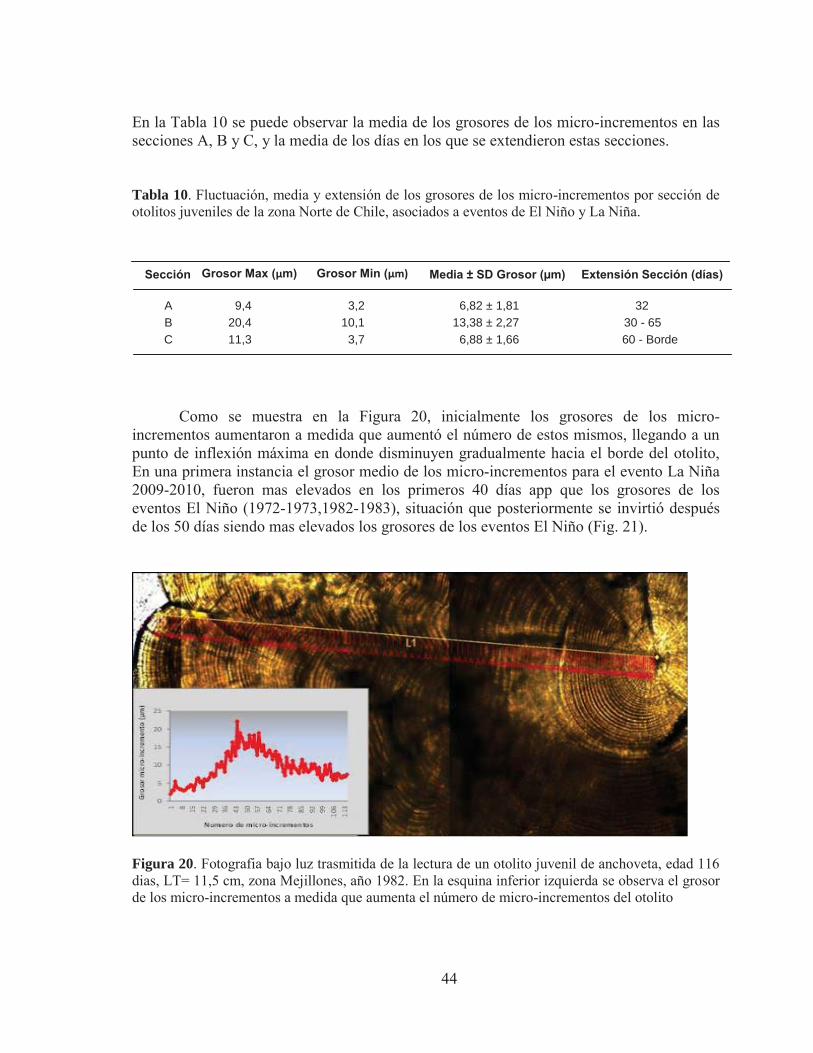
Como se muestra en la Figura 20, inicialmente los grosores de los micro-incrementos aumentaron a medida que aumentó el número de estos mismos, llegando a un punto de inflexión máxima en donde disminuyen gradualmente hacia el borde del otolito, En una primera instancia el grosor medio de los micro-incrementos para el evento La Niña 2009-2010, fueron mas elevados en los primeros 40 días app que los grosores de los eventos El Niño (1972-1973,1982-1983), situación que posteriormente se invirtió después de los 50 días siendo mas elevados los grosores de los eventos El Niño (Fig. 21).
Figura 20. Fotografia bajo luz trasmitida de la lectura de un otolito juvenil de anchoveta, edad 116 dias, LT= 11,5 cm, zona Mejillones, año 1982. En la esquina inferior izquierda se observa el grosor de los micro-incrementos a medida que aumenta el número de micro-incrementos del otolito
Sección Grosor Max (μm) Grosor Min (μm) Media ± SD Grosor (μm) Extensión Sección (días)
A 9,4 3,2 6,82 ± 1,81 32 B 20,4 10,1 13,38 ± 2,27 30 - 65 C 11,3 3,7 6,88 ± 1,66 60 - Borde
45
0
2
4
6
8
10
12
14
1 6
11
16
21
26
31
36
41
46
51
56
61
66
71
76
81
86
91
96
10
1
10
6
11
1
11
6
12
1
Gro
sor
mic
ro-i
ncr
em
en
tos
(μm
)
Numero de micro-incrementos
1973
1982
2009-2010
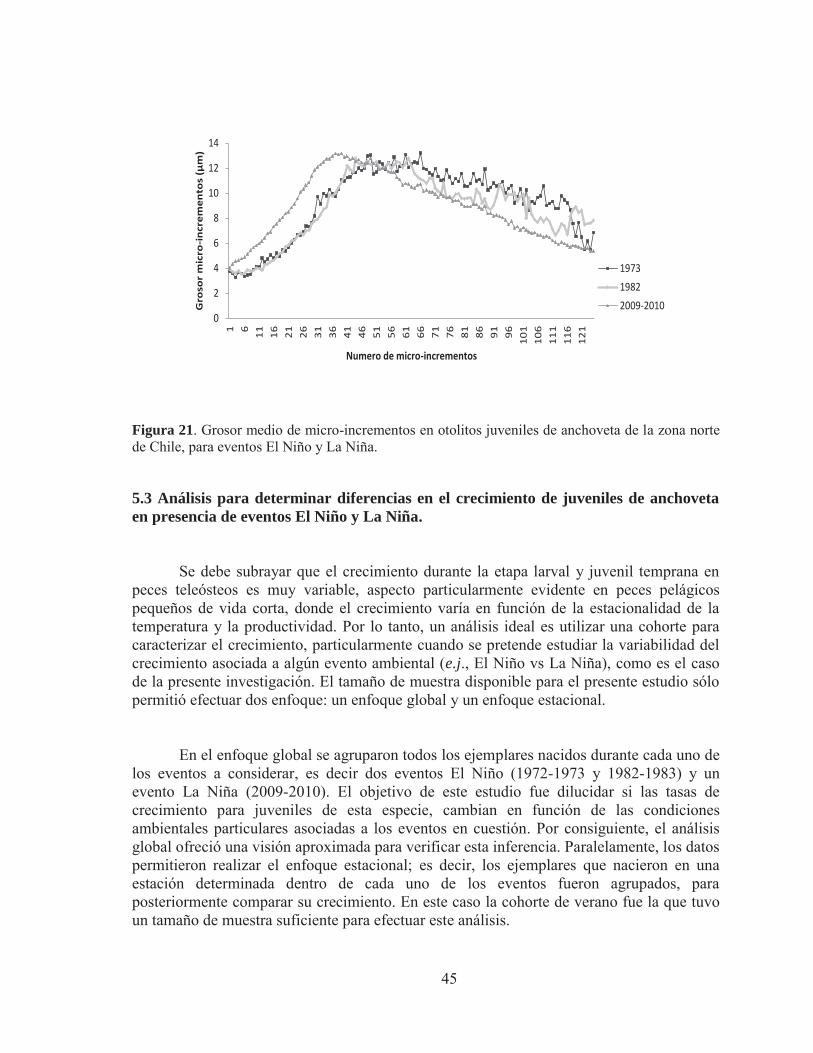
Figura 21. Grosor medio de micro-incrementos en otolitos juveniles de anchoveta de la zona norte de Chile, para eventos El Niño y La Niña. 5.3 Análisis para determinar diferencias en el crecimiento de juveniles de anchoveta en presencia de eventos El Niño y La Niña.
Se debe subrayar que el crecimiento durante la etapa larval y juvenil temprana en peces teleósteos es muy variable, aspecto particularmente evidente en peces pelágicos pequeños de vida corta, donde el crecimiento varía en función de la estacionalidad de la temperatura y la productividad. Por lo tanto, un análisis ideal es utilizar una cohorte para caracterizar el crecimiento, particularmente cuando se pretende estudiar la variabilidad del crecimiento asociada a algún evento ambiental (e.j., El Niño vs La Niña), como es el caso de la presente investigación. El tamaño de muestra disponible para el presente estudio sólo permitió efectuar dos enfoque: un enfoque global y un enfoque estacional.
En el enfoque global se agruparon todos los ejemplares nacidos durante cada uno de los eventos a considerar, es decir dos eventos El Niño (1972-1973 y 1982-1983) y un evento La Niña (2009-2010). El objetivo de este estudio fue dilucidar si las tasas de crecimiento para juveniles de esta especie, cambian en función de las condiciones ambientales particulares asociadas a los eventos en cuestión. Por consiguiente, el análisis global ofreció una visión aproximada para verificar esta inferencia. Paralelamente, los datos permitieron realizar el enfoque estacional; es decir, los ejemplares que nacieron en una estación determinada dentro de cada uno de los eventos fueron agrupados, para posteriormente comparar su crecimiento. En este caso la cohorte de verano fue la que tuvo un tamaño de muestra suficiente para efectuar este análisis.

47
Por otro parte, la evaluación y comparación del crecimiento se efectuó siguiendo los tres procedimientos paralelos, descritos en la metodología con el fin de fortalecer las conclusiones. (i) Relación Talla-Edad utilizando longitudes retro-calculadas (ii) Grosor de los micro-incrementos (iii) Tasas de crecimiento retro-calculadas 5.3.1 Relación talla-edad a) Comparación a nivel global: En este análisis se agrupó toda la información por cohortes anuales para obtener una proximidad del comportamiento de la relación talla-edad para el rango de edad y longitud de los individuos muestreados de cada evento (Tabla 11). Inicialmente se efectuó el ajuste desde el primer día de vida retro-calculada, para luego realizar el mismo procedimiento desde el día 40 en adelante. En la Tabla 12 se puede observar los parámetros del modelo para cada evento. Tabla 11. Rango de longitud y edad analizada en juveniles de anchoveta Engraulis ringens para el evento El Niño 1973 y 1982, y para el evento La Niña 2009-2010.
El proceso de retro-cálculo aplicado permitió reconstruir las longitudes previas y soslayar el problema debido al reducido rango de longitud analizada. Las longitudes retro-calculadas mostraron un patrón de crecimiento asintótico para cada año, que fue ajustado significativamente a un modelo curvilíneo convencional Laird Gompertz para cada evento de forma anual, como sigue:
La tasa de crecimiento instantánea (TICD) a cualquier edad es estimada como:
Evento Rango Lt (cm) Edad (días)
1972-1973 9 - 12 64 - 126 1982-1983 8,5 - 12 64 - 134 2009-2010 5,5 - 12 60 - 178
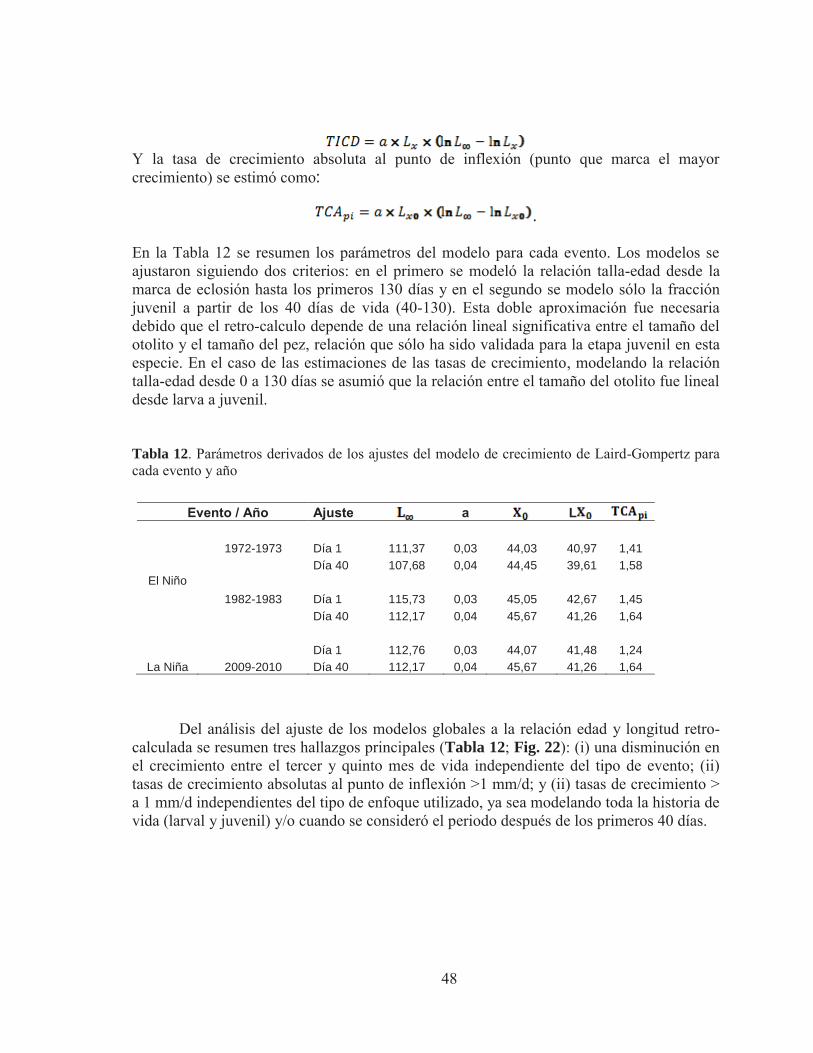
48
Y la tasa de crecimiento absoluta al punto de inflexión (punto que marca el mayor crecimiento) se estimó como:
. En la Tabla 12 se resumen los parámetros del modelo para cada evento. Los modelos se ajustaron siguiendo dos criterios: en el primero se modeló la relación talla-edad desde la marca de eclosión hasta los primeros 130 días y en el segundo se modelo sólo la fracción juvenil a partir de los 40 días de vida (40-130). Esta doble aproximación fue necesaria debido que el retro-calculo depende de una relación lineal significativa entre el tamaño del otolito y el tamaño del pez, relación que sólo ha sido validada para la etapa juvenil en esta especie. En el caso de las estimaciones de las tasas de crecimiento, modelando la relación talla-edad desde 0 a 130 días se asumió que la relación entre el tamaño del otolito fue lineal desde larva a juvenil. Tabla 12. Parámetros derivados de los ajustes del modelo de crecimiento de Laird-Gompertz para cada evento y año
Del análisis del ajuste de los modelos globales a la relación edad y longitud retro-
calculada se resumen tres hallazgos principales (Tabla 12; Fig. 22): (i) una disminución en el crecimiento entre el tercer y quinto mes de vida independiente del tipo de evento; (ii) tasas de crecimiento absolutas al punto de inflexión >1 mm/d; y (ii) tasas de crecimiento > a 1 mm/d independientes del tipo de enfoque utilizado, ya sea modelando toda la historia de vida (larval y juvenil) y/o cuando se consideró el periodo después de los primeros 40 días.
Evento / Año Ajuste a L
1972-1973 Día 1 111,37 0,03 44,03 40,97 1,41 Día 40 107,68 0,04 44,45 39,61 1,58
El Niño 1982-1983 Día 1 115,73 0,03 45,05 42,67 1,45 Día 40 112,17 0,04 45,67 41,26 1,64 Día 1 112,76 0,03 44,07 41,48 1,24
La Niña 2009-2010 Día 40 112,17 0,04 45,67 41,26 1,64
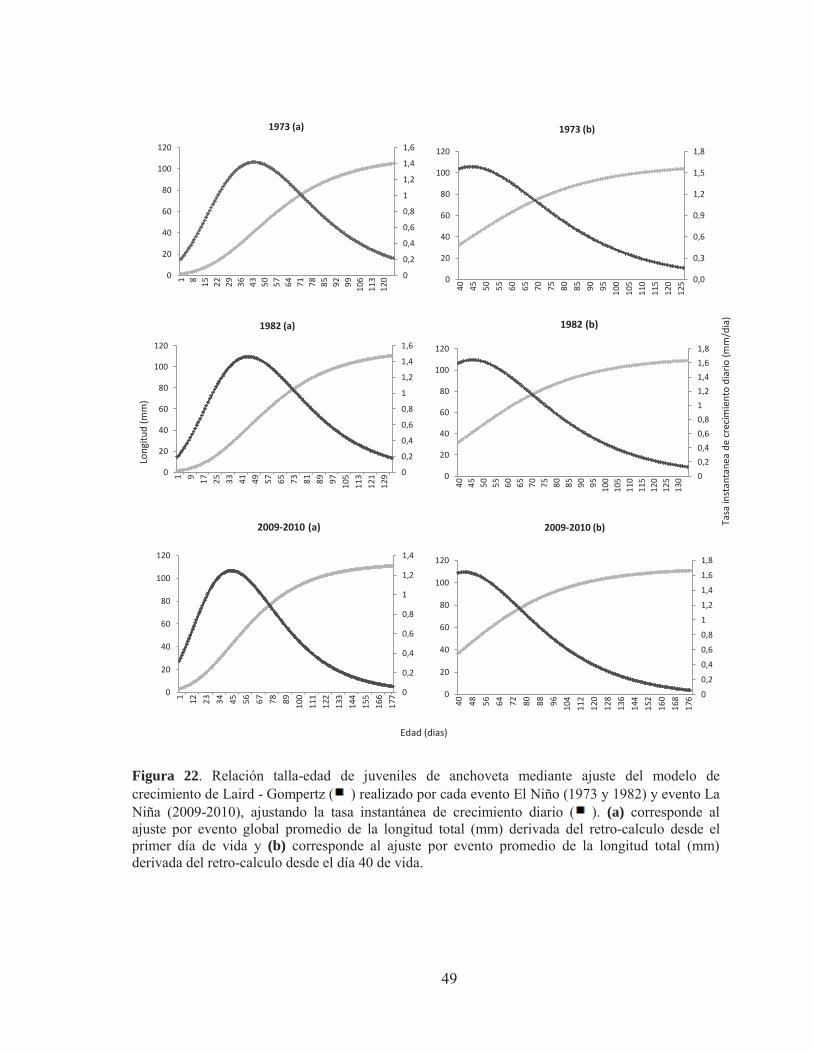
49
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
0
20
40
60
80
100
120
1 8 15 22 29 36 43 50 57 64 71 78 85 92 99 106
113
120
1973 (a)
0,0
0,3
0,6
0,9
1,2
1,5
1,8
0
20
40
60
80
100
120
40 45 50 55 60 65 70 75 80 85 90 95 100
105
110
115
120
125
1973 (b)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
0
20
40
60
80
100
120
1 9 17 25 33 41 49 57 65 73 81 89 97 105
113
121
129
1982 (a)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
0
20
40
60
80
100
120
40 45 50 55 60 65 70 75 80 85 90 95 100
105
110
115
120
125
130
1982 (b)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
0
20
40
60
80
100
120
1 12 23 34 45 56 67 78 89 100
111
122
133
144
155
166
177
2009-2010 (a)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
0
20
40
60
80
100
120
40 48 56 64 72 80 88 96 104
112
120
128
136
144
152
160
168
176
2009-2010 (b)
Edad (dias)
Long
itud
(mm
)
Tasa
inst
anta
nea
de c
reci
mie
nto
diar
io(m
m/d
ia)
Figura 22. Relación talla-edad de juveniles de anchoveta mediante ajuste del modelo de crecimiento de Laird - Gompertz ( ) realizado por cada evento El Niño (1973 y 1982) y evento La Niña (2009-2010), ajustando la tasa instantánea de crecimiento diario ( ). (a) corresponde al ajuste por evento global promedio de la longitud total (mm) derivada del retro-calculo desde el primer día de vida y (b) corresponde al ajuste por evento promedio de la longitud total (mm) derivada del retro-calculo desde el día 40 de vida.
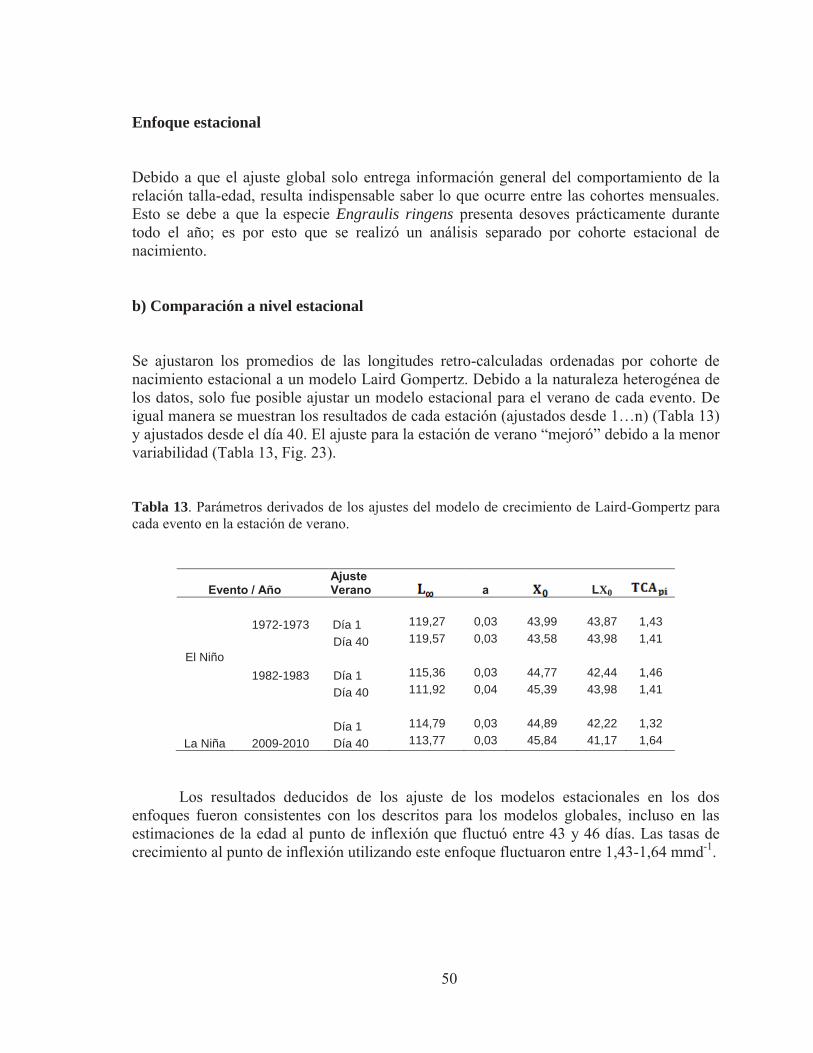
50
Enfoque estacional Debido a que el ajuste global solo entrega información general del comportamiento de la relación talla-edad, resulta indispensable saber lo que ocurre entre las cohortes mensuales. Esto se debe a que la especie Engraulis ringens presenta desoves prácticamente durante todo el año; es por esto que se realizó un análisis separado por cohorte estacional de nacimiento. b) Comparación a nivel estacional Se ajustaron los promedios de las longitudes retro-calculadas ordenadas por cohorte de nacimiento estacional a un modelo Laird Gompertz. Debido a la naturaleza heterogénea de los datos, solo fue posible ajustar un modelo estacional para el verano de cada evento. De igual manera se muestran los resultados de cada estación (ajustados desde 1…n) (Tabla 13) y ajustados desde el día 40. El ajuste para la estación de verano “mejoró” debido a la menor variabilidad (Tabla 13, Fig. 23). Tabla 13. Parámetros derivados de los ajustes del modelo de crecimiento de Laird-Gompertz para cada evento en la estación de verano.
Evento / Año Ajuste Verano a LX0
1972-1973 Día 1 119,27 0,03 43,99 43,87 1,43 Día 40 119,57 0,03 43,58 43,98 1,41
El Niño
1982-1983 Día 1 115,36 0,03 44,77 42,44 1,46
Día 40 111,92 0,04 45,39 43,98 1,41
Día 1 114,79 0,03 44,89 42,22 1,32
La Niña 2009-2010 Día 40 113,77 0,03 45,84 41,17 1,64
Los resultados deducidos de los ajuste de los modelos estacionales en los dos enfoques fueron consistentes con los descritos para los modelos globales, incluso en las estimaciones de la edad al punto de inflexión que fluctuó entre 43 y 46 días. Las tasas de crecimiento al punto de inflexión utilizando este enfoque fluctuaron entre 1,43-1,64 mmd-1.
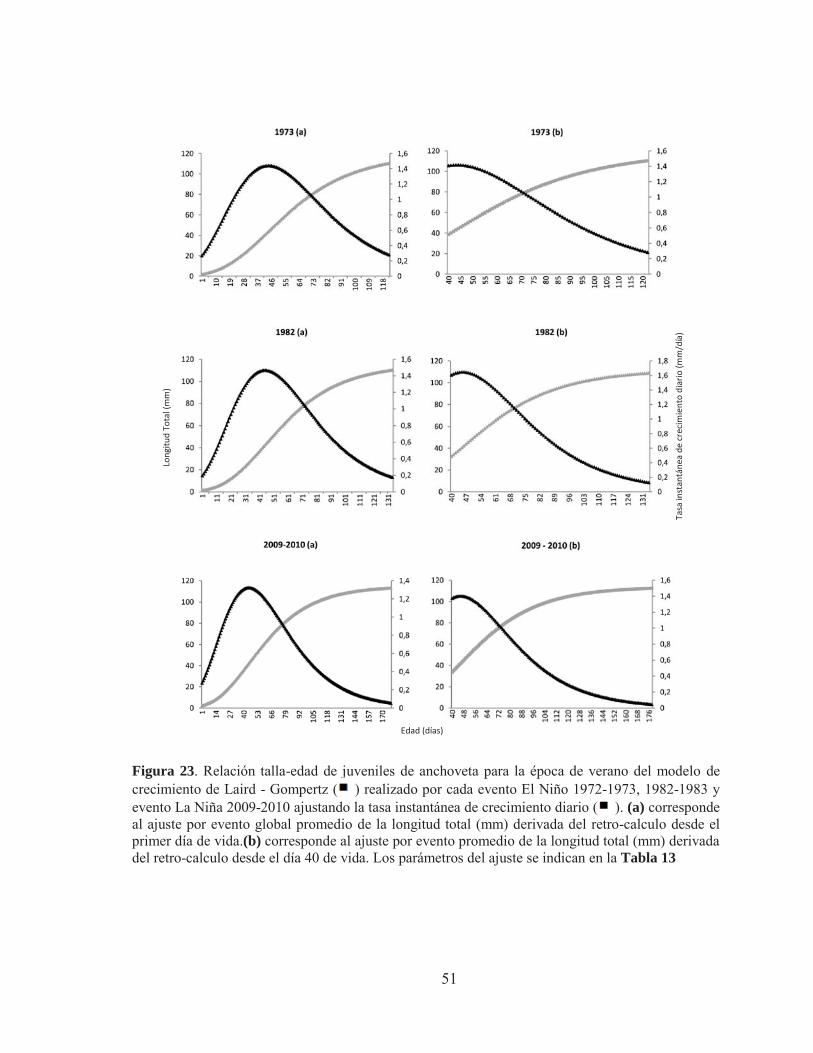
51
Edad (días)
Tasa
inst
antá
nea
de c
reci
mie
nto
diar
io (m
m/d
ía)
L
ongi
tud
Tota
l (m
m)
Figura 23. Relación talla-edad de juveniles de anchoveta para la época de verano del modelo de crecimiento de Laird - Gompertz (
j) realizado por cada evento El Niño 1972-1973, 1982-1983 y
evento La Niña 2009-2010 ajustando la tasa instantánea de crecimiento diario ( ). (a) corresponde al ajuste por evento global promedio de la longitud total (mm) derivada del retro-calculo desde el primer día de vida.(b) corresponde al ajuste por evento promedio de la longitud total (mm) derivada del retro-calculo desde el día 40 de vida. Los parámetros del ajuste se indican en la Tabla 13

52
c) Procedimiento de contraste de curvas de crecimiento
Se contrastaron los parámetros de la curva de Laird-Gompertz del análisis de la estación de verano ajustado desde 1 y 40 dias, realizando la rutina del programa estadístico SAS (procedimiento de contraste de curvas) y los resultados se muestran en la Tabla 14. Tabla 14. Procedimiento de contraste para la curva de crecimiento del modelo Laird–Gompertz derivada del análisis estacional de verano para anchoveta de la zona norte. Subíndices (73, año 1973; 82, año 1982; 10, año 2010).
Los resultados del procedimiento muestran que existe diferencia significativa (PROCNLMIXED, p < 0.05) para todos los parámetros de las curvas analizadas. 5.3.2 Tasas de crecimiento diarias retro-calculadas. Las tasas de crecimiento retro-calculadas analizadas global y estacionalmente evidenciaron un mismo patrón de desarrollo, independiente del tipo de evento. Sin embargo, se pudo observar una característica distintiva en el periodo de mayor crecimiento, donde las tasas de crecimiento retro-calculadas del evento La Niña tendieron a ser mayores que en el evento El Niño (1972-1973 y 1982-1983). Pese a esta diferencia que se produjo en primera instancia, se observó un efecto compensatorio en el crecimiento a partir del día 50 aproximadamente (Fig.24).
1,2 análisis estación verano desde el día 1 3,4 análisis estación verano desde el día 40
Contraste ¹ Valor F ² Valor P ³ Valor F 4 Valor P
L 73 vs L 82 24,27 0,0001 24,22 0,0001 L 82 vs L 10 39,16 0,0001 39,21 0,0001 L 73 vs L 10 3,88 0,0495 3,92 0,0486 a 73 vs a 82 11,93 0,0006 11,89 0,006 a 82 vs a 10 99,75 0,0001 99,83 0,0001 a 73 vs a 10 43,5 0,0001 43,63 0,0001 X0_ 73 vs X0_ 82 24,27 0,0001 24,22 0,0001 X0_ 82 vs X0_10 39,16 0,0001 39,21 0,0001 X0_ 73 vs X0_10 3,88 0,0495 3,92 0,0486
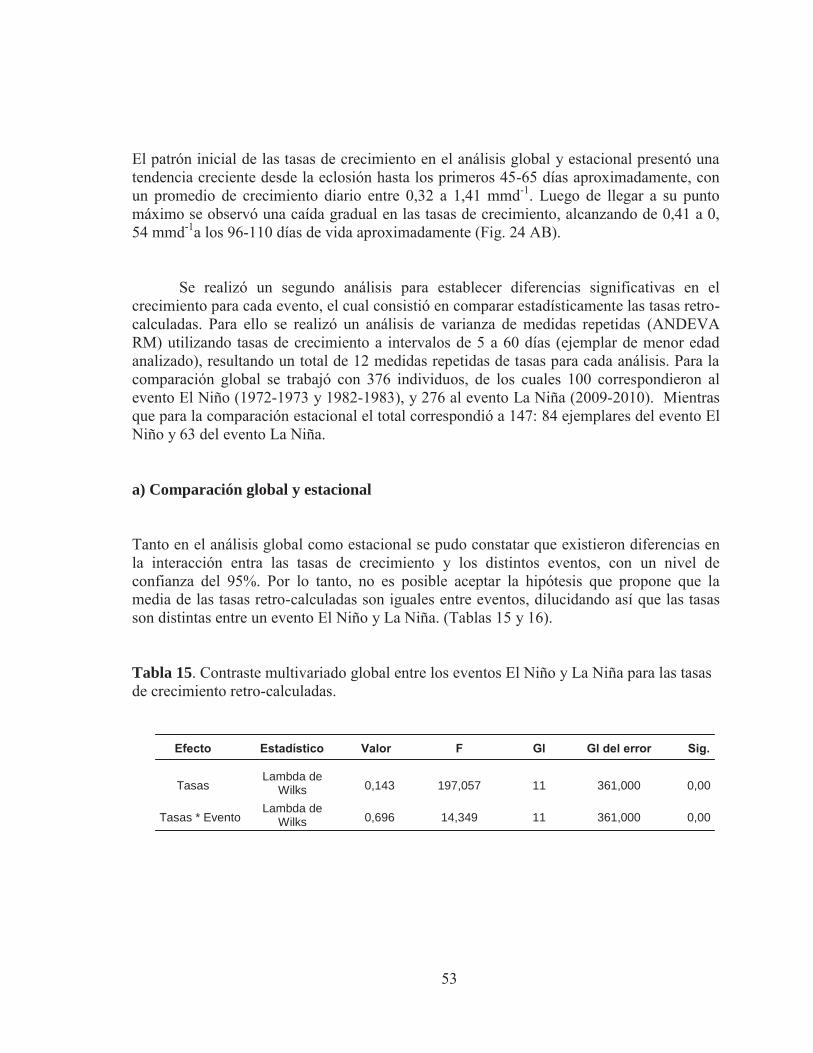
53
El patrón inicial de las tasas de crecimiento en el análisis global y estacional presentó una tendencia creciente desde la eclosión hasta los primeros 45-65 días aproximadamente, con un promedio de crecimiento diario entre 0,32 a 1,41 mmd-1. Luego de llegar a su punto máximo se observó una caída gradual en las tasas de crecimiento, alcanzando de 0,41 a 0, 54 mmd-1a los 96-110 días de vida aproximadamente (Fig. 24 AB).
Se realizó un segundo análisis para establecer diferencias significativas en el
crecimiento para cada evento, el cual consistió en comparar estadísticamente las tasas retro-calculadas. Para ello se realizó un análisis de varianza de medidas repetidas (ANDEVA RM) utilizando tasas de crecimiento a intervalos de 5 a 60 días (ejemplar de menor edad analizado), resultando un total de 12 medidas repetidas de tasas para cada análisis. Para la comparación global se trabajó con 376 individuos, de los cuales 100 correspondieron al evento El Niño (1972-1973 y 1982-1983), y 276 al evento La Niña (2009-2010). Mientras que para la comparación estacional el total correspondió a 147: 84 ejemplares del evento El Niño y 63 del evento La Niña. a) Comparación global y estacional Tanto en el análisis global como estacional se pudo constatar que existieron diferencias en la interacción entra las tasas de crecimiento y los distintos eventos, con un nivel de confianza del 95%. Por lo tanto, no es posible aceptar la hipótesis que propone que la media de las tasas retro-calculadas son iguales entre eventos, dilucidando así que las tasas son distintas entre un evento El Niño y La Niña. (Tablas 15 y 16). Tabla 15. Contraste multivariado global entre los eventos El Niño y La Niña para las tasas de crecimiento retro-calculadas.
Efecto Estadístico Valor F Gl Gl del error Sig.
Tasas Lambda de
Wilks 0,143 197,057 11 361,000 0,00
Tasas * Evento Lambda de
Wilks 0,696 14,349 11 361,000 0,00

54
Tabla 16. Contraste multivariado estacional entre los eventos El Niño y La Niña para las tasas de crecimiento retro-calculadas.
Figura 24.A: Análisis global de la tasa de crecimiento retro-calculada media anual de los micro-incrementos por evento y B: análisis estacional de la tasa de crecimiento retro-calculada media anual para la época de verano por evento. La línea vertical muestra el punto de corte del análisis de medidas repetidas.
Efecto Estadístico Valor F Gl Gl del error Sig.
Tasas
Lambda de Wilks
0,097 114,187a 11 135,000 0,00
Tasas * Evento Lambda de
Wilks 0,619 7,554a 11 135,000 0,00
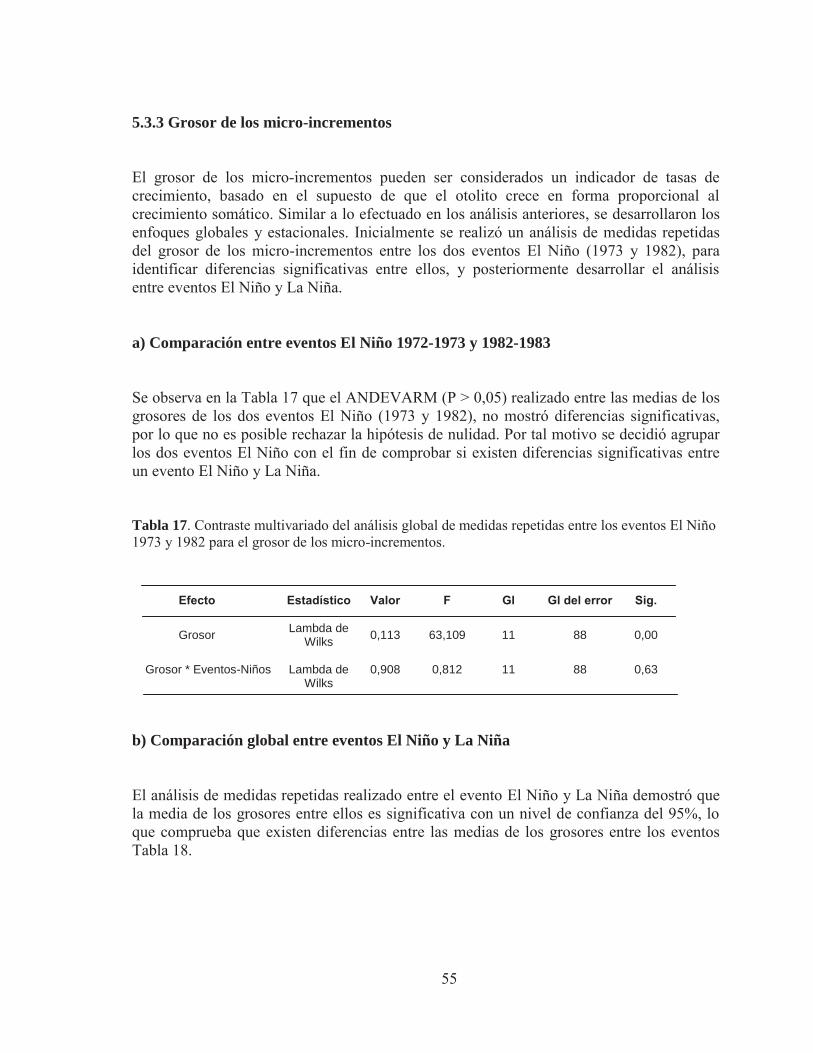
55
5.3.3 Grosor de los micro-incrementos El grosor de los micro-incrementos pueden ser considerados un indicador de tasas de crecimiento, basado en el supuesto de que el otolito crece en forma proporcional al crecimiento somático. Similar a lo efectuado en los análisis anteriores, se desarrollaron los enfoques globales y estacionales. Inicialmente se realizó un análisis de medidas repetidas del grosor de los micro-incrementos entre los dos eventos El Niño (1973 y 1982), para identificar diferencias significativas entre ellos, y posteriormente desarrollar el análisis entre eventos El Niño y La Niña. a) Comparación entre eventos El Niño 1972-1973 y 1982-1983 Se observa en la Tabla 17 que el ANDEVARM (P > 0,05) realizado entre las medias de los grosores de los dos eventos El Niño (1973 y 1982), no mostró diferencias significativas, por lo que no es posible rechazar la hipótesis de nulidad. Por tal motivo se decidió agrupar los dos eventos El Niño con el fin de comprobar si existen diferencias significativas entre un evento El Niño y La Niña. Tabla 17. Contraste multivariado del análisis global de medidas repetidas entre los eventos El Niño 1973 y 1982 para el grosor de los micro-incrementos.
b) Comparación global entre eventos El Niño y La Niña El análisis de medidas repetidas realizado entre el evento El Niño y La Niña demostró que la media de los grosores entre ellos es significativa con un nivel de confianza del 95%, lo que comprueba que existen diferencias entre las medias de los grosores entre los eventos Tabla 18.
Efecto Estadístico Valor F Gl Gl del error Sig.
Grosor
Lambda de
Wilks
0,113
63,109
11
88
0,00
Grosor * Eventos-Niños
Lambda de Wilks
0,908 0,812 11 88 0,63
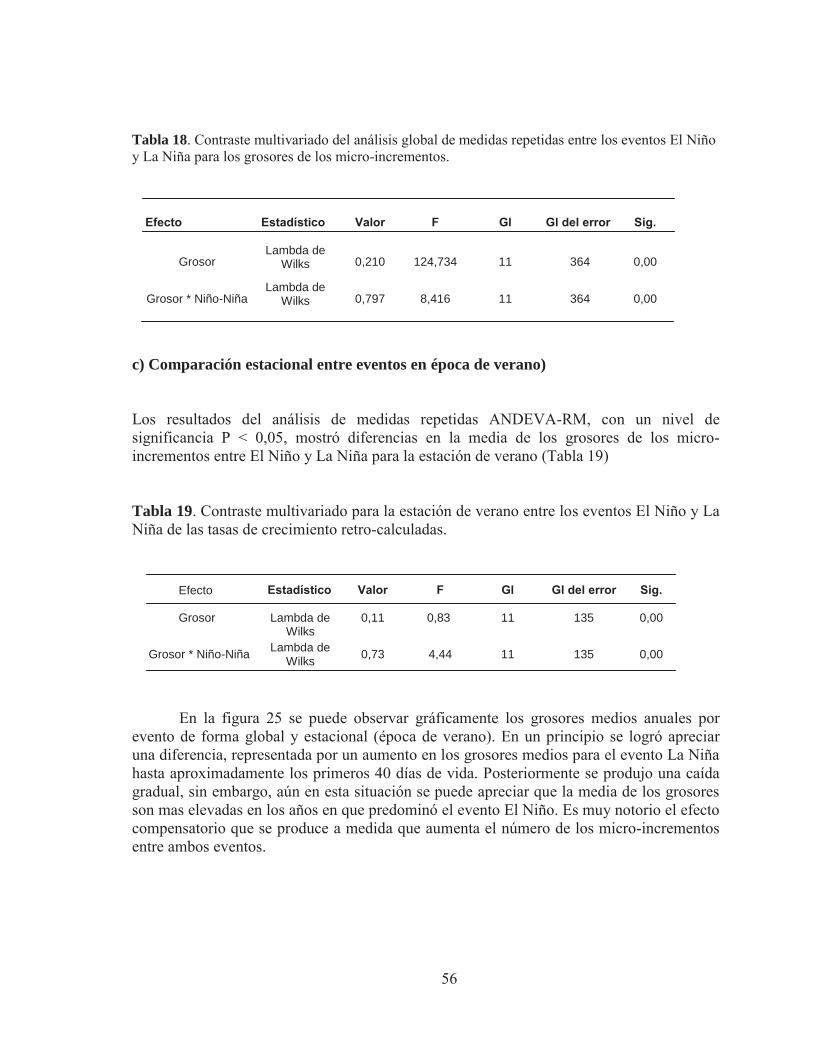
56
Tabla 18. Contraste multivariado del análisis global de medidas repetidas entre los eventos El Niño y La Niña para los grosores de los micro-incrementos.
c) Comparación estacional entre eventos en época de verano) Los resultados del análisis de medidas repetidas ANDEVA-RM, con un nivel de significancia P < 0,05, mostró diferencias en la media de los grosores de los micro-incrementos entre El Niño y La Niña para la estación de verano (Tabla 19) Tabla 19. Contraste multivariado para la estación de verano entre los eventos El Niño y La Niña de las tasas de crecimiento retro-calculadas.
En la figura 25 se puede observar gráficamente los grosores medios anuales por
evento de forma global y estacional (época de verano). En un principio se logró apreciar una diferencia, representada por un aumento en los grosores medios para el evento La Niña hasta aproximadamente los primeros 40 días de vida. Posteriormente se produjo una caída gradual, sin embargo, aún en esta situación se puede apreciar que la media de los grosores son mas elevadas en los años en que predominó el evento El Niño. Es muy notorio el efecto compensatorio que se produce a medida que aumenta el número de los micro-incrementos entre ambos eventos.
Efecto Estadístico Valor F Gl Gl del error Sig.
Grosor
Lambda de Wilks
0,11 0,83 11 135 0,00
Grosor * Niño-Niña Lambda de Wilks 0,73 4,44 11 135 0,00
Efecto Estadístico Valor F Gl Gl del error Sig.
Grosor Lambda de
Wilks 0,210 124,734 11 364 0,00
Grosor * Niño-Niña Lambda de
Wilks 0,797 8,416 11 364 0,00
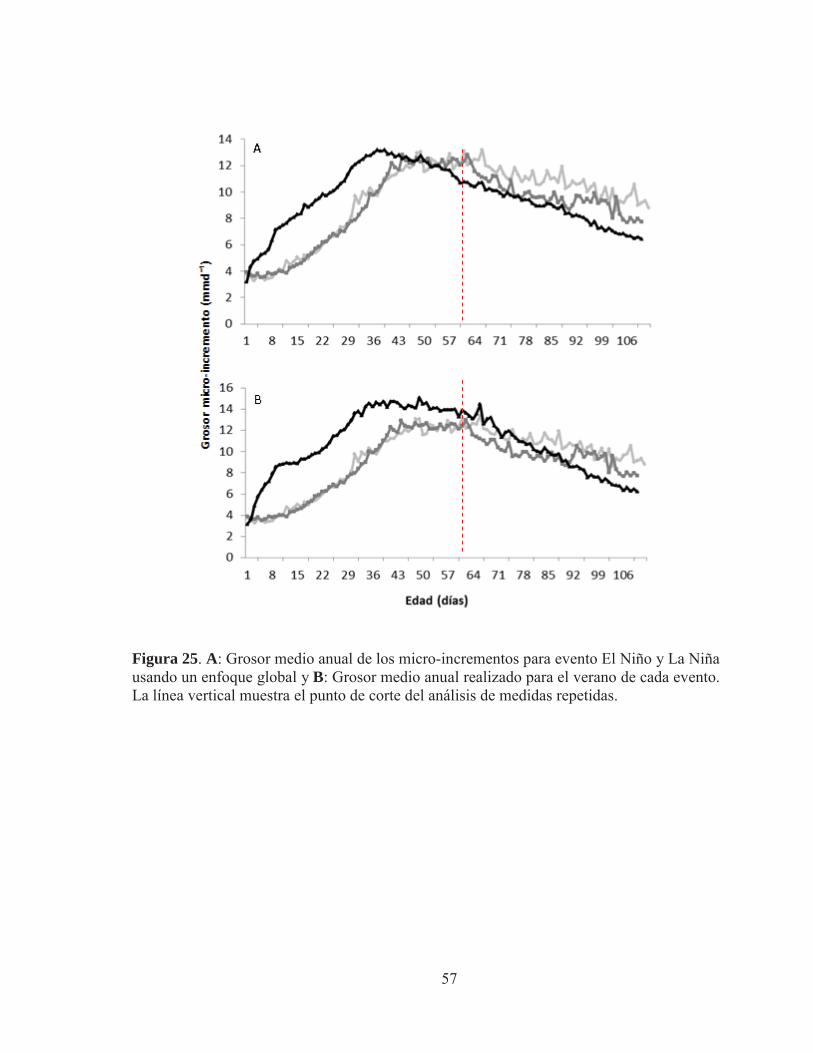
57
Figura 25. A: Grosor medio anual de los micro-incrementos para evento El Niño y La Niña usando un enfoque global y B: Grosor medio anual realizado para el verano de cada evento. La línea vertical muestra el punto de corte del análisis de medidas repetidas.

58
6. DISCUSIÓN
6.1 Micro-estructura de otolitos juveniles de anchoveta
En la microestructura de otolitos (sagitta) de juveniles de anchoveta de la zona norte de Chile se logró distinguir tres secciones, denominadas en este estudio A, B y C. De forma general una de las principales características de estas tres secciones fue la nitidez con la que se observaron los micro-incrementos, revelando visiblemente la diferencia entre la zona hialina y la zona opaca. Esto se puede deber a la exposición de los individuos a las fluctuaciones térmicas diarias naturales. Estudios realizados anteriormente (Neilson et al., 1985; Ojanguren et al., (1999) han demostrado que las fluctuaciones de temperatura provocan una mayor resolución de los micro-incrementos. Las sección A se extendió desde el centro del crecimiento hasta aproximadamente los primeros 30 días. En esta área fue posible identificar claramente el primordio, la marca de primera alimentación y la marca de eclosión, la que coincidió con la formación del primer incremento (Lagardere, 1989).
La zona intermedia B se extendió entre los 30 y 60 días, donde la identificación de
los micro-incrementos diarios fue un poco más compleja debido a la existencia de anillos dobles y/o perturbaciones. Se ha reportado la existencia de anillos dobles y o triples en ciertas regiones del otolito (Zhan & R. hunham, 1992; Palomera (1992), llegando a conclusiones similares a las de Cermeño et al. (2006; 2008) en Engraulis encrasilocus. La identificación de estos anillos dobles como diarios intrínsicamente implica que durante este periodo existiría una reducción después de un periodo de crecimiento larval acelerado. Algunos autores han señalado que esta alternancia de bandas implicaría cambios rápidos en la tasa de crecimiento, difíciles de explicar desde un punto de vista fisiológico para el periodo de transición de larva a juvenil. Estos argumentos han sido esgrimidos para considerar estas estructuras bipartitas como sub-diarias (Tomas & Panfili 2000; Hwang & Lee, 2005; Cermeño et al., 2006; Cermeño et al., 2008). En contraposición a estos argumentos, varios estudios han reportado que el crecimiento en clupeiformes se torna alométrico hacia el final de la etapa larval (Blaxter & Hunter, 1982; Watanabe & Kuroki, 1997; Takahashi et al., 2001; Plaza et al., 2008). Esta inflexión del crecimiento presumiblemente esta vinculado a que el crecimiento en longitud puede ser proporcionalmente menor que en profundidad corporal y/o en peso durante el periodo de transición. Adicionalmente, nuestros resultados concuerdan con el trabajo reciente de Moreno et al. (2010), quienes caracterizaron el periodo de transición de larva a juvenil para esta misma especie en condiciones de confinamiento. Estos autores demostraron que existen dos periodos de inflexión en el crecimiento para Engraulis ringens, uno que ocurre al inicio del periodo de transición entre 32 y 64 días (media 46 dias) y otro al finalizar el mismo entre los 81 y 149 días. Por consiguiente, es razonable aseverar que los anillos dobles, contados como unidades independientes, reflejan los cambios en la ontogenia de esta especie. Sin embargo, resulta necesario incorporar en futuros estudios un análisis micro-estructural de otros pares de otolitos, como lapilus y/o asteriscus, para dilucidar si la

59
existencia de estas estructuras bipartitas son sólo características propias de los otolitos sagitales. En la tercera sección la característica más relevante fue la disminución del grosor de los micro-incrementos hacia al borde del otolito, resultando un poco mas complicado en algunos casos distinguirlos. Esto debido a que la zona posterior del otolito fue en la mayoría de los casos hialina, lo que pudo haber sido provocado por el desgaste de forma desigual al momento de pulirlo, lo que resulta inevitable debido a la irregularidad y diseño cóncavo del otolito, provocando el sobre pulido de esta área.
El grosor de los incrementos no mostró diferencias significativas en las muestras analizadas entre los eventos El Niño 1972-1973 y 1982-1983, situación que puede deberse a que los ejemplares en ambos periodos estuvieron sometidos a similares condiciones océano-atmosféricas. Sin embargo, al comparar los ejemplares entre un evento El Niño y La Niña se pudo constatar diferencias significativas en los grosores de los otolitos pertenecientes al evento La Niña en donde se distinguió un notorio aumento de estos los primeros 40 días de vida, Esta diferencia se sustenta en el hecho de que las principales variables que controlan el crecimiento en los estadios tempranos de peces son la disponibilidad de alimento (Jenkins, 1987) y la temperatura del mar (Miller et al.,1988; Houde & Zastrow, 1993). Este último factor presenta anomalías negativas en presencia del evento La Niña, existiendo una mayor disponibilidad de alimento a diferencia del evento El Niño donde el calentamiento de las aguas provoca un empobrecimiento de estas causando una falla en la cadena trófica. El factor temperatura explica más del 50% de la variabilidad en el crecimiento en los estadios tempranos de peces teleósteos (Shepherd & Cushing, 1980). Esta variable junto con la disponibilidad de alimento se ven afectadas en distinta forma por la presencia de los eventos océano-atmosféricos El Niño y La Niña.
Estudios anteriores han reportado que el crecimiento en clupeiformes se torna alométrico hacia el final de la etapa larval (Blaxter & Hunter, 1982; Watanabe & Kuroki, 1997; Gaughan et al., 2001; Takahashi et al., 2001; Plaza et al., 2008) situación que coincide con los resultados entregados al analizar de forma global y estacional el grosor de los micro-incrementos considerados como un proxy del crecimiento somático. La detención en el crecimiento al finalizar este periodo ha sido atribuida a los cambios producidos por el proceso de transición de larva a juvenil, aspecto que ha sido también identificado en Engraulis ringens en condiciones de confinamiento (Moreno et al., 2005).
Sin embargo, resulta indispensable investigar acuciosamente el comportamiento de
estos en periodos de desarrollo larval y juvenil, ya sea de forma mensual, semanal e incluso diaria, lo que permitiría una visión mucho más completa y detallada de la conducta del crecimiento en los estadios de vida temprana.

60
Uno de los requisitos primordiales para el uso de las técnicas de retro-cálculo es que debe existir una relación lineal entre el tamaño del pez y el tamaño del otolito. La especie Engraulis ringens posee una relación proporcional entre su tamaño corporal y el tamaño del otolito (Hernández & Castro, 2000). Es decir, que se pueda predecir el tamaño del pez mediante el tamaño del otolito esta condición resulta indispensable para así poder determinar la longitud al tiempo de formación de cada una de las marcas presentes en los otolitos y así estimar el crecimiento en etapas anteriores a su captura 6.2 Crecimiento en la etapa juvenil asociado a los eventos El Niño y La Niña Los hallazgos reportados en el presente estudio demuestran que esta especie tiene un elevado crecimiento en su fase larval y juvenil temprana. Las tasas de crecimiento absolutas reportadas en el presente estudio, derivadas de la relación talla-edad retro-calculada, mediante dos enfoques (global y estacional) para cada evento y ajustadas desde edad 1 y 40 días fueron las siguientes: las tasas globales en general para ambos ajustes fluctuaron entre 1.23 y 1.64 mmd-1, alcanzada a una edad promedio de 45 días. Las tasas de crecimiento absolutas estacionales, realizadas para la cohorte de verano de cada evento fluctuaron entre 1.32 y 1.64 mmd-1 , alcanzada a una edad promedio de 44 días. Lo más relevante que se debe destacar de estos resultados, es que independiente del evento en el cual se encuentre la cohorte, su paso por anomalías de temperatura ya sean negativas o positivas, no afectan significativamente (p < 0.05) su crecimiento absoluto, de la cohorte en la estación de verano. El perfil de las tasas de crecimiento diarias retro-calculadas mostró un efecto compensatorio entre eventos, lo que indica que la anchoveta posee gran plasticidad en el crecimiento durante sus fases de vida temprana utilizando su máximo potencial de crecimiento en las primeras etapas de su vida para alcanzar su tamaño optimo lo antes posible, esta estrategia suele considerarse una ventaja al alcanzar tamaños mayores o mayor rapidez en el crecimiento en la etapa larval sobre las larvas menores o de crecimiento mas lento (Meekan & Fortier, 1996; Butler et al., 1997). Según Bailey & Houde (1989) en la supervivencia intervendría una mortalidad selectiva por tamaño, la cual llega a ser uno de los factores de mayor importancia en la vida de las larvas de los peces.
Las tasas de crecimiento reportadas en otros estudios para la familia Engraulidae resultan aparentemente menores a las obtenidas en este estudio, las que fluctuaron entre 1,24 y 1,64 (mm dia-1) y contrastan con las obtenidas por Butler (1989); Methor, Tsuji (1983); Sigel (1988) y Cermeño (2008). En la siguiente Tabla 1 se pueden observar las tasas de crecimiento conseguidas por las investigaciones realizadas. Sin embargo, las tasas de crecimiento absolutas obtenidas a través de los procedimientos realizados, son similares a las alcanzadas para la anchoa europea Engraulis encrasicolus en el golfo de Bizkaia durante los estadios tempranos de su ciclo de vida. En este estudio se realizó un ajuste talla-edad con datos observados y ajustados a un modelo logístico. Al final del primer año de crecimiento, la talla estimada por la anchoa fue de 168,3 mm, con una tasa máxima de crecimiento de 1,85 mmd-1 a los 73 días después de la eclosión (Aldanondo et al., 2011).
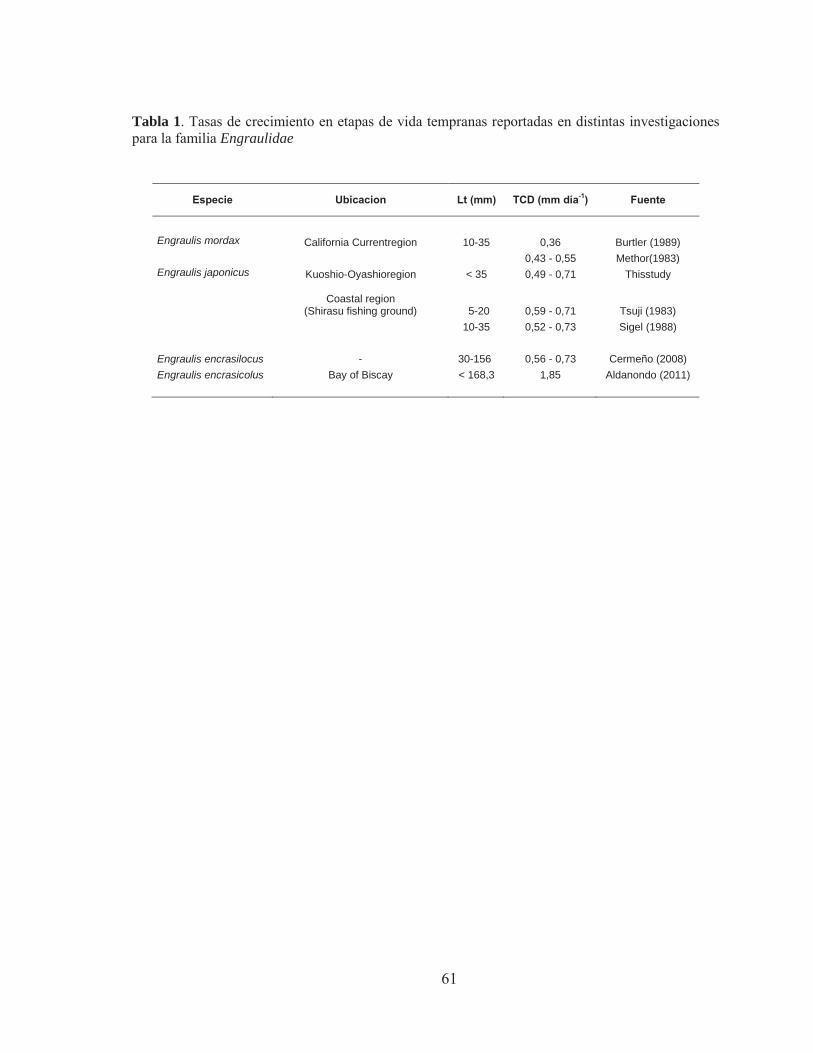
61
Tabla 1. Tasas de crecimiento en etapas de vida tempranas reportadas en distintas investigaciones para la familia Engraulidae
Especie Ubicacion Lt (mm) TCD (mm día-1) Fuente
Engraulis mordax California Currentregion 10-35 0,36 Burtler (1989) 0,43 - 0,55 Methor(1983) Engraulis japonicus Kuoshio-Oyashioregion < 35 0,49 - 0,71 Thisstudy
Coastal region
(Shirasu fishing ground) 5-20 0,59 - 0,71 Tsuji (1983) 10-35 0,52 - 0,73 Sigel (1988) Engraulis encrasilocus - 30-156 0,56 - 0,73 Cermeño (2008) Engraulis encrasicolus Bay of Biscay < 168,3 1,85 Aldanondo (2011)

62
7. CONCLUSIONES
La resolución de los micro-incrementos fue elevada, particularmente en individuos menores a 10 cm, donde un doble pulido en posición sagital fue suficiente para relevar los micro-incrementos diarios desde el primordio hasta el borde del otolito.
El análisis de la micro-estructura de los otolitos sagitta no evidenciaron diferencias entre los ejemplares analizados de los distintos años 1973, 1982 (eventos El Niño) y 2009-2010 (evento La Niña), encontrándose un similar patrón de formación y crecimiento en todos los ejemplares estudiados. Se lograron identificar tres secciones cuya principal características fue la nitidez con la que se observaron los micro-incrementos, revelando visiblemente el primordio, la marca de eclosión y de primera alimentación, y la diferencia entre la zona hialina y la zona opaca. Se identificó una clara diferencia en el grosor de los micro-incrementos entre eventos, resultando significativamente mayores los grosores de los ejemplares pertenecientes al evento La Niña durante el primer mes de vida. Sin embargo, posteriormente se produjo un efecto compensatorio en la etapa juvenil temprana (40-110 días), con grosores mayores durante los eventos El Niño.
La relación talla-edad a nivel global y por cohorte estacional de nacimiento, independiente del evento (El Niño y La Niña) se ajustó significativamente a un modelo Laird-Gompertz, demostrando que para el rango de juveniles analizados (7-12 cm) el crecimiento se torno asintótico hacia el quinto mes de vida aproximadamente a los 11 cm de longitud total, presumiblemente vinculado al proceso inicial de primera madurez sexual.
Se demostró que esta especie alcanzó tasas de crecimiento absolutas al punto de inflexión durante la fase juvenil temprana de 1,24 a 1,64 mm/día, alcanzada a una edad promedio de 45 días. Estas elevadas tasas de crecimiento se obtuvieron tanto en años en que predomino el evento El Niño como La Niña, lo que sugiere que esta especie tiene una gran plasticidad en el crecimiento durante sus estadios tempranos. Con respecto a la especie Engraulis ringens se puede concluir que es un recurso con una tasa de renovación elevada, que maximiza su potencial de crecimiento en los primeros meses de vida. Esta característica puede ser un mecanismo adaptativo para asegurar su supervivencia en un ecosistema de surgencia altamente productivo.
8. REFERENCIAS

63
Aguayo, M. 1976. Edad y Crecimiento de la anchoveta (Engraulis ringens), del norte de
Chile (Arica-Iquique). Ser. Invest. Pesq. Inst Fom. Pesq., Chile 23: 51p. Aldanondo, N., U. Cotano & E. Etxebeste. 2011. Growth of young-of-the-year European
anchovy (Engraulis encrasicolus), in the Bay of Biscay. Sci. Mar., 75: 227-235. Alheit, J. & M. Niquen. 2004. Regime shifts in the Humboldt Current ecosystem. Progress
in Oceanography., 60: 201–222. Aliaga, B., D. Gómez & S. Neira. 2001. Análisis bio-económico de la pesquería de sardina
(Sardinops sagax) y anchoveta (Engraulis ringens) de la zona norte de Chile. Invest. Mar., Valparaíso, 29(2): 15-23.
Araya, M. & L. Cubillos. 2002. The retrospective analysis of the growth in fish and their associated problems. Gayana, 66: 161-179.
Arcos, D., L. Cubillos & S. Núñez. 2004. Efectos de El Niño 1997-1998 Sobre las Principales Pesquerías pelágicas de la zona centro-sur de Chile. En: El Niño-La Niña 1997-2000, Sus Efectos en Chile. S. Avaria, J. Carrasco, J. Rutllant y E. Yáñez (eds.). Comité Oceanográfico Nacional (CONA), Valparaíso, Chile.
Avaria, S., J. Carrasco, J. Rutllant & E. Yáñez. 2004. El Niño-La Niña 1997-2000, Sus Efectos en Chile.
Bailey, K.M. & E.D. Houde. 1989. Predation on eggs and larvae and the recruitment problem. Adv. Mar. Biol., 79: 303–309.
Blaxter, J. H. S. & J. R. Hunter. 1982. The biology of the clupeoid fishes. In: J. H. S. Blaxter, F. S. Rusell & M. Yonge. (eds.). Advances in Marine Biology New York: Academic Press, 1–223 pp.
Brothers, E. B., C. P. Mathews & R. Lasker. 1976. Daily growth increments in otoliths from larval and adult fishes. Fish. Bull., 74: 1–8.
Brothers, E. B. 1978. Exogenous factors and the formation of daily and sub·daily growth increments in fish otoliths. Abstr. Am. Zool. Hl: 631.
Butler, J. L. 1989. Growth during the larval and juvenile stages of the Northern Anchovy, Engraulis mordax, in the California Current during 1980–84. Fish. Bull., 87: 645–652.
Campana, S. E. & J.D. Neilson, 1982. Daily growth increments in otoliths of starry flounder (Platichthys stellatus) and the influence of some environmental variables in their production. Can. J. Fish. Aquat. Sci., 39: 937–942.
Campana, S. E. 1983. La alimentación de la periodicidad y la producción de incremento de crecimiento diario de los otolitos de trucha arco iris Salmo gaidneri y los platija Platichthys tellatus. Can J. Zool., 61(1): 591-597.
Campana, S. E. & J.D. Neilson. 1985. Microstructure of fish otoliths. Can J. Fish. Aquat. Sci., 42: 1014-1032.
Campana, S.E. 1990. How reliable are growth back-calculation based on otoliths? Can. J. Fish. Aquat. Sci., 47: 2219-2227.
Campana, S. E. & C. M. Jones. 1992. Analysis of otolith microstructure data. In: D. K. Stevenson & S. E. Campana (eds.). Otolith microstructure examination and analysis. Can. Spec. Publ. Fish. Aquat. Sci.,117 pp

64
Cañón, J.R. 1986.Variabilidad ambiental en relación con la pesquería pelágica de la zona norte de Chile. In: P. Arana (ed.). La Pesca en Chile. Escuela de Ciencias del Mar, UCV, Valparaíso.
Carlander, K.D. 1981. Caution on the use of the regression method of back-calculating lengths from scale measurements. Fish. Bull., 6(1): 2-4
Carreño, D. 2010. Tesis para optar al título de Ingeniería en Acuicultura. “Reconstruir los
patrones de crecimiento, durante la etapa de incubación de salmón del Atlántico (Salmo salar), mediante el análisis de la microestructura de otolitos” pp 33
Casselman, J.M. 1990. Growth and relative size of calcified structures of fish. Trans. Amer. Fish. Soc., 119: 673-688.
Castellano, J.P. & M.C. Vasconcellos. 1995. Growth rate of anchovy (Engraulis anchoíta) larvae caught off Cape Santa Marta Grande (Brazil). Arch. Fish. Mar. Res., 42: 263- 281.
Castro, L. R., G. R. Salinas & E. H. Hernández. 2000. Environmental influences on winter spawning of the anchoveta Engraulis ringens off central Chile. Mar. Ecol. Progr. Ser., 197: 247-258.
Cazeneuve, H., R. Ozu & C. Rinaldi. 2000. El Niño y La Niña. Rev. Info. Cient. Amb. Ecol., 75: 51-52.
Cermeño, P., B. Morales-Nin & A. Uriarte. 2006. Juvenile European anchovy otolith microstructure. Sci. Mar., 70: 553-557.
Cermeño, P., A. Uriarte, B. Morales-Nin, U. Cotano & P. Alvarez. 2008. Setting up interpretation criteria for ageing juvenile European anchovy otoliths. Sci. Mar.,72: 733-742.
Cerna F., G. Plaza, A. López, C. Machuca, R. Serra, C. Gaspar & G. Moyano. 2011. Revisión de la asignación por grupo de edad en la anchoveta de la zona norte. Informe Pre Final: FIP 2009-17. 52 pp.
Chambers, R.C. & T.J. Miller. 1995. Evaluating fish growth by means of otolith increment analysis: special properties of individual–level longitudinal data. In: J.D. Secor and S.E. Campana (eds.). Recent developments in fish otolith research pp. 155-175. University of South Carolina Press.
Chong, J., M. Aguayo & I. Payá. 2007. Determination of age, growth and natural mortality of Chilean hoki, Macruronus magellanicus, Lönnberg, 1907 (Macruronidae, Gadiformes) from the Southeastern Pacific Ocean. Rev. Biol. Mar., 42(3): 311-333.
Chugunova, N.I. 1963. Age and growth studies in fish. National science foundation Washington D.C., Israel Program for Scientific Translations. Jerusalem. 132 pp
Cubillos, L. & H. Arancibia. 1993b. Análisis de la pesquería de sardina común (Strangomera bentincki) y anchoveta (Engraulis ringens) del área de Talcahuano, Chile. Invest. Mar. Valparaíso, 21: 3-21.
Cubillos, L., M. Canales, A. Hernández. D. Bucarey, L. Vilugrón & L. Miranda. 1998. Poder de pesca, esfuerzo de pesca y cambios estacionales e interanuales en la abundancia relativa de Strangomera bentincki y Engraulis ringens en el área frente a Talcahuano, Chile (1990-97). Invest. Mar. Valparaíso, 26: 3-14.

65
Cubillos, L., M. Canales, D. Bucarey, A. Rojas & R. Alarcón. 1999. Época reproductiva y talla media de primera madurez sexual de Strangomera bentincki y Engraulis ringens en la zona centro-sur de Chile en el período 1993-1997. Invest. Mar. Valparaíso, 27: 73-86.
Cubillos, L., D. Bucarey & M. Canales. 2002. Monthly abundance estimation for common sardine Strangomera bentincki and anchovy Engraulis ringens in the central-southern area off Chile (34–40°S). Fish. Res., 57: 117-130.
Cubillos, L. & D. Arcos. 2002. Is the jack mackerel population affected by ENSO’s variability? In: Extended abstracts of the El Niño Symposium & Workshop. Investigaciones Marinas, Valparaíso, 30: 185- 193.
Cubillos, L. & D. Arcos. 2002. Recruitment of common sardine (Strangomera bentincki) and anchovy (Engraulis ringens) off central-south Chile in the 1990s: There were effects of the 1997-98 El Niño event? Aquat. Liv. Res., 15 pp.
Cury, P. & C. Roy. 1989. Optimal environmental window and pelagic fish recruitment success in upwelling areas. Can. J. Fish. Aquat. Sci., 46: 670-680.
Cushing, D. H. 1975. Marine Ecology and Fisheries. Cambridge: Cambridge University Press.
Degens, E.T., W.G. Deuser & R.L. Haedrich. 1969. Molecular structure and composition of fish otoliths Mar. Biol., 2: 105-113.
Dunkelberger, DG., JM. Dean & N. Watabe. 1980. La ultraestructura de la membrana de los otolitos y de los otolitos en el menor mummichog Fundulus heteroclitus. Morfología Diario, 163: 367-377.
Espino, M. 1999. El Niño 1997-98: Su efecto sobre el ambiente y los recursos pesqueros en el Perú. Rev. Per. Biol., 3: 97-109.
FAO. 1988. Clupeoid fishes of the world Part 2: Engraulididae. Rome, pp. 322. Francis, R.I.C.C. 1990. Back-calculation of fish length: a critical review. J. Fish Biol., 36:
883–902. Fuiman, L. A. 2002. Special considerations of fish eggs and larvae, In: L. A. Fuiman & R.
G. Werner (eds.). The unique contributions of early life stages. Fish. Sci., 2: 1-32. Gaughan, D., W. Fletcher & K. White. 2001. Growth rate of larval Sardinops sagax from
ecosystems with different levels of productivity. Mar. Biol., 139: 831-837. Gauldie, R. W. 1988a. Microscopic growth increments in the otolith of orange roughy
Hoplostethus atlantieus and their potential use in studies of growth, recruitment and otolith structure. (Un- published manuscript held at Fisheries Research Centre, New Zealand Ministry of Agriculture and Fisheries, Wellington)
Geffen, A.J.1982. Otolith ring deposition in relation to growth rate in herring (Clupea harengus) and turbot (Scophthalmus maximus) larvae. Mar. Biol., 71: 317-326.
Gorini, F., V.E. Abachian & A.R. Giussi. 2010. Estimación de la edad y crecimiento de la Merluza austral Merluccius australis del mar Argentino. Rev. Invest. Desarr. Pesq. N° 20: 65-73.
Hare, J. A. & R. K. Cowen. 1994. Ontogeny and otolith microstructure of bluefish Pomatomus saltatrix (Pisces: Pomatomidae). Mar. Biol., 118: 541-550.
Hjort, J. 1914. Fluctuations in the great fisheries of northern Europe viewed in the light of biological research. Rapp. P.v. Reun. Cons. into Explor. Mer., 20: 1-228.

66
Houde, E. D. & C. E. Zastrow. 1993. Ecosystem- and taxon-specific dynamic and energetics properties of fish larvae assemblages. Bull. Mar. Sci., 53: 290–335.
Houde, E.D. 1997. Patterns and consequences of selective processes in teleost early life histories. In: Chambers, R.C., Tripprl, E.A. (eds.). Early Life History and Recruitment in Fish Populations, vol. 21. Fish and Fisheries Series, Chapman & Hall, 173–196 pp.
Hwang, S.D & T.W. Lee. 2005. Spawning dates and early growth of chub mackerel Scomber japonicus as indicated by otolith microstructure of juveniles in the inshore nursery ground. Fish. Sci., 71: 1185-1187.
Irie, T. 1960. El crecimiento de los otolitos de peces. Revista Facultad de Zootecnia Universidad de Hiroshima, 3: 203-221.
Jenkins, G. P. 1987. Age and growth of co-occurring larva of two flounder species, Rhombosolea tapirina and Ammotretis rostratus. Mar. Biol., 95: 157–166.
Jones, C. 1992. Development and application of the otolith increment technique. In: D.K. Stevenson and S.E. Campana (eds.). Otolith Microstructure Examination and Analysis. Can. J. Fish. Aquat. Sci., 117: 1-11.
Lagardere, F. 1989. Influence of feeding conditions and temperature on the growth rate and otolith-increment deposition of larval Dover sole (Solea solea (L.)). Rapp P-V Reun Cons Int Explor Mer, 191: 390-399.
Lasker, R. 1975. Field criteria for survival of anchovy larvae: the relation between inshore chlorophyll maximum layers and successful first feeding. U. S. Nat. Mar. Fish. Serv. Fish. Bull., 73: 453–462.
Legget, W.C. & E. Deblois. 1994. Recruitment in marine fishes: is it regulated by starvation and predation in the eggs and larval stages? Neth. J. of Sea Res., 32(2): 119-134.
Leta, H. & A. Keim. 1982. Lectura de edades en otolitos de peces teleósteos. FAO Documento Técnico de Pesca No. 39. Montevideo, Uruguay
Llanos-Rivera, A. & L. Castro. 2006. Interpopulation difererences in temperature effects on Engraulis ringens yolk-sac larvae. Mar. Ecol. Prog. Ser., 312: 245-253.
Lombarte, A. 1990. Efectos ambientales y fitogenéticos en la morfología del laberinto auditivo de los peces teleósteos. Tesis Doctoral, Universidad Autónoma de Barcelona, 335 pp.
Lough, R.G., M. Pennington, G.R. Bolz & A. Rosenberg. 1982. Age and growth of larval Atlantic herring. Clupea harengus, in the Gulf of Maine- Georges Bank region based on otolith growth increments. Fish. Bull., 80: 187-200.
Mann S, Parker SB, Ross MD, Skarnulis AJ, Williams RJP (1983). The ultrastructure
of the calcium carbonate balance organs of the inner ears. An ultra-high resolution electron microscopy study. Proceeding of the Research Society of London, 218: 187-217.
Marks, R. & D. Conover. 1993. Ontogenetic shift in the diet of young-of-the-year
bluefish, Pomatomus saltatrix, during the oceanic phase of the early life history. Fish. Bull., 91: 97-106.

67
Martínez, C., C. SaIazar, G. Böhm, J. Mendieta & C. Estrada. 1985. Efectos del fenómeno El Niño 1982-83 sobre los principales recursos pelágicos y su pesquería (Arica-Antofagasta). Invest. Pesq. Chile, 32: 129-139.
Maturana, J., M. Bello & M. Manley. 2004. El Niño-La Niña 1997-2000, Sus Efectos en Chile. S. Avaria, J. Carrasco, J. Rutllant y E. Yáñez (eds.). Comité Oceanográfico Nacional (CONA), Valparaíso, Chile.
McPhaden, M.J.1999. Génesis and evolution of the 1997–98 El Niño. Sci., 283: 950-954. Meekan, M.G. & L. Fortier. 1996. Selection for fast growth during the larval life of
Atlantic cod Gadus morhua on the Scotian Shelf. Mar. Ecol. Prog. Ser., 137: 25–37.
Methot R. D. 1983. Seasonal variation in survival of larval northern anchovy, Engraulis mordax, estimated from the age distribution of juveniles. Fish. Bull., 81: 741-750.
Miller, T. J., L. B. Crowder, J. A. Rice & E. A. Marschall. 1988. Larval size and recruitment mechanisms in fishes: toward a conceptual framework. Can. J. Fish. Aquat. Sci., 45: 1657–1670.
Morales-Nin, B. 1986b. Chemical composition of the sea bass (Dicentrarchus labrax, Pisces: Serranidae) otoliths. Cybium, 10 (2): 115-120.
Morales-Nin, B. 1991. Determinación del crecimiento de peces óseos en base a la microestructura de los otolitos. FAO Documento Técnico de Pesca No. 322. Roma, Italia.
Morales-Nin, B. & J. Panfili. 2002. III. Sclerochronological studies: why and how? A. Age estimation. In: J. Panfíli, H. Troadec, H. de Puntual and P.J. Wright (eds.). Manual of Schlerochronology in Fish, 416 pp.
Moreno, P., G. Claramunt, L.R. Castro. 2011. Período de transición de larva a menores en anchoveta Engraulis ringens . Longitud relacionada con la edad?. J. Fish Biol., 78(3): 825-837.
Mugiya, Y., N. Watabe & J. Yamada. 1981. Diurnal rhythm in otolith formation in thegoldfish. Comp. Biochem. Phys, 68: 659-662.
Ojanguren, A. F., F. G. Reyes-Gavilán & R. Rodríguez-Muñoz. 1999. Effects of temperature on growth and efficiency of yolk utilisation in eggs and pre-feeding larval stages of Atlantic salmon. Aqua. Inter., 7: 81–87.
Palomera, I. 1992. Spawning of anchovy Engraulis encracicolus in the Northwestern Mediterranean relative to hydrographic features un the region. Mar. Ecol. Prog. Ser. 79: 215-223.
Pannella, G. 1971. Fish otoliths: Daily growth layers and periodical patterns. Sci., 173: 1124–1127.
Pannella, G. 1974. Otolith growth patterns: An aid in age determination in temperate and tropical fishes. In: T. B. Bagenal (ed.), The ageing of fish, Unwin Brothers, La Surrey, 28-39.pp
Pannella, G. 1980. Growth patterns in fish sagittae. In: D. C. Rhoads & R. A. Lutz. (eds.). Skeletal growth of aquatic organisms: biological records of environmental change. Plenum, New York, 519-560 pp.
Plaza, G., S. Katayama & Omori. M. 2001. Otolith microstructure of the black rockfish, Sebastes inermis. Mar. Biol., 139: 797–805.

68
Plaza, G., H. Honda, H. Sakaji & K. Nashida. 2005. Preparing fish sagittae for examination of daily growth increments of young-of-the-year fishes: A modification of the embed method. J. Fish Biol., 66: 592-597.
Plaza, G., M. Ishida & D. Aoyama. 2008. Temporal patterns of growth in larval cohorts of the Japanese sardine Sardinops melanosticus in a coastal nursey area. J. Fish Biol., 73: 1284-1300.
Quiñones, C. & J. Lopez. 2001. Crecimiento de juveniles de la lisa blanca Mugil curema (Teleostei: Mugilidae) en una laguna costera del suroeste del golfo de California. Lat. Am. J. Aquat. Res., 39: 25-32.
Radtke, R. L. & J. M. Dean. 1982. Increment formation in the otoliths of embryos, larvae and juveniles of the mummichog, Fundulus heteroclitus. Fish. Bull., 80: 201–215.
Ricker, W. 1975. Computation and interpretation of biological statistic of fish population. Department of Environmental Fisheries and Marine Service. Fish. Bull., 191: 1-382.
Secor, D. H., J. M. Dean & E. H. Laban. 1992. Otolith removal and preparation for microstructural examination. In: D. K Stevenson & S. E Campana (eds.). Otolith Microstructural Examination and Analysis. Can. Spe. Publ. Fish. Aquat. Sci. 117 pp.
Serra, J. R. 1983. Changes in the abundance of pelagic resources along the Chilean coast. In: G. Sharp & J Csirke (eds.). Proceeding of the expert consulation to examine changes in abundance and species composition of neritic fish resources. FAO Fish Rep., 291: 255-284.
Shepherd, J. G. & D. H. Cushing, 1980. A mechanism for density dependent survival of larval fish as the basis of a stock-recruitment relationship. Journal du conseil/Conseil permanent international pour l’exploration de la mer, 39: 160–167.
Sigel, R. B. 1988. Population dynamics of the Japanese anchovy, Engraulis japonicus, Shirasu of the Enshu-Nada and Ise Bay fishery, Japan revealed through otolith daily increments. Ph. D. thesis, Univ. Tokyo.
Simkiss, K., 1974. Calcium metabolism of fish in relation to ageing. In T.B. Bagenal, (ed.). Ageing of fish, Unwin Brothers Ltd., Surrey England: 1-12.
Smith, C.L. 1983. Summary of round table discussions on back calculation. U. S. Dep. Commer. NOAA Tech. Rep. NMFS, 8: 45-47.
Sogard, S.M. 1991. Interpretation of otolith microstructure in juvenile winter flounder (Pseudopleuronectes americanus): ontogenetic development, daily increment validation, and somatic growth relationships. Can. J. Fish. Aquat. Sci., 48: 1862-1871.
Sosa, O. 1999. Impactos de "El Niño" en las pesquerías. Página: http://elnino.cicese.mx/impactos/pesquerias.htm. Revisado: 30 de Julio de 2012.
Takahashi, M., Y. Watanabe, T. Knoshita & C. Watanabe. 2001. Growth of larval and early juvenile Japanese anchovy, Engraulis japonicus, in the Kuroshio-Oyashio transition region. Fish. Ocean., 10: 235-247.
Takasuka, A., I. Aoki & I. Mitani. 2003. Evidencia de crecimiento selectivo de la depredación de larvas de anchoveta Engraulis japonicus japoneses en la bahía de Sagami. Ecol. Mar. Progr. Ser., 252: 223-238

69
Takasuka, A., I. Aoki & I. Mitani. 2004. Tres sinérgicos relacionados con el crecimiento en los mecanismos de la supervivencia a corto plazo de las larvas de anchoveta Engraulis japonicus japoneses en la bahía de Sagami. Ecol. Mar. Progr. Ser., 270: 217-228.
Tanaka, K., Y. Mugiya & J. Yamada. 1981. Effects of photoperiod and feeding on daily growth patterns in otoliths of juvenile Tilapia nilotica. Fish. Bull., 79: 459-466.
Tomás, J. & J. Panfili. 2000. Otolith microstructure examination and growth patterns of Vinciguerria nimbaria (Photichthyidae) in the tropical Atlantic Ocean. Fish. Res., 46: 131-145.
Tresierra, A. & Z. Culquichicon. 1993. Biología pesquera. Libertad, Trujillo, Perú, 432pp.
Tsuji, S. 1983. Study on recruitment mechanism of larval anchovy to shirasu fishing grounds in Sagami Bay based upon otolith daily increment analyses. PhD thesis, University of Tokyo.
Watabe, N., K. Tanaka, J. Yamada & J.M. Dean. 1982. Scanning electron microscope observation of the organic matrix in the otolith of the teleos fish Fundulus heteroclitus and Tilapia nilotica. J. Exp. Mar. Biol. Ecol., 58: 127–134.
Watanabe, Y. & T. Kuroki. 1997. Asymptotic growth trajectories of larval sardine Sardinops melanostictus in the costal waters off western Japan. Mar. Biol., 127: 369-378.
Wright, P.J., N.B. Metcalfe & J.E. Thorpe. 1990. Otolith and somatic growth rates in Atlantic salmon parr, Salmo salar L.: evidence against coupling. J. Fish Biol., 36: 241-249.
Yáñez, E., M.A. Barbieri & L. Santillán, 1992. Long-term environmental variability and
pelagic fisheries in Talcahuano, Chile. In: Payne, A. I., Brink, L. Mann, K. H., Hilborn, R. (Eds.), Benguela Trophic Functioning. S. Afr. J. Mar. Sci., 12: 175-188.
Yáñez, E., M.A. Barbieri & A. Montecinos. 1990. Relaciones entre las variaciones del medio ambiente y las fluctuaciones de los principales recursos pelágicos explotados en la zona de Talcahuano, Chile. In: M.A. Barbieri (ed.). Perspectivas de la Actividad Pesquera en Chile, Escuela de Ciencias del Mar, UCV, Valparaíso, 5: 49-62.
Zhang, Z. & N.W. Runham. 1992. Effects of food ration and temperature level on the growth of Oreochromis niloticus (L.) and their otoliths. J. Fish Biol., 40: 341–349.