plant innate immunity
DESCRIPTION
PlantTRANSCRIPT

SUBMITTED BY:Kavita B. Joshi
(Masc. .3rd SEM. BIOCHEMISTRY. ) REGISTRATION
NO: - J4-00629-2010Dept of BiochemistryCollege of Agriculture,
JAU, Junagadh
SUBMITTED TO:
Dr. M.V.Parakhiya,Assistant Professor
Dept of Biotechnology,College of Agriculture,
JAU, Junagadh
Plant Innate Immunity
1
Sub. –Biochem-506 ,IMMUNO CHEMISTRY
,

2
CONTENTIntroductionPlant Defense systems
Physical barriers Trachomas Epidermis
Chemical defenses Entering Plant CellsHow Plants Identify Different Parasites through
General PatternsEffecter-Triggered Susceptibility and Effecter-
Triggered Immunity Receptors Recognizing Pathogens, Blurring the
Borderline between PTI and ETI Stabilization, Signaling and Degradation of R Protein
Complexes Hypersensitive Response and Cell Death SignalingDefense Signal Transduction Remembering Previous Attacks referances

3
Introduction
Innate immunity is an ancient trait where various recognition systems distinguish between ‘self’ and ‘nonself’. This distinction provides the fundamental basis for altruistic behavior in ‘social’ microbes and ultimately how multicellular organisms could arise.
Like all living systems, plants need to protect themselves from parasites. Large, multicellular organisms with a comparably slow generation time must race against small and rapidly evolving parasites.
Despite this, plants and other multicellular organisms are not fighting a losing battle and infectious disease is more an exception than a rule. In fact, the immune response itself is a double-edged sword and must be tightly controlled in order not to cause disease itself or otherwise be detrimental for the organism.
Plant Defense systems
There are main 2 types of innate defense systems in plants
Physical barriers: trichomes, leaf hairs, waxy cuticles, etc
Chemical defenses; Preformed: antimicrobial compounds (e.g. phytoalexins) Induced: Hypersensitive response (HR, a localized host cell death at the site of infection), defense gene/protein expression, oxidative burst, increased levels of antimicrobial compounds.
4

5
Physical barriers

6
6
Trichomes (“leaf hairs”) are specialized epidermal cells found on aerial plant parts that may provide both physical and chemical protection against insect pests. The velvety appearance of dusty miller (Senecio cineraria) is caused by thousands of tiny trichomes covering the plant’s surface. Trichomes on the surface of soybeans (Glycine max) prevent insect eggs from reaching the epidermis and the larvae starve after hatching. The hook-shape of snap bean (Phaseolis vulgaris) trichomes impale caterpillars as they move across the leaf surface, and glandular trichomes in potato and tomato secrete oils that repel aphids. In woody plants, the periderm replaces the epidermis on stems and roots. Outer bark (phellem) is an excellent example of a preformed structural barrier that contains high amounts of water-resistant suberin and prevents many pathogens and insects from reaching the living cells underneath.
Trichomes
7
EpidermisThe epidermis constitutes the outermost protective tissue system of leaves, floral parts, fruits, seeds, stems, and roots of plants until they undergo considerable secondary growth. It is the first line of defense against invading pathogens and consists of both specialized and unspecialized cells. The epidermal cells of aerial plant parts are often covered in a waxy cuticle that not only prevents water loss from the plant, but also prevents microbial pathogens from coming into direct contact with epidermal cells and thereby limits infection. The cuticle can be relatively thin (aquatic plants) or extremely thick (cacti). The hydrophobic nature of the cuticle also prevents water from collecting on the leaf surface, an important defense against many fungal pathogens that require standing water on the leaf surface for spore germination. However, some fungal pathogens including Fusarium solani produce cutinases that degrade the cuticle and allow the fungi to penetrate the epidermis.

8
Chemical defenses

9
Entering Plant Cells
To invade a plant, phytopathogenic bacteria, fungi and oomycetes have evolved strategies to subvert host immunity and actively penetrate plant tissue. Wounded plants are sensitive since wounds constitute an easy entrance for many types of pathogens. Many bacteria such as Ps. syringae swim towards openings like stomata and hydathodes and enter the apoplastic space of plant tissue. From there, bacterial effectors can be injected into the cytoplasm via various secretion systems, like the type III secretion system in Pseudomonas and the type IV secretion system found in Agrobacterium. Several fungi have spores that upon germination on a host form an appressorium. This is a fungal structure that exerts high pressure on the plant cell walls allowing the fungal hyphae to enter and invaginate cell membranes. At this stage pathogens need either to bypass or suppress pathogen-associated molecular patterns (PAMP) triggered immune responses to proliferate and colonize the host tissue.
The plant endomembrane system exhibits a much higher degree of complexity than that of mammals or yeast (Jurgens, 2004). This is accompanied by an increased complexity of genes coding for proteins regulating vesicle trafficking. In plants N-ethylmaleimide-sensitive factor adaptor protein receptors or SNARE (soluble N-ethylmaleimide- sensitive factor attachment protein receptor)- mediated vesicle trafficking are of outmost importance in basal plant defense.

10
Gene-for-gene-type defense Induced upon Based on recognition of highly specific Avr-gene products by specialized, matching R-gene products infection by specialized pathogensHigh individual diversity due to meiotic gene shuffling Rapid reaction including "hypersensitive response"
Elicitor-induced defense ("basal defense") Induced upon infection by almost all microbes Based on recognition of general elicitors (= ubiquitous molecules characteristic for whole groups of microbes) Rapid activation of defense, but generally without a "hypersensitive response"
Gene-for-gene-type and elicitor-induced defense in plants

11
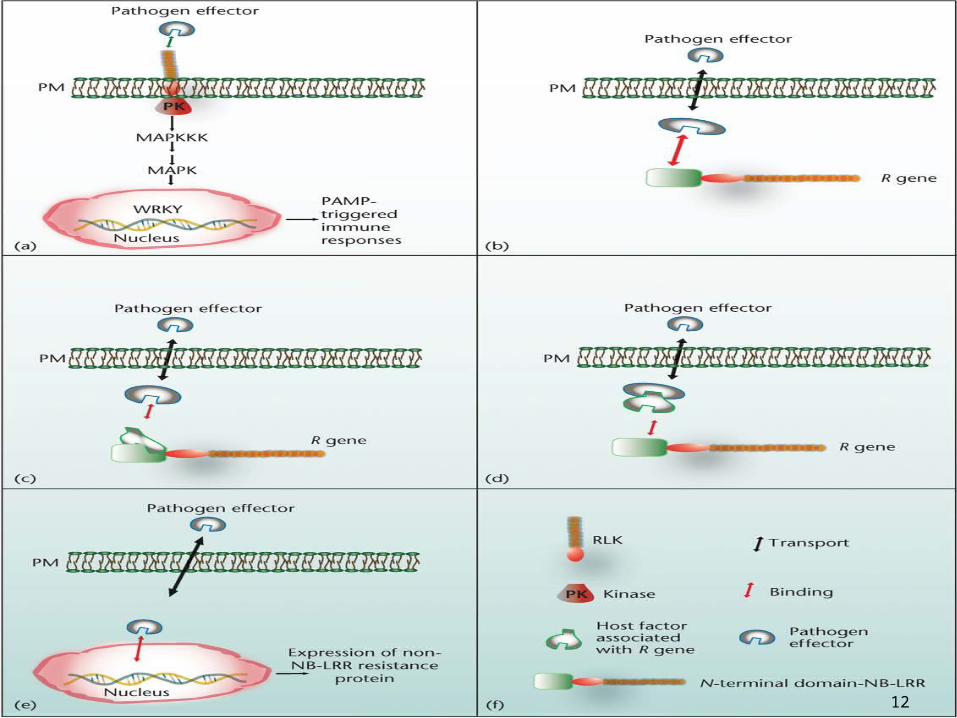
How Plants Identify Different Parasites through General Patterns
Different modes of pathogen recognition. (a)Recognition of microbe-associated molecular patterns (MAMPs)
by extracellular receptor-like kinases (RLKs) triggers basal immunity mediated via MAP kinase signalling and activation of WRKY transcription factors.
(b) Direct recognition can occur between pathogen effectors and an NB-LRR encoded R gene.
(c) NB-LRRs can indirectly recognize pathogens through theN-terminal domain via a host protein (guardee), alternatively
(d) the guardee component can first be associated with the pathogen effector and subsequently become recognized by the N-terminal of a NB-LRR encoding gene.
(e) The most recently found interaction is based on recognition of a pathogen effector that mimics a transcription factor and binds directly to a non-NB-LRR R protein.
(f) Explanation of the symbols used.
12 12
13
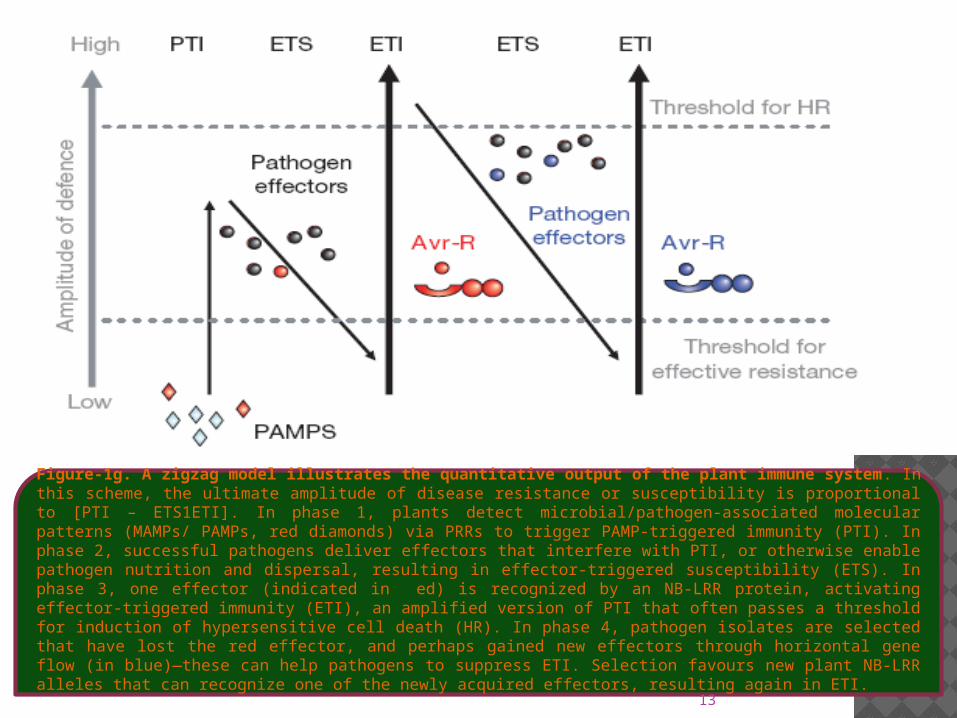
Figure-1g. A zigzag model illustrates the quantitative output of the plant immune system. In this scheme, the ultimate amplitude of disease resistance or susceptibility is proportional to [PTI – ETS1ETI]. In phase 1, plants detect microbial/pathogen-associated molecular patterns (MAMPs/ PAMPs, red diamonds) via PRRs to trigger PAMP-triggered immunity (PTI). In phase 2, successful pathogens deliver effectors that interfere with PTI, or otherwise enable pathogen nutrition and dispersal, resulting in effector-triggered susceptibility (ETS). In phase 3, one effector (indicated in ed) is recognized by an NB-LRR protein, activating effector-triggered immunity (ETI), an amplified version of PTI that often passes a threshold for induction of hypersensitive cell death (HR). In phase 4, pathogen isolates are selected that have lost the red effector, and perhaps gained new effectors through horizontal gene flow (in blue)—these can help pathogens to suppress ETI. Selection favours new plant NB-LRR alleles that can recognize one of the newly acquired effectors, resulting again in ETI.

14
RECEPTORS RECOGNIZING PATHOGENS, BLURRING THE BORDERLINE BETWEEN PTI AND ETI
Receptors detecting microbial patterns can be divided into surface and intracellular receptors. found by Chisholm et al. (2006) and Jones and Dangl (2006).
Since 1992, approximately 40 R genes have been cloned.These genes confer resistance to several classes of pathogens, including viruses, bacteria, fungi, oomycetes, insects and nematodes (Hammond-Kosack and Parker, 2003).
Surprisingly, the protein products of these R genes are structurally similar to each other and contain a few, conserved domains. The LRR domain is the most common domain among R proteins, and it is also found in animal innate immunity molecules, including Toll from Drosophila, and TLRs and nucleotide-binding oligomerization domain proteins (NODs) from mammals (Nu¨ rnberger et al., 2004; Staal and Dixelius, 2007).
Members of the largest class of R proteins comprise, in addition to the LRR, a central nucleotide-binding (NB) site domain that is similar to the NB of the NODs and the animal cell death effector proteins Apaf1 and CED4 (cell death 4), denoted NB-ARC (Apa1, R protein and Ced4 domain homology).
15
The NB-LRR class of R proteins is further subdivided according to the N-terminal domain of these proteins. Some proteins contain a Toll–interleukin 1 receptor (TIR) homology region domain, whereas others possess a coiledcoil domain. Like the LRR and NB domains, the TIR domain is found in animal innate immunity proteins, specifically Toll, the TLRs and their adaptor proteins TRIF (Toll/IL-1 receptor (TIR)-domain-containing adaptor protein-inducing IFN-beta) andMyD88 (Pa˚ lsson- McDermott and O’Neill, 2007).
The surface receptors mainly detect PAMPs and include receptor-like kinases (RLK), receptor-like proteins (RLP) and extracellular binding proteins. In fact the conceptually clear distinction between PAMPs and effectors can be indistinct. For example, the quorum-sensing signal protein AvrXa21, detected by the rice RLK Xa21, is an indispensable structure and at the same time a race-specific elicitor (Lee et al., 2006).
RLKs reside in plasma membranes and are composed of a putative extracellular ligand-binding domain, a single transmembrane domain and an intracellular serine/ threonine kinase domain. Thus, RLKs show to have structural similarities to animal receptor tyrosine kinases.
A recent global phenotyping of mutants in all Arabidopsis RLPs revealed a role in development, hormone sensing and non host resistance towards a bacterial pathogen (Wang et al., 2008).
Conti…

16
Hypersensitive Response and Cell Death Signaling
•Tentative Cell Death Circuitry in Disease Resistance in Arabidopsis.• Many inputs, detailed in the text, lead to cell death. Cell death can result in the accumulation of SA, which in turn leads to the activation of downstream defense pathways that culminate in local and systemic resistance. SA accumulation is blocked by expression of the nahG gene.• Signals depicted feeding into cell death could be in one or more linear pathways rather than independent inputs, as shown here. Question marks denote hypothetical positioning of various mutations. •Arrows imply activation steps; blunt symbols imply negative regulatory roles for the wild-type products of LSD7 and CPR7. Double-mutant analysis of lsd6 and lsdl provides genetic evidence for an amplification loop involved in determining the extent of cell death. Mutants noted here are from Bowling et ai. (1994), Cao et al. (1994), Delaney et al. (1994, 1995), Dietrich et al. (1994), and Century et al. (1995). • Note that the Isd suppressors will be called Phoenix @hx) after the mythological bird that arises from its own ashes. The sup9’Sdi mutant is now phx2‘sdi.

17
Defense Signal Transduction
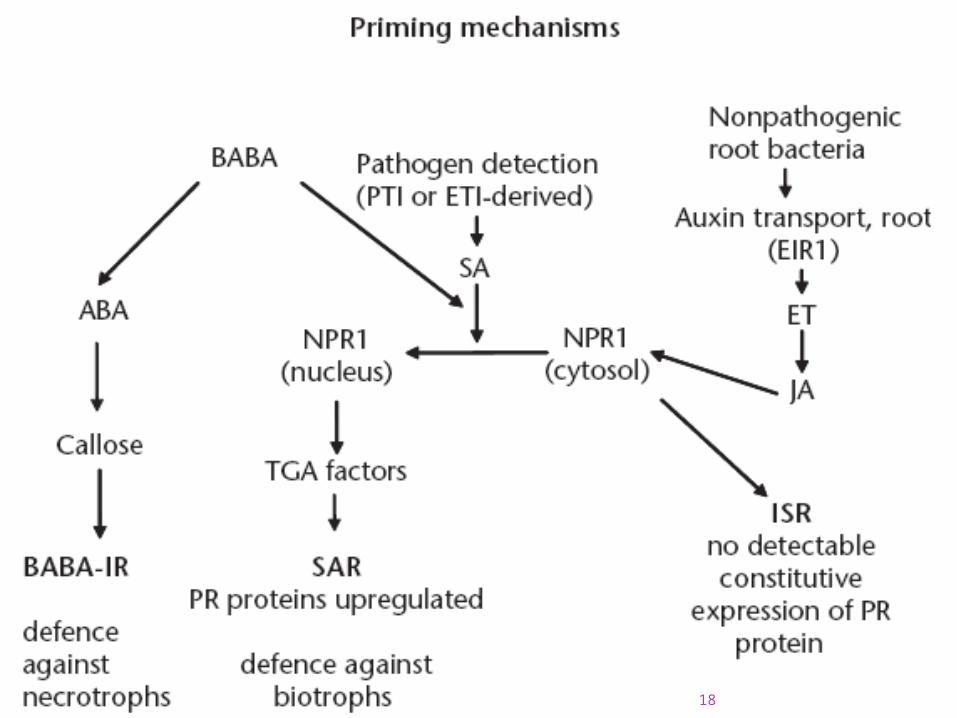
Priming is a nonacute plant pathogen response, which renders the plant more resistant to future attack. In ISR priming models, a role for NPR1 in JA responses has been established, whereas this role is not apparent during infection with necrotrophs – where the detrimental role of NPR1-mediated antagonism by SA is more important.
The chemical agent b-aminobutyric acid (BABA) induces priming of callose deposition via interference of ABA signalling, resulting in BABA-induced resistance (BABA-IR). BABA-IR can act independently of SAR and ISR against some pathogens and via the SAR pathway against others.
18

19
Remembering Previous Attacks
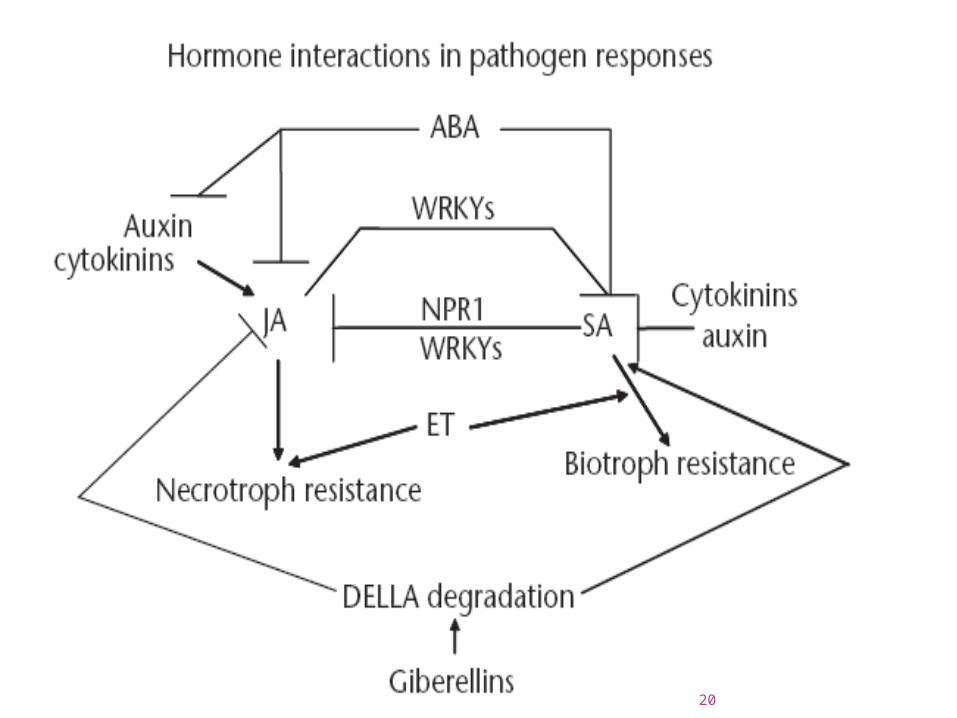
Through observations of disease outcome in various mutants, the mutual antagonism between defenses against necrotrophs and biotrophs was observed. Most classical models have focused on the twomain pathways with SA-dependent resistance against biotrophs and JA and/or ET-dependent responses versus necrotrophs.
As more mutants in other hormone pathways have been studied, a complex webof interactions has become more and more apparent. Some pathogens where the three main plant biotic defense hormones SA, JA and ET play no or aminor role in resistance, mutants in other hormone pathways have rendered susceptibility – indicating that there are yet undiscovered pathogen response pathways that need to be delineated and characterized in relation to the more well-established models.
Each interaction is also more complex than indicated by simplified model drawings, since the combination of signals can cause some subsets of the responses to be antagonistic whereas others are synergistic or unaffected. The models are thus not representative for every response gene, but rather for the final disease outcome in those plant–pathogen systems hitherto studied.
20
Remembering Previous Attacks

21
References
•Thomas Boller1 and Georg Felix2,’ a Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors’, Annu. Rev. Plant Biol. 2009. 60:379–406•Jonathan D. G. Jones1 & Jeffery L. Dangl2,’ The plant immune system’, Vol 444j16 November 2006jdoi:10.1038/nature05286.•Greenberg, J. T. & Yao, N. The role and regulation of programmed cell death in plant–pathogen interactions. Cell. Microbiol. 6, 201–211 (2004).•John M. McDowell and Jeffery L. Dangl,’ Signal transduction in the plant immune responseBoller1 and Georg Felix2,’ Perception of Microbe-Associated Molecular Patterns and Dang’ 2000, Elsevier Science Ltd. All rights reserved.PG-89 TO 82. •Thomas Boller1 and Georg Felix2,’ Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors’,
http://www.apsnet.org/edcenter/intropp/PathogenGroups/Pages/default.aspxhttp://jxb.oxfordjournals.org/content/61/10/2549.shorthttp://genesdev.cshlp.org/search?fulltext=Innate+immunity&sortspec=date&submit=Submit&andorexactfulltext=phrase

22