physiology of the rocky intertidal predator · pdf filephysiology of the rocky intertidal...
TRANSCRIPT

2816
Ecology, 82(10), 2001, pp. 2816–2829q 2001 by the Ecological Society of America
PHYSIOLOGY OF THE ROCKY INTERTIDAL PREDATOR NUCELLAOSTRINA ALONG AN ENVIRONMENTAL STRESS GRADIENT
ELIZABETH P. DAHLHOFF,1 BRADLEY A. BUCKLEY,2 AND BRUCE A. MENGE
Department of Biology and Center for Environmental Studies, Santa Clara University, Santa Clara, California 95053 USA,and Department of Zoology, Oregon State University, Corvallis, Oregon 97331 USA
Abstract. To gain insight into suborganismal mechanisms underlying responses ofpredators to environmental stress, we examined the effects of temperature and food avail-ability on the physiology and feeding activity of the predatory whelk Nucella ostrina(formerly N. emarginata). Studies were conducted in the mid-low transition zone on rockyreefs at two sites on the Oregon coast: Strawberry Hill (SH) and Boiler Bay (BB). SH isdominated by suspension-feeding invertebrates and their predators, while BB is dominatedby macrophytes. In wave-exposed (EX) and wave-protected (PR) habitats at each site, wemeasured air and water temperatures, whelk body temperatures, and indices of physiologicalcondition (malate dehydrogenase [MDH] activities and RNA : DNA ratios) for wild-caughtwhelks. The influence of food (6prey) and thermal conditions (6shelter) on prey con-sumption, physiological performance (oxygen consumption rate), and response to thermalstress (induction of Hsp70-class stress proteins) of whelks was determined in field exper-iments. Low-tide air temperatures, whelk body temperatures, and biochemical indices ofwhelk field condition were consistently different between sites (SH . BB) and wave ex-posures (temperatures: PR . EX, condition: PR , EX). Foraging activity, metabolic rates,and stress protein (Hsp) expression were consistently higher at SH than BB. Respirationrates were lower for all 2prey treatments than for 1prey treatments; however, whelks in2prey 1shade treatments had lower Hsp expression and higher respiration rates (an in-dication of better physiological condition) than whelks in 2prey 2shade treatments. Mostmeasures of performance suggested that wave-exposed individuals were more physiolog-ically robust than wave-protected individuals. Temporal physiological responses to stresssuggested that high levels of heat stress compromise whelk performance later in the summer.These data are consistent with the hypothesis that physiological stress interacts with preyabundance to modify activity of whelks. Detailed studies of the effects of environmentalstress on predator–prey interactions are needed to elucidate the relationship between phys-iological stress and predator–prey interactions. However, present evidence suggests thatwhelks alter activity (behavioral modification) and physiology (biochemical modification)in ways that lead to differential impacts by these predators on prey populations at differentsites.
Key words: community structure; enzymes; food availability; heat shock proteins; mussels; My-tilus; Nucella; oxygen consumption rates; RNA : DNA; rocky intertidal; wave exposure; whelks.
INTRODUCTION
Experimental field studies have taught us much aboutthe ecological processes determining community struc-ture (Paine 1966, 1974, Dayton 1971, Connell 1975,Menge and Sutherland 1976, 1987, Louda and Collinge1992, Bertness and Callaway 1994). These and otherstudies suggest that both biotic interactions and envi-ronmental stress shape the structure of communities(Menge and Olson 1990). Rocky intertidal communi-ties are ideal model systems for such studies becausethey harbor organisms that experience large spatial andtemporal variation in environmental conditions due to
Manuscript received 4 October 1999; revised 14 July 2000;accepted 26 September 2000; final version received 1 November2000.
1 E-mail: [email protected] Present address: Department of Biology, Arizona State
University, Tempe, Arizona 85287 USA.
steep gradients in wave impact, food availability, tem-perature, and desiccation (Paine 1977, Newell 1979,Denny 1988, Menge and Farrell 1989). Furthermore,key species interactions often vary significantly alongthese environmental gradients (Connell 1961a, b, Paine1966, 1974, 1984, Dayton 1971, 1975, Menge 1978a,b, Dayton et al. 1984, Menge and Sutherland 1987,Menge et al. 1994, 1999, Leonard et al. 1998, Bertnesset al. 1999). Environmental stress models predict thatabiotic stress will differentially affect interacting spe-cies, thereby altering the outcome of species interac-tions along environmental gradients (Menge and Olson1990). For example, consumer stress models predictthat foraging predators are more susceptible than ses-sile prey to abiotic stress. When foraging predators and/or sessile prey species have strong direct or indirecteffects on community structure, abiotic stress may playa disproportionately large role in shaping community

October 2001 2817EFFECTS OF STRESS ON WHELK ECOPHYSIOLOGY
structure (Menge 1976, Louda and Collinge 1992, Ar-nott and Vanni 1993).
Recent studies of the physiology of rocky intertidalinvertebrates have begun to yield insight into how en-vironmental stress may impact animals in nature (Bert-ness et al. 1991, Hofmann and Somero 1995, 1996a,b, Dahlhoff and Menge 1996, Roberts et al. 1997, Hof-mann 1999, Tomanek and Somero 1999). These andother studies have used either laboratory manipulationof the ‘‘stress’’ of interest, or have directly measuredphysiological indices of stress and performance in na-ture (Garrity 1984, Stickle et al. 1985, Stickle and Bay-ne 1987, Bayne and Hawkins 1990, Bayne et al. 1993,Abele et al. 1998). Few studies of marine systems todate have used field manipulations to examine the phys-iological mechanisms that ultimately underlie patternsof community structure, although this approach hasbeen used quite successfully in other systems (Mooneyet al. 1987, Bazzaz 1996).
In the present study, we address the issue of howspecies interactions vary with environmental stress bytesting one prediction of environmental stress models:That a mobile consumer will be physiologically com-promised under certain environmental conditions, andthese conditions may alter foraging activity. We mea-sured foraging activity, physiological performance, andresponse to stress in the dogwhelk Nucella ostrina (for-merly N. emarginata; see Palmer et al. 1990, Marko1998) along naturally occurring and experimentally-manipulated gradients of temperature and food avail-ability at two rocky intertidal sites: Strawberry Hill(SH; 448N159 N, 1248079 W) and Boiler Bay (BB;448409 N, 1248039 W).
Nucella ostrina is a common midzone predator thatpreys on mussels (Mytilus trossulus) and barnacles (es-pecially Balanus glandula) in rocky intertidal com-munities along the West Coast of North America. Pre-vious studies of the distribution, abundance, and re-lationships between whelks and other species at SHand BB have shown that N. ostrina can have a moderateto strong effect on community structure (Connell 1970,Dayton 1971, Menge 1992, Menge et al. 1994, Na-varrete 1996, Navarrete and Menge 1996). N. ostrinais thus ideal for examining the effects of stress on thephysiology and activity of a foraging predator.
Patterns of species distribution and abundance differstrikingly between SH and BB (Menge et al. 1994,1997a, b). In the mid-low transition zone, sessile in-vertebrate prey (notably the mussel Mytilus trossulusand the barnacle Balanus glandula) are larger and moreabundant at SH than at BB (Menge et al. 1994, 1997a,b, Sanford 1999a, b, Sanford and Menge 2000). Thewhelks Nucella ostrina and N. canaliculata are alsocommon at this tidal level, due in large part to highprey density (Navarrete et al. 2000). Phytoplanktonabundance and productivity are consistently higher atSH than at BB, and thus suspension feeders living atSH have more food available than do suspension-feed-
ers living at BB (Menge et al. 1996, 1997a, b). Higherfood availability at SH results in higher metabolic ratesand higher capacity for protein synthesis in mussels(Dahlhoff and Menge 1996) and barnacles (E. P. Dahl-hoff, unpublished data).
To assess ecophysiological responses of Nucella os-trina to environmental stress, we measured habitat tem-peratures, whelk body temperatures, and foraging ac-tivity. We also quantified indicators of performanceknown to respond rapidly to variation in food avail-ability and physiological condition: whole animal ox-ygen consumption (V̇O2), activities of the metabolicenzyme malate dehydrogenase, and RNA : DNA ratio(Hawkins and Bayne 1984, Stickle et al. 1985, Stickleand Bayne 1987, Lowery and Somero 1990, Dahlhoffand Menge 1996, Menge et al. 1997a, 1999). To furtherquantify physiological response to thermal stress, wemeasured expression of a 70 kD-class stress protein(Hsp70), as Hsp70 expression responds rapidly to tem-perature variation in rocky intertidal invertebrates(Hofmann and Somero 1996a, b, Roberts et al. 1997,Hofmann 1999, Tomanek and Somero 1999). The ra-tionale behind using these physiological measures issummarized in the next section. Using these indices ofstress and performance, we addressed the followingquestions: (1) Do whelks show evidence of being dif-ferently affected by stress in distinct thermal and di-etary microhabitats? (2) Do whelks compensate for lowprey abundance by decreasing their metabolic ratesand/or increasing their foraging activity? (3) Arewhelks that are ‘‘well-fed,’’ either due to high foragingactivity or higher prey density, better able to toleratethermal stress than those that are deprived access toprey?
BACKGROUND: ECOPHYSIOLOGICAL RESPONSES
Physiological response to food availability
Carnivorous invertebrates such as whelks living inrocky intertidal regions may experience variation infood quality, quantity, and feeding opportunities bothwithin and between sites due to variation in wave ex-posure, substratum topography, and tidal level (Na-varro and Winter 1982, Bayne and Hawkins 1990,Hawkins and Bayne 1992). Variation in food avail-ability can have profound effects on the physiology offoraging invertebrates (Wolcott 1973, Underwood andJernakoff 1984, Stickle et al. 1985, Stickle and Bayne1987, Branch and Moreno 1994, Trussell 1997). Thisvariation could result from a variety of factors, in-cluding wave exposure, availability of shade, compe-tition for food, and food quality. First, whelks livingin wave-exposed areas may have longer foraging timesthan individuals in wave-protected areas because oflonger periods of wetness, and thus lower thermal stressat wave-exposed sites (Lewis 1964, Tomanek and So-mero 1999). Whelks may also experience limitation inforaging activity in areas of high wave impact due to

2818 ELIZABETH P. DAHLHOFF ET AL. Ecology, Vol. 82, No. 10
the fact that foraging whelks are more susceptible todisplacement by heavy waves than nonforaging indi-viduals (Denny 1988). Second, whelks in areas of highprey density may have both more food available andmore shelter (among the prey) from both heavy waveactivity and heat stress at low tide (Davenport et al.1996, 1998). Third, there may be greater competitionfor prey at lower tidal heights, especially when the seastar Pisaster ochraceus is present (Navarrete 1996, Na-varrete and Menge 1996, Navarrete et al. 2000). Fourth,the ‘‘quality’’ of suspension-feeding prey may be high-est in areas where phytoplankton abundance is highest(Bertness et al. 1991, van Erkon Schurink and Griffiths1992, Kreeger 1993, Kreeger and Langdon 1993, Dahl-hoff and Menge 1996).
Variation in food availability may lead to adjust-ments in metabolic rates to bring metabolic demandsof organisms into alignment with available energy sup-plies (Hawkins and Bayne 1984, 1985, van Erkon Schu-rink and Griffiths 1992, Bayne et al. 1993, Kreeger1993). Variation in food availability should thus bereflected in a variation in physiological ‘‘condition,’’indexed by changes in metabolic rate or dietary state.Metabolic rate is most typically determined by mea-suring oxygen consumption rate (Stillman and Somero1996), or can be measured indirectly by assaying levelsof metabolic enzymes (Yang and Somero 1993, Dahl-hoff and Menge 1996, Stillman et al. 1996). Dietarystatus can be assayed using the RNA : DNA ratio, anindex of protein synthetic capacity (Wright and Hetzel1985, Martinez et al. 1992, Foster et al. 1993, Dahlhoffand Menge 1996). Both direct and indirect indicatorsof metabolic activity were important to the presentstudy. Malate dehydrogenase activities and RNA :DNA ratios were used as a ‘‘snapshot’’ of field met-abolic activity and dietary status, whereas oxygen con-sumption measurements were used to assess whole an-imal metabolism for field-manipulated individuals. Weexpected that physiological condition should be cor-related to prey density (SH . BB) and foraging op-portunity (EX . PR). However, at low tide in somemicrohabitats (e.g., Strawberry Hill, wave-protected[SHPR]), thermal stress may ameliorate the benefits ofhigh prey density.
Physiological response to temperature
Temperature has an important and pervasive influ-ence on the distribution and abundance of organismsvia its effects on physiological processes (Hochachkaand Somero 1984, Prosser 1986, Cossins and Bowler1987). Rocky intertidal habitats are often characterizedby steep thermal gradients. Thus intertidal inverte-brates may be particularly susceptible to the physio-logical effects of environmental temperature variation,especially since, as ectotherms, their body temperaturesare determined by environmental temperature (Ho-chachka and Somero 1984, Cossins and Bowler 1987).Stress proteins (heat shock proteins, or Hsp’s) minimize
cellular heat damage by refolding partially unfoldedproteins into their functional state (Lindquist 1986,Parsell and Lindquist 1993, Parsell et al. 1993, Hof-mann 1999). Temperature extremes often induce Hsp’sand enhance thermal tolerance (Parsell et al. 1993,Krebs and Bettencourt 1998, 1999, Krebs and Feder1998). Not surprisingly, the heat shock response ap-pears to be a critical component of adaptational strat-egies used by ectothermic animals to acclimatize tovariations in body temperature in situ (Hofmann andSomero 1995, 1996a, b, Roberts et al. 1997, Chappleet al. 1998, Feder 1999, Hofmann 1999). However, Hspproduction can incur a substantial physiological cost(Feder and Hofmann 1999, Hofmann 1999). One maythus expect a trade-off between Hsp production, phys-iological condition, and foraging activity for animalsexperiencing natural gradients in environmental stress.Unfortunately, this relationship has not been exploredfor any intertidal invertebrate, especially one that haspotentially strong effects on community structure viaits impact on prey distribution and abundance.
METHODS
Quantification of field temperatures
Ambient air and water temperatures were monitoredevery 30 min throughout the cage transplant studies(see Field-manipulated populations) using thermistorsencased in dataloggers (Onset Computer Corporation,Pocasset, Massachusetts). We secured loggers to therock using housings made of stainless-steel mesh, whiteVexar mesh, and cable ties. Loggers were placed at thesame tidal height as the experiments, and were installedaway from algal canopy, rocky ledges, or other sourcesof shade. Whelk body temperatures were measured us-ing a hand-held digital thermometer (Omega HH-82;Omega Vanzetti, Sharon, Massachusetts) equippedwith E-type Teflon-insulated 36 AWG fine-wire ther-mocouples. A small hole was drilled in the posteriorportion of the whelk’s shell, and the thermocouple wasgently inserted into this hole until it was touching, butnot piercing, soft tissue. Modeling clay was used tocover the insertion point, and the whelk was placedback where it was collected and allowed to re-attachto the substratum. Body temperature was recorded ev-ery 15 min. Recordings were initiated on consecutivelow tides as soon as the water receded at the beginningof low tide, and continued until the water returned.
Natural populations
Animal collections for physiological measure-ments.—From July 1993 to July 1994, whelks werecollected every 90–110 d at wave-exposed areas at SHand BB. Ten individuals were collected at randomlydetermined locations along a 10-m transect line laidparallel to the shore. Individuals were immediately dis-sected and frozen on dry ice in the field. Dissectedtissues were returned to the laboratory, where they were

October 2001 2819EFFECTS OF STRESS ON WHELK ECOPHYSIOLOGY
stored at 2708C until biochemical analyses were con-ducted.
RNA : DNA ratios.—Concentrations of RNA andDNA in foot tissue were determined by ethidium bro-mide fluorescence following the method of Bentle etal. (1981) as modified by Dahlhoff and Menge (1996).Tissues were thawed on ice, weighed, and homogenizedin 30 volumes 2 mol/L NaCl with a hand-driven glasshomogenizer (Kontes Duall, Vineland, New Jersey,USA). Fifty mL of each sample were incubated in 1.5mL of 0.005 mg/mL ethidium bromide and 0.10mg/mL Proteinase K at 378C for 90 min. After incu-bation, 0.5 mL buffer (80 mmol/L Tris-Cl, pH 7.5 at208C) was added, and fluorescence was recorded at 365nm excitation and 590 nm emission using a Perkin-Elmer (Oakbrook, Illinois) LS-5B luminescence spec-trofluorometer. RNA and DNA concentrations of tis-sues were estimated from a standard curve calculatedusing known quantities of RNA and DNA (Sigma calfthymus DNA, 1–4 mg; Sigma calf liver RNA, type IV,2–8 mg; Sigma Genosys, The Woodlands, Texas).
Malate dehydrogenase activities.—To assay malatedehydrogenase activities, foot tissue was thawed on ice,weighed, and immediately homogenized in 10 vol ice-cold 50 mmol/L potassium phosphate buffer (pH 6.8at 208C). The homogenate was centrifuged (48C, 5 minat 14 000g [137 293.1 m/s2]) and supernatant collected.Enzymes were assayed spectrophotometrically follow-ing the methods of Dahlhoff and Menge (1996). En-zymatic activities are reported in International Unitsper gram wet mass (IU/g).
Field-manipulated populations
Cage transplants, foraging rates and animal collec-tions.—All experimental whelks (15–20 mm length)used at both sites were collected in May from a locationof intermediate wave exposure at SH away from theexperimental reef area. Time and logistical constraintsdid not allow a reciprocal set of experiments using BBwhelks. Although this constrains generalizations fromour experiments (i.e., we cannot infer that BB whelkswould have necessarily shown similar responses), itdoes reduce the likelihood that any changes in exper-iments were due to genetic differences between whelkpopulations. Moreover, sampling studies (RNA : DNAratios, malate dehydrogenase [MDH]) from naturalpopulations native to each site and exposure providedresults consistent with those observed in experimental(SH-derived) whelks. Nonetheless, we remain sensitivein our interpretations to possible effects of whelksource on our results.
Whelks were taken to the laboratory, and oxygenconsumption rates were measured for 30 randomly se-lected individuals. All remaining individuals weremarked with nail polish, and transplanted into 10 3 103 5 cm stainless-steel wire-mesh cages with 6-mmopenings at BB and SH. Cages were secured to the rockwith stainless-steel screws. Ten marked whelks were
placed in each cage, which were assigned to one ofthree treatments: 1prey 1shelter, 2prey 1shelter, and2prey 2shelter. Whelks in 1prey 1shelter treatmentswere placed in cages into which small Mytilus trossulushad been translocated two weeks beforehand. Musselswere added throughout the experiment to these cagesto maintain continuous availability of food. In 1preycages, mussels were held against the rock with plasticmesh. Whelks had access to mussels at all times be-cause they could feed by extending their proboscisesthrough the mesh. Whelks in 2prey 1shelter treat-ments were placed in cages over bare, scraped rock towhich empty mussel shells had been attached with ma-rine epoxy (Z-spar, Kop-Coat, Pittsburgh, Pennsylva-nia). These shells provided shelter from the sun towhelks, controlling for the fact that M. trossulus pro-vide both food and shelter to the whelks in the 1prey1shelter treatments. Whelks in 2prey 2shelter treat-ments were placed in bare-rock cages without eitherfood or shelter. Each block of three treatments wasreplicated five times at each site 3 exposure combi-nation.
Two whelks were removed from each cage every 30d. Collected individuals were placed in a cooler on ice,and brought immediately to the laboratory at Newport,Oregon. Whelks were placed in seawater for two hr atambient seawater temperature (118–128C) to allowthem to recover from collection and handling. Col-lected whelks were replaced with unmarked, wild-caught individuals to maintain a constant number ofwhelks per cage throughout the experiment. During thefirst collection (June), the total number of empty M.trossulus shells was counted in each 1food 1sheltertreatment to determine foraging activity. Mussel shellscould not fall out of the cages, and most mussel shellsshowed evidence of whelk-induced damage (small drillholes). Foraging rates were calculated using the totalnumber of empty shells in the 1food 1shelter cages.Predation rates reported here for June represent a slightoverestimate of the actual rate because mussels pro-tected from whelks suffer a low level (#5%) of non-predator-caused mortality (E. P. Dahlhoff and B. A.Menge, unpublished data). Background mortality ratesdid not differ between sites, so adjusting rates to ac-count for background mortality was deemed unneces-sary. Post-June foraging rates were not analyzed be-cause nonpredator-related mortality (as determined bythe absence of drill marks) of transplanted mussels washigh and variable.
Oxygen consumption rates.—Oxygen consumptionrates (V̇O2) were determined using a Gilson differentialrespirometer (Gilson Medical Electronics, Middleton,Wisconsin) to measure metabolic rate of whelks heldin cages under different field treatments. Snails wereplaced in glass chambers containing 5 mL sterile-fil-tered seawater, and the chambers were connected to therespirometer. Whelks were allowed to equilibrate to theassay temperature (108C) for 45 min. This assay tem-

2820 ELIZABETH P. DAHLHOFF ET AL. Ecology, Vol. 82, No. 10
TABLE 1. Ambient air temperatures at Strawberry Hill (SH)and Boiler Bay (BB) in wave-protected (PR) and wave-exposed (EX) regions, June–August 1995.
Site andexposure
Air temperature (8C)
Mean 1 SE
No. daysabove 258C
SHPRBBPRSHEXBBEX
25.021.920.315.9
0.70.70.60.6
282219
3
perature was selected to approximate the seawater tem-perature the whelks would experience upon the returnof the water at the end of low tide, when the animalsbecome active in situ. Each measurement was con-ducted in triplicate 60-min time periods, with 10 minbetween each period, during which time the respirom-eter was opened to equilibrate the system with freshair. At the end of each triplicate, snails were removedfrom chambers, dissected, weighed, and frozen at2708C. The consumption of oxygen was linear withtime, suggesting specimens did not experience oxygenlimitation during the experiment. The slope of the re-sulting line was used to calculate mass-specific oxygenconsumption rate (Stillman and Somero 1996).
Tissue levels of Hsp70.—Tissue levels of Hsp70were measured throughout the summer for a subset ofwhelks that were held in the experimental cages underall treatments. Whelks were randomly sampled every30 d from cages in each site 3 exposure combination,and foot tissue was dissected from whelks in the fieldand immediately frozen on an aluminum block chilledon dry ice. Samples were transported back to the labon dry ice and stored at 2708C until processing. Wedetermined tissue levels of Hsp70 for field-manipulatedwhelks by Western Blot analysis following publishedmethods (Roberts et al. 1997, Dahlhoff and Rank2000). We quantified Hsp70 by chemiluminescence us-ing a commercially available anti-Hsp70 antibody thatrecognizes several isoforms of Hsp70 (MA3-001, Af-finity Bioreagents, Golden, Colorado) and a ‘‘second-ary’’ antibody, conjugated with peroxidase, used to lo-cate anti-Hsp70 antibody using x-ray film. We mea-sured band area and density using Sigma Scan Pro (re-lease 4.0 for PC; SPSS Science, Chicago, Illinois), anddetermined Hsp70 levels by linear regression to a serialdilution curve of four concentrations of a known quan-tity of pure Hsp72 (Human recombinant, SPP-855,Stressgen Biotechnologies, Victoria, British Columbia,Canada) on each blot. We routinely observed a singleband at 72 kD on experimental blots. However, bothnoninducible (70 kD) and stress-inducible (72 kD) iso-forms of Hsp70 may have been present. We remainsensitive to this issue in our interpretation of Hsp data.
Statistical analyses.—Regression analysis was usedto analyze diurnal measurements of whelk body tem-perature. For all other data we used analysis of variance(ANOVA), with ‘‘site’’ (SH and BB), ‘‘exposure’’ (EXand PR), time, food (prey), and shelter as treatments.All were regarded as fixed effects; our null hypotheseswere that whelk responses did not differ between thesesites of known ecological conditions, or between con-trasting conditions of exposure, time, food, or shelter.This experimental design constrains statistically basedinferences about whelk responses beyond these sites,wave exposures, or experimental conditions, but doesnot prevent consideration of more general implicationsof our results based on these studies, and other eco-logical and physiological knowledge in this system.
Two-way ANOVA was used to compare seasonal andsite-related variation of RNA : DNA ratios and enzy-matic activities of whelks. Enzyme concentrations weresquare-root transformed for analysis, whereas RNA :DNA ratios were used without transformation becauseassumptions of normality and independence of errorterms were met by the ratio data. Three-way and four-way ANOVA were used to analyze V̇O2 (site, wave-exposure, prey, and time) and Hsp70 (site, exposure,and time) data. Oxygen consumption rates were square-root transformed for analysis, whereas Hsp70 valueswere not transformed because assumptions of ANOVAwere met by the nontransformed data. Homogeneity ofvariances was confirmed using Cochran’s test (Wineret al. 1991). Statistical analyses were conducted usingSYSTAT (Release 7.0 for PC [Wilkinson 1996]) andJmp (Release 3.1 for PC, SAS Institute).
RESULTS
Habitat thermal regime and whelk body temperature
Data obtained from electronic loggers indicate thatthe thermal environment of whelks varies with site andexposure. Table 1 shows mean habitat temperatures(with SE) at each locale, as well as the number of daysthat maximal air temperature exceeded 258C for at least30 min. Strawberry Hill is warmer than Boiler Bay,and wave-protected areas are warmer than wave-ex-posed areas (Two-way ANOVA, Site: F 5 34.71, P ,0.0001; Exposure: F 5 70.74, P , 0.0001; site 3 ex-posure: NS). Air temperatures were higher in July andAugust than in June (data not shown for brevity). Thenumber of days that maximal air temperature exceeded258C, a temperature known to induce Hsp70 expressionfor whelks in the lab, was greatest at wave-protectedareas. Interestingly, the number of days that air tem-perature exceeded 258C was similar at Strawberry Hill,wave-exposed [SHEX] and Boiler Bay, wave-protected[BBPR] (Table 1). Minimum temperature at these siteswas equivalent to sea-surface temperature, which var-ied from 9.5 to 12.28C throughout the study (as judgedfrom high tide temperatures taken from loggers). High-er air temperatures at wave-protected areas are likelydue to lower water flow and lower wave splash. Atwave-exposed sites, wave splash can cool the surfaceof the rock (and thus animals) by 108–158C in a fewminutes (E. P. Dahlhoff, unpublished data). Other con-ditions that influence air and rock temperature, includ-

October 2001 2821EFFECTS OF STRESS ON WHELK ECOPHYSIOLOGY
FIG. 1. Measurements of whelk body temperatures duringlow tide at (A) Strawberry Hill and (B) Boiler Bay, on twoconsecutive days in July (2 July SH, 3 July BB). Body tem-peratures were recorded in randomly selected whelks ap-proximately every 15 min using hand-held digital thermom-eters. Data points are means (6 1 SE) of four individuals ateach wave exposure.
FIG. 2. Mussel-consumption rates of caged whelks at SHand BB. Foraging rates were determined by counting the num-ber of mussels present per cage at the beginning of the ex-periment and those still alive after 14 d, and dividing thenumber of consumed mussels by the number of whelks pre-sent in the cage. Data are means (6 1 SE) of data from fivecages per site 3 exposure. Asterisks indicate significance ofcomparisons between site by exposure (one-way ANOVA;**P , 0.005).
FIG. 3. Differences in (A) malate dehydrogenase activityand (B) RNA : DNA ratio of natural populations of whelksat SH and BB. Data are means (6 1 SE) of 10 individualscollected 90–110 d apart. Statistical analyses are reported inTable 2.
ing fog, rain, and degree of sun exposure, do not differbetween wave exposures, although they vary betweenSH and BB (Menge et al. 1997b).
Whelk body temperatures at low tide mirrored pat-terns observed in air temperature (Fig. 1). SHEXwhelks had higher body temperatures than Boiler Bay,wave-exposed [BBEX] whelks (compare filled symbolson Fig. 1A and B). Regression analysis suggests thatrates of body temperature increase varied with site andexposure (site 3 exposure interaction was significant,F 5 3.116, P , 0.05). EX whelks at BB warmed upmore slowly than did PR whelks or whelks at SH.
Mussel consumption
Mussel consumption by whelks varied with site andexposure (Fig. 2). Consumption rates were higher atEX areas at SH (Fig. 2, site 3 exposure: F 5 5.704,P , 0.05). Recent studies suggest that whelks havesimilar impacts on barnacle abundance at SH and BB
2822 ELIZABETH P. DAHLHOFF ET AL. Ecology, Vol. 82, No. 10
TABLE 2. Two-way ANOVA on the effect of site and timeon enzymatic activities and RNA : DNA ratios of Nucellaostrina.
Source ofvariation df SS F P
MDH activitySiteTimeSite 3 timeError
133
72
72.55178.3515.885
119.16
43.8235.913.198
,0.0001,0.0001,0.05
RNA : DNA ratioSiteTimeSite 3 timeError
133
72
3.0743.6751.2335.7096
38.7415.445.149
,0.0001,0.0001,0.005
Notes: Sites were Strawberry Hill and Boiler Bay; timecorresponds to measurements made approximately every 90d from July 1993 to July 1994. Site and time were treated asfixed variables.
TABLE 3. Four-way ANOVA on the effect of site, exposure, prey availability, and time onoxygen consumption rate (V̇O2) of Nucella ostrina.
Source of variation df SS F P
SiteExposurePrey availabilityTimeSite 3 exposure
11121
14.960.018.30
13.591.74
95.050.05
26.3643.1911.07
,0.000NS
,0.000,0.000,0.00
Site 3 prey availabilitySite 3 timeExposure 3 prey availabilityExposure 3 timeFeeding 3 time
12122
0.725.910.392.501.89
2.3018.78
1.237.943.01
NS
,0.00NS
,0.05NS
Site 3 exposure 3 prey availabilitySite 3 exposure 3 timeSite 3 prey availability 3 timeExposure 3 prey availability 3 timeSite 3 exposure 3 prey availability 3 timeError
12222
237
0.460.791.080.400.92
37.29
1.472.501.720.631.46
NS
NS
NS
NS
NS
Notes: Independent variables were site (Strawberry Hill and Boiler Bay), exposure (wave-exposed and wave-protected), prey availability (1prey and 2prey), and time (every 30 d fromMay to August 1995). Site, exposure, feeding, and time were treated as fixed variables. NS,not significant (P . 0.05).
(B. A. Menge, unpublished data). Whelks at SHEXmay thus feed faster but for less time than at other sitesor exposures.
Biochemical variation in natural populations.—Metabolic activity of N. ostrina, as indexed by malatedehydrogenase activities of foot tissue, was higher atSH than at BB, particularly in summer (Fig. 3, Table2; site 3 time interaction was significant). RNA : DNAratios of foot for SH whelks were also greater thanthose of BB whelks most times of the year (Fig. 3B,Table 2; site 3 time interaction was significant). Thesedata suggest that whelks at SH had higher somatic tis-sue growth rates than whelks at BB, as RNA : DNAratio is an excellent indicator of protein synthetic ca-pacity in animals with indeterminate growth. The ob-servation that whelk MDH activities and RNA : DNAratios are higher at SH than at BB suggests that whelksat SH have higher metabolic rates than BB whelks.
Physiological changes in transplanted whelks
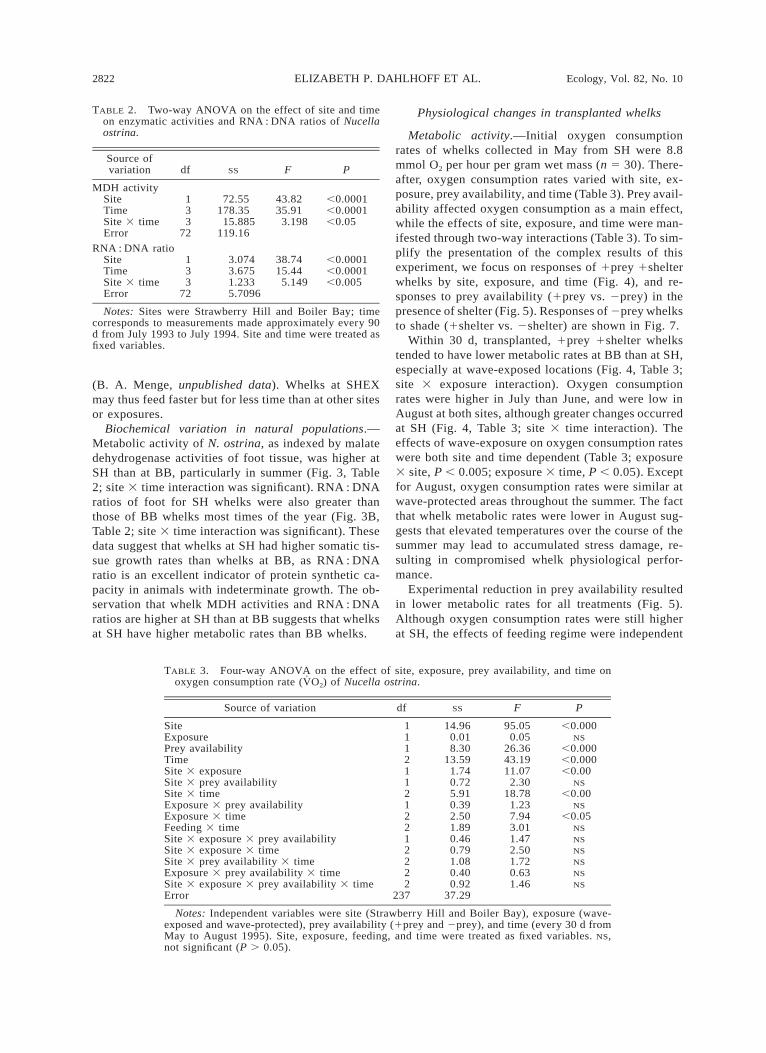
Metabolic activity.—Initial oxygen consumptionrates of whelks collected in May from SH were 8.8mmol O2 per hour per gram wet mass (n 5 30). There-after, oxygen consumption rates varied with site, ex-posure, prey availability, and time (Table 3). Prey avail-ability affected oxygen consumption as a main effect,while the effects of site, exposure, and time were man-ifested through two-way interactions (Table 3). To sim-plify the presentation of the complex results of thisexperiment, we focus on responses of 1prey 1shelterwhelks by site, exposure, and time (Fig. 4), and re-sponses to prey availability (1prey vs. 2prey) in thepresence of shelter (Fig. 5). Responses of 2prey whelksto shade (1shelter vs. 2shelter) are shown in Fig. 7.
Within 30 d, transplanted, 1prey 1shelter whelkstended to have lower metabolic rates at BB than at SH,especially at wave-exposed locations (Fig. 4, Table 3;site 3 exposure interaction). Oxygen consumptionrates were higher in July than June, and were low inAugust at both sites, although greater changes occurredat SH (Fig. 4, Table 3; site 3 time interaction). Theeffects of wave-exposure on oxygen consumption rateswere both site and time dependent (Table 3; exposure3 site, P , 0.005; exposure 3 time, P , 0.05). Exceptfor August, oxygen consumption rates were similar atwave-protected areas throughout the summer. The factthat whelk metabolic rates were lower in August sug-gests that elevated temperatures over the course of thesummer may lead to accumulated stress damage, re-sulting in compromised whelk physiological perfor-mance.
Experimental reduction in prey availability resultedin lower metabolic rates for all treatments (Fig. 5).Although oxygen consumption rates were still higherat SH, the effects of feeding regime were independent
October 2001 2823EFFECTS OF STRESS ON WHELK ECOPHYSIOLOGY
FIG. 4. Oxygen consumption rates (V̇O2) of 1food 1shel-ter whelks at (A) wave-exposed and (B) wave-protected areasat BB and SH. Snails in this treatment were given food (smallmussels) and shelter (shades built from empty mussel shells).Data are least-squares means (6 1 SE) of 10 individuals.Statistical analyses are reported in Table 3.
FIG. 5. Effects of prey availability on the metabolic ratesof SH-derived whelks at SH and BB. Data shown are com-bined least-squares means (6 1 SE) of 20 individuals pertreatment and exposure. All individuals were given shelter.Statistical analyses are reported in Table 3.
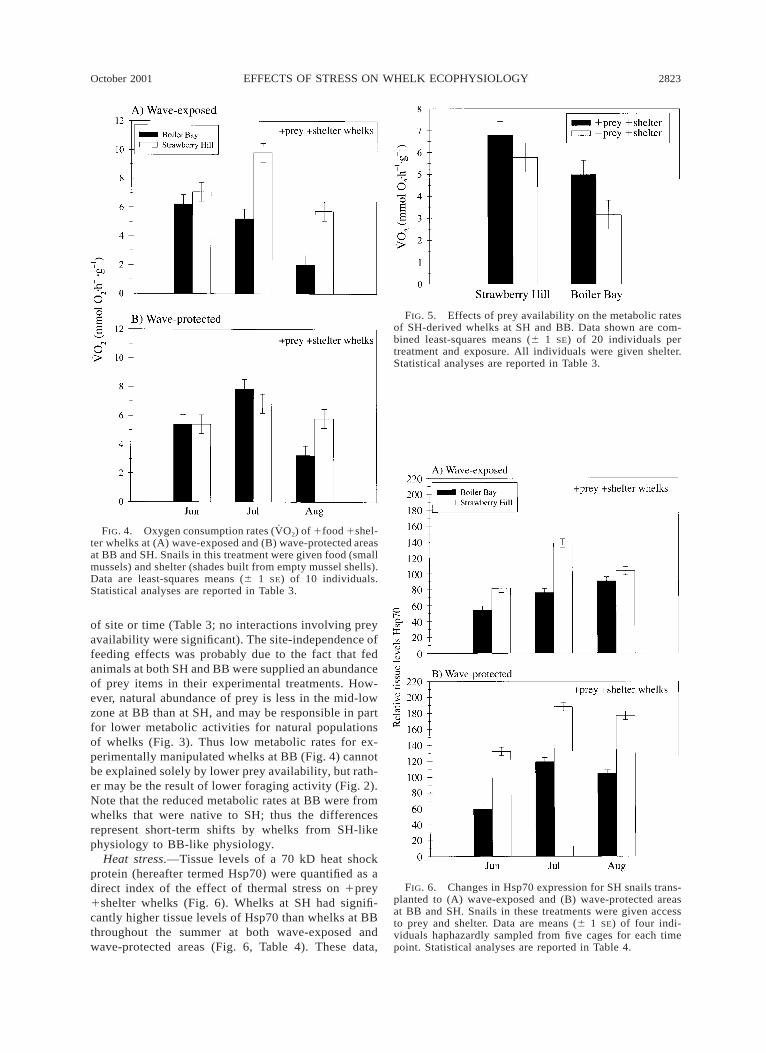
FIG. 6. Changes in Hsp70 expression for SH snails trans-planted to (A) wave-exposed and (B) wave-protected areasat BB and SH. Snails in these treatments were given accessto prey and shelter. Data are means (6 1 SE) of four indi-viduals haphazardly sampled from five cages for each timepoint. Statistical analyses are reported in Table 4.
of site or time (Table 3; no interactions involving preyavailability were significant). The site-independence offeeding effects was probably due to the fact that fedanimals at both SH and BB were supplied an abundanceof prey items in their experimental treatments. How-ever, natural abundance of prey is less in the mid-lowzone at BB than at SH, and may be responsible in partfor lower metabolic activities for natural populationsof whelks (Fig. 3). Thus low metabolic rates for ex-perimentally manipulated whelks at BB (Fig. 4) cannotbe explained solely by lower prey availability, but rath-er may be the result of lower foraging activity (Fig. 2).Note that the reduced metabolic rates at BB were fromwhelks that were native to SH; thus the differencesrepresent short-term shifts by whelks from SH-likephysiology to BB-like physiology.
Heat stress.—Tissue levels of a 70 kD heat shockprotein (hereafter termed Hsp70) were quantified as adirect index of the effect of thermal stress on 1prey1shelter whelks (Fig. 6). Whelks at SH had signifi-cantly higher tissue levels of Hsp70 than whelks at BBthroughout the summer at both wave-exposed andwave-protected areas (Fig. 6, Table 4). These data,
2824 ELIZABETH P. DAHLHOFF ET AL. Ecology, Vol. 82, No. 10
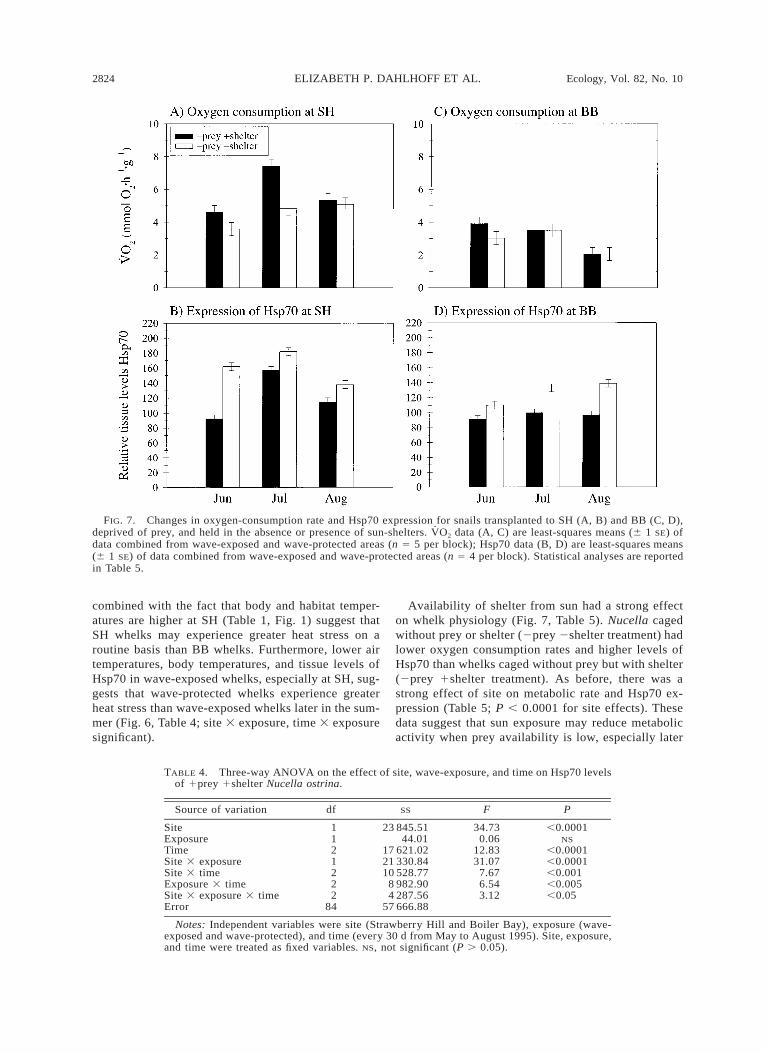
FIG. 7. Changes in oxygen-consumption rate and Hsp70 expression for snails transplanted to SH (A, B) and BB (C, D),deprived of prey, and held in the absence or presence of sun-shelters. V̇O2 data (A, C) are least-squares means (6 1 SE) ofdata combined from wave-exposed and wave-protected areas (n 5 5 per block); Hsp70 data (B, D) are least-squares means(6 1 SE) of data combined from wave-exposed and wave-protected areas (n 5 4 per block). Statistical analyses are reportedin Table 5.
TABLE 4. Three-way ANOVA on the effect of site, wave-exposure, and time on Hsp70 levelsof 1prey 1shelter Nucella ostrina.
Source of variation df SS F P
SiteExposureTimeSite 3 exposure
1121
23 845.5144.01
17 621.0221 330.84
34.730.06
12.8331.07
,0.0001NS
,0.0001,0.0001
Site 3 timeExposure 3 timeSite 3 exposure 3 timeError
222
84
10 528.778 982.904 287.56
57 666.88
7.676.543.12
,0.001,0.005,0.05
Notes: Independent variables were site (Strawberry Hill and Boiler Bay), exposure (wave-exposed and wave-protected), and time (every 30 d from May to August 1995). Site, exposure,and time were treated as fixed variables. NS, not significant (P . 0.05).
combined with the fact that body and habitat temper-atures are higher at SH (Table 1, Fig. 1) suggest thatSH whelks may experience greater heat stress on aroutine basis than BB whelks. Furthermore, lower airtemperatures, body temperatures, and tissue levels ofHsp70 in wave-exposed whelks, especially at SH, sug-gests that wave-protected whelks experience greaterheat stress than wave-exposed whelks later in the sum-mer (Fig. 6, Table 4; site 3 exposure, time 3 exposuresignificant).
Availability of shelter from sun had a strong effecton whelk physiology (Fig. 7, Table 5). Nucella cagedwithout prey or shelter (2prey 2shelter treatment) hadlower oxygen consumption rates and higher levels ofHsp70 than whelks caged without prey but with shelter(2prey 1shelter treatment). As before, there was astrong effect of site on metabolic rate and Hsp70 ex-pression (Table 5; P , 0.0001 for site effects). Thesedata suggest that sun exposure may reduce metabolicactivity when prey availability is low, especially later

October 2001 2825EFFECTS OF STRESS ON WHELK ECOPHYSIOLOGY
TABLE 5. Three-way ANOVA for effects of site and shelter of Hsp70 levels and V̇O2 of2prey whelks at Strawberry Hill and Boiler Bay.
Source of variation df SS F P
Oxygen consumptionSiteTreatmentTimeSite 3 treatmentSite 3 timeTreatment 3 timeSite 3 treatment 3 timeError
1121222
162
12.820.264.880.851.420.001.42
25.74
87.611.80
33.325.839.720.029.67
,0.0001NS
,0.0001,0.05,0.005
NS
,0.005
Levels of Hsp70SiteTreatmentTimeSite 3 treatmentSite 3 timeTreatment 3 timeSite 3 treatment 3 timeError
1121222
84
23 845.5144.01
17 621.0221 330.8410 528.77
8 982.904 287.56
57 666.88
34.730.06
12.8331.07
7.676.543.12
,0.0001NS
,0.0001,0.0001,0.001,0.005,0.05
Notes: Independent variables were site (Strawberry Hill and Boiler Bay), treatment (1shelterand 2shelter), and time (measured every 30 d from May to August 1995). Site, treatment, andtime were treated as fixed variables. NS, not significant (P . 0.05).
in the summer after many consecutive days of stress-fully high temperatures. This reduction in metabolicactivity may be caused by a shift of cellular energyfrom foraging activity to tissue repair, as evidenced byhigh levels of Hsp70 concomitant with lower metabolicrates in 2prey 2shelter whelks in July and August(Table 5; three-way ANOVA, P , 0.001 for three-wayinteraction of site, time, and treatment).
DISCUSSION
Our results suggest that physiological stress is tightlylinked to whelk metabolism over a range of environ-mental conditions at two rocky intertidal sites with dis-tinct community profiles. Modification of whelk activ-ity due to physiological stress may lead to differentialimpacts of these predators on prey abundance, whichmay ultimately impact community dynamics. We feelthat when combined with recent results demonstratingthe effects of physiological stress on major prey species(Hofmann and Somero 1995, Dahlhoff and Menge1996, Roberts et al. 1997), our results set the stage forstudies detailing the physiological mechanisms that un-derlie the community impacts of predator–prey inter-actions.
Environmental conditions and whelk ecophysiology
Habitat thermal regime and whelk body tempera-tures.—The thermal regime of Strawberry Hill (SH)and Boiler Bay (BB) was characterized by determiningthe substratum surface air temperature. Our data re-vealed that temperatures are warmest at SHPR, coolestat BBEX, and intermediate at SHEX and BBPR. Weused near-substratum air temperature rather than watertemperature to characterize thermal habitat because airtemperatures reach greater extremes, and are thus more
likely to be more stressful. For example, overall var-iation in water temperature during our study was smallin magnitude (9.58–12.28C) relative to variation in airtemperature (5.28–36.48C). Our direct measurements ofwhelk body temperatures, while limited in scope, wereconsistent with results using electronic loggers, al-though maximal whelk body temperatures were lowerthan maximal substratum temperatures. Whelks arethus likely to experience physiological stress duringexposure to elevated air temperatures that may com-promise their ability to forage when the tide returns.
The lower body temperatures of whelks comparedto logger temperatures may be due in part to the factthat whelks behaviorally regulate their body tempera-tures. Avoidance of potentially lethal temperatures isa key thermal regulatory strategy for many ectothermicanimals (Prosser 1986, Cossins and Bowler 1987). Atlow tide we observed whelks in microsites that wereboth exposed to and sheltered from thermal stress, in-cluding wedged in cracks, under large mussels, in themidst of barnacles or small mussels, and on open rocksurfaces. In the experimental treatments, whelks wereobserved inside the mussel shelters that were provided,but were also observed either out on open substratumor on top of their mussel prey. Thus although shelterwas available, whelks did not always avoid high sunexposure under either natural conditions or in experi-mental treatments. Our data suggest that whelks canexperience large excursions in body temperature (118–278C) during low tide, between sites, and among wave-exposures, and that they do not always avoid exposureto high air temperatures.
Physiological variation in natural populations.—Malate dehydrogenase activities and RNA : DNA ratioswere higher for whelks at SH than at BB. Differences

2826 ELIZABETH P. DAHLHOFF ET AL. Ecology, Vol. 82, No. 10
in food availability and quality may underlie differ-ences in whelk physiology between sites. Suspensionfeeders at SH are more abundant and have highergrowth rates, RNA : DNA ratios, metabolic activities,and tissue-protein contents than those at BB (Menge1992, Menge et al. 1994, 1997a, b, Dahlhoff and Menge1996, Stillman et al. 1996). These earlier data suggestthat SH prey have higher nutritional content than BBprey, as previous studies of mussel dietary physiologyhave shown that protein content of prey correlates withits dietary value (Bayne and Hawkins 1990, Bayne etal. 1993, Kreeger 1993, Kreeger and Langdon 1993,Kreeger et al. 1995).
Since suspension-feeders and their predators are im-portant players in rocky intertidal food webs, variationin both the density and caloric content of prey couldultimately have important impacts on community struc-ture. Previous research revealed that the overall impactof predation in the low intertidal zone was weaker atBB than SH (Menge 1992, Menge et al. 1994, 1996,Navarrete 1996, Navarrete and Menge 1996). Not sur-prisingly, most of the difference in predation impactwas due to Pisaster ochraceus, not to Nucella. Fieldexperiments suggested that whelk predation, both on aper population and per capita basis, was similar in mag-nitude at wave-exposed sites at BB and SH (Navarreteand Menge 1996). Since foraging time is limited atboth sites by tidal flux, and may be limited by hightemperatures in wave-protected regions, SH whelksmay obtain higher energetic return on their foragingactivity by foraging faster and on prey with highercaloric value. Therefore, our results suggest that phys-iological acclimatization of predators to variation infood quality or prey density may be an important factorunderlying variation in the impact of predation on com-munity dynamics.
Differences in metabolic activity between naturalpopulations of whelks at SH and BB were most pro-nounced during the summer. There was significant sea-sonal variation in both RNA : DNA ratio and MDHactivity at both sites, with metabolic activity being low-est in February and highest in July. Several factors arelikely to contribute to this result. First, like many ec-tothermic animals, whelks become less active duringthe winter, and their metabolic activity is suppressed,independent of spatial variability in physical factors orbiological interactions (Newell 1979). Second, preyabundance and prey quality are highest in the summer(Dahlhoff and Menge 1996, Menge et al. 1997a, b).Third, temperatures are highest in the summer, whichmay lead to elevated activity. We discuss these pos-sibilities below with respect to our measurements fromexperimentally manipulated individuals.
Metabolic activity of experimentally manipulatedwhelks.—Whelks at SH had higher metabolic rates (asindexed by oxygen consumption) than whelks at BB.At wave-exposed areas, this pattern developed within30 d of transplantation, and was especially pronounced
in July. At wave-protected areas, metabolic rates weresimilar for whelks in both BB and SH experiments untilAugust. Whelks are ectotherms, and increased bodytemperatures in ectotherms usually increases activity,at least on a short-term basis (Prosser 1986). The higherbody and habitat temperatures observed for SH whelksmay result in greater foraging activity. Conversely, thelower temperatures and foraging rates at BB may haveproduced the lower metabolic rates there. However,elevated body temperatures cannot directly explainhigher experimental oxygen consumption rates. Al-though whelks were acclimatized to different thermalconditions in the field, temperatures at which oxygenconsumption rates were measured were identical for allspecimens. If whelks differentially altered metabolicrates in response to temperature, we would actuallyexpect lower metabolic rates (at any given temperature)for warm-acclimated animals (the Q10 effect; Prosser1986). A more likely scenario is that high body tem-peratures led to higher activity in SH whelks, whichthen led to high ingestion of prey, which resulted inhigh metabolic rates. However, this dietary model doesnot explain the fact that whelk metabolic rates werehighest in July and dropped dramatically in August.This effect may be due to decreases in prey quality,but is more likely due to accumulated physiologicalstress due to several months of exposure to elevatedair temperatures.
Tissue levels of Hsp70 in experimentally manipu-lated whelks.—Tissue levels of Hsp70 were higher forwhelks at SH than at BB, and were higher at wave-protected areas than wave-exposed areas. Since Hsp70is a direct indicator of physiological stress, Hsp70 pro-duction is likely to have a strong impact on whelkactivity, especially late in the summer after severalmonths of exposure high air temperatures during lowtide. Heat shock proteins, expressed to repair proteinsdamaged by high temperature, are energetically costlyto produce (Lindquist 1986, Parsell and Lindquist1993). High tissue levels of Hsp70 in August whelksmay mean that energy that was being used for activity(and possibly growth) early in the summer has beenshifted to housekeeping and repair functions later inthe summer.
The data presented here are consistent with otherstudies of Hsp70 in natural populations of rocky in-tertidal invertebrates (Hofmann and Somero 1995,1996a, b, Roberts et al. 1997, Tomanek and Somero1999). Those studies showed that Hsp70 expressionwas higher for mussels and snails at wave-protectedsites than wave-exposed sites, and higher in the sum-mer than in the winter. It was somewhat surprising thatwe observed similar patterns of variation for mobilepredators. Although some mobile ectotherms can es-cape high temperatures behaviorally (Prosser 1986),our data are more consistent with the hypothesis thatwhelks are exposed to significant site- and exposure-related gradients in temperature. Recent studies of

October 2001 2827EFFECTS OF STRESS ON WHELK ECOPHYSIOLOGY
snails in the genus Tegula also suggest that mobilitydoes not guarantee a rocky intertidal invertebrate es-cape from physiological heat stress (Tomanek and So-mero 1999). This observation may be due to the factthat mobile foragers are time-limited by tidal flux andwave activity in many rocky intertidal systems. Thusan animal may ‘‘sacrifice’’ its ability to avoid heatstress to remain in an area where prey density is highor abundance of competitors is relatively low. In thesecases, the ability of an individual to recover from heatstress (by launching the heat shock response) may bea key component of its success in any given region onthe rocky intertidal.
CONCLUSIONS
The data presented in this study show strong, con-sistent patterns in physiological variation for the rockyintertidal predator Nucella ostrina over a range of tem-peratures, wave-exposures, and prey densities. Whelksliving in areas with high prey abundance are more phys-iologically ‘‘robust’’ (i.e., have higher energy stores)even when conditions are physiologically stressful.These results are consistent with the hypothesis thatbottom-up ecological effects can be a major determi-nant of the relative success of foraging predators, asthey are for suspension-feeding prey. Although the pre-sent study does not provide definitive evidence thatcommunity differences between, or within, SH and BBare caused by variation in whelk physiology, it suggeststhat whelks are able to alter the intensity of their ac-tivities to respond to heterogeneous environmental con-ditions. This study lays the groundwork for future in-vestigations exploring the differential impacts of pred-ators along environmental stress gradients and how thatmay ultimately affect community dynamics.
ACKNOWLEDGMENTS
We thank two anonymous reviewers and the editor for theirinsightful comments that greatly improved the manuscript.We thank G. Dahlhoff, S. Grist, F. Kemper, R. Scholes, M.Sullivan, and J. Troll for their invaluable assistance in thefield and laboratory. We thank G. Hofmann for her assistancewith heat shock protein techniques. Finally, we especiallyacknowledge J. Clegg and the staff of Bodega Marine Lab-oratory, University of California Davis, for their generoussupport of E. Dahlhoff’s research program. This work wasfunded by Andrew Mellon Foundation grants to B. Mengeand J. Lubchenco, and the Thomas Terry Faculty ResearchEnhancement grant to E. Dahlhoff, Santa Clara University.This is contribution no. 29 of the Partnership for Interdisci-plinary Studies of Coastal Oceans (PISCO): a Long-term Eco-logical Consortium funded by the David and Lucile PackardFoundation.
LITERATURE CITED
Abele, D., B. Burlando, A. Viarengo, and H. O. Poertner.1998. Exposure to elevated temperatures and hydrogenperoxide elicits oxidative stress and antioxident responsein the Antarctic intertidal limpet Nacella concinna. Com-parative Biochemistry and Physiology B 120:425–435.
Arnott, S. E., and M. J. Vanni. 1993. Zooplankton assem-blages in fishless bog lakes: influence of biotic and abioticfactors. Ecology 74:2361–2380.
Bayne, B. L., and A. J. S. Hawkins, editors. 1990. Filter-feeding in bivalve molluscs: controls on energy balance.Karger Press, Basel, Switzerland.
Bayne, B. L., J. P. Iglesias, A. J. Hawkins, E. Navarro, M.Heral, and M. J. Deslous-Paoli. 1993. Feeding behavior ofthe mussel Mytilus edulis: responses to variations in thequantity and organic content of the seston. Journal of theMarine Biological Association of the United Kingdom 73:813–829.
Bazzaz, F. A. 1996. Plants in changing environments: linkingphysiological, population, and community ecology. Cam-bridge University Press, New York, New York, USA.
Bentle, L. A., S. Dutta, and J. Metcoff. 1981. The sequentialenzymatic determination of DNA and RNA. AnalyticalBiochemistry 116:5–16.
Bertness, M. D., and R. Callaway. 1994. Positive interactionsin communities. Trends in Ecology and Evolution 9:191–193.
Bertness, M. D., S. D. Gaines, D. Bermudez, and E. S. San-ford. 1991. Extreme spatial variation in the growth andreproductive output of the acorn barnacle Semibalanus bal-anoides. Marine Ecology Progress Series 75:91–100.
Bertness, M. D., G. H. Leonard, J. M. Levine, and J. F. Bruno.1999. Climate-driven interactions among rocky intertidalorganisms caught between a rock and a hot place. Oecol-ogia 120:446–450.
Branch, G. M., and C. A. Moreno. 1994. Intertidal and sub-tidal grazers. Pages 75–100 in R. W. Siegfried, editor.Rocky shores: exploitation in Chile and South Africa.Springer-Verlag, New York, New York, USA.
Chapple, J. P., G. R. Smerdon, R. J. Berry, and A. J. S.Hawkins. 1998. Seasonal changes in stress protein 70 lev-els reflect thermal tolerance in the marine bivalve Mytilusedulis L. Journal of Experimental Marine Biology and Ecol-ogy 229:53–68.
Connell, J. H. 1961a. Effects of competition, predation byThais lapillus, and other factors on natural populations ofthe barnacle Balanus balanoides. Ecological Monographs31:61–104.
Connell, J. H. 1961b. The influence of interspecific compe-tition and other factors on the distribution of the barnacleChthamalus stellatus. Ecology 42:710–723.
Connell, J. H. 1970. A predator–prey system in the marineintertidal region. I. Balanus glandula and several predatoryspecies of Thais. Ecological Monographs 40:49–78.
Connell, J. H. 1975. Some mechanisms producing structurein natural communities: a model and evidence from fieldexperiments. Pages 460–490 in M. L. Cody and J. M. Di-amond, editors. Ecology and Evolution of Communities.Belknap Press, Cambridge, Massachusetts, USA.
Cossins, A. R., and K. Bowler. 1987. Temperature Biologyof Animals. Chapman and Hall, New York, New York,USA.
Dahlhoff, E. P., and B. A. Menge. 1996. Influence of phy-toplankton concentration and wave exposure on the eco-physiology of Mytilus californianus. Marine Ecology Pro-gress Series 144:97–107.
Dahlhoff, E. P., and N. E. Rank. 2000. Functional and phys-iological consequences of genetic variation at phosphog-lucose isomerase: heat shock protein expression is relatedto enzyme genotype in a montane beetle. Proceedings ofthe National Academy of Sciences USA 97:10056–10061.
Davenport, J., P. G. Moore, and E. Lecomte. 1996. Obser-vations on defensive interactions between predatory dog-whelks, Nucella lapillus L. and mussels, Mytilus edulis L.Journal of Experimental Marine Biology and Ecology 206:133–147.
Davenport, J., P. G. Moore, S. H. Maghill, and L. A. Fraser.1998. Enhanced condition in dogwhelks Nucella lapillus

2828 ELIZABETH P. DAHLHOFF ET AL. Ecology, Vol. 82, No. 10
L. living under mussel hummocks. Journal of Marine Bi-ology and Ecology 230:225–234.
Dayton, P. K. 1971. Competition, disturbance, and commu-nity organization: the provision and subsequent utilizationof space in a rocky intertidal community. Ecological Mono-graphs 41:351–389.
Dayton, P. K. 1975. Experimental evaluation of ecologicaldominance in a rocky intertidal algal community. Ecolog-ical Monographs 45:137–159.
Dayton, P. K., V. Currie, T. Gerrodette, B. D. Keller, R. Ro-senthal, and D. Ven Tresca. 1984. Patch dynamics andstability of some California kelp communities. EcologicalMonographs 54:253–289.
Denny, M. W. 1988. Biology and the mechanics of the wave-swept environment. Princeton University Press, Princeton,New Jersey, USA.
Feder, M. E. 1999. Organismal, ecological, and evolutionaryaspects of heat-shock proteins and the stress response: es-tablished conclusions and unresolved issues. American Zo-ologist 39:857–864.
Feder, M. E., and G. E. Hofmann. 1999. Heat-shock proteins,molecular chaperones and the heat-shock response: evo-lutionary and ecological physiology. Annual Review ofPhysiology 61:243–282.
Foster, A. R., D. F. Houlihan, and S. J. Hall. 1993. Effectsof nutritional regime on correlates of growth rate in juvenileAtlantic cod Gadus morula: comparisons of morphologicaland biochemical measurements. Canadian Journal of Fish-eries and Aquatic Science 50:502–512.
Garrity, S. D. 1984. Some adaptations of gastropods to phys-ical stress on a tropical rocky shore. Ecology 65:559–574.
Hawkins, A. J. S., and B. L. Bayne. 1984. Seasonal variationin the balance between physiological mechanisms of feed-ing and digestion in Mytilus edulis (Bivalvia: Mollusca).Marine Biology 82:233–240.
Hawkins, A. J. S., and B. L. Bayne. 1985. Seasonal variationin the relative utilization of carbon and nitrogen by themussel Mytilus edulis: budgets, conversion efficiencies andmaintenance requirements. Marine Ecology Progress Series25:181–188.
Hawkins, A. J. S., and B. L. Bayne. 1992. Physiologicalinterrelations and the regulation of production. Pages 171–222 in E. Gosling, editor. The mussel Mytilus: ecology,physiology, genetics and culture. Elsevier, Amsterdam, TheNetherlands.
Hochachka, P. W., and G. N. Somero. 1984. Biochemicaladaptation. Princeton University Press, Princeton, New Jer-sey, USA.
Hofmann, G. E. 1999. Ecologically relevant variation in in-duction and function of heat shock proteins in marine or-ganisms. American Zoologist 39:889–900.
Hofmann, G. E., and G. N. Somero. 1995. Evidence for pro-tein damage at environmental temperatures: seasonalchanges in levels of ubiquitin conjugates and hsp70 in theintertidal mussel Mytilus trossulus. Journal of ExperimentalBiology 198:1509–1518.
Hofmann, G. E., and G. N. Somero. 1996a. Protein ubiqui-tination and stress protein synthesis in Mytilus trossulusoccurs during recovery from tidal emersion. Molecular Ma-rine Biology and Biotechnology 5:175–184.
Hofmann, G. E., and G. N. Somero. 1996b. Interspecific var-iation in thermal denaturation of proteins in the congenericmussel Mytilus trossulus and M. galloprovincialis: evidencefrom the heat-shock response and protein ubiquitination.Marine Biology 126:65–75.
Krebs, R. A., and B. R. Bettencourt. 1998. Heat shock proteinvariation and the evolution of thermotolerance. AmericanZoologist 38:72–81.
Krebs, R. A., and B. R. Bettencourt. 1999. Evolution of ther-
motolerance and variation in the heat shock protein, Hsp70.American Zoologist 39:910–919.
Krebs, R. A., and M. E. Feder. 1998. Hsp70 and larval ther-motolerance in Drosophila melanogaster: How much isenough and when is more too much? Journal of InsectPhysiology 44:1091–1101.
Kreeger, D. A. 1993. Seasonal patterns in utilization of di-etary protein by the mussel Mytilus trossulus. Marine Ecol-ogy Progress Series 95:215–232.
Kreeger, D. A., A. J. S. Hawkins, B. L. Bayne, and D. W.Lowe. 1995. Seasonal variation in the relative utilizationof dietary protein for energy and biosynthesis by the musselMytilus edulis. Marine Ecology Progress Series 126:177–184.
Kreeger, D. A., and C. J. Langdon. 1993. Effects of dietaryprotein content on the growth of juvenile mussels, Mytilustrossulus. Biological Bulletin 185:123–139.
Leonard, G. H., J. M. Levine, P. R. Schmidt, and M. D.Bertness. 1998. Flow-driven variation in intertidal com-munity structure in a Maine estuary. Ecology 79:1395–1411.
Lewis, J. R. 1964. The ecology of rocky shores. EnglishUniversities Press, London, UK.
Lindquist, S. 1986. The heat-shock response. Annual Reviewof Biochemistry 55:1151–1191.
Louda, S. M., and S. K. Collinge. 1992. Plant resistance toinsect herbivores: a field test of the environmental stresshypothesis. Ecology 73:153–169.
Lowery, M. S., and G. N. Somero. 1990. Starvation effectson protein synthesis in red and white muscle of the barredsand bass Paralabrax nebulifer. Physiological Zoology 63:630–648.
Marko, P. B. 1998. Historical allopatry and the biogeographyof speciation in the prosobranch snail genus Nucella. Evo-lution 52:757–774.
Martinez, G., M. Torres, E. Uribe, M. A. Diaz, and H. Perez.1992. Biochemical composition of broodstock and earlyjuvenile Chilean scallops, Argopecten purpuratus L. heldin two different environments. Journal of Shellfish Re-search 11:307–313.
Menge, B. A. 1976. Organization of the New England rockyintertidal community: role of predation, competition andenvironmental heterogeneity. Ecological Monographs 46:355–393.
Menge, B. A. 1978a. Predation intensity in a rocky intertidalcommunity. Effect of an algal canopy, wave action anddesiccation on predator feeding rates. Oecologia (Berlin)34:17–35.
Menge, B. A. 1978b. Predation intensity in a rocky intertidalcommunity. Relation between predator foraging activityand environmental harshness. Oecologia (Berlin) 34:1–16.
Menge, B. A. 1992. Community regulation: under what con-ditions are bottom-up factors important on rocky shores?Ecology 73:755–765.
Menge, B. A., E. L. Berlow, C. A. Blanchette, S. A. Navarrete,and S. B. Yamada. 1994. The keystone species concept:variation in interaction strength in a rocky intertidal habitat.Ecological Monographs 64:249–286.
Menge, B. A., B. A. Daley, J. Lubchenco, E. Sanford, E.Dahlhoff, P. M. Halpin, G. Hudson, and J. L. Burnaford.1999. Top-down and bottom-up regulation of New Zealandrocky intertidal communities. Ecological Monographs 69:297–330.
Menge, B. A., B. A. Daley, and P. A. Wheeler. 1996. Controlof interaction strength in marine benthic communities. Pag-es 258–274 in G. A. Polis and K. O. Winemiller, editors.Food webs: integration of pattern and dynamics. Chapmanand Hall, New York, New York, USA.
Menge, B. A., B. A. Daley, P. A. Wheeler, E. Dahlhoff, E.Sanford, and P. T. Strub. 1997a. Benthic-pelagic links and

October 2001 2829EFFECTS OF STRESS ON WHELK ECOPHYSIOLOGY
rocky intertidal communities: bottom-up effects on top-down control? Proceedings of the National Academy ofSciences, USA 94:14530–14535.
Menge, B. A., B. A. Daley, P. A. Wheeler, and P. T. Strub.1997b. Rocky intertidal oceanography: an association be-tween community structure and nearshore phytoplanktonconcentration. Limnology and Oceanography 42:57–66.
Menge, B. A., and T. M. Farrell. 1989. Community structureand interaction webs in shallow marine hard-bottom com-munities: tests of an environmental stress model. Advancesin Ecological Research 19:189–262.
Menge, B. A., and A. M. Olson. 1990. Role of scale andenvironmental factors in regulation of community struc-ture. Trends in Ecology and Evolution 5:52–57.
Menge, B. A., and J. P. Sutherland. 1976. Species diversitygradients: synthesis of the roles of predation, competition,and temporal heterogeneity. American Naturalist 110:351–369.
Menge, B. A., and J. P. Sutherland. 1987. Community reg-ulation: variation in disturbance, competition, and preda-tion in relation to environmental stress and recruitment.American Naturalist 130:730–757.
Mooney, H. A., R. W. Pearcy, and J. Ehleringer. 1987. Plantphysiological ecology today. Bioscience 37:18–20.
Navarrete, S. A. 1996. Variable predation: effects of whelkson a mid-intertidal successional community. EcologicalMonographs 66:301–321.
Navarrete, S. A., and B. A. Menge. 1996. Keystone predationand interaction strength: interactive effects of predators ontheir main prey. Ecological Monographs 66:409–429.
Navarrete, S. A., B. A. Menge, and B. A. Daley. 2000. Spe-cies interactions in a rocky intertidal food web: prey orpredation regulation of intermediate predators? Ecology 81:2264–2277.
Navarro, J. M., and J. E. Winter. 1982. Ingestion rate, assim-ilation frequency and energy balance in Mytilus chilensisin relation to body size and different algal concentrations.Marine Biology 67:255–266.
Newell, R. C. 1979. Biology of intertidal animals. MarineEcological Surveys, Kent, UK.
Paine, R. T. 1966. Food web complexity and species diver-sity. American Naturalist 100:65–75.
Paine, R. T. 1974. Intertidal community structure: experi-mental studies on the relationship between a dominant com-petitor and its principal predator. Oecologia (Berlin) 15:93–120.
Paine, R. T. 1977. Controlled manipulations in the marineintertidal zone, and their contribution to ecological theory.Pages 245–270 in Changing scenes in natural sciences,1776–1976. Special Publication 12, Academy of NaturalSciences, Philadelphia, Pennsylvania, USA.
Paine, R. T. 1984. Ecological determinism in the competitionfor space. Ecology 65:1339–1348.
Palmer, A. R., S. D. Gayron, and D. S. Woodruff. 1990.Reproductive, morphological, and genetic evidence for twocryptic species of Northeastern Pacific Nucella. Veliger 33:325–338.
Parsell, D. A., and S. Lindquist. 1993. The function of heat-shock proteins in stress tolerance: degradation and reacti-vation of damaged proteins. Annual Review of Genetics27:437–496.
Parsell, D. A., J. Taulien, and S. Lindquist. 1993. The roleof heat-shock proteins in thermotolerance. PhilosophicalTranscripts of the Royal Society of London B, BiologicalScience 339:279–285.
Prosser, C. L. 1986. Adaptational biology: molecules to or-ganisms. John Wiley, New York, New York, USA.
Roberts, D. A., G. E. Hofmann, and G. N. Somero. 1997.Heat-shock protein expression in Mytilus californianus: ac-climatization (seasonal and tidal-height comparisons) andacclimation effects. Biological Bulletin 192:30–320.
Sanford, E. 1999a. Oceanographic influences on rocky in-tertidal communities: coastal upwelling, invertebrategrowth rates, and keystone predation. Dissertation. De-partment of Zoology, Oregon State University, Corvallis,Oregon.
Sanford, E. 1999b. Regulation of keystone predation by smallchanges in ocean temperature. Science 283:2095–2097.
Sanford, E., and B. A. Menge. 2001. Spatial and temporalvariation in barnacle growth in a coastal upwelling system.Marine Ecology Progress Series 209:143–157.
Stickle, W. B., and B. L. Bayne. 1987. Energetics of themuricid gastropod Thais (Nucella) lapillus L. Journal ofExperimental Marine Biology and Ecology 107:263–278.
Stickle, W. B., M. N. Moore, and B. L. Bayne. 1985. Effectsof temperature, salinity, predation and lysosomal stabilityof the dogwhelk Thais (Nucella) lapillus L. Journal of Ex-perimental Marine Biology and Ecology 93:235–258.
Stillman, J. H., E. P. Dahlhoff, and G. N. Somero. 1996.Biochemical indicators of physiological state in the inter-tidal mussel Mytilus californianus. Physiologist 37:921.
Stillman, J. H., and G. N. Somero. 1996. Adaptation to tem-perature stress and aerial exposure in congeneric speciesof intertidal porcelain crabs (genus Petrolisthes): correla-tion of physiology, biochemistry and morphology with ver-tical distribution. Journal of Experimental Biology 199:1845–1855.
Tomanek, L., and G. N. Somero. 1999. Evolutionary andacclimation-induced variation in the heat-shock responsesof congeneric marine snails (genus Tegula) from differentthermal habitats: implications for limits of thermotoleranceand biogeography. Journal of Experimental Biology 202:2925–2936.
Trussell, G. C. 1997. Phenotypic plasticity in the foot sizeof an intertidal snail. Ecology 78:1033–1048.
Underwood, A. J., and P. Jernakoff. 1984. The effects of tidalheight, wave-exposure, seasonality and rock-pools on graz-ing and the distribution of intertidal macroalgae in NewSouth Wales. Journal of Experimental Marine Biology andEcology 75:71–96.
van Erkon Schurink, C., and C. L. Griffiths. 1992. Physio-logical energetics of four South African mussel species inrelation to body size, ration, and temperature. ComparativeBiochemistry and Physiology 101A:779–789.
Wilkinson, L. 1996. SYSTAT 7.0 for Windows. SPSS, Chi-cago, Illinois, USA.
Winer, B. J., D. R. Brown, and K. M. Michels. 1991. Sta-tistical principles in experimental design. McGraw-Hill,New York, New York, USA.
Wolcott, T. G. 1973. Physiological ecology and intertidalzonation in limpets (Acmaea): a critical look at ‘limitingfactors.’ Biological Bulletin 145:389–422.
Wright, D. A., and E. W. Hetzel. 1985. Use of RNA : DNAratios as an indicator of nutritional stress in the Americanoyster Crassostrea virginica. Marine Ecology Progress Se-ries 25:199–206.
Yang, T. H., and G. N. Somero. 1993. Effects of feeding andfood deprivation on oxygen consumption, muscle proteinconcentration, and activities of energy metabolism enzymesin muscle and brain of shallow- (Scorpaena guttata) anddeep- (Sebastelobus alascanus) living Scorpaenid fishes.Journal of Experimental Biology 181:213–223.