photosynthese ii „lichtreaktionen“ · 22.05.2016 2 die photosynthese besteht aus zwei...
TRANSCRIPT

22.05.2016
1
Vorlesung „Zellbiologie Physiologie und Genetik“
SoSe 2016
Photosynthese II„Lichtreaktionen“
Prof. Dr. Iris Finkemeier
Institut für Biologie und Biotechnologie der Pflanzen
Photosynthese II
• Photosynthese Überblick
• Linearer Elektronentransport
• Zyklischer Elektonentransport und Wasser-Wasser Zyklus
• Bakterienphotosynthese
• ATP Synthese (Photo-Phosphorylierung)
• Mechanismen der Photosynthese zur Akklimatisierung an Licht

22.05.2016
2
Die Photosynthese besteht aus zweimiteinander verbundenen Reaktionen
Adapted from Kramer, D.M., and Evans, J. R. (2010). The importance of energy balance in improving photosynthetic productivity. Plant Physiol.155: 70–78.
Chloroplast2 H2O O2 + 2 H+ + 2 ADPH+
ATP
Die Lichtreaktionen in der Thylakoidmembran
Die Kohlenstofffixierungim Stroma
2 NADP+
2 NADPH
2 H+e−
e−
Hill, R. (1937): Oxygen evolution by isolated chloroplasts. In: Nature. Bd. 139, S. 881–882.
Sir Robert Hill

22.05.2016
3
„Linearer“ Elektronentransport dient der Erzeugung von Reduktionskraft (NADPH)
und eines Protonengradienten, der die ATP-Synthese antreibt
Buchanan et al., „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
2 H2Oe-
4 H+O2
Photosystem II
e−
e−
e−
e−
Cytochromeb6f complex
Photosystem I
Plastoquinone
Plastocyanin
Ferredoxin
H2O PSII PQ Cyt b6f PC PSI Fd NADP+ NADPH
Reprinted by permission from Baniulis, D., Yamashita, E., Zhang, H., Hasan, S.S. and Cramer, W.A. (2008). Structure–Function of the Cytochrome b6f Complex†. Photochem. Photobiol. 84: 1349-1358.
Photosynthetischer linearer ElektronentransportOxidation von Wasser und Reduktion von NADPH

22.05.2016
4
• Eigentliche endotherme Redoxreaktion, die durch Sonnenenergie ermöglicht wird.
• Elektronentransport über mehrere Komplexe und Redox-Carrier• Elektronen werden am OEC („oxygen-evolving complex“) vom PSII aus
Wasser „extrahiert)
Photosynthetischer linearer ElektronentransportOxidation von Wasser und Reduktion von NADPH
Der Emerson-Effekt (oder Enhancement-Effekt)
Emerson, Robert (1957). "Dependence of yield of photosynthesis in long wave red on wavelength and intensity of supplementary light". Science 125: 746.

22.05.2016
5
Der Cyt b6f -Komplex vermittelt beim Elektronentransport zwischen PSII und PSI
Damit fungiert er gleichzeitig als Protonenpumpe ( ATP-Synthese)
In Pflanzen und Cyanobakterien sind zwei Photosysteme(PSII und PSI) in Serie geschaltet
PSII entzieht dem Wasser Elektronen (e-)
PSI überträgt sie letztendlich auf NADP+ NADPH + H+
Abb. aus: Buchanan et al. „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
Fdx = FerredoxinFNR = Fdx-NADP-Reduktase
Photosynthese II
• Photosynthese Überblick
• Linearer Elektronentransport
• Zyklischer Elektonentransport und Wasser-Wasser Zyklus
• ATP Synthese (Photo-Phosphorylierung)
• Mechanismen der Photosynthese zur Akklimatisierung an Licht

22.05.2016
6
2 mobile redox-Carrier: - PQ (Plastochinon): PSII Cyt b6f (in der Lipidphase) - PC (Plastocyanin): Cyt b6f PSI (im Thylakoid-Lumen)
Cytochrom b6f-Komplex: - vermittelt Elektronentransport (PSIIPQ und PCPSI) - fungiert als Protonenpumpe (StromaLumen)- homolog zum Cytochrom bc-Komplex der Mitochondrien (Atmungskette)
Photosynthetischer Elektronentransport (PET)
FNR = Ferredoxin‐NADP‐Reduktase
Abb. aus: Buchanan et al. „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
Fdx = FerredoxinFNR = Fdx-NADP-Reduktase
Q Zyklus und Cytochrom b6f Komplex
Stroma(electro-
negative side)
Lumen(electro-
positive side)
PSII Reaction Center
Cytochromb6f complex
PSI Reaction Center
Mobiles PQH2
transportiert Elektronenzum Cyt b6f
Reprinted by permission from Baniulis, D., Yamashita, E., Zhang, H., Hasan, S.S. and Cramer, W.A. (2008). Structure–Function of the Cytochrome b6f Complex†. Photochem. Photobiol 84: 1349-1358.

22.05.2016
7
Cytochrom b6f Komplex
Rieske JS, Maclennan DH, Coleman, R (1964). "Isolation and properties of an iron-protein from the(reduced coenzyme Q)-cytochrome C reductase complex of the respiratory chain". Biochem. Biophys. Res. Commun. 15 (4): 338–344
Q-Zyklus am Cytochrom b6f-Komplex
Bilanz: 1 PQH2 + 2 PCox + 4 H+(Stroma) 1 PQ + 2 PCred + 4 H+
(Lumen)Bilanz: 1 PQH2 + 2 PCox + 4 H+(Stroma) 1 PQ + 2 PCred + 4 H+
(Lumen)
Abb. aus: Buchanan et al. „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
2 e- 1 e-
Übergang2 e- 1 e-
Übergang
Die Reduktion von Plastochinon (PQ, 1) zu Plastochinol (PQH2, 3) erfolgt über das Semichinon-Radikal (PQH, 2).
22.05.2016
8
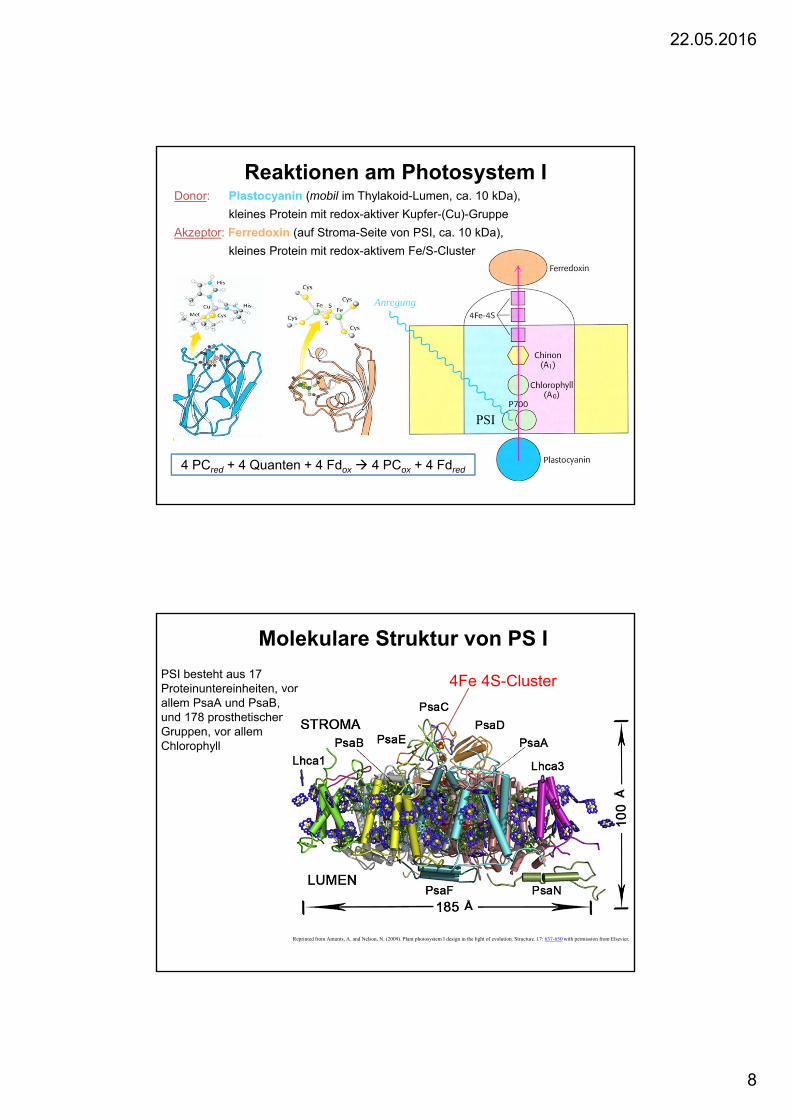
Reaktionen am Photosystem IDonor: Plastocyanin (mobil im Thylakoid-Lumen, ca. 10 kDa),
kleines Protein mit redox-aktiver Kupfer-(Cu)-Gruppe
Akzeptor: Ferredoxin (auf Stroma-Seite von PSI, ca. 10 kDa),
kleines Protein mit redox-aktivem Fe/S-Cluster
4 PCred + 4 Quanten + 4 Fdox 4 PCox + 4 Fdred
PSI
Anregung
Molekulare Struktur von PS I
PSI besteht aus 17 Proteinuntereinheiten, vorallem PsaA und PsaB, und 178 prosthetischenGruppen, vor allemChlorophyll
Å
Å
Reprinted from Amunts, A. and Nelson, N. (2009). Plant photosystem I design in the light of evolution. Structure. 17: 637-650 with permission from Elsevier.
4Fe 4S-Cluster
22.05.2016
9
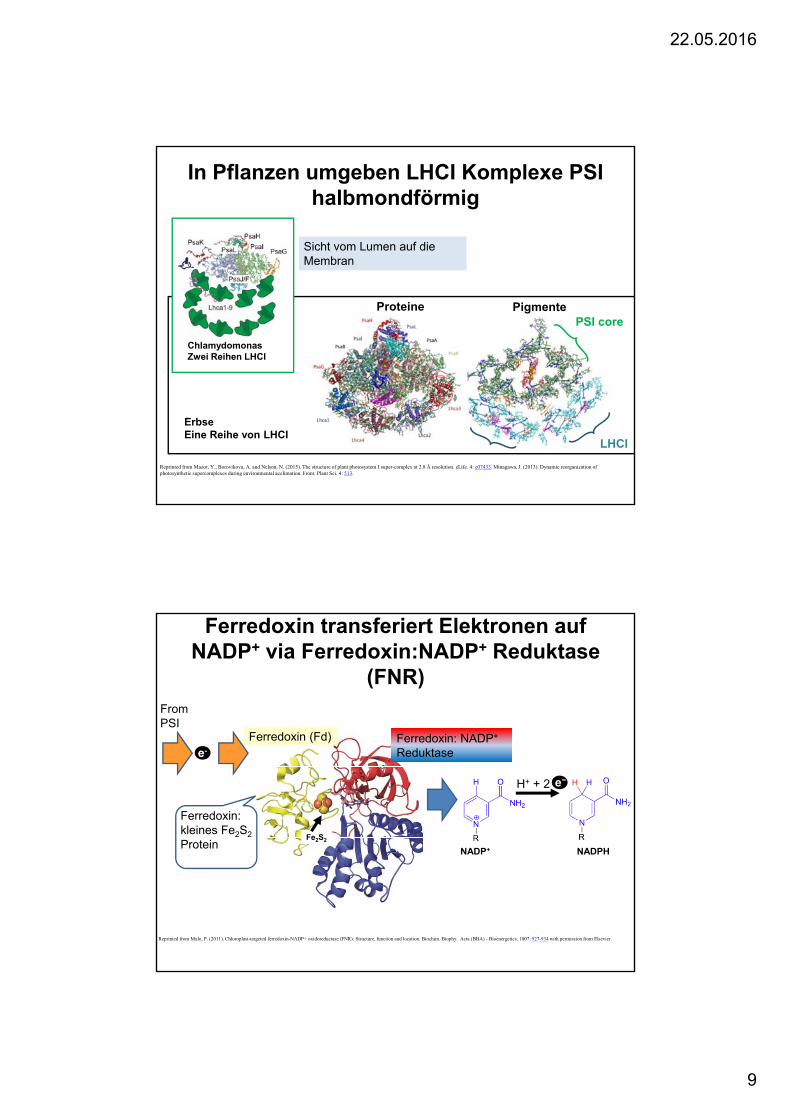
In Pflanzen umgeben LHCI Komplexe PSI halbmondförmig
Reprinted from Mazor, Y., Borovikova, A. and Nelson, N. (2015). The structure of plant photosystem I super-complex at 2.8 Å resolution. eLife. 4: e07433. Minagawa, J. (2013). Dynamic reorganization of photosynthetic supercomplexes during environmental acclimation. Front. Plant Sci. 4: 513.
PSI core
LHCI
Proteine Pigmente
Sicht vom Lumen auf die Membran
ChlamydomonasZwei Reihen LHCI
ErbseEine Reihe von LHCI
Ferredoxin transferiert Elektronen auf NADP+ via Ferredoxin:NADP+ Reduktase
(FNR)
Reprinted from Mulo, P. (2011). Chloroplast-targeted ferredoxin-NADP+ oxidoreductase (FNR): Structure, function and location. Biochim. Biophy. Acta (BBA) - Bioenergetics. 1807: 927-934 with permission from Elsevier.
From PSI
e-
NADP+ NADPH
e−H+ + 2
Ferredoxin: kleines Fe2S2
Protein
Ferredoxin (Fd) Ferredoxin: NADP+
Reduktase
Fe2S2

22.05.2016
10
Ferredoxin:NADP+ Reduktase (FNR)
Ferredoxin verbindet PET mit verschiedenen Stoffwechselreaktionen
22.05.2016
11
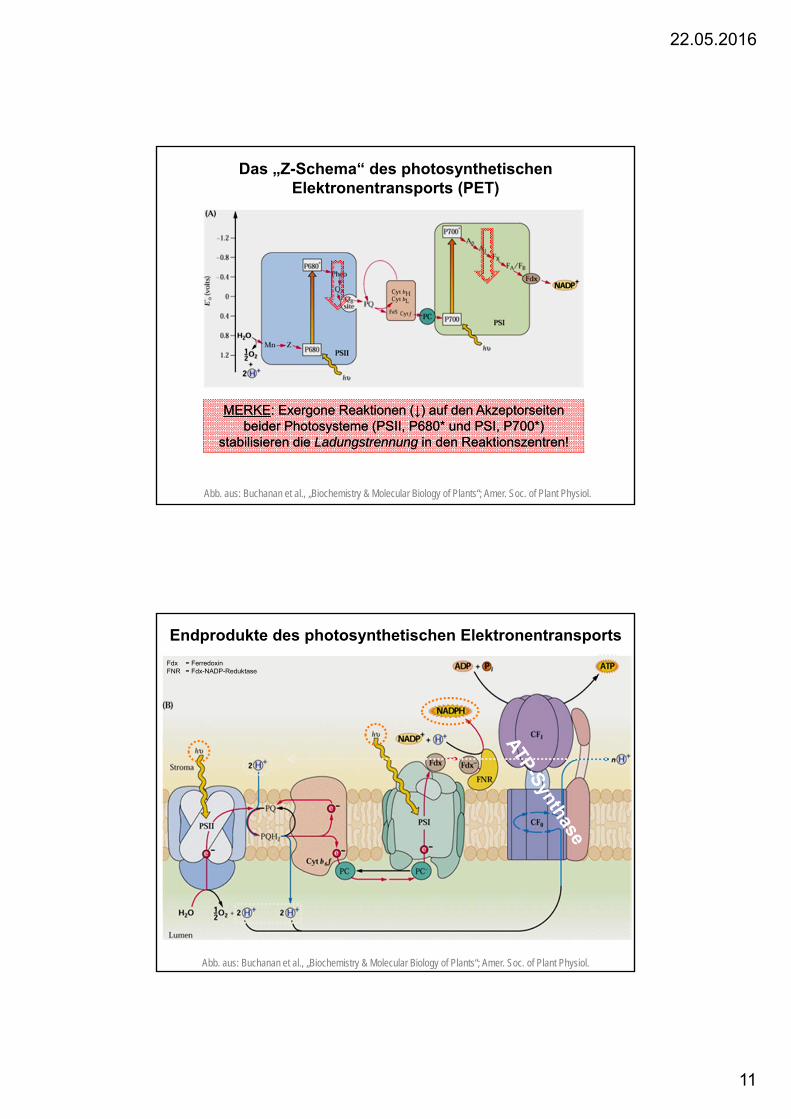
Das „Z-Schema“ des photosynthetischenElektronentransports (PET)
Abb. aus: Buchanan et al., „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
MERKE: Exergone Reaktionen (↓) auf den Akzeptorseitenbeider Photosysteme (PSII, P680* und PSI, P700*)
stabilisieren die Ladungstrennung in den Reaktionszentren!
MERKE: Exergone Reaktionen (↓) auf den Akzeptorseitenbeider Photosysteme (PSII, P680* und PSI, P700*)
stabilisieren die Ladungstrennung in den Reaktionszentren!
Abb. aus: Buchanan et al., „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
Fdx = FerredoxinFNR = Fdx-NADP-Reduktase
Endprodukte des photosynthetischen Elektronentransports

22.05.2016
12
Zusammenfassung
Die Photosysteme PSII und PSI sind in Serie geschaltet (bei Cyanobakterien und Pflanzen).
Photochemische Ladungstrennung am PSII führt zur Spaltung von Wasser.
Die konzertierte Spaltung von 2 Mol H2O wird durch einen Mangan-Komplex vermittelt.
Die dem Wasser entzogenen Elektronen werden auf Plastochinon (PQ); einen mobilen lipophilen Redox-Carrier in der Thylakoidmembran übertragen (2 e- + PQ + 2 H+ PQH2).
Photochemische Ladungstrennung am PSI führt zur Reduktion von NADP+ (NADPH + H+), Elektronendonor ist der mobile Redox-Carrier Plastocyanin (PC) im Lumen der Thylakoide.
Der Cytochrom b6f-Komplex vermittelt bei der Elektronenübertragung zwischen PQ (2 e-) und PC (1 e-) und fungiert gleichzeitig als Protonenpumpe (H+) Q-Zyklus.
Endprodukte des Licht-getriebenen Elektronentransports sind (neben O2 aus der Wasser-spaltung) NADPH und ATP, die den Kohlenstoffreaktionen (z.B. Calvin-Zyklus) im Chloroplasten-Stroma zur Verfügung stehen.
Photosynthese II
• Photosynthese Überblick
• Linearer Elektronentransport
• Zyklischer Elektonentransport und Wasser-Wasser Zyklus
• Bakterienphotosynthese
• ATP Synthese (Photo-Phosphorylierung)
• Mechanismen der Photosynthese zur Akklimatisierung an Licht

22.05.2016
13
Drei Wege des photosynthetischenElektronentransports (PET)
Linearer ET (LET) Zyklischer ET (CET) Wasser-Waser Zyklus
• Elektronentransfer von H2O auf NADPH
• PSII, Cyt b6f, und PSI• NADPH und pmf für ATP
Synthese
• Cyt b6f and PSI• Generiert pmf für ATP
synthese, kein NADPH
H2O
NADPHH+
H+
H+
H+
H2O
H+
H+
H2O
• Elektronentransfer vonH2O to H2O
• PSII, Cyc b6f, and PSI• Generiert pmf für ATP
synthese, kein NADPH
PSI kann ohne PSII arbeitenZyklischer Elektronentransport
dient ausschließlich dem Pumpen von Protonen
P700*
P700+
P700
PSI
Cyt b6f
PQ
PC
Die Elektronentransportkettegeneriert die pmf zur Produktionvon ATP
Zyklischer Elektronentransport: • nur PSI, nicht PSII• Involviert die Elektronentransportkette• zur ATP Produktion• Keine O2 Produktion• Keine NADPH Produktion
Dadurch kann ATP-Mangel ausgeglichen werden.
Der zyklische Elektronentransport ähnelt der bakteriellen Photosynthese!

22.05.2016
14
Beim zyklischen Elektronentransporttransferiert PSI Elektronen auf Cyt b6f
Elektronen werdennicht auf NADP+
transferiert, nur pmfim Lumen aufgebaut.
Das Verhältniszwischen ATP und NADPH wird erhöht.
Wichtig zurVermeidung von Photoinhibition!
Reprinted from Shikanai, T. (2014). Central role of cyclic electron transport around photosystem I in the regulation of photosynthesis. Curr. Opin. Biotechnol. 26: 25-30 by permission from Elsevier..
Reprinted from Shikanai, T. (2014). Central role of cyclic electron transport around photosystem I in the regulation of photosynthesis. Curr. Opin. Biotechnol. 26: 25-30 by permission from Elsevier..
Es gibt mindestens zwei Wege des zyklischen Elektronentransports
(1) PSI Fd PQ Cyt b6f PC PSI
(2) PSI Fd NADP+ PQ Cyt b6f PC PSI
Weg 1 benötigt PGR5 (PROTON GRADIENT
REGULATED5) und PGRL1 (PGR-LIKE1) Weg 2 involviert NAD(P)H
Dehydrogenase (NDH)

22.05.2016
15
1. PGRL1/PGR5-abhängiger zkylischer Elektronentransport
2. NAD(P)H dehydrogenase-abhängigerzyklischer Elektronentransport

22.05.2016
16
Warum zkylischer Elektronentransport?
Stöchiometrie des Ertrags durch den Elektronentransport
Minagawa, J. (2013). Dynamic reorganization of photosynthetic supercomplexes during environmental acclimation. Front. Plant Sci. 4: 513.
1. Bei einer Quanten-Effizienz von ~0.8: für 5 Photonen in PSII gelangen 4 e− in LET
3… und 4 H+
freigesetzt: Oxidation von H2O
4. Die 4 e− von PSII führen zu 8 H+ im Lumen durch den Q-Zyklus
7. Daher werdenwenigstens 2 H+ durchden CEF benötigt um den Calvin-Benson Zyklus zubetreiben
2… 4 e−
reduzieren2 NADP+
+ 2 H+ zuNADPH…
5. Der Calvin-Benson Zyklus benötigt 3 ATP für je 2 NADPH
6. ATP synthase benötigt 14 H+
um 3 ATP zuproduzieren
22.05.2016
17
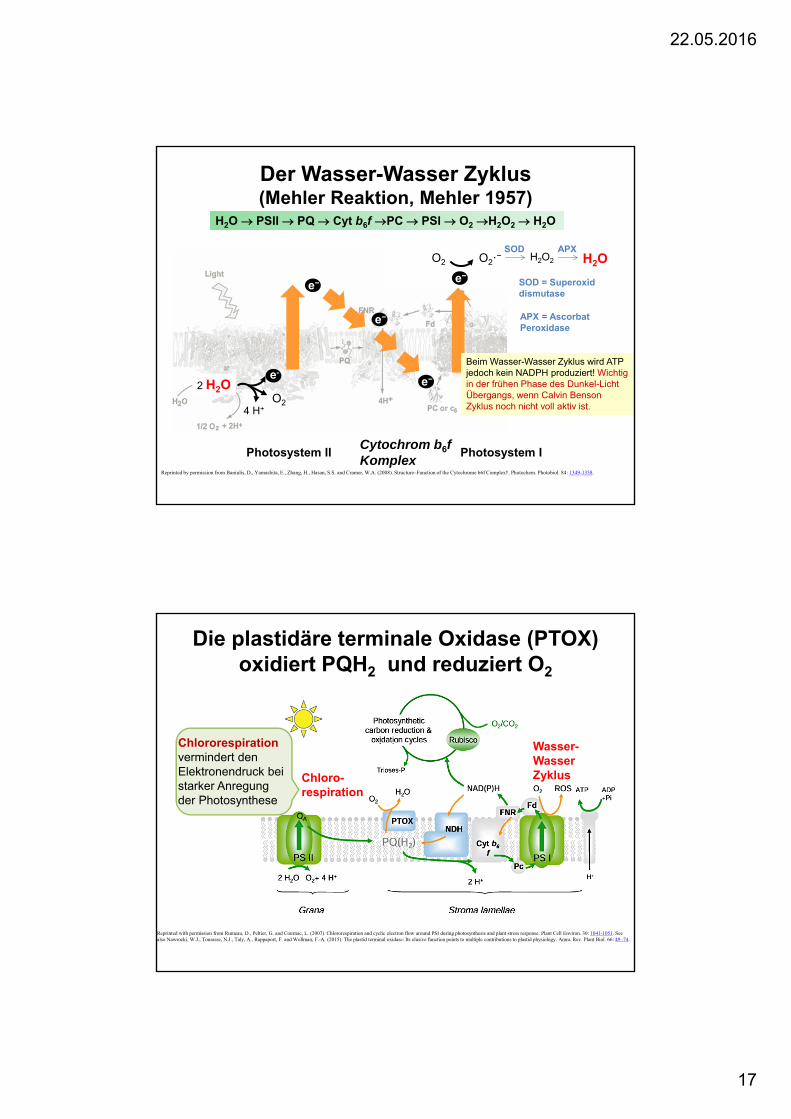
Der Wasser-Wasser Zyklus(Mehler Reaktion, Mehler 1957)
2 H2Oe-
4 H+O2
Photosystem II
e−
e−
e−
Cytochrom b6f Komplex
Photosystem I
e−O2 O2·− H2O2 H2O
SOD APX
SOD = Superoxiddismutase
APX = AscorbatPeroxidase
Beim Wasser-Wasser Zyklus wird ATP jedoch kein NADPH produziert! Wichtigin der frühen Phase des Dunkel-Licht Übergangs, wenn Calvin Benson Zyklus noch nicht voll aktiv ist.
H2O PSII PQ Cyt b6f PC PSI O2 H2O2 H2O
Reprinted by permission from Baniulis, D., Yamashita, E., Zhang, H., Hasan, S.S. and Cramer, W.A. (2008). Structure–Function of the Cytochrome b6f Complex†. Photochem. Photobiol. 84: 1349-1358.
Die plastidäre terminale Oxidase (PTOX) oxidiert PQH2 und reduziert O2
Reprinted with permission from Rumeau, D., Peltier, G. and Cournac, L. (2007). Chlororespiration and cyclic electron flow around PSI during photosynthesis and plant stress response. Plant Cell Environ. 30: 1041-1051. See also Nawrocki, W.J., Tourasse, N.J., Taly, A., Rappaport, F. and Wollman, F.-A. (2015). The plastid terminal oxidase: Its elusive function points to multiple contributions to plastid physiology. Annu. Rev. Plant Biol. 66: 49 -74.
Wasser-Wasser ZyklusChloro-
respiration
Chlororespirationvermindert den Elektronendruck bei starker Anregung der Photosynthese

22.05.2016
18
Zusammenfassung: Wege des photosynthetischen Elektronentransports
Beim LET ist Wasser (H2O) der Elektronendonor und NADP+ der Akzeptor.
Beim CET werden Elektronenvom PSI wieder an PSI übertragen.
Beim Wasser-Wasser Zyklus werden Elektronenvom Wasser auf O2
übertragen und dabei wiederzu Wasser reduziert.
Der Anteil der jeweiligen Wegewird durch den Bedarf des
Metabolismus an ATP und NADPH definiert
RUBISCO
3 GAP
Photosynthese II
• Photosynthese Überblick
• Linearer Elektronentransport
• Zyklischer Elektonentransport und Wasser-Wasser Zyklus
• Bakterienphotosynthese
• ATP Synthese (Photo-Phosphorylierung)
• Mechanismen der Photosynthese zur Akklimatisierung an Licht

22.05.2016
19
1.5 billion years ago
Glaucophytes
Rhodophytes (Red algae)
Chlorophytes (Green algae)
Plants
PlastidPrimary eukaryotic host
Nucleus
Free-livingβ-Cyanobacterium
Mitochondrion
0.06 billion years ago
Free-livingα-Cyanobacterium
Eukaryotic amoeba host
Photosynthetic amoeba Paulinella chromatophora
chromatophores
5 μm
Reprinted by permission from Macmillan Publishers Ltd: from Gould, S.B. (2012). Evolutionary genomics: Algae's complex origins. Nature. 492: 46-48. Reproduced with permission © Annual Reviews Gould, S.B., Waller, R. F., and McFadden, G. (2008). Plastid evolution. Annu. Rev. Plant Biol. 59: 491 – 517; See also Mackiewicz, P., Bodył, A. and Gagat, P. (2012). Possible import routes of proteins into the cyanobacterial endosymbionts/plastids of Paulinella chromatophora. Theory in Biosciences. 131: 1-18.
Beweis für ein weiteres unabhängigesendosymbiotisches Ereignis: Paulinella chromatophora
22.05.2016
20
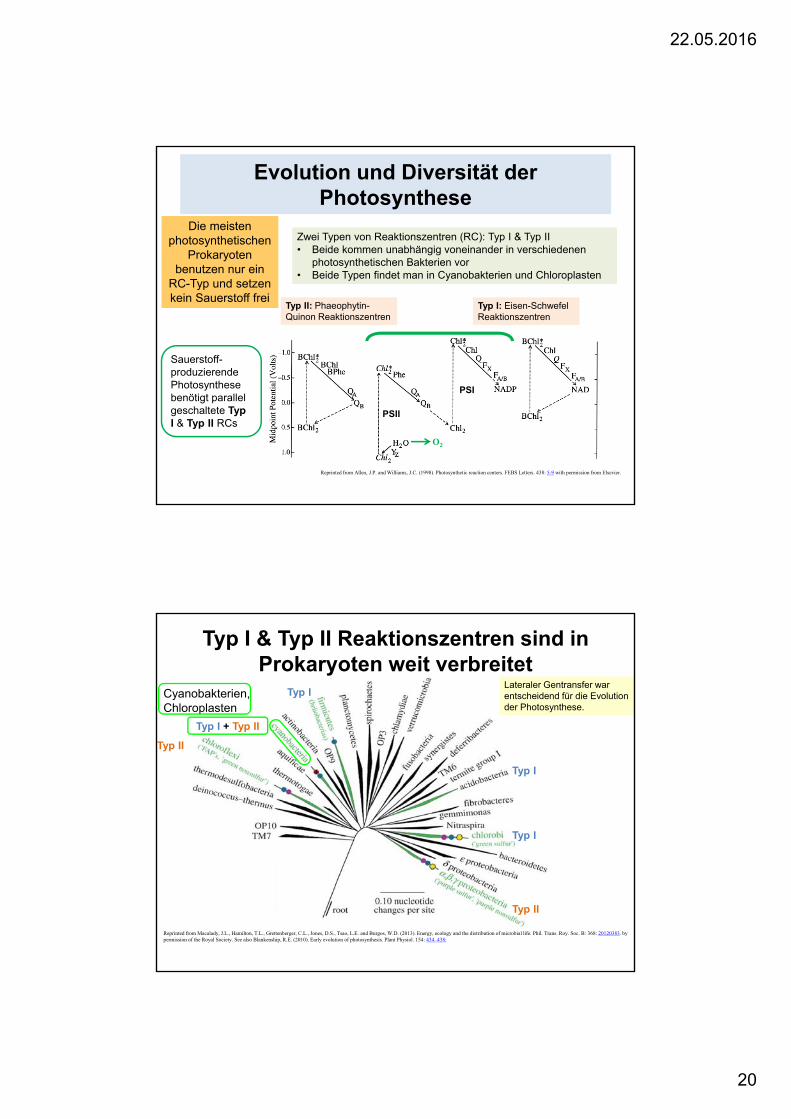
Evolution und Diversität der Photosynthese
Zwei Typen von Reaktionszentren (RC): Typ I & Typ II• Beide kommen unabhängig voneinander in verschiedenen
photosynthetischen Bakterien vor• Beide Typen findet man in Cyanobakterien und Chloroplasten
Typ I: Eisen-SchwefelReaktionszentren
Typ II: Phaeophytin-Quinon Reaktionszentren
PSII
PSI
Sauerstoff-produzierendePhotosynthesebenötigt parallel geschaltete TypI & Typ II RCs
O2
Die meistenphotosynthetischen
Prokaryotenbenutzen nur ein
RC-Typ und setzenkein Sauerstoff frei
Reprinted from Allen, J.P. and Williams, J.C. (1998). Photosynthetic reaction centers. FEBS Letters. 438: 5-9 with permission from Elsevier.
Typ I & Typ II Reaktionszentren sind in Prokaryoten weit verbreitet
Reprinted from Macalady, J.L., Hamilton, T.L., Grettenberger, C.L., Jones, D.S., Tsao, L.E. and Burgos, W.D. (2013). Energy, ecology and the distribution of microbial life. Phil. Trans. Roy. Soc. B: 368: 20120383. by permission of the Royal Society. See also Blankenship, R.E. (2010). Early evolution of photosynthesis. Plant Physiol. 154: 434–438;
Typ I
Typ II
Typ II
Typ I
Typ I
Cyanobakterien, Chloroplasten
Typ I + Typ II
Lateraler Gentransfer war entscheidend für die Evolution der Photosynthese.

22.05.2016
21
Prokaryotische Diversität der Photosynthese
Adapted from Raymond, J. (2008). Coloring in the tree of life. Trends Microbiology. 16: 41-43.
Phylum Entdeckung RC Pigmente
Cyanobakterien 1800s Typ I + Tye II Chl a,b,c,d
Proteobakterienz. B. Purpurbakterien
1800s Typ II BChl a,b
ChlorobiGrüne Schwefelbakterien
1906 Typ I BChl a,c, d, e; Chl a
ChloroflexiFilamentöse anoxische Phototrophe
1974 Typ II BChl a,c
FirmicutesHeliobakterien
1983 Typ I BChl g; Chl a
Acidobakterien 2007 Typ I BChl a,c
Typ I sind die ursprünglchen RCs
With kind permission from Springer Science+Business Media Nelson, N. (2013). Evolution of photosystem I and the control of global enthalpy in an oxidizing world. Photosynth. Res. 116: 145-151.
Model zur Evolution der RCs
Gene duplication: PsA → PsaB
tim
epa
stpr
esen
t
Chlorobi Cyanobacteria
Fe-S cluster as electron acceptor
Ancestral reaction center
ProteobacteriaCyanobacteria Rhodopseudomonas

22.05.2016
22
Bakterien-Photosynthese (außer Cyanobakterien, spalten bereits Wasser)
Photosynthetische Bakterien enthalten:
nur ein Photosystem
einen Cytochrom-bc-Komplex als Protonenpumpe (homolog zum Cytochrom-bf- Komplex der Pflanzen)
eine rotierende ATP-Synthase(homolog zur ATP-Synthase in Thylakoiden und Mitochondrien)
MERKE: Photosynthese hat sich im Anaeroben entwickelt!
Purpurbakterienbetreiben zyklische Photosynthese
Abbildung aus H-W Heldt „Pflanzenbiochemie“

22.05.2016
23
PET und Oxidation von Schwefelwasserstoff (z.B. bei grünen Schwefelbakterien )
Abbildung aus H-W Heldt „Pflanzenbiochemie“
Photosynthese II
• Photosynthese Überblick
• Linearer Elektronentransport
• Zyklischer Elektonentransport und Wasser-Wasser Zyklus
• Bakterienphotosynthese
• ATP Synthese (Photo-Phosphorylierung)
• Mechanismen der Photosynthese zur Akklimatisierung an Licht
22.05.2016
24
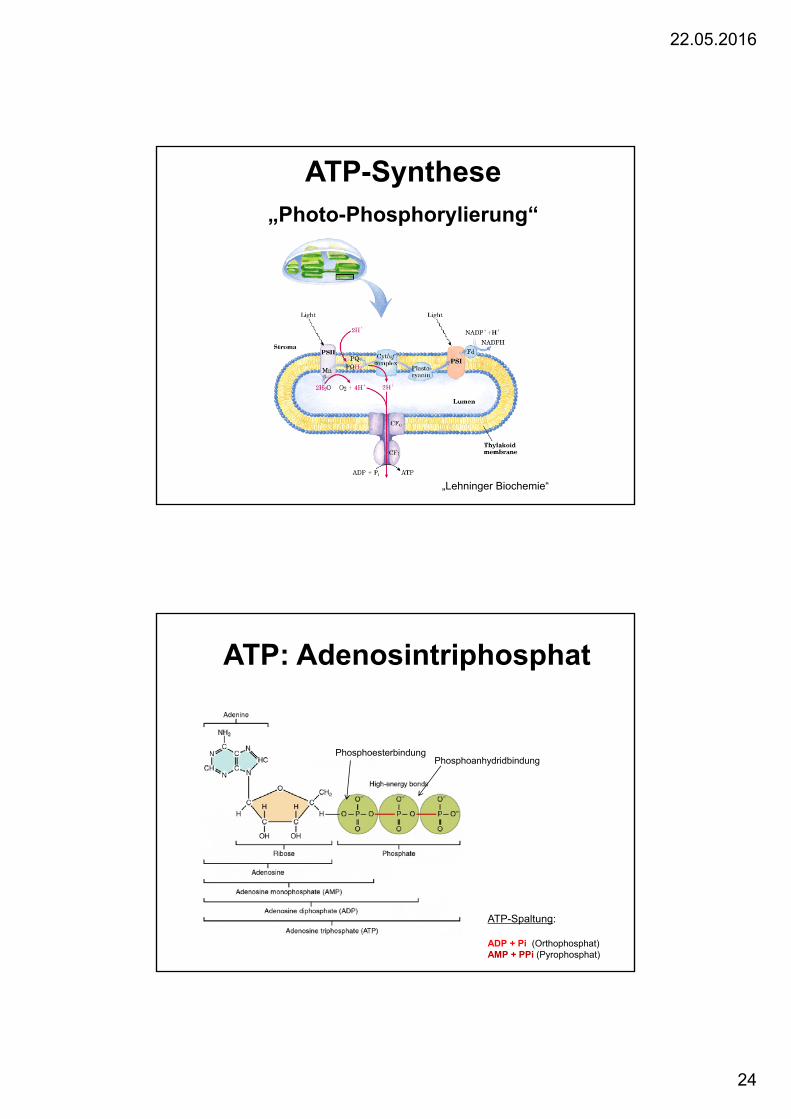
ATP-Synthese „Photo-Phosphorylierung“
„Lehninger Biochemie“
ATP: Adenosintriphosphat
ATP-Spaltung:
ADP + Pi (Orthophosphat)AMP + PPi (Pyrophosphat)
PhosphoesterbindungPhosphoanhydridbindung

22.05.2016
25
Elektrische und H+ Gradienten betreibenATP Synthese
ADP + Pi
PSI
Cyt b6f
2 NADP+
2 NADPH
2 H+ e−
PSII
H+
ATP
[H+]ProtonenGradient von hoch(Lumen)
nach gering(Stroma)
2 H2O e−
4 H+ O2
4
Thylakoid Membran
ATP Synthase
Struktur und Funktion der ATP Synthase
Daum, B., Nicastro, D., Austin, J., McIntosh, J.R. and Kühlbrandt, W. (2010). Arrangement of Photosystem II and ATP synthase in chloroplast membranes of spinach and pea. Plant Cell. 22: 1299-1312.
Thylakoid Lumen
ADP + Pi
H+
ATP
H+
ATP Synthase
H+
H+
H+H+ H+
H+ H+
H+
H+
Die Dissipation des Protonengradienten ist mit ATP Synthese gekoppelt

22.05.2016
26
Plastidäre (CF-ATPase) und mitochondriale (F-ATPase) sind homologe Enzyme und weitgehend identisch.
Plastidäre (CF-ATPase) und mitochondriale (F-ATPase) sind homologe Enzyme und weitgehend identisch.
Die molekulare Struktur der ATP-Synthase ist aufgeklärt.Die molekulare Struktur der ATP-Synthase ist aufgeklärt.
Kopf
Hals
Stator
Membran-anteil
Die Struktur der F- ATPase ist hoch konserviert.
rot Kern-codiert rosa Plastom-codiert
Die Untereinheiten aus Spinat (rot) und E. coli (rosa) können funktionelle Chimären bilden !
Die UE der F- ATPase sind in verschiedenen Genomen kodiert.
22.05.2016
27
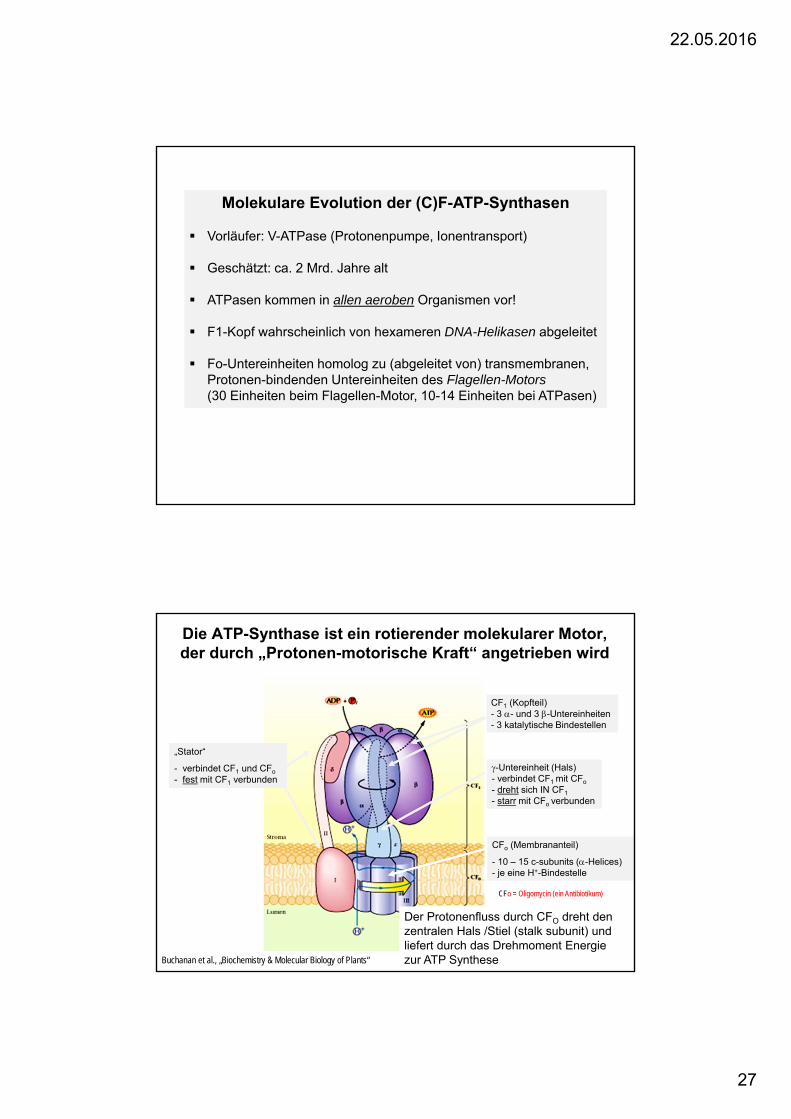
Molekulare Evolution der (C)F-ATP-Synthasen
Vorläufer: V-ATPase (Protonenpumpe, Ionentransport)
Geschätzt: ca. 2 Mrd. Jahre alt
ATPasen kommen in allen aeroben Organismen vor!
F1-Kopf wahrscheinlich von hexameren DNA-Helikasen abgeleitet
Fo-Untereinheiten homolog zu (abgeleitet von) transmembranen, Protonen-bindenden Untereinheiten des Flagellen-Motors(30 Einheiten beim Flagellen-Motor, 10-14 Einheiten bei ATPasen)
Molekulare Evolution der (C)F-ATP-Synthasen
Vorläufer: V-ATPase (Protonenpumpe, Ionentransport)
Geschätzt: ca. 2 Mrd. Jahre alt
ATPasen kommen in allen aeroben Organismen vor!
F1-Kopf wahrscheinlich von hexameren DNA-Helikasen abgeleitet
Fo-Untereinheiten homolog zu (abgeleitet von) transmembranen, Protonen-bindenden Untereinheiten des Flagellen-Motors(30 Einheiten beim Flagellen-Motor, 10-14 Einheiten bei ATPasen)
Die ATP-Synthase ist ein rotierender molekularer Motor, der durch „Protonen-motorische Kraft“ angetrieben wird
„Stator“
- verbindet CF1 und CFo
- fest mit CF1 verbunden
„Stator“
- verbindet CF1 und CFo
- fest mit CF1 verbunden-Untereinheit (Hals) - verbindet CF1 mit CFo
- dreht sich IN CF1
- starr mit CFo verbunden
CFo (Membrananteil)
- 10 – 15 c-subunits (-Helices) - je eine H+-Bindestelle
CFo (Membrananteil)
- 10 – 15 c-subunits (-Helices) - je eine H+-Bindestelle
CFo (Membrananteil)
- 10 – 15 c-subunits (-Helices) - je eine H+-Bindestelle
CF1 (Kopfteil) - 3 - und 3 -Untereinheiten - 3 katalytische Bindestellen
Buchanan et al., „Biochemistry & Molecular Biology of Plants“
CFo = Oligomycin (ein Antibiotikum)
Der Protonenfluss durch CFO dreht den zentralen Hals /Stiel (stalk subunit) und liefert durch das Drehmoment Energiezur ATP Synthese

22.05.2016
28
Summengleichung Photo-Phosphorylierung:
ADP + Pi + pmf ATP + H2O
Die ATP-Synthese wird durch Protonen-motorische Kraft (proton-motive force, pmf) angetrieben, und setzt sich aus dem chemischen Gradienten (pH) und dem elektrischen Potential (y) zusammen:
pmf = mH+ = pH + y
Die pmf wird durch Licht-getriebenen e--Transport aufgebaut (H+ Lumen).
Bei ATP-Synthese fließen 4,67 H+ pro ATP zurück ins Stroma (Abbau der pmf).
Proton motive force (pmf)
Problem: In wässriger Lösung liegt das Gleichgewicht auf Seite der hydrolytischen Spaltung von ATP!
ATP + H2O ADP + Pi
Problem: In wässriger Lösung liegt das Gleichgewicht auf Seite der hydrolytischen Spaltung von ATP!
ATP + H2O ADP + Pi
Buchanan et al., „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
In wässriger Umgebung: ATP + H2O ADP + Pi
In wasserfreier Umgebung: ADP + Pi ATP (+ H2O, gebunden)
In der geschlossenen Bindetasche wird H2O vorübergehend gebunden und damit dem chemischen Gleichgewicht entzogen!
Die ATP-Synthese geht mit zyklischen, pmf-getriebenen Konformationsänderungen der 3 Bindetaschen am CF1 einher. Dabei werden die Bindetaschen vorübergehend „wasserfrei“.
Die ATP-Synthese geht mit zyklischen, pmf-getriebenen Konformationsänderungen der 3 Bindetaschen am CF1 einher. Dabei werden die Bindetaschen vorübergehend „wasserfrei“.
Buchanan et al., „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
F1 binding sites can adopt an „open“ (O), „tight“ (T), or „loose“ (L) conformation (Paul Boyer, 1977)
ATP wird im „Boyer's binding change mechanism” gebildet(Nobelpreis für Chemie 1997, Boyer, Walker, Skou)

22.05.2016
29
Der Protonen-Rückfluss löst eine Drehung von [CFo + g-UE] gegenüber [CF1 + Stator] aus.
Die Drehung der -UE im Kopfteil treibt den katalytischen „binding change cycle“ an.
Pro Drehung (im katalytischen Zyklus) werden 14 Protonen transportiert und 3 ATP gebildet.
Rotation wird durch sukzessive Protonierung (Einbringen positiver Ladung in die 12 -Helices des CFo) ausgelöst.
4 Protonen
Konformations-
änderungen!
Chemiosmotische Kopplung durch Rotation der ATP-Synthase
Buchanan et al., „Biochemistry & Molecular Biology of Plants“; Amer. Soc. of Plant Physiol.
Video-Empfehlung: https://www.youtube.com/watch?v=XI8m6o0gXDY
Zusammenfassung: Struktur und Funktion der Photosynthese Komplexe
ATP Synthase
Die einzelnen Photosynthesekomplexe bestehen aus Dutzenden Proteinen und Hunderten von Pigmenten und Co-Faktoren. Ihre Aufklärung und das Verständnisgehen auf die Forschung von Hunderten von Wissenschaftlern über die letztenJahrzehnte zurück.
Reprinted by permission from Baniulis, D., Yamashita, E., Zhang, H., Hasan, S.S. and Cramer, W.A. (2008). Structure–Function of the Cytochrome b6f Complex. Photochemistry and Photobiology. 84: 1349-1358.

22.05.2016
30
Photosynthese II
• Photosynthese Überblick
• Linearer Elektronentransport
• Zyklischer Elektonentransport
• ATP Synthese (Photo-Phosphorylierung)
• Bakterienphotosynthese
• Mechanismen der Photosynthese zur Akklimatisierung an Licht
Anpassung an Lichtstress
Adapted from Li, Z., Wakao, S., Fischer, B.B. and Niyogi, K.K. (2009). Sensing and responding to excess light. Annu. Rev.Plant Biol. 60: 239-260.
Light intensity
Rat
e o
f p
ho
tosy
nth
esis
Bei geringerLichtintensität:
Lichtintensitätlimitiert
Photosynthese-leistung
Bei hoher Lichtintensität: Zu viel Licht kann die
Photosynthesekomplexe schädigen
Der metabolische und physiologische Status der Pflanze oder Zelledefiniert wieviel Licht
“zu viel” ist
Optimal conditions
Stressed conditions

22.05.2016
31
Zu viel Anregungsenergie kann zu photo-oxidativen Schäden führen
Reprinted by permission from Macmillan Publishers Ltd Demmig-Adams, B. and Adams, W.W. (2000). Photosynthesis: Harvesting sunlight safely. Nature. 403: 371-374.
Photonen-angeregtes Chlorophyll bildet angeregtes SingulettChlorophyll 1Chl*1Chl* kehrt zum Grundzustandzurück durch:
PhotochemieFluoreszenzDissipation
(z.B., Hitze, NPQ)
Alternativ, wird 1Chl* zumangeregten Triplett Chlorophyll 3Chl* umgewandelt, welches seine Energie auf Sauerstoff transferiert. Dabei entsteht Singulett-Sauerstoff, der Zellschädenverursachen kann.
P
F
D
NPQ
Lichtreaktionen
Kohlenstofffixierung
ATP NADPHADP NADP+
Hohe Temp.Erhöhte
Membranpermeabilitätund verringerte pmf
TrockenheitinduziertStomataschluss und verhindert CO2
Aufnahme
KälteverringertGeschwindigkeitvon Enzymreaktionen
Nährstoffmangeloder ÜberschussbeeinflusstElektronentransport
Hitze, Trockenheit, Temperatur & andereFaktoren beeinflussen die Effizienz der
Photosynthese

22.05.2016
32
Photoinhibition (qI) verursacht vor allemSchaden an PSII
Reprinted by permission from Takahashi, S. and Murata, N. (2008). How do environmental stresses accelerate photoinhibition? Trends Plant Sci 178-182.
Das D1 Protein von PSII wirdleicht geschädigt. Das D1 Protein wird daher sehrschnell neusynthetisiert und ausgetauscht. Wenn die Syntheserate mitder Schädigungnicht mithält, trittPhotoinhibitionein.
1. Kurzfristige Strategien zur Vermeidungvon Schäden durch Starklicht
Li, Z., Ahn, T.K., Avenson, T.J., Ballottari, M., Cruz, J.A., Kramer, D.M., Bassi, R., Fleming, G.R., Keasling, J.D. and Niyogi, K.K. (2009). Lutein accumulation in the absence of zeaxanthin restores nonphotochemical quenching in the Arabidopsis thaliana npq1 mutant. Plant Cell. 21: 1798-1812. Havaux, M. and Niyogi, K.K. (1999). The violaxanthin cycle protects plants from photooxidative damage by more than one mechanism. Proc. Natl. Acad. Sci. USA. 96: 8762-8767; Adapted from Li, Z., Wakao, S., Fischer, B.B. and Niyogi, K.K. (2009). Sensing and responding to excess light. Annu. Rev.Plant Biol. 60: 239-260
Dynamische Anpassung der Antennenkomplexe
Reaction center
Antenna complex
Abgabe der Energie als Hitzeoder Fluoreszenz
Quenching von Triplettchlorophyllund Entgiftung von reaktivenSauerstoffspezies
lutein
Npq1 Pflanzen haben einen Defekt beider Disspation von Strahlungsenergie
und bleichen bei Starklicht aus.
zeaxanthin

22.05.2016
33
Oikawa, K., Kasahara, M., Kiyosue, T., Kagawa, T., Suetsugu, N., Takahashi, F., Kanegae, T., Niwa, Y., Kadota, A. and Wada, M. (2003). CHLOROPLAST UNUSUAL POSITIONING1 is essential for proper chloroplast positioning. Plant Cell. 15: 2805-2815. Donald Hoburn; Walter Siegmund; Kristian Peters
Sch
wac
hlic
ht
Sta
rklic
ht
Top view Side view
Chloroplastenwandern an die Seite der Zellen
Selaginella lepidophylla
Änderung des Blattwinkels oderEinrollen von Blättern.
Arctostaphylos patula
Eucalyptus rossii
2. Mittelfristige Strategien zur Vermeidungvon Schäden durch Starklicht
Chloroplastenbewegung

22.05.2016
34
Akklimatisierung durch Veränderung in der Stöchiometrie der Photosynthesekomplexe
Walters, R.G. (2005). Towards an understanding of photosynthetic acclimation. J. Exp. Bot. 56: 435-447.
Schwachlicht: Mehr Antennen(LHCII)
Starklicht: GrößerephotosynthetischeKapazität (mehrPSII etc)
Arabidopsis im Schwachlicht (schwarze Balken) oderStarklicht (weiße Balken).
3. Langfristige Strategien zur Vermeidungvon Schäden durch Starklicht
Dissipation von zu viel Licht via “non-photochemical quenching” NPQ
“Non-photochemical quenching” Setzt sich zusammen aus:
qE = Energie-abhängiges Quenching: Xanthophyll-ZyklusqT = State transition: Bewegung von LHCIIqI = Photoinhibition: Verlust der Photosyntheseleistungdurch Licht-induzierte Schädigung
Chl
1Chl*
Photon Non-photochemical quenching (NPQ)

22.05.2016
35
Energie-abhängiges quenching (qE) macht den Haupteil von NPQ aus
PSII
[H+]
LHC PSIILHC
Normallicht: HoheEffizienz der Photosynthesebei Ausnutzungder vollenLichtenergie
[H+]
[H+]
[H+][H+]
VDEStarklicht:Energie-abhängigesquenching 1. Ansäuerung des Lumens aktiviert dieViolaxanthin De-epoxidase (VDE)
2. VDE wandeltViolaxanthin in Zeaxanthin um. Absorption des Lichts durch Zeaxanthin führt zurDissipation der Lichtenergie
Xanthophyll Zyklus: reversible Umwandlung von Carotenoiden
Violaxanthin de-epoxidase
(VDE)
Zeaxanthin epoxidase
(ZE)
Starklicht/ geringer luminalerpH induziert VDE, welche die Umwandlung von Violaxanthinzu Zeaxanthin katalysiert.Bei Schwachlicht/ hohemluminalen pH, wird die Reaktion durch ZE revertiert.
Hieber, A.D., Kawabata, O. and Yamamoto, H.Y. (2004). Significance of the lipid phase in the dynamics and functions of the xanthophyll cycle as revealed by PsbS overexpression in tobacco and in-vitro de-epoxidation in monogalactosyldiacylglycerol micelles. Plant Cell Physiol 45: 92-102 by permission of Oxford University Press.

22.05.2016
36
Zeaxanthin fördert strukturelle Änderungenund Hitze-Dissipation im Photosystem
Reprinted with permission from Ruban, A.V. (2015). Evolution under the sun: optimizing light harvesting in Photosynthesis. J. Exp. Bot. 66: 7 – 23; Müller-Moulé, P., Conklin, P.L. and Niyogi, K.K. (2002). Ascorbate Deficiency Can Limit Violaxanthin De-Epoxidase Activity in Vivo. Plant Physiol. 128: 970-977.
Die Akkumulation von Zeaxanthin (durch VDE Aktivierung) führt zurUmlagerung von LHCII und RCII. Vermindert Lichttransfer an RCII
Die strukturellen Änderungenführen dazu, dass mehrEnergie als Hitze abgestrahltwird.
Energie-abhängiges quenching (qE) macht den Haupteil von NPQ aus
Neben dem Xantophyll-Zyklus wird auch das PSBS-Protein durch Protonierung aktiviert und hilft bei der Reorganisation von PSII.
Buchanan et al., „Biochemistry & Molecular Biology of Plants“

22.05.2016
37
Zeaxanthin und Lutein sind wichtigeAntioxidantien zum Schutz vor Lichtschäden
Niyogi, K.K., Björkman, O. and Grossman, A.R. (1997). The roles of specific xanthophylls in photoprotection. Proc. Natl. Acad. Sci. USA 94: 14162-14167.
Mangel an Zeaxanthin und Lutein beiChlamydomonas führt zum Ausbleichen
Akkumulation von Zeaxanthin und Lutein (aus dem Essen) in der Makula(orange Farbe), schützen auch das menschliche Auge vor photooxidativenSchäden.
Asymmetrische Verteilung
der Photosynthese-Komplexe
in der Thylakoidmembran

22.05.2016
38
Die Menge an CURT1 Protein bestimmt das Ausmaß der Ausbildung von Granastapeln.
Armbruster et al. (2013) Arabidopsis CURVATURE THYLAKOID1 proteins modify thylakoid architectureby inducing membrane curvature. Plant Cell

22.05.2016
39
CURT1 Oligomere stabilisieren die Grana-Ränder (margins). Protein-Phosphorylierung spielt auch eine große Rolle bei der Ausbildung von
Granastapeln.Pribil M, Labs M, Leister D. 2014 Structure and dynamics of thylakoids in land plants. J Exp Bot.
Die Membranen der Chloroplastensind ungewöhnlich
Lipid-Zusammensetzung
Nur 10-15% Phospholipide
75% Galactolipide
Stark ungesättigt
O
H
HH
OH
H
OH
H OH
O
OH
O
O
O
R
O
R
galactose
linolenoyl
Sehr flüssig
Kleine Kopfgruppe
Keine Ladung

22.05.2016
40
Grana- und Stromathylakoide habenverschiedene Funktionen
Daum, B., et al., (2010). Arrangement of Photosystem II and ATP synthase in chloroplast membranes of spinach and pea. Plant Cell. 22: 1299-1312; Nagy, G., et al. and Minagawa, J. (2014). Chloroplast remodeling during state transitions in Chlamydomonas reinhardtii as revealed by noninvasive techniques in vivo. Proc. Natl. Acad. Sci. USA. 111: 5042-5047.
Lumen
Grana
PSII gepackt zwischen zwei Membranen
State 1
ATP synthase
PSI-LHCI supercomplex
Cyt b6f
LHCIICyt b6f
PSII
PSII findet man bevorzugt in Granathylakoiden, ATP synthase and PSIin StromalamellenCyt b6f überall in der Thylakoide
Wie entsteht laterale Heterogenität der Proteinkomplexe?
- Sterische Hinderung ATP Synthase
- Anziehung durch Lichtsammelkomplexe in den Granamembranen
Pribil, M., Labs, M., and Leister, D. (2014). Structure and dynamics of thylakoids in land plants. J. Exp. Bot.

22.05.2016
41
Die Kernbereiche der Grana-Stapel sind Orte der Wasserspaltung
Von den Grana-Randbereichen („margins“) werden im Licht ATP und NADPH in das Stroma entlassen und zur CO2-Fixierung genutzt
Stroma-Lamellen sind Orte von Biosynthese, Abbau und „Reparatur“ (Protein turn-over)
NADPH
ATP
Abbildung verändert
State transitions: LHCII wandert von Granastapeln zu Stromalamellen nach Belichtung
mit PSII Licht (650 nm)
Ein reduzierter PQH2 Pool aktiviert die Kinase STN7 LHCII wird phosphoryliert. Oxidation aktiviert die Phosphatase TAP38.
State 1: Lhcb2 (dephosphoryliert) vermehrt an PSII
State 2: Lhb2 (phosphoryliert) vermehrt an PSI

22.05.2016
42
Der Redoxstatus des PQ Pools reguliertstate transitions (qT)
Rosso, D., Bode, R., Li, W., Krol, M., Saccon, D., Wang, S., Schillaci, L.A., Rodermel, S.R., Maxwell, D.P. and Hüner, N.P.A. (2009). Photosynthetic redox imbalance governs leaf sectoring in the Arabidopsis thaliana variegation mutants immutans, spotty, var1, and var2. Plant Cell. 21: 3473-3492.
Wenn PSII und PSI imGleichgewicht sind, dann istauch der PQ Pool imGleichgewicht zwischen PQ (oxidiert) und PQH2 (reduziert)
Starklicht (oder PSI Licht) oder Inhibition des Metabolismus führen zueiner Überreduktion des PQ pools
LHCII Phosphorylierung und Migration

22.05.2016
43
Langzeitakklimatisierung an PSII Licht: vermehrte Bildung von PSI (PsaA/B)
Retrograde Signale vom Plastiden an den Zellkern sindentscheidend für die Anpassung der Photosynthese durch
Steuerung der nukleären Genexpression
Reprinted with permission from Macmillan Publishing Ltd from Jarvis, P. and López-Juez, E. (2013). Biogenesis and homeostasis of chloroplasts and other plastids. Nat Rev Mol Cell Biol. 14: 787-802; see also Dietz, K.-J. (2015). Efficient high light acclimation involves rapid processes at multiple mechanistic levels. J. Exp. Bot. 66: 2401-2414l Chi, W., Sun, X. and Zhang, L. (2013). Intracellular signaling from plastid to nucleus. Annu. Rev. Plant Biol. 64: 559-582; Estavillo, G.M., Chan, K.X., Phua, S.Y. and Pogson, B.J. (2013). Reconsidering the nature and mode of action of metabolite retrograde signals from the chloroplast. Frontiers Plant Sci. 3: 300.
Es gibt biogene(Entwicklung) und operative (Funktion)retrograde Signale
Verschiedenste Moleküle fungierenals retrograde Signale, z. B. Produkte
der Hämbiosynthese und reaktiveSauerstoffspezies und andere
Metabolite

22.05.2016
44
Zeitskala der Hochlichtantwort
Sekunden Minuten Stunden Tage
Elektronentransport gestört
∆ pH
reduzierter PQ pool
Protein Degradation und PSII Reparatur
Antennengröße
erhöhter Xanthophyll Pool Änderung der PSI/ PSII Stöchiometrie
Photochemie
Metabolismus
Translatom
Transkriptom
Wachstum
Dark qE
qT
qI10 – 200 s
10 – 60 s
10 – >30 min
10 – 60 min
NPQ Kinetiken
Adapted from Jahns, P. and Holzwarth, A.R. (2012). The role of the xanthophyll cycle and of lutein in photoprotection of Photosystem II. Biochim. Biophys. Acta Bioenergetics. 1817: 182-193. Dietz, K.-J. (2015). Efficient high light acclimation involves rapid processes at multiple mechanistic levels. J. Exp. Bot. 66: 2401-2414. Erickson, E., Wakao, S. and Niyogi, K.K. (2015). Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 82: 449-465.
State transitions
Bedarf an NADPH and ATP regulieren die Lichtreaktionen
from Schöttler, M.A., Tóth, S.Z., Boulouis, A. and Kahlau, S. (2015). Photosynthetic complex stoichiometry dynamics in higher plants: biogenesis, function, and turnover of ATP synthase and the cytochrome b6f complex. J. Exp. Bot. 66: 2373-2400.
Ist Angebot > Nachfrage: erhöhte NADPH & ATP Spiegel induzieren Photoprotektion (rote Pfeile)
NPQ
pH-induzierte Umwandlung von Violaxanthin (V) zu Zeaxanthin (Z)
PhotosynthetischeKontrolle der PlastoquinolReoxidation
Metabolische Ungleichgewichte, Trockenheit, Kälte, Infektionenund andere Umweltfaktorenkönnen den Flux durch den Calvin-Benson Zyklusvermindern.

22.05.2016
45
Zusammenfassung: Photosynthese-Reaktionen sind flexibel und können sich
anpassenVariabel
Lichtintensität, Wellenlänge, Winkel, Dauer
AntwortLichtsammlung ändert sich
Chloroplasten und Blattbewegungen, Akkumulation von Pigmentenund Proteinen
VariabelChloroplasten ChemieTransthylakoid pH/ pmfPQ / PQH2
Energie Dissipation und Photonen Gleichgewicht
Photoprotektion
qE, quenching in LHC
* *
qT, state transition
VariabelPhysiologyUmwelstress
Plastidäre terminale OxidaseAntioxidantien
VariabelLicht und Stress-induzierte ReaktiveSauerstoffspecies
Schutzmechanismen vorphoto-oxidativem
Schaden erhöhen die Photosyntheseleistung