photoinhibition of stimulable bioluminescence in marine dinoflagellates
TRANSCRIPT

Photochemistry and Photobiology Vol. 33, pp. 909 to 914, 1981 Printed in Great Britain. All rights reserved
003 1-8655/81/060909-06502.00/0 Copyright 0 1981 Pergamon Press Ltd
PHOTOINHIBITION OF STIMULABLE BIOLUMINESCENCE IN MARINE DINOFLAGELLATES
J. P. HAMMAN, W. H. BIGGLEY and H. H. SELIGER* McCollum-Pratt Institute and Department of Biology, The Johns Hopkins University,
Baltimore, MD 21218, USA
(Received 6 August 1980; accepted 8 December 1980)
Abstract-Different genera of bioluminescent photosynthetic dinoflagellates exhibit different mechan- isms for the inhibition of stimulable bioluminescence during daylight. These are (a) reduction in bio- luminescence capacity, (b) increased refractoriness to mechanical stimulation, and (c) inhibition of trans- mission of signals from mechanical receptor sites to bioluminescence emission sites. The increase in stimulable bioluminescence that in nature mirrors the decrease in sunlight intensity prior to sunset is dependent upon the logarithm of the ambient irradiation intensity. Photoinhibition of bioluminescence in all species examined except Gonyaulax polyedra is the result of absorption of light in the blue region of the spectrum.
INTRODUCTION
Bioluminescent marine dinoflagellates, upon being subjected to deforming forces, emit flashes of light that may have some survival value against predators (Seliger and McElroy, 1965; Esaias and Curl, 1972; White, 1979). Except for the non-photosynthetic Noc- tiluca miliaris and Polykrikos schwartzii, all of the dinoflagellate species that have been examined emit much less light upon mechanical stimulation during daylight (photophase) than during the night (scoto- phase). This photoinhibition of the mechanically stimulable luminescence (MSL) of dinoflagellates appears to be expressed differently in the genera Pyr- odinium, Gonyaulax, Dissodinium and Pyrocystis.
In nature the MSL of dinoflagellates during the period preceding sunset increases as the ambient sun- light intensity decreases. The question that arose was whether this increase in MSL is induced by a decrease in ambient intensity or whether the level of MSL is dependent directly upon the ambient intensity. In this paper we have effectively 'halted the sun' in its down- ward course and examined the effects of different lower ambient intensities on the MSL of the four genera. We have been able to interpret the mechanical stimulation experiments by comparisons with chemi- cal stimulation which bypasses mechanical stimu- lation and to show three separate mechanisms for photoinhibition of the native stimulability of bio- luminescence in the four genera above. The photo- inhibition of MSL during sunset is shown to be a steady-state photoinhibition of MSL, proportional to the logarithm of the ambient intensity of irradiation and for all species examined except Gonyaulax polyedra blue light is required for the inhibition.
*To whom reprint requests should be sent. TDAL,): time in hours after the onset of darkness (light).
MATERIALS AND METHODS
Data relative to the culture conditions and the charac- teristics of the bioluminescent marine dinoflagellates inves- tigated in this paper are given in Hamman and Seliger (1972). Only log phase cultures were used in the experi- ments. Details of the electronics of the bioluminescence measurements are given in Biggley et a/ . (1969). All data are reported in absolute units of photon emission or pho- ton emission relative to scotophase.
Two methods of mechanically stimulating the dinoflagel- lates to emit light were employed, differing in the degree of the mechanical stimulus and the time period over which the light intensity is measured. A strong mechanical stimu- lus was obtained by stirring a 3 d sample of the dinoflagellate culture in a c~lture tube at 1800 rpm with a wire bent into a U shape at 1 I end. The top of the wire U was positioned below the surriLe of the sample in order to avoid the formation of a vortex during stirring. A weak mechanical stimulus was obtained by bubbling air at various flow rates through a No. 15 cannula, with the can- nula tip extending halfway into the 3m/ sample in the culture tube.
The degree of mechanical stimulability, K, is defined as the maximum intensity of the rapid flash portion of the stimulated emission, divided by the total stimulable light. This ratio, with units of s - ' , analogous to a rate constant, is a maximum for any species during scotophase. The dimensionless quantity, K', where
was used to compare the effects of photoinhibition. Photoinhibition in the middle of scotophase (D4 to D6)t
was achieved by removing the cells from the dark and irradiating them with light of the same spectral quality and intensity as the normal photophase light.
For extended photophase experiments tubes containing the dinoflagellates remained in photophase until 10 min before LI2 (Do). At that time the tubes were placed in 150-mm diam dishes containing water at 25°C andsi;ting on a light table. The light table consisted of a white plastic diffusing plate 30.5 x 122cm below which were 4 cool white fluorescent lamps, each 1.2m in length. Room air' was drawn across the fluorescent lamps to minimize heat- ing of the water bath by convection. The light table was
909

i
L
d
L-)
\ \ 1%
09-G
s3 \
rn
I i i i I I i / /
0'0
;o
m
r
D 4
<
m
-
s-0
--I m
z 4
-i
E
0' I

Dinoflagellate bioluminescence 91 I
divided into 5 light-shielded sections. The intensity of illumination in each section was varied by placing the glass water baths on additional white diffusing plates. The white diffusing plates did not change the spectral quality of the transmitted light. The lamps were turned on at Liz, the time at which the fluorescent lights in the incubator in which the cells had been growing were scheduled to be turned off. Thus, the cells were transferred to an extended photophase of the same spectral quality as in the incuba- tor, but at 5 different ambient light intensities. The cells in the tubes were assayed for mechanically ( M S L ) and chemi- stimulated luminescence.
Experiments to measure the relative effectiveness of dif- ferent spectral regions for photoinhibition of M S L and CSL were carried out by replacing the white diffusing plates by 15 x 15 cm Corning glass filters to separate the blue, yellow + red and red regions of the spectrum. The photon intensities were calculated from thermopile measurements of the energy fluxes, using mean wave- lengths of 436, 580 and 620nm for the three spectral regions. The relative spectral emission of the cool white fluorescent lamps and the transmission bands of the Corn- ing glass filters used to isolate the blue, yellow + red and red regions of the spectra are shown in Fig. I .
RESULTS
For Gonyaulax polyedra, photoinhibited during scotophase, the initial flash emitted during the first few seconds of mechanical stimulation was much reduced from the dark scotophase controls whose total MSL is emitted during the first few seconds. However, upon continued mechanical stimulation, the bioluminescence continued to be emitted at low intensities until after several hundreds of seconds the total MSL of the photoinhibited cells was equal to that of the dark controls. If mechanical stimulation was stopped prior to complete MSL and the same cells were chemically stimulated, then the sum of M S L plus the additional CSL was always equal to the total CSL of the dark (scotophase) control cells.
For Pyrodinium bohamense, the initial flash of photoinhibited cells was also much reduced from that of the scotophase non-irradiated controls. However, no further light was emitted upon continued mechan- ical stimulation and no further light was emitted upon chemical stimulation.
For Dissodinium and Pyrocystis, the initial flash of photoinhibited cells was also much reduced from that
t 1 1 LL 2 3 4 5
LL lo’ - 1 0 1
HOURS
Figure 2. Dependence on ambient white light irradiance of the steady-state level of mechanically stimulated lumi- nescence for P. ucuta. At r = 0, the end of the light phase of the 12:12 photoperiod the cells were illuminated at intensities of 32.0 (0). 62.5 (0). and 150 (A) pW cm-’. The cells represented by the solid circles (0) were allowed to go into the dark at I = 0. The photophase intensity to which all cells were exposed prior to t = 0 was 278 pW cm-‘. MSL was assayed by stirring and the light emission was
integrated for 60 s.
of the scotophase non-irradiated cells. Although no further light was emitted upon continued mechanical stimulation, upon chemical stimulation, an additional bioluminescence was emitted such that
M S L + additional CSt = total CSL
These data are summarized in Table 1 for the four genera.
From the second column in Table 1, it is seen that photoinhibition in all genera reduces the flash re- sponse that would be stimulated in nature by a preda- tor. However, by virtue of the experimental technique for measuring this effect, it is possible to unravel three different mechanisms that are operating. By compari- son of columns 3 and 4, these mechanisms are rep- resented by (Yes, Yes), (No, No) and (No. Yes).
Table 1. Characteristics of MSL and CSL for dinoflagellates photoinhibited during scotophase relative to scotophase
Species
Gonyarr1a.u polyedra Gonyau1a.u catenella Gotiyau1a.u acatenella* Pyrodinium bahamense Dissodinium lunula Pyrocystis noctiluca Pyrocystis jiusiformis Pyrocystis acuta
Initial rapid flash upon mechanical
stimulation
Reduced to 1% Reduced to 1% Reduced to 1% Reduced to 1% Reduced to 1% Reduced to 1% Reduced to 4% Reduced to 3%
Additional M S L upon prolonged mechanical
stimulation
Yes (up to 100%) Yes (up to 100%) Yes (up to 100%) No No No No N o
Additional CSL subsequent to
initial MSL flash
Yes (up to 100%) Yes (up to 100%) Yes (up to 100%) No Yes (up to 80%) Yes (up to 60%) Yes (up to 50%) Yes (up to SOYo)
Bioluminescence capacity (CSL)
Unchanged Unchanged Unchanged Reduced to 1% Unchanged Unchanged Unchanged Unchanged
* Data from Esaias et al. (1973). .
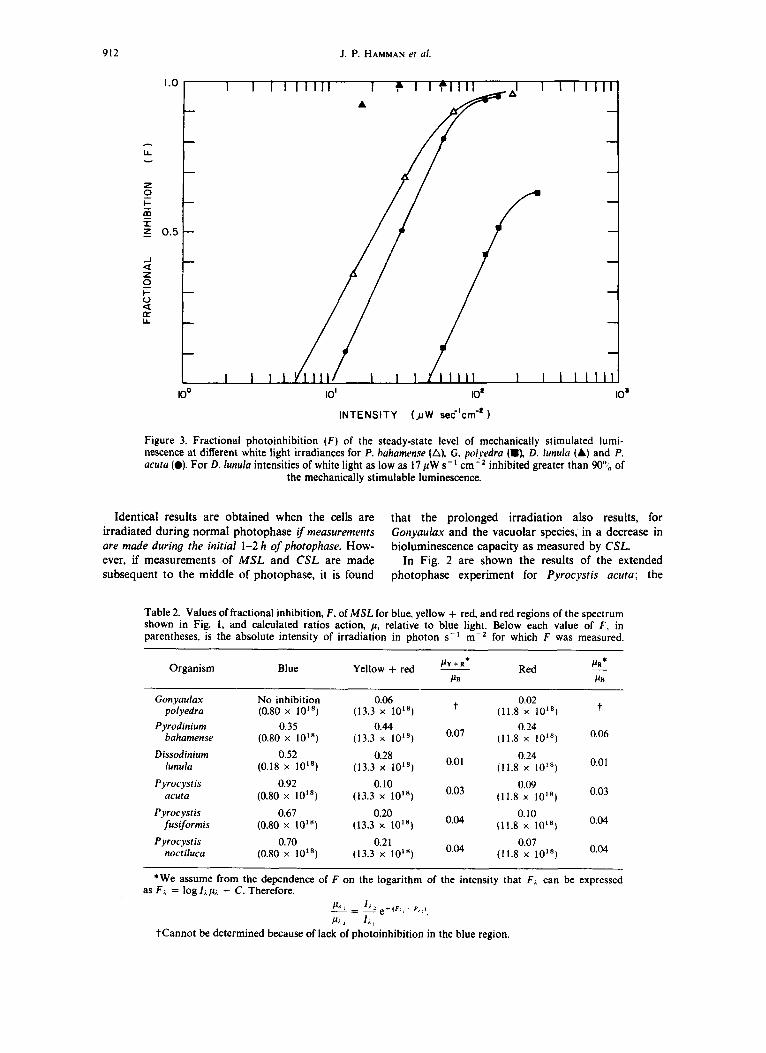
912 J. P. HAMMAN er ul.
INTENSITY (JIW sec'crn-'
Figure 3. Fractional photoinhibition ( F ) of the steady-state level of mechanically stimulated lumi- nescence at different white light irradiances for P. huhamense (A), G. polyedra (m), D. lunulu (A) and P. acuta (0). For D. lunula intensities of white light as low as 17 pW s- I inhibited greater than 90"; of
the mechanically stimulable luminescence.
Identical results are obtained when the cells are that the prolonged irradiation also results, for irradiated during normal photophase if measurements G o n y d a x and the vacuolar species, in a decrease in are made during the initial 1-2 h of photophase. How- bioluminescence capacity as measured by CSL. ever, if measurements of MSL and CSL are made In Fig. 2 are shown the results of the extended subsequent t o the middle of photophase, it is found photophase experiment for Pyrocyst is acuta; the
Table 2. Values of fractional inhibition, F, of M S L for blue, yellow + red, and red regions of the spectrum shown in Fig. 1, and calculated ratios action, p, relative to blue light. Below each value of F , in parentheses, is the absolute intensity of irradiation in photon s-I m-2 for which F was measured.
PR* Organism Blue Red - PY + R* Yellow + red - PE PB
t 0.02 (11.8 x 10")
Pyrodinium 0.35 0.44 0.24
Dissodinium 0.52 0.28 0.24
t Gonyaulax No inhibition 0.06 polyedra (0.80 x 10") (13.3 x 10")
bahumense (0.80 x 10") (13.3 x 10") 0'07 (11.8 x lo1') 0.06
0.01 lunula (0.18 x lot8) (13.3 x lo1*) 0'01 (11.8 x 10")
Pyrocystis 0.92 0.10 0.09 (13.3 x 10'8) 0'03 (11.8 x 10") 0.03 acuta (0.80 x 10")
0.04 Pyrocyst is 0.67 0.20 0.10 fusiformis (0.80 x 10") (13.3 x 10") (11.8 x loL8)
0.04 Pyrocystis 0.70 0.21 0.07 noctiluca (0.80 x 10") (13.3 x 10") (11.8 x 10")
*We assume from the dependence of F on the logarithm of the intensity that F;. can be expressed as F i = log l ~ p i - C. Therefore,
p ' . 6 - ' j . 2 e+(F , , - F , : )
P i : I;. , t Cannot be determined because of lack of photoinhibition in the blue region
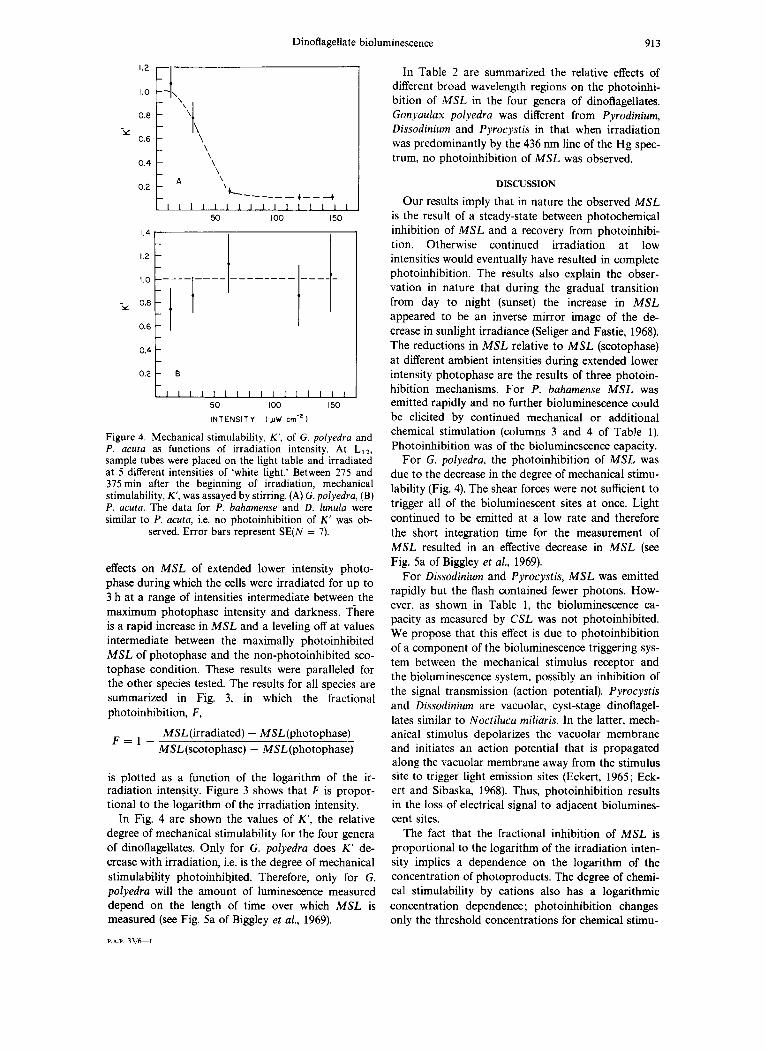
Dinoflagellate bioluminescence 913
F I I
\ Y
0.6 I- \ I
1.4
1.2
1.0
-x 0.8
0.6
0.2
I I I I I , I , I , , , I I 50 I00 I50
INTENSITY ( JIW ern-')
Figure 4. Mechanical stimulability, K‘ , of G. polyedra and P. acuta as functions of irradiation intensity. At LI2, sample tubes were placed on the light table and irradiated at 5 different intensities of ‘white light.’ Between 275 and 375 min after the beginning of irradiation, mechanical stimulability, K‘, was assayed by stirring. (A) G. polyedra, (B) P. acuta. The data for P. bahamense and D . lunula were similar to P. acuta, i.e. no photoinhibition of K’ was ob-
served. Error bars represent SE(N = 7).
effects on MSL of extended lower intensity photo- phase during which the cells were irradiated for up to 3 h at a range of intensities intermediate between the maximum photophase intensity and darkness. There is a rapid increase in MSL and a leveling off at values intermediate between the maximally photoinhibited MSL of photophase and the non-photoinhibited sco- tophase condition. These results were paralleled for the other species tested. The results for all species are summarized in Fig. 3, in which the fractional photoinhibition, F,
MSL (irradiated) - MSL(photophase) MSL(scotophase) - MSL(photophase)
F = l -
is plotted as a function of the logarithm of the ir- radiation intensity. Figure 3 shows that F is propor- tional to the logarithm of the irradiation intensity.
In Fig. 4 are shown the values of K’, the relative degree of mechanical stimulability for the four genera of dinoflagellates. Only for G. polyedra does K’ de- crease with irradiation, i.e. is the degree of mechanical stimulability photoinhibited. Therefore, only for G. polyedra will the amount of luminescence measured depend on the length of time over which MSL is measured (see Fig. 5a of Biggley et a/., 1969).
P.A.P. 3316-1
In Table 2 are summarized the relative effects of different broad wavelength regions on the photoinhi- bition of MSL in the four genera of dinoflagellates. Gonyaulax polyedra was different from Pyrodinium, Dissodinium and Pyrocystis in that when irradiation was predominantly by the 436 nm line of the Hg spec- trum, no photoinhibition of MSL was observed.
DISCUSSION
Our results imply that in nature the observed MSL is the result of a steady-state between photochemical inhibition of MSL and a recovery from photoinhibi- tion. Otherwise continued irradiation at low intensities would eventually have resulted in complete photoinhibition. The results also explain the obser- vation in nature that during the gradual transition from day to night (sunset) the increase in MSL appeared to be an inverse mirror image of the de- crease in sunlight irradiance (Seliger and Fastie, 1968). The reductions in MSL relative to MSL (scotophase) at different ambient intensities during extended lower intensity photophase are the results of three photoin- hibition mechanisms. For P . bahamense MSL was emitted rapidly and no further bioluminescence could be elicited by continued mechanical or additional chemical stimulation (columns 3 and 4 of Table 1). Photoinhibition was of the bioluminescence capacity.
For G. polyedra, the photoinhibition of MSL was due to the decrease in the degree of mechanical stimu- lability (Fig. 4). The shear forces were not sufficient to trigger all of the bioluminescent sites a t once. Light continued to be emitted at a low rate and therefore the short integration time for the measurement of MSL resulted in an effective decrease in MSL (see Fig. 5a of Biggley et al., 1969).
For Dissodinium and Pyrocystis, MSL was emitted rapidly but the flash contained fewer photons. How- ever, as shown in Table l , the bioluminescence ca- pacity as measured by CSL was not photoinhibited. We propose that this effect is due to photoinhibition of a component of the bioluminescence triggering sys- tem between the mechanical stimulus receptor and the bioluminescence system, possibly an inhibition of the signal transmission (action potential). Pyrocystis and Dissodinium are vacuolar, cyst-stage dinoflagel- lates similar to Noctiluca miliaris. In the latter, mech- anical stimulus depolarizes the vacuolar membrane and initiates an action potential that is propagated along the vacuolar membrane away from the stimulus site to trigger light emission sites (Eckert, 1965; Eck- ert and Sibaska, 1968). Thus, photoinhibition results in the loss of electrical signal to adjacent biolumines- cent sites.
The fact that the fractional inhibition of M S L is proportional to the logarithm of the irradiation inten- sity implies a dependence on the logarithm of the concentration of photoproducts. The degree of chemi- cal stimulability by cations also has a logarithmic concentration dependence; photoinhibition changes only the threshold concentrations for chemical stimu-

914 J. P. HAMMAN et al.
lability. Thus, for both stimulation and inhibition there appears t o be a common factor, perhaps a change in membrane potential that changes the per- meability of the membrane t o cations.
In Pyrodinium, Dissodinium and Pyrocystis, blue light of 436 nm was most effective for photoinhibition of M S L , and yellow o r red light intensities were rela- tively ineffective as shown in Table 2. A blue light action spectrum for D. lunula has been confirmed sep- arately (Hamman et al., 1981). The absence of a 436 nm and a red light photoinhibition of MSL in G. polyedra and the positive effect of intermediate wave-
lengths imply a wavelength dependence for photoinhibition of M S L similar t o that reported by Esaias et a / . (1973) for G. catenella and G. acatenella and unlike the chlorophyll-like action spectrum reported by Sweeney et a/. (1959). The action spec- trum for photoinhibition of M S L in G . polyedra needs to be reinvestigated.
Acknowledgements-Contribution No. 1085 of the McCol- lum-Pratt Institute and Department of Biology, The Johns Hopkins University. This work was supported by the U.S. Department of Energy Contract DEAS 02 76 EV03277.
REFERENCES
Biggley, W. H., E. Swift, R. J. Buchanan and H. H. Seliger (1969) J . Gen. Physiol. 54, 96122. Eckert, R. (1965) Science 147, 1142-1145. Eckert, R. and T. Sibaska (1968) J . Gen. Physiol. 52, 258-282. Esaias, W. E. and H. C. Curl, Jr. (1972) Limnol. Oceanogr. 17, 901-906. Esaias, W. E., H. C. Curl, Jr. and H. H. Seliger (1973) J . Cell. Physiol. 82, 363-372. Hamman, J. P. and H. H. Seliger (1972) J . Cell Physiol. 80, 397408. Hamman, J. P., W. H. Biggley and H. H. Seliger (1981) Photochem. Photobiol., in press. Seliger, H. H. and W. G. Fastie (1968) J . Mar. Res. 26, 273-280. Seliger, H. H. and W. D. McElroy Light: Physical and Biologicul Action, p. 276. Academic
Sweeney, B. M., F. T. Haxo and J. W. Hastings (1959) J . Gen. Physiol. 43, 285-299. White, H. H. (1979) J . Exp. Mar. Biol. Ecol. 36, 217-224.
(1965) Press, New York.