patterns of initial versus delayed regeneration of white spruce in boreal mixedwood succession
TRANSCRIPT

Patterns of initial versus delayed regeneration ofwhite spruce in boreal mixedwood succession
Vernon S. Peters, S. Ellen Macdonald, and Mark R.T. Dale
Abstract: The timing of white spruce regeneration in aspen (Populus tremuloides Michx.) – white spruce (Piceaglauca (Moench) Voss) boreal mixedwood stands is an important factor in stand development. We examined borealmixedwood stands representing a 59-year period of time since fire and determined (1) whether and when a delayed re-generation period of white spruce occurred, (2) whether the relative abundance of initial (<20 years) versus delayed(≥20 years postfire) regeneration is related to seed availability at the time of the fire, and (3) what are the important re-generation substrates for initial versus delayed regeneration. Initial regeneration occurred primarily on mineral soil orhumus, while delayed regeneration established primarily on logs and peaked 38–44 years after fire. Of the 20 standsinvestigated, seven were dominated by initial regeneration, six were dominated by delayed regeneration, and seven wereeven mixtures of both. The dominance of a site by initial or delayed regeneration could not be simply explained byburn timing relative to mast years or distance to seed source; our results suggested that fire severity and the competi-tive influence of initial regeneration on delayed regeneration were important at fine scales. Based on our results we de-scribe several possible postfire successional pathways for boreal mixedwood forests.
Résumé : La période au cours de laquelle l’épinette blanche (Picea glauca (Moench) Voss) se régénère dans les peu-plements mixtes boréaux composés de peuplier faux-tremble (Populus tremuloides Michx.) et d’épinette blanche est unfacteur important dans le développement de ces peuplements. Nous avons étudié des peuplements mixtes boréaux cou-vrant une période de 59 ans après feu et avons déterminé (1) si une régénération retardataire d’épinette blanche s’étaitétablie et, le cas échéant, à quel moment elle était apparue, (2) si l’abondance relative de la régénération initiale(<20 ans après feu) et retardataire (≥20 ans après feu) est reliée à la disponibilité des semences au moment du feu,et (3) quels sont les substrats de régénération importants pour la régénération initiale et retardataire. La régénérationinitiale s’est installée principalement sur le sol minéral ou l’humus alors que la régénération retardataire s’est établiesurtout sur des troncs d’arbres et a atteint un sommet d’établissement entre 38 et 44 ans après feu. Sur les 20 peuple-ments étudiés, sept étaient dominés par la régénération initiale, six étaient dominés par la régénération retardataire etles sept autres étaient dominés par un mélange égal des deux types de régénération. La dominance de la régénérationinitiale ou retardataire sur une station ne pouvait pas être expliquée simplement par la relation entre le moment du feu,les années semencières ou la distance des sources de semences. Nos résultats indiquent que la sévérité du feu et lacompétition exercée par la régénération initiale sur la régénération retardataire jouent un rôle important à des échellesfines. Nous décrivons plusieurs trajectoires possibles de succession pour les forêts mixtes boréales en se basant sur nosrésultats.
[Traduit par la Rédaction] Peters et al. 1609
Introduction
The succession of trees in boreal forests after fire is fre-quently characterized by initial floristic composition modelsbecause the presence of serotiny in several coniferous spe-cies, root suckering in broadleaf species, and wind-dispersalof species from remnant unburned patches collectively influ-
ence postfire regeneration (Egler 1954; Bergeron 2000).White spruce (Picea glauca (Moench) Voss) is a non-serotinous species with short-lived seeds. Because regenera-tion of white spruce after fire depends on dispersal fromsurviving seed sources (Galipeau et al. 1997) this species ismore susceptible to being recruitment limited than most bo-real trees (Greene and Johnson 2000). Seed dispersal de-clines rapidly downwind, such that most regeneration occurswithin 100 m of a seed source (Zasada and Lovig 1983;Dobbs 1976).
Successional development of boreal mixedwood forests isenvisioned as following a transition from a broadleaf (pre-dominantly aspen; Populus tremuloides Michx.) to a whitespruce dominated canopy. However, immediate postfire re-generation of white spruce has not been regarded as criticalto this successional pattern because spruce can apparentlyestablish later after fire, often on decayed logs (Youngblood1995; Lieffers et al. 1996b; Simard et al. 1998; Kabzemsand Garcia 2004). Thus, two possible patterns reflecting theextremes of stand development (succession) patterns in bo-
Can. J. For. Res. 36: 1597–1609 (2006) doi:10.1139/X06-020 © 2006 NRC Canada
1597
Received 22 June 2005. Accepted 12 January 2006. Publishedon the NRC Research Press Web site at http://cjfr.nrc.ca on1 June 2006.
V.S. Peters1,2 and M.R.T. Dale. Department of BiologicalSciences, University of Alberta, Edmonton, AB T6G 2E9,Canada.S.E. Macdonald. Department of Renewable Resources,University of Alberta, Edmonton, AB T6G 2H1, Canada.
1Corresponding author (e-mail: [email protected]).2Present address: Department of Biology, The King’sUniversity College, 9125 50th Street, Edmonton,AB T6B 2H3, Canada.

real mixedwoods consist of (1) if spruce regeneration occursonly immediately after fire, stands will be even aged, re-flecting an initial floristics model, (2) if there is only de-layed regeneration, stands will be multiaged, reflecting relayfloristics. These two patterns differ with respect to the rela-tive importance of the mechanisms driving stand dynamics.In both cases seed source and availability of regenerationmicrosites are undoubtedly important but competition (bothintra- and inter-specific) is likely to have more influence ondelayed regeneration; as well, the types of regenerationmicrosites that are important may change over time. Thetiming of delayed regeneration after fire in boreal mixed-woods and its importance to age structures, and thus standsuccessional development, are not well understood.
Provided a seed source exists, most white spruce estab-lishment occurs over a 3- to 5-year period after fire (mainlyon mineral soil seedbeds); this is particularly true followingmast seed years (Zasada and Gregory 1969; Purdy et al.2002; Peters et al. 2005). Abundant initial regeneration ofwhite spruce would subsequently influence the light regimein the stand (Constabel and Lieffers 1996) and thus could in-hibit later white spruce regeneration. Even-aged stands dom-inated by immediate postfire regeneration may, therefore, bemore likely to occur on sites with strong seed sources andsuitable seedbeds or where fires coincide with mast years.Stands burning in nonmast years have significantly less re-generation immediately postfire than stands burning in mastyears; this is because of initial seed limitation and the rapiddeterioration of the seedbed (Peters et al. 2005). It seemslikely that regeneration of white spruce might be limitedonce the forest floor microsites become unfavourable untilthe competitive influence of the aspen canopy declines (dueto self-thinning) and suitably decayed logs become availableas regeneration microsites (i.e., stand exclusion stage; Oliverand Larson 1996). Most snags in mixedwoods fall downwithin 20 years after fire; they must then decay sufficientlyto provide a suitable substrate for spruce to establish (Lee1998).
In this study, we address the following questions regard-ing the regeneration of white spruce in fire-origin borealmixedwood forests of Alberta: (1) Does a delayed regenera-tion pulse occur, and if so, what is the timing of delayed re-generation after a fire? (2) Is dominance of a stand by initialor delayed regeneration related to seed availability at thetime of the fire (mast versus nonmast year, proximity to and
direction from seed source)? (3) What are the important re-generation substrates for initial versus delayed regeneration?(4) Do density-dependent factors influence the relative im-portance of initial versus delayed regeneration withinstands? We also present a rules-based aging approach forwhite spruce that uses cross-dating and yet facilitates rapidaging, such that factors influencing successional patterns canbe assessed across a large number of stands.
Materials and methods
Study areaThe study took place in a 60 000 km2 area of boreal
mixedwood forest in east-central Alberta, between 54°46′N,110°02′W and 57°43′N, 115°08′W. The study area is situatedwithin the Boreal Mixedwood Ecoregion. The average tem-perature is 13.5 °C from May to August and –13.2 °C fromNovember to February. Sixty percent of the mean annualprecipitation occurs as rain during July. Gray Luvisols andEutric Brunisols are the predominant soils on upland sites(Strong and Leggat 1992). Intense crown fires annually burnan estimated 0.42% of the land base within the Boreal PlainsEcozone (Stocks et al. 2003).
The fire-origin stands that we sampled were dominated byan overstory of trembling aspen, balsam poplar (Populusbalsamifera L.), and (or) paper birch (Betula papyriferaMarsh.), and an understory of white spruce (Table 1). Allstands occurred on mesic sites that were typically flat togently sloped (slope <10%).
Timing of delayed regenerationTo determine the timing of delayed regeneration, we sam-
pled white spruce in two stands per fire for fires occurring in1961 and 1941 (mast years) and in 1958 and 1942 (nonmastyears). We sampled one fire from each of 1961 and 1958 (38and 41 years old) and two fires from each of 1942 and 1941(58 and 59 years old; a total of 12 stands for this portion ofthe sampling). Sampled stands were randomly selected from20–30 mixedwood stands per fire that exceeded 95% canopykill and had a mature white spruce seed source (80+ years)at the time of the fire (confirmed with historic aerial photosand forest inventory data). Sampled stands were located300 m to 15 km apart. Mast and nonmast years were identi-fied primarily on the basis of peaks and lows, respectively,in the provincial cone collection records (1959–1996) and
© 2006 NRC Canada
1598 Can. J. For. Res. Vol. 36, 2006
Composition of regeneration (% of stand basal area)
Fire year and namea Size (ha) Start date Location (township-range)bPopulustremuloides
Populusbalsamifera
Betulapapyrifera
Piceaglauca
Otherconifers
1961 House River 2 845 4 June 80-16 21.1 43.9 22.8 6.6 5.71958 Lawrence Lake 414 na 69-24 58.9 15.6 21.4 4.0 0.01942 Logan Lake 2 774 16 May 71-10 63.8 8.0 9.4 13.8 4.91942 Wabasca 1 558 6 May 78-22 56.3 23.1 16.8 2.4 1.41941 Piche Lake 26 871 30 April 70-11, 71-10, 71-11 49.8 10.3 14.6 19.0 6.31941 Conklin Trail 4 355 26 April 71-13 63.3 8.6 13.7 14.1 0.4
Note: Fires from 1991 to 1979 are described in Peters et al. (2005).aFire name, start date, size, and location are from Delisle and Hall (1987) and Alberta Sustainable Resource Development (2005).bIndicates township and range within Alberta; all are west of the 4th meridian.
Table 1. General characteristics of the 1941 to 1961 fires and stands sampled in the boreal mixedwood region of Alberta for age of re-generation and other features.

red squirrel trapping records (1941–1958). From the poten-tial mast and nonmast years identified, we selected fires thatwere accessible, were more than 200 ha in size, and initiatedbetween 26 April and 6 June. The procedures for identifyingmast years, fires, and stands are outlined in more detail inPeters et al. (2005).
Stands were sampled during the summers of 1998–2000.In each stand, sampling was conducted at two sites, 20 and100 m into the burn away from the unburned seed source.Three sampling points were randomly located (at least 8 mapart) at each distance (site). At each sampling point we col-lected the five live white spruce nearest to the plot center foraging, for a total of 15 trees at each site in each stand (360trees in total from the 12 stands). Trees were cut at groundlevel, and the belowground stumps were excavated and col-lected. Regeneration substrate was also recorded (see be-low).
We used the following procedures to determine the age ofeach tree. A ground-level disk was scored with a fine razorblade, and a dissecting microscope (18–110× magnification)was used to count rings along two radii. Trees that by thissimple method dated to within 20 years of the fire were clas-sified as initial regeneration and were used to address the re-maining objectives. Previous cross-dating work verified thatmost trees for which ground-level ring counts aged the treeto within 20 years of the fire could actually be dated towithin 10 years of the fire. Our approach in this study reliesheavily on the fact that most trees initiate either within10 years of the fire or more than 20 years after fires. Evi-dence for this was based on previous cross-dating work thatshowed that in the first 20 years after fire, 89% of the regen-eration established within 5 years of the fire (31 stands, 8fires) (Peters et al. 2005). Furthermore, a delayed regenera-tion pulse did not occur within the first 20 years after fire,as evidenced by many empty age-classes between 8 and20 years (Peters et al. 2005). Thus, it was not necessary inthis study to age these initial recruits more carefully.
If the ground-level ring count indicated that the tree mayhave established as late as 21 years postfire, the tree wascross-dated above and below ground, according to the proce-dures in Peters et al. (2002). Both above- and below-groundcross-dating are necessary for accurate ages of white sprucebecause missing rings are common at ground level, and sub-stantial portions of the trunk may be buried below ground,contributing to further age underestimation. A total of 134trees were finally determined to have regenerated 21 or moreyears postfire, and the cross-dated ages of these trees wereused in analyses of the timing of delayed regeneration.
Initial versus delayed regenerationTo quantify the relative importance of initial versus de-
layed regeneration, and to examine which factors operatingat the time of the fire might influence the relative abundanceof each within a stand, we used trees collected from the twostands in each of the 58- (nonmast) and 59-year-old (mast)fires described above, plus similarly sampled trees from anadditional three stands per fire. Hence a total of 15 exca-vated stumps were typically collected at each of two sites(20 and 100 m from the seed source) in each of five standsfor four fires. One of the sites lacked spruce, and fewer than
15 trees occurred at a few sites, so in the end there were 39sites and 577 trees.
Having identified appropriate threshold ages for classify-ing trees as either initial regeneration or delayed regenera-tion, we used seven steps, as follows, to classify treeswithout the time-consuming process of cross-dating everyone. (1) Aging errors (difference between ground-level ringcount and best estimate of true age) were quantified in onestand from each of the 58- and 59-year-old fires (30 treesper stand, from 4 stands total) using above- and below-ground cross-dating (Peters et al. 2002). (2) For each of theremaining trees, ground-level disks were sanded to 400 gritand the rings were counted. (3) The mean aging error (as de-termined in step 1: 5.5 years; range: 4.8–6.4 years) wasadded to this ground-level ring count. (4) Trees that basedon the age after step 3 originated within 20 years of the firewere classified as initial regeneration. (5) When the age,after step 3, indicated regeneration was between 21 and31 years postfire, the tree was assigned to cross-dating (step7). (6) For the remaining trees (age at step 3 indicated regen-eration 31+ years postfire), we measured the length of theburied trunk. When the length was less than 15 cm (whichrepresents a mean error of 5.1 buried years, based on data inPeters et al. 2002), the tree was assumed to have regeneratedapproximately 26+ years postfire and was classified as de-layed regeneration. When the length of the buried trunk wasmore than 15 cm, the tree was assigned to cross-dating (step7). (7) Trees assigned to cross-dating were cross-dated be-low ground using skeleton plots, a technique that depicts rel-ative ring width for each tree ring and permits cross-datingamong samples (Peters et al. 2002). If the age after cross-dating indicated that the tree originated within 20 years ofthe fire, it was classified as initial regeneration. If the age af-ter cross-dating indicated that the tree regenerated more than20 years after fire, it was classified as delayed regeneration.Based on this process, and on the ring counts and cross-dating of the trees used for the assessment of timing ofdelayed regeneration, we were able to classify each of the577 trees as either initial (within 20 years postfire) or de-layed (20+ years postfire) regeneration.
Regeneration substratesWe recorded the rooting substrate for each of the white
spruce trees classified as initial or delayed regeneration (577trees in the 58- and 59-year-old fires, including those used toassess the timing of delayed regeneration) as well as for thesimilarly sampled trees from an additional three stands fromeach of the four fires (an additional 12 stands and 217 trees).For each site we recorded the position as either downwind ofthe seed source (relative to prevailing winds: 60–150°) ornot downwind of the seed source (all other directions). Wealso had rooting substrate data for 1332 spruce sampled in71 stands from fires that were 19 to 41 years old (see Peterset al. 2005) for comparison. To determine rooting substrate,we removed the leaf litter and humus at the base of each treeand recorded the establishment substrate as log, humus, ormineral soil. Trees rooted on mineral soil and humus weresubsequently pooled into a single mineral soil – humus classbecause the location of the root collar could not be exactlydetermined in the field (see Peters et al. 2002). The condi-tion of logs was recorded as burned or unburned (indicating
© 2006 NRC Canada
Peters et al. 1599

prefire versus postfire origin), and above or below ground,depending on whether the trunk outline was visible withoutexcavation. In fires up to 41 years old, belowground logshad existed as downed wood prior to the fire; in 58- and 59-year-old fires, belowground logs might have been downedwood that existed before the fire or a fire-killed tree thatlater fell and had decayed considerably (V.S. Peters, per-sonal observation).
To quantify the availability of logs as regeneration sub-strates and describe the general vegetation structure forstands at different times since fire, we sampled 135 stands ina total of 17 fires between 4 and 59 years old (included 46stands from the sampled 38-, 41-, 58-, and 59-year-old fires;other fires are described in Peters et al. 2005). In one plot(5 m2) at each of the two sites (20 and 100 m from seedsources) per stand, we recorded percent cover for downedlogs, bryophytes, herbs, grass (live, lying dead, standingdead), and shrubs. Canopy closure was estimated using aconvex spherical densiometer. Shrub cover was estimatedseparately for three height strata (0–0.2, 0.2–0.5, 0.5–2 m).A prism was used to quantify basal area of postfire regenera-tion by species. Postfire regeneration was easily distin-guished from residual trees by their much smaller diametersand heights.
Factors influencing initial versus delayed regenerationDensity and height of regeneration were quantified in the
20 stands (39 sites) from the 58- and 59-year-old fires, forwhich we had classified 577 trees as initial or delayed regen-eration (3 sample points per site, but only 115 in total be-cause we were unable to collect height data at two sites). Ateach of the three sampling points per site, we recorded thedensity and height of trees classified as initial or delayed re-generation within a 28.3 m2 circular plot and within a nestedsubplot (5 m2). We also noted each tree’s location with re-spect to these plots, or as being outside the 28.3 m2 plot. Wecalculated a mean height of initial regeneration within the28.3 m2 plots for use in subsequent data analyses. We calcu-lated the density of initial- and delayed-regeneration treesfrom the total regeneration density and the proportionalabundance of both initial- and delayed-regeneration trees ateach site. Densities were calculated for each sample point(based on density in the 28.3 m2 plot) and also for each site(averaging density for the three 28.3 m2 plots). As an indica-tion of seed-source strength, we counted the number ofstanding and fallen white spruce trees in the adjacent un-burned patch that were seed producing at the time of the fire(for details see Appendix D in Peters et al. 2005).
In these same stands we quantified the basal area of liveresidual deciduous and coniferous trees at each of the 39sites (using a prism) as an indication of surviving tree com-position postfire. We assumed that the proportional represen-tation of deciduous and coniferous trees in basal area wassimilar to what it would have been immediately postfire.Detailed vegetation data had been collected at each of the 39sites as part of the description of general vegetation structure(see above).
To quantify mortality (self-thinning) of postfire regenera-tion, we counted all dead spruce 1 year or older in 5 m2
plots at each of 258 sites in 135 stands from 17 fires be-tween 4 and 59 years old (the same sites we sampled for log
availability and general vegetation structure; see above; firesare described in Peters et al. 2005). This included all sam-pling points in the 39 sites from the 58- and 59-year-oldfires for which we had classified trees as initial or delayedregeneration. Plots were carefully searched for both standingand fallen dead stems.
Data analysisTo test whether delayed regeneration peaked at a particu-
lar time, we compared the age distribution of the 134 cross-dated delayed-regeneration trees to a uniform distribution,using a Kolmogorov–Smirnov (K–S) one-sample test. Wetested the age distributions for the 38- and 41-year-old fires(27 trees, 3-year age-classes) and for the 58- and 59-year-oldfires (107 trees, 1-year age-classes) separately. We alsocompared the regeneration timing (years postfire) of the“delayed-regeneration” trees between stands in 38-year-old(mast) fire and stands in 41-year-old (nonmast) fires (n = 3stands) and between stands in 58-year-old (nonmast) andstands in 59-year-old (mast) fires (n = 6 stands) with a one-way ANOVA test (including only stands that had ≥8delayed-regeneration trees). SPSS version 10.0 (SPSS Inc.,Chicago, Illinois) was used for these statistical analyses.
For analyses of the relative abundance of initial versus de-layed regeneration, we used the 577 trees from the 58- and59-year-old fires (20 stands, 39 sites), each of which wehad classified as either initial or delayed regeneration. AWilcoxon signed rank test (Zar 1996) was used to testwhether initial regeneration was more abundant than delayedregeneration in the 20 stands. To examine the influence ofvarious factors on the relative abundance of initial versus de-layed regeneration, we used the log-likelihood ratio statistic(G) (Bishop et al. 1975). We examined the effect of burntiming (mast versus nonmast year, Gm), fire replicate (Gf),and distance (Gd) and direction from (downwind or not, Gw)seed source. Finally, we categorized each stand as (1) domi-nated by initial regeneration (60% or more); (2) having anequal mix of initial and delayed regeneration (40%–60% ofeach); or (3) dominated by delayed regeneration (60% ormore). From this, we quantified the proportion of stands ineach category for each of the 58- and 59-year-old fires.
To determine whether regeneration was more abundant onmineral soil – humus or on logs we used a Wilcoxon signedrank test and data from the 794 trees (32 stands, 63 sites(one lacked spruce)) in 58- and 59-year-old fires for whichwe had recorded rooting substrate. We calculated the propor-tion of trees regenerating on mineral soil – humus and onlogs for initial- and delayed-regeneration stands in the 58-and 59-year-old fires and also quantified proportional regen-eration for different categories of downed wood (e.g., burnedor unburned, above or below ground). We similarly calcu-lated proportional regeneration on mineral soil – humus andlogs (and type of log) for each of the 19- to 41-year-old fires(71 stands, 1332 trees). Log-likelihood (G) tests were used(as above) to examine differences in the proportional regen-eration between mineral soil – humus and logs, in the 58-and 59-year-old fires, as related to burn timing (mast versusnonmast year, ′Gm), fire replicate ( ′Gf), and distance ( )′Gd anddirection from (downwind or not, ′Gw) seed source.
We used a number of different generalized linear models(GLMs) to examine which factors could explain variation in
© 2006 NRC Canada
1600 Can. J. For. Res. Vol. 36, 2006

© 2006 NRC Canada
Peters et al. 1601
initial versus delayed regeneration at the level of the site andthe sampling point in the 58- and 59-year-old stands. ThreeGLMs were conducted at the level of the site (39 sites in the58- and 59-year-old fires) as follows: (1) Is density of initialregeneration (average of the three 28.3 m2 plots per samplepoint) affected by the basal area of live residual deciduousand (or) coniferous trees (an indicator of stand-history ef-fects)? (2) Is the density of delayed regeneration affected bythe density of initial regeneration (both densities were theaverage of the three 28.3 m2 plots per sample point)? (3) Isthe proportion of delayed regeneration (of 15 trees at eachsite) affected by the density of initial regeneration (averageof the three 28.3 m2 plots per sample point)?
Two GLMs were conducted at the level of the samplingpoints (at the 39 sites there were 115 sampling points forwhich we had height data on trees classified as initial or de-layed regeneration) as follows: (1) Is the density of delayedregeneration affected by the density and (or) height of initialregeneration (all as quantified within the 28.3 m2 plot)?(2) Is density of delayed regeneration (within the 5 m2 plot)affected by density and (or) height of surrounding initial re-generation (quantified in the 28.3 m2 plot), canopy closure,cover of downed logs (both quantified within the 5 m2
plots), seed trees (within 60 m), and proportional basal areaof regeneration (by species)? For this we used the samplingpoints for which we had detailed vegetation data (one sam-ple point for each of the 39 sites).
Two GLMs were used to examine factors influencing thedensity of dead regeneration. At the site level: Is the densityof dead stems affected by the time since fire, fire timing(mast versus nonmast year), and direction from and (or)strength of seed source for the 17 different fires (4 to59 years old). At the sampling-point level: Is the density ofdead regeneration (within the 28.3 m2 plots at each samplepoint in the 58- and 59-year-old fires) affected by the heightand density of initial regeneration?
When a GLM had several independent variables, theywere added using a stepwise procedure (α = 0.05). Categori-cal independent variables (e.g., mast versus nonmast) were
entered as dummy variables. A quasi-likelihood distributionwas used for modeling count data because of overdispersion,whereas a binomial distribution was used for proportions(McCullagh and Nelder 1983). All models were tested fornonlinear effects using generalized additive models; how-ever, they did not improve the model fit and were thereforeomitted. Models were developed using S-Plus 2000 Profes-sional Release version 2.1 (Insightful Corporation, Seattle,Washington).
Results
Timing of delayed regenerationIn the 38- and 41-year-old fires, there were 27 delayed-
regeneration seedlings that established 21 to 41 years afterfire. The overall age distribution of these seedlings did notdiffer significantly from a uniform distribution (Z = 0.986,p = 0.285; Fig. 1a). No significant differences were observedin the timing of delayed regeneration between the mast-year(38-year-old) and nonmast-year (41-year-old) fires (F =0.233, p = 0.794).
In the 58- and 59-year-old fires, 107 trees were identifiedas delayed regeneration (21–59 years after fire), and theirage distribution departed significantly from a uniform distri-bution (Z = 1.710, p = 0.006). There was a distinct peak inregeneration between 38 and 44 years after fire (Fig. 1b).Most trees regenerating during this time did so on logs (ver-sus humus – mineral soil). Of all the regeneration that oc-curred on logs from 21 to 59 years after fire, 57% occurredbetween 38 and 44 years after fire (Figs. 1b and 2). Five ofthe eight stands from 58- and 59-year-old fires showed aregeneration pulse at this time period, whereas two showeda slightly later peak (45–52 years postfire). In 38- and 41-year-old stands, we observed that shrub, herb, grass, andtotal understory cover values were low, but canopy covervalues were high (Table 2). The timing of delayed regenera-tion did not vary significantly among individual stands forthe 58- and 59-year-old fires (p ≥ 0.153 in all cases), al-though the overall test statistic was marginally significant
21 26 31 36 41
0
1
2
3
4
5
6
7
Nu
mb
er
of
trees
Time since fire (years)
0
2
4
6
8
10
12
14
21 26 31 36 41 46 51 56
Humus – mineral
substrate
Log substrate
(a) 38- and 41-year-old fires (b) 58- and 59-year-old fires
Fig. 1. Timing of delayed regeneration on logs versus mineral soil – humus for (a) two stands from each of two 38- and 41-year-oldfires (n = 27 trees, 4 stands, 24 sampling points), and (b) two stands from each of four 58- and 59-year-old fires (n = 107 trees,8 stands, 48 sampling points). Trees that established within the first 20 years following fire were omitted because cross-dating showedthey originated within a few years of the fire (see Peters et al. 2002, 2005).

(F = 2.302, p = 0.051); the mean age of delayed regenera-tion ranged from 36.3 to 41.4 years among tested stands.
Initial versus delayed regenerationOverall, initial regeneration accounted for 53.6% of all
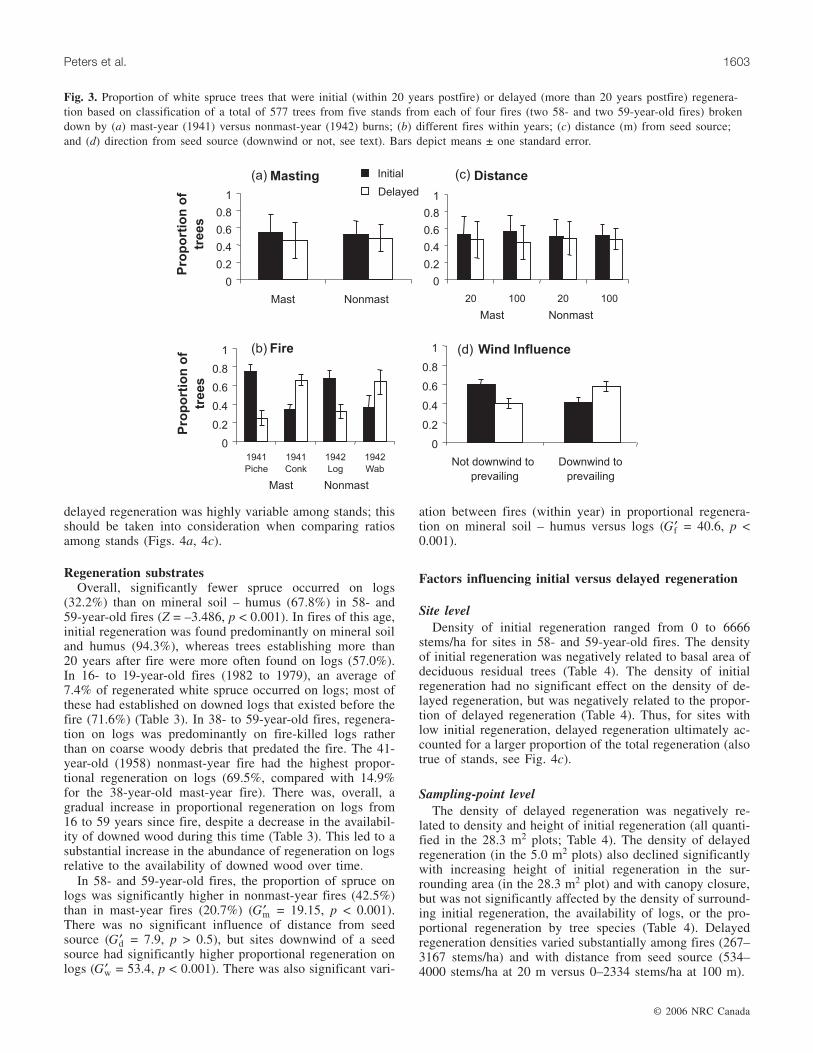
trees in 58- and 59-year-old stands (309 of 577 trees). Forstands of this age, we found no significant difference in thenumber of initial versus delayed trees (Z = –0.894, p =0.371, n = 20 stands). Mast-year fires did not have higherproportions of initial regeneration than nonmast-year fires(Gm = 0.84, p > 0.25; Fig. 3a), nor did sites 20 m from seedsources have higher proportions of initial regeneration thansites 100 m from seed sources (Gd = 1.95, p > 0.5; Fig. 3c).Initial regeneration ranged from 10% to 93% of the total re-generation (167 to 2778 stems/ha) for stands within mast-
year fires (1941) and from 0% to 100% (0–3300 stems/ha)(Fig. 4b) for stands within nonmast-year fires (1942). Therewas significant variability among fires (within year) in therelative abundance of initial versus delayed regeneration(Gf = 158.6, p < 0.001; Fig. 3b). Sites situated downwind ofseed sources had significantly higher proportions of delayedregeneration than sites that were not downwind of seedsources (Gw = 20.59, p < 0.005; Fig. 3d).
There was considerable variability among fires in the fre-quency of stands dominated by initial regeneration, delayedregeneration, or an even mixture of both (40%–60% of each;Figs. 4a, 4c). Overall, for the 58- and 59-year-old fires, 35%of stands were dominated by initial regeneration, 35% byeven mixtures of initial and delayed regeneration, and 30%by delayed regeneration (Fig. 4). The density of initial and
© 2006 NRC Canada
1602 Can. J. For. Res. Vol. 36, 2006
59-year-old fire (1941 Piche Lake) 59-year-old fire (1941 Conklin Trail)
0123456
21
-28
29
-36
37
-44
45
-52
53
-59
Nu
mb
er
of
trees
Stand 1
0123456
21
-28
29
-36
37
-44
45
-52
53
-59
Stand 6
0
1
2
3
4
5
6
21-2
8
29-3
6
37-4
4
45-5
2
53-5
9
Nu
mb
er
of
trees
Stand 2
0
1
2
3
4
5
6
21
-28
29
-36
37
-44
45
-52
53
-59
Stand 8
0
2
4
6
8
10
12
21
-28
29
-36
37
-44
45
-52
53
-59
Stand 4
0
1
2
3
4
5
6
21-2
8
29-3
6
37-4
4
45-5
2
53-5
9
Stand 1
58-year-old fire (1942 Logan Lake)
0
3
6
9
12
15
18
21-2
8
29-3
6
37-4
4
45-5
2
53-5
9
Log
Stand 5
58-year-old fire (1942 Wabasca Lake)
0
4
8
12
16
20
24
21
-28
29
-36
37
-44
45
-52
53
-59
Min–hum
Log
Stand 5
59-year-old fire (1941 Piche Lake) 59-year-old fire (1941 Conklin Trail)
0123456
21
-28
29
-36
37
-44
45
-52
53
-59
Nu
mb
er
of
trees
Stand 1
0123456
21
-28
29
-36
37
-44
45
-52
53
-59
Stand 6
0
1
2
3
4
5
6
21-2
8
29-3
6
37-4
4
45-5
2
53-5
9
Nu
mb
er
of
trees
Stand 2
0
1
2
3
4
5
6
21
-28
29
-36
37
-44
45
-52
53
-59
Stand 8
0
2
4
6
8
10
12
21
-28
29
-36
37
-44
45
-52
53
-59
Stand 4
0
1
2
3
4
5
6
21-2
8
29-3
6
37-4
4
45-5
2
53-5
9
58-year-old fire (1942 Logan Lake)
0
3
6
9
12
15
18
21-2
8
29-3
6
37-4
4
45-5
2
53-5
9
Min–hum
Stand 5
- -
0
4
8
12
16
20
24
21
-28
29
-36
37
-44
45
-52
53
-59
Time since fire (years)
Fig. 2. The timing of delayed regeneration (trees establishing 20+ years postfire) on logs versus mineral soil – humus in eight standsfrom four 59- and 58-year-old fires (a total of 30 trees per stand were aged). The 1941 fires occurred in a mast year, while the 1942fires occurred in a nonmast year.
Shrub cover by height class Grass cover by category
Stand age(years)
n(fires)a 0–0.2 m 0.2–0.5 m 0.5–2.0 m Live
Lyingdead
Standingdead
Herbcover
Total understorycoverb
Canopycover
4–7 3 8.5±1.8 11.9±4.0 8.3±2.3 6.3±3.0 10.9±8.6 5.4±4.4 21.3±10.8 72.8±19.2 25.0±4.116–19 7 6.2±1.8 6.2±1.2 5.0±0.9 1.5±0.5 5.3±2.1 1.2±0.5 10.4±2.1 34.0±4.9 51.9±5.738–41 3 0.8±0.2 7.9±1.3 4.0±1.3 0.9±0.3 1.2±0.6 0.9±0.7 12.8±1.5 27.1±5.5 73.3±10.158–59 4 1.8±0.1 6.9±1.2 6.9±2.6 1.7±0.4 2.6±1.7 1.0±0.6 17.4±5.9 36.3±8.6 55.9±7.2
Note: Values are means ± SEs.aSeven or more stands were sampled per fire.bUnderstory components were summed; however, this elevates the total estimate of cover because the overlap between understory layers is included in
the estimate.
Table 2. Canopy cover and cover of shrubs (by height class), grass (by live, lying dead, standing dead), and herb cover in mixedwoodstands of different postfire ages.
delayed regeneration was highly variable among stands; thisshould be taken into consideration when comparing ratiosamong stands (Figs. 4a, 4c).
Regeneration substratesOverall, significantly fewer spruce occurred on logs
(32.2%) than on mineral soil – humus (67.8%) in 58- and59-year-old fires (Z = –3.486, p < 0.001). In fires of this age,initial regeneration was found predominantly on mineral soiland humus (94.3%), whereas trees establishing more than20 years after fire were more often found on logs (57.0%).In 16- to 19-year-old fires (1982 to 1979), an average of7.4% of regenerated white spruce occurred on logs; most ofthese had established on downed logs that existed before thefire (71.6%) (Table 3). In 38- to 59-year-old fires, regenera-tion on logs was predominantly on fire-killed logs ratherthan on coarse woody debris that predated the fire. The 41-year-old (1958) nonmast-year fire had the highest propor-tional regeneration on logs (69.5%, compared with 14.9%for the 38-year-old mast-year fire). There was, overall, agradual increase in proportional regeneration on logs from16 to 59 years since fire, despite a decrease in the availabil-ity of downed wood during this time (Table 3). This led to asubstantial increase in the abundance of regeneration on logsrelative to the availability of downed wood over time.
In 58- and 59-year-old fires, the proportion of spruce onlogs was significantly higher in nonmast-year fires (42.5%)than in mast-year fires (20.7%) ( ′Gm = 19.15, p < 0.001).There was no significant influence of distance from seedsource ( ′Gd = 7.9, p > 0.5), but sites downwind of a seedsource had significantly higher proportional regeneration onlogs ( ′Gw = 53.4, p < 0.001). There was also significant vari-
ation between fires (within year) in proportional regenera-tion on mineral soil – humus versus logs ( ′Gf = 40.6, p <0.001).
Factors influencing initial versus delayed regeneration
Site levelDensity of initial regeneration ranged from 0 to 6666
stems/ha for sites in 58- and 59-year-old fires. The densityof initial regeneration was negatively related to basal area ofdeciduous residual trees (Table 4). The density of initialregeneration had no significant effect on the density of de-layed regeneration, but was negatively related to the propor-tion of delayed regeneration (Table 4). Thus, for sites withlow initial regeneration, delayed regeneration ultimately ac-counted for a larger proportion of the total regeneration (alsotrue of stands, see Fig. 4c).
Sampling-point levelThe density of delayed regeneration was negatively re-
lated to density and height of initial regeneration (all quanti-fied in the 28.3 m2 plots; Table 4). The density of delayedregeneration (in the 5.0 m2 plots) also declined significantlywith increasing height of initial regeneration in the sur-rounding area (in the 28.3 m2 plot) and with canopy closure,but was not significantly affected by the density of surround-ing initial regeneration, the availability of logs, or the pro-portional regeneration by tree species (Table 4). Delayedregeneration densities varied substantially among fires (267–3167 stems/ha) and with distance from seed source (534–4000 stems/ha at 20 m versus 0–2334 stems/ha at 100 m).
© 2006 NRC Canada
Peters et al. 1603
0
0.2
0.4
0.6
0.8
1
1941
Piche
1941
Conk
1942
Log
1942
Wab
(b) Fire
Pro
po
rtio
no
f
trees
Mast Nonmast
0
0.2
0.4
0.6
0.8
1
Mast Nonmast
(a) MastingP
rop
ort
ion
of
trees
(c) Distance
0
0.2
0.4
0.6
0.8
1
20 100 20 100
Mast Nonmast
0
0.2
0.4
0.6
0.8
1
Not downwind to
prevailing
Downwind to
prevailing
(d) Wind Influence
Initial
Delayed
Fig. 3. Proportion of white spruce trees that were initial (within 20 years postfire) or delayed (more than 20 years postfire) regenera-tion based on classification of a total of 577 trees from five stands from each of four fires (two 58- and two 59-year-old fires) brokendown by (a) mast-year (1941) versus nonmast-year (1942) burns; (b) different fires within years; (c) distance (m) from seed source;and (d) direction from seed source (downwind or not, see text). Bars depict means ± one standard error.

The density of dead white spruce in plots from the 17 dif-ferent fires (aged 4 to 59 years) increased significantly withtime since fire and number of seed trees (within 60 m); itwas also significantly higher on mast-year fires (~six timesthat of nonmast-year fires) and downwind of seed sources(Table 4). Although only one dead spruce was found in 10fires (82 stands) in the 4- to 20-year-old fires, dead spruceincreased to 110 and 479 stems/ha in 38- and 59-year-oldmast-year fires, respectively. In 58- and 59-year-old fires,the density of dead spruce (in the 5 m2 plots) increased sig-nificantly with increasing density of the surrounding initialregeneration (in the 28.3 m2 plots; Table 4).
Discussion
Timing of delayed regenerationOur data from the 58- and 59-year-old fires suggest that
there is a window of opportunity for delayed regeneration ofwhite spruce 38–44 years after fire in mixedwood stands.
This regeneration occurred predominantly on fire-killedlogs. Regeneration of white spruce on logs has been reportedin several studies throughout the boreal forest (Rowe 1955;Lieffers et al. 1996b; Simard et al. 1998), but the timing wasnot previously well documented. The timing of the delayedregeneration pulse coincided with the time when most snagshad fallen (only 8% remain standing after 23 years; Lee1998) and could have reached suitable decay classes forspruce establishment. Further, stands of this age were char-acterized by reduced cover of understory vegetation, buthigh canopy cover. Leaf area index in aspen-dominatedstands in Alberta may peak by age 9 and begin declining af-ter age 25 or 35 (Lieffers and Stadt 1994; Pinno et al. 2001).
In 38- and 41-year-old stands, seedlings were usuallyfound on fallen logs that were above ground but partiallycovered by a thin layer of moss (<1 cm), had no bark re-maining, and were soft with small pieces of wood missing.In our oldest stands (58 and 59 years old), downed woodwas less visible and in a later decay class. Transplant and
© 2006 NRC Canada
1604 Can. J. For. Res. Vol. 36, 2006
(a)
(b)
00.20.40.60.8
1
1941Piche
1941Conk
1942Log
1942Wab
Fire
Pro
po
rtio
n o
fst
and
s
Initial
Delayed
Even
Mast Fires Nonmast Fires
0
1000
2000
3000
4000
5000
6000
0 1000 2000 3000 4000
Initial regeneration
Del
ayed
reg
ener
atio
n(s
tem
s/h
a)
Piche Lake, MConklin Trail M,Logan Lake, NMWabasca Lake, NM
0
1000
(stems/ha)
0
1000
2000
3000
4000
5000
6000
0 1000 2000 3000 4000
Initial regeneration
InitialDelayedEven
(stems/ha)
Del
ayed
reg
ener
atio
n(s
tem
s/h
a)
Fig. 4. (a) Proportion of stands dominated by initial regeneration, delayed regeneration, or an equal mixture of each along with densityof initial versus delayed regeneration for five stands within each fire with (b) stands classified by fire (mast-year (M), 59-year-old fire;or nonmast-year (NM), 58-year-old fire) and (c) stands classified as being dominated by initial regeneration, delayed regeneration, oran even mixture of both. Initial: stands had 60% or more initial regeneration; delayed: stands had 60% or more delayed regeneration;even: stands had 40%–60% of each of initial and delayed regeneration.

© 2006 NRC Canada
Peters et al. 1605
Fireyear
Time sincefire (years)
Cover of downedlogs (%)a
Density(stems/ha)b
Proportion onlogsc,d
Index of treeson logse
No. of treesassessed
1994 4 3.1±0.4 578±266 — — —1992 6 1.7±0.9 0±0 — — —1991 7 8.2±2.3 2089±480 — — —1982 16 7.9±4.5 1996±738 0.063 0.16 4231981 17 7.6±4.8 542±496 0.100 0.07 801980 18 6.6±3.0 1242±919 0.067 0.13 3811979 19 8.9±1.9 4033±1272 0.066 0.30 301961 38 6.3±4.8 2714±333 0.149 0.64 2081958 41 3.8±3.4 762±164 0.695 1.39 2101942 58 3.7±2.6 2083±1000 0.425 2.39 3761941 59 2.5±1.7 2271±354 0.207 1.88 418
aAll means are from at least seven stands from a single fire, except 16-, 18-, 58-, and 59-year-old fires, which are meansof 14 (or more) stands from two fires.
bBased on data published in Peters et al. (2005).cThe root collar of the sampled trees was situated in wood.dThe substrate was not recorded for trees in the 1991, 1992, and 1994 fires, but we noted that most of the seedlings were
rooted in mineral soil or thin humus.eCalculated as (mean density of trees/m2 × proportion on logs × (1 / proportion cover of downed logs)).
Table 3. Cover of downed logs (mean ± SE), white spruce regeneration density (mean ± SE), the proportionof regeneration on logs, and an index of tree regeneration on logs relative to the availability of logs forstands sampled 4 to 59 years postfire.
(a) Site level.
Dependent variable
Independent variableDensity of initialregeneration
Density of delayedregeneration
Proportion delayedregeneration
Density of deadstemsa
Density of initial regeneration —b 2.1, 0.978 28.8, 0.026 (–) —Decid. residual basal area 9.9, <0.001 (+) — — —Conif. residual basal area 0.6, 0.039 (–) — — —Time since fire — — — 26.6, <0.001 (+)Masting — — — 14.9, <0.001c
Direction from seed source — — — 7.9, <0.001d
Initial seed trees within 60 m — — — 11.3, <0.001 (+)(b) Sampling-point level.
Dependent variable
Independent variableDensity of delayedregeneratione
Density of delayedregenerationf
Density of deadstemsg
No. of initial regeneration 2.8, 0.009 (–) 4.5, 0.058 45.0, <0.001 (+)Height of initial regeneration 21.1, <0.001 (–) 21.1, <0.001 (–) 0.1, 0.755Canopy closure 11.6, 0.005 (–) —
Note: Each column within (a) or (b) describes a single GLM. All analyses are from the 58- and 59-year-old stands, except as oth-erwise noted. Values are percent deviance, p value, and the effect of independent variable (positive (+) or negative (–) when signifi-cant, for continuous independent variables only).
aIncluded data from 135 stands in 17 fires from 4 to 59 years old.bBlank cells indicate that the independent variable was not included in that GLM.cMast > nonmast.dDownwind > not downwind.eIncluded data from 115 sample points in 39 stands from 58- and 59-year-old fires.f Included data from one plot at each of 39 sites from 58- and 59-year-old fires (for which detailed vegetation data were avail-
able); variables tested but not significant: cover of downed wood, number of seed trees within 60 m, proportional basal area of re-generation by species.
gIncluded data from 115 sample points in 39 stands from 58- and 59-year-old fires.
Table 4. Results of generalized linear models predicting density of initial or delayed regeneration, proportion of de-layed regeneration, and the density of dead stems at the level of the (a) site and (b) sampling point, as described inthe text.

seeding experiments in similar mixedwood stands in Alberta(43–73 years old) indicated that establishment and over-winter (first year) survival are higher on logs than on leaflitter (Delong et al. 1997). This can be attributed to more fa-vourable light and nutrient conditions, reduced competition,less leaf litter accumulation, and better root growth (Liefferset al. 1996b; Delong et al. 1997; Simard et al. 1998). Thesefactors likely contributed to the proportionally greater regen-eration on logs with time in our study, despite a decrease inexposed downed wood. The window of opportunity for de-layed regeneration on logs may be short because over timethe depth of moss increases, herbs colonize, and further de-cay results in eventual burial by leaf litter. No seedlingswere found on downed wood arising from self-thinning ofpostfire aspen or birch in stands up to 59 years old. Theselogs were likely too small and were too rapidly covered withleaf litter or insufficiently decayed to serve as regenerationmicrosites during the time frame examined. The delayed re-generation pulse we observed also may have benefited fromreduced competition with shrubs, grasses, and herbs, whichcombined had the lowest cover in stands 38 and 41 yearspostfire. Further delayed regeneration of white spruce de-clined by 58 and 59 years after fire, by which time standswere characterized by lower canopy tree cover but increasedtotal understory cover. Overall, our results suggest thatavailability of downed logs as a regeneration microsite andcompetition with understory plants (as opposed to just can-opy trees) both influence the timing of later establishment ofwhite spruce.
The timing of the delayed regeneration pulse could varywith preburn stand structure, burn severity, and availabilityof a seed source. Our sampled stands were likely older than80 years at the time of the fire; thus a supply of large fire-killed snags would have fallen and reached suitable decayclasses for spruce establishment at a similar time in all thestands studied. Stands that burn at a younger age might beless likely to show a distinct pulse of delayed regenerationbecause smaller fire-killed trees could be less suitable as re-generation microsites. In mature stands that experience lessintense burns, delayed regeneration on logs may be moregradual because canopy trees would die slowly over time,gradually recruiting into decay stages suitable for spruce es-tablishment.
Factors influencing the importance of initial versusdelayed regeneration
This study relies heavily on the identification of appropri-ate threshold ages for classifying trees according to mean-ingful successional classes; such thresholds and classeswould likely vary among plant communities and regions.Our approach relies on prior cross-dating work with whitespruce in which we determined the duration of initial regen-eration and showed the errors associated with aging progres-sively older trees in the study area (Peters et al. 2002, 2005).The distinct regeneration periods of initial regeneration (3–5 years postfire), lack of regeneration between 11 and20 years postfire (<1% based on 22 stands from 16- to 20-year-old fires (Peters et al. 2005)), and evidence presentedherein that most delayed regeneration occurred well after20 years postfire (our threshold for classifying trees as initialversus delayed regeneration) provide rigor in our quantifica-
tion of the timing of delayed regeneration. Thus, it was notnecessary in this study to age most initial recruits precisely,nor may it be warranted for studies investigating broadsuccessional patterns across a large number of stands (Peterset al. 2004).
Fire scale versus stand scaleImmediate postfire regeneration of white spruce depends
on immediate availability of mineral soil seedbeds and astrong seed source (Purdy et al. 2002; Zasada and Gregory1969). Our results confirmed that most initial regenerationoccurs on mineral soil or humus. Because mast-year burnshave been shown to have much higher densities of initial re-generation (versus nonmast-year burns) we expected mostindividual stands to be dominated by initial regeneration(Peters et al. 2005). This may not have occurred for threemain reasons: (1) While higher densities of initial regenera-tion on mast-year burns were still evident in fires as old as38–41 years, (Peters et al. 2005; Table 2), our present de-tailed analysis of the age structures from the same 58- and59-year-old fires show that within a few more years, delayedregeneration can constitute proportionally more of the totalregeneration within individual stands of a mast-year burn(e.g., Conklin Trail mast-year fire, Fig. 4b). (2) All of the58- and 59-year-old fires were spring fires (start dates be-tween 26 April and 16 May), which typically consume lesshumus and would likely have less mineral soil exposure(hence less initial regeneration) than summer fires. The fireswhere initial regeneration contributed to significantly higherdensities on mast-year burns (7, 20, and 38 years old; Peterset al. 2005) were primarily summer fires (start dates between30 May and 3 July). This helps explain why the 59-year-oldmast-year spring burns did not have higher overall sprucedensities than the 58-year-old nonmast-year burns (Table 3;Peters et al. 2005). (3) Furthermore, mineral soil exposurecan range from 0% to 100% within the same stand-replacingfire (Zasada and Gregory 1969; Greene et al. 2005), leadingto substantial variation in the density of immediately estab-lishing white spruce among stands (Purdy et al. 2002). Den-sities of both initial and delayed regeneration were highlyvariable among stands within fires (Fig. 4b). Delayed regen-eration was proportionally more abundant in stands with lowinitial regeneration densities (Table 4). The negative rela-tionship between basal area of aspen surviving the fire andinitial regeneration density may suggest that burn severitycontributed to differences in initial regeneration densityamong stands; however, there is little data in the literature tosupport the assumption that canopy mortality is well corre-lated with duff consumption.
Site scaleProximity to seed sources has been observed to contribute
to spatial variation in forest age structure and successionaldevelopment because dispersal limitations influence the den-sity and spatial pattern of regeneration (Palik and Pregitzer1995; Turner et al. 1998). In the eastern boreal forests, whitespruce may disperse seed as early as 40 years after fire, al-lowing later generations to rapidly develop further and fur-ther away from seed sources that were present immediatelyafter fire (Galipeau et al. 1997). We did not observe either ofthese phenomenon in our sites; none of the initial regenera-
© 2006 NRC Canada
1606 Can. J. For. Res. Vol. 36, 2006

tion had produced cones by age 59, nor did the ratio ofinitial to delayed regeneration differ between sites 20 and100 m from seed sources, despite up to 20-fold differencesin total density. The influence of seed availability on de-layed regeneration, however, was demonstrated by thegreater proportional abundance of delayed regeneration onsites downwind of the seed source.
Site versus sampling-point scaleOur results suggest that intraspecific competition can also
influence the relative importance of initial versus delayedwhite spruce regeneration, particularly at the sampling-pointscale. Canopy cover and height of initial regeneration (butnot density) influenced the density of delayed regeneration.Delayed regeneration rarely occurred in 5 m2 plots that hadan initial regeneration tree over 6 m tall within the surround-ing 3 m radius plot (i.e 28.3 m2; 4 of 32 cases). Much initialregeneration in 58- and 59-year old stands was below thisheight (median = 3.32 m) and likely had little effect on re-cruitment or mortality of delayed regeneration. White sprucethat regenerated within 20 years of the fire were tremen-dously variable in height (within 58- and 59-year-old stands:mean = 4.62 m; range = 0.045–18 m); thus competitive ef-fects on delayed regeneration at the sampling-point scalewould have been quite variable. Analyses at the site scaledid not account for the patchy distribution of initial anddelayed regeneration; this may have limited our ability todetect competitive interactions between age-classes at thisscale.
Mortality likely also influenced the relative abundance ofinitial versus delayed regeneration. The dead trees observedin the 58- and 59-year-old stands appeared to be mainly ini-tial regeneration (>1 m in height and virtually none of thedead trees occurred on logs, V.S. Peters, personal observa-tion), although dead stems of delayed regeneration, whichwere usually smaller, may not be recognizable for as longafter death. If initial regeneration experiences higher mortal-ity than delayed regeneration, relative abundances of the twowill change over time, perhaps masking the effect of somefactors influencing the ratio of the two (e.g., mast versusnonmast year). We found a positive relationship betweendensity of dead regeneration and mast-year burns, seedsource strength (site-level factors), and density of survivinginitial regeneration (sampling point scale); this suggests thatdensity-dependent mortality may have reduced the apparentimportance of initial regeneration. As trees grow over time,initial regeneration could have a competitive effect ondelayed regeneration (Lieffers et al. 1996b); as mentionedabove, we found a negative effect of height of initial regen-eration on density of delayed regeneration. The ratio of ini-tial to delayed regeneration is no doubt dynamic, the valueswe observed being a function of the age of our stands.
Successional pathways of mixedwoodsOur results suggest that not all mixedwood stands follow
the same pattern of postfire stand development (i.e., success-ional pathway) even for stand-replacing fires that were simi-lar with respect to seed availability. Models of forestsuccession also support the idea that the same initial condi-tions can lead to different outcomes for stand structure andcomposition (Glenn-Lewin and van der Maarel 1992). There
was greater variation in the ratio of initial to delayed regen-eration among fires (from the same year) than was seen as aresult of differences in seed source (mast versus nonmastyear and distance from seed source). This suggests that otherfactors, such as burn severity or weather conditions in theimmediate postfire period, must also be influencing the tim-ing of white spruce regeneration and, hence, stand success-ional development. Several authors have suggested that fireseverity affects spruce age structure (Rowe 1955; Lieffers etal. 1996a); however, this is difficult to demonstrate in retro-spective studies because fire behaviour cannot readily be re-constructed.
Tree regeneration in the mixedwood stands that we stud-ied does not readily fit either the initial or relay floristicsmodel of succession (Egler 1954). Few sites were stronglydominated by either initial or delayed regeneration of whitespruce; many showed a relatively even mix of both. Thispattern contrasts with much of the boreal forest where theinitial floristics model adequately describes regeneration dy-namics of tree species (Bergeron 2000). Many studies indi-cate that succession within a plant community can occur atdifferent rates and have different dominant species in lateseral stages (Olson 1958; Horn 1981; Pickett et al. 1987). Anonequilibrium picture of succession (Glenn Lewin and vander Maarel 1992) seems appropriate for boreal mixedwoodsbecause the time required for white spruce to achieve domi-nance will vary greatly; indeed, pure hardwood (aspen)stands may persist as such for hundreds of years (Cumminget al. 2000).
Overall, our results suggest two important windows of op-portunity for regeneration of white spruce: the immediatepostfire period (within 5 years; Purdy et al. 2002; Peters etal. 2005) and the period 35–45 years postfire (results pre-sented here). Effects of burn severity, seed source, availabil-ity of suitable decayed logs, and competition can result inextensive variation in density of recruitment and subsequentsurvival and growth in each of these time periods. The sur-vival and growth of delayed regeneration, and thus its contri-bution to future canopy composition, is unclear. Gutsell andJohnson (2002) found that delayed cohorts of white sprucedid not survive more than 50 years in the understory in olderboreal forests in Saskatchewan (78–154 years old). In con-trast, Lieffers et al. (1996b) found that delayed regenerationgrows as well as initial regeneration, and they concluded thatit could recruit into the canopy, given enough time. Our datasuggests that later recruiting trees establish in gaps withoutlarge initial recruits; such recruitment could result in morestands becoming fully stocked with white spruce (i.e., ≥400stems/ha, depending on spacing) than would occur solelyfrom initial regeneration.
We propose the following as possible pathways ofsuccessional development in boreal mixedwood stands domi-nated by hardwood tree species and white spruce. (1) Mixed-wood stands lacking a seed source or available seedbeds willregenerate to even-aged pure hardwood following fire.(2) Even-aged mixedwood stands, including mostly initialregeneration of white spruce, will arise following severefires at which there is a strong seed source. High densities ofinitial establishment may inhibit later regeneration.(3) Uneven-aged mixedwood stands, including mainly de-layed regeneration of white spruce, will occur following less
© 2006 NRC Canada
Peters et al. 1607

severe fires and (or) when seed sources are weak at the timeof the fire but later become strong. (4) Uneven-agedmixedwood stands, including both initial and delayed regen-eration of white spruce, will occur following moderate to se-vere fires and when there is a strong seed source availableover the long term; intraspecific competition and the highermortality of deciduous species will then influence the rela-tive abundance of the initial versus delayed cohort. Whilemost mixedwood stands will eventually become unevenaged, fire severity and seed source strength will influencehow quickly stands exhibit the characteristics of these path-ways (Lieffers et al. 1996a; Chen and Popadiouk 2002). Allof these pathways may be observed within individual firesbecause boreal fires often last many weeks, fire behaviourdepends on daily weather conditions, and both seed sourcesand regeneration are spatially variable.
Acknowledgments
We are grateful for the financial support of the Sustain-able Forest Management Network, Alberta–Pacific ForestIndustries Inc., Natural Sciences and Engineering ResearchCouncil of Canada, and Circumpolar Boreal and Arctic Re-search Grants. Special thanks to field and laboratory assis-tants, and to Lana Laird, Gitte Grover, Susan Peters, HugoAsselin, Ken Stadt, and many other reviewers.
References
Alberta Sustainable Resource Development. 2005. Historical wild-fire information [online]. Forest Protection Branch. Availablefrom http://www.srd.gov.ab.ca/wildfires/fpd/wi_hdhwd.cfm [cited8 December 2003].
Bergeron, Y. 2000. Species and stand dynamics in the mixedwoods of Quebec’s southern boreal forest. Ecology, 81: 1500–1516.
Bishop, Y.M., Fienberg, S.E., and Holland, P.W. 1975. Discretemultivariate analysis: theory and practice. MIT Press, Cam-bridge, Mass.
Chen, H.Y.H., and Popadiouk, R.J. 2002. Dynamics of NorthAmerican boreal mixedwoods. Environ. Rev. 10: 137–166.
Constabel, A.J., and Lieffers, V.J. 1996. Seasonal patterns of lighttransmission through boreal mixedwood canopies. Can. J. For.Res. 26: 1008–1014.
Cumming, S.G., Schmiegelow, F.K.A., and Burton, P.J. 2000. Gapdynamics in boreal aspen stands: is the forest older than wethink? Ecol. Appl. 10: 744–759.
Delisle, G.P., and Hall, R.J. 1987. Forest fire history maps of Al-berta, 1931 to 1983. Canadian Forest Service, Northern ForestryCentre, Edmonton, Alta.
Delong, H.B., Lieffers, V.J., and Blenis, P.V. 1997. Micrositeeffects on first-year establishment and overwinter survival ofwhite spruce in aspen-dominated boreal mixedwoods. Can. J.For. Res. 27: 1452–1457.
Dobbs, R.C. 1976. White spruce seed dispersal in central BritishColumbia. For. Chron. 52: 225–228.
Egler, F.E. 1954. Vegetation science concepts. I. Initial floristiccomposition — a factor in old field vegetation development.Vegetatio, 4: 412–417.
Galipeau, C., Kneeshaw, D., and Bergeron, Y. 1997. White spruceand balsam fir colonization of a site in the southeastern borealforest as observed 68 years after fire. Can. J. For. Res. 27: 139–147.
Glenn-Lewin, D.C., and van der Maarel, E. 1992. Patterns and pro-cesses of vegetation dynamics. In Plant succession: theory andprediction. Edited by D.C. Glenn-Lewin, R.K. Peet, and T.T.Veblen. Chapman & Hall, London. pp. 11–59.
Greene, D.F., and Johnson, E.A. 2000. Tree recruitment from burnedges. Can. J. For. Res. 30: 1264–1274.
Greene, D.F., Macdonald, S.E., Cumming, S., and Swift, L. 2005.Seedbed variation from the interior through the edge of a largewildfire in Alberta. Can. J. For. Res. 35: 1640–1647.
Gutsell, S.L., and Johnson, E.A. 2002. Accurately ageing trees andexamining their height-growth rates: implications for interpret-ing forest dynamics. J. Ecol. 90: 153–166.
Horn, H.S. 1981. Some causes of variety in patterns of secondarysuccession. In Forest succession: concepts and application.Edited by D.C. West, H.H. Shugart, and D.B. Botkin. Springer-Verlag, New York. pp. 24–35.
Kabzems, R., and Garcia, O. 2004. Structure and dynamics oftrembling aspen – white spruce stands near Fort Nelson, B.C.Can. J. For. Res. 34: 384–295.
Lee, P. 1998. Dynamics of snags in aspen-dominated midborealforests. For. Ecol. Manage. 105: 263–272.
Lieffers, V.J., and Stadt, K.J. 1994. Growth of understory Piceaglauca, Calamagrostis canadensis, and Epilobium angustifoliumin relation to overstory light transmission. Can. J. For. Res. 24:1193–1198.
Lieffers, V.J., Macmillan, R.B., MacPherson, D., Branter, K., andStewart, J.D. 1996a. Semi-natural and intensive silviculturalsystems for the boreal mixedwood forest. For. Chron. 72: 286–292.
Lieffers, V.J., Stadt, K.J., and Navratil, S. 1996b. Age structure andgrowth of understory white spruce under aspen. Can. J. For. Res.26: 1002–1007.
McCullagh, P., and Nelder, J.A. 1983. Generalized linear models.Chapman & Hall, London, UK.
Oliver, C.D., and Larson, B.C. 1996. Forest stand dynamics — up-date edition. John Wiley & Sons, Inc., Toronto, Ont.
Olson, J.S. 1958. Rates of succession and soil changes on southernLake Michigan sand dunes. Bot. Gaz. 119: 125–170.
Palik, B.J., and Pregitzer, K.S. 1995. Height growth of advanceregeneration under an even-aged bigtooth aspen (Populusgrandidentata) overstory. Am. Midl. Nat. 134: 166–175.
Peters, V.S., Macdonald, S.E., and Dale, M.R.T. 2002. Aging dis-crepancies on white spruce affect the interpretation of static agestructure in boreal mixedwoods. Can. J. For. Res. 32: 1496–1501.
Peters, V.S., Macdonald, S.E., and Dale, M.R.T. 2004. Reply to thecomment by V.J. Lieffers and K.J. Stadt on “Aging discrepan-cies of white spruce affect the interpretation of static age struc-ture in boreal mixedwoods”. Can. J. For. Res. 34: 1365–1367.
Peters, V.S., Macdonald, S.E., and Dale, M.R.T. 2005. The interac-tion between masting and fire is key to white spruce regenera-tion. Ecology, 86: 1744–1750.
Pickett, S.T.A., Collins, S.L., and Armesto, J.J. 1987. Models,mechanisms and pathways of succession. Bot. Rev. 53: 335–371.
Pinno, B.D., Lieffers, V.J., and Stadt, K.J. 2001. Measuring andmodelling the crown and light transmission characteristics of ju-venile aspen. Can. J. For. Res. 31: 1930–1939.
Purdy, B.G., Macdonald, S.E., and Dale, M.R.T. 2002. The regen-eration niche of white spruce following fire in the mixedwoodboreal forest. Silva Fenn. 36: 289–306.
Rowe, J.S. 1955. Factors influencing white spruce reproduction inManitoba and Saskatchewan. Dep. North. Aff. Nat. Resour. For.Res. Div. Tech. Note 3.
© 2006 NRC Canada
1608 Can. J. For. Res. Vol. 36, 2006

© 2006 NRC Canada
Peters et al. 1609
Simard, M.J., Bergeron, Y., and Sirois, L. 1998. Conifer seedlingrecruitment in a southeastern Canadian boreal forest: the impor-tance of substrate. J. Veg. Sci. 9: 575–582.
Stocks, B.J., Mason, J.A., Todd, J.B., Bosch, E.M., Wotton, B.M.,Amiro, B.D., Flannigan, M.D., Hirsch, K.G., Logan, K.A.,Martell, D.L., and Skinner, W.R. 2003. Large forest fires in Can-ada, 1959–1997. J. Geophys. Res. 108: 1–12.
Strong, W., and Leggat, K.R. 1992. Ecoregions of Alberta. AlbertaForestry, Lands and Wildlife, Land Inf. Serv. Div. Resour. Inf.Br. Publ. T:245.
Turner, M.G., Baker, W.L., Peterson, C.J., and Peet, R.K. 1998.Factors influencing succession: lessons from large, infrequentnatural disturbances. Ecosystems, 1: 511–523.
Youngblood, A.P. 1995. Development patterns in young conifer–hardwood forests of interior Alaska. J. Veg. Sci. 6: 229–236.
Zar, J.H. 1996. Biostatistical analysis. 3rd ed. Prentice-Hall,Englewood Cliffs, N.J.
Zasada, J.C., and Gregory, R.A. 1969. Regeneration of whitespruce with reference to interior Alaska: a literature review.USDA For. Serv. Gen. Tech. Rep. PNW-79.
Zasada, J.C., and Lovig, D. 1983. Observations on primary dis-persal of white spruce, Picea glauca, seed. Can. Field-Nat. 97:104–106.