p-glucan synthesis in the cotton fiber’ - plant physiology · p-glucan synthesis in the cotton...
TRANSCRIPT

Plant Physiol. (1995) 107: 111-123
P-Glucan Synthesis in the Cotton Fiber’
IV. In Vitro Assembly of the Cellulose I Allomorph
Krystyna Kudlicka, R. Malcolm Brown, Jr.*, Likun Li2, Jong H. Lee, Heungsop Shin, and Shigenori Kuga3
Department of Botany, The University of Texas at Austin, Austin, Texas 7871 3-7640
In vitro assembly of cellulose from plasma membrane extracts of the cotton (Gossypium hirsutum) fiber was enriched by a combina- tion of 3 4 N-morpholino)propanesulfonic acid extraction buffer and two independent digitonin solubilization steps consisting of 0.05% digitonin (SE,) followed by 1 % digitonin (SE,). Clucan synthase activity assays revealed that, although the SE, fraction possessed higher activity, only 8.6% of the in vitro product survived aceticl nitric acid treatment. On the other hand, the SE, fraction was less active, but 32.1 % of the total glucan in vitro product was resistant to acetichitric acid. In vitro products synthesized from the SE, fraction contained P-1,3-glucan and fibrillar cellulose I , whereas the SE, fraction produced P-1,3-glucan and cellulose II. Both cel- luloses assembled in vitro were labeled with cellobiohydrolase I - gold complex, and the electron diffraction patterns of both products from SE, and SE, revealed cellulose I and cellulose II, respectively. Contamination of native cellulose was ruled out by extensive evi- dente from autoradiography of the ethanol-insoluble and aceticl nitric acid-insoluble materials, including three different controls.
To understand the mechanism of cellulose biosynthesis, many attempts to synthesize cellulose in vitro have been made with cell-free systems from different sources (Franz and Heininger, 1981; Robinson and Quader, 1981; Carpita, 1982; Maclachan, 1982; Blaschek et al., 1983; Delmer, 1987; Brown, 1989a, 1989b; Dhugga and Ray, 1991; Read and Delmer, 1991). The greatest progress has been made using Acetobacter xylinum as an experimental model system, and sufficiently high rates of synthesis of P-1,4-glucan from UDP-Glc were achieved (Aloni et al., 1983; Bureau and Brown, 1987; Ross et al., 1987; Brown, 1989a; Lin and Brown, 1985, 1989; Lin et al., 1990; Saxena et al., 1990, 1991; Wong et al., 1990; Mayer et al., 1991). This enabled further in vitro synthesis experiments leading to the purification of two polypeptides involved in cellulose biosynthesis
’ This work was supported by the Johnson & Johnson Endow- ment, the Texas Advanced Technology Program (TATP 121), and Welch grant F-1217.
Present address: Department of Biochemistry and Cell Biol- ogy, Weiss School of Natural Sciences, Rice University, Houston, TX 77051.
Present address: Department of Forest Products, Faculty of Agriculture, The University of Tokyo, Yayoi Bunkyo-ku, Tokyo 113, Japan.
* Corresponding author; e-mail mbbrown8ccwf.cc.utexas.edu; fax 1-512-471-3573.
(93 and 83 kD) (Lin and Brown, 1989; Lin et al., 1990) and cloning of the genes of a cellulose synthase (Saxena et al., 1990, 1991, 1994; Wong et al., 1990).
Although the Acefobacfer system has yielded excellent progress toward a fundamental perception of in vitro cel- lulose synthesis, much confusion has been caused by the lack of adequate product analysis from higher plants, since high levels of /3-1,3-glucan (callose) are synthesized from UDP-Glc (Delmer, 1991). Now, it is realized that no single technique is sufficient for the identification and quantita- tion of P-glucans produced in vitro, and the product must be analyzed by a combination of both physical and bio- chemical techniques (Delmer, 1991; Read and Delmer, 1991; Okuda et al., 1993).
From comprehensive product analyses, we have posi- tively identified cellulose synthesized in vitro from cotton (Gossypium hirsutum) fibers (Okuda et al., 1993). The achievement of in vitro cellulose synthesis became possible through an intensive study to test and find a combination of co-factors that led to P-1,4-glucan assembly (Li and Brown, 1993). The photoaffinity-labeling study provided preliminary indication that a 37-kD polypeptide is the most likely candidate for the UDP-Glc-binding subunit of cellu- lose synthase. Furthermore, a 52-kD polypeptide was con- sidered to be the most likely candidate for the catalytic subunit of P-1,3-glucan synthase (Li et al., 1993).
Although cellulose is a P-lP-linked polymer of D-G~c, it occurs in severa1 distinct crystalline forms known as allo- morphs or polymorphs (Sarko, 1978). Nearly a11 organisms that synthesize cellulose generate the allomorph called cel- lulose I or ”native cellulose.” Only a few exceptional or- ganisms are known to synthesize cellulose I1 in vivo (Frey- Wyssling, 1976; Roberts et al., 1989; Kuga et al., 1993).
The in vitro product derived from digitonin-solubilized membranes of Acefobacfer gave diffraction patterns typical of cellulose I1 (Bureau and Brown, 1987; Lin and Brown, 1989). Similarly, ANIP synthesized from digitonin-solubi- lized membranes of cotton fibers yielded an x-ray diffrac-
Abbreviations: AN reagent, acetic/nitric acid reagent; ANIP, acetic/nitric acid reagent-insoluble product; CBH I, cellobiohydro- lase I; c-3’:5’-GMP, cyclic 3’:5’-GMP; ED, electron diffraction; EIP, ethanol-insoluble product; ES, protein fraction released into super- natant after entrapment; PM, plasma membrane; SE,, enzyme solubilized in the first solubilization step by using 0.05% digitonin; SE,, enzyme solubilized in the second solubilization step by using 1% digitonin.
111 www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.

112 Kudlicka et al. Plant Physiol. Vol. 107, 1995
PM SE-, SE2
MWS1ES 2ES
Figure 1. Protein profiles of different fractions. A, Gel stained withCoomassie blue. Lane 1 (MWS), molecular mass standards; lane 2,PM; lane 3, SE,; lane 4, SE2 (lanes 2-4, 20 /xg). B, Gel stained withsilver. Lane 1 (MWS), molecular mass standards; lane 2 (1ES), proteinreleased from SE, fraction after entrapment (note the prominent37-kD band); lane 3 (2ES), protein released after entrapment from SE2
fraction (the 37-kD band is attenuated).
tion pattern characteristic of cellulose II (Okuda et al.,1993). Thus, there has been no conclusive evidence for thesynthesis of cellulose I from an in vitro preparation, al-though S. Kuga has occasionally found in the in vitroproduct mixture fibrils that gave a typical cellulose I EDpattern. Because these fibrils were so rare and because wewere using the methodology of Okuda et al. (1993), it wasimpossible to believe at that time that we may have syn-thesized cellulose I in vitro, even though this product wasnever detected in the controls lacking substrate or en-zymes. This seemed all the more unlikely since cellulose
synthesized in vitro from UDP-Glc always resulted in theformation of cellulose II (Bureau and Brown, 1987; Lin andBrown, 1989; Okuda et al., 1993). Furthermore, it has beenargued that living organisms synthesizing cellulose I mustcontrol glucan chain crystallization in a manner not dupli-cated under acellular conditions (Ranby, 1952; Stipanovicand Sarko, 1976; Blackwell, 1982; Sawyer and George,1982).
In this study the conditions essential for in vitro celluloseI assembly have been revealed. Use of Mops buffer withSue and two digitonin extraction protocols led to an in-crease of the quantity of /3-1,4-glucan produced in vitro.Although a low concentration of digitonin (0.05%) in thefirst solubilization protocol released a small quantity ofproteins, this SEl fraction synthesized a larger percentageof ANIP (32.1%) than reported in our previous publication(4% by SE fraction) (Okuda et al., 1993). This SE, fractionalso is prerequisite for the synthesis of cellulose I. A higherconcentration of digitonin (1%) solubilized more mem-brane proteins, but this SE2 fraction produced a productthat contained less cellulose (8.6% of the total glucan prod-uct), which is essentially cellulose II.
MATERIALS AND METHODS
Chemicals
UDP-[Glc-U-14C] (318 mCi/mmol) was purchased fromICN Biochemical Inc. (Costa Mesa, CA). UDP-[Glc-l-3H](15.3 Ci/mmol) was purchased from NEN Research Prod-ucts (Dupont Co., Doraville, GA). Digitonin was purchasedfrom Serva (Heidelberg, Germany). Cellobiose, c-3':5'-GMP, protease inhibitors, and other chemicals were ob-tained from Sigma. Gold suspension (Auro Beads G5),average particle size 5 nm, was purchased from Amer-sham. CBHI enzyme was a gift from Dr. Martin Schiilein ofNovo Industri (Copenhagen, Denmark). The molecularmass standards, which contain phosphorylase b (97 kD),BSA (66 kD), ovalbumin (45 kD), carbonic anhydrase (31kD), soybean trypsin inhibitor (21 kD), and hen egg whitelysozyme (14 kD), were purchased from Bio-Rad.
Plant Materials
The cotton strain, Gossypium hirsutum Texas marker 1(TM-1; [Kohel, 1970]), provided by Barbara Triplett (South-ern Regional Research Laboratory, U.S. Department of Ag-
Table I. Glucan synthase activity in different fractionsReactions were carried out for 30 min at 25°C during which the reaction rate was found to be linear. At the end of the reactions, [14C]Glu
incorporation was measured. One unit = nmol of [14C]Glu incorporated min"1. The so values were calculated from five values obtained fromfive separate preparations.
Glucan Synthase Activity Cellulose Synthase ActivityFraction Total Protein
Total activity Specific activity Total activity Specific activity
Percentage of CelluloseSynthase Activity
versus TotalGlucan Synthase Activity
PMSE,SE2
mg
33.61.43.5
units760.5
8.052.8
units/mg22.7 ± 1.9
5.7 ± 1.215.6 ± 4.6
units50.62.54.3
units/mg1.5 ± 0.31.8 ± 0.31.3 ±0.3
6.6 ± 1.232.1 ± 2.28.6 ± 2.6
www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.

In Vitro Cellulose I Synthesis from Cotton 113
Figure 2. Negatively stained and CBH [-goldlabeled in vitro product synthesized by SE, andSE2 fractions mixed 1:1 under conditions favor-ing synthesis of /3-1,4-glucan. A, Synthesizedproduct in the reaction mixture. Fibrillar struc-tures are uniformly dispersed on the grid, andCBH l-gold particles also are dispersed in be-tween the products. X107,000 (bar = 100 nm).B, In vitro products after protease treatment.Note that the products appear in three differentmorphological forms: a, fibrillar material ar-ranged into rodlets not labeled with CBHI-goldcomplex, indicating j3-1,3-glucans; b, longfibrils; and c, disorganized aggregates. Both band c are strongly labeled with CBH l-gold par-ticles, indicating j3-1,4-glucans. XI 07,000. (Bar= 100 nm.)
riculture, New Orleans, LA), was used for the experiments.Bolls were harvested and the locules were removed andstored in liquid nitrogen as described previously (Okuda etal., 1993).
Preparation of the PM-Enriched Fractions
The PM-enriched fraction from cotton fibers was pre-pared as described previously (Okuda et al., 1993), exceptthat the extraction buffer consisting of 50 mM Mops, pH7.5, 5 mM EDTA, 0.25 M Sue, and a combination of proteaseinhibitors (0.5 mM PMSF, 10 /LIM leupeptin, 0.1 mM N-a-p-tosyl-L-Lys chloromethyl ketone, and 0.1 mM L-1-tosylam-ide-2-phenyl-ethyl chloromethyl ketone) were used. Themembranes from 50 locules were resuspended in 10 mL ofresuspension buffer consisting of 50 mM Mops, pH 7.5, and0.25 M Sue to obtain approximately 4.5 mg of protein in1 mL.
Solubilization of Membrane Proteins
Solubilized enzyme fractions were obtained separatelythrough a two-step digitonin solubilization. In the firststep, the PM-enriched fraction (approximately 5 mL) wasmixed with an equal volume (approximately 5 mL) of thefirst solubilization buffer containing 50 mM Mops, pH 7.5,0.25 M Sue, and 0.1% digitonin. The mixture was carefullylayered on a 30% (w/v) glycerol cushion and centrifuged at100,000£ for 1 h at 4°C. The supernatant was collected anddenoted as SE,. In the second solubilization step, the pelletbelow the glycerol cushion was resuspended in the second
solubilization buffer (the same volume as in the first solu-bilization protocol) containing 50 mM Mops, pH 7.5, 0.25 MSue, and 1% digitonin, and the mixture was sonicated onthe surface of ice for 5 min and then incubated on ice (totalsolubilization time was 1 h). After solubilization, the mix-ture was layered again on a glycerol cushion and centri-fuged under the same conditions as in the first solubiliza-tion step. The supernatant was collected and was denotedas SE2.
SE, and SE2 were concentrated with a Centriprep-10Concentrator (Amicon Division of W.R. Grace and Co.,Beverly, MA). The proteins in each fraction were assayedusing a modification of the Lowry procedure (Markwell etal., 1978). Electrophoresis was performed as described byPorzio and Pearson (1977), using 10% acrylamide in theseparation gel and 4.5% in the stacking gel. The loadingbuffer contained 10% 2-mercaptoethanol. The gel wasstained with 0.125% Coomassie blue R-250 in 50% metha-nol and 10% acetic acid, or with silver reagent according tothe Bio-Rad silver-staining procedure.
Preparation of the in Vitro Product for EM
The condition favoring synthesis of 0-1,4-glucan reactionmixture contained 10 mM bis-tris-propane-Hepes buffer(pH 7.6), 20 HIM cellobiose, 8 mM MgCl2, 1 HIM CaCl2, 100JU.M c-3':5'-GMP, 0.05% digitonin, the solubilized enzymefractions, 3 mM NaN3, and 1 mM UDP-Glc (in 1 mL vol-ume) and was incubated at 25°C for about 15 h (overnight).The synthesized product was centrifuged at 15,000g for 10 www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.

114 Kudlicka et al. Plant Physiol. Vol. 107, 1995
Figure 3. Autoradiography of EIP and ANIP synthesized by SE, and SE2 mixed 1:1. A, Low magnification view of EIP showingvery localized labeling of product with silver grains. X92,000 (bar = 1 ^.m). B, Higher magnification of a small fragmentof A. Silver grains and CBH l-gold particles coincide with the product. X34,700 (bar = 0.5 jim). C and D, Morphologicalappearance of ANIP. X67,500 (bar = 0.2 ju,m). C, Long fibrils indicating cellulose I, labeled with CBH l-gold and silvergrains. D, Disorganized aggregates, indicating cellulose II.
min. For the total product analysis, the pellet was washedwith deionized water, treated with protease (consisting ofpronase C [1 mg] and trypsin [1 mg] in 1 mL) for 30 min atroom temperature, washed three times in water, resus-pended in a small volume of water, sonicated for 30 s(Fisher Sonic Dismembrator model 300), and used for EMstudies. For ANIP, the pellet was resuspended in AN re-agent, placed for 30 min in a boiling water bath (Updegraff,1969), washed three times with deionized water, resus-pended in a small volume of water, and sonicated for 30 s.Both product suspensions were placed on EM grids thatwere shielded between two Petri dishes (to avoid anypossible lint from the air) and allowed to dry. In addition,the room air was filtered with 0.2-ju,m HEPA filters. TheCBH I-gold complex was prepared as described by Chanzyet al. (1984). Gold labeling of the in vitro products and the
negative staining were performed as described by Okudaet al. (1993).
For autoradiographic studies, the reaction mixture wasessentially the same as described above, except that 0.5 m.MUDP-H-^HlGlc (10 mCi/mmol) was used as the substrate.Reactions were performed overnight at 25°C. Three controlexperiments were conducted: (a) zero-time incubation, (b)incubation without substrate, and (c) incubation with allcomponents but without enzyme. After incubation, the invitro product was collected by centrifugation for 10 min at15,000g. For total product analysis, the pellet was washedthree times with water. For ANIP, the pellet was resus-pended in a small volume of AN reagent, placed for 30 minin a boiling water bath, and then washed three times withwater. The water suspensions of both radioactive products(total and ANIP) were mounted on Formvar-coated grids, www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.
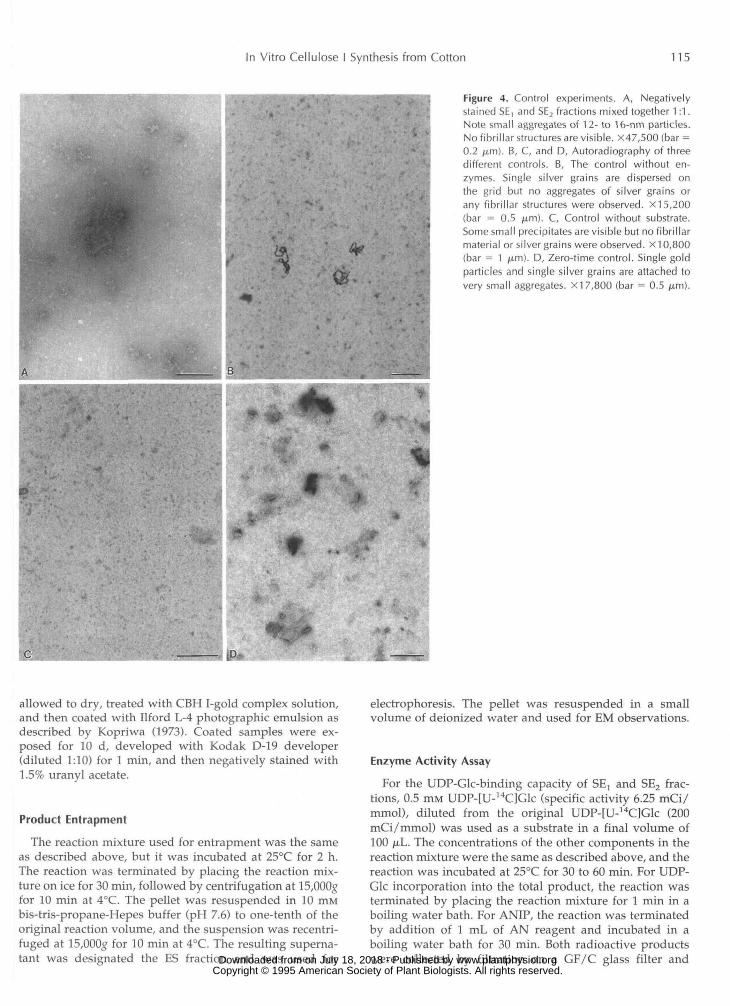
In Vitro Cellulose I Synthesis from Cotton 115
Figure 4. Control experiments. A, Negativelystained SE, and SE2 fractions mixed together 1:1.Note small aggregates of 12- to 16-nm particles.No fibrillar structures are visible. X47,500 (bar =0.2 fj.m). B, C, and D, Autoradiography of threedifferent controls. B, The control without en-zymes. Single silver grains are dispersed onthe grid but no aggregates of silver grains orany fibrillar structures were observed. X15,200(bar = 0.5 fim). C, Control without substrate.Some small precipitates are visible but no fibrillarmaterial or silver grains were observed. XI 0,800(bar = 1 urn). D, Zero-time control. Single goldparticles and single silver grains are attached tovery small aggregates. X17,800 (bar = 0.5 jum).
allowed to dry, treated with CBH I-gold complex solution,and then coated with Ilford L-4 photographic emulsion asdescribed by Kopriwa (1973). Coated samples were ex-posed for 10 d, developed with Kodak D-19 developer(diluted 1:10) for 1 min, and then negatively stained with1.5% uranyl acetate.
Product EntrapmentThe reaction mixture used for entrapment was the same
as described above, but it was incubated at 25°C for 2 h.The reaction was terminated by placing the reaction mix-ture on ice for 30 min, followed by centrifugation at 15,000gfor 10 min at 4°C. The pellet was resuspended in 10 mMbis-tris-propane-Hepes buffer (pH 7.6) to one-tenth of theoriginal reaction volume, and the suspension was recentri-fuged at 15,000g for 10 min at 4°C. The resulting superna-tant was designated the ES fraction and was used for
electrophoresis. The pellet was resuspended in a smallvolume of deionized water and used for EM observations.
Enzyme Activity AssayFor the UDP-Glc-binding capacity of SE, and SE2 frac-
tions, 0.5 mM UDP-[U-14C]Glc (specific activity 6.25 mCi/mmol), diluted from the original UDP-[U-14C]Glc (200mCi/mmol) was used as a substrate in a final volume of100 /nL. The concentrations of the other components in thereaction mixture were the same as described above, and thereaction was incubated at 25°C for 30 to 60 min. For UDP-Glc incorporation into the total product, the reaction wasterminated by placing the reaction mixture for 1 min in aboiling water bath. For ANIP, the reaction was terminatedby addition of 1 mL of AN reagent and incubated in aboiling water bath for 30 min. Both radioactive productswere collected by filtration on a GF/C glass filter and www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.

116 Kudlicka et al. Plant Physiol. Vol. 107, 1995
Figure 5. ED patterns and the irradiated fields ofthe exposed areas selected for ED. A, Defocusedimage of native cotton. B, ED pattern of nativecotton. Note the narrow meridian 004 (a) andthree equatorial reflections, 200, 110, and 110(b), indicating a high degree of orientation andrelatively high crystallinity. The characteristic re-flection 002 (d) for cellulose 1/3 is present. C,Defocused image of ANIP synthesized in vitro bythe SE, fraction. D, ED pattern of in vitro productsynthesized by the SE, fraction. Note the simi-larity of these reflections to those of the nativecellulose I pattern in B.
Figure 6. ED patterns and the irradiated fields ofthe exposed area of the in vitro products treatedfor only 5 min with AN reagent. A, Defocusedimage of the product synthesized by the SE, frac-tion. B, ED pattern of the product synthesized bythe SE, fraction. The cellulose I reflections areobscured by incomplete removal of ^-l,3-glucanafter a 5-min treatment with AN reagent eventhough some hint of cellulose I reflections (arrow-heads) can be seen. C, Defocused image of theproduct synthesized by the SE2 fraction. D, EDpattern of the product synthesized by the SE2
fraction. Note powder pattern with reflectionspartially obscured by incomplete removal ofj3-1,3-glucan, after 5 min of treatment with ANreagent (characteristic for cellulose II).
www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.

In Vitro Cellulose I Synthesis from Cotton 117
Figure 7. Negatively stained and CBH l-gold la-beled in vitro product synthesized by the SE2
enzyme fraction. A, Abundant fibrillar materialarranged linearly or overlapping to form a fi-brous mass not labeled with CBH l-gold com-plex, indicating /3-1,3-glucan chains, and a smalldisorganized aggregate with CBH l-gold particlesattached. X60,300 (bar = 0.1 /xm). B, Highermagnification view of A. A zig-zag structure in-dicative of possible folded glucan chains is visi-ble. XI 37,000 (bar = 100 nm).
washed three times with distilled water (ANIP was firstwashed once with 0.5 N NaOH) and once with methanol.The radioactivity retained on the filters was dissolved in aReady Organic Cocktail (Beckman Instruments, Inc., Ful-lerton, CA) and counted with an LS 6800 Liquid Scintilla-tion System (Beckman Instruments, Inc.).
RESULTS
A comparison of protein profiles and /3-1,4-glucan syn-thase activity was made for the PM, solubilized enzymefractions, and the enzymes released after the entrapment.Figure 1A shows two bands at approximately 60 to 70 kDand four prominent bands (31, 52, 54, and 56 kD) in the PMfraction (lane 2). The same bands were present in the SE2fraction (lane 4). In these two fractions, a 37-kD band isweakly stained. By contrast, in the SE, fraction (lane 3),prominent 37-, 54-, and 56-kD bands were present, whereasthe 52-kD band was diminished. After entrapment purifi-cation from the SE, fraction, a prominent 37-kD polypep-tide band was revealed (Fig. IB, lane 2). This polypeptidewas much weaker after the product entrapment from theSE2 fraction (lane 3). The enzyme activity assay of the PM,SE,, and SE2 fractions revealed that the PM and SE2 en-zyme fractions had higher glucan synthase specific activity,
and each incorporated more radioactive UDP-Glc into thetotal product than did the SE, fraction; however, when theincorporation into ANIP was compared with the totalproduct produced by each fraction, it was evident that theSE, fraction produced the highest amount of ANIP mate-rial (Table I), which is believed to be cellulose (Okuda et al.,1993). Table I shows that about 32.1% of the total productproduced by the SE, fraction consisted of cellulose,whereas with the SE2 enzyme fraction, only about 8.6% ofthe total product was cellulose, compared to 6.6% for cel-lulose in the PM.
Since the total amount of protein in the SE, fraction waslow, both SE, and SE2 fractions were mixed together for thein vitro product synthesis. When a drop of this reactionproduct mixture was placed on an EM grid, reacted withthe CBH I-gold complex, and negatively stained, it wasdifficult to distinguish any differences among the products.The product was very disperse and relatively homoge-neous, and gold particles also were found dispersed in thismaterial (Fig. 2A). When the same product was centrifugedand resuspended in water, it became so strongly aggre-gated with proteins that it was difficult to distinguish bytransmission EM any product labeled with gold particles(data not shown). After this reaction product was treatedwith protease, three different structures could be identified(Fig. 2B): fibrillar material aggregating into long rodlets, www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.

118 Kudlicka et al. Plant Physiol. Vol. 107, 1995
Figure 8. Autoradiography of the in vitro prod-ucts (EIP and ANIP) synthesized by the SE2 frac-tion. A, EIP is very heavily labeled with silvergrains, and CBH l-gold particles are dispersedbetween these grains. X31,700 (bar = 0.5 jam).B, ANIP is more dispersed that EIP, and smallaggregates of product are covered by silvergrains and labeled with CBH l-gold complex.X61,600 (bar = 0.1 urn).
not labeled with CBH I-gold, indicated by "a"; long fibrils,indicated by "b"; and disorganized aggregates, indicatedby "c." The long fibrils and disorganized aggregates wereboth very strongly labeled with CBH I-gold, and theirmorphology indicated that there were two different struc-tural forms of cellulose synthesized.
The incorporation of UDP-[3H]Glc into the in vitro prod-uct synthesized from mixed SE, and SE2 fractions, followedby ethanol treatment, CBH I-gold labeling, autoradiogra-phy, and negative staining, indicated that the CBH I-goldparticles and silver grains both coincided with the EIPproducts, whereas the background is free of labeling (Fig.3, A and B). Similar results were obtained when the radio-active product was treated with the AN reagent, followedby autoradiography, CBH I-gold labeling, and negativestaining (Fig. 3, C and D). In this case, there were twodifferent types of aggregates: (a) long fibrils labeled withCBH I-gold particles and silver grains (Fig. 3C) and (b)disorganized aggregates also strongly labeled with CBHI-gold and silver (Fig. 3D).
To verify that the cellulose in the reaction product wasnot due to contamination, the mixed enzyme fractions wereexamined with transmission EM. No fibrillar material wasobserved (Fig. 4A). Additional evidence that the fibrillarproduct was synthesized in vitro came from autoradio-graphic studies of three controls: (a) all components mixed
together without any of the enzyme fraction added (Fig.4B); (b) all components mixed together but without sub-strate (Fig. 4C); and (c) zero-time reaction with all compo-nents added (Fig. 4D).
In the control without enzyme (Fig. 4B), silver grainswere dispersed on the grids, indicating a typical back-ground of applied radioactive material, but no aggregationof silver grains was observed. In the second control inwhich the substrate was omitted from the reaction mixture(Fig. 4C), it was difficult to find any silver grains at all. Inthe case of the zero-time control (Fig. 4D), single goldparticles from CBH I-gold labeling and single silver grainswere occasionally visible, indicating that the reaction mix-ture placed for 1 min in the boiling water bath did notimmediately block the enzyme activity completely, but thistreatment was sufficient to inhibit the continued synthesisof the product.
EM of the product synthesized in vitro by digitonin-solubilized mixed fractions (SE, plus SE2) clearly indicatedthat there were three different kinds of morphologies in-dicative of j3-l,3-glucans as well as 0-1,4-glucans corre-sponding to cellulose I and cellulose II (Fig. 2B). The rela-tive amounts of these allomorphs were difficult to assess.Only the ratio between (3-1,3- and 0-1,4-glucans could beestimated by UDP-[14C]Glc incorporation into the totalproduct and into ANIP (Table I). In the early course of this www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.

In Vitro Cellulose I Synthesis from Cotton 119
Figure 9. Negatively stained in vitro productsynthesized by the SE, enzyme fraction, treatedwith protease, and labeled with CBH l-goldcomplex. A, Low magnification view showinglong fibrils in the form of linear aggregates, la-beled with CBH l-gold complex embedded in/3-1,3-glucan (dark mass of material), which isnot labeled with CBH l-gold complex, x 10,750(bar = 1 fi.m). B, Higher magnification viewof A. The structures of the /3-1,3-glucans and/3-1,4-glucans are clearly distinguishable.X47,000 (bar 0.2 = /im). C, High magnificationview of the fibrils arranged into a large bun-dle, spanning the background of /3-1,3-glucan.X 107,000 (bar = 0.1 /j.m).
work, it was puzzling why two different morphologicalforms of cellulose were synthesized, whereas in the earlierwork only cellulose II was produced (Okuda et al., 1993).
In an attempt to pinpoint more accurately which extrac-tion protocol gave an active cellulose I assembly, we beganto use the two solubilized fractions independently. It was asurprise to discover that the SE, fraction produced not onlya higher percentage of ANIP (Table I) but also it madecellulose I (Fig. 5D). The SE2 fraction produced more /3-1,3-glucan, and 8.6% of the total product of this fraction wascellulose II (see also Fig. 6D).
The morphology of products synthesized by the separateSE, and SE2 fractions was completely different. Figure 7 Ashows the product synthesized by the SE2 enzyme fraction.The amount of j3-l,3-glucan (material not labeled with CBHI-gold) is much greater than the quantity of cellulose.Higher magnification views of the cellulose produced bythe SE2 fraction (Fig. 7B) show that this product is verysimilar to the cellulose II reported earlier (Okuda et al.,1993).
Autoradiography of the product produced by the SE2fraction shows that EIP was very specifically associatedwith silver grains, and CBH I-gold particles were relativelyhighly dispersed in between grains (Fig. 8A). ANIP pro-
duced by the SE2 fraction showed a small quantity ofdisorganized aggregates abundantly labeled by CBH I-goldand silver grains (Fig. 8B). It was very difficult to localizeany product on the grids for ED studies after a typical30-min boiling with AN reagent; therefore, the treatmenttime was reduced to 5 min only. After a 5-min treatmentwith AN, the recorded ED pattern was similar to the pat-tern of cellulose II, but it was not very clear (Fig. 6D). Theregion from which the ED pattern was recorded shows thatthere is still an abundant quantity of other material present,which probably is /3-1,3-glucan not completely removedafter the short treatment with AN (Fig. 6C). This materialobscured the pattern of cellulose II.
In contrast, the product synthesized by the SEj fractionhad a different morphology (Fig. 9, A-C). Low magnifica-tion views (Fig. 9A) indicated that the ratio of /3-1,3-glucan:/3-1,4-glucan is not as high as that produced by the SE2fraction (Fig. 7A) and that the material associated withCBH I-gold particles had a distinctive fibrillar structureand appeared dispersed among /3-1,3-glucans. Higher mag-nification views of the product showed that the fibrils arevery long, and their dimensions are 1.2 to 1.8 nm (Fig. 9, Band C), slightly larger than the size of native cotton fibrils(not shown). www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.

120 Kudlicka et al. Plant Physiol. Vol. 107, 1995
Figure 10. Autoradiography of the in vitro prod-uct synthesized by the SE, enzyme fraction. A,Low magnification of ANIP. Silver grains andCBH l-gold particles label only the product. Thebackground is free of labeling. X7,100 (bar = 1p.m). B, EIP. In addition to the long fibrils withCBH l-gold particles attached, other material (/3-1,3-glucan) is labeled only by silver grains.X71,000 (bar = 0.1 /Am).
Autoradiography of the product synthesized by the SElfraction after ethanol (Fig. 10B) and AN reagent treatments(Figs. 10A, 11, A and B) showed that in both cases theproduct was very specifically labeled with silver grains andCBH I-gold particles. High magnification of the EIP re-vealed that, in addition to long fibrils with attached CBHI-gold particles, there was also a product not labeled withCBH I-gold, indicating /3-1,3-glucan (Fig. 10B). After ANreagent treatment, only the elongated fibrils remained, alllabeled with CBH I-gold and silver (Figs. 10A and 11, Aand B). The ED pattern of the same ANIP product showedreflections typical of cellulose I (Fig. 5D), comparing favor-ably with the ED pattern of native cotton cellulose (Fig. 5B).
DISCUSSION
Our results show that by a modification of the extractionand solubilization procedures, cellulose I can be synthe-sized in vitro from cotton membrane extracts. Evidencethat the fibrillar material synthesized in vitro from the SEafraction is cellulose I comes from morphological studies ofthe ANIP and from ED analysis. The morphology of theproduct synthesized by this fraction is completely differentfrom the product synthesized by the SE2 fraction. Celluloseproduced by the SEj fraction is in the form of extendedfibrils, which are labeled with CBH I-gold. In contrast, themorphology of cellulose produced from the SE2 fraction is
similar to that of cellulose II reported earlier (Okuda et al.,1993), and it is also similar to the morphology of foldedglucan chains from an A. xylinum mutant, which wereidentified recently as cellulose II (Kuga et al., 1993).
Conclusive evidence for the identity of cellulose from thein vitro product comes from our ED studies. A diffractionpattern of native cotton fibers is shown in Figure 5B. Ac-cording to the indexing convention of Woodcock and Sarko(1980), the following diagnostic reflections of cellulose I areobserved: the 004 meridian reflection (labeled a); threeequatorial reflections, 200, 110, and 110 (labeled b), the 012reflections (labeled c), and the 002 reflection (labeled d).The diffraction pattern of the in vitro product synthesizedby the SE, fraction is shown in Figure 5D and is virtuallyidentical with that of native cotton. Furthermore, a reflec-tion characteristic for cellulose Ij3 is present, which sug-gests that the in vitro cellulose probably contains a fractionof this suballomorph (Sugiyama et al., 1991). Whether cel-lulose Ij3 is enriched over that found in native cellulose isunknown at present; however, these results are not unlikethose obtained from synthetic cellulose I recently producedin this laboratory (Lee et al., 1994).
It is interesting that only the SE, fraction produces cel-lulose I, whereas the SE2 produces cellulose II. This isprobably due to the gentle solubilization process in the firststep, which uses a low concentration of digitonin. In thiscase, the enzyme complex may remain intact, with its www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.
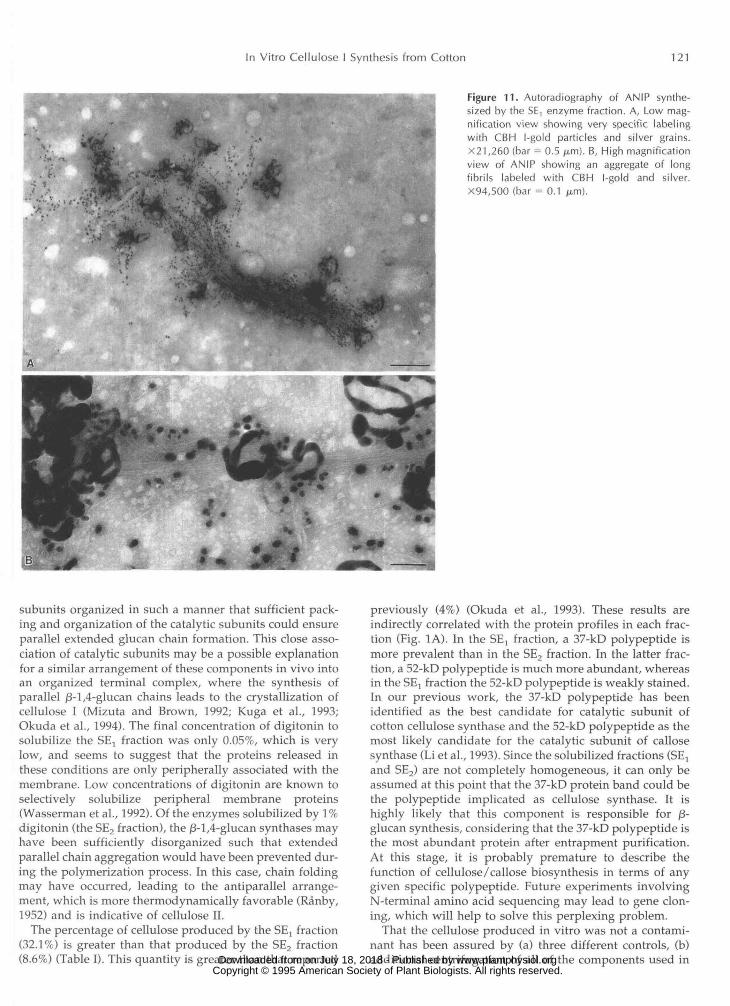
In Vitro Cellulose I Synthesis from Cotton 121
Figure 11. Autoradiography of ANIP synthe-sized by the SE, enzyme fraction. A, Low mag-nification view showing very specific labelingwith CBH l-gold particles and silver grains.X21,260 (bar = 0.5 /j.m). B, High magnificationview of ANIP showing an aggregate of longfibrils labeled with CBH l-gold and silver.X94,500 (bar = 0.1 pm).
subunits organized in such a manner that sufficient pack-ing and organization of the catalytic subunits could ensureparallel extended glucan chain formation. This close asso-ciation of catalytic subunits may be a possible explanationfor a similar arrangement of these components in vivo intoan organized terminal complex, where the synthesis ofparallel /3-1,4-glucan chains leads to the crystallization ofcellulose I (Mizuta and Brown, 1992; Kuga et al., 1993;Okuda et al., 1994). The final concentration of digitonin tosolubilize the SE, fraction was only 0.05%, which is verylow, and seems to suggest that the proteins released inthese conditions are only peripherally associated with themembrane. Low concentrations of digitonin are known toselectively solubilize peripheral membrane proteins(Wasserman et al., 1992). Of the enzymes solubilized by 1%digitonin (the SE2 fraction), the j3-l,4-glucan synthases mayhave been sufficiently disorganized such that extendedparallel chain aggregation would have been prevented dur-ing the polymerization process. In this case, chain foldingmay have occurred, leading to the antiparallel arrange-ment, which is more thermodynamically favorable (Ranby,1952) and is indicative of cellulose II.
The percentage of cellulose produced by the SE! fraction(32.1%) is greater than that produced by the SE2 fraction(8.6%) (Table I). This quantity is greater than that reported
previously (4%) (Okuda et al., 1993). These results areindirectly correlated with the protein profiles in each frac-tion (Fig. 1A). In the SE] fraction, a 37-kD polypeptide ismore prevalent than in the SE2 fraction. In the latter frac-tion, a 52-kD polypeptide is much more abundant, whereasin the SE, fraction the 52-kD polypeptide is weakly stained.In our previous work, the 37-kD polypeptide has beenidentified as the best candidate for catalytic subunit ofcotton cellulose synthase and the 52-kD polypeptide as themost likely candidate for the catalytic subunit of callosesynthase (Li et al., 1993). Since the solubilized fractions (SE]and SE2) are not completely homogeneous, it can only beassumed at this point that the 37-kD protein band could bethe polypeptide implicated as cellulose synthase. It ishighly likely that this component is responsible for )3-glucan synthesis, considering that the 37-kD polypeptide isthe most abundant protein after entrapment purification.At this stage, it is probably premature to describe thefunction of cellulose/callose biosynthesis in terms of anygiven specific polypeptide. Future experiments involvingN-terminal amino acid sequencing may lead to gene clon-ing, which will help to solve this perplexing problem.
That the cellulose produced in vitro was not a contami-nant has been assured by (a) three different controls, (b)additional centrifugation of all of the components used in www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.

122 Kudlicka et al. Plant Physiol. Vol. 107, 1995
the in vitro reaction mixture, and (c) additional centrifuga- tion of enzyme fractions before addition to other compo- rients. No fibrillar structures were observed when UDP-Glc was omitted from the reaction mixture (Fig. 4C) or when enzyme was not added to the other components of the reaction mixture (Fig. 4B). In the zero-time control, single CBH I-gold particles and single silver grains were visible (Fig. 4D), indicating that the reaction mixture placed for 1 min in the boiling water bath did not completely block the enzyme activity immediately, but it was sufficient to pre- vent continued synthesis of product. We were also very careful about any particulate contamination from the air and have taken a11 possible precautions to avoid it. More- over, contamination was ruled out by observing the solu- bilized enzyme fractions with EM, by which means no fibrillar material was ever detected (Fig. 4A).
The addition of 3H-labeled UDP-Glc to the reaction mix- ture, followed by CBH I-gold labeling, autoradiography, and negative staining, shows that the CBH I-gold and silver grains both coincide with the product. The dual labeling provides reasonable evidence that the product is a glucan derived from the supplied UDP-Glc and that this product does not represent any preexisting fibrils in the preparation. Taken together, these observations confirm that cellulose I has been synthesized in vitro by using new extraction and solubilization protocols.
These results validate and extend our previous work (Okuda et al., 19931, which was questioned by Delmer et al. (1993) as being due to contamination and on the basis of a low yield of cellulose. The present work further confirms that cellulose is synthesized in vitro using improved solu- bilization protocols. The percentage of cellulose synthe- sized in vitro has been increased from 4% to at least 32.1% of the total in vitro product. Even this latter figure is conservative, since we were measuring only the ANIP, which does not retain low mo1 wt or poorly crystallized P-1,4-glucans.
Recently, the cellulose I allomorph has been assembled in an abiotic reaction mixture containing cellobiosyl fluo- ride as the substrate and a highly purified endoglucanase as the catalyst (Lee et al., 1994). The reaction proceeds in the presence of acetonitrile and acetate buffer. Presumably, micelles of the catalyst are formed in such a manner that multiple parallel glucan chains are polymerized, which can associate to form extended fibrils. These results comple- ment the findings presented here. It is surprising that the metastable allomorph, cellulose I, has been assembled both in vitro and abiotically when the synthesis of only cellulose I1 had been expected. The digitonin-Mops buffer may serve to produce micelles of the solubilized PM-associ- ated polypeptides that can organize them in such a man- ner as to maintain parallel extended glucan chain poly- merization as a prerequisite for cellulose I synthesis. It is remarkable that, in these two very different reactions in which the substrate and the catalyst are distinct, the crys- tallization mechanisms may be very similar. They pro- vide an additional means for investigating the fundamen- tal mechanisms governing the crystallization of nature's most abundant macromolecule.
ACKNOWLEDCMENTS
We thank Richard Santos for reading the manuscriF t and exten- sive assistance throughout the conduct of the researc I. We thank Henri Chanzy for helpful suggestions concerning EI) interpreta- tion and Dr. Martin Schiilein for the CBH I.
Received July 6, 1994; accepted September 23, 1994. Copyright Clearance Center: 0032-0889/95/l07/0ll1/13.
LITERATURE ClTED
Aloni Y, Cohen R, Benziman M, Delmer DP (1983) Shbilization of the UDP-glucose: 1,4-~-~-glucosyltransferase (c(4lulose syn- thase) from Acetobacter xylinum. J Biol Chem 258: 44194423
Blackwell J (1982) The macromolecular organization of cellulose and chitin. In RM Brown Jr, ed, Cellulose and Other Natural Polymer Sys tems. Biogenesis, Structure, and Degr 3dation. Ple- num Press, New York, pp 403427
Blaschek W, Haass D, Koehler H, Semler U, Franz G (1983) Demonstration of a P-lP-primer glucan in cellulos2-like glucan synthesized in vitro. Z Pflanzenphysiol 111: 357-3154
Brown RM Jr (1989a) Cellulose biogenesis and ,I decade of progress: a personal perspective. In C Schueerch, ed, Cellulose and Wood-Chemistry and Technology. John Wiky and Sons, New York, pp 639-657
Brown RM Jr (1989b) Bacterial cellulose. In JF Kennedy, GO Philips, PA Williams, eds, Cellulose: Structural and Functional Aspects. Ellis Horwood Ltd, London, UK, pp 145-151
Bureau TE, Brown RM Jr (1987) In vitro synthesis cd cellulose I1 from a cytoplasmic membrane fraction of Acetobacter xylinum. Proc Natl Acad Sci USA 8 4 6985-6989
Carpita NC (1982) Cellulose synthesis in detached cotton fibers. In RM Brown Jr, ed, Cellulose and Other Natural Polymer Systems. Biogenesis, Structure and Degradation. Plenum Press, New York, pp 225-242
Chanzy H, Henrissat B, Vuong R (1984) Colloidal gold labelling of 1,4-P-o-glucan cellobiohydrolase adsorbed on cellulose sub- strates. FEBS Lett 172 193-197
Delmer DP (1987) Cellulose biosynthesis. Annu Rev Plant Physiol
Delmer DP (1991) The biochemistry of cellulose synthesis. In CW Lloyd ed, The Cytoskeletal Basis of Plant Growth and Form. Academic Press, London, pp 101-107
Delmer DP, Ohana P, Gonen L, Benziman M (1993) In vitro synthesis of cellulose in plants: still a long way to go! Plant Physiol 103: 307-308
Dhugga KS, Ray PM (1991) A 55 kDa plasma membrime-associaed polypeptide is involved in P-1,3-glucan synthase activity in pea tissue. FEBS Lett 278 283-286
Franz G, Heininger U (1981) Biosynthesis and metabolism: cellu- lose and non-cellulosic cell wall glucans. In FA Loewus, W Tanner, eds, Encyclopedia of Plant Physiology, Vol 13. Springer Verlag, New York, pp 47-67
Frey-Wyssling A (1976) The Plant Cell Wall. Grebriider Born- trager, Stuttgart, Germany, pp 1-294
Kohel RJ (1970) Texas Marker-1. Description of a gertetic standard for Gossypium hirsutum L. Crop Sci 1 0 670-671
Kopriwa BM (1973) A reliable, standardized metl-.od for ultra- structural electron microscopic radioautography. Histochemie
Kuga S , Takagi S, Brown RM Jr (1993) Native foltled-chain cel- lulose 11. Polymer 3 4 3293-3297
Lee JH, Brown RM Jr, Kuga S, Shoda S , Kobayashi. S (1994) The first assembly of synthetic cellulose I. Proc Natl Acad Sci USA
Li L, Brown RM Jr (1993) P-Glucan synthesis in the cotton fiber. 11. Regulation and kinetic properties of P-glucan synthases. Plant Physiol 101: 1143-1 148
Li L, Drake R Jr, Clement S, Brown RM Jr (1493) P-Glucan synthesis in the cotton fiber. 111. Identification of UDP-glucose-
38: 259-290
37: 1-17
91: 7425-7429
www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.

In Vitro Cellulose I Synthesis from Cotton 123
binding subunits of P-glucan synthases by photoaffinity labeling with [p- 3ZP15'-N,-UDP-glucose. Plant Physiol 101: 1149-1 156
Lin FC, Brown RM Jr (1989) Purification of cellulose synthase from Acetobacter xylinum. In C Schuerch, ed, Cellulose and Wood-Chemistry and Technology. John Wiley and Sons, New York, pp 473492
Lin FC, Brown RM Jr, Cooper JB, Delmer D (1985) Synthesis of fibrils in vitro by a solubilized cellulose synthase from Aceto- bacter xylinum. Science 230 822-825
Lin FC, Brown RM Jr, Drake RR Jr, Haley BE (1990) Identification of the uridine 5'-diphosphoglucose (UDP-Glc) binding subunit of cellulose synthase in Acetobacter xylinum using the photoaf- finity probe 5-azido-UDP-Glc. J Biol Chem 265: 47824784
Maclachan G (1982) Does p-glucan synthesis need a primer? In RM Brown Jr, ed, Cellulose and Other Natural Polymer Systems. Plenum Press, New York, pp 327-339
Markwell MAK, Hass SM, Bieber LL, Tolbert NE (1978) A mod- ification of the Lowry procedure to simplify protein determina- tion in membrane and lipoprotein samples. Anal Biochem 87:
Mayer R, Ross I', Weinhouse H, Amikam D, Volman G, Ohana P, Calhoon RD, Wong HC, Emerick W, Benziman M (1991) Polypeptide composition of bacterial cyclic diguanylic acid- dependent cellulose synthase and the occurrence of immunolog- ically cross-reacting proteins in higher plants. Proc Natl Acad Sci USA 88: 5472-5476
Mizuta S, Brown RM Jr (1992) High resolution analysis of the formation of cellulose synthesizing complexes in Vaucheria hamata. Protoplasma 166: 187-199
Okuda K, Li L, Kudlicka K, Kuga S, Brown RM Jr (1993) p- Glucan synthesis in the cotton fiber. I. Identification of P-1,4- and p-1,3- glucans synthesized in vitro. Plant Physiol 101:
Okuda, K, Tsekos J, Brown RM Jr (1994) Cellulose microfibril assembly in Erythrocladia subintegra Rosenv.: an ideal model system for understanding the relationship between synthesizing complexes (TCs) and microfibril crystallization. Protoplasma
Porzio MA, Pearson AM (1977) Improved resolution of myofibril- lar proteins with sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Biochim Biophys Acta 490 27-34
RHnby BG (1952) The mercerisation of cellulose. 111. A phase transition study using electron diffraction. Acta Chem Scand 6
Read SM, Delmer DP (1991) Biochemistry and regulation of cel- lulose synthesis in higher plants. In CH Haigler, P Weimer, eds, Biosynthesis and Biodegradation of Cellulose. Marcel Dekker, New York, pp 177-200
206-210
1131-1142
180 49-58
128-133
Roberts EM, Saxena IM, Brown RM Jr (1989). Biosynthesis of cellulose I1 in Acetobacter xylinum. In C Schuerch, ed, Cellulose and Wood-Chemistry and Technology. John Wiley and Sons, New York, pp 689-704
Robinson DG, Quader H (1981) Structure, synthesis, and orienta- tion of microfibrils. IX. A freeze-fracture investigation of the oocystis plasma membrane after inhibitor treatment. J Cell Biol
Ross P, Weinhouse H, Aloni Y, Michaeli D, Weinberger-Ohana P, Mayer R, Braun S, de Vroom E, van der Marel GA, van Boom JH, Benziman M (1987) Regulation of cellulose synthesis in Acetobacter xylinum by cyclic diguanylic acid. Nature 325:
Sarko A (1978) What is the crystalline structure of cellulose?
Sawyer LH, George W (1982) Comparison between synthetic and natural microfiber systems. rn RM Brown Jr, ed, Cellulose and Other Natural Polymer Systems. Plenum Press, New York, pp 429-453
Saxena IM, Kudlicka K, Okuda K, Brown RM Jr (1994) Charac- terization of genes in the cellulose synthesizing operon (acs operon) of Acetobacter xylinum: implications for cellulose crys- tallization. J Bacteriol 176: 5735-5752
Saxena IM, Lin FC, Brown RM Jr (1990) Cloning and sequencing of the cellulose synthase catalytic subunit gene of Acetobacter xylinum. Plant Mo1 Biol 1 5 673-683
Saxena IM, Lin FC, Brown RM Jr (1991) Identification of a new gene in an operon for cellulose biosynthesis in Acetobacter xyli- num. Plant Mo1 Biol 16: 947-954
Stipanovic AJ, Sarko A (1976) Packing analysis of carbohydrates and polysaccharides. VI. Molecular and crystal structure of re- generated cellulose 11. Macromolecules 9: 851-857
Sugiyama J, Person J, Chanzy H (1991) Combined infrared and electron diffraction study of the polymorphism of native cellu- loses. Macromolecules 2 4 2461-2466
Updegraff DM (1969) Semimicro determination of cellulose in biological materials. Anal Biochem 3 2 420424
Wasserman BP, Wu A, Harriman RW (1992) Probing the molec- ular architecture of (1,3)-P-glucan (callose) synthase: polypep- tide depletion studies. Biochem SOC Trans 2 0 18-22
Woodcock C, Sarko A (1980) Packing analysis of carbohydrates and polysaccharides. 11. Molecular and crystal structure of na- tive ramie cellulose. Macromolecules 9: 1183-1 187
Wong HC, Fear AL, Calhoon RD, Eichinger GH, Mayer R, Amikam D, Benziman M, Gelfand DH, Meade JH, Emerick AW, Bruner R, Ben-Bassat A, Tal R (1990). Genetic organization of the cellulose synthase operon in Acetobacter xylinum. Proc Natl Acad Sci USA 87: 8130-8134
25: 278-288
279-281
TAPPI 61: 59-61
www.plantphysiol.orgon July 18, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.