overlapping and nonoverlapping cortical projections to cortex of the superior temporal sulcus in the...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 370:173-190 (1996)
Overlapping and Nonoverlapping Cortical Projections to Cortex of the Superior
Temporal Sulcus in the Rhesus Monkey: Double Anterograde Tracer Studies
BENJAMIN SELTZER, MONIQUE G. COLA, CAROLINA GUTIERREZ, MICHAEL MASSEE, CHRISTOPHER WELDON, AND CATHERINE G. CUSICK
Departments of Psychiatry and Neurology (B.S.) and Anatomy (B.S., M.G.C, M.M., C.W., C.G.C.) and the Neurosciences Training Program (B.S., C.G., C.G.C.),
Tulane University School of Medicine, Department of Veterans Affairs Medical Center, New Orleans, Louisiana 70112
ABSTRACT To examine how fibers from functionally distinct cortical zones interrelate within their
target areas of the superior temporal sulcus (STS) in the rhesus monkey, separate anterograde tracers were injected in two different regions of the same hemisphere known to project to the STS. Paired injections were placed in dorsal prearcuate cortex and the caudal inferior parietal lobule (IPL), interconnected regions that are part of a hypothesized distributed network concerned with visuospatial analysis or directed attention; in a presumed auditory region of the superior temporal gyrus (STG) and in extrastriate visual cortex, the caudal IPL and lower rim of the intraparietal sulcus; and in dorsal prearcuate cortex and the STG. Overlapping and nonoverlapping projections were then examined in STS visual and polysensory areas.
Prefrontal and parietal fibers directly overlapped extensively in area MST and all subdivisions of presumed polysensory cortex (areas TPOc, TPOi, and TPOr), although nonoverlapping connections were also found. Although STG and IPL fibers targeted all TPO subdivisions, connections were to nonoverlapping, but often adjacent, columns. Paired prefrontal and STG injections revealed largely nonoverlapping vertical columns of connections but substantial overlap within layers VI and I of areas TPOc and TPOi. The findings suggest that area TPO contains differently connected modules that may maintain at least initial segregation of visual versus auditory inputs. Other modules within area TPO receive directly converging input from the posterior parietal and the prefrontal cortices and may participate in a distributed cortical network concerned with visuospatial functions. , 1996 WiIey-I,iss, Inc.
Indexing terms: visual cortex, superior temporal polysensory area, middle temporal area, medial superior temporal area, primates
Cortex of the superior temporal sulcus (STS) in the rhesus monkey has connections with multiple sensory and high-order areas of the cerebral cortex (Jones and Powell, 1970; Seltzer and Pandya, 1978, 1994; Ungerleider and Desimone, 1986; Andersen et al., 1990; Blatt et al., 1990; Boussaoud et al., 1990; Suzuki and Amaral, 1994; Cusick et al., 1995; Schall et al., 1995; Stanton et al., 1995) and contains, within its upper bank, cells that respond to visual, auditory, and somatic sensory stimuli, including some that are activated by two or three types of sensory stimulation (Benevento et al., 1977; Desimone and Gross, 1979; Bruce et al., 1981, 1986; Baylis et al., 1987; Hikosaka et al., 1988; Hietanen and Perrett, 1993; Oram et al., 1993). This region, the “superior temporal polysensory (STP)” cortex,
is presumably concerned, at least in part, with integrating different types of cortical sensory information.
The association of the STS region with complex behavior is supported by observations that experimental lesions, which include the upper bank of the STS result in visual, auditory, somatic sensory, and spatial hemineglect and deficits in hand and eye movements to the contralateral hemispace (Heilman et al., 1970; Luh et al., 1986; Omata et al., 1988; Watson et al., 1994; Scalaidhe et al., 1995). The
Accepted February 7, 1996 Address reprint requests to Catherine G. Cusick, Ph.D., Department of
Anatomy, Tulane University School of Medicine, 1430 Tulane Avenue, New Orleans, LA 70112.
c 1996 WILEY-LISS, INC.

174 B. SELTZER ET AL.
Fig. 1. Overlapping and nonoverlapping label within the superior temporal sulcus (STS) cortex following tracer deposits in the prefrontal ([”SI-methionine) and posterior parietal (wheatgerm agglutinin- horseradish peroxidase; WGA-HRP) cortex in case TMQ. The loca- tions of injections are shown in black on the lateral view of the brain surface. On the unfolded map of STS cortex, horizontal hatching represents label from the prefrontal injection, vertical hatching denotes label from the posterior parietal injection, and cross hatching denotes overlapping label from both sites. The black region represents slight spread of the WGA-HRP injection and its halo (shading) to the caudal upper bank of the STS. The long dashed line represents the midpoint of
upper bank of the sulcus has reciprocal connections with the dorsal prearcuate cortex of the frontal lobe and the caudal inferior parietal lobule (IPL; Jones and Powell, 1970; Seltzer and Pandya, l984,1989a, 1994; Selemon and Goldman-Rakic, 1988; Andersen et al., 1990), in which lesions also impair awareness of and motor interactions with the contralateral extrapersonal hemispace (Lynch, 1992; Funahashi et al., 1993; Goldman-Rakic et al., 1992; Milner and Goodale, 1993). Thus, the upper bank of the STS may be part of a hypothesized hemisphere wide
the depth of the STS. The shorter, horizontally oriented dashed line separates the middle temporal area (MT) from the dorsally adjacent region thought to contain its peripheral representation (MTp: Unger- leider and Desimone, 1986). FST, fundal superior temporal area: MST, medial superior temporal area; V4t, transitional part of V4; IPa, area of fundal cortex from Seltzer and Pandya (1978), as revised by Cusick et al. (1995). TPOc, TPOi, and TPOr, architectonically determined subdi- visions of area TPO (Cusick et al., 1995). The short numbered vertical lines indicate the locations of sections shown in Figures 2-4; the plane of section is approximately the same as the vertical hatching.
distributed network of cortical areas concerned with mecha- nisms controlling attention and exploratory movements (e.g., Luh et al., 1986; Goldman-Rakic, 1988; Selemon and Goldman-Rakic, 1988; Goldman-Rakic and Friedman, 1991; Mesulam, 1990; Seltzer, 1992; Morecraft et al., 1993).
In addition to several well-characterized extrastriate visual areas, the upper bank of the STS contains a distinct architectonic zone, termed “area TPO,” that presumably corresponds in large part to the STP region (Cusick et al., 1995). Area TPO has exceptionally diverse cortical connec-

OVERLAP AND NONOVERLAP OF STS AFFERENTS 175
tions, including input from auditory, visual, and, possibly, somatic sensory areas (Seltzer and Pandya, 1978, 1984, 1991, 19941, and receives “ascending” and ‘‘lateral’’ inputs from the posterior parietal and prefrontal cortices, respec- tively (Felleman and Van Essen, 1991; Cusick et al., 1995). Architectonic and connectional studies suggest a further division of area TPO into at least three different rostrocau- dal zones (Seltzer and Pandya, 1989b, 1994; Cusick et al., 1995).
Previous studies of the connections of STS cortex typi- cally involved series of tracer experiments in different animals so that data directly comparing different afferent projections in the same hemisphere are lacking. Because the sulcal configuration differs from case to case, the precise relation among different afferent fiber systems remains unknown. Yet, different connectional relations have poten- tially different functional consequences for how informa- tion from separate cortical sources is integrated within the STS. The strategy of using two different anterograde tracers, e.g., radiolabeled amino acids and wheatgerm agglu- tinin conjugated to horseradish peroxidase (WGA-HRP), in a single hemisphere permits a direct comparison of the areal and laminar distribution of termination of two differ- ent corticocortical pathways. Such an approach was used by Selemon and Goldman-Rakic (1988) to contrast the projec- tions of the posterior parietal and prefrontal cortex in subcortical and cortical areas, including portions of the STS.
In the present study, separate anterograde tracers were injected into two different areas of the same hemisphere to study patterns of overlap and nonoverlap of different cortical pathways targeting the STS. To investigate the position of STS cortex in the hypothesized distributed network of cortical areas pertaining to attention, we com- bined injections in the dorsal prearcuate and caudal IPL cortex. To examine the morphological basis for potential sensory-sensory integration in STP cortex, an injection of the superior temporal gyrus (STG), a presumed unimodal auditory area, was paired with one in the extrastriate visual cortex of the caudal IPL and adjacent lower rim of the intraparietal sulcus (IPS). An injection within the dorsal prearcuate cortex was paired with one in the STG to compare sites within the STS participating in presumed higher-order versus presumed unimodal sensory circuits.
METHODS Five rhesus monkeys (Macaca rnulatta) received injec-
tions of different anterograde tracers in two separate cortical areas of the same hemisphere known from previous studies to project to cortex of the STS. Followingpremedica- tion with ketamine (10 mgikg, i.m.), the animals were anesthetized by a mixture of isoflurane and nitrous oxide and continuously monitored throughout the procedure so that heart rate, expired COz, and body temperature re- mained within physiological limits for surgical anesthesia. Using aseptic surgical technique, 7-15 injections of 0.3 p1 tritiated proline and/or leucine (50-75 kCi/pl) were made into one cortical region, and, 7-8 days later, 6-12 injections of approximately 0.25 k1 of 1.0% WGA-HRP were placed in another zone of the same hemisphere in three monkeys. In two other cases, similar WGA-HRP injections in one corti- cal region were combined, a t the same operation, with 5 or 8 injections of 0.2 p1 [WJ-methionine (200 kCi/kl) in an- other region of the same hemisphere.
. . A
B
TMQ 524 - 1 rnrn
Fig. 2. Overlapping and nonoverlapping parietal (HRP-labeled) and prefrontal (autoradiographically labeled) fibers within the STS at the levels of area TPOc (A,B) and caudal STS visual areas (C,D) in case TMQ (Fig. 1). Camera lucida drawings of WGA-HRP (HRP) and autoradiographic (AFtG) label from single sections that were first treated for HRP histochemistry and then exposed to X-ray film. The outline and fiducial artifacts were first drawn from the section under darkfield optics. This outline was then used independently to chart each pattern of label. A,C: For WGA-HRP label, cells are depicted as larger dots, and stippling indicates terminals. B and D show ARG-labeled terminals. Asterisks mark some of the overlapping patches of fibers labeled by the two methods. Arrows indicate architectonic borders that were transferred, also by camera lucida, from adjacent sections. Thin lines within cortex denote layer IV, and small ovals denote blood vessels used for reference. Sections are numbered consecutively from caudal to rostral. The numbered section levels for Figures 2-4 are indicated on Figure 1.
Forty-eight to 72 hours following the last injection, the animals were euthanatized by an intravenous overdose of sodium pentobarbital and perfused with saline followed by 1.5 liters of 4% paraformaldehyde in 0.1 M phosphate buffer, and, finally, 0.5 liter of the same solution with 10% sucrose added. The brains were then stored in 30% buffered sucrose

176 B. SELTZER ET AL.
TMQ 654 n
k Prefrontal injection
Fig. 3. Overlapping and nonoverlapping parietal (A$) and prefron- tal (B,D) projections shown on the same section, at the level of TPOi and MST, in case TMQ (Fig. 1). Conventions as in Figure 2. Asterisks mark a few sites in which the two labels interdigitate rather than
A
b
C
overlap in layer IV. In this and in subsequent figures, gaps placed in the outline of layer IV mark where foci of label cross the border of layer IV on the other drawing of the pair.
. .
Fig. 4. Overlapping and nonoverlapping parietal (A,C) and prefrontal (B,D) fibers shown on the same section, at the level of TPOr and IPa in case TMQ (Fig. 1). Asterisks in A and B mark some of the sites labeled in layer IV by one method only (nonoverlapping label).
solution before being cut coronally on a freezing microtome at intervals of 25-50 pm. Parallel series of closely spaced sections were processed for autoradiography (ARG; Cowan et al., 19721, WGA-HRP neurohistochemistry (Mesulam, 19781, cytoarchitecture (Nissl), myeloarchitecture (Gallyas, 19791, and chemoarchitecture. For chemoarchitectonic analyses of cortical architecture, the calcium-binding pro- tein parvalbumin was localized immunocytochemically by
using a monoclonal antibody (Sigma) and standard avidin- biotin peroxidase complex (ABC) methods (Vector; Hsu et al., 1981); another series of sections was immunostained for a nonphosphorylated form of neurofilament protein utiliz- ing the SMI-32 antibody (Sternberger Monoclonals, Inc.; Campbell and Morrison, 1989; Hof and Morrison, 1995). Details of these procedures are presented elsewhere (Cusick et al., 1995). A third series was prepared for cytochrome
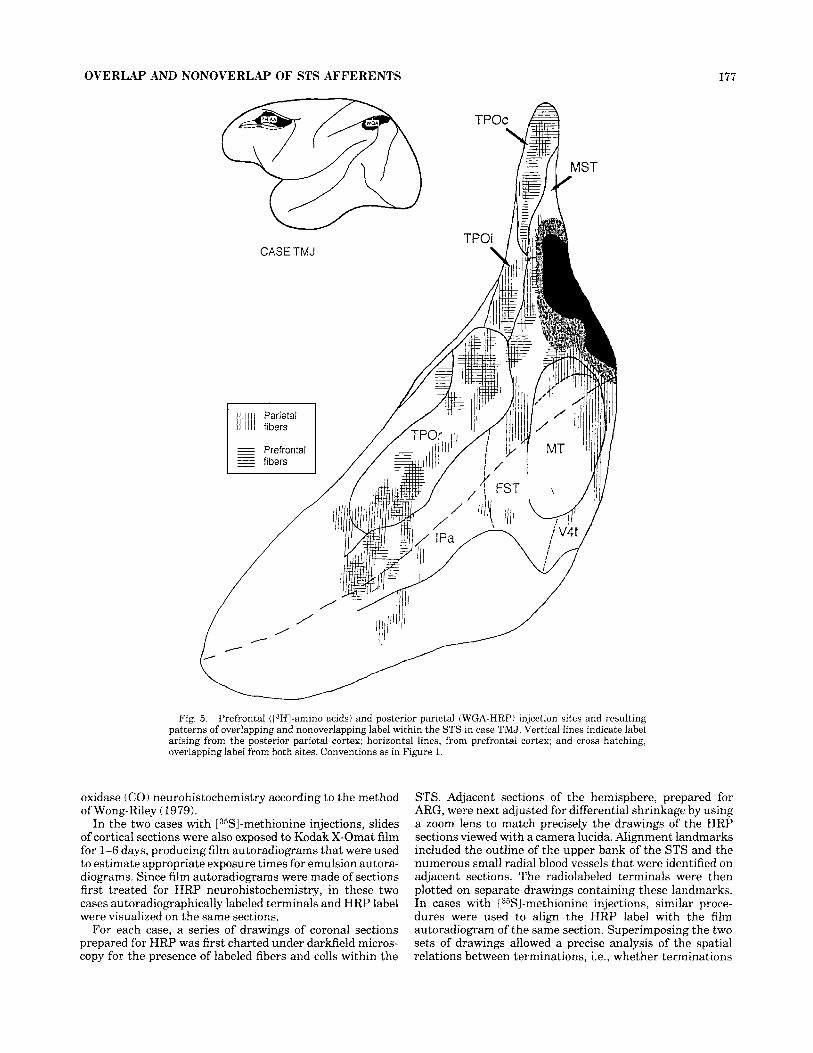
OVERLAP AND NONOVERLAP OF STS AFFERENTS 177
CASE TMJ
Fig. 5. Prefrontal (I3HI-amino acids) and posterior parietal (WGA-HRP) injection sites and resulting patterns of overlapping and nonoverlapping label within the STS in case TMJ. Vertical lines indicate label arising from the posterior parietal cortex; horizontal lines, from prefrontal cortex; and cross hatching, overlapping label from both sites. Conventions as in Figure 1.
oxidase (CO) neurohistochemistry according to the method of Wong-Riley (1979).
In the two cases with [35SJ-methionine injections, slides of cortical sections were also exposed to Kodak X-Omat film for 1-6 days, producing film autoradiograms that were used to estimate appropriate exposure times for emulsion autora- diograms. Since film autoradiograms were made of sections first treated for HRP neurohistochemistry, in these two cases autoradiographically labeled terminals and HRP label were visualized on the same sections.
For each case, a series of drawings of coronal sections prepared for HRP was first charted under darkfield micros- copy for the presence of labeled fibers and cells within the
STS. Adjacent sections of the hemisphere, prepared for ARG, were next adjusted for differential shrinkage by using a zoom lens to match precisely the drawings of the HRP sections viewed with a camera lucida. Alignment landmarks included the outline of the upper bank of the STS and the numerous small radial blood vessels that were identified on adjacent sections. The radiolabeled terminals were then plotted on separate drawings containing these landmarks. In cases with [35SJ-methionine injections, similar proce- dures were used to align the HRP label with the film autoradiogram of the same section. Superimposing the two sets of drawings allowed a precise analysis of the spatial relations between terminations, i.e., whether terminations
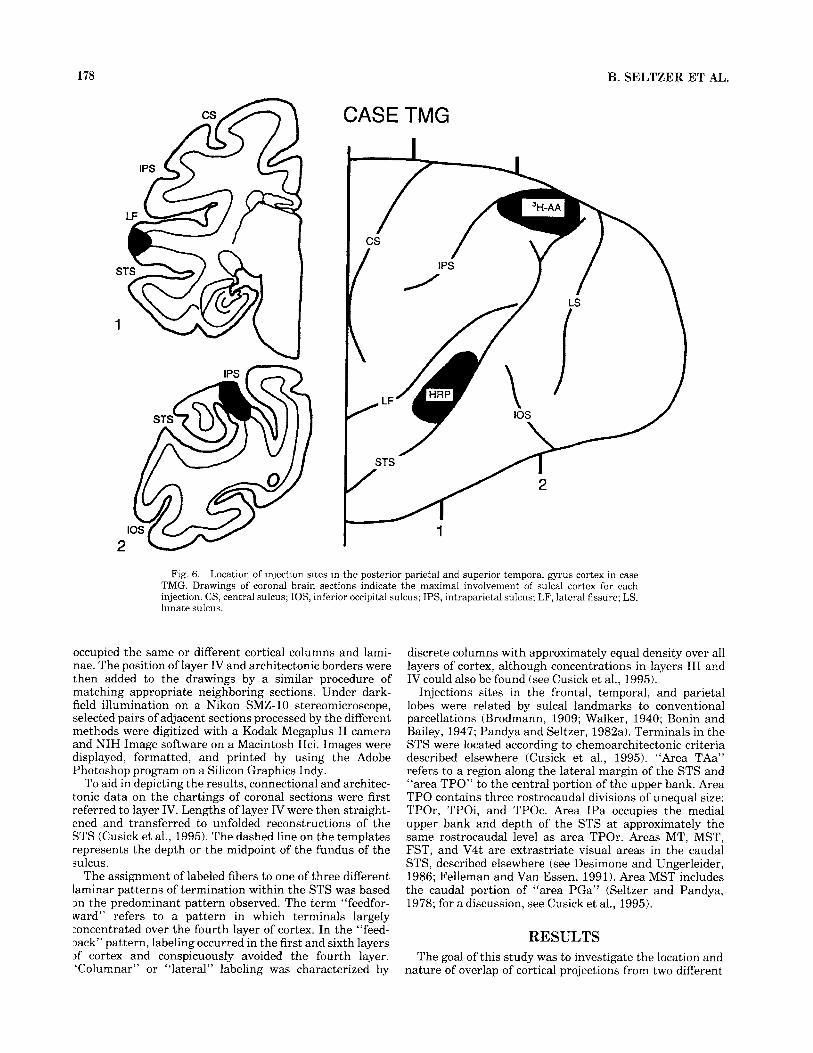
178 B. SELTZER ET AL.
CASE TMG
1
Fig. 6. Location of injection sites in the posterior parietal and superior temporal gyrus cortex in case TMG. Drawings of coronal brain sections indicate the maximal involvement of sulcal cortex for each injection. CS, central sulcus; IOS, inferior occipital sulcus; IPS, intraparietal sulcus; LF, lateral fissure; LS, lunate sulcus.
occupied the same or different cortical columns and lami- nae. The position of layer IV and architectonic borders were then added to the drawings by a similar procedure of matching appropriate neighboring sections. Under dark- field illumination on a Nikon SMZ-10 stereomicroscope, selected pairs of adjacent sections processed by the different methods were digitized with a Kodak Megaplus I1 camera and NIH Image software on a Macintosh IIci. Images were displayed, formatted, and printed by using the Adobe Photoshop program on a Silicon Graphics Indy.
To aid in depicting the results, connectional and architec- tonic data on the chartings of coronal sections were first referred to layer IV. Lengths of layer IV were then straight- ened and transferred to unfolded reconstructions of the STS (Cusick et al., 1995). The dashed line on the templates represents the depth or the midpoint of the fundus of the sulcus.
The assignment of labeled fibers to one of three different laminar patterns of termination within the STS was based 3n the predominant pattern observed. The term “feedfor- ward” refers to a pattern in which terminals largely :oncentrated over the fourth layer of cortex. In the “feed- back” pattern, labeling occurred in the first and sixth layers 3f cortex and conspicuously avoided the fourth layer. ‘Columnar” or ‘‘lateral’’ labeling was characterized by
discrete columns with approximately equal density over all layers of cortex, although concentrations in layers I11 and IV could also be found (see Cusick et al., 1995).
Injections sites in the frontal, temporal, and parietal lobes were related by sulcal landmarks to conventional parcellations (Brodmann, 1909; Walker, 1940; Bonin and Bailey, 1947; Pandya and Seltzer, 1982a). Terminals in the STS were located according to chemoarchitectonic criteria described elsewhere (Cusick et al., 1995). “Area TAa” refers to a region along the lateral margin of the STS and “area TPO” to the central portion of the upper bank. Area TPO contains three rostrocaudal divisions of unequal size: TPOr, TPOi, and TPOc. Area IPa occupies the medial upper bank and depth of the STS at approximately the same rostrocaudal level as area TPOr. Areas MT, MST, FST, and V4t are extrastriate visual areas in the caudal STS, described elsewhere (see Desimone and Ungerleider, 1986; Felleman and Van Essen, 1991). Area MST includes the caudal portion of “area PGa” (Seltzer and Pandya, 1978; for a discussion, see Cusick et al., 1995).
RESULTS The goal of this study was to investigate the location and
nature of overlap of cortical projections from two different

OVERLAP AND NONOVERLAP OF STS AFFERENTS
CASE TMG
179
n
Fig. 7. Largely nonoverlapping patterns of label within the STS resulting from injection sites in the posterior parietal (13HJ-amino acids; vertical hatching) and superior temporal gyrus cortex (WGA- HRP; horizontal hatching) in case TMG (Fig. 6). Architectonic area
sources in architectonic areas of the STS. When two projection systems target the same cortical area, two broad patterns are possible: (1) the differently labeled afferents may remain widely separated from each other (“topo- graphic” pattern), or (2) they may terminate in close proximity to each other. Within the second pattern, the different afferents may interleave with each other without actually overlapping (“interdigitated” pattern), occupy ar- chitectonically distinct subfields, or be directly superim- posed (“overlapping” pattern). If the differently labeled fibers target the same cortical column, they may occupy either the same or different (“complementary”) cortical layers. “Mixed” patterns, with overlap in certain cortical layers and interdigitation in others, are another possibility.
TAa, terminology of Seltzer and Pandya (1978). DMZ, densely myelin- ated zone (Ungerleider and Desimone, 1986). Numbered vertical lines refer to coronal sections shown in Figures 8 and 9.
Because of the complexity of the detailed relations be- tween two neighboring terminal fields, the data are dis- played as unfolded reconstructions and as pairs of coronal sections at representative levels. In Figure 1 and in subse- quent figures showing unfolded reconstructions, regions of the STS are depicted as having overlapping projections (cross hatching) as follows: (1) for two ‘‘lateral’’ or “feedfor- ward” projections, if the overlap occurred in layers 111 and IV; (2) for two “feedback” connections, if direct overlap was found over layers I and VI; and (3) for projections to complemen- tary “feedforward” and “feedback” connections, if the overlapping connections targeted the same radial columns of cortex. For clarity, mixed patterns, with overlap in layers I and VI and nonoverlap in layers 111 and IV, are

180 B. SELTZER ET AL.
A C
STG injection
Fig. 8. Largely nonoverlapping temporal (HRP-labeled) and parietal (AFGlabeled) fibers shown on sections of the STS separated by 25 pm at the levels of areas TPOc (A,B) and TPOi (C,D) in case TMG (Fig. 6) . Asterisks mark sites of largely nonoverlapping label.
shown as nonoverlapping. The camera lucida drawings of coronal sections provide the most accurate information on the precise spatial relations between two cortical pathways.
Frontal and parietal injections Two monkeys (cases TMQ and TMJ) had paired injec-
tions of anterograde tracers in portions of the prefrontal and posterior parietal cortices of the same hemisphere. Coronal sections illustrating the injection sites are pre- sented in a previous study (Cusick et al., 1995). The areas injected are known from previous studies to project to polysensory and extrastriate areas of the STS and to each other (e.g., Seltzer and Pandya, 1978, 1984, 1989a, 1994; Cavada and Goldman-Rakic, 1989; Andersen et al., 1990; Schall et al., 1995; Stanton et al., 1995).
In case TMQ (Fig. I), the frontal lobe injection of [35SJ-methionine involved most of the cortex between the upper bank of the principal sulcus and the lower bank of the upper spur and concavity of the arcuate sulcus (dorsal area 46 and areas 8a and 8b). The WGA-HRP injection involved the apex of the IPL (area 7a or Opt of Pandya and Seltzer, 1982a), from which there was slight spread into the ex- treme caudal depth of the STS. As predicted, the frontal injection produced terminal label in the posterior parietal cortex (area 7a and LIP), whereas the parietal injection resulted in terminal label in the depth and upper bank of, and cortex immediately dorsal to, the principal sulcus (dorsal area 46 and area 8b) and the rostra1 bank of the arcuate sulcus (area 8a).
Within the STS, the following architectonic areas re- ceived projections from the prefrontal (Fig. 1, horizontal lines) and posterior parietal (Fig. 1, vertical lines) cortex: TPOc, TPOi, TPOr, V4t, MT, MST, FST, and IPa. In area TPOc, there was a precise correspondence between the sites of termination of ARG- and HRP-labeled fibers (Figs. 1, 24B). Parietal projections were focused on layer IV, and prefrontal terminations tended to be of the “columnar type” through all
cortical layers. Both projections formed patches of fibers in layers I-IV, and these patches directly overlapped each other. There was also widespread overlap in layer VI. In area TPOi, ARG- and HRP-labeled fibers were arranged in patches in layers I11 and IV that largely overlapped but sometimes interdigitated (Fig. 3). Overlapping projections were simultaneously found in layers VI and I. Finally, area TPOr displayed overlap (e.g., Fig. 4C,D) and interdigitation (Fig. 4A,B) of fibers from the frontal and parietal lobes terminating in layers I11 and IV. Widespread overlap was also found in layers VI and I.
Within areas V4t, MT, and FST, radiolabeled and HRP- labeled fibers were often found to be segregated from each other, displaying the “topographic” pattern. At certain sites within areas MT and V4t, however, frontal and parietal terminals directly overlapped each other in the first and sixth cortical layers (see Fig. 2C,D). Considerable overlap also occurred in area MST. As might be expected, where both projection systems terminated largely in a feedback pattern, frontal- and parietal-derived fibers over- lapped in layers I and VI (Fig. 2C,D).
Caudodorsally, in area MSTd (Komatsu and Wurtz, 1988) the pattern of overlap was “complementary,” with parietal-derived terminals clustered in and around layer IV (Fig. 2A) and frontal fibers focused on layers I and VI (Fig. 2B). A focus of overlap in area IPa also had a complemen- tary pattern. Here parietal fibers showed a feedforward pattern, whereas frontal fibers were concentrated in layers I and VI (Fig. 4C,D).
In case TMJ (Fig. 51, tritiated amino acids were injected into the frontal lobe above the caudal half of the principal sulcus and below the upper segment of the arcuate sulcus (dorsal area 46). The injection did not extend as far caudoventrally as that in case TMQ. The WGA-HRP injec- tion in TMJ involved the caudal one-third of the IPL (area 7a), with somewhat more spread into the caudal upper bank of the STS (area MST) than in case TMQ. On the lateral

-
A 1 m
rn
287
B
IPL
inje
ctio
r I
Fig.
9.
Lar
gely
non
over
lapp
ing
tem
pora
l (A
,C; H
RP-
labe
led)
and
par
ieta
l (B
,D; A
RG
- in
A a
nd B
poi
nt t
o si
tes
of n
onov
erla
ppin
g la
bel,
i.e.,
to u
nlab
eled
int
erva
ls o
f la
yer
IV t
hat
labe
led)
fibe
rs s
how
n on
dir
ectly
adj
acen
t sec
tions
of t
he S
TS,
at t
he le
vel o
f are
a T
POr,
in c
ase
cont
ain
labe
l on
the
othe
r dra
win
g of
the
pair
. TM
G (
Fig.
6).
In A
and
C, t
he S
TG
inje
ctio
n is
sho
wn
in b
lack
, and
its
halo
is s
tippl
ed. A
ster
isks

182 B. SELTZER ET AL.
Fig. 10. Case TMG: Digitized microscopic images of largely nonoverlapping (A) temporal (HRP-labeled) and (B) parietal (ARG-labeled) fibers shown on directly adjacent sections of the STS at the level of area TPOr. The plane of cut is slightly nonradial. Thus, the medial nonoverlapping sets of HRP and ARG label are actually in different cortical columns. Arrows mark the same blood vessels. Scale bar = 1 mm.
surface of the parietal lobe, radiolabeled fibers were identi- fied in the caudal parietal operculum and lower bank of the IPS, including the caudal IPL. HRP-labeled fibers were found in the prefrontal cortex extending from the caudal principal sulcus to the upper segment of the arcuate sulcus (dorsal area 46 and areas 8a and 8b). These observations confirmed that the two injection sites were interconnected.
Within the STS, frontal and parietal fibers targeted areas TPOc, TPOi, TPOr, MST, and IPa (Fig. 5 ) . The spatial relations between the two afferent systems in area TPO of this case were very similar to those demonstrated for case TMQ. Area TPOc contained several sites where columns of frontal- and parietal-derived afferent fibers directly over- lapped each other. In area TPOi, columns of radiolabeled fibers often tightly interdigitated feedforward columns of
HRP-labeled fibers, but sites of direct overlap between the differently labeled fibers also occurred in layers I11 and IV. Within area TPOr, several foci of panlaminar overlap were found. The pattern of overlap in areas MST and IPa also resembled that seen in case TMQ. In area MST, to which the prefrontal and posterior parietal cortices projected in feedback fashion, there was a small focus of overlap over layers I and VI. In area IPa, frontal fibers occupied the first and sixth cortical layers, and parietal fibers occupied the fourth.
Parietal and temporal injections Two monkeys had paired injections in the IPL and STG,
regions known to project to the STS (e.g., Seltzer and

OVERLAP AND NONOVERLAP OF STS AFFERENTS 183
CASE TMH I
Fig. 11. Location of injection sites in the posterior parietal and superior temporal gyrus cortex in case TMH. Conventions as in Figure 6 .
Pandya, 1978, 1984, 1994; Galaburda and Pandya, 1983; Andersen et al., 1990). The former is considered an extrastri- ate visual area (e.g., Felleman and Van Essen, 1991), the latter, an auditory-related region of the hemisphere (Gala- burda and Pandya, 1983; Morel et al., 1993). As expected from previous studies (Pandya and Sanides, 1973; Seltzer and Pandya, 1984; Cavada and Goldman-Rakic, 1989; Andersen et al., 1990; Morel and Bullier, 1990; Baizer et al., 1991), the injection sites did not significantly connect with each other.
Case TMG had paired injections of tritiated amino acids in the caudal IPL and of WGA-HRP in the middle and caudal STG (Fig. 6). The parietal injection extended into the lower bank of the IPS (area LIP) and caudalmost upper bank of the STS (area MST); the temporal injection spread slightly into the upper rim of the STS (area TAa). The parietal injection in this experiment and in case TMH (see below) did not produce label in areas MT or V4t. The variable results from parietal injections may be due to differences in sensitivity of the respective techniques used (L3H]-amino acids in cases TMG and TMH vs. WGA-HRP in cases TMQ and TMJ) or to differences in the extent of the caudal STS included in the injection sites.
Both injections produced label in area TPOc near the border with area TPOi, and in areas TPOi and TPOr (Fig. 7). At most sites within area TPO, the majority of IPL- derived projections tended to lie medial to those emanating
from the STG (Figs. 7-9), and the most lateral portion of area TPO tended to receive fibers from the STG but not the IPL (Figs. 8, 9). Occasionally, the differently labeled fibers lay close to each other, and, although columns of STG- derived fibers interdigitated with those coming from the parietal lobe, they did not actually overlap (Figs. 8-10). At certain individual levels in TPOr (e.g., Fig. 9A,B), the HRP-labeled fibers appeared to occupy deeper layers of cortex, and radiolabeled fibers were more superficial, but analysis of serial sections showed that this apparent “over- lap” was due to a nonradial plane of cut and that the two afferent systems were actually segregated into adjacent nonoverlapping columns.
Case TMH had an injection of tritiated amino acids involving approximately the caudal half of the IPL (area 7a) and of WGA-HRP involving the mid-STG (Fig. 11). There was minimal involvement of cortex in the caudalmost upper bank (area MST) and mid-upper rim (area TAa) of the STS by the amino acid and HRP injections, respectively. Within the STS, both projection systems were directed to areas TPOi and TPOr (Fig. 12). Although they sometimes inter- digitated, as in case TMG, they remained separate from each other and did not overlap.
Prefrontal and temporal injection Another monkey (case TMP) had an injection of [W]-
methionine in the prefrontal cortex (caudodorsal area 46

184 B. SELTZER ET AL.
CASE TMH
Fig. 12. Largely nonoverlapping patterns of label within the STS resulting from injection sites into the posterior parietal ([?HI-amino acids; vertical hatching) and superior temporal gyrus cortex (WGA-HRP; horizontal hatching) in case TMH (Fig. 11). Architectonic area Tpt, terminology of Pandya and Sanides (1973).
and area 8a) combined with an injection of WGA-HRP into the caudal STG (areas Ts3, paAlt, and, to a lesser extent, Tpt; Pandya and Sanides, 1973) (Fig. 13). Coronal sections of these injection sites are shown in Cusick et al. (1995). The latter injection spread slightly into cortex along the upper rim of the STS (area TAa) and involved subjacent white matter. Although prefrontal and temporal areas injected are known to project to the STS (e.g., Seltzer and Pandya, l978,1989a, 19941, they have comparatively sparse interconnections with each other (reviewed by Pandya and Yeterian, 1990). Indeed, the frontal injection produced only a few labeled terminals in the most caudal segment of the STG (area Tpt) posterior to the site of the HRP injection;
and the STG injection produced minimal label in the extreme rostra1 upper bank of the principal sulcus.
As expected, extrastriate visual areas V4, V4t, MT, MST, and FST received projections from the prefrontal injection alone, and area TAa only from the STG (Fig. 13). By contrast, both injections produced labeled terminals within areas TPOr, TPOi, and TPOc of the STS (Figs. 13-16). The presence of STG-derived fibers within TPOc in cases TMP and TMG, and their absence in case TMH, may be a consequence of differences in rostrocaudal placement of the injection sites. The ARG- and HRP-labeled fibers in case TMP often lay immediately adjacent to each other (e.g., TPOc, Fig. 14; TPOr, Fig. 161, but, for the most part,

OVERLAP AND NONOVERLAP OF STS AFFERENTS
404
185
Fig. 13. Overlapping and largely nonoverlapping (in layer IV) patterns of label within the STS resulting from injection sites into the prefrontal (I”Sl-methionine; vertical lines) and superior temporal gyrus cortex (WGA-HRP; horizontal lines) in case TMP. Numbered vertical lines refer to section levels in Figures 14-16.
connections were focused on nonoverlapping segments of layer IV and were thus interpreted as showing segregated “feedforward” connections (Figs. 13-16). At a number of sites, however, there was extensive overlap over layers VI and I (Figs. 14, 15).
DISCUSSION The double anterograde tracer stratem employed in this
study showed that selected frontal, temporal, and parietal projections interrelate in several different ways within area TPO, which presumably corresponds to the STP region (Desimone and Gross, 1979; Bruce et al., 1981; Baylis et al.,
1987; Hikosaka et al., 1988). Prefrontal and parietal, i.e., caudal IPL, fibers overlapped in all three divisions of area TPO, with the differently labeled columns of fibers directly superimposed upon each other. At some sites, the parietal and prefrontal overlap involved nearly the entire vertical extent of a given cortical column, including layer IV (e.g., in areas TPOc and TPOr; Figs. 2, 4). Thus, prefrontal and parietal afferents may provide presumably excitatory driv- ing inputs to the same cortical circuits within area TPO.
By contrast, parietal and temporal fibers were noted to interleave each other within different subdivisions of area TPO (e.g., Fig. 91, but there was little direct overlap of these projections. Although frontal and temporal fibers

186
.\ TMP404 q i
B. SELTZER ET AL.
Prefrontal injection
TPOc
c
D
I * TPOc Fig. 14. A-D: Overlapping and nonoverlapping temporal (HRP-labeled) and prefrontal ( AFLG-labeled)
fibers shown on the same section of the STS at the level of TPOc and caudal STS visual areas in case TMP (Fig. 13). The pattern ofoverlap in area TPOc varies by layer and is termed “mixed.” Asterisks mark sites of nonoverlap in layer IV.
tended to remain segregated from each other in area TPO, there were several sites where adjacent columns of fibers coming from the two different cortical areas overlapped extensively in layer VI, while, in the same region, alternat- ing in layers I11 and IV (e.g., Figs. 14C,D, 15). Thus, whereas connections in layers I and VI were widespread and xerlapping, prefrontal and STG fibers in layer IV may be Sistributed mainly to different anatomical modules of area r p o .
Although principally focused on the STP region, the present study also examined potential overlap in extrastri- ate visual areas of the STS following the relevant parietal and prefrontal injections. In portions of area MT, in which feedback terminations from prefrontal cortex and sparse connections from IPL were found in one case, there was both overlap and topographic segregation of fibers (Fig. 11, but at other sites, notably MST and MTp (Fig. 1) and IPa (Fig. 5), overlap appeared to predominate. The overlapping
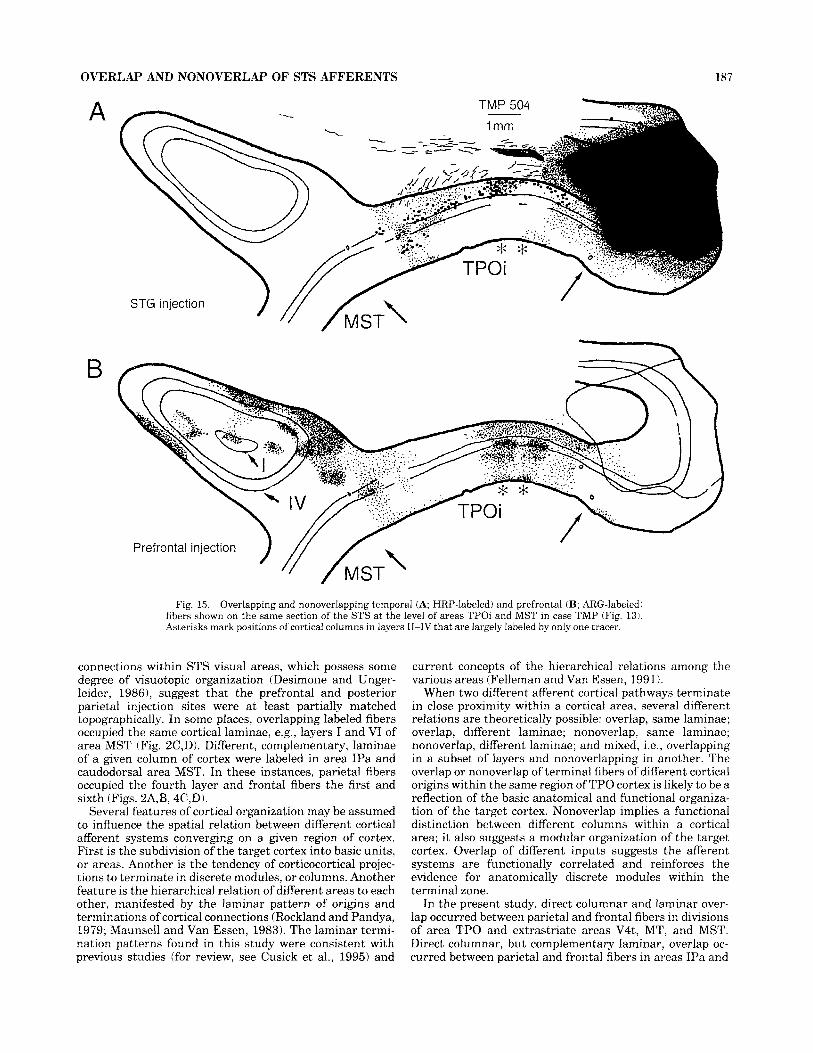
OVERLAP AND NONOVERLAP OF STS AFFERENTS 187
A
B
STG injection
7
Prefrontal injection
Fig. 15. Overlapping and nonoverlapping temporal (A; HRP-labeled) and prefrontal (B; ARG-labeled) fibers shown on the same section of the STS at the level of areas TPOi and MST in case TMP (Fig. 13). Asterisks mark positions of cortical columns in layers 11-IV that are largely labeled by only one tracer.
connections within STS visual areas, which possess some degree of visuotopic organization (Desimone and Unger- leider, 19861, suggest that the prefrontal and posterior parietal injection sites were at least partially matched topographically. In some places, overlapping labeled fibers occupied the same cortical laminae, e.g., layers I and VI of area MST (Fig. 2C,D). Different, complementary, laminae of a given column of cortex were labeled in area IPa and caudodorsal area MST. In these instances, parietal fibers occupied the fourth layer and frontal fibers the first and sixth (Figs. 2A,B, 4C,D).
Several features of cortical organization may be assumed to influence the spatial relation between different cortical afferent systems converging on a given region of cortex. First is the subdivision of the target cortex into basic units, or areas. Another is the tendency of corticocortical projec- tions to terminate in discrete modules, or columns. Another feature is the hierarchical relation of different areas to each other, manifested by the laminar pattern of origins and terminations of cortical connections (Rockland and Pandya, 1979; Maunsell and Van Essen, 1983). The laminar termi- nation patterns found in this study were consistent with previous studies (for review, see Cusick et al., 1995) and
current concepts of the hierarchical relations among the various areas (Felleman and Van Essen, 1991).
When two different afferent cortical pathways terminate in close proximity within a cortical area, several different relations are theoretically possible: overlap, same laminae; overlap, different laminae; nonoverlap, same laminae; nonoverlap, different laminae; and mixed, i.e., overlapping in a subset of layers and nonoverlapping in another. The overlap or nonoverlap of terminal fibers of different cortical origins within the same region of TPO cortex is likely to be a reflection of the basic anatomical and functional organiza- tion of the target cortex. Nonoverlap implies a functional distinction between different columns within a cortical area; it also suggests a modular organization of the target cortex. Overlap of different inputs suggests the afferent systems are functionally correlated and reinforces the evidence for anatomically discrete modules within the terminal zone.
In the present study, direct columnar and laminar over- lap occurred between parietal and frontal fibers in divisions of area TPO and extrastriate areas V4t, MT, and MST. Direct columnar, but complementary laminar, overlap oc- curred between parietal and frontal fibers in areas IPa and
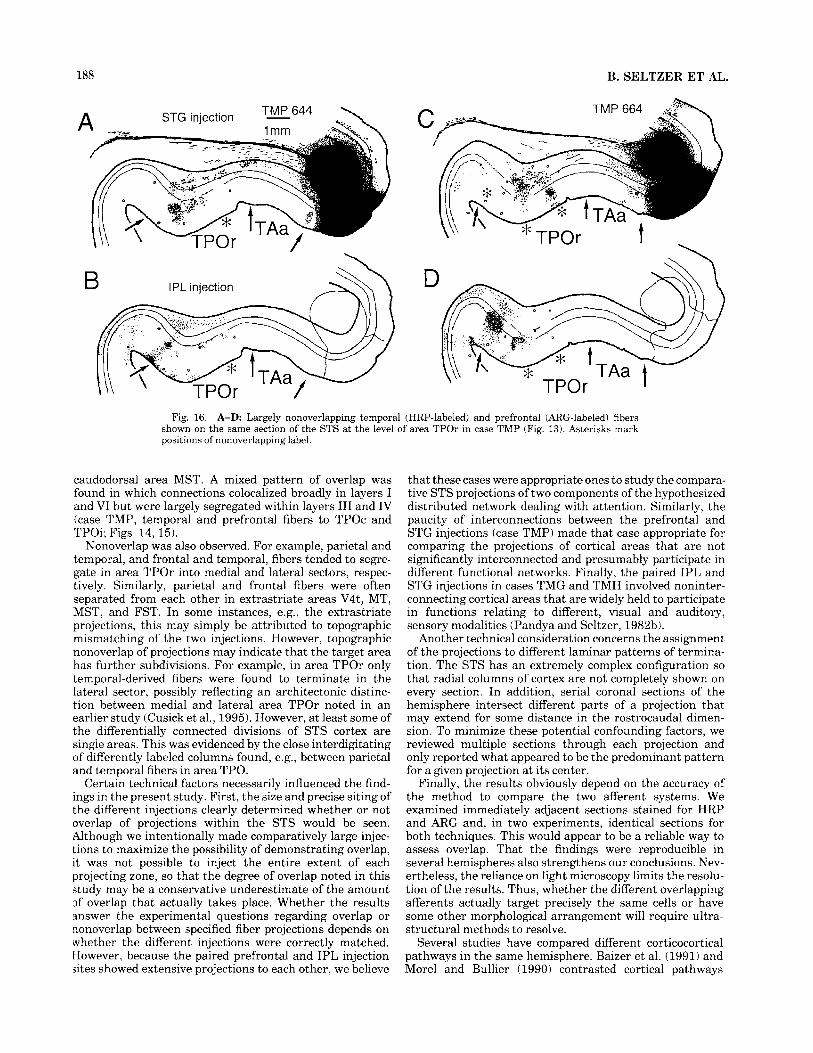
188 B. SELTZER ET AL.
\ \ \ . TPOr \
t
Fig. 16. A-D Largely nonoverlapping temporal (HRP-labeled) and prefrontal (ARG-labeled) fibers shown on the same section of the STS at the level of area TPOr in case TMP (Fig. 13). Asterisks mark positions of nonoverlapping label.
caudodorsal area MST. A mixed pattern of overlap was found in which connections colocalized broadly in layers I and VI but were largely segregated within layers 111 and IV (case TMP, temporal and prefrontal fibers to TPOc and TPOi; Figs. 14, 15).
Nonoverlap was also observed. For example, parietal and temporal, and frontal and temporal, fibers tended to segre- gate in area TPOr into medial and lateral sectors, respec- tively. Similarly, parietal and frontal fibers were often separated from each other in extrastriate areas V4t, MT, MST, and FST. In some instances, e.g., the extrastriate projections, this may simply be attributed to topographic mismatching of the two injections. However, topographic nonoverlap of projections may indicate that the target area has further subdivisions. For example, in area TPOr only temporal-derived fibers were found to terminate in the lateral sector, possibly reflecting an architectonic distinc- tion between medial and lateral area TPOr noted in an earlier study (Cusick et al., 1995). However, at least some of the differentially connected divisions of STS cortex are single areas. This was evidenced by the close interdigitating of differently labeled columns found, e.g., between parietal and temporal fibers in area TPO.
Certain technical factors necessarily influenced the find- ings in the present study. First, the size and precise siting of the different injections clearly determined whether or not overlap of projections within the STS would be seen. Although we intentionally made comparatively large injec- tions to maximize the possibility of demonstrating overlap, it was not possible to inject the entire extent of each projecting zone, so that the degree of overlap noted in this study may be a conservative underestimate of the amount of overlap that actually takes place. Whether the results answer the experimental questions regarding overlap or nonoverlap between specified fiber projections depends on whether the different injections were correctly matched. However, because the paired prefrontal and IPL injection sites showed extensive projections to each other, we believe
that these cases were appropriate ones to study the compara- tive STS projections of two components of the hypothesized distributed network dealing with attention. Similarly, the paucity of interconnections between the prefrontal and STG injections (case TMP) made that case appropriate for comparing the projections of cortical areas that are not significantly interconnected and presumably participate in different functional networks. Finally, the paired IPL and STG injections in cases TMG and TMH involved noninter- connecting cortical areas that are widely held to participate in functions relating to different, visual and auditory, sensory modalities (Pandya and Seltzer, 1982b).
Another technical consideration concerns the assignment of the projections to different laminar patterns of termina- tion. The STS has an extremely complex configuration so that radial columns of cortex are not completely shown on every section. In addition, serial coronal sections of the hemisphere intersect different parts of a projection that may extend for some distance in the rostrocaudal dimen- sion. To minimize these potential confounding factors, we reviewed multiple sections through each projection and only reported what appeared to be the predominant pattern for a given projection at its center.
Finally, the results obviously depend on the accuracy of the method to compare the two afferent systems. We examined immediately adjacent sections stained for HRP and ARG and, in two experiments, identical sections for both techniques. This would appear to be a reliable way to assess overlap. That the findings were reproducible in several hemispheres also strengthens our conclusions. Nev- ertheless, the reliance on light microscopy limits the resolu- tion of the results. Thus, whether the different overlapping afferents actually target precisely the same cells or have some other morphological arrangement will require ultra- structural methods to resolve.
Several studies have compared different corticocortical pathways in the same hemisphere. Baizer et al. (1991) and Morel and Bullier (1990) contrasted cortical pathways

OVERLAP AND NONOVERLAP OF STS AFFERENTS 189
and STG fibers. The nonoverlap of fibers from the STG and caudal IPL in area TPO is not unexpected, given that these areas are not themselves interconnected (Seltzer and Pan- dya, 1984; Cavada and Goldman-Rakic, 1989; Andersen et al., 1990; Morel and Bullier, 1990; Baizer et al., 1991). Furthermore, up to the present time, cortical regions relating to presumably different sensory modalities have generally been associated with segregated corticocortical pathways. Nevertheless, pathways relating to hearing and vision do reach area TPO in the STS, where they presum- ably may interact, and cells in this vicinity have clearly been shown to have bimodal, and even trimodal, sensory re- sponse properties (Desimone and Gross, 1979; Bruce et al., 1981). The present data suggest that this takes place, at least in part, by an interaction between contiguous auditory and visual modules within the STP rather than in discrete convergence columns.
relating to the dorsal and ventral visual streams (Unger- leider and Mishkin, 1982) following injections of retrograde tracers into the IPL and inferotemporal cortex. Within the STS, they showed a predominant segregation of these two major visual pathways. A notable focus of dorsal and ventral stream overlap was found, not within STP cortex, but rostrally in the fundus of the STS. Selemon and Goldman-Rakic (1988) explicitly contrasted the cortical and subcortical projections of prefrontal and posterior parietal (IPL) cortex by using the double anterograde tracer method. They found complementary overlap within area IPa, simi- lar to what was found in this study, but did not indicate any overlap of projections in the polysensory cortex of the upper bank of the STS.
Although based of necessity on injections involving only a few selected cortical areas, the data may nevertheless reveal certain organizing principles of cortical connections. Thus, areas of the cortex that are themselves interconnected, such as the dorsal prearcuate cortex and the caudal IPL, appear to send terminal fibers that overlap each other when they both target a third cortical area such as area TPO. Pairs of areas such as the IPL and STG, which have few interconnections, show little, if any, direct overlap of fibers within the STS, even though both precursor zones do send projections to the same target area. An intermediate pat- tern, showing overlap within layer VI and mainly nonover- lap in layers I11 and IV, was noted following paired injec- tions of prefrontal and STG cortex, which also were largely noninterconnected areas.
Dorsal prearcuate and posterior parietal cortices have been hypothesized to be part of a hemispherewide distrib- uted system of cortical areas having to do with mechanisms of directed attention and related functions (Mesulam, 1990; Friedman and Goldman-Rakic, 1994). That both areas project in an overlapping fashion to area TPO in the STS suggests that at least portions of area TPO also belong to the same distributed network. The network concept does not, however, imply that all component areas of a given circuit are functionally equal. Rather, each is considered to have a special role with respect to the function subserved by the network (Mesulam, 1990). Columns within area TPO receive converging input from the posterior parietal and prefrontal cortex that may relate to visuospatial sensory and eye movement information, respectively. However, the particular role played by STP cortex (area TPO) in spatial attention remains unknown.
The frontal-STG pattern is principally nonconvergent, which might be predicted because the injected portions of these regions are not significantly interconnected. That there was significant overlap of frontal and temporal fibers in layers I and VI of area TPO suggests, however, an integration of the two different types of afferent informa- tion within cortical layers I and VI. Furthermore, although the prefrontal and temporal fibers tended to segregate within layers 111 and IV, there were some instances of slight overlap within these layers, raising the possibility that, had we made an injection into some other, perhaps more rostral, area of prefrontal cortex, more overlap might have been found. Previous studies of STG connections to prefron- tal cortex (Barbas, 1988; see Pandya and Yeterian, 1990) suggest that frontal-STG-STP distributed networks may exist.
The results imply that, in STP cortex, functional integra- tion of information carried by parietal and prefrontal fibers is achieved by a different mode than that relayed by parietal
ACKNOWLEDGMENTS We thank C’Lita Henry for technical assistance. This
study was supported by NIH grant EY08906, the Depart- ment of Veterans Affairs Medical Center, New Orleans, Louisiana, and the Administrators of the Tulane Educa- tional Fund.
LITERATURE CITED Andersen, R.A., C. Asanuma, G. Essick, and R.M. Siege1 (1990) Corticocorti-
cal connections of anatomically and physiologically defined subdivisions within the inferior parietal lobule. J. Comp. Neurol. 296:65-113.
Baizer, J.S., L.G. Ungerleider, and R. Desimone (1991) Organization of visual inputs to the inferior temporal and posterior parietal cortex in macaques. J. Neurosci. 11:168-190.
Barbas, H. ( 1988) Anatomic organization of basoventral and mediodorsal visual recipient prefrontal regions in the rhesus monkey. J. Comp. Neurol. 276:313-342.
Baylis, C.G., E.T. Rolls, and C.M. Leonard (1987) Functional subdivisions of the temporal neocortex. J. Neurosci. 7:330-342.
Benevento, L.A., J. Fallan, B.J. Davis, and M. Rezak (1977) Auditory- visual interactions in single cells in the cortex of the superior temporal sulcus and the orbital frontal cortex of the macaque monkey. Exp. Neurol. 575349-872.
Blatt, G.J., R.A. Andersen, and G.R. Stoner (1990) Visual receptive field organization and cortico-cortical connections of the lateral intraparietal area (area LIP) in the macaque. J. Camp. Neurol. 299:421-445.
Bonin, G. von, and P. Bailey (1947) The Neocortex of Macaca rnulatta. Urbana: University of Illinois Press.
Boussaoud, D., L.G. Ungerleider, and R. Desimone (1990) Pathways for motion analysis: Cortical connections of the medial superior temporal and fundus of the superior temporal visual areas in the macaque. J. Comp. Neurol. 296:463495.
Brodmann, K. ( 1909) Vergleichende Lokalisationslehre der Grosshirnrinde, 2nd ed. Leipzig: Barth.
Bruce, C.J., R. Desimone, and C.G. Gross (1981) Visual properties of neurons in a polysensory area in superior temporal sulcus of the macaque. J. Neurophysiol. 46:369-384.
Bruce, C.J., R. Desimone, and C.G. Gross (1986) Both striate cortex and superior colliculus contribute to visual properties of neurons in superior temporal polysensory area of macaque monkey. J. Neurophysiol. 55:1057- 1075.
Campbell, M.J., and J.H. Morrison ( 1989) Monoclonal antibody to neurofila- ment protein (SMI-32) labels a subpopulation of pyramidal neurons in the human and monkey neocortex. J. Comp. Neurol. 282: 191-205.
Cavada, C., and P.S. Goldman-Rakic (1989) Posterior parietal cortex in the rhesus monkey: 11. Evidence for segregated corticocortical networks linking sensory and limbic areas with the frontal lobe. J. Comp. Neurol. 287:422445.
Cowan, W.M., D.I. Gottlieb, A.E. Hendrickson, J.L. Price, and T.A. Woolsey (1972) The autoradiographic demonstration of axonal connections in the central nervous system. Brain Res. 37.2-51

190
Cusick, C.G., B. Seltzer, M. Cola, and E. Griggs (1995) Chemoarchitectonics and cortico-cortical terminations within the superior temporal sulcus of the rhesus monkey: Evidence for subdivisions of superior temporal polysensory cortex. J . Comp. Neurol. 360513-535.
Desimone, R., and C.G. Gross 11979) Visual areas in the temporal cortex of the macaque. Brain Res. 178:363-380.
Desimone, R., and L.G. Ungerleider 11986) Multiple visual areas in the caudal superior temporal sulcus of the macaque. J . Comp. Neurol. 248: 164-189.
Felleman, D.J., and D.C. Van Essen ( 1991) Distributed hierarchical process- ing in the primate cerebral cortex. Cereb Cortex 1:1-47.
Funahashi, S., C.J. Bruce, and P.S. Goldman-Rakic (1993) Dorsolateral prefrontal lesions and oculomotor delayed-response performance: Evi- dence for mnemonic “scotomas.” J . Neurosci. 13:1479-1497.
Friedman, H.R., and P.S. Goldman-Rakic (1994) Coactivation of prefrontal cortex and inferior parietal cortex in working memory tasks revealed by 2DG functional mapping in the rhesus monkey. J . Neurosci. 14:2775- 2788.
Galaburda, A.M., and D.N. Pandya (1983) The intrinsic architectonic and connectional organization of the superior temporal region of the rhesus monkey. J . Comp. Neurol. 221:169-184.
Gallyas, F. 11979) Silver staining of myelin by means of physical develop- ment. Neurol. Res. 1:203-209.
Goldman-Rakic, P.S. 11988) Topography of cognition: Parallel distributed networks in primate association cortex. Annu. Rev. Neurosci. 1 I:137- 156.
Goldman-Rakic, P.S., and H. Friedman (1991) Circuitry of working memory revealed by anatomy and metabolic imaging. In H.S. Levin, H.M. Eisenberg, and A.L. Benton ieds): Frontal Lobe Function and Dysfunc- tion. New York: Oxford University Press, pp. 72-91.
Goldman-Kakic, P.S., J.F. Bates, and M.V. Chafee (1992) The prefrontal cortex and internally generated motor acts. Curr. Opin. Neurobiol. 6r830-835.
Heilman, K.M., D.N. Pandya, and N. Geschwind 11970) Trimodal inatten- tion following parietal lobe ablations. Trans. Am. Neurol. Assoc. 95:259- 261.
Hietanen, J.K., and D.I. Perrett (1993) Motion sensitive cells in the macaque superior temporal polysensory area. I. Lack of response to the sight of the animal’s own limb movement. Exp. Brain Res. 93:117-128.
Hikosaka, K., E. Iwai, H. Saito, and K. Tanaka (1988) Polysensory properties of neurons in the anterior bank of the caudal superior temporal sulcus of the macaque monkey. J . Neurophysiol. 60:1615-1637.
Hof, P.R., and J .H. Morrison (1995) Neurofilament protein defines regional patterns of cortical organization in the macaque monkey visual system: A quantitative immunohistochemical analysis. J . Comp. Neurol. 352: 161- 186.
Hsu, S.M., L. Raine, and H. Fanger (1981) Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: A comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cyto- chem. 29577-580.
Jones, E.G., and T.P.S. Powell (1970) An anatomical study of converging sensory pathways within the cerebral cortex of the monkey. Brain 93: 793-820.
Komatsu. H., and R.H. Wurtz 119881 Relation ofcortical areas MT and MST to pursuit eye movements. I. Localization and visual properties of neurons. J. Neurophysiol. 60:580-603.
Luh, K.E., C.M. Butter, and H.A. Buchtel (1986) Impairments in orienting to visual stimuli in monkeys following unilateral lesions of the superior sulcal polysensory cortex. Neuropsychologia 24:461470.
Lynch, J .C. 11992) Saccade initiation and latency deficits after combined lesions of the frontal and posterior eye fields in monkeys. J . Neuro- physiol. 68:1913-1916.
Maunsell. J.H.R., and D.C. Van Essen (1983) The connections of the middle temporal visual area IMT) and their relationship to a cortical hierarchy in the macaque monkey. J . Neurosci. 3:2563-2586.
Mesulam, M.-M. i 1978) Tetramethylbenzidine for horseradish peroxidase neurohistochemistry: A non-carcinogenic blue reaction product with superior sensitivity for visualizing neural afferents and efferents. J . Histochem. Cytochem. 26:106-117.
Mesulam, M.-M. 11990) Large scale neurocognitive networks and distributed processing for attention, language, and memory. Ann. Neurol. 28:597- 613.
Milner, A.D., and M A Goodale 11993) Visual pathways to perception and action. Prog. Brain Res. 95:317-337.
Morecraft. K.J.. C. Geula. and M.-M. Mesulam 11993) Architecture of
B. SELTZER ET AL.
Morel, A., and J . Bullier (1990) Anatomical segregation of two cortical visual pathways in the macaque monkey. Vis. Neurosci. 4555-578.
Morel, A., P.E. Garraghty, and J .H. Kaas (1993) Tonotopic organization, architectonic fields, and connections of auditory cortex in macaque monkeys. J . Comp. Neurol. 335:437-459.
Omata, K., Y. Osawa, H. Okuda, and E. Iwai (1988) Behavioral impairment following unilateral lesions of caudodorsal superior temporal sulcus in monkey. Nokenkyukai Kaisji/Neuroscience 14: 168-170.
Oram, M.W., D.I. Perrett, and J.K. Hietanen (1993) Directional tuning of motion-sensitive cells in the anterior superior temporal polysensory area of the macaque. Exp. Brain Res. 97274-294.
Pandya, D.N., and F. Sanides (1973) Architectonic parcellation of the temporal operculum and its projection pattern. Zeitschr. Anat. Entwick- lungsgesch. 139: 127-161.
Pandya, D.N., and B. Seltzer i1982aj Intrinsic connections and architecton- ics of posterior parietal cortex in the rhesus monkey. J . Comp. Neurol. 204:196-210.
Pandya, D.N., and B. Seltzer (1982b) Association areas of the cerebral cortex. Trends Neurosci. 5.386-390.
Pandya, D.N., and E.H. Yeterian 11990) Prefrontal cortex in relation to other cortical areas in rhesus monkey: Architecture and connections. Prog. Brain Res. 8563-94.
Rockland, K.S., and D.N. Pandya (1979) Laminar origins and terminations of cortical connections of the occipital lobe in the rhesus monkey. Brain Res. 179:3-20.
Scalaidhe, S.P., T.D. Albright, and H.R. Rodman (1995) Effects of superior temporal polysensory area lesions on eye movements in the macaque monkey. J. Neurophysiol. 73:l-19.
Schall, J.D., A. Morel, D.J. King, and J . Bullier (1995) Topography of visual cortex connections with frontal eye field in macaque: Convergence and segregation of processing streams. J . Neurosci. 15:4464-4487.
Selemon, L.D., and P.S. Goldman-Rakic (1988) Common cortical and subcortical targets of the dorsolateral prefrontal and posterior parietal cortices in the rhesus monkey: evidence for a distributed neural network subsewing spatially guided behavior. J. Neurosci. 8:4049-4068.
Seltzer, B. 11992) An anatomy of parallel distributed processing. Behav. Brain Sci. 15:796.
Seltzer, B., and D.N. Pandya (1978) Afferent cortical connections and architectonics of the superior temporal sulcus and surrounding cortex in the rhesus monkey. Brain Res. 149:l-24.
Seltzer, B., and D.N. Pandya (1984) Further observations on parieto- temporal connections in the rhesus monkey. Exp. Brain Res. 55:301- 312.
Seltzer, B., and D.N. Pandya 11989a) Frontal lobe connections of the superior temporal sulcus in the rhesus monkey. J . Comp. Neurol. 281:97-1 13.
Seltzer, B., and D.N. Pandya i1989b) Intrinsic connections and architecton- ics of the superior temporal sulcus in the rhesus monkey. J. Comp. Neurol. 290:45 1 4 7 1 .
Seltzer, B., and D.N. Pandya (1991) Post-Rolandic cortical projections of the superior temporal sulcus in the rhesus monkey. J . Comp. Neurol. 312:625-640.
Seltzer, B., and D.N. Pandya 11994) Parietal, temporal, and occipital projections to cortex of the superior temporal sulcus in the rhesus monkey: A retrograde tracer study. J. Comp. Neurol. 343:445463.
Stanton, G.B., C.J. Bruce, and M.E. Goldberg 11995) Topography of projec- tions to posterior cortical areas from the macaque frontal eye fields. J . Comp. Neurol. 353:291-305.
Suzuki, W.A., and D.G. Amaral i 1994) Perirhinal and parahippocampal cortices of the macaque monkey: Cortical afferents. J . Comp. Neurol. 350:497-533.
Ungerleider, L.G., and R. Desimone (1986) Cortical connections of visual area MT in the macaque. J . Comp. Neurol. 248:190-222.
Ungerleider, L.G., and M. Mishkin (1982) Two cortical visual systems. In D.J. Ingle, M.A. Goodale, and R.J.W. Mansfield (eds): Analysis of Visual Behavior. Cambridge, MA: MIT Press, pp. 549-586.
Walker, A.E. (1940) A cytoarchitectural study of the prefrontal area of the macaque monkey. J . Comp. Neurol. 73:59-86.
Watson, R.T., E. Valenstein, A. Day, and K.M. Heilman 11994) Posterior neocortical systems subserving awareness and neglect. Arch. Neurol. 5I:1014-1021.
Wong-Riley, M.T.T. (1979) Changes in the visual system of monocularly sutured or enucleated cats demonstrable with cvtochrome oxidase
, ,
connectivity within a cingulo-fronto-parietal neurocognitive network for directed attention. Arch. Neurol. 50:279-284. histochemistry. Brain Res. 171:11-28