orangutan behaviour in captivity: activity … · orangutan behaviour in captivity: activity...
TRANSCRIPT

O R A N G U T A N B E H A V I O UR IN C APT I V I T Y :
A C T I V I T Y BUD G E TS, E N C L OSUR E USE A ND
T H E V ISI T O R E F F E C T
C H O O Y U A N T IN G
B.Sc. (Hons.), NUS
A THESIS SUBMITTED FOR THE DEGREE OF
MASTER OF SCIENCE
DEPARTMENT OF BIOLOGICAL SCIENCES
NATIONAL UNIVERSITY OF SINGAPORE
2011

ii
A C K N O W L E D G E M E N TS
Just as I was thinking that my affair with the hairy creatures would never end, I finally found
myself having to bid them farewell. The following thanks are kept brief, as no words adequately
express the relationships between people. I had fun and much chance to grow during my research,
and to everyone listed here, I hope I have made just a bit of difference in your life as well.
As with any routine done regularly over time, one starts to develop what I call specialized quirks.
Some people call them occupational hazards. Being in the unique situation of watching apes in a
zoo setting, where I have both hairy as well as hairless ones to watch, I cannot help but draw
comparisons between the two.
During the first few months of watching orangutans, I started seeing orangutans everywhere I
went. The way my dad sat with his leg up on the sofa looked like an orangutan. The way a
stranger on the street scratched himself reminded me of a certain old orangutan in the zoo. The
crowning touch came one day when after exiting the zoo washroom, I passed through a crowded
restaurant on the way back to the orangutan exhibit. To my horror/amazement, I saw Bento, one
of the 2 year old (orangutans) rolling around on the floor! How on earth did he get out, and there
he was, frolicking in the midst of humans! I was stunned. But on taking a second look, I realized
that it was actually a very tanned human child in an orange singlet, rolling around on the floor.
That incident proved really hilarious, even to myself.
Other humou s of time include
hearing the zoo show jingle, in the toilet. And once, I thought I heard
the mating/territorial call of a mature male orangutan in my house. I wondered: Why is Charlie
long-calling in my house??
The following thesis is a summary and analysis of my past months with these apes. They have
brought me only fun, laughter and constant opportunities to self-introspect. On the other hand, the
heartache and sweat came solely from the efforts of this human, to want to watch them in order to
write a long report that humans call a thesis. For all of this, I am grateful, and would like to thank
the following people.

iii
Dr Li, my ever present source of advice and support. Thank you for taking in the young and
inexperienced girl back then, and nurturing her into the competent, confident researcher she is
today.
Dr Todd, my grammar master. Always ready to pick up where I have left off, and pushing me that
extra step to do a better job. Thank you.
Of course, with any captive animals, come their ever faithful keepers. Jack, Kumaran, Gabriel,
Prakash, Md Noor, Arshad and Marzuki. I sincerely thank you for all you have taught me and I
hope your lives have been at least somewhat enriched,
rt and advice you have given
me, I have really enjoyed knowing you!
Eunice. I felt that choosing to go on with Masters, rather than working after my B.Sc, was made
in part worthwhile because I got to know you ^^
My labmates with them, I understand why humans are truly social creatures. Even the intensely
focused task of analyzing data or writing a thesis re
company better to revel in, than those of my ever fun, ever game fellow researchers. Thank you
Huishan, Diego, Stanley, Junhao, Seok Ping, Shichang, Mindy, Joelyn, Ganison and my other
Spider Lab colleagues who make me look forward to going to lab.
Giam, Dr Chan Yiong Huak, Kia Chong, Yangchen and Lainie. Thank you for your invaluable
statistical advice and comments on my project.
My father, who paces me both in life, and as a researcher. He, who I must attribute much of who I
am today to. My family, always supportive, despite my regular crankiness. Thank you.
And last but definitely not least, myself.
best and grown lots through this experience. Well done.
And to you, dear reader, I hope you enjoy the following account of my time spent with the hairy,
as well as not so hairy apes of the Singapore Zoo.

iv
T A B L E O F C O N T E N TS
Page
Acknowledgements ii
Table of Contents iv
Summary vii
L ist of Tables ix
L ist of F igures x
Chapter 1: General Introduction 1
Orangutans 1
Zoos then, and now 4
Singapore Zoo 6
Study objectives 8
Chapter 2: Activity Budgets of O rangutans in Two Naturalistic Enclosures and Comparisons to Wild O rangutans
11
Introduction 12
Materials and Methods 15
Results 23
Overall activity budgets and activity budgets across age groups 23
Comparing activity budgets across enclosures 27
Comparing captive to wild activity budgets 28
Discussion 29
Comparing activity budgets across age groups 29
Comparing activity budgets across enclosures 31
Comparing captive to wild activity budgets 33
Recommendations 37

v
Chapter 3: Use of Enclosure Structures and Vertical Space in Two Naturalistic O rangutan Exhibits
39
Introduction 40
Materials and Methods 44
Results 50
Overall structure use 50
Structure use across age groups in both exhibits 51
Behaviours at each structure type 53
Use of vertical space in enclosure 56
Discussion 57
Use of enclosure structures across age groups and exhibits 57
Behaviours at each structure type 61
Use of vertical space in enclosure 64
Limitations of study 67
Recommendations 68
Chapter 4: V isitor effects on zoo orangutans in two novel, naturalistic
enclosures
71
Introduction 72
Materials and Methods 76
Results 82
Effect of visitors on orangutan behaviour in the Boardwalk exhibit 83
Effect of visitors on orangutan behaviour in the Island exhibit 83
Discussion 87
Effects of visitor number 87
Effects of visitor activity 88
Effects of visitor proximity 90
Other influences on the visitor-animal interactions 91
Limitations of study 92
Recommendations 96

vi
Chapter 5: General Discussion 98
Common factors across the three chapters 99
Recommendations on enclosure design 102
Recommendations on husbandry routine 104
Other considerations and recommendations 106
Study limitations and future directions 109
References 111
Appendix 124

vii
SU M M A R Y
Orangutans are great apes that are found in the forests of Borneo and Sumatra. In the wild, they
live in complex rainforest habitats, and travel great distances daily for food. As a result of habitat
destruction and poaching, these apes are now endangered. In captivity, orangutans become obese
if not provided with sufficient arboreal stimulation. They are also known for their intelligence,
and keeping them mentally occupied is a challenge. For captive orangutans, little is known about
their activity budgets, enclosure use and how zoo visitors affect them. Hence, more knowledge is
required to maintain the welfare of these intelligent apes.
In zoos, there is a current and ongoing interest for naturalistic exhibits, as such exhibits may
provide greater stimulation for captive animals. Using features such as vegetation and rockwork,
naturalistic enclosures are designed to increase species-specific behaviours by simulating wild
habitats. Such exhibits have also been found to improve visitor appreciation of captive animals. In
Singapore Zoo, the presence of two naturalistic orangutan exhibits provides the chance to study
the behaviour of this ape in such enclosures. Hence, the activity budgets, enclosure use, and
visitor effects on Singapore Zoo orangutans were investigated.
The results showed that captive orangutan activity budgets were age-specific, differed across
enclosures, and were not dissimilar from that of wild orangutans. Exhibit use was influenced by
both biological and environmental factors. Biological factors included age and dominance
hierarchy in orangutans, and environmental factors included the availability and arrangement of
structures within an exhibit, as well as features surrounding the exhibit. For visitor effects, large
crowds, visitors with food, visitors who were looking or taking photographs, and visitors who
were close by, all affected orangutan behaviour. On the whole however, the effects of visitors on

viii
Interestingly, possible
sources of visitor stress appeared to be alleviated by the large, naturalistic enclosure designs and
the unusual husbandry routines implemented at Singapore Zoo. Being the first study on structure
use in a naturalistic orangutan exhibit, and amongst the few existing studies on orangutan activity
budgets and visitor effects, this research provides useful information for zoo management, and
sets possible direction for future studies.

ix
L IST O F T A B L ES
Table 2.1.
Individual details of study animals. 20
Table 2.2.
Definitions for orangutan behaviours.
20
Table 3.1.
Individual details and age groups of study animals. 49
Table 3.2.
Definitions for orangutan behaviours. 49
Table 4.1.
Individual details of study animals. 78
Table 4.2.
Definitions for orangutan and visitor variables.
79
Table 4.3.
Definitions and levels of variables for multinomial logistic regression 81
Table 4.4. Likelihood ratio test results for both enclosures.
82
Table 4.5. Results from multinomial logistic regression testing effects of visitor number, activity and proximity on orangutan behaviour in the two exhibits.
85
Table 4.6. Effects of visitor variables on likelihood of each orangutan behaviour for both exhibits.
86

x
L IST O F F I G UR ES
Fig 1.1. Top: adult female orangutan. Bottom: juvenile and adolescent males feeding on leaves.
3
Fig. 2.1. Schematic diagram of Boardwalk Free-ranging exhibit.
16
Fig. 2.2. Schematic diagram of Island Free-ranging exhibit.
17
Fig. 2.3. Plan diagram showing proximity of Boardwalk and Island exhibits to surrounding features.
18
Fig. 2.4. Overall activity budgets for all orangutans in Boardwalk exhibit and Island exhibit.
23
Fig. 2.5. Activity budgets for different age groups in Boardwalk exhibit.
24
Fig. 2.6. Activity budgets for different age groups in Island exhibit.
25
Fig. 2.7. Activity budgets for Boardwalk and Island exhibits.
27
Fig. 2.8. Activity budgets for wild and captive orangutans.
28
Fig. 3.1. Schematic diagram of Boardwalk Free-ranging exhibit
45
Fig. 3.2. Schematic diagram of Island Free-ranging exhibit.
46
Fig. 3.3. Plan diagram showing proximity of Boardwalk and Island exhibits to surrounding features.
47
Fig. 3.4. Overall structure use for all orangutans in Boardwalk and Island exhibits.
50
Fig. 3.5. Structure use for different age groups in Boardwalk and Island exhibits.
51
Fig. 3.6. Behaviours of different age groups at structures in the Boardwalk and Island exhibits.
54
Fig. 3.7. Use of vertical space by different age groups in Boardwalk and Island exhibits.
56
Fig. 4.1. Island exhibit and Boardwalk exhibit showing possible locations of visitor-orangutan interaction.
77
Fig 5.1. Log that facilitates entry into Island orangutan exhibit.
109

1
C H APT E R 1
G E N E R A L IN T R O DU C T I O N
O rangutans
The orangutan (Pongo pygmaeus, Pongo abelii) is an enigmatic ape which lives in the forests of
Southeast Asia. It belongs to the family Hominidae, and is the only great ape found outside of
Africa (Groves et al., 2005). Orangutans are currently distributed on the islands of Borneo and
Sumatra (P. pygmaeus, P. abelii respectively), and from fossil records, were thought to
previously range through Java, Vietnam and Southern China (Delgado and Van Schaik, 2000;
Rijksen and Meijaard, 1999). These apes show sexual dimorphism, where adult females weigh an
average of 36 kg, and males, 78 kg (Smith and Jungers, 1997). Males come in two forms, mature
flanged males with disc-like cheek pads (Kingsley, 1988), and unflanged subadult males, which
weigh approximately the same as adult females (Galdikas, 1985b).
In the wild, orangutans are mostly arboreal, and have home ranges spanning several kilometers
(males: 0.6 25 km2, females: 0.4 9km2) (Utami Atmoko et al., 2009). Because they are highly
adapted to life in the trees, orangutans have relatively longer upper limbs and shorter lower limbs.
Hook-like hands and opposable toes in combination with extremely mobile hip joints allow all
four limbs to grip tree branches, providing ease of movement in the spatially complex tree canopy
(see Fig 1.1) (Morbeck and Zihlman, 1988; Rodman and Mitani, 1987). These slow moving
creatures are frugivores, and feed opportunistically according to seasons in the rainforest. When
fruits are abundant (e.g. during masting season), orangutans spend more time feeding, and are
thought to store fat reserves for seasons of unpredictable fruit availability. When fruits are less

2
abundant, orangutans compensate by increasing the amounts of bark and leaves in their diet
(Delgado and Van Schaik, 2000). Because their food sources tend to be distributed irregularly
throughout the forest, these apes spend more than half of their time foraging and travelling
(Morrogh-Bernard et al., 2009)
Besides being physically adapted to life in the rainforest, orangutans are also intelligent creatures
with distinct cognitive abilities. As mentioned, orangutan food sources are located patchily in the
forest (Oates, 1987). Before an orangutan can feed, it has to first recall where the nearest food
source is, whether it is currently in season, and the shortest way to get there through the spatially
complex tree canopy. All these requirements point to capacities for higher mental abilities (Wich
et al., 2009). Orangutans in the wild also have distinct cultures and are known to innovate in their
behaviour and food extraction techniques (van Schaik et al., 2009). Although not as well studied
as in chimpanzees (Pan troglodytes), tool use is present in orangutans; more than twenty different
tool types have been fashioned for a variety of purposes (van Schaik et al., 2003). Captive
orangutans are also known to be expert tool users, and have been anecdotally reported to use tools
in escaping from zoo enclosures (Marks, 2009; Pope, 2009; Sonti, 2009). In captivity, orangutans
consistently score amongst the highest for primates in cognition tests (Tomasello and Call, 1997).
Together with the other great apes, they are capable of abilities seldom found in other primates,
such as mirror self-recognition, intentional deception and proto-language (Russon, 1998; Russon
and Bard, 1996). Studies have also shown that orangutans are capable of communicating their
intention to human observers and are able to modify their gestures to match the understanding of
the person (Cartmill and Byrne, 2007). Lastly, anecdotes from orangutan caretakers illustrate how
animals under their care actively observe their surroundings and show great interest in the

3
happenings beyond their enclosure (Hebert, 2011, pers. comm.1; Nantha, 2009, pers. comm.2). All
these poi
F ig 1.1. Top: adult female orangutan. Hand-like feet and mobile hip joints allow arboreal flexibility in orangutans. Bottom: juvenile and adolescent males feeding on leaves. Feeding typically occurs in pronograde suspension; the orangutan is suspended by one arm and one leg, and its body is horizontal to the ground.
1 Patricia Hebert, Orangutan Researcher, Fort Wayne Children's Zoo. 2 Gabriel Nantha, Orangutan Keeper, Singapore Zoo

4
In the 1700s, when wild orangutans were first discovered and captured by explorers, little was
known about their behaviour or ecology. As a result, captive animals rarely survived for long
(Maple, 1979). With present knowledge gained from wild populations, the reproductive success
and welfare of captive orangutans has improved greatly (Markham, 1990). Because orangutan
populations in the wild are declining rapidly due to habitat loss and poaching, much effort has
been turned to captive rehabilitation and release (Delgado and Van Schaik, 2000). Many zoos
have also started captive orangutan breeding programs to ensure the survival of this endangered
ape. As pointed out by Maple (1979) and Gippoliti (2000), orangutan exhibits still suffer from the
difficulties of creating suitable habitats for a large, arboreal primate. This is in comparison to
gorilla and chimpanzee exhibits, where excellent enclosures exist in a number of zoos. The main
challenges of keeping orangutans in captivity are providing adequate physical and mental
stimulation (Markham, 1990). Because orangutans tend to become obese if not given sufficient
climbing opportunities (Maple, 1979; Maple, 1980), and their intelligence and immense strength
lends to great investigative and destructive capabilities (Maple, 1980); it is essential to have good
understanding of both wild and captive orangutan behaviour for their welfare in zoos to be
optimized (Snowdon, 1989).
Zoos then, and now
Since olden times, zoos (and their historic equivalents) have been a source of fascination for
people of all cultures. The earliest records of exotic animals in captivity stem from 2500 BC,
where ancient Egyptians kept many species of birds and mammals as pets or in royal gardens
(Kisling, 2001; Strouhal, 1992). Menageries were also common in China and Greece where
animals were housed in private collections (Hosey et al., 2009; Schafer, 1968). During mediaeval
times, zoos (or their equivalents) did not feature as prominently in the historical records (AD 600
1450). However, in the seventeenth century, they proliferated again. A turning point came in

5
the eighteenth century when growth of European empires encouraged travel to distant places.
Enthused explorers returned from faraway lands, bringing with them an influx of exotic animals
and stimulating a subsequent interest in natural history. With the growing public interest, zoos
were no longer private menageries for the rich, but started opening to members of the public. By
the late nineteen century, the creation of zoological parks had also spread to the USA, and zoos
began to be seen as places of conservation and education. Nowadays, zoos are perceived as
conservation parks with missions to conduct research and create awareness on captive animal
welfare (Hosey et al., 2009).
With changes in the philosophy and mission statements of zoos, the way animals are housed has
also evolved. The mid-eighteenth century saw the use of first-generation exhibits which were
barren cages with steel bars and concrete floors. Subsequently, second-generation exhibits, still
prevalent today, are often cement enclosures surrounded by dry or wet moats. And as animal
welfare came under scrutiny in the 1970s, third-generation, and the most recent exhibits, came
into being (Hosey et al., 2009; Shettel-Neuber, 1988). Such exhibits are enclosures of greater
complexity which make use of naturalistic features to improve the welfare of captive creatures.
-nat
visitor appreciation of animal behaviour and ecology (Maple and Stine, 1982; Shettel-Neuber,
1988). Existing variants of naturalistic displays include free-ranging and immersion
(walkthrough) exhibits, as well as ecosystem zoos, wildlife parks, and bioparks (Hosey et al.,
2009).
Even though the shift towards naturalistic enclosures has proliferated worldwide, rigorous studies
on the benefits of such exhibits are still underway. To date, it is known that naturalistic exhibits
benefit both animals and zoo visitors, but scientific understanding is currently limited.

6
Naturalistic exhibits have been found to decrease abnormal behaviours and increase species-
specific behaviours in a variety of primates (Brent et al, 1991; Maple and Stine, 1982, Little and
Sommer, 2002). They can also improve visitor understanding of animal behaviour, increase
positive feelings towards the animals (Finlay et al., 1988; Ford and Burton, 1991) and have
greater visitor holding power (Ogden et al., 1994; Price et al., 1994). In addition, potential stress
from visitors is thought to be dampened in a large exhibit with natural features (Davey, 2007).
Despite the overall positive benefits of naturalistic enclosures, how animals interact with
individual features in their environment is poorly understood (Ogden et al., 1990) and the study
of visitor-animal interactions in such enclosures is still in its infancy (Davey, 2007; Ogden et al.,
1990). Hence, the following work will focus on addressing existing gaps in zoo ecology.
Singapore Zoo
Singapore Zoo, opened in 1973, is a local zoo under the management of the Wildlife Reserves
Singapore. Set in tropical surroundings, the zoo spans 28 hectares and holds 315 animal species
in its collection (website, Singapore Zoo) - ed for many of
the species, including the orangutans, Pongo pygmaeus and Pongo abelii.
In 2006, Singapore Zoo introduced two -
design from the older island-style exhibit, these enclosures each comprise a row of tall trees,
joined by artificial vines, nets, platforms and logs. Using trees which were originally situated next
to main visitor tracks, the orangutan enclosures were constructed by adding vines and structures
to the tree canopy and pruning tree foliage to maintain a minimum gap from surrounding trees.
Low-current electrical wires were also installed at the base of tree canopies to prevent orangutans
from descending to the ground.

7
- -ranging
(see Chapter 2, page 16). The former enclosure has an elevated boardwalk, so visitors
can view orangutans from both the main track (road), and the boardwalk. Visitors to the Island
exhibit, however, can only view orangutans from the main track below. These enclosures were
designed to encourage species-specific behaviours,
- website, Singapore Zoo). The enclosures are unique in
several ways and these include possible effects on both orangutans and visitors. The following are
several examples.
First, the majority of orangutan enclosures worldwide have animals displayed in exhibits at the
eye level of visitors, and climbing structures provided are often made of timber or inanimate
materials. Singapore Zoo orangutan exhibits are unique because orangutans are displayed on live
trees, under which visitors walk. Because the height at which animals are viewed has been found
or she is looking down at an animal, looking up instead at orangutans may instead evoke different
responses (Coe, 1985). Personal observation has shown that visitor responses range from one of
pleasant surprise, to obliviousness of the large apes moving overhead, and most comically of all,
sudden realization as the unsuspecting visitor is showered by excrement from an orangutan
above! In addition, it has also been suggested that viewing an animal amidst a natural backdrop of
greenery may promote a different viewing experience, because visitors gain a greater appreciation
of t and how it moves about in its natural environment (Shettel-Neuber,
1988).
Secondly, orangutans at Singapore Zoo have been observed to move from tree to tree using
species-specific behaviours which may not be possible in traditional orangutan exhibits. One
-

8
weight is used to bend tree branches to decrease the distance and cross gaps between
neighbouring trees (Thorpe et al., 2007). The presence of live trees in the exhibits may in this way
provide additional physical and mental stimulation for this group of orangutans.
Last but not least, when the effects of enclosure design on visitor-animal interactions are
considered, the elevated boardwalk in one of the exhibits allows visitors to come very close, up to
a distance of 3 m, from the orangutans. This close proximity permits animal observation in great
detail, and has been seen to evoke many excited responses from visitors. As a result, visitor-
animal interactions, such as food solicitation, are believed to occur more frequently in the exhibit
with the elevated boardwalk. For these reasons, Singapore Zoo orangutan exhibits are thought to
be unique.
Study objectives
Having discussed the presence of a unique study site, and the need for more information on
certain aspects of captive orangutan welfare, my study will focus on three areas of captive
orangutan behaviour, specifically, activity budgets, enclosure use and visitor effects.
Activity budgets
To date, published studies on captive orangutans have focused on cognition, play,
communication, plus a few on enrichment and enclosure modification (Cartmill and Byrne, 2010;
Leavens, 2007; Pizzutto et al., 2008; Tripp, 1985; Zucker et al., 1986). Although data are
available for many basic aspects of orangutan biology, there is a lack of information on captive
orangutan activity budgets. The activity budget of an animal can be defined as an adaptation to its
environment (Daan and Aschoff, 1975; Jaman and Huffman, 2008). For example, captive animals

9
are known to vary their activity budgets when changes are made to their enclosure or husbandry
routine (Hoff et al., 1997; Shepherdson et al., 1993). By comparing activity budgets between two
captive conditions, or between wild and captive populations, behavioural data can be used to
gauge if the welfare of the animals has been adequately provided for (Hosey et al., 2009). Such
data can also be used to ensure that behavioural diversity has not been lost through generations of
captive breeding (Hosey et al., 2009). Because orangutans are slow moving and are known to be
less expressive than other primates (Maple, 1980), specific indicators of stress in these captive
apes may be more difficult to identify. Hence, the proportion of time spent on different activities
can serve as a broad index of welfare. To date, there are only a few instances of published activity
budgets in orangutan literature (Forthman et al., 1993; Pearson et al., 2010; Pizzutto et al., 2008).
comparisons to wild conspecifics as well as those in other zoos, is the first objective of this thesis.
Enclosure use
As mentioned previously, there is a current trend worldwide to build naturalistic zoo exhibits.
These enclosures, with their greater complexity and more natural features, have been found to
improve the general welfare of captive animals. Aggressive and abnormal behaviours have been
reported to decrease, and species-specific behaviours increase, in naturalistic, complex,
enclosures (see review in Stoinski et al., 2001). However, specific interactions of animals with
features in the enclosure are not well understood, and more knowledge is required to optimize
exhibit use. Hence, the second objective of this study is to learn more about how captive
orangutans interact with structures in a naturalistic exhibit. The presence of two novel,
naturalistic enclosures at Singapore Zoo presents a good opportunity to research this topic.

10
Visitor effects
One factor that is common to all zoos is the presence of visitors. The effects of visitors on
animals have been studied since the 1970s (Oswald and Kuyk, 1977; Thompson, 1976) and have
been found to have positive, neutral or negative effects on animal welfare (Hosey, 2000). Such
welfare indications include: increases in active behaviour (positive) (Hosey, 2000), increases in
aggressive behaviour (negative) (Chamove et al., 1988; Sellinger and Ha) or no response to
varying visitor conditions (neutral) (see review in Davey, 2007). Most visitor research has
focused on simple presence-absence or visitor numbers. The limited knowledge on other aspects
of visitor presence, the lack of detailed definitions for some visitor variables, and the study of
visitor effects in isolation from other exhibit variables, all need to be addressed (Davey, 2007).
Hence, the third objective of this study is to investigate the visitor effects on orangutans in two
naturalistic, novel enclosures. The study will include a relatively wide range of visitor variables,
as well as investigate how visitor effects may interact with exhibit design.

11
C H APT E R 2
Activity Budgets of O rangutans in Two Naturalistic Enclosures
and Comparisons to Wild O rangutans
Abstract. Activity budgets can be used as indices of welfare, and also to check if captive animals
possess the behavioural diversity of their wild counterparts. Because keeping orangutans both
physically and mentally occupied in captivity is challenging, knowledge of their basic behaviour
is essential. However, detailed activity budgets for captive orangutans are currently lacking in zoo
literature. This study was designed to provide baseline data on orangutan activity budgets in two
naturalistic, novel enclosures. Activity budgets were collected for animals of different ages, and
compared against that of wild orangutans and animals from other zoos. A comparison of activity
budgets across two different enclosures was also made. Activity budgets differed significantly
across age groups, with younger animals showing more play and movement, and older animals
idling and paying more attention to their surroundings. Enclosure design and surrounding features
also appeared to influence activity budgets. Interestingly, activity budgets of this group were
similar to that of other orangutans, and also wild orangutans, especially those in masting
forests. The findings of this study are discussed in relation to captive welfare, and relevant
recommendations are made for zoo management.
K eywords: orangutan, activity budget, welfare, wild-captive, enclosure design, naturalistic

12
IN T R O DU C T I O N
The activity budget of an animal is the proportion of time it spends on different behaviours, such
as feeding, resting, and travelling. In captivity, many factors like restricted space, husbandry
practices or social grouping may affect an anima (Hosey et al., 2009). For
example, gorillas which were kept in an indoor enclosure showed more aggression, lay down
more, and had significantly different activity budgets from animals kept in outdoor naturalistic
enclosures (Hoff et al., 1997). Similarly, when feeding routines of leopard cats were changed to
include multiple feedings of hidden food, the cats paced less and their exploratory behaviour and
movement increased (Shepherdson et al., 1993). Because the activity budget of an animal can be
considered as an adaptation to its environment (Daan and Aschoff, 1975; Jaman and Huffman,
2008), a creature in captivity may have a budget that differs from that of the wild. In order to
provide optimal welfare, animal keepers have to understand how the species under their care may
respond to different captive conditions (Kleiman, 1994).
In the wild, orangutans are arboreal creatures which travel great distances daily (Galdikas, 1988;
Singleton and van Schaik, 2001). Because their food sources tend to be temporally and spatially
separate, free-living orangutans have to make their way through complex forest environments
(Oates, 1987) in search of food (Wich et al., 2009). Wild orangutans have distinct cultures in
different populations (van Schaik et al., 2003), and their ability to use and fashion tools for a
variety of purposes suggests a high level of intelligence (Parker and Gibson, 1977; van Schaik et
al., 1999). However, due to habitat loss and poaching, both Bornean and Sumatran orangutans are
endangered in the wild (IUCN, 2010). As a result, much effort has been focused on ensuring
welfare and reproductive success in captive populations (Gould, 1983; Mallinson, 1984;
Markham, 1990).

13
In captivity, orangutans are inclined towards obesity due to protein-rich diets (Delgado and Van
Schaik, 2000; Gippoliti, 2000) and a lack of physical activity (Maple, 1979; 1980). Orangutans
tend to become inactive and confined to the ground if not provided with adequate structures for
arboreal activity (Gippoliti, 2000; Pizzutto et al., 2008). Coupled with their curious nature which
lends to their destructive propensity, it is agreed that keeping orangutans both physically and
mentally occupied in captivity is challenging (Markham, 1990). Hence, it is important to have
sufficient knowledge about captive orangutan behaviour, to ensure adequate welfare for this
intelligent ape (Snowdon, 1989).
To date, behavioural studies on captive orangutans tend to focus on cognition (Leavens, 2007),
communication (Cartmill and Byrne, 2010), social behaviour (Poole, 1987; Tobach et al., 1989),
or the effects of enrichment or enclosure modification (Perkins, 1992; Pizzutto et al., 2008; Tripp,
1985). Detailed activity budget studies are rarely published; of the few available studies, the
majority are internal zoo reports or unpublished student projects which are not easily accessible
by the zoo or research community. So far, there have only been a few published papers which
detail the activity budgets of captive orangutans (Forthman et al., 1993; Pearson et al., 2010;
Pizzutto et al., 2008). As behavioural data can be used to gauge if animals have adequate welfare,
as well as ensure that behavioural diversity has not been lost through generations of captive
breeding (Hosey et al., 2009), it is essential to have records of captive orangutan activity that can
be used for further evaluation (Snowdon, 1989). In addition, even though many existing
environmental enrichment and management routines attempt to simulate wild-type behaviour in
captive primates (Britt, 1998; Honess and Marin, 2006; Marriner and Drickamer, 1994), there
have been surprisingly few comparative studies on wild versus captive primates (Melfi and
Feistner, 2002; Todd et al., 2008). Hence, by comparing captive orangutan activity budgets with
those of wild animals, we can shed light on how the zoo environment has affected orangutan
behaviour (Hosey et al., 2009). Therefore, this study is designed to elucidate the activity budgets

14
of captive orangutans as well as provide baseline data for future comparisons. The activity
budgets of two captive orangutan groups were studied in Singapore Zoo, and comparisons were
made against wild orangutan data as well as orangutans from other zoos. The activity budgets of
animals in different age groups were also compared.
In addition, there is currently a trend in zoos worldwide to provide naturalistic exhibits for
animals. , are designed
to encourage wild-type behaviours in captive animals, as well as educate visitors by fostering an
appreciation of animals in their natural setting (Coe, 1989; Hosey et al., 2009). Although such
exhibits have burgeoned worldwide; to date, there have been only a handful of studies (mostly on
primates) on how such enclosures influence animal behaviour (Stoinski et al., 2001). There is a
need to learn how these naturalistic and often, more structurally complex enclosures (Ogden et
al., 1990) may affect the behaviour of animals (Stoinski et al., 2001). Currently, only a few
published studies exist on orangutan behaviour in naturalistic exhibits (Forthman et al., 1993;
Hebert and Bard, 2000; Pearson et al., 2010). The orangutan exhibit in Singapore Zoo is not only
naturalistic, but also novel in that there is no physical barrier between animals and visitors;
visitors can walk directly under the trees on which the orangutans are found. As a result, the
presence of two novel, naturalistic treetop exhibits in Singapore Zoo, as well as a unique
husbandry routine in which two groups of animals are rotated regularly between two enclosures,
allows us to study not only orangutan behaviour, but also shed light on how naturalistic exhibits
may influence animal behaviour. Hence, this study also investigated the differences in activity
budgets between apes in two naturalistic enclosures.

15
Objectives of study
1. What is the activity budget of the orangutans? How does it differ for animals of different
ages?
2. Does activity budget differ between two naturalistic enclosures?
3. How does the activity budget of Singapore Zoo orangutans compare to that of wild
orangutans and orangutans in other zoos?
M A T E RI A LS A ND M E T H O DS
Study site and data collection
The study was conducted at orangutan enclosures in Singapore Zoo. The zoo introduced two
- naturalistic exhibits in 2006, - (Boardwalk
exhibit) - (Island exhibit) (Raj, 2009, pers. comm.3). Each
exhibit was situated next to main tracks in the zoo, and consisted a row of tall trees joined by
vines, nets, logs, and platforms. Some differences were present between the exhibits. In the Island
exhibit, visitors could only view the orangutans from the main road next to the trees, but in the
Boardwalk exhibit, visitors could see the orangutans from both the main road and an elevated
boardwalk in the exhibit. This boardwalk allowed visitors to come into much closer proximity (up
to 3m) from the animals (see Figs. 2.1, 2.2).
3 Jackson Raj, Head orangutan keeper, Singapore Zoo
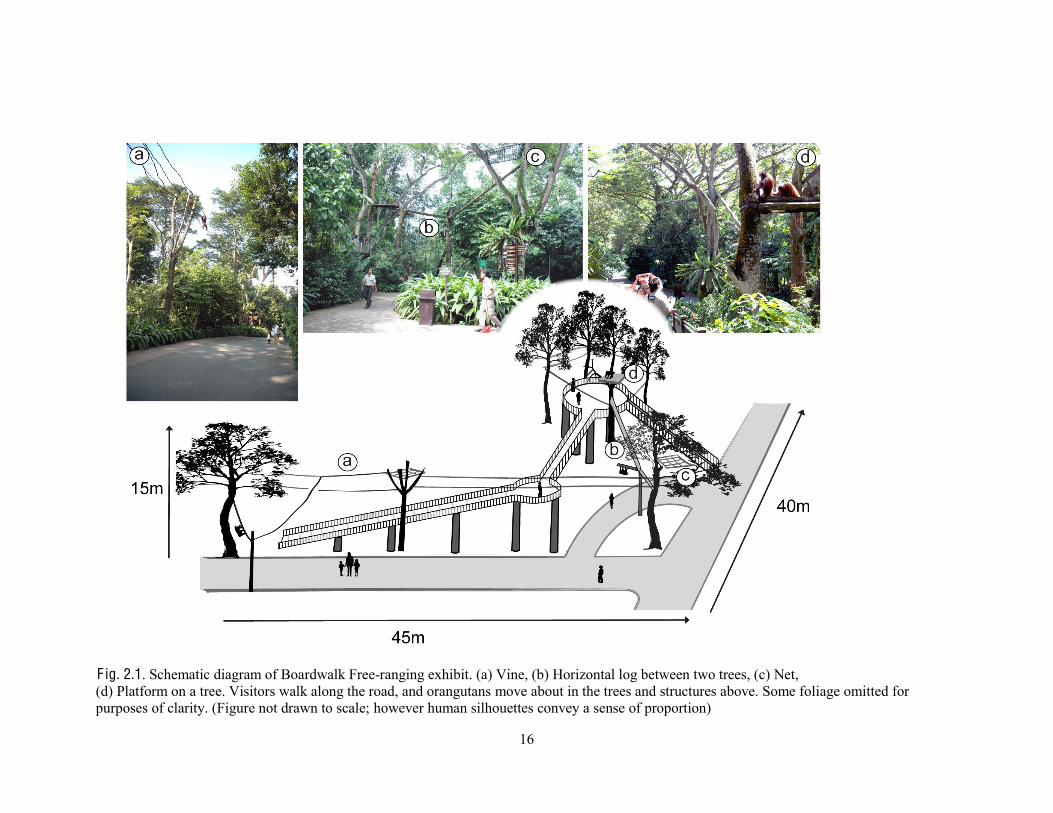
16
F ig. 2.1. Schematic diagram of Boardwalk Free-ranging exhibit. (a) Vine, (b) Horizontal log between two trees, (c) Net, (d) Platform on a tree. Visitors walk along the road, and orangutans move about in the trees and structures above. Some foliage omitted for purposes of clarity. (Figure not drawn to scale; however human silhouettes convey a sense of proportion)

17
F ig. 2.2. Schematic diagram of Island Free-ranging exhibit. (a) Large net, (b) Vine, (c) Horizontal log, (d) Platforms. Visitors walk along the road, and orangutans move about in the trees and structures above. Some foliage omitted for purposes of clarity. (Figure not drawn to scale; however human silhouettes convey a sense of proportion)

18
The other main difference between the enclosures was that visual stimulus from surrounding
features was greater for orangutans in the Boardwalk exhibit than in the Island exhibit. Features
in the vicinity of both exhibits were generally closer to the Boardwalk exhibit (Fig. 2.3). The
latter was situated directly alongside a third orangutan enclosure and a busy restaurant. On the
other hand, the Island exhibit was further from the restaurant and there were trees between the
Island exhibit and the third enclosure, which obstructed the view of the latter from the Island
exhibit. Hence, orangutans when in the Boardwalk exhibit, received more visual stimuli than
when in the Island exhibit.
Busy restaurant with outdoor dining area
ISL
AN
D E
xhib
it
T hird orangutan exhibit
B O ARDWA L K Exhibit
Main track for visitors and vehicles
Tree
F ig. 2.3. Plan diagram showing proximity of Boardwalk and Island exhibits to surrounding features. Orangutans received greater visual stimuli from surrounding features when in the Boardwalk exhibit.

19
Two groups of orangutans were rotated daily between the free-ranging exhibits. The groups were
made up of individuals of varying ages, and each group had one mother-infant pair (Table 2.1).
As the infants were still dependent on their mothers, they were excluded from the observations.
The composition of the groups remained the same throughout the study period. This display
arrangement presented a unique opportunity: usually, studies that compare animal behaviour
across different enclosures use data from different animals; however, in Singapore Zoo, because
the same animals were regularly rotated between two exhibits, I had the rare chance to study the
same subjects in different enclosures, simulating a manipulative experiment.
Data was collected on both weekdays and weekends from October 2009 to February 2010,
between 09:30 h to 17:00 h. Instantaneous scan sampling, with the aid of binoculars, was used for
taken at 10 min intervals were sufficiently far apart enough to avoid autocorrelation. A total of
192 hours of observations (48 hours for each group-exhibit combination) were made. As regular
feeding sessions were held twice daily, at 11:30 h and 15:30 h for the Island exhibit and 14:15 h
and 16:30 h for the Boardwalk exhibit, data collection was paused 15 min before each session
started and only resumed 15 min after the feeding ended. During each scan, the behaviour and
location of individual orangutans was recorded (see Table 2.2 for behaviour categories). All
observational procedures in this study complied with the guidelines of the Institutional Biosafety
Committee (National University of Singapore, NUS) and the NUS Institutional Animal Care and
Use Committee, risk assessment number OSHE/RA/03/04/ FOSo-152.
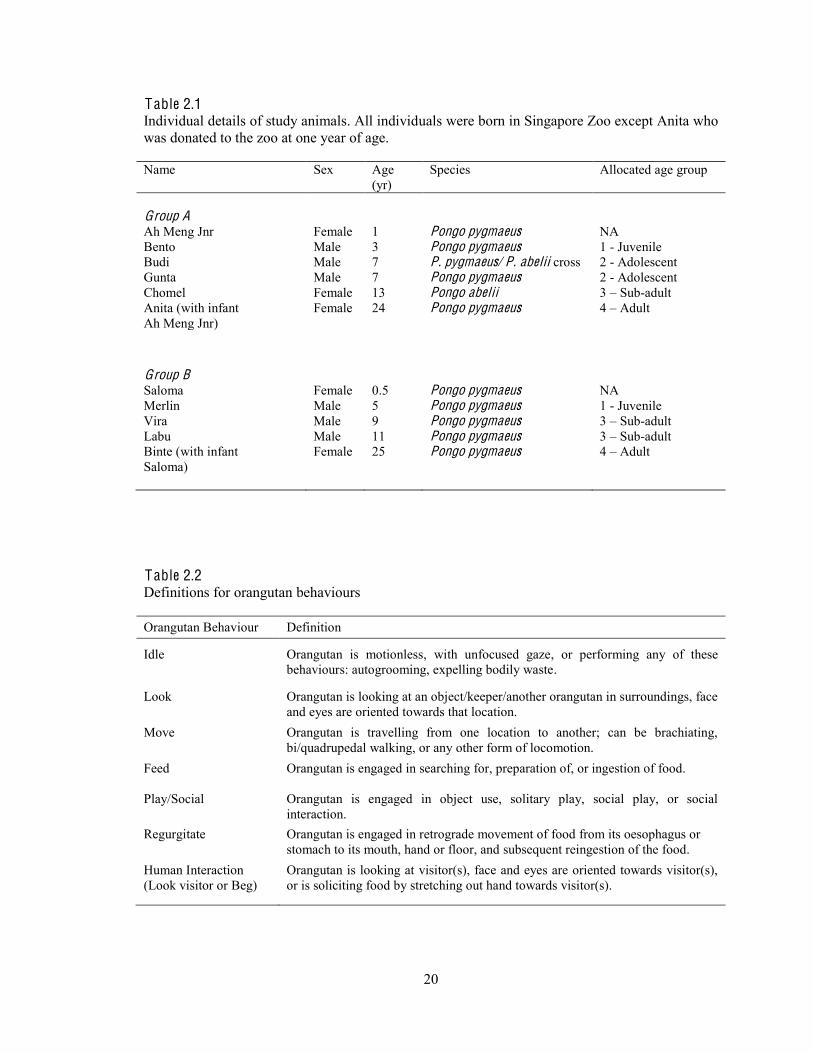
20
Table 2.1 Individual details of study animals. All individuals were born in Singapore Zoo except Anita who was donated to the zoo at one year of age. Name Sex Age
(yr) Species Allocated age group
Group A
Ah Meng Jnr Female 1 Pongo pygmaeus NA Bento Male 3 Pongo pygmaeus 1 - Juvenile Budi Male 7 P. pygmaeus/ P. abelii cross 2 - Adolescent Gunta Male 7 Pongo pygmaeus 2 - Adolescent Chomel Female 13 Pongo abelii 3 Sub-adult Anita (with infant Ah Meng Jnr)
Female 24 Pongo pygmaeus 4 Adult
Group B Saloma Female 0.5 Pongo pygmaeus NA Merlin Male 5 Pongo pygmaeus 1 - Juvenile Vira Male 9 Pongo pygmaeus 3 Sub-adult Labu Male 11 Pongo pygmaeus 3 Sub-adult Binte (with infant Saloma)
Female 25 Pongo pygmaeus 4 Adult
Table 2.2 Definitions for orangutan behaviours Orangutan Behaviour Definition
Idle Orangutan is motionless, with unfocused gaze, or performing any of these behaviours: autogrooming, expelling bodily waste.
Look Orangutan is looking at an object/keeper/another orangutan in surroundings, face and eyes are oriented towards that location.
Move Orangutan is travelling from one location to another; can be brachiating, bi/quadrupedal walking, or any other form of locomotion.
Feed Orangutan is engaged in searching for, preparation of, or ingestion of food.
Play/Social Orangutan is engaged in object use, solitary play, social play, or social interaction.
Regurgitate Orangutan is engaged in retrograde movement of food from its oesophagus or stomach to its mouth, hand or floor, and subsequent reingestion of the food.
Human Interaction (Look visitor or Beg)
Orangutan is looking at visitor(s), face and eyes are oriented towards visitor(s), or is soliciting food by stretching out hand towards visitor(s).

21
Data processing and analysis
The data from both orangutan groups were pooled for each exhibit. The activity budgets for
individual orangutans were calculated by counting the number of scan samples for each
behaviour and converting it to a percentage of the total number of scans. A total activity budget
was calculated for each enclosure by averaging across all orangutans.
To compare behaviour between animals of different ages, the orangutans in the study were
grouped into four age groups based on classifications from Mackinnon (1974) (see Table 2.1).
To investigate if there was any relationship between age group and activity budget (i.e. proportion
of time spent on different activities), a chi-square test of independence was run for age
group against counts of behaviour in each activity. Separate chi-square analyses were run for both
enclosures. Subsequent chi-square tests of independence were then run for each behaviour to see
if the amount of that behaviour different significantly across age groups.
When comparing activity budgets between the Boardwalk and Island exhibits, a Pe -
square test of independence was run for exhibit versus behaviour. Again, individual chi-square
tests were conducted for each behaviour.
For the comparison of captive to wild orangutan behaviour, data was gathered from orangutan
studies which had been conducted in other zoos (Jersey Zoo, UK, Cleveland Metroparks Zoo,
USA, and Zoo Atlanta, USA). Both published (Pearson et al., 2010) and unpublished data
(Cassella, 2010; Marchal, 2004) were used. Captive orangutan behaviour was recategorized for
comparison against the wild orangutan data. The data for wild orangutans came from Morrogh-
Bernard et al. (2009), which was a compilation of orangutan activity budgets from various study
sites in Borneo and Sumatra. The study sites comprised two types of forests, one with regularly

22
fruiting trees, and one with irregularly fruiting trees (masting forests). One activity budget was
calculated from the data for each forest type by averaging across all sites in that forest type. These
data were then used for qualitative comparisons of the captive and wild orangutan groups.
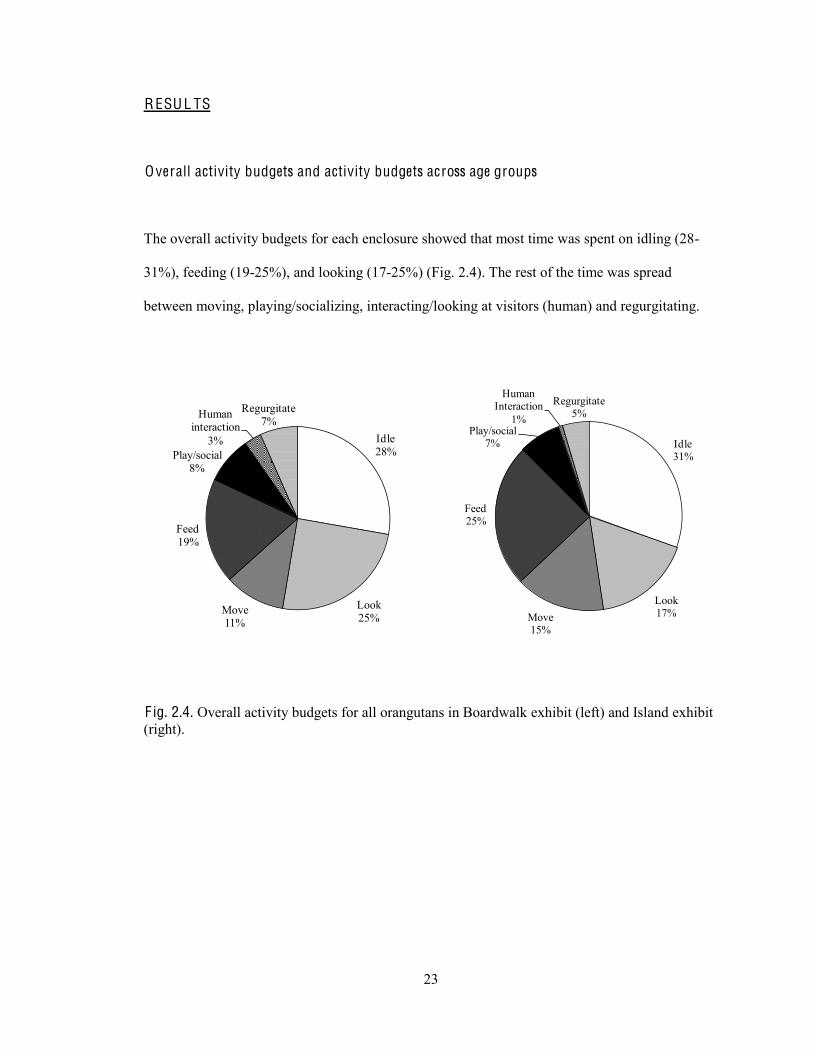
23
R ESU L TS
Overall activity budgets and activity budgets across age groups
The overall activity budgets for each enclosure showed that most time was spent on idling (28-
31%), feeding (19-25%), and looking (17-25%) (Fig. 2.4). The rest of the time was spread
between moving, playing/socializing, interacting/looking at visitors (human) and regurgitating.
s
Idle28%
Look25%
Move11%
Feed19%
Play/social8%
Human interaction
3%
Regurgitate7%
Idle31%
Look17%Move
15%
Feed25%
Play/social 7%
Human Interaction
1%
Regurgitate5%
F ig. 2.4. Overall activity budgets for all orangutans in Boardwalk exhibit (left) and Island exhibit (right).
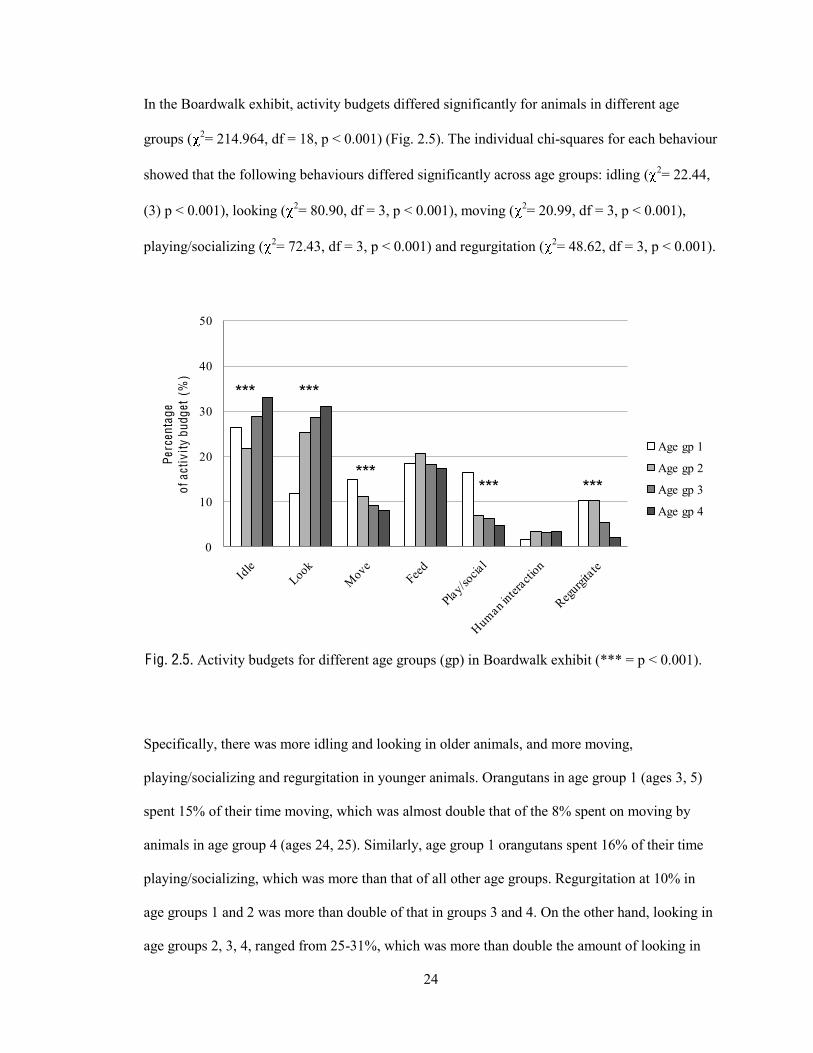
24
In the Boardwalk exhibit, activity budgets differed significantly for animals in different age
groups ( 2= 214.964, df = 18, p < 0.001) (Fig. 2.5). The individual chi-squares for each behaviour
showed that the following behaviours differed significantly across age groups: idling ( 2= 22.44,
(3) p < 0.001), looking ( 2= 80.90, df = 3, p < 0.001), moving ( 2= 20.99, df = 3, p < 0.001),
playing/socializing ( 2= 72.43, df = 3, p < 0.001) and regurgitation ( 2= 48.62, df = 3, p < 0.001).
0
10
20
30
40
50
Perc
enta
ge
of a
ctiv
ity b
udge
t (%
)
Age gp 1
Age gp 2
Age gp 3
Age gp 4
*** ***
*** *** ***
F ig. 2.5. Activity budgets for different age groups (gp) in Boardwalk exhibit (*** = p < 0.001).
Specifically, there was more idling and looking in older animals, and more moving,
playing/socializing and regurgitation in younger animals. Orangutans in age group 1 (ages 3, 5)
spent 15% of their time moving, which was almost double that of the 8% spent on moving by
animals in age group 4 (ages 24, 25). Similarly, age group 1 orangutans spent 16% of their time
playing/socializing, which was more than that of all other age groups. Regurgitation at 10% in
age groups 1 and 2 was more than double of that in groups 3 and 4. On the other hand, looking in
age groups 2, 3, 4, ranged from 25-31%, which was more than double the amount of looking in
25
group 1 (12%). Similarly, idling was highest in age groups 3 and 4 at around 30%. Feeding ( 2=
2.90, df = 3, p = 0.407) and human interaction ( 2= 5.27, df = 3, p = 0.153) did not differ
significantly across the age groups (Fig. 2.5).
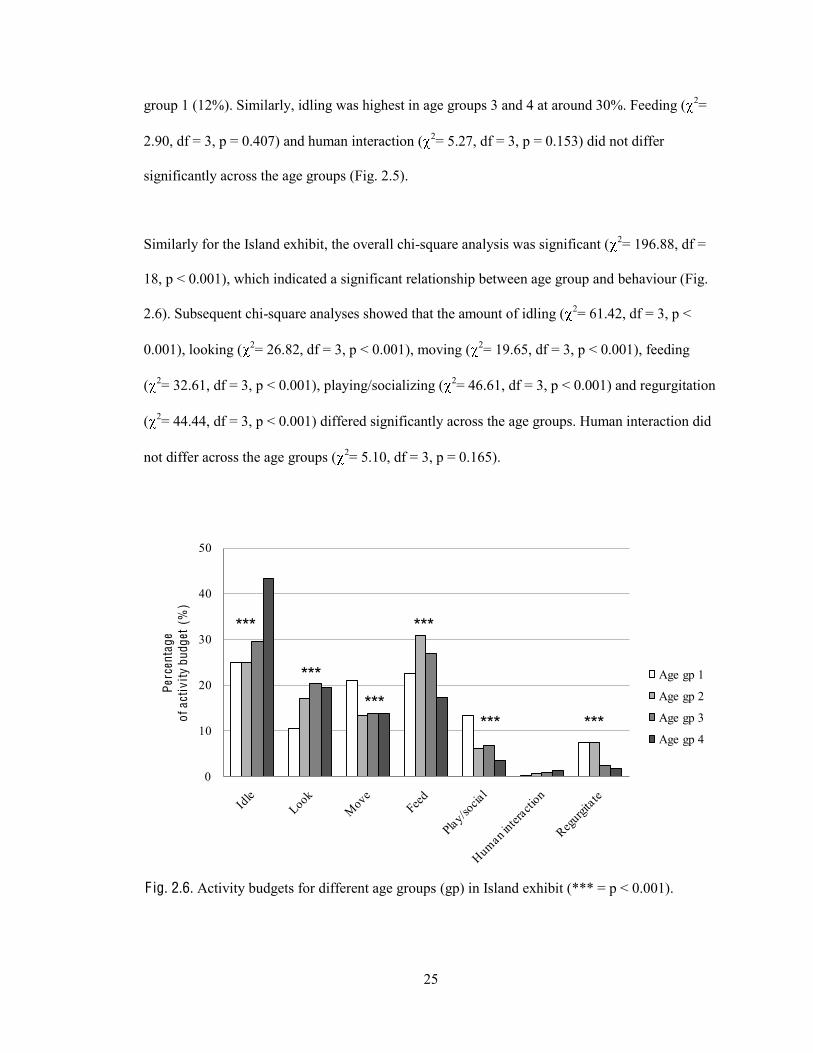
Similarly for the Island exhibit, the overall chi-square analysis was significant ( 2= 196.88, df =
18, p < 0.001), which indicated a significant relationship between age group and behaviour (Fig.
2.6). Subsequent chi-square analyses showed that the amount of idling ( 2= 61.42, df = 3, p <
0.001), looking ( 2= 26.82, df = 3, p < 0.001), moving ( 2= 19.65, df = 3, p < 0.001), feeding
( 2= 32.61, df = 3, p < 0.001), playing/socializing ( 2= 46.61, df = 3, p < 0.001) and regurgitation
( 2= 44.44, df = 3, p < 0.001) differed significantly across the age groups. Human interaction did
not differ across the age groups ( 2= 5.10, df = 3, p = 0.165).
0
10
20
30
40
50
Perc
enta
ge
of a
ctiv
ity b
udge
t (%
)
Age gp 1
Age gp 2
Age gp 3
Age gp 4
*** ***
*** *** ***
***
F ig. 2.6. Activity budgets for different age groups (gp) in Island exhibit (*** = p < 0.001).

26
Again, older orangutans idled and looked more while younger animals moved, played/socialized
and regurgitated more. Feeding was highest in age groups 2 and 3. These results are similar to
that from the Boardwalk exhibit, except for feeding.
Although there were trends in behaviour across age groups, it is also interesting to note that some
orangutans in the same age group differed greatly in their behaviour. See the Appendix for more
information on individual orangutan behaviour.
27
Comparing activity budgets across enclosures
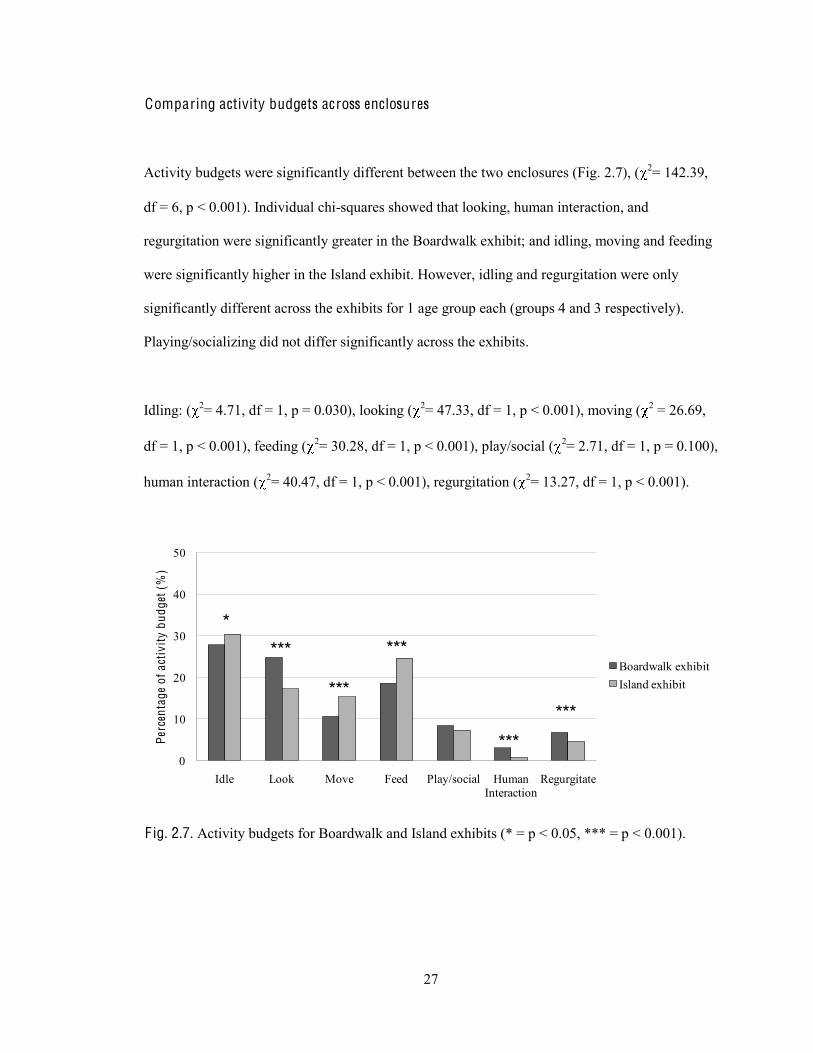
Activity budgets were significantly different between the two enclosures (Fig. 2.7), ( 2= 142.39,
df = 6, p < 0.001). Individual chi-squares showed that looking, human interaction, and
regurgitation were significantly greater in the Boardwalk exhibit; and idling, moving and feeding
were significantly higher in the Island exhibit. However, idling and regurgitation were only
significantly different across the exhibits for 1 age group each (groups 4 and 3 respectively).
Playing/socializing did not differ significantly across the exhibits.
Idling: ( 2= 4.71, df = 1, p = 0.030), looking ( 2= 47.33, df = 1, p < 0.001), moving ( 2 = 26.69,
df = 1, p < 0.001), feeding ( 2= 30.28, df = 1, p < 0.001), play/social ( 2= 2.71, df = 1, p = 0.100),
human interaction ( 2= 40.47, df = 1, p < 0.001), regurgitation ( 2= 13.27, df = 1, p < 0.001).
0
10
20
30
40
50
Idle Look Move Feed Play/social Human Interaction
Regurgitate
Perc
enta
ge o
f act
ivit
y bu
dget
(%
)
Boardwalk exhibitIsland exhibit
****
***
***
******
F ig. 2.7. Activity budgets for Boardwalk and Island exhibits (* = p < 0.05, *** = p < 0.001).
28
Comparing captive to wild activity budgets
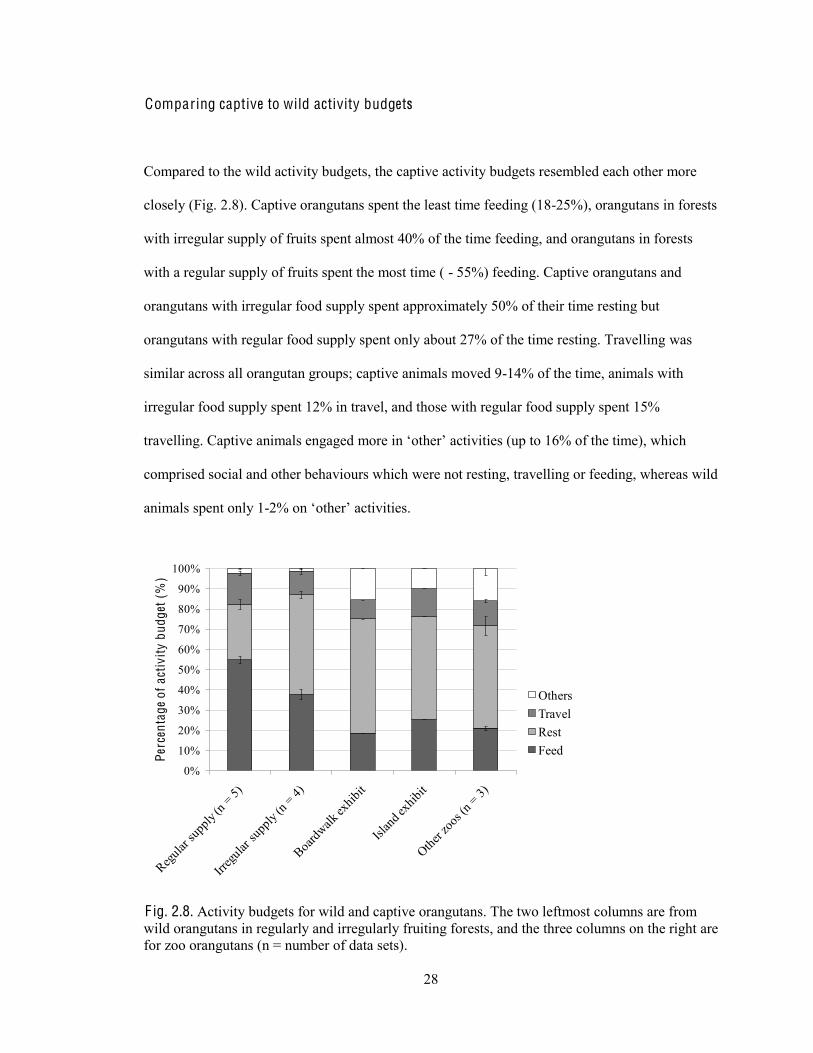
Compared to the wild activity budgets, the captive activity budgets resembled each other more
closely (Fig. 2.8). Captive orangutans spent the least time feeding (18-25%), orangutans in forests
with irregular supply of fruits spent almost 40% of the time feeding, and orangutans in forests
with a regular supply of fruits spent the most time ( - 55%) feeding. Captive orangutans and
orangutans with irregular food supply spent approximately 50% of their time resting but
orangutans with regular food supply spent only about 27% of the time resting. Travelling was
similar across all orangutan groups; captive animals moved 9-14% of the time, animals with
irregular food supply spent 12% in travel, and those with regular food supply spent 15%
comprised social and other behaviours which were not resting, travelling or feeding, whereas wild
animals spent only 1-
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Perc
enta
ge o
f act
ivity
bud
get (
%)
OthersTravelRestFeed
F ig. 2.8. Activity budgets for wild and captive orangutans. The two leftmost columns are from wild orangutans in regularly and irregularly fruiting forests, and the three columns on the right are for zoo orangutans (n = number of data sets).

29
DISC USSI O N
This study investigated orangutan activity budgets across age groups, enclosure designs, and
between wild and captive orangutans. Because it is challenging to keep orangutans in captivity,
and detailed activity budget data for captive animals are rare, activity budget data were collected
on a group of captive orangutans to improve existing knowledge and provide baseline data for
future comparisons. The results showed that orangutan activity budgets differ across age groups,
and are affected by enclosure design. Even though captive activity budgets were not exactly the
same as wild activity budgets, the data from captive animals were surprisingly similar to that of
wild orangutans in irregularly fruiting forests.
Comparing activity budgets across age groups
When comparing behaviours across age groups, the younger orangutans in this study played and
moved significantly more than older animals. Specifically, juveniles (aged 3 and 5) spent the
most amount of time moving and playing, as compared to older animals which tended to idle and
look more at their surroundings. Comparing this study to the few existing captive studies with
detailed activity budgets (Cassella, 2010; Marchal, 2004; Pearson et al., 2010), we see the same
general trends: idling and looking is generally predominant in the older groups, and movement
and play is greater in younger orangutans.
In the wild, adult orangutans are semi-solitary. They have wide home ranges which may
sometimes overlap with that of conspecifics. Therefore, social contact is irregular between mature
animals (Mackinnon, 1974; Rijksen, 1975) and they are not known to play (Mackinnon, 1974).
However in captivity, adult orangutans have been found to engage in social behaviour under
group settings (Poole, 1987; Zucker et al., 1978; 1986). On the other hand, sub-adult orangutans

30
are known to play under free-living conditions (Galdikas, 1985b), and play generally allows
young animals to develop motor and social skills important for survival (Fagen, 1981; Smith,
1978). Hence, we would expect more play and social interactions in immature orangutans
(Mackinnon, 1974). Conversely, alertness to surroundings (looking) was more prevalent in older
animals, and at very low levels in the juvenile orangutans. This is similar to what has been found
in a variety of other species. Dominant wild vervet monkeys in several studies were found to scan
their surroundings more than subordinate animals. This was thought to be a consequence of
having a greater need to protect their kin, as well greater involvement in intergroup resource
competition (Isbell and Young, 1993). Such age-related patterns may explain why attentiveness to
surroundings was found to be lowest in the youngest age group of the present study.
The findings from this study mirror that of previous investigations on wild orangutans, as well as
sociality and play in captive apes. It has been found that younger orangutans are generally more
active, under both captive (Marchal, 2004; Perkins and Bradfield, 1989) and wild settings
(Davenport, 1967; Morrogh-Bernard et al., 2009). Immature animals are also more gregarious
(Galdikas, 1985a; Mackinnon, 1974; Rijksen, 1975) and spend more time playing (Edwards and
Snowdon, 1980; Mackinnon, 1974; Zucker et al., 1986). On the other hand, older orangutans tend
to play less in captivity (Edwards and Snowdon, 1980; Poole, 1987; Zucker et al., 1986) and are
less active both in captivity (Marchal, 2004) and in the wild (Morrogh-Bernard et al., 2009).
The results presented here show that the orangutans in Singapore Zoo display age-specific levels
of behaviour, which reflects positively on the quality of management in this zoo. Such results will
be useful for comparisons with other captive populations to see if captive behaviour deviates
significantly from wild-type behaviour. Such comparisons can be used as welfare indicators for
captive animals (Hosey et al., 2009; Melfi and Feistner, 2002).

31
Comparing activity budgets across enclosures
For this group of orangutans, activity budgets seem to be influenced by two types of factors:
structures within the exhibit, as well as features surrounding the exhibit. Comparing activity
budgets across the two exhibits, orangutans seem to interact more with their surroundings in the
Boardwalk exhibit. As mentioned previously, the Boardwalk exhibit was directly alongside a
(busy) restaurant and a third orangutan exhibit.
further from the restaurant, and the view of the third orangutan exhibit was obscured by several
trees. Because the same environmental features, i.e. the restaurant and the third exhibit were
closer to the Boardwalk than Island exhibit, this resulted in more visitor traffic and orangutan
activity (from the third exhibit) around the Boardwalk exhibit. Hence, the significantly higher
incidences of looking and human interaction when orangutans were in the Boardwalk enclosure
appear to be responses to this particular environment. In addition, the presence of the elevated
visitor boardwalk in this exhibit, which allowed closer proximities between orangutans and
visitors, may also have contributed to the higher human interactions here. Similar results have
been found in other primate studies, where cages nearer the exit and main paths had higher visitor
attendance than cages further from the exit or off the main paths (Mitchell et al., 1990). Also, the
presence of more visitors has been associated with responses from primates which manifest as
greater incidences of aggression (Chamove et al., 1988; Wells, 2005) or the increased use of
enclosure areas near visitors . Not only does enclosure design affect animal behaviour,
environmental features seem to have an effect as well. Information of this nature is especially
useful when designing exhibits for species which may be vulnerable to high levels of noise, or are
easily affected by visitors.
Besides environmental features, the differing arrangement of structures within each exhibit also
seems to have contributed to the dissimilar activity budgets. The higher amounts of regurgitation

32
in the Boardwalk exhibit were most likely due to the location of the platform in this enclosure.
Orangutans in this zoo almost always regurgitated on platforms. The platform in the Boardwalk
exhibit was at the centre, whereas the same structure in the Island exhibit was at one end of the
60m long enclosure. Hence, ease of accessing the platform would have had a strong influence on
amount of regurgitation in each exhibit. Regurgitation occurred more in the Boardwalk exhibit
possibly because the platform was at the centre of the enclosure. Similarly, the spacing and
availability of structural features in the Island exhibit may have contributed to why more
movement was present in that exhibit. The Island exhibit consisted of a straight row of trees, and
the nets, platforms and log in this exhibit were located only at either ends of the exhibit (see Fig.
2.2). In addition, the trees in the centre of the row had less branches and foliage, as compared to
trees at both ends which had larger crown diameters and more foliage and branches. As a result,
the orangutans were observed to spend more time at the sides of the exhibit. Because the row of
trees spanned a good 60m in length, we would expect the orangutans to move more in this exhibit
when getting from one end to the other. In contrast, structural features in the Boardwalk exhibit
were spread more evenly throughout. Similarly, because a huge net was present in the Island
exhibit, but not in Boardwalk exhibit, and heavy utilization of this net was observed, it is possible
that the presence of this structural feature contributed to higher rates of idling (for only age group
4) in the Island exhibit. Lastly, the only plausible reason for more feeding in the latter exhibit, is
that, as compared to the Boardwalk exhibit, the relative lack of environmental stimuli in the
Island exhibit may have resulted in a decrease in other behaviours and a subsequent increase in
feeding behaviour.
It is known that the availability and arrangement of features in an exhibit can affect animal
behaviour. Many studies have found that great apes are more active and display more species-
specific behaviours when in structurally complex enclosures with climbing features, as compared
to barren exhibits (Clarke et al., 1982; Maple and Stine, 1982; Pfeiffer and Koebner, 1978). The

33
location of enclosure features, though not well understood, has also been found to affect how
heavily a feature is used; for example, caimans predominantly used pools in their enclosures and
areas further from the pools were used significantly less (Verdade et al., 2006). From this, we can
see how animal behaviour can be modified providing certain structures or arranging features in a
certain way within an exhibit. This knowledge can be used by zoo management to encourage or
discourage certain behaviours, such as increasing movement or decreasing regurgitation (an
undesirable behaviour, Akers and Schildkraut, 1985, see Chapter 5), or to promote greater use of
the entire exhibit in the species on display (Stoinski et al., 2001). Similarly, features in the
surrounding of an exhibit can be either enrichment or a form of stress for animals. Orangutans are
known to be inquisitive animals, and are often very interested in their surroundings (Hebert,
2011, pers. comm.4; Nantha, 2009, pers. comm.5; pers. obs.). Many anecdotes from zoo keepers,
as well as a study by Bloomfield et al. (2010) suggest that these apes do not actively avoid, and in
fact often spend time watching visitors, as well going-ons outside their enclosure. Hence, features
in the vicinity of an orangutan enclosure could be both enrichment and sources of stress for
captive animals.
Comparing captive to wild activity budgets
Interestingly, the activity budgets of orangutans in captivity were more similar to that of animals
in irregularly fruiting forests than regularly fruiting forests. The amount of time spent feeding
ranged from 18 to 25% in captive animals, as compared to 38% in orangutans of irregularly
fruiting forests and 55% in orangutans of regularly fruiting forests. In the wild, foraging and
ity budget (see review
in Morrogh-Bernard et al., 2009). Orangutans have to locate food sources which are spread
4 Patricia Hebert, Orangutan researcher, Fort Wayne Children's Zoo 5 Gabriel Nantha, Orangutan keeper, Singapore Zoo

34
patchily throughout the forest, and many fruits in the wild require extensive preparation to
separate out the edible from inedible portions (Mackinnon, 1974). However, in captivity, foraging
is reduced to concentrated bouts of feeding during mealtimes where provided foods can usually
be ingested with little preparation. Therefore, it is reasonable to expect that, because orangutans
in Singapore Zoo received only sporadic feedings of leaves and small fruit snacks while on
exhibit, the time spent feeding would be considerably lower than that of wild animals. Also,
because orangutans in the other three zoo studies received only one main meal and sometimes
fresh leaves (browse) during observation periods (Cassella, 2010; Marchal, 2004, Pearson, 2010,
pers. comm.6), feeding behaviours may also be expected to decrease. These results are analagous
to those from another study which compared wild to captive Sulawesi macaques (Macaca nigra),
in which the authors also found that zoo macaques spent less time feeding and more time resting
(Melfi and Feistner, 2002).
Conversely, the amount of resting behaviour in animals from all captive groups, as well as those
from irregularly fruiting forests, was higher than the amount of resting behaviour in animals from
regularly fruiting forests. Due to the lack of foraging opportunities, it is understandable that
captive orangutans spent more time inactive than their wild counterparts (Zucker et al., 1986).
However, what is interesting is that the amount of inactive rest in orangutans with irregular food
supply was not significantly different from that of captive animals. In fact, the main differences in
activity budgets between captive orangutans and wild orangutans with irregular food supply
greater in captive animals and lesser in wild animals. Hence, it appears that the time not spent on
vities such as social and solitary play.
Because all of the captive orangutans in this study were kept in groups of two or more, there were 6 Elissa Pearson, Orangutan researcher, Adelaide Zoo

35
regular opportunities for social interactions which may not be present for wild animals (Zucker et
al., 1986). As a result of this inactivity brought about by the lack of foraging opportunities, as
well as the presence of social partners in captivity, there is a corresponding change in captive
orangutan activity budgets, i.e. less feeding and more play in captive animals. Such reallocation
of activity budgets has been observed in the comparison of provisioned and unprovisioned
primate groups. Savannah baboons (Papio cynocephalus) and vervet monkeys (Cercopithecus
aethiops) which had access to human sources of food spent only about 20% of the time feeding,
and about 50-60% of the time resting. This was in contrast to unprovisioned groups which spent
40-60% of the time feeding, and 10% of their time resting (Altmann and Muruthi, 1988; Brennan
et al., 1985; Lee et al., 1986). Because human food sources such as tourists or garbage dumps
provide easily accessible, spatially concentrated, and relatively nutrition-rich sources of food; as a
result, provisioned animals spend less time procuring the same amount of nutrition. Naturally,
provisioned groups had reduced feeding times, and could afford more time on other activities
such as socializing and resting. Social stimulation may serve as an alternative to reduce inactivity
in captive animals (Perkins, 1992; Wilson, 1982) and may compensate for the lack of captive
environmental complexity relative to that of free-ranging conditions (Poole, 1987).
Lastly, it is encouraging to see that the amount of time spent travelling in captivity was
comparable to that of wild conditions. Although animals in captivity have restricted living space,
these results show that captive orangutans may still have the opportunity to maintain similar
levels of movement as wild counterparts. One possible factor could have been the naturalistic
design of the orangutan enclosures under study. All the orangutan enclosures were large, between
700-1500 m2 in area, and had structures which provided arboreal opportunities (Cassella, 2010;
Marchal, 2004; Pearson et al., 2010). It is known that the presence of complex enclosures, social
partners, and manipulatable objects can increase activity in captive orangutans (Perkins, 1992;
Tripp, 1985; Wilson, 1982), and my results also support this. Because captive apes have a

36
propensity for inactivity and obesity in captivity (Gippoliti, 2000), such findings are encouraging
for zoo managers. Hence, systematic research on the influence of factors such as enclosure usage
may be an important next step to take in optimizing captive animal welfare. The following
chapter investigates the relationship between enclosure structures and animal behaviour to shed
further light on this topic.
Overall, the activity budgets of the captive orangutans seem to be comparable to that of the wild,
which is a good indication for captive orangutan management. Activity budgets give a broad,
although simplistic overview of behavioural expression, which can be used to determine if
captive individuals are deviating from the wild-type pattern, as well as serve as indices of captive
welfare (Hosey et al., 2009).

37
R E C O M M E ND A T I O NS
It should be possible to increase desirable behaviours, and reduce undesirable behaviours
by manipulating the location of structures in an exhibit. For example, placing a platform
at a far end of an enclosure may reduce regurgitation. Also, placing structures far apart in
an enclosure may encourage more movement and less inactivity in captive animals.
It may be feasible to use features surrounding the exhibit as a form of enrichment, and
factor this into exhibit design when deciding where to locate an orangutan exhibit.
Similarly, if the animals are sensitive to noise or visitors, care can be taken in deciding
where to locate an exhibit.
Reduce inactivity by increasing opportunities for feeding, foraging and movement. For
example, increase the complexity, variety and frequency of foods provided. Opportunities
for arboreal movement could also be provided, for example by using live trees in the
exhibit, or providing structures such as sway poles, which allow orangutans to move
between these flexible poles by bending the poles with their body weight. (See next
chapter)

38
C O N C L USI O NS
The activity budgets of the orangutans were found to differ significantly across age
groups. Younger animals were more active and played more, older animals idled and
looked more. Singapore Zoo orangutans show age-specific behaviours, which is a
positive note on orangutan welfare.
Activity budgets differed significantly across enclosures. Both exhibit structures and
environmental features influenced activity budgets.
The activity budgets of zoo orangutans were similar in some ways to that of wild
orangutans. Captive orangutans fed less, rested more, and had comparable amounts of
movement with wild orangutans. Records of captive activity budgets are useful
references for zoo management.

39
C H APT E R 3
Use of Enclosure Structures and Vertical Space
in Two Naturalistic O rangutan Exhibits
Abstract. Animals in captivity are subjected to spatial and behavioural limitations. Research has
examined space use in zoo exhibits, but how animals interact with structures is still poorly
understood. The majority of space use studies have been conducted on chimpanzees and gorillas,
and little is known about the space use of other species. In this study, I investigated the use of
enclosure structures, such trees, vines, nets, platforms and logs in two naturalistic orangutan
exhibits. Proportion of time spent, and behaviours occurring at each structure type were noted.
The time spent by orangutans at different vertical heights in the enclosure was also recorded.
Results showed that structure use varied with age and dominance status. The properties,
availability and location of structures in an exhibit could affect the behaviours which occurred at
each structure type, as well as how frequently the structure was used. Husbandry routines and
features in the surrounding of the exhibits also influenced structure use. Vertical space use was
affected by certain structures, primarily, the properties of tree branches and availability of nets
and platforms at different heights. Recommendations are made on how to encourage behavioural
diversity and optimize exhibit use in captive orangutans.
K eywords: orangutan, naturalistic exhibit, enclosure use, space use, structure use, exhibit design, vertical space

40
IN T R O DU C T I O N
Animals in captivity are often subjected to spatial and behavioural limitations. The arrangement
of structural features in a zoo enclosure can determine how animals move or behave (Jensvold et
al., 2001), and biologically relevant environments can only be created when we understand the
requirements of each species (Estevez and Christman, 2006). However to date, knowledge on
space use in many captive species is still lacking (Leighty et al., 2010; Stoinski et al., 2001).
Over the past two centuries, zoos have begun to incorporate the behavioural and psychological
needs of animals in exhibit design. First-generation exhibits of small, barren cages and deep
smooth walled pits have given way to second-generation exhibits which are more spacious, but
still usually cement enclosures surrounded by dry or wet moats. The most recent third-generation
exhibits, however, are more complex, and aim to display animals in naturalistic settings (Shettel-
Neuber, 1988). Such enclosures simulate the natural habitats of the species by making extensive
use of plants and natural features and housing animals in social groups similar to that of their
free-ranging counterparts (Shettel-Neuber, 1988). On the whole, there has been a distinct
movement to increase the size and environmental complexity of zoo exhibits (Hosey et al., 2009).
Complex exhibits have been shown to promote higher levels of activity and species-typical
behaviours, as well as decrease abnormal behaviours in a variety of species. These include
chimpanzees (Brent et al, 1991; Jensvold et al 2001; Pfeiffer and Koebner, 1978), orangutans
(Maple and Stine, 1982, Perkins, 1992; Tripp, 1985; Wilson, 1982), gorillas (Hoff et al, 1994;
Maple and Stine, 1982, Wilson, 1982; Bowen, 1980), hanuman langurs (Little and Sommer,
2002) and Indian leopards (Mallapur and Chellam, 2002). Similarly, naturalistic exhibits also
increase species specific behaviours (Maple and Stine, 1982; Pfeiffer and Koebner, 1978),
decrease stereotypic behaviours (Akers and Schildkraut, 1985; Erwin and Deni, 1979; Gould and

41
Bres, 1986), improve aggression and affiliation in a variety of primates (Clarke et al., 1982;
Maple and Finlay, 1989; Maple and Stine, 1982), and improve visitor appreciation of animal
ecology and behaviour (Price et al., 1994; Stoinski et al., 2001). Although we can see how
naturalistic and complex exhibits benefit both zoo animals and visitors, there is still a lack of
information on how animals interact with individual exhibit features. Because naturalistic exhibits
are often more costly to build and require more maintenance (Shettel-Neuber, 1988; Stoinski et
al., 2001), and with the increase in naturalistic habitats worldwide, there is a need for further
study on this topic (Ogden et al., 1990). Knowledge gained from enclosure use studies can then
be incorporated into future exhibit design to optimize the welfare of captive animals
(Seidensticker and Doherty, 1996; Shettel-Neuber, 1988).
To date, the scientific study of space use is still in its infancy (Ogden et al., 1990; Stoinski et al.,
2001). Of the existing research, the majority has focused on chimpanzees (Bettinger and Carter,
1994; Goff et al., 1994; Ross et al., 2009; 2011) and gorillas (Ogden et al., 1993; Ross et al.,
2009; 2011; Stoinski et al., 2001), with a few on orangutans (Hebert and Bard, 2000; Pearson et
al., 2010). There have also been single studies on a variety of other mammals (wild boars:
Blasetti et al., 1988; sloth bears: Forthman and Bakeman, 1992; manatees: Horikoshi-Beckett and
Schulte, 2006; South American tapir: Mahler , 1984; American bison: Robitaille and Prescott,
1993). Findings have shown that primates have preferences for walls, corners and vertical
structures (James-Aldridge and Gorena, 1991; Ogden et al., 1993) and adult and juvenile animals
have differential space use (Blasetti et al., 1988; Perkins and Bradfield, 1989; Traylor-Holzer and
Fritz, 1985). Certain exhibit structures or areas tend to be associated with particular behaviours,
such as Indian leopards which used enriched areas in their exhibit for active behaviours (Mallapur
and Chellam, 2002) and wild boars which used shaded and muddy areas for resting (Blasetti et
al., 1988). For the majority of species, however, there is still a lack of understanding on detailed
space use and how animals interact with structures (Leighty et al., 2010; Stoinski et al., 2001).

42
In Singapore Zoo, two naturalistic orangutan exhibits of novel design, believed to be the first of
their kind in the world, were established in 2006. Unlike typical exhibits which contain apes by
using indoor enclosures or moated islands, these exhibits are based entirely in the outdoor canopy
of several trees. Orangutans move about in the canopy above the visitors, and low-current
electrical wires at the base of trees as well as large gaps between exhibit and surrounding
vegetation aid in containing the orangutans. Because such a design is new as well as naturalistic,
it provides the unique opportunity to study space use in an orangutan exhibit that is entirely
arboreal. Orangutans are animals with large home ranges in the wild, and they are known to
traverse large distances over tree canopies in search of food (Wich et al., 2009). Correspondingly,
arboreal use of space has been found to be an important factor in captive orangutan enclosures
(Maple, 1979; Maple and Stine, 1982). Because a lack of activity space can lead to lethargy and
obesity (Gippoliti, 2000; Pizzutto et al., 2008), there is a need to provide adequate climbing
opportunities in captive orangutans (Gippoliti, 2000; Hebert and Bard, 2000; Maple and Stine,
1982). However, most studies on enclosure use so far have focused on chimpanzees and gorillas.
Because these apes live in terrestrial habitats, unlike the forest-dwelling orangutans (Wich et al.,
2009); to date, we still have insufficient knowledge on orangutan space use.
Of the few orangutan studies on enclosure use, most have focused on the percentage of enclosure
utilized and time spent off the ground or at certain structures (Manning, 2002; Perkins and
Bradfield, 1989), with no details on what behaviours go on at different structures. Only one study
has systematically observed the behaviour of orangutans at different vertical heights in an
enclosure (Hebert and Bard, 2000). There is a dearth of information on structure use in
orangutans; hence the presence of a naturalistic exhibit with nine animals provides an excellent
opportunity to learn more about space use in captive orangutans. This study is the first to
systematically investigate the use of structures (trees, vines, nets, platforms, logs) in a naturalistic
enclosure, and the behaviours which occur at each type of exhibit structure. The use of different

43
exhibit heights is also elucidated in this novel exhibit to see if there are any preferences for
vertical heights between age groups. Such information will be useful for captive orangutan
management, and can aid in the design of future orangutan enclosures.
Objectives of study
1. What is the proportion of use for different structure types? Does structure use differ
across age groups and exhibits?
2. What activities/behaviours occur at different structures? Does it differ across age groups
and exhibits?
3. What is the proportion of time spent at different vertical heights of the enclosure? Does it
differ across age groups and exhibits?

44
M A T E RI A LS A ND M E T H O DS
Study site and subjects
The study site and subjects were the same as in the previous chapter. To recap, a group of
orangutans in two treetop, naturalistic enclosures of novel design were studied, to see how they
utilized different structures in each exhibit. The two enclosures, the Boardwalk and Island
exhibits, each consisted trees, vines, nets, platform(s) and a log. However there were differences
in the arrangement of structures within each exhibit, as well as dissimilarities in the
environmental features surrounding the two exhibits (Figs. 3.1, 3.2). The Boardwalk exhibit was
made of an L-shaped row of trees. The structures in this exhibit were spread evenly throughout,
with a log and platform at the centre of the exhibit, and two other small nets spread throughout
the exhibit. On the other hand, the Island exhibit was made up of a straight row of trees. The log
and platforms in this exhibit were located at one end together with a small net, and at the other
end of the exhibit were two other nets, one large, one small. Hence the structures in the Island
exhibit were concentrated at either end of the exhibit, as compared to the Boardwalk exhibit,
which had structures evenly throughout.
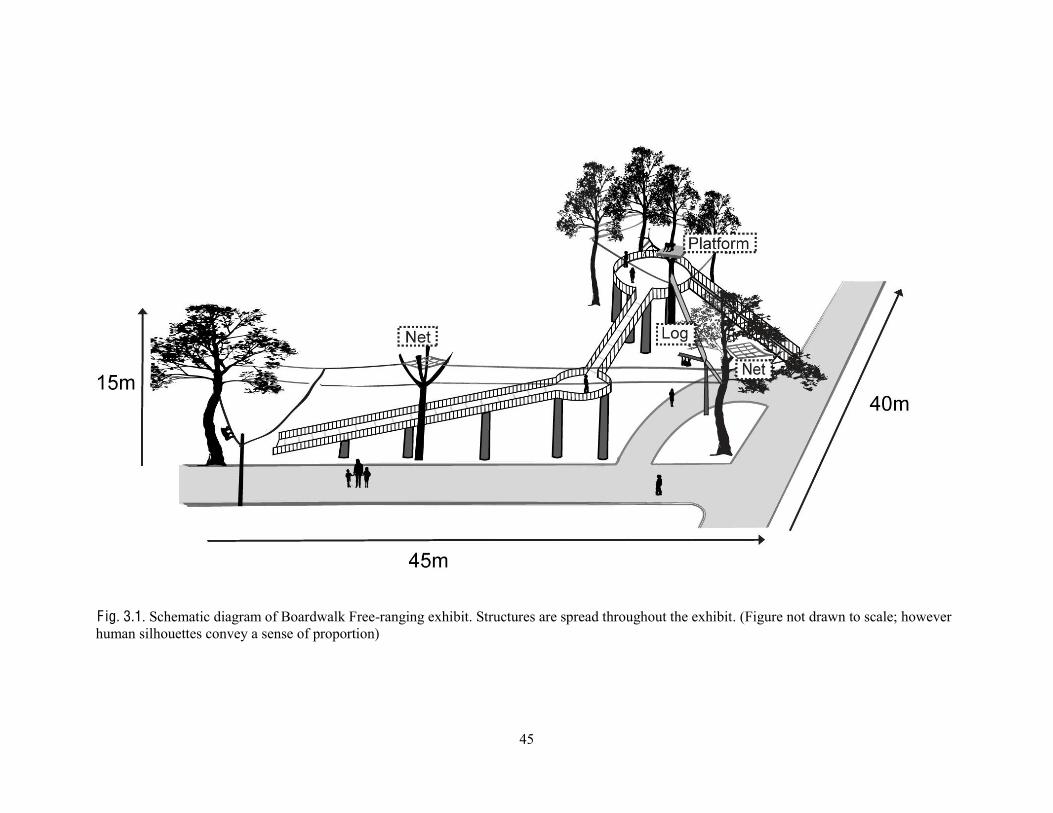
45
F ig. 3.1. Schematic diagram of Boardwalk Free-ranging exhibit. Structures are spread throughout the exhibit. (Figure not drawn to scale; however human silhouettes convey a sense of proportion)

46
F ig. 3.2. Schematic diagram of Island Free-ranging exhibit. Structures are clustered on either end of the exhibit. (Figure not drawn to scale; however human silhouettes convey a sense of proportion)

47
Again, the other main difference between the enclosures was that visual stimulus from
surrounding features was greater for orangutans in the Boardwalk exhibit than in the Island
exhibit. Features in the vicinity of both exhibits were generally closer to the Boardwalk exhibit
(Fig 3.3). The latter was situated directly alongside a third orangutan enclosure and a busy
restaurant. On the other hand, the Island exhibit was further from the restaurant and there were
trees between the Island exhibit and the third enclosure, which obstructed the view of the latter
from the Island exhibit. Hence, orangutans when in the Boardwalk exhibit, received more visual
stimuli than when in the Island exhibit.
Busy restaurant with outdoor dining area
ISL
AN
D E
xhib
it
T hird orangutan exhibit
B O ARDWA L K Exhibit
Main track for visitors and vehicles
Tree
F ig. 3.3. Plan diagram showing proximity of Boardwalk and Island exhibits to surrounding features. Orangutans received greater visual stimuli from surrounding features when in the Boardwalk exhibit.

48
Data collection and processing
The data collected in the previous chapter was analyzed to investigate enclosure use patterns in
the same group of orangutans. Orangutans were again classified into four age groups based on
Mackinnon (1974) (see Table 3.1). As mentioned, each exhibit was made up of several structures,
namely trees, vines, nets, platform(s) and a log. Orangutan location was classified according to
the type of structure that each animal was found on. Use of enclosure structures was then
summarized by counting the number of scan samples for each structure type and converting it to a
percentage of the total scans. A summary of overall structure use was done for each enclosure.
Data analysis
Structure use of the different age groups was compared with a hi-square test of
independence, and separate chi-square analyses were run for each enclosure. Following that, the
behaviours at each structure were investigated. For each structure type (tree, vine, net, platform,
or log), the activity budgets of different age groups were plotted. The behaviour categories used
here were similar to those used in Chapter 2, except that the category of play/social was spilt into
two separate categories for more detailed analysis (see Table 3.2).
Lastly, to investigate vertical use of the exhibits, each exhibit was divided vertically into three
leve
the bottom one-
one-
structures at that height. A P -square test was run for age group against height use, for
both enclosures.

49
Table 3.1 Individual details and age groups of study animals. All individuals were born in the Singapore Zoo except Anita who was donated to the zoo at one year of age. Name Sex Age
(yr) Species Allocated age group
Group A
Ah Meng Jnr Female 1 Pongo pygmaeus NA Bento Male 3 Pongo pygmaeus 1 - Juvenile Budi Male 7 P. pygmaeus/ P. abelii cross 2 - Adolescent Gunta Male 7 Pongo pygmaeus 2 - Adolescent Chomel Female 13 Pongo abelii 3 Sub-adult Anita (with infant Ah Meng Jnr)
Female 24 Pongo pygmaeus 4 Adult
Group B Saloma Female 0.5 Pongo pygmaeus NA Merlin Male 5 Pongo pygmaeus 1 - Juvenile Vira Male 9 Pongo pygmaeus 3 Sub-adult Labu Male 11 Pongo pygmaeus 3 Sub-adult Binte (with infant Saloma)
Female 25 Pongo pygmaeus 4 Adult
Table 3.2 Definitions for orangutan behaviours Orangutan Behaviour Definition Idle Orangutan is motionless, with unfocused gaze, or performing any of these
behaviours: autogrooming, expelling bodily waste. Look Orangutan is looking at an object/keeper/another orangutan in surroundings,
face and eyes are oriented towards that location. Move Orangutan is travelling from one location to another; can be brachiating,
bi/quadrupedal walking, or any other form of locomotion. Feed Orangutan is engaged in searching for, preparation of, or ingestion of food.
Autoplay Orangutan is engaged in object use or solitary play.
Social Orangutan is engaged in social play, or social interactions, inclusive of
Regurgitate Orangutan is engaged in retrograde movement of food from its oesophagus or stomach to its mouth, hand or floor, and subsequent reingestion of the food.
Human Interaction Orangutan is looking at visitor(s), face and eyes are oriented towards visitor(s), or is soliciting food by stretching out hand towards visitor(s).
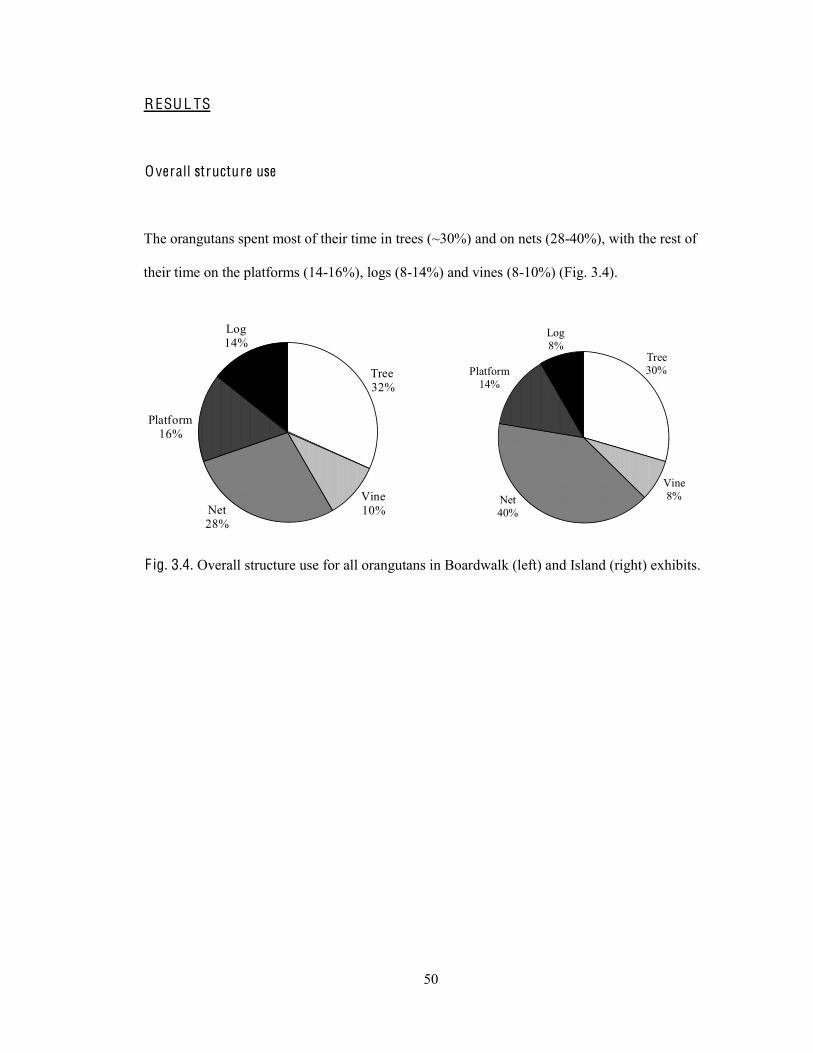
50
R ESU L TS
Overall structure use
The orangutans spent most of their time in trees (~30%) and on nets (28-40%), with the rest of
their time on the platforms (14-16%), logs (8-14%) and vines (8-10%) (Fig. 3.4).
Tree32%
Vine10%Net
28%
Platform16%
Log14%
Tree30%
Vine8%Net
40%
Platform14%
Log8%
F ig. 3.4. Overall structure use for all orangutans in Boardwalk (left) and Island (right) exhibits.
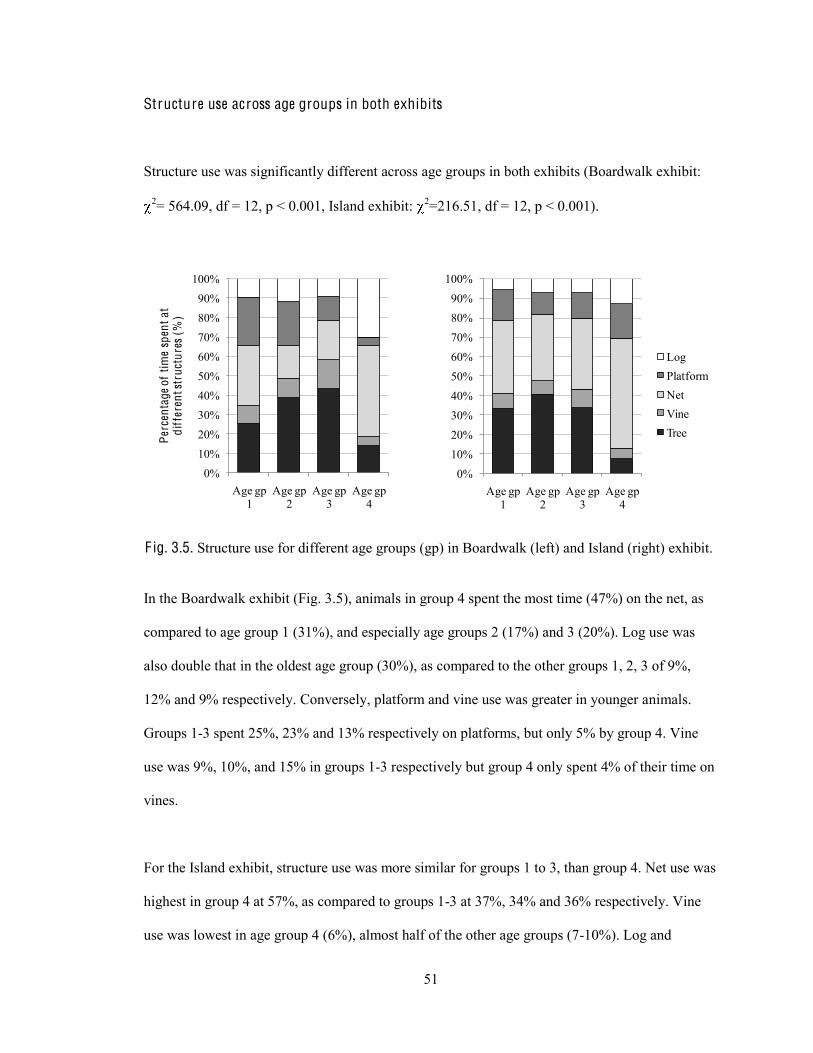
51
Structure use across age groups in both exhibits
Structure use was significantly different across age groups in both exhibits (Boardwalk exhibit:
2= 564.09, df = 12, p < 0.001, Island exhibit: 2=216.51, df = 12, p < 0.001).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Perc
enta
ge o
f tim
e sp
ent a
t di
ffer
ent s
truc
ture
s (%
)
0%10%20%30%40%50%60%70%80%90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
LogPlatformNetVineTree
F ig. 3.5. Structure use for different age groups (gp) in Boardwalk (left) and Island (right) exhibit.
In the Boardwalk exhibit (Fig. 3.5), animals in group 4 spent the most time (47%) on the net, as
compared to age group 1 (31%), and especially age groups 2 (17%) and 3 (20%). Log use was
also double that in the oldest age group (30%), as compared to the other groups 1, 2, 3 of 9%,
12% and 9% respectively. Conversely, platform and vine use was greater in younger animals.
Groups 1-3 spent 25%, 23% and 13% respectively on platforms, but only 5% by group 4. Vine
use was 9%, 10%, and 15% in groups 1-3 respectively but group 4 only spent 4% of their time on
vines.
For the Island exhibit, structure use was more similar for groups 1 to 3, than group 4. Net use was
highest in group 4 at 57%, as compared to groups 1-3 at 37%, 34% and 36% respectively. Vine
use was lowest in age group 4 (6%), almost half of the other age groups (7-10%). Log and

52
platform use was distributed more evenly across the age groups and utilized less than in the
Boardwalk exhibit, at 6-13% for log and 10-18% for platform use.

53
Behaviours at each structure type
For the trees in the Boardwalk exhibit, the major activities were idling and moving in group 1 and
idling and looking in groups 2, 3 and 4 (Fig. 3.6). In the Island exhibit, idling, moving and
feeding were the major activities in age group 1. Idling, looking and feeding were the major
activities in age groups 2 and 3. Looking and moving were most prevalent in the oldest group 4
animals. Looking was higher in the Boardwalk exhibit, and took between 16-54%, but only
between 12-33% in the Island exhibit. Conversely, feeding was higher in the Island exhibit trees
with from 14-38% but only between 5-20% in the Boardwalk exhibit.
On vines, movement was the major activity across all age groups and exhibits, taking up to 30-
45% in the Boardwalk exhibit and 67-88% in the Island exhibit. More play, human interaction,
and looking were also present on the Boardwalk exhibit vines.
On nets, major activities tended to be idling and feeding in both exhibits. There was slightly more
looking in the Boardwalk exhibit than Island exhibit, double the amount of movement in the
Island exhibit than Boardwalk exhibit, and more play in groups 1 and 2 in the Boardwalk exhibit.
Regurgitation, play and idling were the major activities on platforms. There was human
interaction on the platforms in the Boardwalk exhibit, but none at the Island exhibit platforms.
At the logs, there was an even distribution of behaviours. Movement and feeding made up a
larger proportion of the activity budget in the Boardwalk exhibit than in the Island exhibit. Play
and idling were greater on the Island exhibit log.
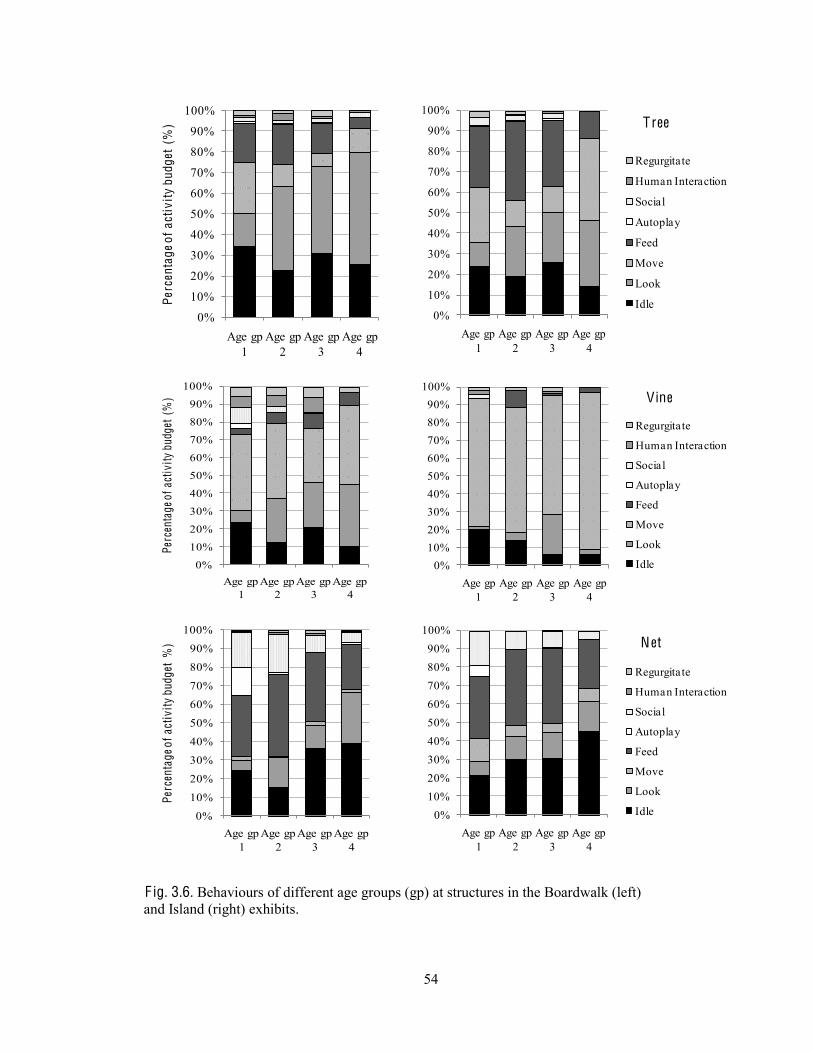
54
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Perc
enta
ge o
f act
ivity
bud
get
(%)
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Tree
Regurgitate
Human Interaction
Social
Autoplay
Feed
Move
Look
Idle
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Perc
enta
ge o
f act
ivity
bud
get (
%)
0%10%20%30%40%50%60%70%80%90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Vine
Regurgitate
Human Interaction
Social
Autoplay
Feed
Move
Look
Idle
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Perc
enta
ge o
f act
ivity
bud
get
%)
0%10%20%30%40%50%60%70%80%90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
N et
Regurgitate
Human Interaction
Social
Autoplay
Feed
Move
Look
Idle
F ig. 3.6. Behaviours of different age groups (gp) at structures in the Boardwalk (left) and Island (right) exhibits.
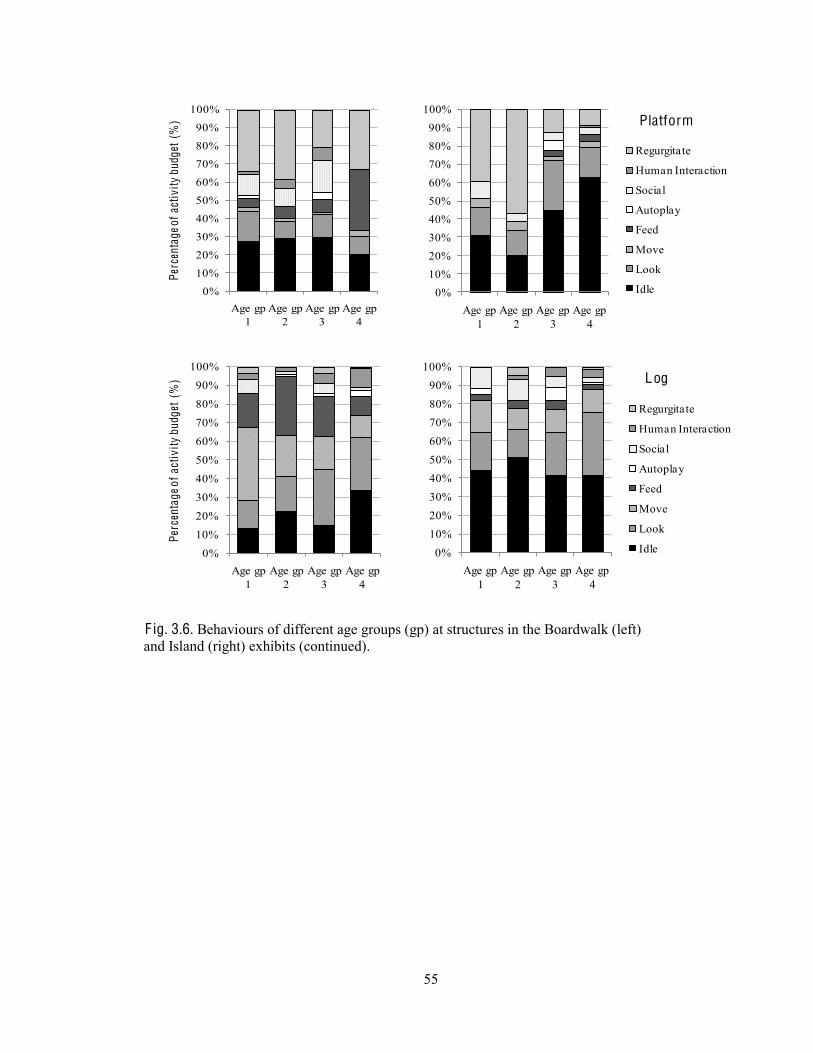
55
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Perc
enta
ge o
f act
ivity
bud
get (
%)
0%10%20%30%40%50%60%70%80%90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Platform
Regurgitate
Human Interaction
Social
Autoplay
Feed
Move
Look
Idle
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Perc
enta
ge o
f act
ivity
bud
get
(%)
0%10%20%30%40%50%60%70%80%90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Log
Regurgitate
Human Interaction
Social
Autoplay
Feed
Move
Look
Idle
F ig. 3.6. Behaviours of different age groups (gp) at structures in the Boardwalk (left) and Island (right) exhibits (continued).
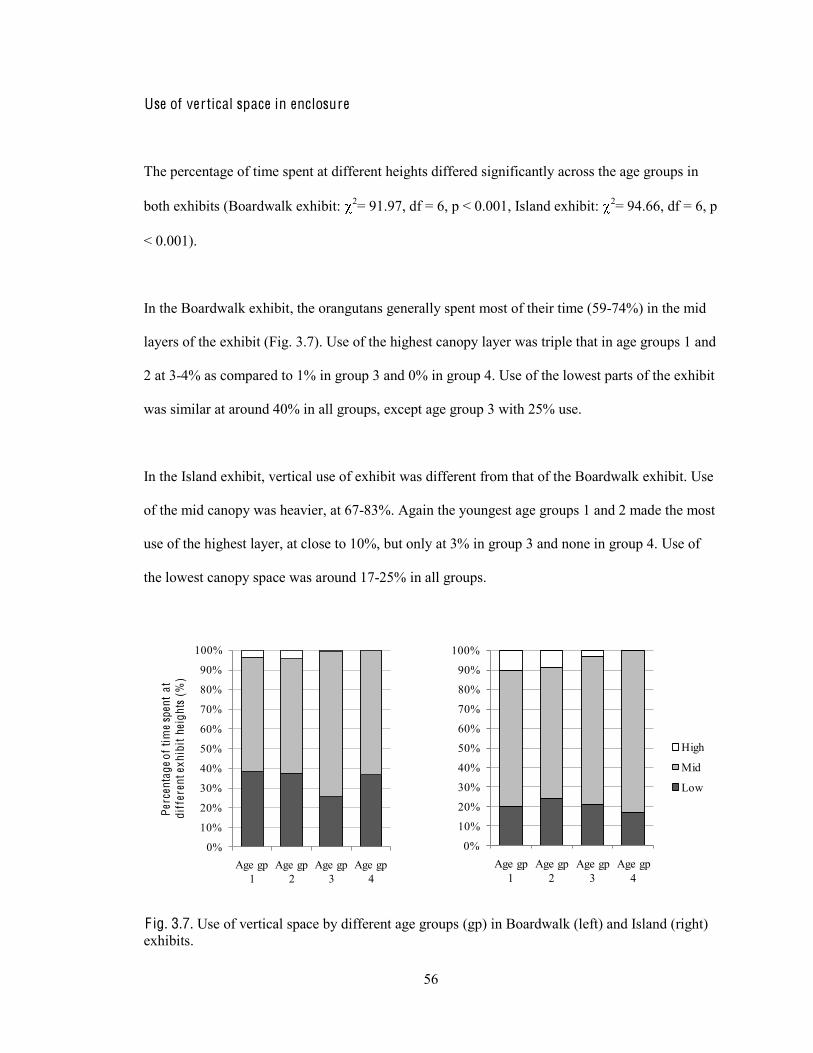
56
Use of vertical space in enclosure
The percentage of time spent at different heights differed significantly across the age groups in
both exhibits (Boardwalk exhibit: 2= 91.97, df = 6, p < 0.001, Island exhibit: 2= 94.66, df = 6, p
< 0.001).
In the Boardwalk exhibit, the orangutans generally spent most of their time (59-74%) in the mid
layers of the exhibit (Fig. 3.7). Use of the highest canopy layer was triple that in age groups 1 and
2 at 3-4% as compared to 1% in group 3 and 0% in group 4. Use of the lowest parts of the exhibit
was similar at around 40% in all groups, except age group 3 with 25% use.
In the Island exhibit, vertical use of exhibit was different from that of the Boardwalk exhibit. Use
of the mid canopy was heavier, at 67-83%. Again the youngest age groups 1 and 2 made the most
use of the highest layer, at close to 10%, but only at 3% in group 3 and none in group 4. Use of
the lowest canopy space was around 17-25% in all groups.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
Perc
enta
ge o
f tim
e sp
ent
atdi
ffer
ent e
xhib
it he
ight
s (%
)
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Age gp 1
Age gp 2
Age gp 3
Age gp 4
High
Mid
Low
F ig. 3.7. Use of vertical space by different age groups (gp) in Boardwalk (left) and Island (right) exhibits.

57
DISC USSI O N
This study investigated structure use and vertical space use in naturalistic orangutan exhibits to
improve understanding of how captive animals use their enclosures. Previous research suggested
that primates have preferences for certain exhibit structures, but studies have been limited to a
few species only. Here, the time spent by orangutans on the different structure types and their
activity budgets at each structure were investigated. Vertical space use was also elucidated. The
results showed that structure use varied with age and dominance status, as well as the
arrangement and availability of structures within the exhibit. Features surrounding the exhibit as
well as husbandry routine also affected structure use.
Use of enclosure structures across age groups and exhibits
From the results of the structure use study, we can see how two types of factors, i.e. biological
factors, such as age, behavioural profile, dominance hierarchy; and environmental features, i.e.
structure availability and location, can affect structure use (Leuck, 1977).
First, let us consider how biological factors affect structure use. In both exhibits, the largest
amount of time spent on any structure across the age groups was approximately 50% on nets by
age group 4. For net use in the Boardwalk exhibit, age groups 1 and 4 spent double the amount of
time on nets as compared to the other 2 age groups. Considering the results from Chapter 2 on
activity budgets, that age group 4 spent the most (a third of their) time idling and age group 1
spent the most time playing, it makes sense that these two groups had the greatest net use. This is
because idling and playing usually occurred on flat surfaces. Unpublished data by Marchal (2004)
showed that older animals (aged 15 - 40) spent the greatest proportion of their time on the floor, a
large, flat surface, as compared to the arboreal structures in their exhibit. As the nearest

58
equivalent of the floor would be the net in the Boardwalk exhibit, the results from the present
study seem to
been found to spend more time on moveable structures (ropes) than older animals (aged 10 - 32),
and were thought to prefer the moveable structures for play (Perkins and Bradfield, 1989). This is
similar to how the age group 1 orangutans in the Boardwalk exhibit spent more time on the
suspended net as compared to age groups 2 and 3, possibly because group 1 animals played more.
On the other hand, age group 4 dominated use of the log. More than 30% of the time was spent by
age group 4 on the log in the Boardwalk exhibit, whereas other groups only spent about half that
amount of time there. As the log was located in the centre of the exhibit and joined two trees on
which a net and the platform (core areas of activity) were found, this structure may have been a
major route or passageway for movement. It also appears the position of the log allowed the
animal which sat there a good view of the visitors coming up the boardwalk. This may suggest
why the older animals who looked more (see previous chapter), tended to spend a large
proportion of their time there. As a result, similar to what was found in captive bison (Robitaille
and Prescott, 1993) and gorillas (Fischer and Nadler, 1978; 1977; Hedeen, 1982), when older and
-
ranking individuals would actively avoid close interactions with them. Hence, lower rates of log
use are apparent in younger animals. Alternatively, similar to what was found in captive gorillas
(Ross and Lukas, 2006), older animals might also have used their location on the pathway as a
form of control over other individuals in their group.
Conversely, platform use in the Boardwalk exhibit was higher in the younger age groups 1 and 2,
possibly because these age groups played and regurgitated more, and playing and regurgitation
generally are facilitated by flat surfaces. Across both exhibits, vine use was double in all other
age groups, as compared to age group 4. This reflects the relative lack of movement in this age
group. Overall, the oldest orangutans spent more time on nets and logs, and the least on vines,

59
which suggests sedentary activities for these animals. These results mirror those found by studies
on wild (Morrogh-Bernard et al., 2009) and captive orangutans (Cassella, 2010; Marchal, 2004;
Perkins and Bradfield, 1989), which show that older animals generally tend to be less active.
younger animals used all structures more evenly. These results mirrors those from Perkins and
Bradfield (1989) who also found that older orangutans have clumped usage of exhibit and
younger animals were dispersed more evenly throughout the enclosure. Similar findings, although
not statistically significantly, are available for gorillas in a naturalistic exhibit (Stoinski et al.,
2001). From the results, it is clear that age, dominance, and subsequently behavioural profile of
different age groups can affect structure use.
Secondly, the results show how environmental factors, such as the location and availability of
structures within the exhibit, as well as the features in the vicinity of the exhibit can also affect
structure use. This can be seen in the differential use of nets, platforms, and logs across the
exhibits. Unlike the Boardwalk exhibit, net use in the Island exhibit was higher across all age
groups. This may be due to the presence of a huge cargo net which was present only in this
exhibit. Such a finding suggests that the availability of structures can affect choice of orangutan
location. Also, unlike in the Boardwalk exhibit where log and platform use was different across
age groups, we find that use of these structures was more evenly spread, as well as lower across
all groups in the Island exhibit. This was because in the Island exhibit, the log and platforms were
at one end of the exhibit, further away from the other core area of activity (i.e. the large cargo net)
(see Fig. 3.2). Conversely in the Boardwalk exhibit, the log and platform were in the centre.
Hence, orangutans spent more time at the log and platform when in the Boardwalk exhibit.
Finally, vine use was generally higher in Boardwalk exhibit than in Island exhibit. As mentioned,
the two exhibits differed in their proximity to surrounding features, such a restaurant and a third
orangutan exhibit in the vicinity. Orangutans while in the Boardwalk exhibit, had been frequently

60
observed to watch other orangutans which were housed in a third exhibit alongside the visitor
boardwalk. This too occurred in the Island exhibit, but to a lesser extent as the view of the third
orangutan exhibit from the Island exhibit was obscured by some trees (see Fig. 3.3). In addition,
have stimulated more looking behaviour. Such behaviour was often carried out from particular
vines in the exhibit.
It is apparent that the arrangement and availability of structures within an enclosure as well as the
presence of features in the surrounding of the enclosure can affect structure use. Manning (2002)
found that orangutans used rope and platform elements in their enclosure more when such
structures were available; similarly, higher rates of structure use were present when gorillas,
orangutans and chimpanzees were moved from barren cages to more complex, naturalistic
habitats (Maple and Stine, 1982; Pfeiffer and Koebner, 1978). The arrangement of features
within an exhibit, although not well studied, has also been found to affect how heavily a feature is
used. For example, caimans, for the most part, used pools in their enclosures; areas further from
pools were used significantly less (Verdade et al., 2006). Also, it has been found that primate
cages nearer the zoo exit and main paths had higher visitor attendance than cages further away or
off the main paths (Mitchell et al., 1990). The presence of more visitors was also associated with
increased aggression from primates (Chamove et al., 1988; Wells, 2005) and increased use of
enclosure areas near visitors (Fa, 1989). From these, we can appreciate how structure
arrangement, availability, and features in the vicinity of enclosures can affect animal behaviour
and subsequently structure use.
Because there is currently very little information available on the use of enclosure space across
many animal species (Stoinski et al., 2001), this knowledge on how different factors affect
structure use will serve as useful baseline data. Such information will come in useful during the

61
regular renovations to Singapore Zoo orangutan exhibits, or for the design of future orangutan
exhibits. Specifically, this knowledge can be used to improve animal welfare by ensuring lowest
possible levels of social conflict and sufficient structure availability for all age groups. Also, it
can be used to optimize enclosure use (greater use of entire exhibit), encourage desirable or
discourage undesirable behaviours in animals, or ensure that animals which are more sensitive to
noise or visitors are placed in appropriate locations. Some recommendations are provided at the
end of this chapter.
Behaviours at each structure type
In trees, major activities tended to be idling and looking in older animals, and idling and moving
in younger animals. Hebert and Bard (2000) also found that orangutans in their study (of ages
similar to age group 3 in this study) were solitary inactive (idle and looking) in the upper
canopy. This may be because trees are tall, provide solid surfaces for sitting, and serve as a good
vantage point to observe the surroundings. Therefore, looking and idling were the major activities
there. On the other hand, vines tend to connect structures; hence movement was predominant at
vines. Hebert and Bard (2000) also showed that orangutans in their study were solitary active
(moved and played) for 50% of the time on vines. In the Boardwalk exhibit, some vines were
directly alongside the third orangutan exhibit, with no visual obstructions in between, and some
vines were near the visitor walkway, whereas vines in the Island exhibit were generally higher off
the ground. This might explain why there was more looking and human interaction on Boardwalk
than Island exhibit vines. Feeding was higher at trees in the Island exhibit as fresh leaves
(browse) was always provided at a particular tree, whereas it was not so in the Boardwalk exhibit.
Higher usage of exhibit areas linked to husbandry and feeding has also been found in gorillas
(Stoinski et al., 2001) and tapirs (Mahler, 1984). It was observed that the use of particular exhibit
areas was much heavier when feeding by keepers was carried out there. Such information may be

62
useful to know if zoo managements wish to optimize use of exhibit space in captive animals. It is
also interesting to note that orangutans sometimes engaged in species-specific behaviours such as
tree-sway (Thorpe et al., 2007). Tree-
bend the topmost branches of one tree towards the next tree, to enable it to reach out, grab and get
onto the neighbouring tree. Such behaviours could only occur on flexible materials such as the
topmost branches of live trees, which is a positive consequence of having live trees as part of the
exhibit.
The spread of activities in nets was very different from that in trees. Playing, feeding and idling
were the major activities for younger age groups, whereas the older age groups tended to feed,
idle and look more. On the other hand, the platform seemed to facilitate regurgitation and idling
in all age groups. While both surfaces were flat, the nature of the surfaces encouraged different
activities. The orangutans were usually observed to spread browse out on the net and engage in
feeding, whereas the solid surface of the platform facilitated regurgitation and reingestion of
regurgitated material. Both platform and nets encouraged play in animals, as large, flat surfaces
facilitated wrestling and tumbling movements which were characteristic of orangutan play
(Maple, 1980). Again human interaction was more at the Boardwalk exhibit because of the closer
proximities of visitors to the orangutans while on the elevated boardwalk.
Even though there was a more even distribution of behaviours at logs, the log at the Boardwalk
exhibit seemed to be dominated by age group 4. More than 50% of the activities by group 4 at the
log were inactive and longer lasting behaviours such as looking and idling. As discussed
previously, the animals in age group 4 were often observed to sit for long periods at the centre of
the log. Because older animals were dominant in the group, younger animals actively avoided
using the log while older animals were there. Hence, activities in the other age groups at this
structure tended to be more transient, such as movement, feeding, etc. On the other hand, because

63
the log in the Island exhibit was not a route between two trees, nor at the centre of the exhibit, use
of the log by all age groups was more even. Idling was the dominant activity here, possibly
because the orangutans would wait at the log in the Island exhibit before feeding times, because
the log was the point of exit from the free-ranging exhibit.
Generally, flat surfaces are used for feeding, playing, idling and regurgitation, vines are used for
movement and looking occurs mainly from trees and vines. It is apparent that behaviour at each
type of structure, other than being dictated by properties of the structure, can also be influenced
by husbandry routine, social factors, location of structures within the exhibit as well as features in
the environment surrounding the exhibit. For example, feeding was more prevalent at trees in the
Island exhibit but the same behaviour occurred more frequently at logs in the Boardwalk exhibit.
This was because browse (fresh leaves) was always provided at a particular tree and at the log in
the respective exhibits. More movement occurred at the log in the Boardwalk exhibit but not the
Island exhibit because the log was a major pathway in the former but not the latter. Also, more
looking took place on vines at the Boardwalk exhibit, but not the Island exhibit because of more
and closer features in the surrounding of the Boardwalk exhibit. Once such knowledge on
behaviours at different structure types is available, this can be used to optimize animal welfare
and improve on future exhibit designs.
In conclusion, a good mix of structure types, at different heights and distances from visitors in an
exhibit, may encourage behavioural diversity in captive orangutans. Some recommendations for
improving orangutan welfare are provided at the end of the chapter.

64
Use of vertical space in enclosure
The results showed that the orangutans favoured the middle layers of the exhibit across both
enclosures. This may have been because the branches in the middle of the canopy were broader
and easier to move on, as compared to thinner branches found at the top of the trees. The results
of this study are similar to another which found that captive marmosets preferred relatively thick
to thin perches (Dettling, 1997). Also, many structures such as nets in both exhibits, and the
platforms in the Island exhibit, were located in the middle layer of the canopy.
The top parts of the exhibit were used mostly by the younger animals (observed to forage there
for seed pods). Although animals from age group 3 also used the topmost layer of the exhibit, it
may have been more difficult for them to move about on the thinner branches, hence, we see less
activity from group 3 animals. The lower usage of the top layer in the Boardwalk than Island
exhibit was because there were more low-current electrical wires to restrict access to the tops of
trees in the Boardwalk exhibit. These wires served to prevent orangutans from escaping the
Boardwalk exhibit as trees there were closer to surrounding trees than those in the Island exhibit.
Hence, access to treetops was limited in the former. Additionally, on occasions that the
orangutans managed to slip past the electrical wires, the presence of more fruiting trees in Island
exhibit than Boardwalk exhibit also encouraged more foraging activity at the top layer.
There was heavier usage of low areas in Boardwalk exhibit as compared to Island exhibit. This
may have been because the log and platform, which were heavily utilized (about 30% of structure
use in total) were both at the low areas in the Boardwalk exhibit, but only the log was at the low
part of the Island exhibit. Similar to the only other study on use of vertical exhibit space (Hebert
and Bard, 2000), the orangutans in that study also used the vertical sections in their exhibit
differently. The exhibit in Hebert and Bard (2000) was divided into four layers. Exhibit skylights

65
were the uppermost layer, followed by the upper canopy and lower canopy (of the artificial trees
in the exhibit), and the lowermost layer being a flooded floor which simulated a constantly
flooded peat forest. However, because the vertical sections are defined differently in this study
and the other study, direct comparisons cannot be made.
The animals in Hebert and Bard (2000) spent most of their time (35-40%) in the upper canopy of
their exhibit, which was about 8.5m off the ground. Similarly, the orangutans in this study spent
the most time (60-80%) in the middle layers of the canopy which was 10 to 15m off the ground.
As suggested by Hebert and Bard (2000), this height approximates the height at which wild
orangutans build their day and night nests, i.e. 10 to 20m off the ground (Wich et al., 2009),
hence the vertical level frequented by the captive orangutans can be called day nest sites.
Orangutans in Hebert and Bard (2000) were said to prefer the upper canopy because it afforded
more privacy and greater distance from visitors, as well as being close to where keepers would
drop food into the exhibit. Conversely, orangutans in the Singapore Zoo may have preferred the
middle layer because majority of the structures, such as nets, vines and a platform was found
there.
Use of the lower canopy was about 25-30% in Hebert and Bard (2000), and similar, at about 17-
38% in this study. Across both studies, the lower canopy, although at different heights, provided
places to sit and rest, like a metal ledge leading to the holding area in Hebert and Bard (2000) and
logs and platforms in this study.
The topmost layer in both studies, i.e. the skylights in Hebert and Bard (2000) and the top layer in
Singapore Zoo free-ranging exhibits, had very different purposes. The topmost branches in the
Island and Boardwalk exhibits were thin and only used for foraging purpose by younger

66
could be out of public view, be closest to the keepers who were dropping food into the exhibit, as
well as be close to the source of natural sunlight in the exhibit. As a result, we see much less use
of this topmost layer in the free-ranging exhibits (0-10%) as compared to the study by Hebert and
Bard (2000) (16-24%).
Although animals in group 4 of the Singapore Zoo group did not go to the top layer during the
data collection period, one of the adult females, Anita, was once observed to chase a younger
animal to the top layer in an agonistic interaction. This suggests that older animals are still
capable of using thin branches, even in captivity, which is encouraging in terms of species-
specific behaviours. Zoo management can consider encouraging species-specific behaviours, such
- -sway is a method where orangutans rock flexible tree
trunks from side to side with increasing magnitude until they can cross gaps in the canopy
(Thorpe et al., 2007). Only with access to the terminal branches of trees, which are thin and
flexible enough, can this behaviour be performed. Such behaviours require planning and
animals. One method to encourage tree crossings would be to include trees with luxuriant foliage
in the exhibit, or to provide thin, flexible poles which simulate tree-sway in the canopy. Such
sway poles have been implemented successfully at other zoos to simulate arboreal tree-crossing
behaviour in orangutans, and also to foster appreciation for orangutan behaviour in zoo visitors
(Oklahoma City Zoological Park: Grisham et al., 2000; Melbourne Zoo: website, Zoolex).
Overall, all layers of the canopy were utilized in this group of captive orangutans; however, there
was a preference for the middle layer of the exhibit.

67
Limitations of study
The investigation of enclosure use in this study focused on types of structures used, behaviours at
each structure, and vertical space use. It may also have been possible to look at whether
orangutans had differential (horizontal) use of areas in their exhibit. A preliminary study of space
use in this group of orangutans (unpublished data from this study) showed that orangutans spent
more than 70% of their time in half of the exhibit space. This may have been due to the nature of
the exhibit structures or proximity to surrounding features in different parts of the exhibit. A
more detailed analysis may be useful in elucidating how captive orangutans use a treetop,
naturalistic enclosure.
Vines in the exhibit were present in two forms; vines joining neighbouring trees, or vines strung
within the branches of the same tree. There was no distinction made between the location of
orangutans when they were on branches of a tree, or vines in a tree. Both were
for orangutan location. A more detailed categorization may have been useful.

68
R E C O M M E ND A T I O NS
Animals in age groups 2 and 3 may not be able to access nets as frequently as desired,
due to dominance effects from age group 4 animals. Hence it may be helpful to include
more nets in the Boardwalk exhibit. The presence of more nets may also stimulate play
and social behaviour.
Ensure that there are adequate pieces of each structure to avoid social conflict.
The presence of easily accessible platforms in the Boardwalk exhibit seems to encourage
regurgitation (an undesirable behaviour) in younger animals. It may be good to place
platforms at less accessible parts of the exhibit, or reduce the number of platforms to
reduce regurgitation. Although both play and regurgitation both take place at platforms,
and the platform was more out of the way in the Island exhibit, play was not significantly
lower in the Island exhibit, but regurgitation was. This may have been because of the
large net present in the Island exhibit. Hence, making platforms less accessible may
reduce regurgitation but not play, as long as sufficient net space is provided.
Ensure that there are always alternative routes, and no dead ends in the exhibit pathways
to prevent cornering of less dominant animals by dominant animals. This has already
been done for the free-ranging exhibits (Nantha, 2009, pers. comm.7). Domination of the
log (a major pathway) by older animals may cause frustration in lower-ranking animals if
there is no alternative through route.
7 Gabriel Nantha, Orangutan keeper, Singapore Zoo

69
Orangutans in this study do not seem to be adversely affected by proximity to human
establishments (see next chapter), hence close proximity to restaurants and roads may
actually be a source of enrichment. Also, some individuals were observed to actively
watch visitors at the restaurant (pers. obs.; Nantha, 2009, pers. comm.8). An anecdotal
piece of information provided by one of the keepers: Once, when an adolescent orangutan
(Budi) escaped from the exhibit, it was found holding a pair of utensils and sitting at one
of the outdoor tables in the restaurant, which suggests that it had actively observed
visitors at the restaurant beforehand. Hence, the presence of visitors may possibly be a
form of enrichment for this group of orangutans.
Exhibits can be designed with surrounding features in mind, to see whether they can have
potential as enrichment or may disturb the animals.
Enable and provide feeding at varying locations in each exhibit as a form of enrichment.
Provide opportunities for species-specific behaviours, such as tree-sway, by allowing
access to mature trees with thin, terminal branches. Alternatively, thin, flexible poles can
be provided to simulate tree-sway in the canopy.
8 Gabriel Nantha, Orangutan keeper, Singapore Zoo

70
C O N C L USI O NS
Structure use can be affected by the following biological and environmental factors:
Age and dominance status of the orangutans
The properties of the structure.
The availability and location of a structure within the exhibit.
The features in the surroundings of the exhibit.
Husbandry routines.
Such information can be used in future exhibit design to maximize animal welfare by reducing
social conflict and encouraging behavioural diversity.

71
C H APT E R 4
V isitor effects on zoo orangutans in two novel, naturalistic enclosures
Abstract. Visitors are known to affect zoo animals, and such effects may be stressful, neutral, or
enriching. The majority of research has focused on visitor number or visitor presence-absence,
yet few studies have examined effects of other variables such as sound volume, visitor activity,
and whether visitors interact with animals. In this study, the effects of visitor number, activity and
proximity to animals were investigated on a group of captive orangutans in two t -
relationships between visitor and orangutan behaviours. Results from these analyses revealed a
significant overall effect of the three visitor variables on orangutan behaviour. Interestingly,
visitor number had little effect on the orangutans, except at one of the exhibits where the
likelihood of food soliciting and looking at visitors increased when the number of people was
more than 40. Visitor activity was generally not associated with any obvious signs of stress in the
orangutans; visitors with food could even be a form of enrichment. However, visitors at close
proximity decreased play behaviour and increased the chances of animals looking at the visitors.
Enclosure design and habituation could have alleviated visitor effects for these two groups of
orangutans. This study shows how investigation of a wider range of visitor variables may allow
for more meaningful conclusions about the visitor effect, and that other factors such as enclosure
design and habituation to visitors may also influence captive animal welfare.
K eywords: visitors, captive, welfare, orangutan, behaviour
A modified version of this chapter was submitted to the journal Applied Animal Behaviour Science. Yuanting Choo, Peter Alan Todd, Daiqin Li. (2011) Visitor effects on zoo orangutans in two novel, naturalistic enclosures. Applied Animal Behaviour Science 133, 78 86.

72
IN T R O DU C T I O N
In the context of captive animal research, the ter
possible influences of visitors on the welfare of zoo animals. Although visitor effect studies have
been conducted since the 1970s (Oswald and Kuyk, 1977; Thompson, 1976), there are still
certain aspects of visitor effects which are relatively unstudied (Davey, 2007; Hosey, 2000).
Here, I will discuss what the visitor effect is, factors which may contribute to it, the consequences
of visitor effects on zoo animals and areas where more research is required.
it is necessary to consider how visitors may impact zoo animals. A visitor crowd can have a
multitude of characteristics. It can be large or small, noisy or quiet. It can comprise of adults
and/or children, who are male or female. They may be near or far away, and may or may not
interact with animals. It has been found that all of these characteristics can affect the behaviour of
zoo animals. Studies have shown that when there are more visitors, animals are found to be more
aggressive (Chamove et al., 1988; Sellinger and Ha, 2005; Simpson, 2004; Wells, 2005), play less
(Glatston et al., 1984; Jones, 2003), and increase their use of enclosure areas near the visitors (Fa,
1989). When visitors are noisier, animals direct more attention towards them (e.g., orangutans:
Birke, 2002). When visitors attempt to interact with the animals, some species of primates display
more visitor-directed behaviours, which can be aggressive, submissive (Chamove et al., 1988;
Hosey and Druck, 1987), or food soliciting behaviours (Cook and Hosey, 1995; Mitchell et al.,
1992a). If visitors appear smaller (by crouching), they appear to create less stress than standing
visitors for some species of primates (Chamove et al., 1988). Interestingly, even the gender of
visitors may play a role in the response of animals. It has found that male mangabeys are more
likely to show aggression towards men, and female mangabeys towards women (Mitchell et al.,

73
1992b). From the existing research, it is apparent that many different visitor characteristics can
affect zoo animal behaviour.
? The effect of visitors on
animals has been perceived in one of three ways negative, neutral, or positive (Hosey, 2000). A
negative visitor effect is usually defined by an increase in aggressive and stereotypic behaviours,
and a decrease in affiliative behaviours. A neutral visitor effect indicates that there is no
difference in the behaviour of the animals between varying visitor conditions. A positive effect
providing a source of variability (Davey, 2007). From previous studies, visitors may have one or
more of these effects negative, neutral or enriching.
As reviewed by Davey (2007), gaps in visitor effect studies can be classified into (i) a
disproportionate amount of research across different visitor variables and animal groupings, (ii)
vague and insufficient descriptions of visitor variables, (iii) limitation of conclusions due to
statistical methods used, or (iv) independence from other areas of visitor research. The present
study on orangutans in Singapore Zoo will attempt to address these gaps.
Of the existing visitor effect studies, a wide range of variables have been investigated. These
include variables from visitor presence or number (Anderson et al., 2002; Davis et al., 2005;
Glatston et al., 1984; Mallapur et al., 2005; Mitchell et al., 1990; 1991; 1992a; Shen-Jin et al.,
2010; Thompson, 1989; Wells, 2005), to visitor sound (Birke, 2002; Cooke and Schillaci, 2007),
visitor activity (Hosey and Druck, 1987; Mitchell et al., 1992a), as well as the height (Chamove et
al., 1988), and gender (Mitchell et al. 1992b) of visitors. However, there is an imbalance of
research, with most studies focusing on visitor number or presence, and few on visitor sound,
activity, proximity or crowd composition (Davey, 2007). Therefore, besides the commonly

74
studied variable of visitor number/presence, my study will highlight the less studied effects of
visitor activity and proximity on zoo orangutans.
Visitor activity has been investigated only by Mitchell et al. (1992a) and Hosey and Druck
looking, holding food, etc), more meaningful conclusions may possibly be drawn about the
effects of visitor activity. Similarly, visitor proximity has not been well studied, possibly because
majority of zoo exhibits have limited the distance of visitors from animals using physical barriers
(Hosey et al., 2009) and there is little variation in minimum animal-visitor distance. However, in
an exhibit like the orangutan enclosures in this study, visitor proximity may be of greater concern.
This is because visitors are not separated from the animals by any physical barriers. Instead, the
animals are restricted to a row of trees above the visitors, and people may approach as close as
three metres from the animals. Hence, my study will break down visitor activity into different
sub-categories and also address visitor proximity.
Visitor effect studies usually focus on one or two variables at a time and data is often analyzed
using univariate analyses. This could be due to the practical limitations of collecting several
visitor variables at once or the difficulties of using appropriate multivariate analyses. Other
reasons may be the small sample sizes or lack of independence between subjects, which are
characteristic of zoo studies (Kuhar, 2006; Plowman, 2008). By collecting data on only one or
two aspects, this leaves out the possible effects of other visitor variables. Here, a multivariate
analysis will be used here to simultaneously investigate three visitor variables.

75
Visitor effects have traditionally been considered independently from other areas of visitor
research and animal welfare. As reviewed by Davey (2005) and Fernandez et al. (2009), factors
such visitor circulation and responses to exhibit design have rarely been considered. In light of
this, I will investigate visitor effects across two different exhibits, and explain how enclosure
design and captive breeding history could also have influenced visitor effects.
Finally, and of particular relevance to the present study, even though most visitor studies have
focused on primates, there exists only two published reports on orangutans (Birke, 2002;
Bloomfield et al., 2010).
I hypothesized that larger groups of visitors, visitors who were more active (e.g. taking
photographs), and visitors who were closer in proximity would be stressful to the orangutans.
This may lead to lowered incidences of feeding, playing and social behaviours, and increased
incidences of undesirable behaviour such as regurgitation. I also hypothesized that visitors with
food would result in more begging behaviour.

76
M A T E RI A LS A ND M E T H O DS
Enclosure design, subjects and data collection
- orangutans in 2006 (Raj, 2009, pers.
comm.9). These exhibits each comprises a row of tall trees connected by artificial vines,
hammocks and platforms, and are designed to encourage brachiation and other species-specific
behaviour. During the 2009/10 study
and visitors passed by regularly. Visitors on the main track were able to look up into the trees to
view the orangutans. The other free-
Island exhibit but visitors could choose to view the orangutans from the main track, or from an
elevated boardwalk. This boardwalk allowed visitors to come into much closer proximity with the
animals (as close as 3 m).
Two groups of orangutans were rotated daily between the two free-ranging exhibits. The groups
were made up of individuals of varying ages, and each group had one mother-infant pair (Table
4.1). As the infants were still dependent on their mothers, they were excluded from the
observations. The composition of the groups remained the same throughout the study period. This
display arrangement presented a unique opportunity: usually, studies that compare animal
behaviour across different enclosures use data from different animals; however, in Singapore
Zoo, because the same animals were regularly rotated between two exhibits, I had the rare chance
to study the same subjects in different enclosures, simulating a manipulative experiment.
9 Jackson Raj, Head orangutan keeper, Singapore Zoo

77
F ig. 4.1. Island exhibit (top) and Boardwalk exhibit (bottom) showing possible locations of visitor-orangutan interaction. Island exhibit: (a) large net across trees, (b) ground-to-foliage view of exhibit trees, (c) low vine across main track where orangutans often interacted with visitors. Boardwalk exhibit: (d) low vine across main track where visitor-orangutan interaction often occurred, (e) elevated visitor boardwalk, (f) orangutan platform next to visitor boardwalk, (g) low log next to visitor track where animals would interact with visitors.

78
Table 4.1 Individual details of study animals. All individuals were born in Singapore Zoo except Anita who was donated to the zoo at one year of age. Name Sex Age (yr) Species Group A
Ah Meng Jnr Female 1 Pongo pygmaeus Bento Male 3 Pongo pygmaeus Budi Male 7 P. pygmaeus/ P. abelii cross Gunta Male 7 Pongo pygmaeus Chomel Female 13 Pongo abelii Anita (with infant Ah Meng Jnr) Female 24 Pongo pygmaeus Group B Saloma Female 0.5 Pongo pygmaeus Merlin Male 5 Pongo pygmaeus Vira Male 9 Pongo pygmaeus Labu Male 11 Pongo pygmaeus Binte (with infant Saloma)
Female 25 Pongo pygmaeus
In order to capture visitor crowds of varying sizes, I collected data on both weekdays and
weekends from October 2009 to February 2010, between the hours of 09:30 h to 17:00 h.
Instantaneous scan sampling, with the aid of binoculars, was used for both the orangutans and
(1996) protocol, it was calculated that scans taken at 10 min intervals
were sufficiently far apart enough to avoid autocorrelation. A total of 192 hours of observations
(48 hours for each group-exhibit combination) were made. As regular feeding sessions were held
twice daily, at 11:30 h and 15:30 h for the Island exhibit and 14:15 h and 16:30 h for the
Boardwalk exhibit, data collection was paused 15 min before each session started and only
resumed 15 min after the feeding ended. The visitor viewing areas (designated as the areas
directly below the exhibit trees) were divided into arbitrary subsections for the purposes of
specifying visitor location. During each scan, I recorded the behaviour and location of individual
orangutans (see Table 4.2 for behaviour categories). For visitors, I recorded the number of
individuals in each subsection of the viewing area and the activity of which the majority (>50%)
of the visitors in each subsection were performing. The visitor data were then processed to obtain

79
values of total visitor number, visitor activity and visitor proximity per orangutan per scan,
according to the definitions in Table 4.2.
Table 4.2 Definitions for orangutan and visitor variables (also see Table 4.3) Orangutan Behaviour Definition
Idle Orangutan is motionless, or performing any of these behaviours: autogrooming, expelling bodily waste or looking around (excluding looking at visitors).
Look at visitor(s) Orangutan is looking at visitor(s), face and eyes are oriented towards visitor(s).
Move Orangutan is travelling from one location to another; can be brachiating, bi/quadrupedal walking, or any other form of locomotion.
Feed Orangutan is engaged in searching for, preparation of, or ingestion of food.
Play/Social Orangutan is engaged in object use, solitary play, social play, or social interaction.
Regurgitate Orangutan is engaged in retrograde movement of food from its oesophagus or stomach to its mouth, hand or floor, and subsequent reingestion of the food.
Beg Orangutan is soliciting food from visitors by stretching out hand towards visitor(s).
Location Each tree/vine/structure in the exhibit was labelled with a name to identify the
location of each orangutan Processed variables Definition
Visitor number Visitor numbers from all subsections were summed to obtain the total number of visitors in the viewing area.
Visitor activity Classified as walking, standing, looking, taking photographs, holding food or no visitors. Only visitors within direct sight of the orangutan were considered. For each scan, I recorded the visitor activity as the activity that the majority (>50%)
visitor behaviours such as looking and taking photographs were thought to draw more attention from the
behaviour was recorded as the activity for that scan sample. The only exception was that visitors with food took precedence over all other visitor activities, regardless of how many such visitors there were, because we noted that food
Visitor proximity Classified into <10 m or >10 m from orangutan. If there were visitors at varying distances from the orangutan, only the closest visitors were considered. Distances were estimated using a rangefinder. Categories were defined as such, because preliminary observations showed that all interactions which involved visitors throwing food occurred at distances of <10 m.

80
Data analysis
Multinomial logistic regression was used to investigate possible visitor effects on orangutan
behaviour. Logistic regression was the preferred option as it can analyse dependent variables with
categorical outcomes (Field, 2009, p265). The output from this regression is expressed as the
likelihood (also known as odds ratio, OR) that a particular outcome category (in relation to the
reference category) will occur when a particular independent variable is present. Here, the
analysis provided the OR for each orangutan behaviour when a particular visitor variable was
present. I performed two separate multinomial logistic regressions, one for each exhibit, using
data pooled from both orangutan groups, to assess the associations between the independent
(visitor) and dependent (orangutan) variables. To avoid cells of zero count in the multivariate
analysis, all data points with no visitors were excluded from the analyses. The analysis was run
using PASW Statistics 18 (SPSS Inc. USA) and statistical significance was set at P < 0.05.
The reference categories for all variables are presented in Table 4.3. All levels of total visitor
number were compared agai -
each combination of predictor-dependent variable, multinomial logistic regression assumes that
all other variables are constant. Hence, an OR of more than one meant that the orangutan
behaviour was more likely to occur when a particular visitor variable was present, and an OR of
less than one meant that the orangutan behaviour was less likely to occur when that visitor
variable was present.

81
Table 4.3 Definitions and levels of variables for multinomial logistic regression Variable Levels Reference value Visitor number 11-20 visitors 1-10 visitors 21-30 visitors 31-40 visitors >40 visitors Visitor activity Standing Walking Looking Taking photographs Holding food Visitor proximity <10 m from orangutan >10m from orangutan Orangutan behaviour Feed Idle Play/Social Move Regurgitate Look Visitor Beg

82
R ESU L TS
Multinomial logistic regressions revealed significant overall effects of the three visitor variables
on orangutan behaviour for both the Boardwalk exhibit ( 2 = 318.902, P < 0.001) and the Island
exhibit ( 2 = 154.435, P < 0.001). For the Boardwalk exhibit, visitor number, activity and
proximity were all significant predictors of orangutan behaviour. For the Island exhibit, visitor
activity and proximity, but not visitor number, were significant predictors of orangutan behaviour
(Table 4.4).
The OR for each of the significant visitor variables in both enclosures are provided in Table 4.5.
For ease of comparison, Table 4.6 presents a summary of the effects of all visitor variables on the
orangutan behaviour decreased (OR < 1), and NS means the visitor variable did not significantly
affect the orangutan behaviour (P
Table 4.4. Likelihood ratio test results for both enclosures Enclosure Variable 2 test
Boardwalk exhibit Visitor number 2 = 37.856, P < 0.05 Visitor activity 2 = 140.222, P < 0.001 Visitor proximity 2 = 114.621, P < 0.001
Island exhibit Visitor number 2 = 34.840, P = 0.071 Visitor activity 2 = 63.059, P < 0.001 Visitor proximity 2 = 39.628, P < 0.001

83
Effect of visitors on orangutan behaviour in the Boardwalk exhibit
Only visitor numbers of > 40 increased the likelihood that orangutans would look at the visitors
(OR = 5.936) or beg (OR = 3.799). All other orangutan behaviours were not significantly affected
by visitor number (Table 4.5, 4.6).
Visitors who stood had no significant effect on the likelihood of any orangutan behaviours.
Visitors who looked at the orangutans significantly increased the chances that the orangutans
would feed (OR = 1.676), play/socialize (OR = 1.757), move (OR = 1.569) or beg (OR = 9.521),
but did not affect the other behaviours. Similarly, visitors who were taking photographs of the
orangutans significantly increased the chances that the animals would feed (OR = 1.370),
play/socialize (OR = 2.034) or move (OR = 1.494), but had no significant effect on the other
behaviours. Visitors with food had a significant effect only on the likelihood of begging (OR =
254.754) and looking at visitors (OR = 12.344). No other orangutan behaviours were affected by
visitors with food (Table 4.5, 4.6).
Visitors who were nearer (<10 m away) significantly increased the chances that the orangutans
would regurgitate (OR = 2.610), beg (OR = 3.014) or look at the visitors (OR = 2.924). However,
visitors who were <10 m away significantly decreased the chances that the orangutans would feed
(OR = 0.481) or play/socialize (OR = 0.669) (Table 4.5, 4.6).
Effect of visitors on orangutan behaviour in the Island exhibit
Visitor activity and proximity had significant effects on orangutan behavior in the Island exhibit,
but not visitor number (Table 4.4).

84
Visitors who were standing decreased the chances of orangutans feeding (OR = 0.599), but had
no other effects on orangutan behaviour. Visitors who were looking at the orangutans did not
significantly affect any of their behaviours; however, visitors who were photographing the
orangutans significantly decreased the chances of play/social behaviour (OR = 0.218), and
increased the chances of begging behaviour (OR = 31.094). Visitors with food significantly
increased the likelihood that the orangutans would be looking at the visitors (OR = 14.869) or
begging (OR = 256.210) (Table 4.5, 4.6).
Visitors who were <10 m away increased the chances that the animals would feed (OR = 1.544)
or beg (OR = 7.307). Such visitors also decreased the chances that the orangutans would
regurgitate (OR = 0.327) (Table 4.5, 4.6).
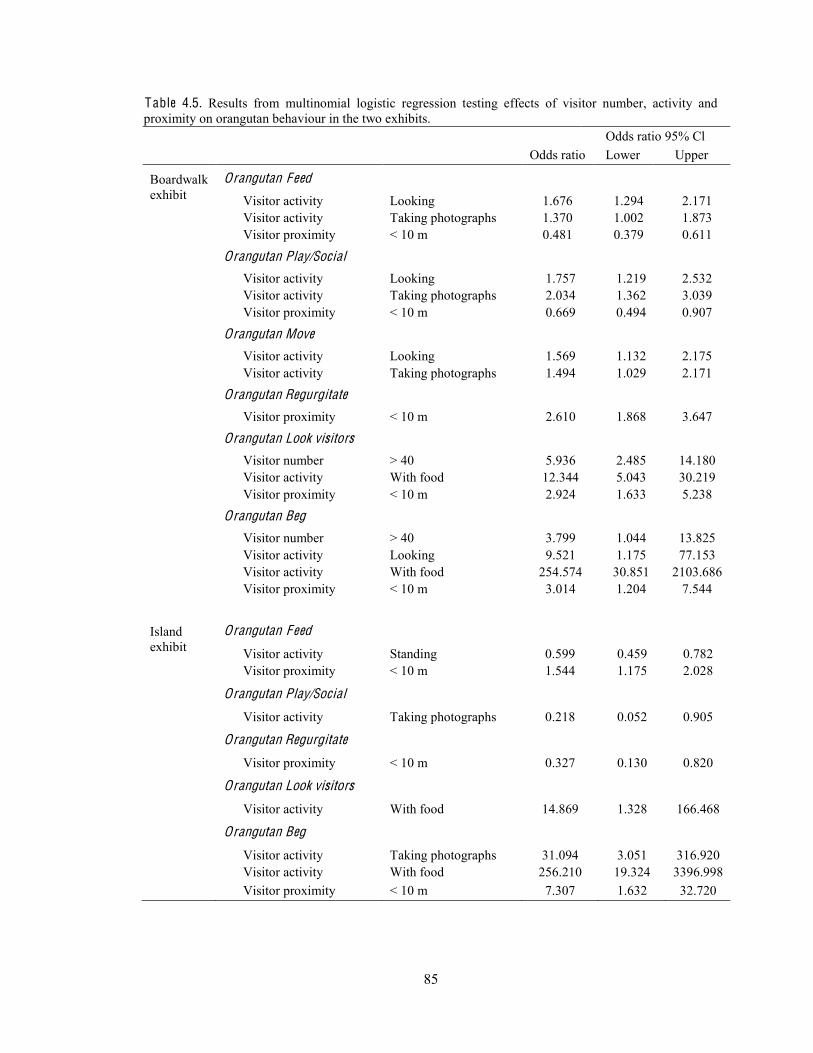
85
Table 4.5. Results from multinomial logistic regression testing effects of visitor number, activity and proximity on orangutan behaviour in the two exhibits. Odds ratio 95% Cl Odds ratio Lower Upper
Boardwalk exhibit
Orangutan F eed
Visitor activity Looking 1.676 1.294 2.171 Visitor activity Taking photographs 1.370 1.002 1.873 Visitor proximity < 10 m 0.481 0.379 0.611 Orangutan Play/Social Visitor activity Looking 1.757 1.219 2.532 Visitor activity Taking photographs 2.034 1.362 3.039 Visitor proximity < 10 m 0.669 0.494 0.907 Orangutan Move Visitor activity Looking 1.569 1.132 2.175 Visitor activity Taking photographs 1.494 1.029 2.171 Orangutan Regurgitate Visitor proximity < 10 m 2.610 1.868 3.647 Orangutan Look visitors Visitor number > 40 5.936 2.485 14.180 Visitor activity With food 12.344 5.043 30.219 Visitor proximity < 10 m 2.924 1.633 5.238 Orangutan Beg Visitor number > 40 3.799 1.044 13.825 Visitor activity Looking 9.521 1.175 77.153 Visitor activity With food 254.574 30.851 2103.686 Visitor proximity < 10 m 3.014 1.204 7.544
Island exhibit
Orangutan F eed
Visitor activity Standing 0.599 0.459 0.782 Visitor proximity < 10 m 1.544 1.175 2.028 Orangutan Play/Social
Visitor activity Taking photographs 0.218 0.052 0.905 Orangutan Regurgitate
Visitor proximity < 10 m 0.327 0.130 0.820 Orangutan Look visitors
Visitor activity With food 14.869 1.328 166.468 Orangutan Beg
Visitor activity Taking photographs 31.094 3.051 316.920 Visitor activity With food 256.210 19.324 3396.998 Visitor proximity < 10 m 7.307 1.632 32.720
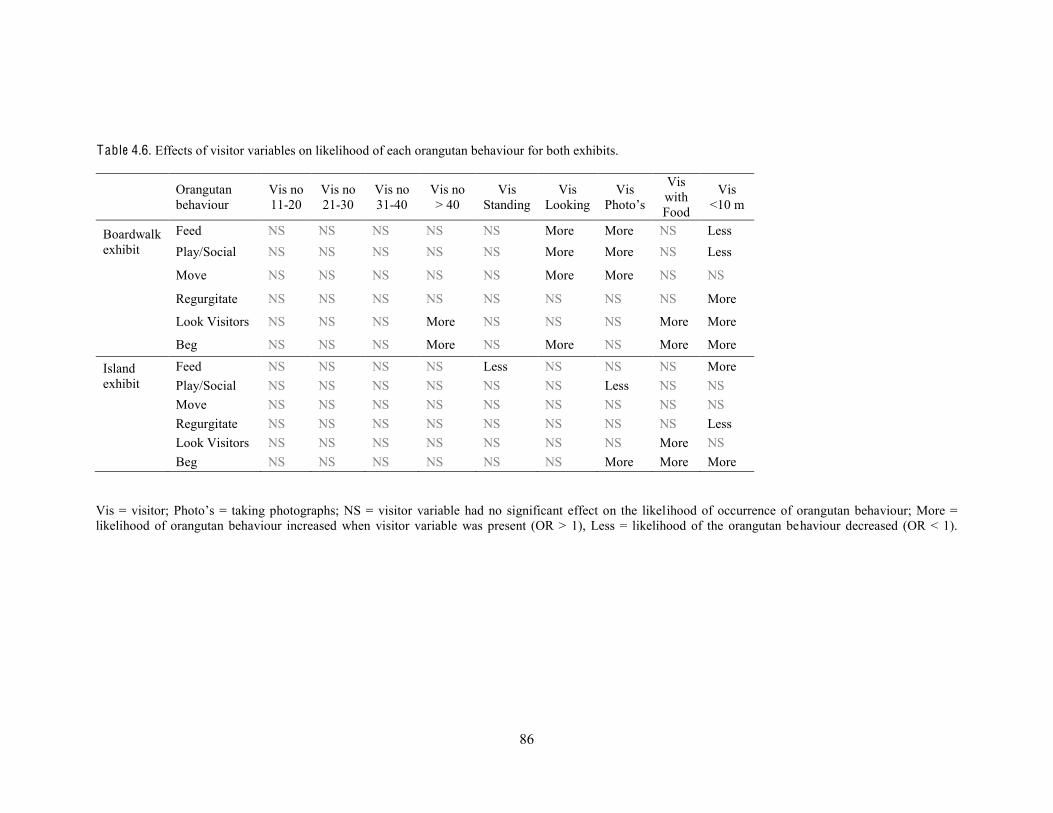
86
Table 4.6. Effects of visitor variables on likelihood of each orangutan behaviour for both exhibits.
Orangutan behaviour
Vis no 11-20
Vis no 21-30
Vis no 31-40
Vis no > 40
Vis Standing
Vis Looking
Vis
Vis with Food
Vis <10 m
Boardwalk exhibit
Feed NS NS NS NS NS More More NS Less Play/Social NS NS NS NS NS More More NS Less
Move NS NS NS NS NS More More NS NS Regurgitate NS NS NS NS NS NS NS NS More Look Visitors NS NS NS More NS NS NS More More Beg NS NS NS More NS More NS More More
Island exhibit
Feed NS NS NS NS Less NS NS NS More Play/Social NS NS NS NS NS NS Less NS NS
Move NS NS NS NS NS NS NS NS NS Regurgitate NS NS NS NS NS NS NS NS Less Look Visitors NS NS NS NS NS NS NS More NS Beg NS NS NS NS NS NS More More More
ihood of occurrence of orangutan behaviour; More = likelihood of orangutan behaviour increased when visitor variable was present (OR > 1), Less = likelihood of the orangutan behaviour decreased (OR < 1).

87
DISC USSI O N
This study incorporated several variables of visitor number, activity and proximity, in an attempt
to improve our understanding of how visitors affect orangutans. Previous research has suggested
that active visitors elicit more audience-directed behaviours from primates (Hosey and Druck,
1987; Mitchell et al., 1992a), but there has been little research on the effects of specific visitor
activities. Here, visitor activity was broken down into distinct categories. I also exploited the fact
that the two groups of orangutans were rotated daily between two exhibits. This regular rotation
formed a natural experiment and allowed comparison of visitor effects across the different
enclosures while controlling for between-group variation in behaviour. Overall, the results
showed that large crowds, visitors with food, visitors who were looking or taking photographs,
and visitors who were close by, all affected orangutan behaviour. On the whole however, the
hypothesized. The free-ranging exhibit design, habituation to humans, or both, may explain this.
Effects of visitor number
As compared to other studies of visitor effects on orangutans (Birke, 2002; Jones, 2003), my
research reveals little effect of visitor number on orangutan behaviour. The likelihood of begging
and looking at visitors increased only when there were crowds of more than 40 people. No other
behaviours were affected by visitor number, suggesting that human presence induces little or no
orangutans covered themselves with paper sacks more and foraged less when more (> 8) visitors
were present. Visitor-induced stress such as reduced social play and mobile feeding activities has
also been reported in orangutans when there were more than 100 visitors (Jones, 2003). However,
as the visitor numbers in this study rarely went above 80, whether higher visitor numbers (e.g., >

88
80) could induce stress on orangutans at Singapore Zoo is unknown. Nevertheless, because the
exhibits were positioned along tracks with heavy human traffic, it is possible that orangutans in
Singapore Zoo have been habituated to large visitor crowds. Also, being at a higher position
(usually in trees) above visitors may have provided a sense of security or dominance (Coe, 1985).
The fact that a large visitor crowd increased the chances of visitor-oriented behaviour only in the
Boardwalk exhibit, where an elevated boardwalk allowed visitors to get closer to the animals,
indicates that exhibit design may influence how visitor numbers affect orangutan behaviour.
Effects of visitor activity
Changes in orangutan behaviour were more strongly associated with active visitors (those looking
or taking photographs) than passive (standing) visitors. Similar results have been reported in other
studies (e.g. Chamove et al., 1988; Hosey and Druck, 1987; Mitchell et al., 1992a), in which
significantly more behaviours were directed towards active than passive audiences. However, in
); whereas in this
study, visitor activity was divided into five categories. It was found that the association between
active visitors and orangutan behaviour was more pronounced at the Boardwalk exhibit than the
Island exhibit, which again suggests that proximity of visitors to animals can affect the visitor-
animal interaction. The greater probability that the orangutans would be feeding,
playing/socializing, or moving when visitors looked or took photographs could be explained by
hypothesis, where active animals attract more attention from visitors
(Hosey, 2000)
visitors in the Boardwalk exhibit who looked at the orangutans, and visitors who photographed
orangutans in the Island exhibit, were linked to a higher chance of begging.

89
Overall, the increase in likelihood of feeding, play and social behaviours when visitors were
looking or taking photographs in the Boardwalk exhibit suggests that the visitors were not a
source of stress. At the Island exhibit, visitors standing were linked to lowered incidences of
feeding in orangutans while those taking photographs were linked to lowered incidences of
playing/socializing in the animals. The latter result was the opposite of what we found at the
Boardwalk exhibit but we suggest that, because play/social behaviour in the Boardwalk exhibit
occurred mostly in highly visible locations near the raised visitor walkway, these behaviours were
more likely to attract visitor attention and hence more photography. On the other hand,
playing/socializing at the Island exhibit occurred at less obvious places (i.e. cargo nets in the
upper branches of the trees), hence less photography. The association between visitors standing
and the lowered incidence of feeding was unexpected and cannot be easily explained by either the
visitor attraction or visitor effect hypothesis.
As hypothesized, food was a very strong stimulus of orangutan behaviour, with the likelihood of
begging and looking at visitors increasing significantly in both exhibits when food was present.
This result contributes to the consensus from other studies where apes were also motivated to
interact with visitors for food (Birke, 2002; Cook and Hosey, 1995; Jones, 2003; Wood, 1998).
The lives of zoo animals may often be routine; therefore, novel occurrences of visitors with food
may serve as a source of variability and enrichment (van Rooijen, 1991; Wiepkema and
Koolhaas, 1993). All orangutans would immediately and fixatedly look at visitors who were
holding food; they would also occasionally solicit food by hanging from a branch directly above
and extending their hand towards the visitor. In some cases, the orangutan would clap its hands
repeatedly, then stretch out a hand in request for food. (See video:
http://www.youtube.com/watch?v=L1bU9rHlGZ0 ) Because the orangutans have been trained to
perform certain behaviours for food rewards, repeated personal observations and communication
with the keepers confirmed that this clapping, then hand-out gesture was an active behaviour to

90
solicit food. Food solicitation has also been recorded with other primates (Cook and Hosey, 1995;
Jones, 2003; Wood, 1998). Barring that visitors may throw unsuitable foods such as sweets and
biscuits (pers. obs.), such incidences may otherwise serve as a source of enrichment for the
orangutans.
Interestingly, begging behaviour only occurred consistently with some of the orangutans, namely
Chomel (a 13 year old female) and Labu (a 11 year old male), whereas some individuals, such as
Anita (a 24 year old female) never displayed such behaviour . An obvious difference in individual
begging styles was also observed. Labu would only beg when food was visible; and he was ever
seen scanning passing visitors, and paying more attention to visitors who looked as though they
were holding food. Chomel would sometimes beg regardless of whether food was present, as long
as there were large visitor crowds. She was sometimes observed hang from a branch for up to half
an hour, waving her hands in a presumed attempt to get food. Her begging behaviour sometimes
- . obs.; pers. comm., orangutan keepers). The impression was
given that she did not really care if she got food, she just wanted to occupy herself. It has been
shown previously that personality may influence an o (Uher et al.,
2008) and Fraser (2009) highlighted this phenomenon for further study.
Effects of visitor proximity
I also examined the effects of visitor proximity on orangutans, a rarely studied variable. The
results showed interesting trends between exhibits, and a possibly stressful visitor effect at closer
proximities. The likelihood of both begging and looking at visitors were increased in the
Boardwalk exhibit when visitors were less than10 m away, but in the Island exhibit, only the
chances of begging were increased. This meant that orangutans looked more at visitors when they
were near in the Boardwalk exhibit, but not in the Island exhibit. This difference between

91
enclosures may have been due to the elevated boardwalk in the Boardwalk exhibit which allowed
visitor-animal interactions at eye level, and at much closer proximity (3-7 m) between orangutans
and visitors. This was in contrast to the Island exhibit, where the orangutans were always in trees
above the crowds, hence, at least 7 m above the visitors. Consequently, any effects of visitor
proximity may have been reduced in the Island exhibit. When visitors were nearer (less than 10 m
away), regurgitation increased in the Boardwalk exhibit but decreased in the Island exhibit. We
noted that regurgitation almost always took place at platforms; while the platform in the
Boardwalk exhibit was less than 10 m away from visitor viewing areas, the platform in the Island
exhibit was further than 10 m from all visitor viewing areas. Hence, the results for regurgitation
may have been influenced by the location of the platforms where regurgitation usually occurred.
The differences in feeding in relation to visitor proximity can also be explained by where the food
(fresh leaves) was usually provided (closer to visitors at the Island exhibit and further from them
in the Boardwalk exhibit). The association between visitor proximity and decreased
play/socializing in the orangutans was also limited to the Boardwalk exhibit. To date, there has
only been one other study on visitor proximity, Bloomfield et al. (2010), who did not find
evidence that orangutans avoid close visitor contact. In this study, however, visitors at <10 m
proximity decreased the chances of play/socializing, suggesting a stressful effect.
Other influences on the visitor-animal interactions
It has been shown how visitor number, activity, proximity and exhibit design influenced visitor-
orangutan interactions. The relatively low level of visitor effects on these two groups of
orangutans may be due to the large and naturalistic enclosure designs. There are many factors
influencing stress in captive animals, including: a lack of control over their environment,
restricted movement, and forced proximity to humans (Morgan and Tromborg, 2007). The large
and complex design of the free-ranging exhibits possibly reduced stress from visitors by allowing

92
for a retreat space, species-specific behaviour, and control over the degree of interaction with
visitors. Generally, the orangutans in this study seem to be habituated to humans, for instance,
active food solicitation by several individuals suggests a lack of fear towards visitors (Hosey et
al., 2009, p. 498). This might be due in part to daily photography sessions, where visitors sit
within one metre of the orangutans; or the regular handling by keepers, during which there is
keeper-animal contact. It has been observed that stimulation during early stages of life enables
some species to cope better with stress when they are older (Moodie and Chamove, 1990). The
orangutans at Singapore Zoo have been handled by keepers up to 12 years of age for males, and
for more than 20 years of age for females, which is not the standard practice at most zoos (Raj,
2008, pers. comm.10). As a result, these captive-born animals have had frequent opportunities to
interact with keepers and observe visitors at close proximity.
Limitations of study
Exclusion of another potential variable
Initially, visitor sound was designed as part of the study. However, preliminary trials showed that
visitor sound died off within 7 m of the source, probably due to the outdoor nature of the exhibit.
Considering that the orangutans could be at a range of distances (3 60 m) from the visitors, this
otherwise potential variable was excluded from the study.
10 Jackson Raj, Head orangutan keeper, Singapore Zoo

93
Definitions of visitor activity
For the definition of visitor activity, the activity that was being performed by most of the visitors
was recorded for each sample point. But for activities of looking and photo taking, they were not
always the majority activities. It was assumed that the effects of looking and photo taking were
dominant over that of walking or st
walking or standing. This assumption was necessary to break down visitor activity into more
detailed categories, however, the specific effects of this assumption are unknown. Further studies
can be carried out to investigate visitor activity in detail.
Limitations of behavioural observations
This study focused on measuring the association between visitor variables and orangutan
behaviour. There are limitations to using only correlational behaviour measurements for welfare
assessment in zoo animals (Davey, 2005; Millman, 2009). It may be possible to supplement
behavioural observations with the collection of physiological indicators, such as measuring
cortisol levels in saliva, urine (Smith, 2004), or any other non-invasive methods to confirm
direction of causality between visitor and orangutan variables. Alternatively, as Bloomfield et al.
(2010) has demonstrated, preference tests in which animals are allowed to choose their level of
interaction with visitors may shed more light into how visitors really affect zoo animals.
Lack of consideration of individual personalities
In many visitor effect studies, the data from several individuals is pooled (Carder and Semple,
2008; Chamove et al., 1988; Fa, 1992; Hosey and Druck, 1987, Mallapur et al., 2005; Mitchell et

94
al., 1992b; Shen-jin et al., 2010; Todd et al., 2007). However, it has been shown that individual
orangutans may differ in their responses to stimuli (Uher et al., 2008), and naturally, also to
visitors (Birke, 2002; pers. obs.; orangutan keepers, pers. comm.). It would be interesting to see if
the visitor effect varied between members of this orangutan group, as suggested earlier by the
difference in begging responses between individuals.
A more holistic approach
The design of most visitor effect studies, including this study, focuses only on short term
responses to visitors, and reduces visitor effects to a set of conveniently quantifiable variables
(Fraser, 2009). However, recent discussion in the field has refocused attention on how a more
holistic approach is necessary (Bloomsmith, 2009; Fraser, 2009; Melfi, 2009; Millman, 2009).
Fraser (2009) discussed how animal welfare often involves understanding the affective states of
animals (pain, fear, distress, etc). This understanding requires observation of complex behaviours,
and postulations about intangible entities, such as emotions (Duncan, 1970; Fraser, 1975). But
because philosophical influences in the history of scientific research maintained that science deals
only with tangible, observable phenomenon (Kolakowski, 1968), the development of science over
the mid 20th century has favoured the collection of quantitative over qualitative data (Fraser,
2009). As a result, controlled experiments which attempt to generalize across a species or type
have become commonplace (Fraser, 2009; Millman et al., 2009). However, to answer questions
about animal welfare and behaviour, it may be necessary to adopt a more holistic approach
(Fraser, 2009). As seen in the work of Jane Goodall (1971) and Barbara Smuts (1999), these
scientists collected both quantitative and qualitative data; they described complex behaviours in
addition to isolated phenomenon, and besides pooling data for groups, they also attempted to
consider individual differences (Fraser, 2009). Such adoption of a wide range of measures may
allow a better understanding of animal welfare.

95
In addition, a recent conference highlighted the need to use a wider range of responses, in
combination with long term, epidemiological studies to better understand the factors influencing
(Bloomsmith, 2009). However, this type of data collection is understandably more challenging
due to its inter-institutional nature; perhaps that explains why most visitor effect studies choose
instead to focus on subjects from one or few institutions.

96
R E C O M M E ND A T I O NS
It will be useful for zoo management to know that visitors who were very close to the
animals may have reduced play behaviour. This is especially so, if there are orangutans
with babies in the exhibit group. This is because young orangutans tend to play more than
adults (Nash, 1993; Zucker et al., 1986), and any reduction in play may be detrimental for
their development. Although no observations were made on the two infant orangutans
who accompanied the females, further research could be done to see if visitors have any
stressful effects on the development or behaviour of young orangutans. As shown by
Mather (1999), visitor presence decreased the amount of time orangutan infants spent
suckling from their mothers. The presence of such effects may then be taken into
consideration when deciding which exhibit to display mother-infant pairs in. This would
be important for the captive breeding program which is part of the conservation efforts at
Singapore Zoo.
It is interesting to note how close proximity at certain points in both exhibits allows
visitors to throw food to the orangutans. Regular food soliciting, although possibly
enriching, may or more not be desirable. This is especially so when we consider that
visitors may throw unsuitable foods to the animals.

97
C O N C L USI O NS
The results show relatively minor visitor effects on captive-bred orangutans in free-ranging
enclosures at Singapore Zoo. This may be due to enclosure design, or habituation to humans.
However, some visitor effects were observed, for instance large crowds, visitors with food,
visitors who were paying attention to the animals, or visitors who were close by all affected
orangutan behaviour to some degree. Of note, visitors who were close by were associated with
reduced play behaviour. This study highlights how enclosure design and captive history of study
subjects may also influence the visitor effect.

98
C H APT E R 5
G E N E R A L DISC USSI O N
The purpose of this study was to investigate the behaviour of captive orangutans in two
naturalistic exhibits. These results represent a substantial cache of information on the activity
budgets, structure use, and possible visitor influences on captive orangutans. This is the first
comprehensive study on orangutan behaviour in a naturalistic exhibit, and can serve as a
reference for zoo management, as well as lay the foundation for future studies.
In summary, the orangutans at Singapore Zoo showed age-specific activity budgets; differing
activity budgets across enclosures, and somewhat similar budgets to that of wild orangutans.
Activity budgets were influenced by husbandry routine, enclosure design, and features in the
vicinity of the exhibit. Structure use and behaviours at each structure varied according to
biological factors and environmental factors. Biological factors included orangutan age,
behavioural profile and dominance hierarchy. Environmental factors consisted of structure
properties, structure availability, location in exhibit, features in exhibit surroundings as well as
husbandry routines. The study on visitor effects showed that large crowds of more than 40,
visitors with food, visitors who were looking or taking photographs, and visitors who were close
by, all affected orangutan behaviour. Of note was the fact that visitor effects may have been
alleviated by the large and naturalistic exhibit design, or habituation to humans.

99
Common factors across the three chapters
Across all three studies, enclosure design and husbandry routines seem to have consistent
influence on orangutan behaviour. For enclosure design, the location, availability and
arrangement of structures within an exhibit affected the activity budgets of orangutans by
encouraging or discouraging particular behaviours. These factors also appeared to play a role in
bridging differences between captive and wild orangutan behaviour. The large enclosure design
also possibly alleviated stress from visitors by allowing for retreat space. Likewise, husbandry
routines seemed to improve the welfare of this orangutan group. The provision of regular food
throughout the day lowered inactivity in the animals. Regular keeper-animal contact, in
combination with daily visitor photography sessions, appeared to be a source of enrichment and
helped habituate the animals to visitors.
The importance of exhibit design and keeper interaction on the quality of captive animal welfare
has previously been highlighted by Shepherdson et al. (1998). Unlike their wild counterparts
which are free to roam, animals in captivity have to adapt to and make the best of enclosure
structures. This may result in abnormal (stereotypic) behaviour if there is insufficient stimulation.
One example is a solitary male gorilla which developed regurgitation and reingestion behaviours
when housed in an enclosure with only concrete walls and a wooden structure (Cipreste et al.,
2010). The abnormal behaviours stopped when its exhibit was renovated to become one of greater
complexity, with added vegetation, rockwork and water features. Similarly, many decisions made
by animals in the wild are taken over by keepers in captivity; for example choosing a mate, or
when and what to eat. Free- ver these
-
(Shepherdson, 2010). As a result, increasing feeding activity by varying food presentation or

100
types of food available has been found to promote species-specific behaviours, reduce stereotypic
behaviour and improve captive animal welfare (Young, 2003).
Regurgitation an artefact of captivity
Throughout this study, regurgitation and reingestion behaviour was referred to as an undesirable
behaviour. Regurgitation and reingestion is the voluntary movement of food from the oesophagus
or stomach back into the mouth or substrate. Such behaviour is considered abnormal in primates
because it is observed almost only in captivity. Regurgitation has not been reported in wild
orangutans (Knott, 2011, pers. comm.11, Wich, 2011, pers. comm. 12) or gorillas (Hill, 2009), and
has only been noted in two individual instances in wild primates (proboscis monkeys, Nasalis
larvatus: Matsuda et al., 2011; vervet monkeys, Cercopithecus aethiops: Struhsaker, 1977). On
the other hand, it is found widely in captive primates, including orangutans (Maple, 1980),
gorillas (Akers and Schildkraut, 1985; Gould and Bres, 1986; Lukas, 1999; Lukas et al., 1999),
chimpanzees (Baker and Easley, 1996; Struck et al., 2007), macaques (Bayne et al., 1991),
siamangs (Fox, 1971), as well as in humans, in particular with infants who have undergone
psychological problems such as stress, lack of stimulation or the loss of a parent (Blinder et al.,
1988; Mayes, 1988).
Because orangutans, similar to gorillas, are hindgut fermenters with simple stomachs (Caton et
al., 1999; Knott, 1998), they are not physiologically designed to ruminate food during digestion
(Elftman and Atkinson, 1950). A recent study on gorillas indicates that regurgitated food contains
stomach acid (Hill, 2009). Given that a behaviour similar to regurgitation, the human rumination
syndrome, has been observed in humans, and associated consequences include dental erosion,
11 Dr Cheryl Knott, Field Researcher, Associate Professor, Department of Anthropology, Boston University 12 Dr Serge Wich, Field Researcher, Post-doc, Anthropological Institute & Museum, Universität Zürich

101
oesophagal motor disorders and ulcers (Thame et al., 2000 ), this indicates potential health
consequences for animals that regularly regurgitate. As such, regurgitation and reingestion is
thought to be an undesirable behaviour found only in captive primates (Lukas, 1999).
There are several factors which may contribute to regurgitation in captivity. These include
psychological factors (stress or boredom), housing factors (space restriction or the lack of
environmental/social stimulation), as well as nutritional factors (species-inappropriate diets or
lack of foraging opportunities) (Akers and Schildkraut, 1985; Lukas, 1999; Lukas et al., 1999).
For example, studies on captive gorillas and chimpanzees have shown that increasing the amount
of browse (leafy vegetation) provided throughout the day can reduce regurgitation and reingestion
behaviour (Gould and Bres, 1986; Lukas, 1999; Struck et al, 2007). This may be because in the
wild, gorillas and chimpanzees spend between 47-60% of their time feeding (Goodall, 1986;
Watts, 1988). However in captivity, feeding behaviour often is limited to two or three meals per
day. Hence, regurgitation and reingestion could possibly be an adaptive mechanism to extend
feeding time and approximate more closely the feeding behaviours of wild primates (Lukas,
1999; Lukas et al., 1999). Interestingly, a study on regurgitation in captive gorillas showed that
removing milk from captive diets also resulted in a 28% decrease in regurgitation/reingestion,
which may indicate that appropriate choice of foods by animal management can also reduce this
undesirable behaviour (Lukas et al., 1999). Therefore, regurgitation may be a coping response to
captivity (Lukas, 1999).
In view of the findings from my study, the following are some recommendations for captive
orangutan welfare.

102
Recommendations on enclosure design
Structures within the exhibit
It is interesting to note how the availability, and especially location, of structures in an exhibit
may affect structure use and behaviour. Only one study to date has discussed structure location
for a caiman exhibit (Verdade et al., 2006), and little is known about the effects of this factor on
other species. The results of my study suggest that it may be possible to encourage or discourage
particular behaviours simply by rearranging the structures in an exhibit. For example,
regurgitation was less frequent in the Island exhibit than Boardwalk exhibit. This suggests that
orangutans may have regurgitated (an undesirable behaviour) less in the Island exhibit because
platforms, structures where regurgitation normally took place, were at the far end of the
enclosure. Similarly, movement may be increased by placing core areas of activity far apart in an
enclosure, as in the Island exhibit. As there have been almost no studies in this area; such
knowledge should be of use for future enclosure design.
To prevent social conflict for orangutans housed in groups, it is advisable to provide sufficient
pieces of each structure. Alternative routes between structures in each exhibit is also necessary.
This will prevent frustration in less dominant animals when access to a resource/structure is
restricted by dominant animals (Robitaille and Prescott, 1993).
F eatures surrounding the exhibit
Before construction for a new exhibit begins, features surrounding the proposed site should first
be considered. Orangutans are generally known to be intelligent and observant; therefore visitor
establishments or other animal exhibits in the surroundings may serve as enrichment. For

103
Singapore Zoo orangutans, there was a restaurant as well as a third orangutan exhibit next to their
enclosures. Constant exposure to visitors and keepers via daily photography sessions and regular
handling is expected to have habituated them to humans; hence, the increased amount of visitor
traffic due to the nearby restaurant may have been less stressful for this group of animals.
Orangutans while in both enclosures were often observed to look at the restaurant; presumably
they were watching the dining visitors. An escaped orangutan was once found seated at an
outdoor restaurant table, holding a spoon and a fork in either hand (Nantha 2009, pers. comm.13).
This suggests that it had previously observed visitors who were using utensils. Hence, the
opportunity to watch visitors may serve as a source of enrichment for these orangutans. Similarly,
the orangutans were often observed to watch some older orangutans which were found in a third
orangutan exhibit between the two free-ranging enclosures (see Fig 3.3). The free-ranging
orangutans showed particular interest when the older orangutans were manipulating objects, or
processing novel foods. Therefore, the presence of the third orangutan exhibit next to the two
free-ranging enclosures is thought to be especially enriching. Such a concept has also been used
in other zoos. For example, the Adelaide Zoo in Australia is building overhead lines and towers to
allow orangutans to pass over and rest above the adjacent tiger display (Coe and Dykstra, 2010).
Other animal exhibits in the surrounding of orangutan exhibits may hence serve as a form of
enrichment. However, response to environmental stimuli ultimately depends on individual
temperament and captive rearing history (Hosey et al., 2009).
orangutans first have to be considered before deciding if environmental stimuli will be enriching
or not.
13 Gabriel Nantha, Orangutan keeper, Singapore Zoo

104
Recommendations on husbandry routine
F eeding routine and regurgitation
Husbandry routines are generally known to have strong influence on captive animal behaviour
(Shepherdson et al., 1998). By varying the frequency and types of food provided, primate
caretakers can encourage species-specific behaviours and reduce inactive or stereotypic
behaviours (Kerridge, 2005). Most of the individuals in this study were noted to immediately feed
once browse (fresh leaves) or other foods were provided. Because wild orangutans spend more
than 50% of their time feeding (Morrogh-Bernard et al., 2009), reduced feeding opportunities in
captivity may lead to behaviours such as regurgitation, which is thought to be a way of increasing
feeding time (Akers and Schildkraut, 1985; Gould and Bres, 1986). Interestingly, regurgitation
was present in this group of orangutans, even though they were housed in a large, naturalistic
exhibit. Of the orangutans observed, only a few individuals performed regurgitation and
reingestion regularly. Two young orangutans in particular, Budi and Merlin (aged seven and five
respectively), regularly regurgitated after being fed. They spent on average, 15% of their activity
budget on this behaviour (see Appendix). Adhoc observations also showed that another young
orangutan, Bento (aged three), seemed to be learning from the other orangutans, and showed
increasing levels of regurgitation over the course of this study. Conversely, an older individual,
Chomel (aged 13), seemed to be making attempts to mimic this behaviour, but usually only
managed to regurgitate a bit of food, before moving on to another behaviour. Because
regurgitation can be caused by a multitude of psychological, housing or nutritional factors, and
also acquired via social learning (Lukas, 1999) especially when the animal is young, any
measures to address this behaviour need to be multi-pronged (Hill, 2009, Lukas, 1999). One
recommendation is to investigate the possible causes of regurgitation in this group of orangutans,
especially in Budi and Merlin, because the presence of this behaviour may indicate underlying

105
welfare issues. In particular, it may be interesting to look at captive rearing history for this group
of orangutans, for example, the age at which the orangutan was separated from its mother, and the
conditions under which it was reared when young. Also, the daily feedings of milk and Milo may
be interesting to consider, as regurgitation in captive gorillas was reduced by 28% when milk was
removed from their diet (Lukas et al., 1999).
An alternative way of varying the feeding routine would be to provide food at different areas in
the exhibit. Fresh leaves and snacks were usually provided at fixed areas in the exhibit; as a
result, feeding behaviour usually occurred at the same locations. By varying where food is
provided, greater enclosure use can be encouraged; this can also serve as a source of physical
exercise as animals have to move towards the keeper. In addition, besides providing leaves, foods
which require greater processing time may also be enriching. In the wild, orangutans are known
to feed on foods which have tough skins or require longer processing times (Mackinnon, 1974).
Food items of such nature (e.g. novel fruits, insects) should be incorporated into captive animal
feeding routines as much as possible.
Animal handling
The presence of visitor photography sessions and animal shows involving orangutans gives rise to
unusual husbandry routines in Singapore Zoo. Unlike many European and American zoos, hands-
on contact with orangutans is much more common in Singapore Zoo (Raj, 2010, pers. comm.14).
During visitor photography sessions which occur several times a day, orangutans and visitors sit
at close proximity from one another. For this to be possible, good keeper-animal relationships are
first necessary. A model of human-animal relationships by Hosey (2008) suggests that animals
14 Jackson Raj, Head orangutan keeper, Singapore Zoo

106
keepers, a strong bond of trust is present between the orangutans and their caretakers (pers. obs.;
orangutan keepers, pers. comm.). Anecdotal observations (in Singapore Zoo), such as orangutan
mothers which voluntarily hand their infants to the keepers provide evidence of the trust that
these orangutans have in their caretakers. This excellent keeper-animal relationship possibly
reduces fear of unfamiliar humans in the orangutans, hence visitor photography sessions, whereby
strange humans sit in close proximity to the orangutans, are possible. Similarly, any stressful
effects from visitors viewing the exhibits may be reduced. This particularly hands-on routine may
therefore improve welfare in this group of captive orangutans.
Other considerations and recommendations
Social housing for semi-solitary apes
Even though orangutans in the wild are semi-solitary, it is hypothesized that such social structures
are influenced by food availability. Orangutan food sources are located patchily throughout the
forest, and association in groups may increase costs of foraging (Setia et al., 2009). In captivity,
however, food availability is no longer a concern, and captive orangutans are known to be more
social (Zucker et al., 1986). Social stimulation may substitute for a lack of variability in captive
environments. This appears to be why orangutans in this study, despite having less feeding
activity than wild orangutans (in irregularly fruiting forests), were not more inactive than the
latter. The time not spent on feeding by captive animals was instead spent on social and play
behaviour. Because orangutans are intelligent animals (Tomasello and Call, 1997), housing
captive apes in social settings may be a good alternative which provides physical and mental
stimulation.

107
Visibility of orangutans in the Island exhibit
I observed that in the Island exhibit, more than half of the visitors passed under the trees without
noticing the orangutans. Considering the effort and costs spent on maintaining naturalistic
exhibits, it is suggested that more signage be put up to draw visitor attention to the arboreal apes.
Even though the orangutans are sometimes difficult to spot in the tree canopy, it has been found
that visitors enjoy searching for free-ranging animals, even when they do not find them (Price et
al., 1994). Hence, opportunities to experience the orangutans as arboreal animals may further
enhance visitor appreciation of this endangered ape (Coe, 1989; Finlay et al., 1988).
F easibility of replicating Singapore Zoo orangutan exhibit design
The orangutan enclosures in this study are complex and provide exposure to environmental
stimuli. They appear to reduce visitor stress in the animals, and allow for species-specific
behaviours. The orangutan keepers have also noted that the fur coats of the orangutans have
become glossier, and their muscles more well-defined, after the introduction of the exhibits. In
addition, because the exhibits were built upon existing trees, they do not take up additional space.
In view of these benefits, the replication of such an exhibit design in other zoos may be worthy of
consideration. However, some features of these enclosures must first be addressed.
One aspect of these orangutan exhibits which was not discussed earlier is how orangutans are
introduced into the enclosures. Unlike traditional enclosures where a sliding door or partition is
opened to let animals enter an exhibit, keeper-animal contact is required to introduce orangutans
into the free-ranging exhibits. Keepers hand-hold the orangutans and walk them out from their
night dens to specific areas in each exhibit. At a special point near each exhibit, a log can be
lowered to form a bridge onto the exhibit (see Fig. 5.1). When orangutans are to be introduced

108
into the enclosure, the log, operated by hydraulic mechanism, is lowered to join the ground and a
branch in one of the exhibit trees. At all other times, the log stands vertical, away from the exhibit
trees. Orangutans are prevented from leaving the exhibit by low-current electrical wires midway
up each tree. Such an exhibit design requires keepers to handle the animals on a daily basis. The
orangutan husbandry routine in Singapore Zoo is such that female orangutans are handled up to
more than 40 years of age by the keepers, and male orangutans have been safely handled up to 17
years of age. Because mature males may not be handled due to safety considerations, only
females and younger males, of below certain weights to prevent branch breakage, may be
displayed in the treetop exhibits. For purposes of replication elsewhere, this exhibit design could
be modified so that keeper-animal contact is not required for animal release. However,
orangutans have been seen on occasion to jump down from the tree canopy. These events are
sporadic, and are due to aggressive interactions between conspecifics. Occasionally, orangutans
also manage to exit the enclosure and escape onto surrounding trees. In view of this, there is
always at least one keeper watching the animals. In the event of enclosure escape, the keeper is
required to retrieve the animal and if necessary, return it to the night dens. This suggests that for
-
animals. This is so that the orangutans, which can jump down from the enclosures if they choose
to, are less likely to travel far, before being called back by their keepers.

109
F ig 5.1. Log that facilitates entry into Island orangutan exhibit. Left: Log not lowered, standing vertically away from exhibit (denoted by white arrow). Right: Log is lowered and bridges ground and exhibit.
Study limitations and future directions
In this study, results were pooled for animals in the same age group. When individual data were
examined (see Appendix), some orangutans in the same age group showed very different levels of
the same behaviour. Animal welfare has been defined as being specific for each individual, hence
consideration of individual (Hosey et al., 2009). Such unique
characteristics occur within a species and are something that animal caretakers are generally
aware of. It was discussed in the visitor effects chapter how individual orangutans differed in
their responses to visitors. Studies on rats also show that differing degrees of neophobia (fear of
novelty) in rats were linked to different probabilities of illness and death. Animals which were
more active in exploring their surroundings were more likely to live longer than their neophobic
siblings. (Cavigelli et al., 2006; Hosey et al., 2009). Considering such personality effects, zoo

110
management could try to take into account individual orangutan preferences when designing their
husbandry routine and enrichment programmes. Future studies that match great ape personalities
via keeper surveys to empirical behavioural observations (Uher and Asendorpf, 2008; Uher et al.,
2008), may also be useful in improving captive orangutan welfare.
C O N C L USI O NS
The study of activity budgets, enclosure use, and visitor effects in a group of captive orangutans
has provided considerable information on behaviour in two naturalistic, novel enclosures. This
knowledge will be useful for zoo management and can provide guidelines for future study on
captive orangutan behaviour.

111
R E F E R E N C ES Akers JS, Schildkraut DS. 1985. Regurgitation/reingestion and coprophagy in captive gorillas.
Zoo Biology 4(2):99-109.
Altmann J, Muruthi P. 1988. Differences in daily life between semiprovisioned and wild-feeding baboons. American Journal of Primatology 15(3):213-221.
Anderson US, Benne M, Bloomsmith MA, Maple TL. 2002. Retreat space and human visitor density moderate undesirable behavior in petting zoo animals. Journal of Applied Animal Welfare Science 5:125 137.
Baker KC, Easley SP. 1996. An analysis of regurgitation and reingestion in captive chimpanzees. Applied Animal Behaviour Science 49(4):403-415.
Bayne K, Mainzer H, Dexter S, Campbell G, Yamada F, Suomi S. 1991. The reduction of abnormal behaviors in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology 23(1):23-35.
Bettinger T, Carter JWT. 1994. Spatial selection in captive adult female chimpanzees. Zoo Biology 13(2):167-176.
Birke JF. 2002. Effects of browse, human visitors and noise on the behaviour of captive orangutans. Animal Welfare 11:189-202.
Blasetti A, Boitani L, Riviello MC, Visalberghi E. 1988. Activity budgets and use of enclosed space by wild boars (Sus scrofa) in captivity. Zoo Biology 7(1):69-79.
Blinder BJ, Goodman SL, Goldstein R, editors. 1988. Rumination: a critical review of diagnosis and treatment. New York, NY: PMA Publishing. 315-329 p.
Bloomfield RC, Gillespie G, Hemsworth PH. 2010. The Effects of Zoo Visitors on the Behaviour of Orang-utans. Proceedings of the 2010 ARAZPA Conference. Healesville, Victoria, Australia.
Bloomsmith MA. 2009. Measuring Zoo Animal Welfare. Journal of Applied Animal Welfare Science 12(3):273 - 275.
Bowen RA. 1980. The behaviour of three hand-reared lowland gorillas, with emphasis on the response to a change in accomodation. Dodo 17:63-79.
Brennan EJ, Else JG, Altmann J. 1985. Ecology and behaviour of a pest primate: vervet monkeys in a tourist-lodge habitat. African Journal of Ecology 23(1):35-44.
Brent L, Lee DR, Eichberg JW. 1991. Evaluation of a chimpanzee enrichment enclosure. Journal of Medical Primatology, 20:29 34.
Britt A. 1998. Encouraging natural feeding behavior in captive-bred black and white ruffed lemurs (Varecia variegata variegata). Zoo Biology 17(5):379-392.

112
Carder G, Semple S. 2008. Visitor effects on anxiety in two captive groups of western lowland gorillas. Applied Animal Behaviour Science 115:211 220.
Cartmill E, Byrne R. 2010. Semantics of primate gestures: intentional meanings of orangutan gestures. Animal Cognition 13(6):793-804.
Cartmill EA, Byrne RW. 2007. Orangutans modify their gestural signaling according to their audience's comprehension. Current Biology 17(15):1345-1348.
Cassella C. 2010. Activity Budgets, Use of Space, and Undesirable Behaviors in Cleveland Metroparks Zoo Orangutans On- and Off-Exhibit. Cleveland, OH, USA: Cleveland Metroparks Zoo.
Caton J, Hume I, Hml D, Harper P. 1999. Digesta retention in the gastro-intestinal tract of the orang utan (Pongo pygmaeus) Primates 40(4):551-558.
Cavigelli SA, Yee JR, McClintock MK. 2006. Infant temperament predicts life span in female rats that develop spontaneous tumours. Hormones and behavior 50:454-462.
Chamove AS, Hosey GR, Schaetzel P. 1988. Visitors excite primates in zoos. Zoo Biology 7(4):359-369.
Cipreste CF, de Azevedo CS, Young RJ. 2010. How to develop a zoo-based environmental enrichment program: incorporating environmental enrichment into exhibits. In: Kleiman DG, Thompson KV, Baer CK, editors. Wild Mammals in captivity: principles and techniques for zoo management. Chicago and London: The University of Chicago Press.
Clarke AS, Juno CJ, Maple TL. 1982. Behavioral effects of a change in the physical environment: A pilot study of captive chimpanzees. Zoo Biology 1(4):371-380.
Coe JC. 1985. Design and perception: Making the zoo experience real. Zoo Biology 4(2):197-208.
Coe JC. 1989. Naturalizing habitats for captive primates. Zoo Biology 8(S1):117-125.
Coe JC, Dykstra G. 2010. New and sustainable directions in zoo exhibit design. In: Kleiman DG, Thompson KV, Baer CK, editors. Wild Mammals in captivity: principles and techniques for zoo management. Chicago and London: The University of Chicago Press.
Cook S, Hosey GR. 1995. Interaction sequences between chimpanzees and human visitors at the Zoo. Zoo Biology 14(5):431-440.
Cooke CM, Schillaci MA. 2007. Behavioral responses to the zoo environment by white handed gibbons. 106(1):125-133.
Daan S, Aschoff J. 1975. Circadian rhythms of locomotor activity in captive birds and mammals: Their variations with season and latitude. Oecologia 18(4):269-316.
Davenport RK, Jr. 1967. The orang-utan in Sabah. Folia Primatologica 5:247-263.
1903.

113
Davey G. 2007. Visitors' effects on the welfare of animals in the zoo: A review. Journal of Applied Animal Welfare Science 10(2):169-183.
Davis N, Schaffner C, Smith T. 2005. Evidence that zoo visitors influence HPA activity in spider monkeys (Ateles geoffroyii rufiventris). Applied Animal Behaviour Science 90:131 141.
Delgado RA, Van Schaik CP. 2000. The behavioral ecology and conservation of the orangutan (Pongo pygmaeus): A tale of two islands. Evolutionary Anthropology: Issues, News, and Reviews 9(5):201-218.
Dettling A. Physical environment and its influence on behaviour in captive common marmosets (Callithrix jacchus). In: Pryce C, Scott I, Schnell C, editors; 1997; Salisbury, UK. p 54-59.
Duncan IJH. 1970. Frustration in the Fowl. In: Freeman BM, Gordon RF, editors. Aspects of Poultry Behaviour. Edinburgh: British Poultry Science Ltd. p 15-31.
Edwards S, Snowdon C. 1980. Social behavior of captive, group-living Orangutans. International Journal of Primatology 1(1):39-62.
Elftman H, Atkinson WB, editors. 1950. The abdominal viscera of the gorilla. New York, NY: Columbia University Press. 197-201 p.
Engel J. 1996. Choosing an appropriate sample interval for instantaneous sampling. Behavioural Processes 38:11-17.
Erwin J, Deni R. 1979. Strangers in a strange land: abnormal behaviors or abnormal environments? Erwin J, Maple T, Mitchell G, editors. New York: Van Nostrand Reinhold.
Estevez I, Christman MC. 2006. Analysis of the movement and use of space of animals in confinement: The effect of sampling effort. Applied Animal Behaviour Science 97(2-4):221-240.
Fa J. 1992. Visitor-directed aggression among the Gibraltar macaques. Zoo Biology 11:43-52.
Housing, care and Psychological Well-Being of Captive and Laboratory Primates. Park Ridge, USA: Noyes Publications. p 270 290.
Fagen R. 1981. Animal Play Behavior. New York: Oxford University Press.
Fernandez EJ, Tamborski MA, Pickens SR, Timberlake W. 2009. Animal-visitor interactions in -8.
Field A. 2009. Discovering Statistics Using SPSS. 3rd ed. London: Sage Publications Ltd.
environment. Environment and Behavior 20:508-528.
Fischer R, Nadler R. 1978. Affiliative, playful, and homosexual interactions of adult female lowland gorillas. Primates 19(4):657-664.

114
Fischer RB, Nadler RD. 1977. Status interactions of captive female lowland gorillas. Folia Primatol 28:122-133.
uralistic approach. Thylac 16:12 17.
Forthman DL, Bakeman R. 1992. Environmental and social influences on enclosure use and activity patterns of captive sloth bears (Ursus ursinus). Zoo Biology 11(6):405-415.
Forthman DL, Brown MT, Perkins LA, Gold KC. Factors influencing orangutan (Pongo pygmaeus abelii) behavior in indoor holding facilities and outdoor exhibits. American Association of Zoological Parks and Aquariums Annual Proceedings; 1993; Wheeling. p 269 274.
Fox MW. 1971. Psychopathology in man and lower animals. Journal of the American Veterinary Medicine Association 159(1):66-77.
Fraser D. 1975. Vocalizations of isolated piglets. I. Sources of variation and relationships among measures. Applied Animal Ethology 1:387-394.
Fraser D. 2009. Animal behaviour, animal welfare and the scientific study of affect. Applied Animal Behaviour Science 118:108-117.
Galdikas BMF. 1985a. Orangutan sociality at Tanjung Puting. American Journal of Primatology 9(2):101-119.
Galdikas BMF. 1985b. Subadult male orangutan sociality and reproductive behavior at Tanjung Puting. American Journal of Primatology 8:87 99.
Galdikas BMF. 1988. Orangutan diet, range and activity at Tanjung Puting, Central Borneo. International Journal of Primatology 9(1):1-35.
Gippoliti S. 2000. Orang-utans in Zoos: Husbandry, Welfare and Management in an Atypical Arboreal Solitary Mammal. International Zoo News 47(6):356-368.
Glatston AR, Geilvoet-Soeteman E, Hora-Pecek E, Van Hooff JARAM. 1984. The influence of the zoo environment on social behavior of groups of cotton-topped tamarins, Saguinus oedipus oedipus. Zoo Biology 3(3):241-253.
Goff C, Howell SM, Fritz J, Nankivell B. 1994. Space use and proximity of captive chimpanzee (Pan troglodytes) mother/offspring pairs. Zoo Biology 13(1):61-68.
Goodall
Goodall J. 1986. The Chimpanzees of Gombe:Patterns of Behavior. Cambridge: Harvard University Press.
Gould E, Bres M. 1986. Regurgitation and reingestion in captive gorillas: Description and intervention. Zoo Biology 5(3):241-250.
Gould KG. 1983. Diagnosis and treatment of infertility in male great apes. Zoo Biology 2(4):281-293.

115
Grisham J, Lyon F, Pearson P, Macfarlane C. 2000. Great EscApe: the great ape facility at Oklahoma City Zoological Park. International Zoo Yearbook 37(1):366-374.
Groves C, Wilson DE, Reeder DM. 2005. Mammal Species of the World. Baltimore: Johns Hopkins University Press.
Hebert PL, Bard K. 2000. Orangutan use of vertical space in an innovative habitat. Zoo Biology 19(4):239-251.
Hedeen SE. 1982. Utilization of space by captive groups of lowland gorillas (Gorilla g. gorilla). Ohio Journal of Science 82:27-30.
Hill SP. 2009. Do gorillas regurgitate potentially-injurious stomach acid during 'regurgitation and reingestion?'. Animal Welfare 18:123-127.
Hoff MP, Forthman DL, Maple TL. 1994. Dyadic interactions of infant lowland gorillas in an outdoor exhibit compared to an indoor holding area. Zoo Biology 13(3):245-256.
Hoff MP, Powell DM, Lukas KE, Maple TL. 1997. Individual and social behavior of lowland gorillas in outdoor exhibits compared with indoor holding areas. Applied Animal Behaviour Science 54(4):359-370.
Honess PE, Marin CM. 2006. Enrichment and aggression in primates. Neuroscience & Biobehavioral Reviews 30(3):413-436.
Horikoshi-Beckett C, Schulte BA. 2006. Activity patterns and spatial use of facility by a group of captive female manatees (Trichechus manatus latirostris). Zoo Biology 25(4):285-301.
Hosey G, Melfi V, Pankhurst S. 2009. Zoo Animals: Behaviour, Management and Welfare. New York: Oxford University Press Inc.
Hosey GR. 2000. Zoo animals and their human audience: what is the visitor effect? Animal Welfare 9:343-357.
Hosey GR. 2008. A preliminary model of human-animal relationships in the zoo. Applied Animal Behaviour Science 109:105 127.
Applied Animal Behaviour Science 18:19 29.
Isbell L, Young T. 1993. Social and ecological influences on activity budgets of vervet monkeys, and their implications for group living. Behavioral Ecology and Sociobiology 32(6):377-385.
IUCN. 28 December 2010. IUCN Red List of Threatened Species. Version 2010.4. http://www.iucnredlist.org.
Jaman MF, Huffman MA. 2008. Enclosure environment affects the activity budgets of captive Japanese macaques (Macaca fuscata). American Journal of Primatology 70(12):1133-1144.

116
James-Aldridge V, Gorena R. 1991. Use of a naturalistic environment by a group of captive chimpanzees. American Journal of Primatology 24:109-110.
Jensvold MLA, Sanz CM, Fouts RS, Fouts DH. 2001. Effect of Enclosure Size and Complexity on the Behaviors of Captive Chimpanzees (Pan troglodytes). Journal of Applied Animal Welfare Science 4(1):53 - 69.
Jones J. 2003. The physiology and behavioural changes in a group of orang-utans in response to different enclosure settings and visitor levels. B. Sc. thesis. Bolton, UK: University of Manchester (Bolton Institute).
Kerridge FJ. 2005. Environmental enrichment to address behavioral differences between wild and captive black-and-white ruffed lemurs (Varecia variegata). American Journal of Primatology 66(1):71-84.
Kingsley S. 1988. Physiological development of male orangutans and gorillas. Schwartz JH, editor. Oxford: Oxford University Press.
Kisling VN. 2001. Zoo and aquarium history: ancient animal collections to zoological gardens. New York, NY/London: CRC Press.
Kleiman DG. 1994. Foreword: Animal behavior studies and zoo propagation programs. Zoo Biology 13(5):411-412.
Knott CD. 1998. Changes in orangutan caloric intake, energy balance, and ketones in response to fluctuating fruit availability. International Journal of Primatology 19(6):1061-1079.
Kolakowski L. 1968. The alienation of reason: a history of positivist thought Guterman N, translator. New York: Doubleday. (cited in Fraser, 2009).
Kuhar CW. 2006. In the deep end: pooling data and other statistical challenges of zoo and aquarium research. Zoo Biology 25(4):339-352.
Leavens DA. 2007. Animal cognition: Multimodal tactics of Orangutan communication. Current Biology 17(17):R762-R764.
Lee PC, Brennan EJ, Else JG, Altmann J. 1986. Ecology and behavior of vervet monkeys in a tourist lodge habitat. Else JG, Lee PC, editors. Cambridge: Cambridge University Press.
Leighty KA, Soltis J, Savage A. 2010. GPS Assessment of the Use of Exhibit Space and Resources by African Elephants (Loxodonta africana). Zoo Biology 29(2):210-220.
Leuck BE. 1977. Differential use of space by eight species of birds in a free-flight zoological park aviary. Applied Animal Ethology 3(2):105-126.
Little KA, Sommer V. 2002. Change of enclosure in langur monkeys: implications for the evaluation of environmental enrichment. Zoo Biology 21(6):549-559.
Lukas KE. 1999. A review of nutritional and motivational factors contributing to the performance of regurgitation and reingestion in captive lowland gorillas (Gorilla gorilla gorilla). Applied Animal Behaviour Science 63(3):237-249.

117
Lukas KE, Hamor G, Bloomsmith MA, Horton CL, Maple TL. 1999. Removing milk from captive gorilla diets: the impact on regurgitation and reingestion (r/r) and other behaviors Zoo Biology 18:515-528.
Mackinnon J. 1974. The behaviour and ecology of wild orang-utans (Pongo pygmaeus). Animal Behaviour 22(1):3-74.
Mahler AE. 1984. Activity budgets and use of exhibit space by South American tapir (Tapirus terrestris) in a Zoological Park setting. Zoo Biology 3(1):35-46.
Mallapur A, Chellam R. 2002. Environmental influences on stereotypy and the activity budget of Indian leopards (Panthera pardus) in four zoos in southern India. Zoo Biology 21(6):585-595.
lion-tailed macaques (Macaca silenus) housed in Indian zoos. Applied Animal Behaviour Science 94:341 352.
Mallinson JJC. 1984. The breeding of great apes at the Jersey Wildlife Preservation Trust and a look into the future. Zoo Biology 3(1):1-11.
Manning C. 2002. Enclosure utilisation and behaviour in the female orang utans (Pongo pygmaeus) of Paignton Zoo. In: Dow S, editor. Annual Symposium on Zoo Research. Bristol Zoo Gardens, Bristol: The Federation of Zoological Gardens of Great Britain and Ireland, London. p 44-50.
Maple T. 1979. Great apes in captivity: the good, the bad, and the ugly. Erwin J, Maple T, Mitchell G, editors. New York: Van Nostrand Reinhold.
Maple TL. 1980. Orang-utan behavior. New York: Van Nostrand Reinhold.
Maple TL, Finlay TW. 1989. Applied primatology in the modern zoo. Zoo Biology 8(S1):101-116.
Maple TL, Stine WW. 1982. Environmental variables and great ape husbandry. American Journal of Primatology 3(S1):67-76.
Marchal V. 2004. Social interactions among a group of Sumatran Orangutans (Pongo pygmaeus abelii) at Jersey Zoo. Jersey, Channel Islands: Durrell Wildlife Conservation Trust. p 1-28.
Markham R, J. 1990. Breeding orangutans at Perth Zoo: Twenty years of appropriate husbandry. Zoo Biology 9(2):171-182.
Marks K. 2009 Orangutan's great escape causes zoo evacuation. The Independent.
Marriner LM, Drickamer LC. 1994. Factors influencing stereotyped behavior of primates in a zoo. Zoo Biology 13(3):267-275.
Mather L. 1999. Response of captive orang utans to human audiences. MPhil thesis. Bolton, UK: University of Manchester (Bolton Institute).

118
Matsuda I, Murai T, Clauss M, Yamada T, Tuuga A, Bernard H, Higashi S. 2011. Regurgitation and remastication in the foregut-fermenting proboscis monkey (Nasalis larvatus). Biology Letters.
Mayes SD, Humphrey, F.J., Handford, H.A., Mitchell, J.F. 1988. Rumination disorder: differential diagnosis. Journal of the American Academy of Child and Adolescent Psychiatry 27:300-302.
Melfi VA. 2009. There are big gaps in our knowledge, and thus approach, to zoo animal welfare: a case for evidence-based zoo animal management. Zoo Biology 28:574 588.
Melfi VA, Feistner A. 2002. A Comparison of the Activity Budgets of Wild and Captive Sulawesi Crested Black Macaques (Macaca Nigra). Animal Welfare 11:213-222.
Millman ST. 2009. Animal welfare scientific approaches to the issues. Journal of Applied Animal Welfare Science 12:88 96.
- across species, across disciplines, and across borders. Journal of Applied Animal Welfare Science 12:83 87.
Mitchell F, Tromborg C, Kaufman J, Bargabus S, Simoni R, Geissler V. 1992a. More on the
Behaviour Science 35:189-198.
Mitchell G, Herring F, Obradovich S. 1992b. Like threaten like in mangabeys and people? Anthrozoös 5:106 112.
Mitchell G, Herring F, Obradovich S, Tromborg C, Dowd B, Neville LE, Field L. 1991. Effects of visitors and cage changes on the behaviors of mangabeys. Zoo Biology 10(5):417-423.
Mitchell G, Obradovich S, Sumner D, DeMorris K, Lofton L, Minor J, Cotton L, Foster T. 1990. Cage location effects on visitor attendance at three Sacramento Zoo mangabey enclosures. Zoo Biology 9(1):55-63.
Biology 9:275-286.
Morbeck ME, Zihlman AL. 1988. Body Composition and Limb proportions. Schwartz JH, editor. Oxford: Oxford University Press.
Morgan KN, Tromborg CT. 2007. Sources of stress in captivity. Applied Animal Behaviour Science 102:262-302.
Morrogh-Bernard HC, Husson SJ, Knott CD, Wich SA, van Schaik CP, van Noordwijk MA, Lackman-Ancrenaz I, Marshall AJ, Kanamori T, Kuze N, bin Sakong R. 2009. Orangutan activity budgets and diet: a comparison between species, populations and habitats. In: Wich SA, Utami Atmoko SS, Mitra Setia T, van Schaik CP, editors. Orangutans : geographic variation in behavioral ecology and conservation. New York, US: Oxford University Press. p 119-133.

119
Nash LT. 1993. Juveniles in nongregarious primates. In: Pereira ME, Fairbanks LA, editors. Juvenile primates: Life history, development, and behavior. Oxford, UK: Oxford University Press. p 119 - 137.
Oates JF, editor. 1987. Food distribution and foraging behaviour. Chicago: Chicago University Press. 197 209 p.
Ogden JJ, Capanzano C, Maple TL. Immersion exhibits: How are they proving as educational exhibits?; 1994. p 224 228.
Ogden JJ, Finlay TW, Maple TL. 1990. Gorilla adaptations to naturalistic environments. Zoo Biology 9(2):107-121.
Ogden JJ, Lindburg DG, Maple TL. 1993. Preference for structural environmental features in captive lowland gorillas (Gorilla gorilla gorilla). Zoo Biology 12(4):381-395.
Orangutan. Singapore Zoo. http://www.zoo.com.sg/l2_t2.aspx?l1=5&l2=39&langid=1. Accessed 13 Jan 2011.
Oswald M, Kuyk K. 1977. The behavior of three lorisoid primate species before and after the public opening of the nocturnal house. In: Crockett C, Hutchins M, editors. Applied behavioral research the Woodland Park Zoological Gardens. Seattle, WA: Pika Press. p 81 100.
Parker ST, Gibson KR. 1977. Object manipulation, tool use and sensorimotor intelligence as feeding adaptations in cebus monkeys and great apes. Journal of Human Evolution 6(7):623-641.
Pearson EL, Davis JM, Litchfield CA. 2010. A Case Study of Orangutan and Siamang Behavior Within a Mixed-Species Zoo Exhibit. Journal of Applied Animal Welfare Science 13(4):330-346.
Perkins L, Bradfield E. 1989. Orangutans of Ketambe: Enclosure Utilization. American Association of Zoological Parks and Aquariums Regional Conference Proceedings. Wheeling, WV: American Association of Zoological Parks and Aquariums. p 564-670.
Perkins LA. 1992. Variables that influence the activity of captive orangutans. Zoo Biology 11(3):177-186.
Pfeiffer AJ, Koebner LJ. 1978. The resocialization of single-caged chimpanzees and the establishment of an island colony. Journal of Medical Primatology, 7(2):70-81.
Pizzutto CS, Nichi M, Sgai MGFG, Corrêa SHR, Viau P, Beresca AM, de Oliveira CA, Barnabé RC, de Barros Vaz Guimarães MA. 2008. Effect of Environmental Enrichment on Behavioral and Endocrine Aspects of a Captive Orangutan (Pongo pygmaeus). Laboratory Primate Newsletter 47(2):10-14.
Plowman AB. 2008. BIAZA statistics guidelines: toward a common application of statistical tests for zoo research. Zoo Biology 27:226-323.

120
Poole TB. 1987. Social behavior of a group of orangutans (Pongo pygmaeus) on an artificial island in Singapore Zoological Gardens. Zoo Biology 6(4):315-330.
Pope J. 2009 Ape escape: Audubon Zoo orangutan breaks out for a few minutes freedom. The Times.
Price EC, Ashmore LA, McGivern A-M. 1994. Reactions of zoo visitors to free-ranging monkeys. Zoo Biology 13(4):355-373.
Rijksen HD. 1975. Social structure in a wild orang-utan population in Sumatra. In: Kondo S, Kawai M, Ehara A, editors. Contemporary Primatology. Basel: S. Karger. p 373-379.
Rijksen HD, Meijaard E. 1999. Our vanishing relative: the status of wild orangutans at the close of the twentieth century. Dordrecht: Kluwer Academic Publishers.
Robitaille JF, Prescott J. 1993a. Use of space and activity budgets in relation to age and social-status in a captive herd of American Bison, Bison-bison. Zoo Biology 12(4):367-379.
Robitaille JF, Prescott J. 1993b. Use of space and activity budgets in relation to age and social status in a captive herd of American bison, Bison bison. Zoo Biology 12(4):367-379.
Rodman PS, Mitani JC. 1987. Orangutans: sexual dimorphism in a solitary species. Smuts B, Cheney DL, Seyfarth RM, Struhsaker T, Wrangham RW, editors. Chicago: University of Chicago Press.
Ross SR, Calcutt S, Schapiro SJ, Hau J. 2011. Space use selectivity by chimpanzees and gorillas in an indoor outdoor enclosure. American Journal of Primatology 73(2):197-208.
Ross SR, Lukas KE. 2006. Use of space in a non-naturalistic environment by chimpanzees (Pan troglodytes) and lowland gorillas (Gorilla gorilla gorilla). Applied Animal Behaviour Science 96(1-2):143-152.
Ross SR, Schapiro SJ, Hau J, Lukas KE. 2009. Space use as an indicator of enclosure appropriateness: A novel measure of captive animal welfare. Applied Animal Behaviour Science 121(1):42-50.
Russon A. 1998. The nature and evolution of intelligence in orangutans (Pongo pygmaeus). Primates 39(4):485-503.
Russon AE, Bard K. 1996. Exploring the minds of great apes: issues and controversies. In: Russon AE, Bard K, Parker S, editors. Reaching into thought: the minds of great apes. Cambridge: Cambridge University Press. p 1 20.
Schafer EH. 1968. Hunting parks and animal enclosures in ancient China. Journal of the Economic and Social History of the Orient 11:318-343.
Seidensticker J, Doherty J. 1996. Integrating animal behavior and exhibit design. Kleiman DG, Allen ME, Thompson KV, Lumpkin S, editors. Chicago, IL: University of Chicago Press.
Sellinger R, Ha J. 2005. The effects of visitor density and intensity on the behavior of two captive jaguars (Panthera onca). Journal of Applied Animal Welfare Science 8:233 244.

121
Setia TM, Delgado RA, Utami Atmoko SS, Singleton I, van Schaik CP. 2009. Social organization and male-female relationships. In: Wich SA, Utami Atmoko SS, Setia TM, van Schaik CP, editors. Orangutans: Geographic Variation in Behavioral Ecology and Conservation. Oxford: Oxford University Press.
Shen-Jin L, Todd PA, Yan Y, Lin Y, Hongmei F, Wan-Hong W. 2010. The effects of visitor density on sika deer (Cervus nippon) behaviour in Zhu-Yu-Wan Park, China. Animal Welfare 19:61-65.
Shepherdson DJ. 2010. Principles of and research on environmental enrichment for mammals. In: Kleiman DG, Thompson KV, Baer CK, editors. Wild Mammals in captivity: principles and techniques for zoo management. Chicago and London: The University of Chicago Press.
Shepherdson DJ, Carlstead K, Mellen JD, Seidensticker J. 1993. The influence of food presentation on the behavior of small cats in confined environments. Zoo Biology 12(2):203-216.
Shepherdson DJ, Mellen JD, Hutchins M, editors. 1998. Second nature: environmental enrichment for captive animals. Washington, DC: Smithsonian Instituition Press.
Shettel-Neuber J. 1988. Second- and third-generation zoo exhibits: a comparison of visitor, staff, and animal responses. Environment and Behavior 20(4):452-473.
Simpson L. 2004 The effect of visitors on captive non-human primates. Zoo Federation Research Newsletter:6.
Singleton I, van Schaik CP. 2001. Orangutan home range size and its determinants in a Sumatran swamp forest. International Journal of Primatology 22(6):877-911.
Smith EO. 1978. Social Play in Primates. Annual Meeting of the Animal Behavior Society. New York: Academic Press.
Smith RJ, Jungers WL. 1997. Body mass in comparative primatology. Journal of Human Evolution 32(6):523-559.
Smith T. 2004. Zoo Research Guidelines: Monitoring Stress in Zoo Animals. In: BIAZA, editor. London.
Smuts BB. 1999. Sex and Friendship in Baboons. Cambridge, USA: Harvard University Press.
Snowdon CT. 1989. The criteria for successful captive propagation of endangered primates. Zoo Biology 8(S1):149-161.
Sonti C. 2009 Orang-utan's great escape from Perth Zoo. WA News.
Stoinski TS, Hoff MP, Maple TL. 2001. Habitat Use and Structural Preferences of Captive Western Lowland Gorillas (Gorilla gorilla gorilla): Effects of Environmental and Social Variables. International Journal of Primatology 22(3):431-447.
Strouhal E. 1992. Life in ancient Egypt. Cambridge: Cambridge University Press.

122
Struck K, Videan EN, Fritz J, Murphy J. 2007. Attempting to reduce regurgitation and reingestion in a captive chimpanzee through increased feeding opportunities: a case study. Lab Anim 36(1):35-38.
Struhsaker TT, editor. 1977. Auditory communication among vervet monkey (Cercopithecus aethiops). Chicago: University of Chicago Press.
Thame MM, Burton KA, Forrester TE. 2000 The human ruminant. West Indian Medical Journal 49:172-174.
Thompson V. 1976. Observation of the great apes in a naturalistic zoo environment. Chicago: Lincoln Park Zoo.
Thompson V. 1989. Behavioral response of 12 ungulate species in captivity to the presence of humans. Zoo Biology 8:275 297.
Thorpe SKS, Crompton RH, Alexander RM. 2007. Orangutans use compliant branches to lower the energetic cost of locomotion. Biology Letters 3(3):253-256.
Tobach E, Greenberg G, Radell P, McCarthy T. 1989. Social behavior in a group of orang-utans (Pongo pygmaeus abelii) in a zoo setting. 23(1):141-154.
Todd P, Macdonald C, Coleman D. 2008. Within-group differences in captive Diana monkey (Cercopithecus diana diana) behaviour. Journal of Ethology 26(2):273-278.
Todd PA, Macdonald C, Coleman D. 2007. Visitor-associated variation in captive Diana monkey (Cercopithecus diana diana) behaviour. Applied Animal Behaviour Science 107:162-165.
Tomasello M, Call J. 1997. Primate Cognition. New York: Oxford University Press.
Traylor-Holzer K, Fritz P. 1985. Utilization of space by adult and juvenile groups of captive chimpanzees (Pan troglodytes). Zoo Biology 4(2):115-127.
Tripp JK. 1985. Increasing activity in captive orangutans: Provision of manipulable and edible materials. Zoo Biology 4(3):225-234.
Uher J, Asendorpf JB. 2008. Personality assessment in the Great Apes: Comparing ecologically valid behavior measures, behavior ratings, and adjective ratings. Journal of Research in Personality 42(4):821-838.
Uher J, Asendorpf JB, Call J. 2008. Personality in the behaviour of great apes: temporal stability, cross-situational consistency and coherence in response. Animal Behaviour 75(1):99-112.
Utami Atmoko SS, Singleton I, van Noordwijk MA, van Schaik CP, Setia TM. 2009. Male-male relationships in orangutans. In: Wich SA, Utami Atmoko SS, Setia TM, van Schaik CP, editors. Orangutans: Geographic Variation in Behavioral Ecology and Conservation. Oxford: Oxford University Press. p 227.
van Rooijen J. 1991. Predictability and boredom. Applied Animal Behaviour Science 31:283-287.

123
van Schaik CP, Ancrenaz M, Borgen G, Galdikas B, Knott CD, Singleton I, Suzuki A, Utami SS, Merrill M. 2003. Orangutan Cultures and the Evolution of Material Culture. Science 299(5603):102-105.
van Schaik CP, Ancrenaz M, Djojoasmoro R, Knott CD, Morrogh-Bernard H, Nuzuar, Odom K, Utami Atmoko SS, van Noordwijk MA. 2009. Orangutan cultures revisited. In: Wich SA, Utami Atmoko SS, Setia TM, van Schaik CP, editors. Orangutans: Geographic Variation in Behavioral Ecology and Conservation. Oxford: Oxford University Press.
van Schaik CP, Deaner RO, Merrill MY. 1999. The conditions for tool use in primates: implications for the evolution of material culture. Journal of Human Evolution 36(6):719-741.
Verdade LM, Piña CI, Araújo JLO. 2006. Diurnal use of space by captive adult broad-snouted caiman (Caiman latirostris): Implications for pen design. Aquaculture 251(2-4):333-339.
Watts DP. 1988. Environmental influences on mountain gorilla time budgets. American Journal of Primatology 15(3):195-211.
Wells DL. 2005. A note on the influence of visitors on the behaviour and welfare of zoo-housed gorillas. Applied Animal Behaviour Science 93(1-2):13-17.
Wich SA, Utami Atmoko SS, Setia TM, van Schaik CP, editors. 2009. Orangutans: Geographic Variation in Behavioral Ecology and Conservation. Oxford: Oxford University Press.
Wiepkema PR, Koolhaas JM. 1993. Stress and animal welfare. Animal Welfare 2:195-218.
Wilson S, F. 1982. Environmental influences on the activity of captive apes. Zoo Biology 1(3):201-209.
Wood W. 1998. Interactions among environmental enrichment, viewing crowds, and zoo chimpanzees (Pan troglodytes). Zoo Biology 17:211-230.
Young RJ. 2003. Environmental enrichment for captive animals. Kirkwood JK, Hubrecht RC, Roberts EA, editors. Oxford: Blackwell Science Ltd.
Zoolex. 9 January 2011. Melbourne Zoo Orang-utan Sanctuary. http://www.zoolex.org/zoolexcgi/view.py?id=858.
Zucker E, Mitchell G, Maple T. 1978. Adult male-offspring play interactions within a captive group of orang-utans (Pongo pygmaeus). Primates 19(2):379-384.
Zucker EL, Clarke MR. 1992. Developmental and Comparative Aspects of Social Play of Mantled Howling Monkeys in Costa Rica. Behaviour 123(1/2):144-171.
Zucker EL, Dennon MB, Puleo SG, Maple TL. 1986. Play profiles of captive adult orangutans: A developmental perspective. Dev Psychobiol 19(4):315-326.
124
APPE NDI X
1. Activity budgets for individual orangutans
0
10
20
30
40
50
60
Perc
enta
ge o
f Act
ivity
bud
get (
%)
(1) Bento(1) Merlin(2) Budi(2) Gunta(3) Vira(3) Labu(3) Chomel(4) Binti(4) Anita
0
10
20
30
40
50
60
Per
cent
age o
f A
ctiv
ity B
udge
t (%
)
(1) Bento(1) Merlin(2) Budi(2) Gunta(3) Vira(3) Labu(3) Chomel(4) Binti(4) Anita
F ig. 1. Activity budgets for individual orangutans in Boardwalk (top) and Island (bottom) exhibits (number in bracket indicates age group).

125
Although there were trends in behaviour across age groups, it is interesting to note that
orangutans of similar age can differ greatly in their behaviour. The following are some examples.
Although Budi and Gunta were both 7 years of age, Gunta was much more attentive to his
idled less as well. Similarly, comparing Anita and Binti, who were around the same age (25 and
24 respectively), Anita spent about 50% of her activity budget idling (which was double of almost
ovement was
rearing history, in which she was the only orangutan amongst the nine studied to be human-
reared, rather than mother-reared. Unlike the other animals, especially Binti, who all appear to be
on the slim slide, it is interesting that Anita is the only orangutan that looks slightly obese. Hence,
rearing history may play an important role in individual behaviour.
Alertness to surrounding, i.e. looking was distinctly higher at 30-40% in three individuals, Gunta,
Labu (aged 11) and Binti. They looked almost 150% of that in other orangutans. Similarly, Budi
and Merlin showed the highest amounts of regurgitation, between 15-20%, which was triple or
quadruple that of other orangutans. And human interaction (i.e. begging for food from visitors)
was mostly performed by Chomel, Labu and Budi.
The amount of play by Bento (age 2) was double that of Merlin (age 4). Although this difference
in play could be due to age differences, it was also possible that the presence of younger
immature animals play more than adults, it is known that the proportion of immature individuals
in the group is critical factor. (Zucker and Clarke, 1992). Additional evidence comes when we
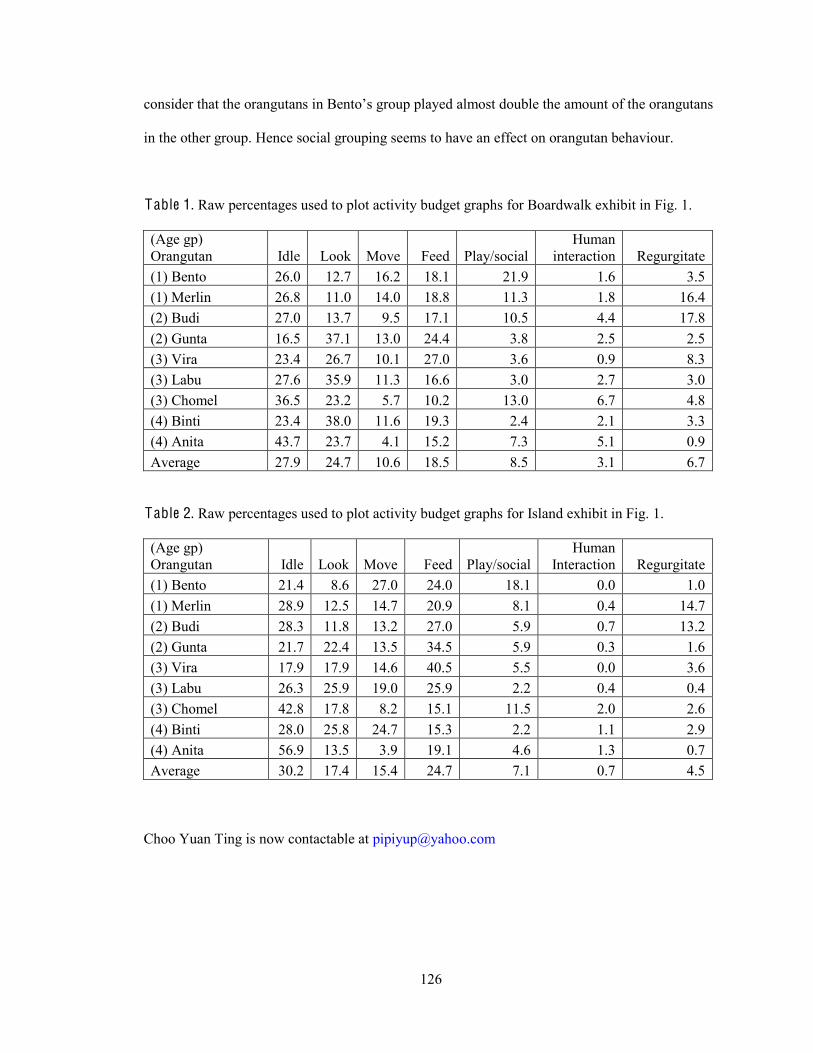
126
in the other group. Hence social grouping seems to have an effect on orangutan behaviour.
Table 1. Raw percentages used to plot activity budget graphs for Boardwalk exhibit in Fig. 1. (Age gp) Orangutan Idle Look Move Feed Play/social
Human interaction Regurgitate
(1) Bento 26.0 12.7 16.2 18.1 21.9 1.6 3.5 (1) Merlin 26.8 11.0 14.0 18.8 11.3 1.8 16.4 (2) Budi 27.0 13.7 9.5 17.1 10.5 4.4 17.8 (2) Gunta 16.5 37.1 13.0 24.4 3.8 2.5 2.5 (3) Vira 23.4 26.7 10.1 27.0 3.6 0.9 8.3 (3) Labu 27.6 35.9 11.3 16.6 3.0 2.7 3.0 (3) Chomel 36.5 23.2 5.7 10.2 13.0 6.7 4.8 (4) Binti 23.4 38.0 11.6 19.3 2.4 2.1 3.3 (4) Anita 43.7 23.7 4.1 15.2 7.3 5.1 0.9 Average 27.9 24.7 10.6 18.5 8.5 3.1 6.7
Table 2. Raw percentages used to plot activity budget graphs for Island exhibit in Fig. 1. (Age gp) Orangutan Idle Look Move Feed Play/social
Human Interaction Regurgitate
(1) Bento 21.4 8.6 27.0 24.0 18.1 0.0 1.0 (1) Merlin 28.9 12.5 14.7 20.9 8.1 0.4 14.7 (2) Budi 28.3 11.8 13.2 27.0 5.9 0.7 13.2 (2) Gunta 21.7 22.4 13.5 34.5 5.9 0.3 1.6 (3) Vira 17.9 17.9 14.6 40.5 5.5 0.0 3.6 (3) Labu 26.3 25.9 19.0 25.9 2.2 0.4 0.4 (3) Chomel 42.8 17.8 8.2 15.1 11.5 2.0 2.6 (4) Binti 28.0 25.8 24.7 15.3 2.2 1.1 2.9 (4) Anita 56.9 13.5 3.9 19.1 4.6 1.3 0.7 Average 30.2 17.4 15.4 24.7 7.1 0.7 4.5
Choo Yuan Ting is now contactable at [email protected]