monoclonal immunoglobulin antibody japanese encephalitis … · infection with japanese...
TRANSCRIPT

Vol. 41, No. 2INFECTION AND IMMUNITY, Aug. 1983, P. 774-7790019-9567/83/080774-06$02.00/0Copyright © 1983, American Society for Microbiology
Monoclonal Immunoglobulin M Antibody to JapaneseEncephalitis Virus that Can React with a Nuclear Antigen in
Mammalian CellsERNEST A. GOULD,'* ALEXANDER C. CHANAS,' ALAN BUCKLEY,' AND CHRISTOPHER S.
CLEGG2
Arbovirus Research Unit, Winches Farm Field Research Station, St. Albans,' and Special PathogensReference Laboratory, Porton Down, Salisbury, Wiltshire,2 United Kingdom
Received 28 December 1982/Accepted 6 May 1983
An immunoglobulin M (IgM) class monoclonal antibody raised against Japaneseencephalitis virus reacted with an epitope on the nonstructural virus protein P74(NV4 in the old nomenclature) of several flaviviruses and also with an antigenpresent in the nuclei of a variety of mammalian cell types. This antigen had acharacteristic granular distribution by immunofluorescence and may correspondto a polypeptide of molecular weight 56,000 seen in nitrocellulose transfers ofsodium dodecyl sulfate-polyacrylamide gels. Cross-reactivity with nuclear antigenwas also occasionally observed in the IgM antibody fraction of mice early afterinfection with Japanese encephalitis virus and also in acute sera from someclinical cases of encephalitis containingvirus.
IgM antibody to Japanese encephalitis
Japanese encephalitis (JE) virus is a memberof the mosquito-borne flaviviruses. It producesepidemics, often of a severe nature, in easternAsia from Japan and the southeastern USSR toIndonesia and India (1). Its replication cycle inmammalian cells is thought to be confined to thecytoplasm, and nuclear fluorescence has neverbeen reported (25). There is, however, evidenceof a nucleus-associated process in its replicativecycle, since the yield of JE virus was severelydepressed in enucleated chicken embryo cells(10). It was suggested that either loss of nuclearmembranes during enucleation or loss of a host-mediated process involving modification of theviral genome could be responsible for the de-pressed yield of virus.On the basis of hemagglutination inhibition
and neutralization tests, cross-reactivity hasbeen detected between JE virus and severalother flaviviruses (5, 17, 22); nevertheless, thesignificance of this antigenic cross-reactivity andits relevance to the pathogenesis of the flavivi-ruses are unknown. We are currently producingmonoclonal antibodies against several of theflaviviruses. Here we describe the properties ofone of the monoclonal antibodies preparedagainst JE virus. It produced bright nuclearimmunofluorescence in both infected and nonin-fected vertebrate cell cultures as well as thecharacteristic cytoplasmic immunofluorescenceseen with flaviviruses. The possible significanceof virus-cell related antigens in the pathogenesisof flavivirus infections is discussed.
MATERIALS AND METHODS
Cells and cell culture. The cells were grown inLeibovitz L15 medium containing 5% fetal calf serum,10% tryptose phosphate broth, penicillin, and strepto-mycin. Vertebrate cells were incubated at 37°C,whereas invertebrate cells were incubated at 28°C.Vero, HEp-2, BHK-21, MDCK, CCL 9.1, and PS cellswere obtained from the Microbiology Department ofthe London School of Hygiene and Tropical Medicine.MRC-5 cells were purchased from Flow Laboratories(Irvine, Scotland). Primary mouse embryo fibroblasts(strain TO or BALB/c) were prepared by trypsiniza-tion of embryos at or near full term. Primary chickenembryo fibroblasts were prepared by trypsinization of10-day-old embryos. The skeletal cells were seeded at106 cells per ml into sterile disposable tissue cultureflasks. Mosquito (AP61) (24) and Xenopus laevis (XL-2) (19) cell cultures were kindly supplied by C. J.Leake (Department of Entomology, London School ofHygiene and Tropical Medicine).Monoclonal antibody. A cloned hybridoma culture
producing the monoclonal antibody designated 62.4awas isolated by standard fusion procedures (9) withthe modifications previously described (2). Immunesplenocytes were obtained from BALB/c mice giventwo intraperitoneal inoculations of live JE virus (Na-kayama strain; 0.1 ml of 20% suckling mouse brainsuspension) followed by one intravenous dose a weeklater. Fusion with the nonsecreting myeloma cell lineP3-X63-Ag8-653 (8) was carried out 2 days later. Theantibody was obtained as mouse ascitic fluid and had atiter of at least 1/10,000 by indirect immunofluores-cence.
Immunodiffusion. To identify the immunoglobulinclass secreted by the cloned culture 62.4a, the super-natant culture medium was concentrated 10-fold with
774
on July 26, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

JE VIRUS AND CELL ANTIGEN CROSS-REACTIVITY
Lyphogel (Gelman Sciences Ltd., United Kingdom).Double immunodiffusion tests were then performedagainst monospecific anti-mouse immunoglobulins.
Immunofluorescence. Cells on glass cover slips wereinfected with JE virus at an estimated input multiplic-ity of 0.01 and incubated at 37°C for 48 h. They werewashed in phosphate-buffered saline (PBS), fixed inchilled acetone, and stored dry at - 20°C. For immuno-fluorescence tests the appropriate dilution of antibodywas incubated on the cells at 37°C for 40 min. Theywere then washed in PBS and treated with eitherfluorescein-conjugated anti-mouse immunoglobulin M[IgM(Fc)], immunoglobulin G (IgG; Nordic Immuno-logical Reagents), or protein A (Pharmacia Fine Chem-icals, Uppsala, Sweden).Western blot analysis. Confluent cultures of Vero
cells were infected with JE virus at a multiplicity of 10to 20. After 30 h of incubation, actinomycin D wasadded to the medium (20) at a final concentration of 1,ug/ml. At 48 h postinfection the cells were washed inwarm methionine-free medium and incubated for 90min in this medium supplemented with 2% dialyzedfetal calf serum and 10 j±Ci of [35S]methionine (1,100Ci/mmol; Amersham International, Amersham, Unit-ed Kingdom) per ml. The cells were then washed twicein cold PBS and lysed in 1% sodium dodecyl sulfate-1% 2-mercaptoethanol-100 mM Tris-hydrochloride(pH 6.8)-15% glycerol-0.01% bromophenol blue (0.5ml per 25-cm2 flask). The lysates were heated at 100°Cfor 2 min and pipetted rapidly to reduce the viscosity.Samples were analyzed on 8 to 15% polyacrylamidegradient gels (11), and the separated proteins wereelectrophoretically transferred to nitrocellulose sheet(23) at 200 to 300 mA for 12 to 16 h in 96 mM glycine-12.5 mM Tris-20% isopropanol. The transferred pro-tein was stained briefly in amido black (23) and de-stained in 10% acetic acid-10% isopropanol, and pro-tein binding sites were blocked by incubation in PBS-2.5% bovine serum albumin-0.05% NaN3 for 60 min atroom temperature with constant agitation. The trans-fers were incubated with a 1/5,000 dilution of 62.4aascitic fluid in PBS-2.5% bovine serum albumin-5%fetal calf serum-1% Triton X-100-0.1% sodium dode-cyl sulfate for 2 h at room temperature with constantagitation. The protein was washed three times for 5min each in PBS-0.05% Triton X-100 and then incu-bated with a 1/1,000 dilution of peroxidase-conjugatedanti-mouse globulin (Miles Laboratories) for 1 h,washed again, and then incubated in 50 mM sodiumacetate (pH 5.0) containing 200 ,ug of aminoethyocar-bazole per ml and 0.015% H202 for 30 min in the dark.
Viruses. The flaviviruses were obtained from eitherJ. Porterfield (Sir William Dunn School of Pathology,University of Oxford) or from C. J. Leake. Each viruswas prepared as a stock culture by intracerebralinoculation of newborn mice (virus diluted 1:100 inPBS). When 20 to 40% of the mice were moribund, a20% suspension of the brains was prepared in PBS andstored at -70°C.
RESULTSDetermination of the antibody subclass secreted
by hybridoma 62.4a. Double-immunodiffusiontests were performed on concentrated hybri-doma culture medium against monospecificmouse immunoglobulins. A single precipitation
line was obtained with antibody 62.4a and anti-mouse IgM(Fc). No lines were obtained witheither anti-IgGl, anti-IgG2a, anti-IgG2b, anti-IgG3, or anti-IgA. The same result was obtainedwith hybridoma culture fluid obtained fromcloned cultures that had been cloned again.
Antigenic specificity of antibody 62.4a for bothvirus and cellular antigens. Indirect immunofluo-rescence experiments with 62.4a antibody onacetone-fixed Vero cells cultured on cover slipsfor 48 h after infection with JE virus (multiplicityof infection, 0.1) showed diffuse stainingthroughout the cytoplasm in approximately 10 to15% of the cells. This corresponded to theknown proportion of cells that contained JEvirus-specific antigen. In addition, a granular orspeckled type of fluorescent staining within thenuclei, but not the nucleoli, of approximately95% of the cells was also observed (Fig. 1).These patterns of fluorescence were observedonly when using anti-IgM(Fc) or whole anti-mouse globulin fluorescent conjugates, andthese results were reproduced after recloningthe hybridoma culture. Control, noninfectedcells showed only the nuclear type of fluores-cence. No cytoplasmic or nuclear fluorescencewas observed when either anti-mouse IgG orfluorescein-conjugated protein A was used asthe second-step reactant. When the 62.4a ascitesfluid was titrated the cytoplasmic antigen wasdetectable at dilutions of 10-4, whereas thenuclear antigen usually had a titer of 1.0 to 2.0log10 units less. Hybridoma culture fluid alsoproduced both patterns of immunofluorescenceand absorption of either 62.4a ascites fluid orhybridoma culture fluid with excessive quanti-ties of acetone-fixed, noninfected Vero cellsremoved their ability to produce nuclear andcytoplasmic immunofluorescence.
Indirect immunofluorescence tests were alsocarried out in Vero cells to determine the anti-genic cross-reactivity of the monoclonal anti-body with a selection of flaviviruses that areantigenically cross-reactive with JE virus. The62.4a antibody was reactive (bright fluorescencein infected cells) with the following flaviviruses:JBE, KUN, USU, WN, ZIKA, TMU, DEN-1,DEN-2, and DEN-4. The 62.4a antibody was notreactive (no fluorescence) with LGT, LI, NEG,POW, CR, DB, MOD, MML, BSQ, KOK, SLE,WSL, YF, BAN, EH, UGS, or DEN-3. (Theabbreviations used are as recommended in theCatalogue ofArthropod-Borne Viruses [1]). Thecharacteristic diffuse virus-specific immunofluo-rescence was observed in the cytoplasm of 9 of26 different flaviviruses tested. In addition, sev-eral different strains of JE and of yellow fever(YF) virus were tested, (data not shown); ineach case the JE strains reacted with 62.4a, butthere was no reaction with the YF virus strains.
VOL. 41, 1983 775
on July 26, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

776 GOULD ET AL.
FIG. 1. Indirect immunofluorescence with JE virus-infected Vero cells and 62.4a monoclonal antibody.Noninfected cells show fluorescent dots only in the nuclei. Infected cells show both nuclear and cytoplasmicfluorescence.
To determine the specificity of the nuclearstaining, a broad spectrum of both vertebrateand invertebrate cell cultures was examined byindirect immunofluorescence with 62.4a mono-clonal antibody and anti-mouse-IgM(Fc) conju-gate. All cells of mammalian origin, which in-cluded human (MRC-5 and HEp-2) monkey(Vero), hamster (BHK-21), canine (MDCK),porcine (PS), and mouse (CCL 9.1, primaryBALB/c, and primary TO) cultures were foundto contain the same nuclear antigen (showedbright fluorescence in the nuclei of the cells).The percentage of positive cells varied from 25to 95%, depending on the density of cells andtheir age since subculturing. Subconfluent cellmonolayers generally showed a lower percent-age of nuclear antigen-positive cells. An in-crease of positive nuclei was observed as celldensity increased. Cultures of primary chickenembryo fibroblasts, mosquito (AP61), and X.laevis (XL-2) cells, on the other hand, did notproduce nuclear fluorescence despite the factthat JE virus can replicate and produce cytoplas-mic fluorescence in these cells.
Determination of molecular specificity. Thespecificity of antibody 62.4a was determined byexamining the ability of nitrocellulose transfersof electrophoretically separated proteins of celllysates ("Western blots") to bind the antibody.Virus-specific proteins on the transfers wereidentified by comparing the proteins labeled in a90-min pulse of [ 5S]methionine in infected and
uninfected cells. The larger virus-specific pro-teins were clearly visible against a backgroundof continuing host protein synthesis (Fig. 2,tracks a and b). Antibody binding was testedunder two sets of conditions: (i) with low levelsof cell protein loaded on the gel (equivalent toabout 2.5 x 104 cells), and high dilutions ofmonoclonal antibody (1/5,000 dilution of asciticfluid), or (ii) with higher levels of cell protein (5X 104 cells) and lower dilution of antibody(1/500). The only virus-specific protein capableof binding antibody under either set of condi-tions was the nonstructural protein P74 (previ-ously known as NV4 [20]) (Fig. 2, tracks c ande). This protein is thus unequivocally identifiedas the virus-specific target for antibody 62.4a.The antibody was also capable of binding to twoclasses of cellular proteins which were found ininfected and uninfected cells. Under conditionsof high antigen and antibody concentration, sev-eral proteins appeared to bind antibody (Fig. 2,tracks c and d). However, with lower antigenand antibody concentrations, only a single cellu-lar protein with an apparent molecular weight of56,000 was able to bind observable quantities ofantibody (Fig. 2, tracks e and f), indicating thatthis protein has a higher affinity for the antibodythan the other cellular proteins. It was unusualin that it penetrated the nitrocellulose matrix,rather than binding to the surface as most pro-teins do, and did not bind monoclonal antibody30.22, which is specific for Sindbis virus nucleo-
INFECT. IMMUN.
on July 26, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

JE VIRUS AND CELL ANTIGEN CROSS-REACTIVITY 777
P93-
P74- _
GP53(E) _'
a b c d e fFIG. 2. Molecular specificity of antibody 62.4a de-
termined by Western blot analysis. Infected (tracks a,
c, and e) and uninfected (tracks b, d, and f) cells werelabeled with [35S]methionine; the proteins were ana-
lyzed on an 8 to 15% polyacrylamide gradient gel andtransferred to nitrocellulose. Radioactive proteinswere revealed by autoradiography (tracks a and b).Antibody-binding proteins were located by incubationwith a 1/500 dilution (tracks c and d) or a 1/5,000dilution (tracks e and f) of ascitic fluid containingantibody 62.4a. Protein loaded on the gel corre-
sponded to about 2.5 x 104 cells (tracks a, b, e, and f)or 5 x 104 cells (tracks c and d).
capsid protein (2) and also active on Westernblots. These experiments suggest that the char-acteristic nuclear fluorescence seen with anti-body 62.4a is probably associated with the56,000-molecular-weight protein, but the possi-bility that the other cellular proteins with lowerbinding affinity may be involved cannot be ruledout.
Antibody production in mice infected with JEvirus. Conventional JE virus hyperimmune anti-sera produce cytoplasmic fluorescence, but donot produce nonspecific nuclear staining ofmammalian cells (data not shown). We have,however, qualitatively reexamined the chrono-logical aspects of the antibody response in JEvirus-infected mice. Adult BALB/c mice inocu-lated intraperitoneally with 103 PFU of JE viruswere bled daily, and their sera were examinedby immunofluorescence for IgM antibodies toJE virus. With JE virus-infected Vero cells,cytoplasmic fluorescence was first detected withanti-IgM antibody on the day 3 postinfection.The sera from 3 of 15 mice produced bothnuclear and virus-specific cytoplasmic fluores-cence 3 to 5 days postinfection (Fig. 3a). Serafrom uninfected mice gave no fluorescent stain-
ing. JE virus-specific IgG antibody producingonly cytoplasmic fluorescence was detectablefrom day 4 postinfection (Fig. 3b). Finally, 18 of26 human sera from cases of suspected JEencephalitis in Nepal produced IgM-specific nu-clear fluorescence similar to that describedabove. This was shown not to be due to rheuma-toid factor by using a Rheum-Wellcotest kit(Wellcome Laboratories, Beckenham, En-gland).
DISCUSSIONThe monoclonal antibody 62.4a described
here gives a characteristic diffuse cytoplasmicfluorescence in cells infected with JE virus andwith a number of other flaviviruses. In addition,the antibody gives rise to speckled nuclear fluo-rescence in uninfected or infected cells of mam-malian origin. These two activities were notseparable either by repeated cloning of the hy-bridoma line or by absorption experiments, sug-gesting that both reside on the same immuno-globulin, and thus that the virus-specifiedprotein and the protein found in mammaliannuclei possess identical or closely related epi-topes.The virus-specified protein carrying the epi-
tope is definitively identified by Western blotanalysis as P74, a protein found in abundance inJE virus-infected cells (3), but which does notform part of the extracellular virus structure. Asfar as we are aware this is the first monoclonalantibody directed at a nonstructural protein of atogavirus to be found. Identification of the hostprotein recognized by antibody 62.4a in nitrocel-lulose transfers of sodium dodecyl sulfate-poly-acrylamide gels cannot be so positive, since thereaction with the candidate 56,000-molecular-weight protein is much weaker, and other cellu-lar proteins also bind the antibody, althoughwith lower affinity. The relative weakness of thereaction with the 56,000-molecular-weight pro-tein reflects the difference in apparent antibodytiter for viral and nuclear antigen seen by immu-nofluorescence and may be due to relatively lowlevels of the host protein as compared with viralP74 in the cells or to a lower affinity of the hostprotein for the antibody due to differences in theepitope structure. The ability of the host proteinto penetrate the nitrocellulose membrane alsosuggests that it does not bind efficiently and maybe partially lost on transfer.The existence of monoclonal antibodies that
react with more than one antigen is expected ontheoretical grounds (4, 13), and host cell proteinssharing a common determinant with a virus-coded protein have indeed been discovered bythe use of these reagents (4, 12). Whether or notproteins carrying such common determinantsshare some common function, as suggested by
VOL. 41, 1983
on July 26, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
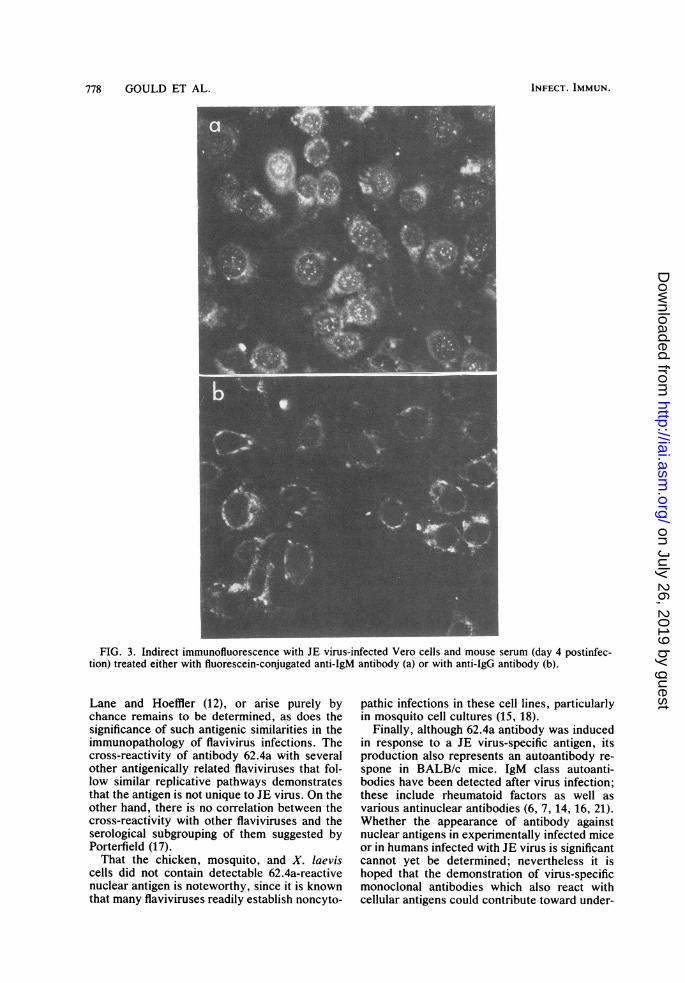
778 GOULD ET AL.
FIG. 3. Indirect immunofluorescence with JE virus-infected Vero cells and mouse serum (day 4 postinfec-tion) treated either with fluorescein-conjugated anti-IgM antibody (a) or with anti-IgG antibody (b).
Lane and Hoeffler (12), or arise purely bychance remains to be determined, as does thesignificance of such antigenic similarities in theimmunopathology of flavivirus infections. Thecross-reactivity of antibody 62.4a with severalother antigenically related flaviviruses that fol-low similar replicative pathways demonstratesthat the antigen is not unique to JE virus. On theother hand, there is no correlation between thecross-reactivity with other flaviviruses and theserological subgrouping of them suggested byPorterfield (17).That the chicken, mosquito, and X. laevis
cells did not contain detectable 62.4a-reactivenuclear antigen is noteworthy, since it is knownthat many flaviviruses readily establish noncyto-
pathic infections in these cell lines, particularlyin mosquito cell cultures (15, 18).
Finally, although 62.4a antibody was inducedin response to a JE virus-specific antigen, itsproduction also represents an autoantibody re-spone in BALB/c mice. IgM class autoanti-bodies have been detected after virus infection;these include rheumatoid factors as well asvarious antinuclear antibodies (6, 7, 14, 16, 21).Whether the appearance of antibody againstnuclear antigens in experimentally infected miceor in humans infected with JE virus is significantcannot yet be determined; nevertheless it ishoped that the demonstration of virus-specificmonoclonal antibodies which also react withcellular antigens could contribute toward under-
INFECT. IMMUN.
on July 26, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

JE VIRUS AND CELL ANTIGEN CROSS-REACTIVITY 779
standing how some self-limiting autoantibodyresponses are triggered and their possible role inviral pathogenesis.
ACKNOWLEDGMENTSPart of this work was supported by the Wellcome Trust.We are indebted to D. Newman for her excellent assistance
and for typing the manuscript.
LITERATURE CITED
1. Berge, T. 0. 1975. International catalogue of arboviruses,2nd ed., U.S. Department of Health, Education andWelfare publication no. (CDC) 75-8301. Public HealthService, Washington, D.C.
2. Chanas, A. C., E. A. Gould, J. C. S. Clegg, and M. G. R.Varma. 1982. Monoclonal antibodies to Sindbis virusglycoprotein El can neutralize, enhance infectivity andindependently inhibit haemagglutination or haemolysis. J.Gen. Virol. 58:37-46.
3. Clegg, J. C. S. 1982. Glycoprotein detection in nitrocellu-lose transfers of electrophoretically separated proteinmixtures using concanavalin A and peroxidase: applica-tion to arenavirus and flavivirus proteins. Anal. Biochem.127:389-394.
4. Crawford, L., K. Leppard, D. Lane, and E. J. Harlow.1982. Cellular proteins reactive with monoclonal antibod-ies directed against simian virus 40 T-antigen. J. Virol.42:612-620.
5. de Madrid, A. T., and J. S. Porterfield. 1974. The flavivi-ruses (group B arboviruses): a cross-neutralization study.J. Gen. Virol. 23:91-96.
6. Dresner, E., and P. Trombly. 1959. The latex-fixationreaction in nonrheumatic diseases. N. Engl. J. Med.261:981-988.
7. Johnson, R. E., and A. P. Hall. 1958. Rubella arthritis:Report of cases studied by latex tests. N. Engl. J. Med.258:743-745.
8. Kearney, J. F., A. Radbruch, B. Liesegand, and K. Ra-jewsky. 1979. A new mouse myeloma cell line that has lostimmunoglobulin expression but permits the constructionof antibody-secreting hybrid cell-lines. J. Immunol.123:1548-1550.
9. Kohler, G., and C. Milstein. 1975. Continuous cultures offused cells secreting antibody of predefined specificity.Nature (London) 256:495-497.
10. Kos, K. A., B. A. Osborne, and R. A. Godsby. 1975.Inhibition of group B arbovirus antigen production andreplication in cells enucleated with cytochalasin B. J.Virol. 15:913-917.
11. Laemmli, U. K. 1970. Cleavage of structural proteinsduring the assembly of the head of bacteriophage T4.Nature (London) 227:680-685.
12. Lane, D., and W. K. Hoeffier. 1980. SV40 large T sharesan antigenic determinant with a cellular protein of molecu-lar weight 68,000. Nature (London) 288:167-170.
13. Lane, D., and H. Koprowski. 1982. Molecular recognitionand the future of monoclonal antibodies. Nature (London)296:200-202.
14. Langenhuysen, M. M. A. C. 1971. Antibodies against y-globulin after blood transfusion and cytomegalovirus-infection. Clin. Exp. Immunol. 9:393-398.
15. Leake, C. J., M. G. R. Varma, and M. Pudney. 1977.Cytopathic effect and plaque formation by arboviruses ina continuous cell line (XTC-2) from the toad Xenopuslaevis. J. Gen. Virol. 35:335-339.
16. Markenson, J. A., C. A. Daniels, A. L. Notkins, J. H.Hoofnagle, J. Gerety, and L. F. Barker. 1975. The interac-tion of rheumatoid factor with hepatitis B surface antigen-antibody complexes. Clin. Exp. Immunol. 19:209-217.
17. Porterfield, J. S. 1980. Antigenic characteristics and clas-sification of togaviridae, p. 13-46. In R. W. Schlesinger(ed.), The togaviruses. Academic Press, Inc., New York.
18. Pudney, M., C. J. Leake, and S. M. Buckley. 1982.Replication of arboviruses in arthropod in vitro systems:an overview, p. 159-194. In K. Maramarosch and J.Mitsuhashi (ed.), Invertebrate cell culture applications.Academic Press, Inc., New York.
19. Pudney, M., M. G. R. Varma, and C. J. Leake. 1973.Establishment of a cell line from the South African clawedtoad Xenopus laevis. Experientia 29:466-467.
20. Shapiro, D., K. Kos, and P. K. Russell. 1973. Proteinsynthesis in Japanese encephalitis virus-infected cells.Virology 56:95-109.
21. Svec, K. H., and J. H. Dingle. 1%5. The occurrence ofrheumatoid factor in association with antibody responseto influenza A2 (Asian) virus. Arthritis Rheum. 8:524-529.
22. Theiler, M., and W. G. Downs. 1973. The arthropod-borneviruses of vertebrates. Yale University Press, New Ha-ven.
23. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electro-phoretic transfer of proteins from polyacrylamide gels tonitrocellulose sheets: procedure and some applications.Proc. Natl. Acad. Sci. U.S.A. 76:4350-4354.
24. Varma, M. G. R., M. Pudney, C. J. Leake, and P. H.Peralta. 1976. Isolations in a mosquito (Aedes pseudoscu-tellaris) cell line (Mos 61) of yellow fever virus strainsfrom original field material. Intervirology 6:50-56.
25. Westaway, E. G. 1980. Replication of flaviviruses, p. 531-577. In R. W. Schlesinger (ed.), The togaviruses. Aca-demic Press, Inc., New York.
VOL. 41, 1983
on July 26, 2019 by guesthttp://iai.asm
.org/D
ownloaded from