mössbauer and epr study of a cytochrome b modeldedicated to professor f. dörr on the occasion of...
TRANSCRIPT

Mossbauer and EPR study of a cytochrome b modely
Olga Iakovleva,a Michael Reiner,a Harald Rau,b Wolfgang Haehnelb and Fritz Parak*a
a Physik Department E17, Technische Universitat Munchen, 85747, Garching, Germanyb Institut fur BiologieII=Biochemie, Albert-Ludwigs-Universitat Freiburg, Schanzlestr. 1,79104, Freiburg, Germany
Receivved 19th October 2001, Accepted 19th Novvember 2001First published as an Advvance Article on the web 17th January 2002
Mossbauer and EPR measurements were performed with MOP1, a novel cytochrome b model compound withtwo heme binding sites. The measurements show for the first site well-resolved normal B-hemichrome EPRsignals with characteristic g-values of 2.95, 2.27 and 1.50 which are a feature of the parallel planes orientation of
axial histidines. The second site shows a poorly resolved EPR signal with a broad peak at g� 3.5, although bothsites are occupied by the same amount of heme as found in the Mossbauer spectra. This site is correlated withthe axial symmetry of the g-tensor and with the perpendicular alignment of the histidine planes. The Mossbauerspectra are simulated with due account for the spin–spin relaxation taking place within one molecular unit. It
shows that the Fe(III) ion in the second site is in a previously not characterised spin-mixed state and not in apure low-spin state, such as a HALS state. The redox potential of this site is about 65 mV more positive thanthat of the first site.
1 Introduction
Many enzymes have a rather complex structure assembledfrom protein domains with different functions and non-proteincofactors. Understanding of the catalytic mechanism is oftenobscured by this complexity and by the lack of structuraldetails. The de novo design and the synthesis of the specificparts of an enzyme allows one to construct chemically simplepeptides and to establish the minimal requirements for thefunction of their highly complex natural counterparts. Hemeproteins are a large group of proteins with a wide diversity offunctions controlled by the coordination of the heme iron andits nearest environment. In this paper a model compound ofthe heme binding region of cytochrome b is investigated. Itmodels the core of the mitochondrial cytochrome bc1-complexand of the homologous photosynthetic cytochrome bf-com-plex. The template assembled anti-parallel four-helix bundlecarries two bis-histidine ligated heme groups in the hydro-phobic interior.1,2 The final goal of such investigations is tolearn how nature tunes the redox potential of the heme iron indifferent types of cytochrome b.The electronic properties and the resulting redox potentials
of heme proteins are modulated by the nature of the axial hemeligands. Cytochromes b, generally identified with bis(imida-zole) coordination of Fe-protoporphyrin IX, differ in theorientation of the axial ligands and can be divided into twogroups from a spectroscopic point of view.3 The first groupconsists of b2 and b5 cytochromes which have redox potentialsin the range from 0 to +30 mV, and show well characterisedEPR spectra of class ‘‘B ’’ heme proteins with resonance atg1� 2.9, g2� 2.2 and g3� 1.5. These signals are typical for apure low-spin ferric ion. In this case the parallel orientation ofthe axial imidazole planes is proved to take place. The secondgroup includes a variety of the membrane-bound cytochromesb, as in mitochondria and chloroplasts, with a wider range ofredox potentials than in the b2 and b5 case. This group ischaracterised by a peculiar EPR signal at g� 3.5 and a broadtail extending up to at least 1 T in the X-band. This so-called
‘‘ strong gmax ’’ EPR signal (see ref. 3 and references therein) issupposed to originate from an orientation of the imidazoleplanes perpendicular to each other. Despite the fact that thestrong gmax-value is out of the range typical for the conven-tional low-spin state, the complexes showing such a signal werestill classified as low-spin species and denoted as HALS (highlyanisotropic low spin).4 It was assumed that the HALS state wasformed by the axial crystal field distortion, intermediate inmagnitude between that for high spin and for low spin, and thatthe cubic field strength in the HALS is significantly weaker thanin the low-spin complexes. This implies that the excited dz2orbital is closer in energy to the t2g orbitals compared to the LSstate where the influence of the eg excited state is negligible.Such an electronic state could be close to the intermediate spinstate (dxy)
2 (dxz , dyz)2(dz2)
1.4 In this contribution we show thatthe discussed EPR signal can be understood as a result of thespin admixture between the intermediate- and the low-spinstates. The EPR spectrum gives no information on the gx , gy-components, so that the Mossbauer measurement is essentialfor the complete identification of this state. The simulation ofthe Mossbauer low-temperature spectra has taken into accountthe spin–spin relaxation between the non-equivalent heme ironswithin one molecular unit.
2 Experimental
The first step was to synthesise the modular protein on thebasis of the template assembling of the synthetic proteins(TASPs).5 Two types of a-helical peptides were synthesized,reproducing the structure of the native binding region of theheme groups in cytochrome b. The helices were designed asamphiphilic ones with the hydrophilic amino acid residues onone side and the hydrophobic residues on the opposite side.One helix carries two histidines on the hydrophobic side tobind the heme groups. Four helices, two of each kind, werefixed at diagonal positions to a cyclic decapeptide template.This method provides a well defined structural arrangement ofthe four helical segments in antiparallel orientation. In thesecond step, the two heme groups were incorporated into thisy Dedicated to Professor F. Dorr on the occasion of his 80th birthday.
DOI: 10.1039/b109570f Phys. Chem. Chem. Phys., 2002, 4, 655–660 655
This journal is # The Owner Societies 2002
Publ
ishe
d on
17
Janu
ary
2002
. Dow
nloa
ded
by U
nive
rsid
ad d
e O
vied
o on
27/
10/2
014
10:3
6:35
. View Article Online / Journal Homepage / Table of Contents for this issue

modular assembled protein (MOP).2 Fig. 1 shows the assumedstructure of MOP.The X-band CW EPR experiment was run on a Brucker
ESP380 set-up equipped with a continuous-flow helium cryo-stat (Oxford), at the microwave frequency of 9.47 MHz. Thespectrum was recorded in a conventional way as a first deri-vative dw00=dB of the imaginary part of the magnetic suscept-ibility w00.The Mossbauer measurements were performed in order to
characterise two heme iron centres in the described artificialprotein unit. Incorporation of 57Fe3+ into protoporphyrin IXfollowed the procedure of Inubushi and Yonetani.6 0.31 mmoliron enriched to more than 95% 57Fe was dissolved in 3 mlHClconc=HNO3 conc (3=1 v=v) and dried in vacuum. The resi-due was dissolved in 5 ml AcOH and added to a solution of0.41 mmol photoporphyrin IX–Na2 in 100 ml AcOH=2 mlpyridine under Ar at 90 �C. After stirring for 30 min at 90 �C ina flask with a reflux condenser, 5 ml saturated NaCl(aq) wasadded. After cooling to room temperature, 500 ml ethyl acet-ate were added and washed with 10% (w=v) HCl solution untilthe washing became colourless. The ethyl acetate layer waswashed three times with 25% (w=v) NaCl, twice with water,dried with Na2SO4 and evaporated to dryness. The yield of thecrude product was 200 mg. Further purification was achievedby column chromatography with silica gel and with a mobileof 10=5=2 (v=v=v) pyridine, water, CHCl3 . The collectedfractions of 50 ml were mixed with 250 ml of ethyl acetate. Theorganic phase was washed three times with 5% HCl, threetimes with 25% (w=v) NaCl(aq), and twice with water. Thevolume was reduced to 20 ml by a vacuum rotator evaporator,the precipitate being collected by centrifugation. The volumewas reduced again and the precipitate collected. The combinedprecipitates were washed once with 30% (v=v) AcOH, twicewith 25% (w=v) NaCl(aq), and three times with water. Dryingunder vacuum yielded 0.195 mmol. Electrospray mass spec-trometry showed the correct mass of 617.4 Da, and not themass of 616.4 Da expected for natural Fe-protoporphyrin IX.Bis-heme MOP1 was synthesized as described in ref. 7 but with57Fe-protoporphyrin IX chloride freeze dried and dissolved in0.1 M sodium phosphate buffer, pH 7, at a concentration of1.9 mM. The sample contained 450 ml of a 1.9 mM aqueoussolution of MOP1-bis-57Fe-heme in the oxidized form. Thesample holder was of 0.5 cm thickness and 1.0 cm diameter. A57CoRh source was mounted on a drive to vary the energy ofthe 14.4 keV Mossbauer quanta. The spectra were taken with aHalder MCA3=1 multichannel analyser. While the source wasat room temperature, the sample was cooled in a bath cryostat.Measurements were performed at 167 K, at 78 K in liquidnitrogen and at 2.4 K and 5 K (not shown) in liquid helium. Aweak magnetic field of about 20 mT was applied perpendicularto the g-beam. The redox potentials of the two hemes in MOP1
were determined elsewhere2,7 and found to be equal to �106mV and �170 mV respectively. In addition, it was recentlyshown that the redox potentials vary significantly due to thedifferences in the protein environment.8
3 Theoretical considerations
The structural scheme of MOP1 with two heme groups is givenin Fig. 1. There are two heme groups, bL and bH , wheresubscripts L and H refer to the relatively low and high redoxpotentials. Iron 1 is ligated by two histidines with parallelimidazole rings and is in the pure low-spin (LS) state with spinS¼ 1=2.9 We ascribe the low potential (LP) form to iron 1 andplace it in the bL heme for the following reason: recently weinvestigated the Mossbauer spectra of the iron centers inD1=D2=Cyt b559 which is characterized by the LP form.
10 TheMossbauer spectrum of the low spin component of MOP1turned out to be practically identical to the spectrum ofD1=D2=Cyt b559 . Iron 2 in the bH heme is ligated by twohistidines with nearly perpendicular imidazole rings, and itsspin state is discussed below.In the further analysis we follow the low-spin state
description discussed in detail by Oosterhuis,11 the spin-mixedstate analysis of Harris12,13 and the method developed for thedescription of Mossbauer spectra in a weak external magneticfield by Afanas’ev et al.14
3.1 Electronic model
In a strong crystal field of axial symmetry the ferric ion com-plexes can exist in the low spin (LS) state with S¼ 1=2, thehigh-spin (HS) state with S¼ 5=2, the intermediate-spin (IS)state with S¼ 3=2, and substantially spin-mixed statesdepending on the crystal field and spin–orbit coupling para-meters. In a strong crystal field of cubic symmetry (Dq> 36 000cm�1) only LS and IS electronic states are possible, the formerwith strong and the latter with weaker tetragonal distortion. Inother words, the spin–orbit interaction can give rise to a sig-nificantly large spin-mixed region between the pure LS and thepure IS state if the ratio of axial=in-plane interaction is rela-tively small. An extended calculation of the eigenfunctions andeigenvalues, as well as of the localized properties of thesestates, has been done elsewhere.12,13 The 12 multicomponentbasis states obtained are the Kramers doublets, and their g-values (unmixed) change markedly over a wide range due tothe spin–orbit coupling. We should keep in mind that for apure low-spin doublet three anisotropic g-values are observedbetween 2.9 and 1.5, while only two values, gz� 2 and g?� 4are observed for a pure IS state. In the spin-mixed region the g-values of the first excited state vary over a wider range com-pared to the ground state. In addition, the larger the g?-value,the larger is the resonance transition probability. Therefore, ifthe g?-value of the excited state is substantially larger thanthat of the ground state, it is possible to see both the groundand excited doublet transitions though the excited state can beseveral hundred cm�1 above the ground state.We begin with the description of the pure low-spin ferric
state developed by Griffith9 to explain the observed g-values.In a strong octahedral field the five d-electrons of the ferriciron occupy the t2g manifold giving a state which can beregarded as a t2g hole with spin S¼ 1=2. In the rhombic sym-metry the system is described by the spin-orbit coupling, ll�s,between the orbital angular momentum and the spin of thehole, and by two ligand field parameters, D and V. Theseparameters account for the axial and rhombic distortionsrespectively, and are conventionally measured in the units of l.Spin–orbit coupling mixes the t2g states and the resultingeigenstates are three Kramers doublets with the eigenenergiesgiven in Table 1 in ref. 9. Since the magnitude of the l is about
Fig. 1 Schematic drawing of the backbone structure of the de novoprotein MOP1 bis-heme complex. The modular protein is synthesizedon the basis of TASPs, see Experimental section. The backbone isshown as a ribbon. The His residues which bind the two heme groupsand the Phe residue in the center of helix H1 are shown as stickstructures. Recently generated atomic co-ordinates describe thisstructure.8
656 Phys. Chem. Chem. Phys., 2002, 4, 655–660
Publ
ishe
d on
17
Janu
ary
2002
. Dow
nloa
ded
by U
nive
rsid
ad d
e O
vied
o on
27/
10/2
014
10:3
6:35
. View Article Online

150–600 cm�1, only the ground doublet is populated at lowtemperatures and only the ground doublet will be consideredbelow:11
cþ ¼ ajxyiaþ bjyzibþ icjxzibc� ¼ ajxyib� bjyziaþ icjxzia:
ð1Þ
Here a and b are the usual notations for the spin states withspin projections ms¼ 1=2 and �1=2 respectively. The occupa-tion numbers of the hole a, b, c are defined unambiguously bythe crystal field parameters D and V. In turn, they uniquelydetermine the g-tensor, the magnetic hyperfine tensor and theelectric field gradient (EFG) tensor of the ground state.Therefore, once D and V are specified, the electronic low-spinstate is characterized.We proceed with the description of an IS–LS spin-mixed
state as a ground state. At 2.4 K the mixed state contains the4A2 and the
2E states:
w� ¼ x3j4A2;�3=2i þ x6j2E�1;�1=2i þ x9j4A2;�1=2iþ x12j2E�1;�1=2i
ð2Þ
The coefficients xi , are defined by the relation between thecrystal field parameters.12
3.2 Spin Hamiltonian
It is convenient to substitute the real Hamiltonian operatingon the real states (1) or (2) with an effective spin Hamiltonianacting on the eigenstates j±1=2i of the effective spin S ¼ 1=2.Then the spin Hamiltonian determining the Mossbauer spec-trum is given in a common way.
~H ¼ beBextgSþ IASþ eQqzz4
I2z �IðIþ 1Þ
3þ Z3ðI2x � I2yÞ
� �ð3Þ
The Hamiltonian contains the g-tensor g, the tensor of thehyperfine magnetic interaction A, and the tensor of the electricfield gradient q, with the principal components qxx , qyy , qzz ,Z ¼ qxx�qyy
qzzis the asymmetry parameter.I is the nuclear spin
with the components Ix , Iy and Iz . The Bohr magneton isdenoted by be , and the nuclear quadrupole moment by Q. e isthe elementary charge, and Bext is an externally applied mag-netic field. It is common to denote the electric quadrupoleinteraction by DEQ ¼ 1
2 eQqzzffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffið1þ Z2=3Þ
p. In the case of the
LS doublet, we define the g-, A- and q-tensors via the valuesof the crystal field parameters D and V following the paper ofOosterhuis,11 Appendix A. In order to calculate the magnetichyperfine interaction, an overall scaling factor P and the Fermicontact factor k are taken at their typical LS ferric state valuesof �4.19 mm s�1 (for the excited state) and 0.35 respectively.15
In the case of the spin-mixed doublet, the values of the g-tensor are calculated as in the paper of Harris,13 and thecomponents of the A-tensor as well as the parameters of thenuclear quadrupole interaction, DEQ and Z, are obtained fromthe simulation of the Mossbauer spectra. Last but not least, inour analysis we assume that the g-tensor and A-tensor of eachKramers doublet share a common principal axis system,determined by the geometry around the iron center. Thecoordinate system of the corresponding q-tensor can be rota-ted vs. this system by Euler angles a1 , a2 and a3 .The application of the weak external magnetic field (�20
mT) results in the splitting of a Kramers doublet which ismuch larger than the hyperfine interaction. The analysis of thehyperfine structure in such fields was given in ref. 14, and itwas successfully applied to the Mossbauer study of variousbiological objects afterwards(see, e.g., ref. 16). In this case thehyperfine interaction is treated as a perturbation to the Zee-man splitting of a Kramers doublet. Then the effectiveHamiltonian of the hyperfine interaction for each Zeemansublevel is obtained by averaging Hhf ¼ IAS over the wavefunctions of the corresponding sublevels:14
H �hf ¼ � 1
2G�1ðGxAxIx þ GyAyIy þ GzAzIzÞ ð4Þ
with
Gz ¼ gz cos W; Gx ¼ gx sin W cosj; Gy ¼ gy sinW sinj
G� ¼ Gx � iGy; G ¼ ðG2x þ G2
y þ G2zÞ1=2 ð5Þ
The angles W, f define the orientation of Bext relative to themolecular coordinate system, and the tensor componentsAi(i¼ x, y, z) of each doublet are given in the units of theoverall scaling factor P.Since the distance between the two heme groups in a MOP1
molecule is 46+A (see, e.g. Fig. 2 in ref. 8), a magnetic inter-
action (exchange or dipole–dipole) may occur between thespins of two non-equivalent heme irons. We treat this influenceas the relaxation process which involves mutual spin flips atneighboring irons. In the case when the spin species partici-pating in the relaxation process are not identical, the doubleflip-flops of their spins give rise to the so-called cross
Fig. 2 EPR spectrum of MOP1-bis-heme at 4.2 K. The signal atg¼ 6.0 comes from the HS Fe3+ impurity, the signal at g¼ 4.3 isascribed to the insignificant amount of the rhombic HS Fe3+ ion. Bothimpurities are not resolved in the Mossbauer spectra. The signals at2.95, 2.27 and 1.5 indicate Fe LS.
Table 1 Crystal field and hyperfine parameters of MOP1-bis-hemeobtained from the simulation of the 2.4 K Mossbauer spectrum withthe assumption that the electronic ground state of the bL heme groupiron is a LS state, and that of the bH heme group iron is the HALSstate
Low-spin, iron 1 HALS, iron 2
D 3.25 1.97V 1.87 0.41gx , gy , gz 1.50, 2.25, 2.93 0.08, 1.53, 3.52Ax , Ay , Az=mm s�1a � 2.74, 1.19, 3.38 2.36, 2.25, 6.20qxx , qyy , qzz=mm s�1 � 1.12, 0.51, 0.61 � 0.78, 0.05, 0.73DEQ=mm s�1 2.25; 1.74;Z¼ (qxx� qyy)=qzz � 2.68 � 1.13d=mm s�1 0.29 0.21g=MHz 3.53 3.53a1 , a2 , a3=� 120, 0, 0 90, 30, 90
a Given for the excited state.
Phys. Chem. Chem. Phys., 2002, 4, 655–660 657
Publ
ishe
d on
17
Janu
ary
2002
. Dow
nloa
ded
by U
nive
rsid
ad d
e O
vied
o on
27/
10/2
014
10:3
6:35
. View Article Online

relaxation. The spin flip of one species is accompanied by thesimultaneous spin flop of the other one in the reverse directionso that the energy is conserved or nearly conserved.17 Oursystem consists of two Kramers doublets with different g-tensors. For the transitions between the Zeeman sublevels ofeach particular doublet, the relaxation rate g enters multipliedby a function of the values Gð1Þ
±z,Gð2Þ±z from eqn. (5).
However, for the polycrystalline sample the influence of theanisotropy in the g-values introduced via this function is sig-nificantly smoothed by the averaging over the possible orien-tations of the molecular unit. T hen the matrix representationof the hyperfine interaction and the relaxation transitionsin the basis of the Zeeman sublevels of the low-spin doubletand of the Zeeman sublevels of the spin-mixed doublet can begiven as
where the superoperator ±L hf( i) describes the magnetic
hyperfine interaction of doublet i with the positive(negative)projection of the effective hyperfine field (4), L Q
( i) describes thequadrupole nuclear interaction, and e is a unit matrix in theLiouville space.
4 Results and discussion
4.1 EPR
Fig.2 shows the first-derivative EPR spectrum of MOP1 takenat 4.2 K. The typical low-spin signals with g-values at 2.95,2.27 are well resolved and the signal at 1.5 is very broad asusual. As was mentioned, these signals are characteristic forthe bis-histidine ligated heme-complexes LS with nearly par-allel orientation of the two histidine planes. In addition, thereis a shoulder at g� 3.5. A similar spectrum was reported inref.18 where the simulation of the LS contribution was donewithout accounting for this shoulder. The absence of theENDOR signal of the HALS species discussed there comesfrom choice of the field position, namely, at gy¼ 2.28 HALSgives no ENDOR response. The most striking fact about thesignal at g� 3.5 in the EPR spectra of MOP1 is that theresonance is extremely broadened and only tentatively identi-fiable compared to its observation in other compounds, see,e.g., Fig. 1 in ref. 3 and 4. The preparation of the sample, onthe other hand, prohibits the presence of both heme irons inthe same subunit with unequal partial contributions, with thisinequality being responsible for the low area of HALS signal inthe EPR spectrum. The plausible change in the ratio HALS toLS heme form due to the presence of the HS Fe3+ signal atg¼ 6.0 and of the HS Fe3+ rhombic signal at g¼ 4.3 can besafely ruled out of consideration due to the much greatertransition probability for the HS impurities than for the LSspecies. It means that the amount of these impurities is almostnegligible.3,18 What is more, the equal account for HALS ironand for the LS iron contribution in the Mossbauer spectrumdoes not result in a satisfactory picture, see below. So far, theobservation of the EPR signal at g¼ 3.5 was correlated withthe presence of a LS iron in nearly axial symmetry with per-pendicular alignment of the ligand planes. All these factsmotivated us to investigate a possible ground state of thesecond species as a spin-mixed state between a ground inter-mediate spin state which has an axial symmetry and an excitedlow spin state.
4.2 Mossbauer spectra
Fig. 3 shows the spectrum measured at 2.4 K in the transverseapplied magnetic field of 20 mT fitted with different assump-tions. In accordance with the chemical preparation, it wasassumed in further analysis that both irons contribute equallyto the area of the spectrum. Iron 1 is supposed to have the pureLS g-values at 2.93, 2,25 and 1.50 (calculated) and parallelorientation of the histidine planes. This assumption is presevedin all further considerations of the present work. For iron 2 weassumed in a first approach that its ground state is also thelow-spin, namely the HALS state. In order to obtain thecrystal field parameters D and V of the both LS and HALSstates, we varied these parameters in our program till weobtained the g-values that most closely match the EPR data.
The program is based on the crystal field Hamiltonian andexpressions for g-values given in ref. 9 and 19. The optimizedcrystal field parameters were kept constant in further simula-tions. It is worthy to note that in this way the components ofthe A- and q-tensors are also defined unambiguously.11 Theresult of this simulation is given in Fig. 3(a). The crystal fieldparameters, the isomer shift, d, and the quadrupole splitting,DEQ , of iron 1 and iron 2 are given in Table 1. The line widthGexp was equal to 0.27 mm s�1. The spin–spin relaxation wastaken into account. We allowed the rotation of both EFGtensors with respect to the molecular coordinate system in
Fig. 3 Mossbauer spectrum of MOP1-bis-heme measured at 2.4 Kwith an externally applied magnetic field of 20 mT applied perpendi-cular to the g–beam. In the following simulations relaxation is takeninto account. (a)The simulation assumes that iron 1 ion is in the LSstate (dot-dashed line), and iron 2 ion is in the HALS state (dottedline). The thick solid line represents the contribution of iron 1 and iron2. (b)This simulation assumes that iron 1 ion is in the LS state (dot-dashed line), and iron 2 ion is in the spin-mixed state (dotted line) andgives a much better explanation of the experimental data. The solidline gives the sum of both contributions. For comparison, the thinsolid line is calculated in the absence of relaxation.
L ¼
Lð1Þhf þ L
ð1ÞQ � ge ge
ge �Lð1Þhf þ L
ð1ÞQ � ge
Lð2Þhf þ L
ð2ÞQ � ge ge
ge �Lð2Þhf þ L
ð2ÞQ � ge
0BBBB@
1CCCCA; ð6Þ
658 Phys. Chem. Chem. Phys., 2002, 4, 655–660
Publ
ishe
d on
17
Janu
ary
2002
. Dow
nloa
ded
by U
nive
rsid
ad d
e O
vied
o on
27/
10/2
014
10:3
6:35
. View Article Online

which the z-axis is parallel to the backbone of the helices of theunit, and the y-axis lies in the heme plane. Nevertheless, theresult is as unsatisfactory as the simulation of the Mossbauerspectrum of (TPP)Fe(2-MeImH)+2 done with the help of theHALS species and shown in Fig.5 of ref. 3. Fig. 3(b) shows thesame Mossbauer spectrum of MOP1 but simulated withthe assumption that iron 2 is in the spin-mixed state. Thehyperfine parameters of both species are summarized inTable 2. The fit was performed with and without spin–spincross relaxation. For the IS state mixed with LS doublet, theg-values are calculated in accordance with eqn. (10) and (11),and (16) and (17) in ref. 13. In the first order of approximation,the j2E�1 ,�1=2i state gives no contribution to the g-values ofthe ground state. Then a particularly simple form is valid:
gz ¼ 2x29; g? ¼ 4x29; ð7Þ
here, x9 is the weight of the j4A2 ,±1=2i state, as defined ineqn. (2). If the IS were in its pure state, x9 would be one, and gzand g? would have their conventional values 2 and 4 respec-tively. It is obvious that in our case the spin state is mixed,though still of predominantly IS nature, with g?¼ 3.46 andx9¼ 0.93. The corresponding value of gz in eqn. (7) should beequal to 1.73, which differs from the measured value of 1.52.However, this discrepancy cannot be observed in either of theexperimental spectra for the following reasons. On the onehand, the Mossbauer spectrum is insensitive to such a differ-ence in the g-value. On the other hand, the diffuse structure ofthe EPR spectrum in the region between 2 and 1.5 hinders thedetermination of the g-value with the required accuracy. Inaddition, the ‘‘ strong gmax ’’ complexes show the characteristicdecrease of the EPR signal at g� 3.5 with increasing tem-perature, and the whole EPR spectrum becomes undetectableabove 45 K.4 That is why it is rather complicated to get par-ticular information on the temperature dependent admixtureof the excited 2E- and 2B2-levels into the spin-mixed state. Thehyperfine values of the spin-mixed iron show the pronouncedaxial symmetry, with Az being significantly smaller than A? .This result, as well as the small isomer shift, is in agreementwith the IS data obtained in ref. 20. In addition, the z-axis ofthe A-tensor is parallel to the x-axis of the g-tensor, in otherwords, it lies in the bH heme plane.Finally, we dwell briefly on the Euler rotation of the EFG
tensors of iron 1 and iron 2. In the case of the LS state, therotation is defined primarily by the angle a1 around the axisnormal to the bL heme plane, with the two other angles beingmuch smaller.21 In the case of the spin-mixed state, the z-axisof the q-tensor, similar to that of the A-tensor, is rather closeto the bH heme plane.The spin–spin relaxation rate g corresponds to the char-
acteristic time value of about 0.1 MHz.22 The spin–spin originof the observed relaxation is confirmed by the fact that theMossbauer spectra of MOP1 measured between 2.4 K and 15K (not shown here) are temperature independent. In Fig. 3(b)
the influence of the relaxation on the simulation of theMossbauer spectrum is shown. In the case where the relaxationwas not taken into consideration, line broadening was intro-duced yielding the line width Gexp equal to 0.49 mm s�1. Onecan easily see that the simulation without relaxation is worsethan that with relaxation.Fig. 4(a) shows the spectrum of MOP1 measured at 78 K in a
transverse magnetic field of 20 mT. It consists of an asymmetricdoublet typical for the Fe(III) relaxed state which is well knownin the case of cytochromes. The simulation is done with thecontribution of both irons with hyperfine parameters beingfixed at the values obtained from the low-temperature spec-trum, and with varied relaxation rate g. In addition, we alloweda change in the quadrupole splitting and in the isomer shift ofboth iron 1 and iron 2. The parameters are given in Table 3. Theline width Gexp was equal to 0.37 mm s�1. The isomer shifts ofboth irons show no temperature dependence (compare Table2). The quadrupole splitting of the LS species is temperatureindependent also, in full agreement with the strong crystal fieldmodel. The quadrupole splitting of the spin-mixed species isidentified predominantly with the IS typical splitting which ischaracterized by the strong descending dependence.20 How-ever, the observed change in the magnitude of quadrupolesplitting from 3.06 mm s�1 to 1.60 mm s�1 in the spin-mixedspecies is really drastic. Keeping in mind the temperatureinstability of the spin-mixed state mentioned above, whichmight originate from the structural changes with temperature,we propose as a possible explanation the significant increase ofthe admixture of the low-spin levels into this state at the tem-perature of liquid nitrogen and higher temperatures. For the
Fig. 4 Mossbauer spectrum of MOP1-bis-heme at 78 K (a) and at167 K (b). Iron 1 ion is in the LS state (dot-dashed line), and iron 2 ionis in the spin-mixed state (dotted line). The solid line gives the sum ofboth contributions.
Table 2 Crystal field and hyperfine parameters of MOP1-bis-hemeobtained from the simulation of the 2.4 K Mossbauer spectrum withthe assumption that the electronic ground state of the bL heme groupiron is a LS state, and of the bH heme group iron is a spin-mixed state
Low-spin, iron 1 Spin-mixed, iron 2
gx , gy , gz 1.50, 2.25, 2.93 3.46, 3.46, 1.52Ax , Ay , Az=mm s�1a � 2.74, 1.19,3.38 1.27, 6.12, 5.20d=mm s�1 0.29 0.21DEQ=mm s�1 2.29 3.06Z (from fit) � 1.68 � 1.58g=MHz 1.62 1.62a1 , a2 , a=� 132, 14, 18 101,124, 94
a Given for the excited state.
Table 3 Mossbauer parameters of MOP1-bis-heme obtained from thesimulation of the Mossbauer spectra measured at 78 K and at 167 KThe parameters of the 167 K spectrum are given in brackets
Low-spin, iron 1 Spin-mixed, iron 2
d=mm s�1 0.29 (0.23) 0.21 (0.21)DEQ=mm s�1 2.26 (2.25) 1.6 (1.74)g=MHz 164.4 164.4
Phys. Chem. Chem. Phys., 2002, 4, 655–660 659
Publ
ishe
d on
17
Janu
ary
2002
. Dow
nloa
ded
by U
nive
rsid
ad d
e O
vied
o on
27/
10/2
014
10:3
6:35
. View Article Online
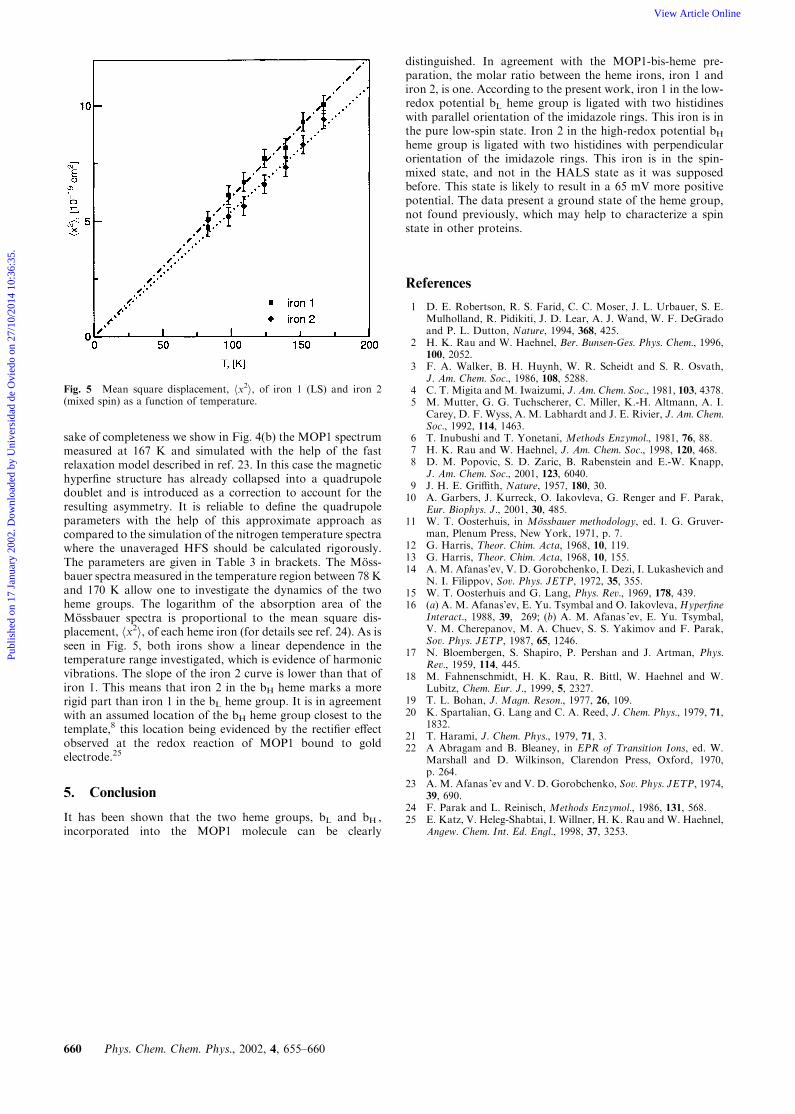
sake of completeness we show in Fig. 4(b) the MOP1 spectrummeasured at 167 K and simulated with the help of the fastrelaxation model described in ref. 23. In this case the magnetichyperfine structure has already collapsed into a quadrupoledoublet and is introduced as a correction to account for theresulting asymmetry. It is reliable to define the quadrupoleparameters with the help of this approximate approach ascompared to the simulation of the nitrogen temperature spectrawhere the unaveraged HFS should be calculated rigorously.The parameters are given in Table 3 in brackets. The Moss-bauer spectra measured in the temperature region between 78 Kand 170 K allow one to investigate the dynamics of the twoheme groups. The logarithm of the absorption area of theMossbauer spectra is proportional to the mean square dis-placement, hx2i, of each heme iron (for details see ref. 24). As isseen in Fig. 5, both irons show a linear dependence in thetemperature range investigated, which is evidence of harmonicvibrations. The slope of the iron 2 curve is lower than that ofiron 1. This means that iron 2 in the bH heme marks a morerigid part than iron 1 in the bL heme group. It is in agreementwith an assumed location of the bH heme group closest to thetemplate,8 this location being evidenced by the rectifier effectobserved at the redox reaction of MOP1 bound to goldelectrode.25
5. Conclusion
It has been shown that the two heme groups, bL and bH ,incorporated into the MOP1 molecule can be clearly
distinguished. In agreement with the MOP1-bis-heme pre-paration, the molar ratio between the heme irons, iron 1 andiron 2, is one. According to the present work, iron 1 in the low-redox potential bL heme group is ligated with two histidineswith parallel orientation of the imidazole rings. This iron is inthe pure low-spin state. Iron 2 in the high-redox potential bHheme group is ligated with two histidines with perpendicularorientation of the imidazole rings. This iron is in the spin-mixed state, and not in the HALS state as it was supposedbefore. This state is likely to result in a 65 mV more positivepotential. The data present a ground state of the heme group,not found previously, which may help to characterize a spinstate in other proteins.
References
1 D. E. Robertson, R. S. Farid, C. C. Moser, J. L. Urbauer, S. E.Mulholland, R. Pidikiti, J. D. Lear, A. J. Wand, W. F. DeGradoand P. L. Dutton, Nature, 1994, 368, 425.
2 H. K. Rau and W. Haehnel, Ber. Bunsen-Ges. Phys. Chem., 1996,100, 2052.
3 F. A. Walker, B. H. Huynh, W. R. Scheidt and S. R. Osvath,J. Am. Chem. Soc., 1986, 108, 5288.
4 C. T. Migita and M. Iwaizumi, J. Am. Chem. Soc., 1981, 103, 4378.5 M. Mutter, G. G. Tuchscherer, C. Miller, K.-H. Altmann, A. I.
Carey, D. F. Wyss, A. M. Labhardt and J. E. Rivier, J. Am. Chem.Soc., 1992, 114, 1463.
6 T. Inubushi and T. Yonetani, Methods Enzymol., 1981, 76, 88.7 H. K. Rau and W. Haehnel, J. Am. Chem. Soc., 1998, 120, 468.8 D. M. Popovic, S. D. Zaric, B. Rabenstein and E.-W. Knapp,
J. Am. Chem. Soc., 2001, 123, 6040.9 J. H. E. Griffith, Nature, 1957, 180, 30.10 A. Garbers, J. Kurreck, O. Iakovleva, G. Renger and F. Parak,
Eur. Biophys. J., 2001, 30, 485.11 W. T. Oosterhuis, in Mossbauer methodology, ed. I. G. Gruver-
man, Plenum Press, New York, 1971, p. 7.12 G. Harris, Theor. Chim. Acta, 1968, 10, 119.13 G. Harris, Theor. Chim. Acta, 1968, 10, 155.14 A. M. Afanas’ev, V. D. Gorobchenko, I. Dezi, I. Lukashevich and
N. I. Filippov, Sov. Phys. JETP, 1972, 35, 355.15 W. T. Oosterhuis and G. Lang, Phys. Rev., 1969, 178, 439.16 (a) A. M. Afanas’ev, E. Yu. Tsymbal and O. Iakovleva, Hyperfine
Interact., 1988, 39, 269; (b) A. M. Afanas ’ev, E. Yu. Tsymbal,V. M. Cherepanov, M. A. Chuev, S. S. Yakimov and F. Parak,Sov. Phys. JETP, 1987, 65, 1246.
17 N. Bloembergen, S. Shapiro, P. Pershan and J. Artman, Phys.Rev., 1959, 114, 445.
18 M. Fahnenschmidt, H. K. Rau, R. Bittl, W. Haehnel and W.Lubitz, Chem. Eur. J., 1999, 5, 2327.
19 T. L. Bohan, J. Magn. Reson., 1977, 26, 109.20 K. Spartalian, G. Lang and C. A. Reed, J. Chem. Phys., 1979, 71,
1832.21 T. Harami, J. Chem. Phys., 1979, 71, 3.22 A Abragam and B. Bleaney, in EPR of T ransition Ions, ed. W.
Marshall and D. Wilkinson, Clarendon Press, Oxford, 1970,p. 264.
23 A. M. Afanas ’ev and V. D. Gorobchenko, Sov. Phys. JETP, 1974,39, 690.
24 F. Parak and L. Reinisch, Methods Enzymol., 1986, 131, 568.25 E. Katz, V. Heleg-Shabtai, I. Willner, H. K. Rau and W. Haehnel,
Angew. Chem. Int. Ed. Engl., 1998, 37, 3253.
Fig. 5 Mean square displacement, hx2i, of iron 1 (LS) and iron 2(mixed spin) as a function of temperature.
660 Phys. Chem. Chem. Phys., 2002, 4, 655–660
Publ
ishe
d on
17
Janu
ary
2002
. Dow
nloa
ded
by U
nive
rsid
ad d
e O
vied
o on
27/
10/2
014
10:3
6:35
. View Article Online