modelling aerobic granular sludge reactors through ...evolcke/pdf/2018 baeten wr... · sludge...
TRANSCRIPT

lable at ScienceDirect
Water Research 146 (2018) 134e145
Contents lists avai
Water Research
journal homepage: www.elsevier .com/locate/watres
Modelling aerobic granular sludge reactors through apparent half-saturation coefficients
Janis E. Baeten a, *, Mark C.M. van Loosdrecht b, Eveline I.P. Volcke a
a Department of Green Chemistry and Technology, Faculty of Bioscience Engineering, Ghent University, Coupure Links 653, 9000, Gent, Belgiumb Department of Biotechnology, Faculty of Applied Sciences, Delft University of Technology, Van der Maasweg 9, 2629 HZ, Delft, the Netherlands
a r t i c l e i n f o
Article history:Received 29 June 2018Received in revised form6 September 2018Accepted 7 September 2018Available online 12 September 2018
Keywords:Half-saturation coefficientMonodDiffusionGranular sludgeBiofilm modellingASM
* Corresponding author.E-mail address: [email protected] (J.E. Baeten
https://doi.org/10.1016/j.watres.2018.09.0250043-1354/© 2018 Elsevier Ltd. All rights reserved.
a b s t r a c t
During biological wastewater treatment, substrates undergo simultaneous diffusion and reactions insidemicrobial aggregates, creating microscale spatial substrate gradients and limiting the macroscale reactionrates. For flocculent and anaerobic granular sludge, this rate-limiting effect of diffusion is often lumped inmodel parameters, like the half-saturation coefficients of Monod kinetics in activated sludge models (ASM).Yet, an explicit description of the reaction-diffusion process with biofilm models is more common foraerobic granular sludge. This work investigates whether apparent half-saturation coefficients could haveapplications for aerobic granular sludge as well and examines the implications of this simplification. To thisend, the macroscopic reaction rates predicted with a one-dimensional biofilm (1D) model were fitted withMonod kinetics. The results showed that the macroscale rates could indeed be described using apparentkinetics, at the very least over a time scale where the microbial population distribution stays fixed.However, the coefficients were sensitive to changes in the microbial population distribution, which can beaffected by long-term changes in operating conditions. Also the activity of organisms that compete for thesame substrates affect the parameter value. Be that as it may, apparent kinetics also depend on theoperating conditions for flocculent and anaerobic granular sludge, but they have still been used successfullyfor design and optimization. Therefore, the last section of this work illustrates that they may also haveapplications for aerobic granular sludge. A simple model for ammonium removal using apparent half-saturation coefficients for oxygen and ammonium is applied to a full-scale reactor, taking advantage ofthe batch-wise operation and on-line monitoring data for regular recalibration.
© 2018 Elsevier Ltd. All rights reserved.
1. Introduction
Wastewater treatment with aerobic granular sludge has severaladvantages compared to conventional activated sludge. The highsettling velocity of granules enables efficient sludge separationinside the reactor itself, reducing the plant footprint by 75% (deBruin et al., 2004). The removal of organic material (often ex-presses as chemical oxygen demand or COD), nitrogen (N) andphosphorus (P) can be achieved in the same tank because of thebatch-wise reactor operation and different redox zones insidegranules. The integration of COD, N and P removal together withsludge separation in a single tank avoids the need for recycle andreturn pumps, which decreases energy demands (Pronk et al.,2015). The distinct chemical composition of the extracellular
).
polymers forming the granules opens up possibilities for the re-covery of biopolymers (Lin et al., 2015). Over the last two decades,the yearly number of publications on aerobic granular sludge haveincreased by 19% per year on average (Fig. S1), while full-scale re-actors have become available on the market.
Given the increasing interest in aerobic granular sludge tech-nology, the need for reliable and user-friendly mathematicalmodels is growing. So far, modelling efforts have mainly focussedon lab-scale systems. The most popular approach is to explicitlydescribe diffusion of substrates inside granules with 1D (Beun et al.,2001; Su and Yu, 2006; Winkler et al., 2015) or 2D biofilm models(Xavier et al., 2007). The reaction rate at every point in a granule isthen typically calculated with Monod kinetics (Eq. (1)).
m ¼ mMaxSlim
Kþ Slim(1)
where m is the specific growth rate (d�1), mMax the maximal specific

J.E. Baeten et al. / Water Research 146 (2018) 134e145 135
growth rate (d�1), Slim the limiting substrate concentration (g.m�3)and K the intrinsic half-saturation coefficient (g.m�3).
Such biofilm models can predict spatial gradients of solutes andmicrobial populations inside granules and are thus useful to gaininsight in the microscale behaviour. Yet, their complexity canhinder their adoption and effective use by the industry for reactordesign and optimization. One important barrier is the search forreliable parameter values. In literature, strongly different values formicroscale parameters can be found, as illustrated in Fig. 1 for theoxygen diffusion coefficient inside biofilms. A second option is tomeasure parameter values, but easily measurable parameters suchas the number, density and size of granules depend on the oper-ating conditions (de Kreuk et al., 2005) and their changes are stillchallenging to predict (Baeten et al., unpublished results). Finally,calibration can be performed, but a rational selection of a limitednumber of parameters to adjust is difficult with so many parame-ters to choose from. A guideline for calibration of biofilm modelshas only recently been published and thus the practical applica-bility is still to be confirmed (Rittmann et al., 2018), especially forgranular sludge with its own particularities.
To develop practically applicable models, the guideline formu-lated by Wanner et al. (2006) should be used: “A model should beas simple as possible, and only as complex as needed”. This para-phrase of Occam's razor also lay at the basis of the widely appliedactivated sludge models (ASM). The primary goal of these modelswas to predict the macroscale behaviour of wastewater treatmentplants, more specifically the effluent quality, oxygen requirementsand sludge production (Henze et al., 2000). These goals weredeemed achievable without explicit consideration of microscaleprocesses like diffusion, even though it was known that substrategradients can exist and affect removal rates (Benefield and Molz,1983; Hamdi, 1995; Matson and Characklis, 1976). In ASM, therate-limiting effect of diffusion is simply accounted for by using“apparent” half-saturation coefficients (Kapp) in Monod kinetics.These have higher values than the intrinsic coefficients (K) (Eq. (1)),which are the substrate concentrations at which half of themaximal growth rate is achieved in well-mixed reactors withperfectly suspended cells (Arnaldos et al., 2015).
Apparent half-saturation coefficients increase with the floc size(P�erez et al., 2005). Because the floc size depends on the degree ofmixing in a reactor, the parameter values can change up to a factor 3or more when the aeration intensity changes (Chu et al., 2003). Thesize of micro-colonies inside flocs also influences apparent half-saturation coefficients (Picioreanu et al., 2016). The micro-colonysize depends on the solid retention time, among others, so this isanother reason to recognize apparent half-saturation coefficients assystem specific and variable over time. For example, values be-tween 0.074 and 3 g O2.m�3 have been found for the apparent half-saturation coefficient for oxygen of ammonium oxidizing organ-isms in flocculent sludge (Vannecke and Volcke, 2015). Neverthe-less, all commercial modelling software such as Biowin, SUMO andWEST apply them for modelling flocculent sludge processes. Thesuccessful application of activated sludge models (Brdjanovic et al.,2015) demonstrates that models for design and optimization do notneed a full description of all microscale processes.
Fig. 1. Variation of measured oxygen diffusion coefficients inside biofilms relative to the valuinside aerobic granules in different biofilm models (Beun et al., 2001; Kagawa et al., 2015;
Models with apparent half-saturation coefficients instead ofreaction-diffusion kinetics have not only been used for flocculentsludge, but also for aerobic (Lübken et al., 2005; Zhou et al., 2013),anammox (Jones et al., 2007) and particularly often for anaerobicgranular sludge reactors (Chen et al., 2015; Fuentes et al., 2011;Kalyuzhnyi et al., 2006; Lohani et al., 2016). It is not immediatelyclear why apparent kinetics are less popular for aerobic granularsludge than for flocculent and anaerobic granular sludge, whichindicates that the implications of this approach are not yetcompletely understood (Baeten et al., unpublished results). There-fore, this work investigates the implications and applicability ofapparent kinetics for modelling aerobic granular sludge throughthe following three research questions.
1. Can apparent half-saturation coefficients lump the reaction-diffusion process inside granules? It is important to verifywhether Monod kinetics are able to capture the effect of short-term (minutes to hours) changes in limiting substrate concen-trations on macroscale removal rates, as they can for flocs.Simulation results of a 1D biofilm model were used as areference.
2. Which factors influence apparent half-saturation coefficients?Reactions that compete for the same substrates, e.g. ammoniumand nitrite oxidation compete for oxygen, could alter intra-granule substrate gradients and thus influence apparent half-saturation coefficients (Picioreanu et al., 2016). Long-term(days to weeks) changes in operating conditions could inducechanging microbial population distributions, on its turn alsoaffecting the coefficients. A 1D biofilm model was used as areference for this question as well.
3. Could apparent half-saturation coefficients be applicable inpractice? Possible applications are illustrated by confronting asimple model for ammonium removal based on apparent half-saturation constants with on-line measurements from a full-scale aerobic granular sludge reactor.
2. Methodology
2.1. Can apparent half-saturation coefficients lump reaction anddiffusion inside granules?
2.1.1. Reference operating conditions and 1D biofilm modelThe operating conditions of the laboratory aerobic granular
sludge reactor described by de Kreuk et al. (2007) were used as areference scenario for the simulations. The sequencing batchreactor with a total volume of 3 L was operated in cycles of 3 h,divided in 60min unaerated feeding, 112min aeration, 3minsettling and 5min effluent discharge with a volume exchange ratioof 50%. During the aeration phase, the dissolved oxygen concen-trationwas controlled at 2 g O2.m�3 and the pH at 7. About 600,000granules with an average diameter of 0.55mm were present. Theinfluent contained acetate, ammonium and phosphate in concen-trations of 396 g COD.m�3, 50 g N.m�3 and 20 g P.m�3.
A model of this reactor was set up in Aquasim (Reichert, 1994).
e in water (data from Stewart (1998)) and assumed densities of heterotrophic biomassNi et al., 2010; Xavier et al., 2007).

J.E. Baeten et al. / Water Research 146 (2018) 134e145136
This software comprises a dynamic 1D biofilm model, which cal-culates the microbial population distribution over the granuledepth resulting from competition for space and substrates (Wannerand Gujer, 1986). The biological reactions were described withASM2d (Henze et al., 2000) as corrected by Hauduc et al. (2010) andextended with two-step nitrification and temperature corrections.As such, a basic representation of the metabolism of ordinaryheterotrophic organisms (OHO), phosphate accumulating organ-isms (PAO), ammonium oxidizing organisms (AOO) and nitriteoxidizing organisms (NOO) was obtained to describe the COD, Nand P removal. Mass transfer resistance from the bulk liquid to thegranule surfacewas neglected. Tomodel the intragranule transport,a compound-specific and temperature-dependent estimation ofthe effective diffusion coefficient inside a biofilm matrix was used(Stewart, 2003).
In Aquasim, the granule depth was divided into 20 grid points.The total reactor volume was split up into a biofilm and a mixedcompartment, coupled with a diffusive link (exchange coefficient1000m3 h�1) to let the liquid phases behave as one perfectly mixed,fluctuating water volume without numerical errors (Baeten et al.,2017). Transformation processes in the mixed compartment wereinactivated and the biofilm compartment was chosen small (0.45 L)so that it contained only 0.03 L of bulk liquid (1% of the total reactorvolume). This makes the contribution of suspended biomass tomacroscale reaction rates negligible, facilitating the interpretationof the results. More details on the model are provided in the sup-plementary material: the stoichiometric matrix (Table S2),composition matrix (Table S3), process rate equations (Table S4),parameter values (Table S5, 6 and 7) and model validation results.
2.1.2. Calculation of apparent half-saturation coefficientsApparent half-saturation coefficients of the growth reactions of
all microbial groups in the 1D biofilmwere determined for all theirlimiting soluble substrates (Table 1) using the method schematizedin Fig. 2. In summary, the method comprises an initial prediction ofthe microbial population distribution and then a fixation of thisdistribution to study the response of these granules to changingsubstrate concentrations, analogous to the method of Picioreanuet al. (2016). The model in Aquasim was first run with the refer-ence operating conditions for 750 days to ensure a steady-statemicrobial population distribution (step 1). Next, the stoichio-metric coefficients of all biomass variables were set to zero toobtain a fixed population distribution during the determination ofthe apparent half-saturation coefficient, independent from theduration of the simulations. This was done because changes in thedistribution are also not expected during short-term changes ofsubstrate concentrations, as occur during one reactor cycle. All re-actions except for one growth reaction from Table 1 were subse-quently deactivated to avoid competition for the limiting substrate.This is analogous to experimental assessments of half-saturationcoefficients, where the activity of competing organisms is mini-mized by depriving them of substrates or adding deactivatingagents (Manser et al., 2005). After these preparations, the granuleswere subjected to a step-wise decreasing concentration of one
Table 1Overview of determined apparent half-saturation coefficients for the respectively sevenefficients were determined individually, by keeping only one reaction active at a time (thesupplementary material). VFA: volatile fatty acids. NHx: ammonium and ammonia nitro
F VFA PO4 O2
OHO KF,OHO,app (4) KVFA,OHO,app (5) KPO4,OHO,app (4) KO2,OH
PAO KPO4,PAO,app (16) KO2,PAO
AOO KO2,AOO
NOO KO2,NO
substrate, while all other substrates for the active growth reactionwere kept available in excess (1000 gm�3) (step 2).
The simulation results were exported to Excel to calculate theamount of biomass of the microbial group of interest by integrationof the local concentration over the granule depth (Eq. (2)).
MXtot;i ¼ZR
0
Xi43pr3dr (2)
where MXtot,i is the total biomass of the microbial group of interestin a granule (g COD), Xi the local biomass concentration at a dis-tance r from the granule core (g COD.m�3), R the granule radius (m)and r the distance from the granule core (m).
Then, the total growth rate (macroscale) of this group wascalculated for every applied limiting substrate concentration bynumerical integration of the local rates over the granule depth(microscale) and successive division by the amount of biomass (Eq.(3)).
mtot;j ¼
Z R
0rj43pr3dr
MXtot;i(3)
where rj is the local rate of the growth reaction of interest at adistance r from the granule core (g COD.m�3.d�1), as calculated byAquasim as the local specific growth rate times the local biomassconcentration, and mtot,i is the total specific growth rate in a granule(d�1).
After every step change of the substrate concentration, a newsteady-state total growth rate was reached as soon as the substrategradients stabilized. Monod kinetics (Eq. (4)) were finally fitted tothis steady-state total specific growth rate as a function of thelimiting substrate concentration in the bulk liquid with the solverin Excel by adjusting Kapp to minimize the sum of squared errors(step 3). mMax,i was not calibrated, but kept the same as in thebiofilm model.
mtot;jzmMax;jSlim
Kapp þ Slim(4)
where mMax,j is the maximal specific growth rate (d�1), Slim thelimiting substrate concentration in the bulk liquid (g.m�3) and Kappthe apparent half-saturation coefficient (g.m�3).
To find the other apparent half-saturation coefficients (Table 1),only step 2 and 3 were repeated for the other reactions and limitingsubstrates.
To determine the apparent half-saturation coefficients of PAO,an excess of the storage compound PHA had to be introduced tominimize limitations by this intracellular limiting substrate duringstep 2. To this end, the precursor VFA was fed with a high con-centration (10,000 g COD.m�3) to the granules for more than 2 hbefore changing the concentration of the limiting soluble substrateof interest. As such, there was time for PHA to accumulate.
, five and two limiting substrates for OHO, PAO and AOO and NOO growth. The co-number in between brackets refers to the reaction under concern, as specified in thegen. PO4: soluble inorganic phosphorus.
NHx NO2� NO3
�
O,app (4) KNHx,OHO,app (4) KNO2,OHO,app (6) KNO3,OHO,app (7),app (16) KNHx,PAO,app (16) KNO2,PAO,app (17) KNO3,PAO,app (18),app (22) KNHx,AOO,app (22)
O,app (24) KNO2,NOO,app (24)

Fig. 2. Method to estimate the apparent half-saturation coefficient Kapp of one microbial group for one substrate using the 1D biofilm model. Only step 2e3 were repeated toestimate this parameter for other microbial groups and substrates. *For research question three, the operating conditions were different from the reference case to evaluate theeffect of long-term changes in operation on the microbial population distribution and associated apparent half-saturation coefficients. **For research question two, more than onegrowth reaction was kept active to assess the effect of competition.
J.E. Baeten et al. / Water Research 146 (2018) 134e145 137
2.2. Which factors influence half-saturation coefficients?
The factors that influence half-saturation coefficients werestudied by means of an example, more specifically the effect ofoxygen on the growth of ammonium oxidizing organisms (AOO),characterised by KO2,AOO,app.
2.2.1. Influence of competitionThe effect of oxygen on the macroscale AOO growth rate was
again determined according to Fig. 2, but this time the aerobicgrowth reactions of other microbial groups were kept active one byone during step 2 (reactions number 5, 16 and 24 in thesupplementary material). The required substrates for the othergrowth reactions were provided in excess to get an idea of themaximum possible interference. For example, the effect of simul-taneous aerobic OHO growth was assessed by keeping the reactionactive in the model and providing VFA, NHx and PO4 at a concen-tration of 1000 gm�3.
2.2.2. Influence of long-term changes in operating conditionsAgain, KO2,AOO,app was determined with the method shown in
Fig. 2, but this time after reaching the steady-state microbial pop-ulation distribution for other operating conditions in step 1. Theinfluent VFA, NHx and PO4 concentrations, temperature, O2 set-point and the granule radius were varied individually,
to �30%, �20%, �10% and þ10%, þ20% and þ30% of their referencevalue. For every new steady-state thus obtained, KO2,AOO,app wasdetermined. The observed changes of KO2,AOO,app were related totwo measures that characterize the microbial population distribu-tion. The first one is the total AOO biomass present in the granuleMXtot,AOO, calculated with Eq. (2). The second indicator was theaverage depth of the AOO population, calculated by numericalintegration in excel (Eq. (5)).
Average population depth ¼ R �
Z R
0r�XAOO
43pr3
�dr
MXtot;AOO(5)
2.3. Are apparent-half saturation coefficients practicallyapplicable?
On-linemonitoring data from a 9600m3 aerobic granular sludgereactor treating 14,300m3 d�1 of sewage in Garmerwolde, TheNetherlands, were used. Every reactor cycle starts with a feedingphase of 1e1.5 h, during which influent is fed from the bottom tocreate a plug-flow, which enables simultaneous effluent dischargefrom the top. Afterwards, aeration is provided during a reactionphase of 1e5 h. Finally, a short settling phase allows the granules to

J.E. Baeten et al. / Water Research 146 (2018) 134e145138
stay in the reactor, before the cycle is repeated. Details on the plantoperation and performance can be found in Pronk et al. (2015).Dissolved oxygen was measured in-situ and ammonium ex-situ(5e10min interval; Hach Lange; Filtrax, AMTAX). Data fromAugust 2017 were extracted from the database as average valuesper minute. Signals from the ammonium sensor during automaticcleaning and calibration of the device were filtered out.
A model for ammonium removal with Monod kinetics wasapplied to the data from the aeration phases. The model describesthe removal rate as a function of the ammonium concentration SNHx(g N.m�3) and dissolved oxygen concentration SO2 (g O2.m�3) (Eq.(6)), using a maximal ammonium removal rate rmax (g N.m�3.d�1)and apparent half-saturation coefficients KNHx,app (g N.m�3) andKO2,app (g O2.m�3).
dSNHxdt
¼ rmaxSNHx
KNHx;app þ SNHx
SO2KO2;app þ SO2
(6)
For each cycle, the simulation was started at the moment wherethe NHx concentration reaches a maximum, indicating that the fedinfluent and reactor content were mixed thoroughly by aeration.This measured maximum NHx concentration was used as initialcondition. The parameters rmax, KNHx,app and KO2,app for every cyclewere obtained by minimizing the sum of squared errors (SSE, Eq.(7)) simultaneously for the preceding 10 cycles (Fig. 3). By fittingseveral cycles at once, a wide variety of combinations of NHx andO2 concentrations was taken into account, which benefits param-eter identifiability. Themean absolute error (MAE, Eq. (8)) was usedto quantify the goodness-of-fit for every cycle, which compensatesfor the different number of measurements per cycle (n)
SSE ¼Xni
�SNHx;measured;i � SNHx;predicted;i
�2(7)
MAE ¼Pn
i
���SNHx;measured;i � SNHx;predicted;i���
n(8)
Calculations were performed in Matlab: ‘ode45’ was used tosolve the differential equation and for calibration ‘lsqnonlin’ wasused in combinationwith ‘multistart’ to find the global minimumofthe SSE.
3. Results and discussion
3.1. Can apparent half-saturation coefficients lump reaction anddiffusion inside granules?
3.1.1. Effect of the substrate concentrationThe dependency of aerobic growth rates on the bulk oxygen
concentration is displayed in Fig. 4 for the steady-state microbialpopulation distribution under the reference conditions (Figure S4).An increasing bulk oxygen concentration resulted in an initial steep
Fig. 3. Moving window used for calibration and prediction of the ammonium removalin the full-scale reactor.
increase of the growth rate, followed by a slower increase towardsthe maximal specific growth rate. This saturation behaviour couldbe described reasonably well with Monod kinetics upon calibrationof the apparent half-saturation coefficient. The fit for NOO is almostperfect. For OHO, Monod kinetics seem the least appropriate, butstill the qualitative behaviour is the same. This could be sufficientlyaccurate, given the comparison is nowmade with another model (a1D biofilm), with its own uncertainty. In practice, OHO also onlyplay a secondary role for the total COD removal and oxygen con-sumption in an aerobic granular sludge reactor, given the highabundancy of PAO (Pronk et al., 2015). Overall, Monod kinetics withapparent half-saturation coefficients seem suitableapproximations.
Apparent half-saturation coefficients have been used before tomodel granular sludge reactors. For example, Zhou et al. (2013) andLübken et al. (2005) used ASM3 without explicit consideration ofdiffusion. Since stable granules were already formed in themodelled reactors and no significant unidirectional changes in theinfluent composition or reactor temperature occurred during thestudied time-frame, a steady microbial population distributionwasprobably present throughout their experiments. This correspondswith our results, that for a fixed microbial population distribution,constant apparent half-saturation coefficients could indeed be usedto incorporate diffusion resistances.
The dependencies of the total specific growth rates on the otherlimiting substrates were also calculated and fitted with Monodkinetics for the reference conditions (Figure S5). The ratio of theestimated apparent over the used intrinsic half-saturation coeffi-cient varies between 1 and 6 (Fig. 5). This wide range can beexplained by different substrate diffusivities, microbial populationdistributions and intrinsic kinetics (P�erez et al., 2005). For example,for OHO, the same intrinsic kinetics were used for fermentablesubstrate (F) and VFA (KVFA,OHO ¼ KF,OHO¼ 4 g COD.m�3; Table S6),but the effective diffusion coefficient of fermentable substrate islower (DF_e20¼1.6 10�5m2 d�1<DVFA_e20¼ 2.6 10�5m2 d�1;Table S7), leading to more diffusion resistance and thus a higherapparent half-saturation coefficient. The same intrinsic half-saturation coefficient for O2 was used for OHO and PAO in the 1Dbiofilmmodel (KO2,PAO¼ KO2,OHO¼ 0.20 g O2.m�3), yet the apparentcoefficients are different because the intragranule distribution andmaximal growth rate of these microbial groups is different (Fig. 4S).Finally, a similar effective diffusion coefficients of O2 and NHx wasused (DO2,e¼ 1.37 10⁻4m2 d�1 and DNHx,e¼ 1.45 10�4m2 d�1), butthe ratio of the apparent to the intrinsic half-saturation coefficientof OHO differs strongly for these substrates because of the differentintrinsic half-saturation coefficient (KO2,OHO¼ 0.2 g O2.m�3 andKNHx,OHO¼ 0.05).
Arnaldos et al. (2015) hypothesized that half-saturation co-efficients of organic molecules are more strongly affected bydiffusion than inorganic molecules. The results from this studyshow that many more factors influence the ratio of the apparent tothe intrinsic half-saturation coefficient apart from the diffusioncoefficient, such as the maximal growth rate, microbial populationdistribution and the intrinsic half-saturation coefficient. The widerange of values for the Kapp/K ratio also indicates that a reduction ofthe maximal specific growth rates of all microbial groups with thesame factor, as applied by Pons et al. (2008), is an inadvisablemethod to account for the effect of diffusion (Fig. 5).
3.1.2. Time delayWhen the concentration of a limiting substrate changes in the
bulk liquid, the rates inside granules do not immediately reach anew steady-state since the formation of new substrate gradientsthrough simultaneous diffusion and reaction takes time.
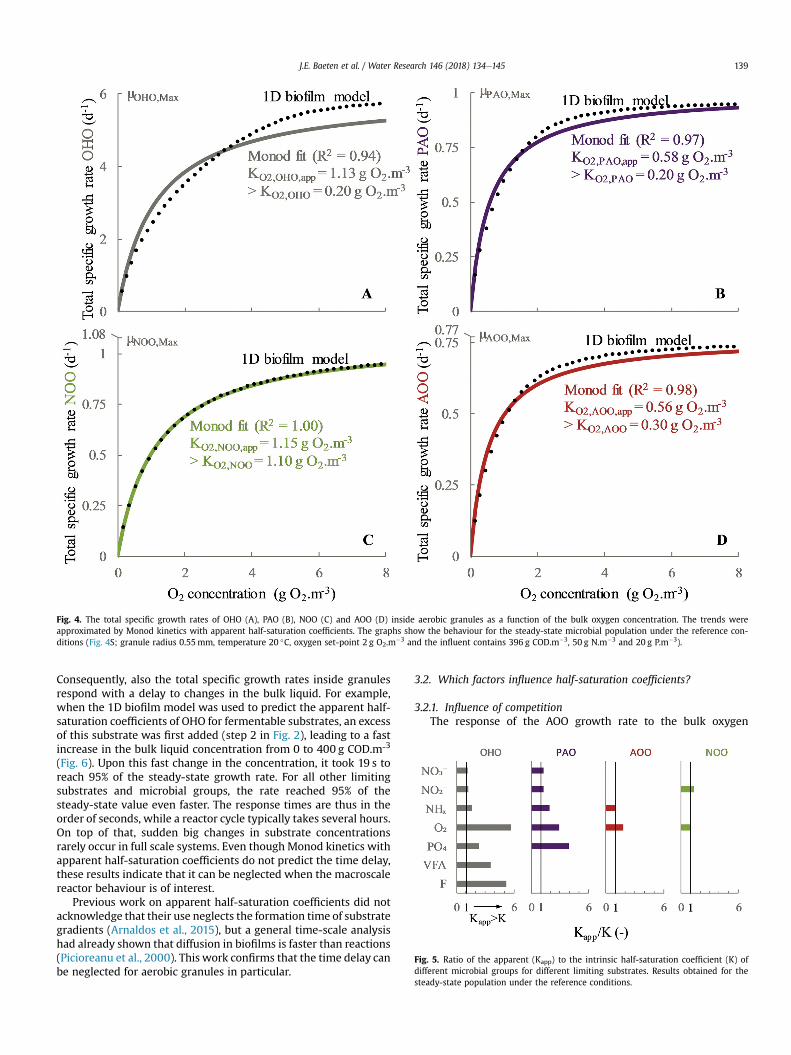
Fig. 4. The total specific growth rates of OHO (A), PAO (B), NOO (C) and AOO (D) inside aerobic granules as a function of the bulk oxygen concentration. The trends wereapproximated by Monod kinetics with apparent half-saturation coefficients. The graphs show the behaviour for the steady-state microbial population under the reference con-ditions (Fig. 4S; granule radius 0.55mm, temperature 20 �C, oxygen set-point 2 g O2.m�3 and the influent contains 396 g COD.m�3, 50 g N.m�3 and 20 g P.m�3).
Fig. 5. Ratio of the apparent (Kapp) to the intrinsic half-saturation coefficient (K) ofdifferent microbial groups for different limiting substrates. Results obtained for the
J.E. Baeten et al. / Water Research 146 (2018) 134e145 139
Consequently, also the total specific growth rates inside granulesrespond with a delay to changes in the bulk liquid. For example,when the 1D biofilm model was used to predict the apparent half-saturation coefficients of OHO for fermentable substrates, an excessof this substrate was first added (step 2 in Fig. 2), leading to a fastincrease in the bulk liquid concentration from 0 to 400 g COD.m⁻3
(Fig. 6). Upon this fast change in the concentration, it took 19 s toreach 95% of the steady-state growth rate. For all other limitingsubstrates and microbial groups, the rate reached 95% of thesteady-state value even faster. The response times are thus in theorder of seconds, while a reactor cycle typically takes several hours.On top of that, sudden big changes in substrate concentrationsrarely occur in full scale systems. Even though Monod kinetics withapparent half-saturation coefficients do not predict the time delay,these results indicate that it can be neglected when the macroscalereactor behaviour is of interest.
Previous work on apparent half-saturation coefficients did notacknowledge that their use neglects the formation time of substrategradients (Arnaldos et al., 2015), but a general time-scale analysishad already shown that diffusion in biofilms is faster than reactions(Picioreanu et al., 2000). This work confirms that the time delay canbe neglected for aerobic granules in particular.
3.2. Which factors influence half-saturation coefficients?
3.2.1. Influence of competitionThe response of the AOO growth rate to the bulk oxygen
steady-state population under the reference conditions.
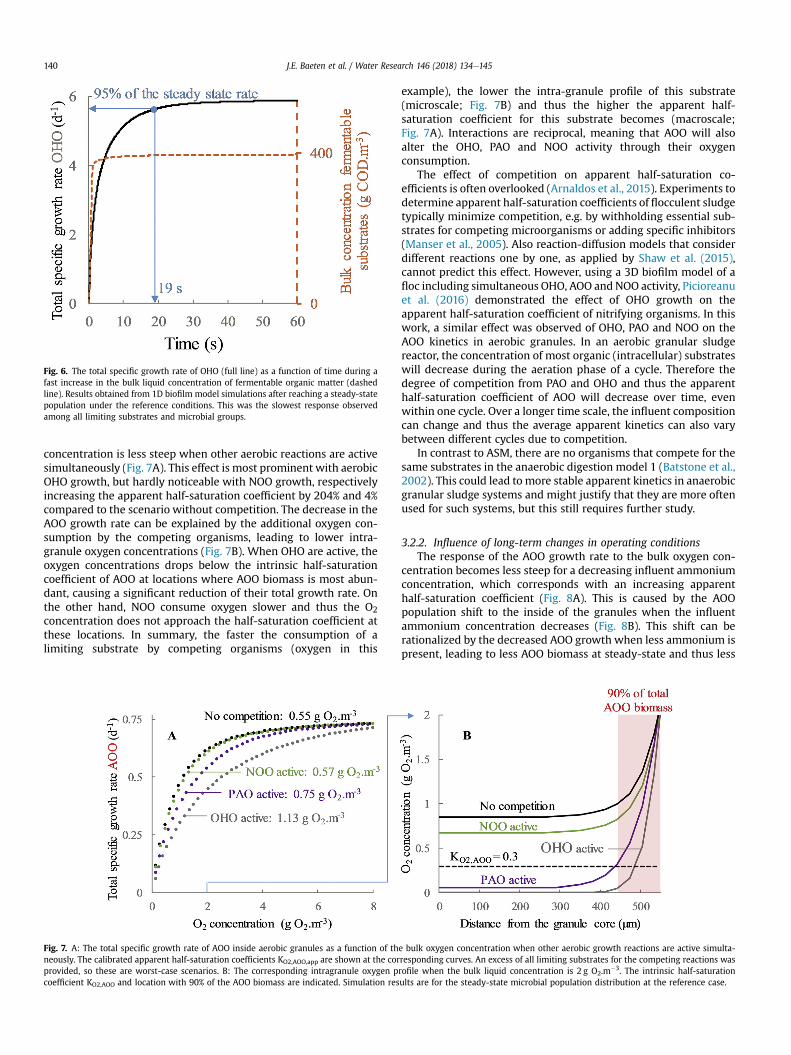
Fig. 6. The total specific growth rate of OHO (full line) as a function of time during afast increase in the bulk liquid concentration of fermentable organic matter (dashedline). Results obtained from 1D biofilm model simulations after reaching a steady-statepopulation under the reference conditions. This was the slowest response observedamong all limiting substrates and microbial groups.
J.E. Baeten et al. / Water Research 146 (2018) 134e145140
concentration is less steep when other aerobic reactions are activesimultaneously (Fig. 7A). This effect is most prominent with aerobicOHO growth, but hardly noticeable with NOO growth, respectivelyincreasing the apparent half-saturation coefficient by 204% and 4%compared to the scenario without competition. The decrease in theAOO growth rate can be explained by the additional oxygen con-sumption by the competing organisms, leading to lower intra-granule oxygen concentrations (Fig. 7B). When OHO are active, theoxygen concentrations drops below the intrinsic half-saturationcoefficient of AOO at locations where AOO biomass is most abun-dant, causing a significant reduction of their total growth rate. Onthe other hand, NOO consume oxygen slower and thus the O2concentration does not approach the half-saturation coefficient atthese locations. In summary, the faster the consumption of alimiting substrate by competing organisms (oxygen in this
Fig. 7. A: The total specific growth rate of AOO inside aerobic granules as a function of thneously. The calibrated apparent half-saturation coefficients KO2,AOO,app are shown at the corprovided, so these are worst-case scenarios. B: The corresponding intragranule oxygen pcoefficient KO2,AOO and location with 90% of the AOO biomass are indicated. Simulation res
example), the lower the intra-granule profile of this substrate(microscale; Fig. 7B) and thus the higher the apparent half-saturation coefficient for this substrate becomes (macroscale;Fig. 7A). Interactions are reciprocal, meaning that AOO will alsoalter the OHO, PAO and NOO activity through their oxygenconsumption.
The effect of competition on apparent half-saturation co-efficients is often overlooked (Arnaldos et al., 2015). Experiments todetermine apparent half-saturation coefficients of flocculent sludgetypically minimize competition, e.g. by withholding essential sub-strates for competing microorganisms or adding specific inhibitors(Manser et al., 2005). Also reaction-diffusion models that considerdifferent reactions one by one, as applied by Shaw et al. (2015),cannot predict this effect. However, using a 3D biofilm model of afloc including simultaneous OHO, AOO and NOO activity, Picioreanuet al. (2016) demonstrated the effect of OHO growth on theapparent half-saturation coefficient of nitrifying organisms. In thiswork, a similar effect was observed of OHO, PAO and NOO on theAOO kinetics in aerobic granules. In an aerobic granular sludgereactor, the concentration of most organic (intracellular) substrateswill decrease during the aeration phase of a cycle. Therefore thedegree of competition from PAO and OHO and thus the apparenthalf-saturation coefficient of AOO will decrease over time, evenwithin one cycle. Over a longer time scale, the influent compositioncan change and thus the average apparent kinetics can also varybetween different cycles due to competition.
In contrast to ASM, there are no organisms that compete for thesame substrates in the anaerobic digestion model 1 (Batstone et al.,2002). This could lead to more stable apparent kinetics in anaerobicgranular sludge systems and might justify that they are more oftenused for such systems, but this still requires further study.
3.2.2. Influence of long-term changes in operating conditionsThe response of the AOO growth rate to the bulk oxygen con-
centration becomes less steep for a decreasing influent ammoniumconcentration, which corresponds with an increasing apparenthalf-saturation coefficient (Fig. 8A). This is caused by the AOOpopulation shift to the inside of the granules when the influentammonium concentration decreases (Fig. 8B). This shift can berationalized by the decreased AOO growth when less ammonium ispresent, leading to less AOO biomass at steady-state and thus less
e bulk oxygen concentration when other aerobic growth reactions are active simulta-responding curves. An excess of all limiting substrates for the competing reactions wasrofile when the bulk liquid concentration is 2 g O2.m�3. The intrinsic half-saturationults are for the steady-state microbial population distribution at the reference case.
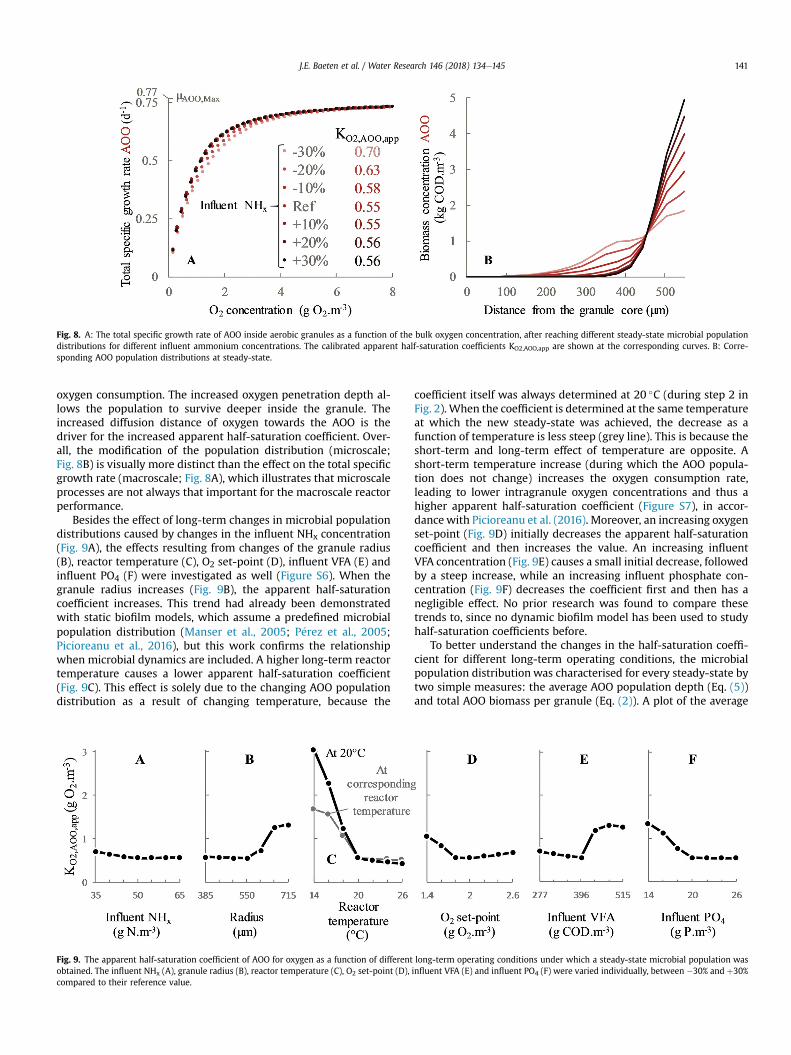
Fig. 8. A: The total specific growth rate of AOO inside aerobic granules as a function of the bulk oxygen concentration, after reaching different steady-state microbial populationdistributions for different influent ammonium concentrations. The calibrated apparent half-saturation coefficients KO2,AOO,app are shown at the corresponding curves. B: Corre-sponding AOO population distributions at steady-state.
J.E. Baeten et al. / Water Research 146 (2018) 134e145 141
oxygen consumption. The increased oxygen penetration depth al-lows the population to survive deeper inside the granule. Theincreased diffusion distance of oxygen towards the AOO is thedriver for the increased apparent half-saturation coefficient. Over-all, the modification of the population distribution (microscale;Fig. 8B) is visually more distinct than the effect on the total specificgrowth rate (macroscale; Fig. 8A), which illustrates that microscaleprocesses are not always that important for the macroscale reactorperformance.
Besides the effect of long-term changes in microbial populationdistributions caused by changes in the influent NHx concentration(Fig. 9A), the effects resulting from changes of the granule radius(B), reactor temperature (C), O2 set-point (D), influent VFA (E) andinfluent PO4 (F) were investigated as well (Figure S6). When thegranule radius increases (Fig. 9B), the apparent half-saturationcoefficient increases. This trend had already been demonstratedwith static biofilm models, which assume a predefined microbialpopulation distribution (Manser et al., 2005; P�erez et al., 2005;Picioreanu et al., 2016), but this work confirms the relationshipwhen microbial dynamics are included. A higher long-term reactortemperature causes a lower apparent half-saturation coefficient(Fig. 9C). This effect is solely due to the changing AOO populationdistribution as a result of changing temperature, because the
Fig. 9. The apparent half-saturation coefficient of AOO for oxygen as a function of differentobtained. The influent NHx (A), granule radius (B), reactor temperature (C), O2 set-point (D),compared to their reference value.
coefficient itself was always determined at 20 �C (during step 2 inFig. 2). When the coefficient is determined at the same temperatureat which the new steady-state was achieved, the decrease as afunction of temperature is less steep (grey line). This is because theshort-term and long-term effect of temperature are opposite. Ashort-term temperature increase (during which the AOO popula-tion does not change) increases the oxygen consumption rate,leading to lower intragranule oxygen concentrations and thus ahigher apparent half-saturation coefficient (Figure S7), in accor-dance with Picioreanu et al. (2016). Moreover, an increasing oxygenset-point (Fig. 9D) initially decreases the apparent half-saturationcoefficient and then increases the value. An increasing influentVFA concentration (Fig. 9E) causes a small initial decrease, followedby a steep increase, while an increasing influent phosphate con-centration (Fig. 9F) decreases the coefficient first and then has anegligible effect. No prior research was found to compare thesetrends to, since no dynamic biofilm model has been used to studyhalf-saturation coefficients before.
To better understand the changes in the half-saturation coeffi-cient for different long-term operating conditions, the microbialpopulation distribution was characterised for every steady-state bytwo simple measures: the average AOO population depth (Eq. (5))and total AOO biomass per granule (Eq. (2)). A plot of the average
long-term operating conditions under which a steady-state microbial population wasinfluent VFA (E) and influent PO4 (F) were varied individually, between �30% and þ30%

J.E. Baeten et al. / Water Research 146 (2018) 134e145142
population depth versus the value of the coefficient shows a clearpositive correlation (Figure S8A; R2¼ 0.74). This confirms theintuitive idea that an apparent half-saturation coefficient is ameasure for the diffusion resistance. However, there is also a pos-itive correlation between the total amount of biomass per granuleand the half-saturation coefficient (Figure S8B; R2¼ 0.85). This canbe explained by the faster oxygen consumption and thus lowerintragranule oxygen concentrations whenmore biomass is present.This correlation was already found with a static biofilm modelassuming a homogeneous microbial population distribution (P�erezet al., 2005). During the simulations for this work, the averagedepth and size of the population changed simultaneously, butFig. 10 shows that a multiplication of both measures suffices toexplain most of the variation (R2¼ 0.96).
3.2.3. Implications for modelling with constant apparent half-saturation coefficients
Among all steady-state simulations, KO2,AOO,app varied between0.42 and 3.04 g O2.m�3 (Fig. 10). Consequently, apparent kineticsare expected to differ between plants and even for the same plant,simulations of long-term changes in operating conditions madewith constant values should be interpreted with care. And even ona shorter term, the results indicate that apparent kinetics are notconstant, because the degree of competition can fluctuate overtime, e.g. when there is a sudden increase in influent organics, thecompetition for AOO by PAO and OHO will increase. However, thisdoes not mean that apparent kinetics are never useful. First of all,the 1D biofilm model always predicted a smooth saturation curvewith increasing substrate concentrations, whatever the microbialpopulation distribution or degree of competition. Therefore, thequalitative behaviour could still be simulated, if an appropriate newset of default apparent ASM2d parameters (or similar bioconver-sion model) is found based on the average behaviour of several full-scale plants. Apparent kinetics have also been successfully used forflocculent sludge, even though they depend on the operatingconditions as well, as discussed in the introduction. Furthermore,also current 1D or 2D biofilm models for aerobic granular sludge
Fig. 10. The apparent half-saturation coefficient of AOO for oxygen obtained fordifferent long-term operating conditions as a function of the average population depthtimes the population size of AOO per granule. The distribution of all values is projectedas red lines on the y-axis. (For interpretation of the references to colour in this figurelegend, the reader is referred to the Web version of this article.)
reactors have lot of uncertainties for long-term changing condi-tions. For example, it is still challenging to accurately predictchanges in granule size distribution or microscale parameters likethe effective diffusion coefficient. Therefore, it is also not guaran-teed that these biofilm models provide reliable quantitative pre-dictions on long-term. Finally, the batch-wise operation of aerobicgranular sludge reactors allows on-line extraction of apparent ki-netics from typical monitoring data, which might enable quanti-tative predictions after all, as illustrated in the last section of thiswork.
3.3. Are apparent-half saturation coefficients practicallyapplicable?
The ammonium removal during the aeration phases of 100consecutive full-scale reactor cycles (24 days) was simulated usinga model with apparent half-saturation coefficients (Eq. (6)), fedevery cycle with parameter values that were estimated based onthe ten preceding cycles (Fig. 11). The mean absolute error indicatesthat the predictive accuracy of themodel stays quite low and stable.The average parameter values were rmax¼ 549 g N.m�3.d�1,KNHx,app¼ 1.1 g N.m�3 and KO2,app¼ 4.0 g O2.m�3, but they showedsome fluctuations, especially from cycle 80 onwards. These fluc-tuations are partly due the limited identifiability when half-saturation coefficients and maximal rates are calibrated simulta-neously (Chouakri et al., 1994), but also actual changes in apparentkinetics could have occurred. Apparent kinetics could have changeddue previously discussed reasons, such as changes in the microbialpopulation distribution or average influent organics concentration,which can alter the degree of competition. Also changes of theintrinsic ammonium half-saturationcoefficient could haveoccurred, e.g. due to a decrease in the reactor pH (Wiesmann,1994).Carstensen et al. (1995) calibrated exactly the same model for aflocculent sludge plant and found apparent half-saturation co-efficients that were lower than the ones found in this work, espe-cially for KO2,app (KNHx,app¼ 0.76 g N.m�3 and KO2,app¼ 0.71 gO2.m�3). This is in line with the expected higher diffusion resis-tance in granular sludge systems, including the bigger impact onthe oxygen-related parameter (Fig. 5).
Even though this model with apparent kinetics neglects manyphenomena, it can be useful. First of all, the fact that the ammo-nium removal could be quite accurately predicted with the modelprovides an important insight: the ammonium removal rate at anygiven time is mostly determined by the oxygen and ammoniumconcentration. This fundamental insight can be less evident frombiofilm models with dozens of parameters. Secondly, the modelgives insight in the rate limiting factors. The estimated maximalammonium removal rates (rmax) were all higher than the fastestobserved removal rate (dSNHx/dt) of 170 g N.d�1.m�3 in this reactor(Pronk et al., 2015). This indicates that the rate is limited by a toolow oxygen and/or ammonium concentration in the reactor. Themodel can be used to find the most limiting factor at any time byplotting the Monod term of oxygen and ammonium during a cycle.As such, this plant was found to be often most limited by oxygen(Figure S9), so an increased aeration capacity could further increasethe ammonium removal rate. Should ammonium be most limiting,this could be remediated by increasing the batch size to obtainhigher ammonium concentrations in the reactor during the aera-tion phase. However, the batch size cannot be too high aswell, sincefilling and discharge occur simultaneously and the dischargedliquid should not be polluted with fresh influent. Another possibleapplication of the model is on-line control. The oxygen set-pointcould be adjusted using model predictions to obtain the requiredammonium concentration within a predefined time frame.Conversely, if the aeration strategy is fixed, themodel could be used
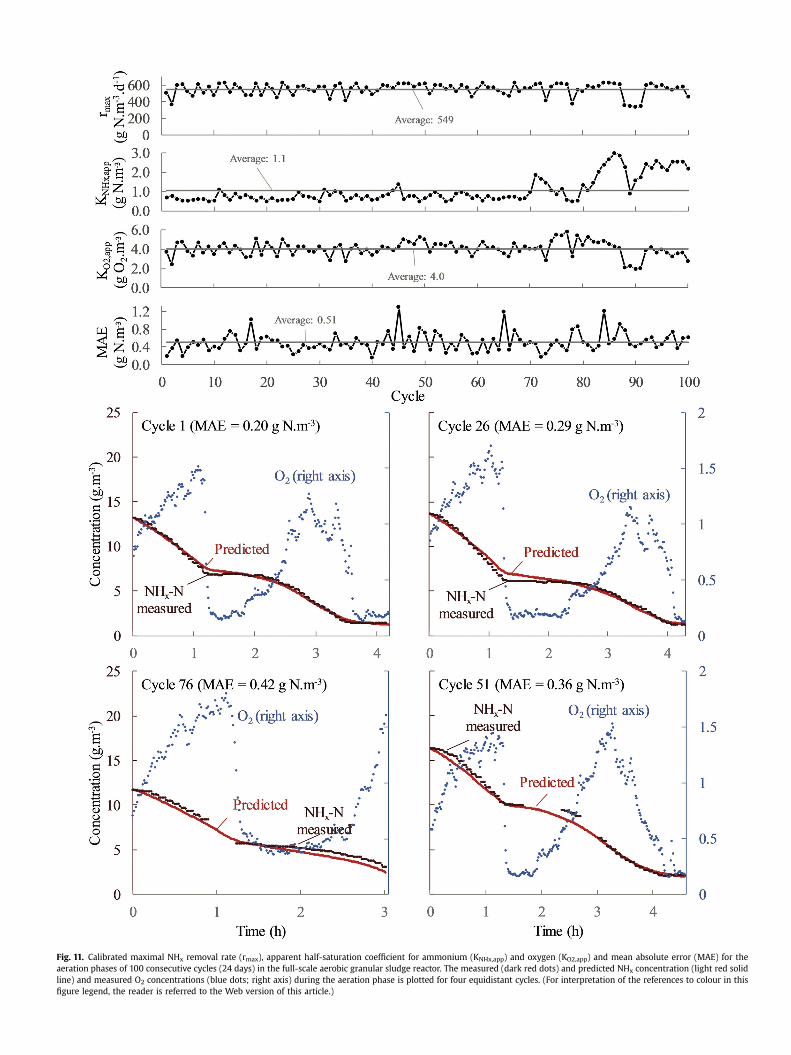
Fig. 11. Calibrated maximal NHx removal rate (rmax), apparent half-saturation coefficient for ammonium (KNHx,app) and oxygen (KO2,app) and mean absolute error (MAE) for theaeration phases of 100 consecutive cycles (24 days) in the full-scale aerobic granular sludge reactor. The measured (dark red dots) and predicted NHx concentration (light red solidline) and measured O2 concentrations (blue dots; right axis) during the aeration phase is plotted for four equidistant cycles. (For interpretation of the references to colour in thisfigure legend, the reader is referred to the Web version of this article.)

J.E. Baeten et al. / Water Research 146 (2018) 134e145144
to predict how long it will take to reach a certain ammoniumconcentration.
4. Conclusions
Since apparent kinetics that lump diffusional resistances arepopular for flocculent and anaerobic granular sludge, this workassessed the implications of using an analogous approach for aer-obic granular sludge.
� For a fixed microbial population distribution, macroscale reac-tion rates in aerobic granular sludge reactors could be approx-imated with Monod kinetics using apparent half-saturationcoefficients
� All estimated apparent half-saturation coefficients were biggerthan their respective intrinsic values, but their exact ratio variesstrongly between microbial groups and limiting substrates.
� Apparent half-saturation coefficients were influenced by theactivity of reactions that compete for the same substrates and bylong-term changes in the influent composition, granule size,oxygen set-point and temperature.
� The changes of the apparent half-saturation coefficients causedby different long-term operating conditions could be largelyexplained by the product of the changing average populationdepth and size.
� A single equation model describing ammonium removal basedon apparent kinetics could be calibrated using typical moni-toring data from a full-scale plant and offers perspectives forprocess control and identifying the rate-limiting substrate.
Acknowledgements
The doctoral research work of Janis Baeten has been financiallysupported by the Research Foundation Flanders (FWO PhDfellowship Janis Baeten). We are grateful to Royal HaskoningDHVfor the collaboration and for sharing the full-scale data.
Appendix A. Supplementary data
Supplementary data to this article can be found online athttps://doi.org/10.1016/j.watres.2018.09.025.
References
Arnaldos, M., Amerlinck, Y., Rehman, U., Maere, T., Van Hoey, S., Naessens, W.,Nopens, I., 2015. From the affinity constant to the half-saturation index: un-derstanding conventional modeling concepts in novel wastewater treatmentprocesses. Water Res. 70, 458e470. https://doi.org/10.1016/j.watres.2014.11.046.
Baeten, J.E., Batstone, D., Schraa, O., van Loosdrecht, M. and Volcke, E.I.P., unpub-lished results. Modelling Anaerobic, Aerobic and Partial Nitritation-anammoxGranular Sludge Reactors - a Review.
Baeten, J.E., van Loosdrecht, M.C.M., Volcke, E.I.P., 2017. Improving the accuracy ofgranular sludge and biofilm reactor simulations in Aquasim through artificialdiffusion. Biotechnol. Bioeng. 114 (9), 2131e2136. https://doi.org/10.1002/bit.26323.
Batstone, D.J., Keller, J., Angelidaki, I., Kalyuzhnyi, S., Pavlostathis, S., Rozzi, A.,Sanders, W., Siegrist, H., Vavilin, V., 2002. Anaerobic Digestion Model No.1(ADM1). IWA Publishing, London, UK.
Benefield, L., Molz, F., 1983. A kinetic-model for the activated sludge- process whichconsiders diffusion and reaction in the microbial flocs. Biotechnol. Bioeng. 25(11), 2591e2615. https://doi.org/10.1002/bit.260251109.
Beun, J.J., Heijnen, J.J., van Loosdrecht, M.C.M., 2001. N-removal in a granular sludgesequencing batch airlift reactor. Biotechnol. Bioeng. 75 (1), 82e92. https://doi.org/10.1002/bit.1167.
Brdjanovic, D., Meijer, S.C.F., Lopez-Vazquez, C.M., Hooijmans, C.M., VanLoosdrecht, M.C.M., 2015. Applications of Activated Sludge Models. IWA pub-lishing, London, UK. https://doi.org/10.2166/9781780404660.
Carstensen, J., Harremoes, F., Madsen, H., 1995. Statistical identification of monod-kinetic parameters from online measurements. Water Sci. Technol. 31 (2),125e133. https://doi.org/10.1016/0273-1223(95)00186-q.
Chen, Y., He, J., Mu, Y., Huo, Y.C., Zhang, Z., Kotsopoulos, T.A., Zeng, R.J., 2015.
Mathematical modeling of upflow anaerobic sludge blanket (UASB) reactors:simultaneous accounting for hydrodynamics and bio-dynamics. Chem. Eng. Sci.137, 677e684. https://doi.org/10.1016/j.ces.2015.07.016.
Chouakri, N., Fonteix, C., Marc, I., Corriou, J.P., 1994. Parameter estimation of amonod-type model. 1. Theoretical identifiability and sensitivity analysis. Bio-technol. Tech. 8 (10), 683e688. https://doi.org/10.1007/bf00151469.
Chu, K.H., van Veldhulzen, H.M., van Loosdrecht, M.C.M., 2003. Respirometricmeasurement of kinetic parameters: effect of activated sludge floc size. WaterSci. Technol. 48 (8), 61e68. https://doi.org/10.2166/wst.2003.0453.
de Bruin, L.M.M., de Kreuk, M.K., van der Roest, H.F.R., Uijterlinde, C., vanLoosdrecht, M.C.M., 2004. Aerobic granular sludge technology: an alternative toactivated sludge? Water Sci. Technol. 49 (11e12), 1e7. https://doi.org/10.2166/wst.2004.0790.
de Kreuk, M., Heijnen, J.J., van Loosdrecht, M.C.M., 2005. Simultaneous COD, ni-trogen, and phosphate removal by aerobic granular sludge. Biotechnol. Bioeng.90 (6), 761e769. https://doi.org/10.1002/bit.20470.
de Kreuk, M.K., Picioreanu, C., Hosseini, M., Xavier, J.B., van Loosdrecht, M.C.M.,2007. Kinetic model of a granular sludge SBR: influences on nutrient removal.Biotechnol. Bioeng. 97 (4), 801e815. https://doi.org/10.1002/bit.21196.
Fuentes, M., Scenna, N.J., Aguirre, P.A., 2011. A coupling model for EGSB bioreactors:hydrodynamics and anaerobic digestion processes. Chem. Eng. Process: ProcessIntensification 50 (3), 316e324. https://doi.org/10.1016/j.cep.2011.01.005.
Hamdi, M., 1995. Biofilm thickness effect on the diffusion limitation in the bio-process reaction: biofloc critical diameter significance. Bioprocess Eng. 12 (4),193e197. https://doi.org/10.1007/BF01767466.
Hauduc, H., Rieger, L., Takacs, I., Heduit, A., Vanrolleghem, P.A., Gillot, S., 2010.A systematic approach for model verification: application on seven publishedactivated sludge models. Water Sci. Technol. 61 (4), 825e839. https://doi.org/10.2166/wst.2010.898.
Henze, M., Gujer, W., Mino, T., van Loosdrecht, M.C.M., 2000. Activated SludgeModels ASM1, ASM2, ASM2d and ASM3. IWA Publishing, London, UK. https://doi.org/10.2166/9781780402369.
Jones, R.M., Dold, P.L., Tak�acs, I., Chapman, K., Wett, B., Murthy, S.,Shaughnessy, M.O., 2007. Simulation for operation and control of reject watertreatment processes. In: Proceedings of the Water Environment Federation,pp. 4357e4372. San Diego, USA. https://doi.org/10.2175/193864707787974599.
Kagawa, Y., Tahata, J., Kishida, N., Matsumoto, S., Picioreanu, C., vanLoosdrecht, M.C.M., Tsuneda, S., 2015. Modeling the nutrient removal process inaerobic granular sludge system by coupling the reactor- and granule-scalemodels. Biotechnol. Bioeng. 112 (1), 53e64. https://doi.org/10.1002/bit.25331.
Kalyuzhnyi, S., Fedorovich, V., Lens, P., 2006. Dispersed plug flow model for upflowanaerobic sludge bed reactors with focus on granular sludge dynamics. J. Ind.Microbiol. Biotechnol. 33 (3), 221e237. https://doi.org/10.1007/s10295-005-0217-2.
Lin, Y.M., Nierop, K.G.J., Girbal-Neuhauser, E., Adriaanse, M., van Loosdrecht, M.C.M.,2015. Sustainable polysaccharide-based biomaterial recovered from wasteaerobic granular sludge as a surface coating material. Sustain. Mater. Technol. 4,24e29. https://doi.org/10.1016/j.susmat.2015.06.002.
Lohani, S.P., Wang, S., Lackner, S., Horn, H., Khanal, S.N., Bakke, R., 2016. ADM1modeling of UASB treating domestic wastewater in Nepal. Renew. Energy 95,263e268. https://doi.org/10.1016/j.renene.2016.04.014.
Lübken, M., Wichern, M., Schwarzenbeck, N., Wilderer, P.A., 2005. Modellingnutrient removal of an aerobic granular sludge lab-scale SBR using ASM3. In:Bathe, S., De Kreuk, M., Mcswain, B., Schwarzenbeck, N. (Eds.), Aerobic GranularSludge, pp. 103e110 (München, Germany).
Manser, R., Gujer, W., Siegrist, H., 2005. Consequences of mass transfer effects onthe kinetics of nitrifiers. Water Res. 39 (19), 4633e4642. https://doi.org/10.1016/j.watres.2005.09.020.
Matson, J.V., Characklis, W.G., 1976. Diffusion into microbial aggregates. Water Res.10 (10), 877e885. https://doi.org/10.1016/0043-1354(76)90022-1.
Ni, B.-J., Sheng, G.-P., Li, X.-Y., Yu, H.-Q., 2010. Quantitative simulation of the gran-ulation process of activated sludge for wastewater treatment. Ind. Eng. Chem.Res. 49 (6), 2864e2873. https://doi.org/10.1021/ie901252k.
P�erez, J., Picioreanu, C., van Loosdrecht, M.C.M., 2005. Modeling biofilm and flocdiffusion processes based on analytical solution of reaction-diffusion equations.Water Res. 39 (7), 1311e1323. https://doi.org/10.1016/j.watres.2004.12.020.
Picioreanu, C., P�erez, J., van Loosdrecht, M.C.M., 2016. Impact of cell cluster size onapparent half-saturation coefficients for oxygen in nitrifying sludge and bio-films. Water Res. 106, 371e382. https://doi.org/10.1016/j.watres.2016.10.017.
Picioreanu, C., van Loosdrecht, M.C.M., Heijnen, J., 2000. Effect of diffusive andconvective substrate transport on biofilm structure formation: a two-dimensional modeling study. Biotechnol. Bioeng. 69 (5), 504e515. https://doi.org/10.1002/1097-0290(20000905)69:5<504::AID-BIT5>3.0.CO;2-S.
Pons, M.N., da Silva, M.D.L., Potier, O., Arnos, E., Battaglia, P., 2008. Modelling of anhybrid wastewater treatment plant. In: Braunschweig, B., Joulia, X. (Eds.), Eu-ropean Symposium on Computer Aided Process Engineering, pp. 907e912.Lyon, France. https://doi.org/10.1016/S1570-7946(08)80157-5.
Pronk, M., de Kreuk, M.K., de Bruin, B., Kamminga, P., Kleerebezem, R., vanLoosdrecht, M.C.M., 2015. Full scale performance of the aerobic granular sludgeprocess for sewage treatment. Water Res. 84, 207e217. https://doi.org/10.1016/j.watres.2015.07.011.
Reichert, P., 1994. AQUASIM e a tool for simulation and data analysis of aquaticsystems. Water Sci. Technol. 30 (2), 21e30. https://doi.org/10.2166/wst.1994.0025.
Rittmann, B.E., Boltz, J.P., Brockmann, D., Daigger, G.T., Morgenroth, E.,

J.E. Baeten et al. / Water Research 146 (2018) 134e145 145
Sørensen, K.H., Tak�acs, I., van Loosdrecht, M., Vanrolleghem, P.A., 2018.A Framework for Good Biofilm Reactor Modeling Practice (GBRMP). WaterScience and Technology. https://doi.org/10.2166/wst.2018.021.
Shaw, A., Takacs, I., Pagilla, K., Riffat, R., DeClippeleir, H., Wilson, C., Murthy, S., 2015.Toward universal half-saturation coefficients: describing extant K-S as a func-tion of diffusion. Water Environ. Res. 87 (5), 387e391. https://doi.org/10.2175/106143015X14212658614072.
Stewart, P.S., 1998. A review of experimental measurements of effective diffusivepermeabilities and effective diffusion coefficients in biofilms. Biotechnol. Bio-eng. 59 (3), 261e272. https://doi.org/10.1002/(SICI)1097-0290(19980805)59:3<261::AID-BIT1>3.0.CO;2-9.
Stewart, P.S., 2003. Diffusion in biofilms. J. Bacteriol. 185 (5), 1485e1491. https://doi.org/10.1128/JB.185.5.1485-1491.2003.
Su, K.-Z., Yu, H.-Q., 2006. A generalized model for aerobic granule-based sequencingbatch reactor. 1. Model development. Environ. Sci. Technol. 40 (15), 4703e4708.https://doi.org/10.1021/es060141m.
Vannecke, T.P.W., Volcke, E.I.P., 2015. Modelling microbial competition in nitrifyingbiofilm reactors. Biotechnol. Bioeng. 112 (12), 2550e2561. https://doi.org/10.1002/bit.25680.
Wanner, O., Eberl, H., Morgenroth, E., Noguera, D., Picioreanu, C., Rittmann, B.E., vanLoosdrecht, M.C.M., 2006. Mathematical Modeling of Biofilms. IWA publishing,London, UK.
Wanner, O., Gujer, W., 1986. A multispecies biofilm model. Biotechnol. Bioeng. 28(3), 314e328. https://doi.org/10.1002/bit.260280304.
Wiesmann, U., 1994. Biological nitrogen removal from wastewater. Adv. Biochem.Eng. Biotechnol./Biotechnology 51, 113e154. https://doi.org/10.1007/BFb0008736.
Winkler, M.K.H., Hong, Q.L., Volcke, E.I.P., 2015. Influence of partial denitrificationand mixotrophic growth of NOB on microbial distribution in aerobic granularsludge. Environ. Sci. Technol. 49 (18), 11003e11010. https://doi.org/10.1021/acs.est.5b01952.
Xavier, J.B., de Kreuk, M.K., Picioreanu, C., van Loosdrecht, M.C.M., 2007. Multi-scaleindividual-based model of microbial and bioconversion dynamics in aerobicgranular sludge. Environ. Sci. Technol. 41 (18), 6410e6417. https://doi.org/10.1021/es070264m.
Zhou, M., Gong, J., Yang, C., Pu, W., 2013. Simulation of the performance of aerobicgranular sludge SBR using modified ASM3 model. Bioresour. Technol. 127,473e481. https://doi.org/10.1016/j.biortech.2012.09.076.