mn(ii) regulation lignin peroxidases and manganese ... · fiechter (23) induced lignin peroxidase...
TRANSCRIPT

Vol. 56, No. 1APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Jan. 1990, p. 210-2170099-2240/90/010210-08$02.00/0Copyright © 1990, American Society for Microbiology
Mn(II) Regulation of Lignin Peroxidases and Manganese-DependentPeroxidases from Lignin-Degrading White Rot Fungi
P. BONNARMEt AND T. W. JEFFRIES*
Institute for Microbial and Biochemical Technology, Forest Products Laboratory,One Gifford Pinchot Drive, Madison, Wisconsin 53705
Received 24 July 1989/Accepted 6 October 1989
Two families of peroxidases-lignin peroxidase (LiP) and manganese-dependent lignin peroxidase (MnP)-are formed by the lignin-degrading white rot basidiomycete Phanerochaete chrysosporium and other white rotfungi. Isoenzymes of these enzyme families carry out reactions important to the biodegradation of lignin. Thisresearch investigated the regulation of LiP and MnP production by Mn(II). In liquid culture, LiP titers variedas an inverse function of and MnP titers varied as a direct function of the Mn(II) concentration. Theextracellular isoenzyme profiles differed radically at low and high Mn(II) levels, whereas other fermentationparameters, including extracellular protein concentrations, the glucose consumption rate, and the accumula-tion of cell dry weight, did not change significantly with the Mn(II) concentration. In the absence of Mn(II),extracellular LiP isoenzymes predominated, whereas in the presence of Mn(II), MnP isoenzymes weredominant. The release of "'Co2 from "4C-labeled dehydrogenative polymerizate lignin was likewise affected byMn(II). The rate of "'CO2 release increased at low Mn(II) and decreased at high Mn(II) concentrations. Thisregulatory effect of Mn(II) occurred with five strains of P. chrysosporium, two other species of Phanerochaete,three species of Phkbia, Lentinula edodes, and Phellinus pini.
Phanerochaete chrysosporium is the most extensivelystudied lignin-degrading white rot basidiomycete. It has beenused to elucidate lignin biodegradation with model com-pounds (17, 37), and it was the first organism for which ligninperoxidases (LiPs) were described (8, 21, 33, 35). Morerecently, P. chrysosporium has been shown to produce twoother types of extracellular enzymes, Mn(II)-dependent per-oxidases (2, 3, 11, 29), and glyoxal oxidase, an enzymeinvolved in extracellular H202 production (14). Both LiPsand manganese-dependent lignin peroxidases (MnPs) areglycosylated heme proteins containing protoporphyrin IX,and both are believed to play roles in the biodegradation oflignin (30).LiP and MnP differ in their catalytic mechanisms. LiP acts
by abstracting single electrons from aromatic rings of ligninand lignin model compounds, leading to the formation of acation radical (15) and subsequent cleavage reactions (10).MnP acts by generating Mn (III), a highly reactive interme-diate, which, when stabilized by chelators (7), can diffusefrom the enzyme active site to attack and oxidize the ligninstructure in situ (9, 30). LiP and MnP have been demon-strated in a number of white rot fungi (7, 20, 27, 36).
Previous studies on the lignin-degrading basidiomycete P.chrysosporium have shown the importance of nutritionalfactors in the appearance of ligninolytic activity (13, 19).Using assays based on the release of 14CO2, Keyser et al.(16) demonstrated that the ligninolytic enzyme system of P.chrysosporium is synthesized under nitrogen starvation.Nitrogen repression of ligninolytic activity has been wellstudied in P. chrysosporium (5, 6, 31), and this regulatoryeffect has been observed in other white rot fungi as well (22).Carbon source and sulfur limitations likewise induce ligni-nolytic activity in P. chrysosporium (13). Jager et al. (12) firstreported the production of lignin peroxidases in agitated
* Corresponding author.t Present address: Laboratoire de Microbiologie, INRA-PG,
78850 Thiverval-Grignon, France.
submerged cultures with various detergents, and Leisola andFiechter (23) induced lignin peroxidase production in agi-tated cultures by the addition of veratryl alcohol.Trace elements are important in the production of lignin
peroxidases by P. chrysosporium (18). An earlier studyseeking an optimum mineral nutrient balance showed thatligninolytic activity increases with decreasing Mn(II) con-centrations (13). In reassessing the micronutrient balancerequired to obtain an optimum medium for enzyme produc-tion, we noted that Linko (24) used unusually low Mn(II)concentrations in achieving high LiP activity. We thereforeinvestigated the effects of Mn(II) on LiP and MnP produc-tion in P. chrysosporium and in fungi that had been shown tobe lignin degraders (1, 28, 36).
MATERIALS AND METHODSOrganisms. The following white rot fungi were obtained
from the culture collection at the Center for Forest Mycol-ogy Research at the Forest Products Laboratory, ForestService, U.S. Department of Agriculture, Madison, Wis.:Phanerochaete chrysosporium BKM-F-1767 (ATCC 24725);Phanerochaete chrysosporium SC26 (a mutant of BKM-F-1767); Phanerochaete chrysosporium ME 446 (ATCC34541); Phanerochaete chrysosporium K3; Phanerochaetechrysosporium HHB-6251; Phanerochaete flavido alba FP106507; Phanerochaete magnoliae JHG-366-sp; Phlebia ra-diata MJL-1198-sp; Phlebia tremellosa PRL-2845; Phlebiasubserialis RLG-6074-sp; Lentinula (Lentinus) edodes RA-3-2E (ATCC 48085); Phellinus pini FP-53236-sp. Unlessotherwise noted, P. chrysosporium BKM-F-1767 was used.
Culture conditions. P. chrysosporium strains were culti-vated at 39°C, and all other fungi were cultivated at 30°C. Allthe organisms were maintained on yeast extract-malt ex-tract-peptone-glucose (YMPG) slants at 30°C. YMPG con-tained (per liter): 10 g of glucose, 10 g of malt extract, 2 g ofpeptone, 2 g of yeast extract, 1 g of asparagine, 2 g ofKH2PO4, 1 g of MgSO4 7H20, 1 mg of thiamine hydrochlo-ride, and 20 g of agar. Agitated cultures were grown in 2-liter
210
on January 1, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

LIGNIN-DEGRADING WHITE ROT FUNGI 211
Erlenmeyer flasks containing 750 ml of medium or in 125-mlErlenmeyer flasks containing 40 ml of medium. When amycelial inoculum was used, 45 ml of medium was used in a125-ml flask. The 2-liter flasks were shaken at 120 rpm(2.5-cm-diameter stroke), and the 125-ml flasks were shakenat 180 rpm.The basal medium contained (per liter) 10 g of glucose, 0.2
g of D-diammonium tartrate, 20 mM sodium tartrate (pH 4.5)(25), 2 g of KH2PO4, 0.5 g of MgSO4 7H20, 0.1 g ofCaCl2 .2H20, 1 mg of thiamine hydrochloride, 0.5 g ofTween 20, 2.5 mM veratryl alcohol (24), and 70 ml of a traceelements solution without MnSO4. This trace element solu-tion contained (per liter) 1.5 g of nitriloacetic acid, 3.0 g ofMgSO4 7H20, 1.0 g of NaCl, 0.1 g of FeSO4 7H20, 0.1 gof CoSO4, 0.1 g of CaCl2- 2H20, 0.1 g of ZnSO4- 7H20,0.01 g of CuSO4 5H20, 0.01 g of AlK(SO4)2. 12H20, 0.01g of H3BO3, and 0.01 g of Na2MoO4- 2H20. Mn(II) wasadded separately as MnSO4. H20 or MnCl2- H20 to obtainthe specified concentrations of Mn(II). Unless stated other-wise, Mn(II) was added as its sulfate form. All Mn(II)concentrations given in parts per million (milligrams perliter) are for the free ion and not the salt.
Unless stated otherwise, cultures of P. chrysosporiumBKM 1767 were inoculated with conidiospores from a 7- to10-day-old slant (5.5 x 104 conidiospores per ml final con-centration). For cultivation of the other fungi, the inoculumwas prepared from 5- to 15-day-old mycelial mats fromnonagitated cultures (75 ml volume) by blending them twicefor 15 s in a Waring blender. Mats were not washed prior toblending. From 1 to 4 ml of this mycelial homogenate wasused as the inoculum.
In the experiment designed to test the effect of Mn(II) onvarious fungi, high Mn(II) corresponded to 39.8 ppm and lowMn(II) to 0.32 ppm. The basal level of Mn(II) used was 11.15ppm, as given by Kirk et al. (18). Mn(II) was added after 48h of growth. At the same time, cultures were flushed for 1min with pure oxygen (gas flow rate, 6.5 liters/min for 125-mlflasks and 10.5 liters/min for 2-liter flasks, 760 mm Hg, 21°C)and then for 1 min every day thereafter. Each experimentwas done in triplicate and repeated at least once.To determine the isoenzyme profile, 750-ml cultures were
grown either without Mn(II) or with the basal level, andtriplicate cultures were harvested at 4 and at 6 days. Myce-lial pellets were separated from the extracellular fluid byfiltration, and the culture fluid was concentrated by ultrafil-tration with an Amicon membrane (Amicon YM 10; 10,000Mr cut-off). Fast protein liquid chromatography (FPLC)analyses of LiP and MnP isoenzymes were performed by themethod of Kirk et al. (18).Enzymatic assays. Lignin peroxidase activity was deter-
mined spectrophotometrically at room temperature by themethod of Tien and Kirk (34) with veratryl alcohol as asubstrate.
Mn(II)-dependent peroxidase activity was determinedspectrophotometrically at room temperature by the methodof Paszczynski et al. (30) with vanillylacetone as a substrate.Reaction mixtures contained 755 j.l of diluted culture fluid(50 to 100 [ul of enzyme sample plus water), 0.1 mMvanillylacetone, 100 mM sodium tartrate (pH 5), 0.1 mMMnSO4, and 0.05 mM H202 in a final volume of 1 ml. Thereaction was started by adding H202, and the rate ofdisappearance of vanillylacetone was monitored at 334 nm.The extinction coefficient used for vanillylacetone was18,300 M1 cm-1. LiP and MnP assays were performeddaily.
In the experiment designed to test the effect of Mn(II) on
{ 200
100
.5.
1000qb
~80
2400
0.__
0 50 100 150
'rue fh)
FIG. 1. Regulation of LiP and MnP production by Mn(II). (a) LiPactivity and (b) MnP activity with basal level of Mn(II) (O) andwithout Mn(II) (U).
production of MnP by various fungi, possible interferencefrom laccase was checked by using the protocol for MnP asdescribed, but without the addition of hydrogen peroxide.
Analytical. Glucose analyses were performed on a high-pressure liquid chromatography (HPLC) carbohydrate anal-ysis column with Aminex resin HPX-87C (Bio-Rad Labora-tories, Richmond, Calif.). Water was used as the eluant.Protein concentrations were determined by the Quantigoldmethod as described in the supplier's guidelines (DiversifiedBiotech, Newton Center, Mass.). Residual nitrogen concen-tration was determined with the Kjeldahl procedure (3a).Dry weight was measured by filtering and drying the pelletsovernight at 105°C.['4C]DHP degradation. In the studies of 14C-labeled dehy-
drogenative polymerizate (DHP) degradation, experimentswere done in six replicate cultures per condition. After 48 hof cultivation, [14C]DHP was added (50,600 dpm per flask).Flushing with pure oxygen was started at this time (38mi/mn, 15 min, 760 mm Hg, 21°C). This flushing wasperformed every day thereafter, and the 14CO2 released wastrapped in a scintillation cocktail (16).
RESULTS
Effect of basal Mn(II) concentration on enzymatic activity.Initial studies examined the effect of removing Mn(II) fromthe basal medium. Titers of LiP (Fig. la) and MnP (Fig. lb)were measured. In Mn(II)-free cultures, LiP activity ap-peared earlier and was 2.5 times higher than in culturesgrown under the basal conditions [11.15 ppm Mn(II)]. On theother hand, cultures grown in a medium containing noMn(II) produced only very low MnP activity (Fig. lb). At thebasal Mn(II) concentration, MnP activity was 24 timeshigher.
Nutrient nitrogen and protein concentration. With or with-out Mn(II) in the culture medium, nutrient nitrogen wasconsumed at the same rate, and essentially all nitrogen was
VOL. 56, 1990
on January 1, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
212 BONNARME AND JEFFRIES
tS
w-S.b..
2
2
vv2vw
30
_
205
Go
10a
r.toPS
Time (h)
FIG. 2. Residual nitrogen and extracellular protein during en-
zyme production. Residual nitrogen was assayed with basal level ofMn(II) ([1) and without Mn(II) (-); extracellular protein was as-
sayed with basal level of Mn(II) (0) and without Mn(II) (0).
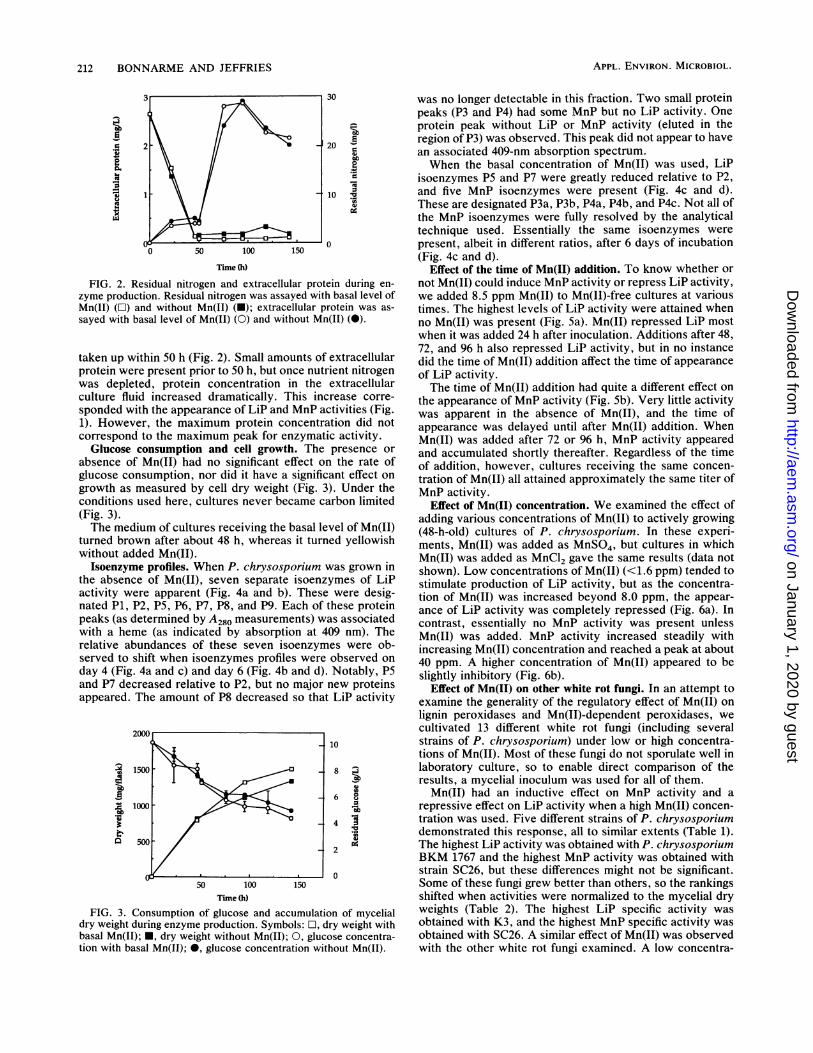
taken up within 50 h (Fig. 2). Small amounts of extracellularprotein were present prior to 50 h, but once nutrient nitrogenwas depleted, protein concentration in the extracellularculture fluid increased dramatically. This increase corre-
sponded with the appearance of LiP and MnP activities (Fig.1). However, the maximum protein concentration did notcorrespond to the maximum peak for enzymatic activity.
Glucose consumption and cell growth. The presence or
absence of Mn(II) had no significant effect on the rate ofglucose consumption, nor did it have a significant effect on
growth as measured by cell dry weight (Fig. 3). Under theconditions used here, cultures never became carbon limited(Fig. 3).The medium of cultures receiving the basal level of Mn(II)
turned brown after about 48 h, whereas it turned yellowishwithout added Mn(II).
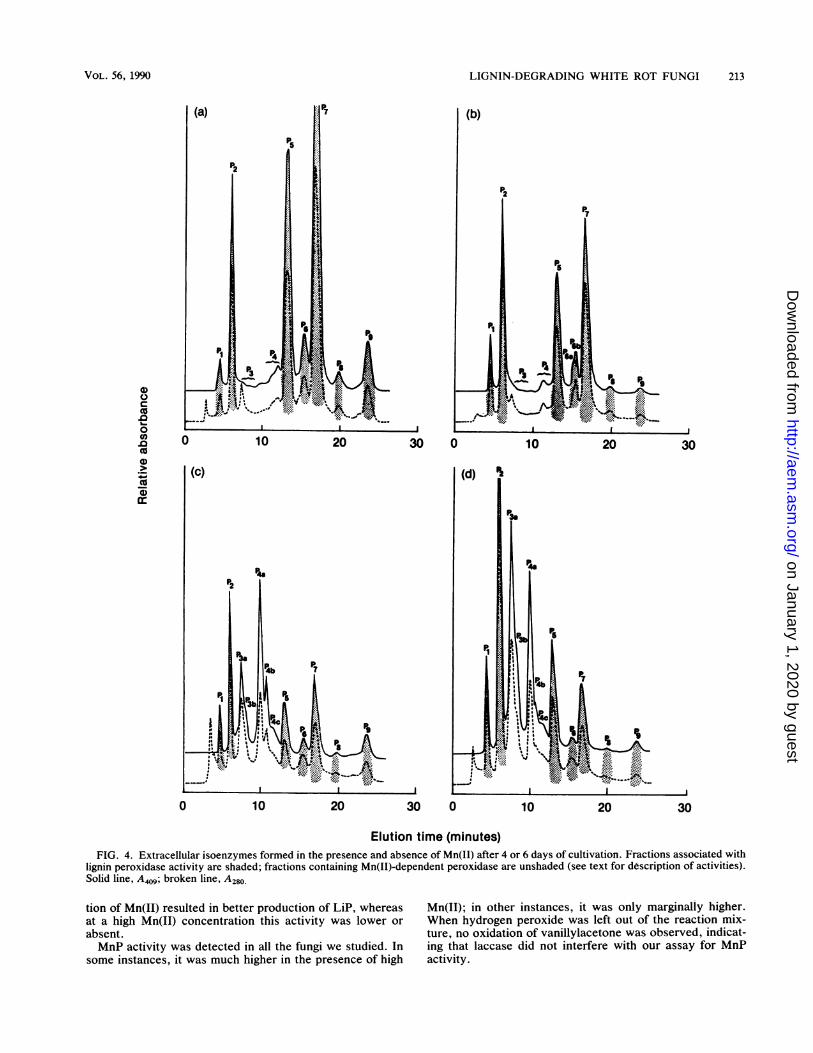
Isoenzyme profiles. When P. chrysosporium was grown inthe absence of Mn(II), seven separate isoenzymes of LiPactivity were apparent (Fig. 4a and b). These were desig-nated P1, P2, P5, P6, P7, P8, and P9. Each of these proteinpeaks (as determined by A280 measurements) was associatedwith a heme (as indicated by absorption at 409 nm). Therelative abundances of these seven isoenzymes were ob-served to shift when isoenzymes profiles were observed on
day 4 (Fig. 4a and c) and day 6 (Fig. 4b and d). Notably, P5and P7 decreased relative to P2, but no major new proteinsappeared. The amount of P8 decreased so that LiP activity
10
8 jw
4 0
.H
2
0
50 100 150
Time (h)
FIG. 3. Consumption of glucose and accumulation of mycelialdry weight during enzyme production. Symbols: 0, dry weight withbasal Mn(II); *, dry weight without Mn(II); 0, glucose concentra-tion with basal Mn(II); 0, glucose concentration without Mn(II).
was no longer detectable in this fraction. Two small proteinpeaks (P3 and P4) had some MnP but no LiP activity. Oneprotein peak without LiP or MnP activity (eluted in theregion of P3) was observed. This peak did not appear to havean associated 409-nm absorption spectrum.When the basal concentration of Mn(II) was used, LiP
isoenzymes P5 and P7 were greatly reduced relative to P2,and five MnP isoenzymes were present (Fig. 4c and d).These are designated P3a, P3b, P4a, P4b, and P4c. Not all ofthe MnP isoenzymes were fully resolved by the analyticaltechnique used. Essentially the same isoenzymes werepresent, albeit in different ratios, after 6 days of incubation(Fig. 4c and d).
Effect of the time of Mn(II) addition. To know whether ornot Mn(II) could induce MnP activity or repress LiP activity,we added 8.5 ppm Mn(II) to Mn(II)-free cultures at varioustimes. The highest levels of LiP activity were attained whenno Mn(II) was present (Fig. 5a). Mn(II) repressed LiP mostwhen it was added 24 h after inoculation. Additions after 48,72, and 96 h also repressed LiP activity, but in no instancedid the time of Mn(II) addition affect the time of appearanceof LiP activity.The time of Mn(II) addition had quite a different effect on
the appearance of MnP activity (Fig. 5b). Very little activitywas apparent in the absence of Mn(II), and the time ofappearance was delayed until after Mn(II) addition. WhenMn(II) was added after 72 or 96 h, MnP activity appearedand accumulated shortly thereafter. Regardless of the timeof addition, however, cultures receiving the same concen-tration of Mn(II) all attained approximately the same titer ofMnP activity.
Effect of Mn(II) concentration. We examined the effect ofadding various concentrations of Mn(II) to actively growing(48-h-old) cultures of P. chrysosporium. In these experi-ments, Mn(II) was added as MnSO4, but cultures in whichMn(II) was added as MnCl2 gave the same results (data notshown). Low concentrations of Mn(II) (<1.6 ppm) tended tostimulate production of LiP activity, but as the concentra-tion of Mn(II) was increased beyond 8.0 ppm, the appear-ance of LiP activity was completely repressed (Fig. 6a). Incontrast, essentially no MnP activity was present unlessMn(II) was added. MnP activity increased steadily withincreasing Mn(II) concentration and reached a peak at about40 ppm. A higher concentration of Mn(II) appeared to beslightly inhibitory (Fig. 6b).
Effect of Mn(II) on other white rot fungi. In an attempt toexamine the generality of the regulatory effect of Mn(II) onlignin peroxidases and Mn(II)-dependent peroxidases, wecultivated 13 different white rot fungi (including severalstrains of P. chrysosporium) under low or high concentra-tions of Mn(II). Most of these fungi do not sporulate well inlaboratory culture, so to enable direct comparison of theresults, a mycelial inoculum was used for all of them.
Mn(II) had an inductive effect on MnP activity and arepressive effect on LiP activity when a high Mn(II) concen-tration was used. Five different strains of P. chrysosporiumdemonstrated this response, all to similar extents (Table 1).The highest LiP activity was obtained with P. chrysosporiumBKM 1767 and the highest MnP activity was obtained withstrain SC26, but these differences might not be significant.Some of these fungi grew better than others, so the rankingsshifted when activities were normalized to the mycelial dryweights (Table 2). The highest LiP specific activity wasobtained with K3, and the highest MnP specific activity wasobtained with SC26. A similar effect of Mn(II) was observedwith the other white rot fungi examined. A low concentra-
APPL. ENVIRON. MICROBIOL.
.9
D
on January 1, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
LIGNIN-DEGRADING WHITE ROT FUNGI 213
(b)
10 20 30 0 10 20 30
(d)
p7
10 20 30 0 10 20 30
Elution time (minutes)FIG. 4. Extracellular isoenzymes formed in the presence and absence of Mn(II) after 4 or 6 days of cultivation. Fractions associated with
lignin peroxidase activity are shaded; fractions containing Mn(II)-dependent peroxidase are unshaded (see text for description of activities).Solid line, A409; broken line, A280.
tion of Mn(II) resulted in better production of LiP, whereasat a high Mn(II) concentration this activity was lower orabsent.MnP activity was detected in all the fungi we studied. In
some instances, it was much higher in the presence of high
Mn(II); in other instances, it was only marginally higher.When hydrogen peroxide was left out of the reaction mix-ture, no oxidation of vanillylacetone was observed, indicat-ing that laccase did not interfere with our assay for MnPactivity.
0)
.00coDoenCo0)
co01)
0
(C)
0
VOL. 56, 1990
on January 1, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

214 BONNARME AND JEFFRIES
I
.1
0
I
-J
0
.5
U,U)3
EUW'E
U,U)4U,
EUEU
600 -
400
200
00 50 100 150 200
Time (h)
FIG. 5. Effect of the time of Mn(II) addition on the production ofLiP activity (a) and MnP activity (b) at 24 h (U), 48 h (O), 72 h (0),and 96 h (0) or after no addition (A).
Effect of Mn(II) on 14Co2 release. We investigated theeffect of Mn(II) on ligninolytic activity as determined by therelease of 14CO2 from 14C-labeled DHP. As shown in Fig. 7,Mn(II) strongly repressed ligninolytic activity in agitatedcultures of P. chrysosporium grown under nitrogen limita-tion. Ligninolytic activity of P. chrysosporium was morethan seven times higher with low Mn(II) than with highMn(II). High LiP and low MnP activities were measured incultures grown under low Mn(II). High MnP activity but noLiP activity was detected in the high-Mn(II) cultures. There-fore, low Mn(II) correlated with low MnP, high LiP, and ahigh rate of 14CO2 release, while high Mn(II) correlated withhigh MnP, low LiP, and a low rate of 14CO2 release.
DISCUSSION
These results show that in nitrogen-limited cultures,Mn(II) plays an important role in regulating the appearanceof LiP and MnP activity in several different white rot fungiand that Mn(II) can also regulate the overall mineralizationof [14C]DHP lignin. Because Mn(II) is a substrate for MnP,the effect could be substrate-level induction, but this needsfurther study. Unlike the previously reported effects ofnitrogen, carbon, and sulfur, Mn(II) affects the cell in a
relatively specific manner. The production of mycelial dryweight and extracellular protein is not affected, nor are therates of consumption of carbon and nitrogen sources.The effect of Mn(II) is also relatively easy to demonstrate.
Sulfur limitation, for example, is effective only over a
'E
.
I2
bO.5
.C
E'aU0
or
U,
Go
U,laEU4
Time (h)FIG. 6. Effect of the concentration of Mn(II) on the production
of LiP activity (a) and MnP activity (b) with no Mn(II) (U), 0.3 ppm(O), 1.6 ppm (0), 8.0 ppm (0), 40.0 ppm (A), and 199.0 ppm (A).
narrow range and only advances the time of onset ofligninolytic activity. Carbon limitation effects a rapid onset,but the resulting mycelial mat is so frail that the burst ofactivity lasts only a few days and the mycelia appear to beautocatabolized (13).
Nitrogen limitation has often been used to induce ligni-nolytic activity. The onset of lignin biodegradation undernitrogen limitation is delayed by about 24 h following nitro-gen depletion (13). During this time, a rapid turnover ofnitrogen occurs (5). Lignin biodegradation can be sustainedas long as carbohydrate is present, but it is very difficult tomaximize extracellular enzyme production while restrictingavailable nitrogen.Both carbohydrate and nitrogen regulation of lignin bio-
degradation by white rot fungi are generalized metabolicresponses that might be mediated through a secondaryeffector. The regulation by Mn(II) appears to be morespecific. No loss of mycelial dry weight occurs, and differenteffects are observed with different components of the lignin-degrading enzyme system. Indeed, Mn(II) could be a pow-erful tool to study LiP and MnP regulation.Our results show that when Mn(II) is low, mineralization
of DHP proceeds rapidly, whereas when Mn(II) is high,mineralization of DHP is slower. The results suggest thatMnP activity is not rate limiting in DHP mineralization,because 14CO2 production was highest when MnP waslowest and vice versa. Previous studies have shown that LiPactivity and ligninolytic activity (DHP degradation) aresimultaneous (20), and our results are in agreement.
APPL. ENVIRON. MICROBIOL.
on January 1, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

LIGNIN-DEGRADING WHITE ROT FUNGI 215
TABLE 1. Effect of Mn(II) on the titer of extracellular LiPs andMnPs produced by selected white rot fungia
Activity (nmol/min per ml)
Species and Lip MnPstrain
Low High Low HighMn(II) Mn(II) Mn(II) Mn(II)
P. chrysosporiumBKM 251 (7)b 0 86 (8) 1,789 (6)SC26 233 (7) 0 95 (7) 1,915 (6)ME446 208 (7) 0 157 (7) 1,690 (7)K3 197 (6) 0 230 (6) 1,548 (5)HHB6251 63 (8) 0 29 (7) 696 (7)
Lentinula edodes 217 (9) 0 40 (9) 1,146 (9)Phanerochaeteflavido alba 25 (9) 0 324 (6) 781 (6)Phanerochaete magnoliae 22 (9) 0 46 (9) 79 (9)Phellinus pini 27 (8) 0 25 (8) 90 (9)Phlebia radiata 33 (9) 0 167 (7) 835 (7)Phlebia subserialis 0 0 26 (9) 146 (9)Phlebia tremellosa 0 0 20 (10) 56 (10)
a All experiments were performed in triplicate, and the standard deviationwas less than 10%o of the mean.
b Numbers in parentheses indicate the time of maximum activity (in days).
We noted in comparing the time courses for extracellularprotein accumulation with those for enzymatic activity thatenzyme activity lagged behind protein accumulation. Thedelayed appearance of enzymatic activity might mean thatonce excreted, the enzymes need a "maturation period" tobecome active. We do not know whether extracellularprocessing occurs.Our observations on extracellular isoenzymes agree fairly
well with those of other researchers. In cultures of BKMgrown with Mn(II), we separated five MnPs and six to sevenLiPs. These results are in good accordance with previousstudies by Kirk et al. (18) (Table 3). In cultures grownwithout Mn(II) we observed one to two minor MnPs andseven LiPs. These results are consistent with recent work
TABLE 2. Effect of Mn(II) on the specific activity ofextracellular LiPs and MnPs produced by selected white rot fungi
Sp acta (nmol/min per mg[dry wt] of mycelia)
Species and LiP MnPstrain
Low High Low HighMn(II) Mn(II) Mn(II) Mn(II)
P. chrysosporiumBKM 150 ndb 51 1,308SC26 114 nd 47 1,348ME446 103 nd 78 896K3 173 nd 202 1,144HHB6251 26 nd 12 267
Lentinula edodes 180 nd 33 691Phanerochaeteflavido alba 31 nd 397 378Phanerochaete magnoliae 20 nd 41 50Phellinus pini 24 nd 22 73Phlebia radiata 27 nd 134 432Phlebia subserialis nd nd 14 93Phlebia tremellosa nd nd 16 52
a Specific activity was determined on the day of maximum activity (seeTable 1).
b nd, Activity not detected.
~30
LI
20
0
10
100 200 300
rtme 0)
FIG. 7. Time course of '4CO2 release by P. chrysosporium at low(U) and high (l) Mn(II) levels.
published by Linko (24), in which she reported seven LiPsand two MnPs in the extracellular fluid of P. chrysosporiumgrown under glucose limitation instead of nitrogen limita-tion. As noted above, Linko also used relatively low levelsof Mn(II).Our results demonstrate that Mn(II) plays a key role in the
regulation of LiP and MnP of P. chrysosporium and otherwhite rot fungi that are effective lignin degraders in wood(28). All studies reported here were performed under condi-tions optimized for P. chrysosporium. MnP activity wasproduced by all the fungi studied. LiP was detected in mostbut not all other organisms tested. It is possible that theconditions we used were simply not appropriate for LiPproduction by these other white rot fungi.Manganese could regulate the appearance of these en-
zymes when they are growing on wood. Because of thecomplexity of native lignin and its inaccessibility in wood, itis difficult to discern which enzyme systems actually func-tion in lignin biodegradation. Our results clearly show thatLiP is formed in liquid cultures of P. chrysosporium whenMn(II) is present at less than 8 ppm. The concentration ofMn varies greatly with the wood species (26, 32, 38), butgenerally it is present in trunk wood at concentrationsranging from 20 to 200 ppm. It can be even higher in bark.We do not know whether this Mn is bound or otherwiseinaccessible or if it is available to the organism, but it ispossible that it could play a role in suppressing LiP activity.At such Mn concentrations, MnP would be induced. MnPacts by oxidizing Mn(II) to Mn(III), which then diffuses intothe wood. After a period of time, Mn might be depleted inthe region of the fungal hypha, and LiP would be dere-pressed. LiP acts directly on lignin substructures, and fol-
TABLE 3. Comparison of heme proteins from extracellularfluid of P. chrysosporium BKM with isozymes
identified in previous work
Peak no. LiP activity MnP activity Tentative(409 nm) assignment'
P1 + - HlP2 + - H2P3a,b - + H3P4a,b - + H4P4c - + H5P5 + - H6P6 + - H7P7 + - H8P8 + - H9P9 + - H10a See reference 18.
VOL. 56, 1990
on January 1, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

216 BONNARME AND JEFFRIES
lowing the activity of MnP, the lignin might be more suscep-tible to LiP attack. Wood decayed by white rot fungi containblack regions and flecks with large concentration of Mn (4),indicating that these organisms mobilize manganese inwood, so we would expect Mn to play a role in regulation insitu.
In conclusion, our results show that Mn(II) regulatesproduction of LiPs and MnPs. More work must be done todemonstrate the role of these enzymes in the actual in vivodegradation of lignin. This finding provides a very simplemeans of control over the production of either LiPs or MnPs,over the ligninolytic ability of P. chrysosporium, and overthe enzymatic activity of various other white rot fungi aswell. It might also provide a way to produce high titers ofthese enzymes in liquid culture. Our results also indicate thatMn(II) might regulate ligninolytic activity in wood.
ACKNOWLEDGMENTS
We are grateful to others at the Forest Products Laboratory(Madison), J. Popp, P. Kersten, and T.K. Kirk, for valuablediscussions, and M. Wesolowski for nitrogen analyses. P.B. thanksT. K. Kirk, Forest Products Laboratory (Madison), and G. Goma,Centre de Transfert en Biotechnologie Microbiologie (Toulouse,France), for the support necessary to complete these experiments.
LITERATURE CITED1. Agosin, E., J.-J. Daudin, and E. Odier. 1985. Screening of
white-rot fungi on (14C) whole-labelled wheat straw. Appl.Microbiol. Biotechnol. 22:132-138.
2. Anderson, L. A., V. Renganatham, A. A. Chiu, T. M. Loehr, andM. H. Gold. 1985. Spectral characterization of dairylpropaneoxygenase, a novel peroxide-dependent, lignin-degrading hemeenzyme. J. Biol. Chem. 260:6080-6087.
3. Asada, Y., M. Miyabe, M. Kikkawa, and M. Kuwahara. 1987.An extracellular NADH-oxidizing peroxidase produced by alignin-degrading basidiomycete, Phanerochaete chrysosporium.J. Ferment. Technol. 65:483-487.
3a.Association of Official Analytical Chemists. 1980. Total nitrogen,p. 14-15. Sections 2.055-2.058. Official methods of analyis, 13thed. Association of Official Analytical Chemists, Washington, D.C.
4. Blanchette, R. A. 1984. Manganese accumulation in wood de-cayed by white-rot fungi. Phytopathology 6:725-730.
5. Fenn, P., S. Choi, and T. K. Kirk. 1981. Ligninolytic activity ofPhanerochaete chrysosporium: physiology of suppression byNH4' and L-glutamate. Arch. Microbiol. 130:66-71.
6. Fenn, P., and T. K. Kirk. 1981. Relationship of nitrogen to theonset and suppression of ligninolytic activity and secondarymetabolism in Phanerochaete chrysosporium. Arch. Microbiol.130:59-65.
7. Forrester, I. T., A. C. Grabski, R. R. Burgess, and G. F.Leatham. 1988. Manganese, Mn-dependent peroxidases and thebiodegradation of lignin. Biochem. Biophys. Res. Commun.157:992-999.
8. Glenn, J. K., M. A. Morgan, M. B. Mayfield, M. Kuwahara, andM. H. Gold. 1983. An extracellular H202-requiring enzymepreparation involved in lignin biodegradation by the white-rotfungi basidiomycete Phanerochaete chrysosporium. Biochem.Biophys. Res. Commun. 114:1077-1083.
9. Gold, M. H., H. Wariishi, L. Akileswaran, Y. Mino, and T. M.Loehr. 1987. Spectral characterization of Mn-peroxidase, anextracellular heme enzyme from Phanerochaete chrysos-porium, p. 113-118. In E. Odier (ed.), Lignin enzymic andmicrobial degradation. INRA Publications, Versailles, France.
10. Hammel, K. E., M. Tien, B. Kalyanaraman, and T. K. Kirk.1985. Mechanism of oxidative CQ,-C, cleavage of a lignin modeldimer by Phanerochaete chrysosporium linginase. J. Biol.Chem. 260:8348-8353.
11. Huynh, V.-B., and R. L. Crawford. 1985. Novel extracellularenzymes (ligninases) of Phanerochaete chrysosporium. FEMSMicrobiol. Lett. 28:119-123.
12. Jager, A., S. Croan, and T. K. Kirk. 1978. Production ofligninases and degradation of lignin in agitated submergedcultures of Phanerochaete chrysosporium. Appl. Environ. Mi-crobiol. 50:1274-1278.
13. Jeifries, T. W., S. Choi, and T. K. Kirk. 1981. Nutritionalregulation of lignin degradation by Phanerochaete chrysos-porium. Appl. Environ. Microbiol. 42:290-296.
14. Kersten, P. J., and T. K. Kirk. 1987. Involvement of a newenzyme, glyoxal oxidase, in extracellular H202 production byPhanerochaete chrysosporium. J. Bacteriol. 169:2195-2201.
15. Kersten, P. J., M. Tien, B. Kalyanaraman, and T. K. Kirk. 1985.The ligninase of Phanerochaete chrysosporium generates cationradicals from methoxybenzenes. J. Biol. Chem. 260:2609-2612.
16. Keyser, P., T. K. Kirk, and J. G. Zeikus. 1978. Ligninolyticenzyme system of Phanerochaete chrysosporium synthesized inthe absence of lignin in response to nitrogen starvation. J.Bacteriol. 135:790-797.
17. Kirk, T. K., W. J. Connors, R. D. Bleam, W. F. Hackett, andJ. G. Zeikus. 1975. Preparation and microbial decomposition ofsynthetic ['4C]lignins. Proc. Natl. Acad. Sci. USA 72:2515-2519.
18. Kirk, T. K., S. Croan, M. Tien, K. E. Murtagh, and R. L.Farrel. 1985. Production of multiple ligninases by Phanerocha-ete chrysosporium: effect of selected growth conditions and useof a mutant strain. Enzyme Microb. Technol. 8:27-32.
19. Kirk, T. K., E. Schultz, W. J. Connors, L. F. Lorenz, and J. G.Zeikus. 1978. Influence of culture parameters on lignin metabo-lism by Phanerochaete chrysosporium. Arch. Microbiol. 117:277-285.
20. Kirk, T. K., M. Tien, S. C. Johnsrud, and K.-E. Eriksson. 1985.Lignin-degrading activity of Phanerochaete chrysosporiumBurds.: comparison of cellulase-negative and other strains.Enzyme Microb. Technol. 8:75-80.
21. Kirk, T. K., M. Tien, P. J. Kersten, M. D. Mozuch, and B.Kalyanaraman. 1986. Ligninase of Phanerochaete chrysos-porium: mechanism of the degradation of the non-phenolicarylglycerol 3-aryl ether substructure of lignin. Biochem. J.236:279-287.
22. Leatham, G. F., and T. K. Kirk. 1983. Regulation of ligninolyticactivity by nutrient nitrogen in white-rot basidiomycetes. FEMSMicrobiol. Lett. 16:65-67.
23. Leisola, M. S. A., and A. Fiechter. 1985. Ligninase production inagitated conditions by Phanerochaete chrysosporium. FEMSMicrobiol. Lett. 29:33-36.
24. Linko, S. 1988. Production and characterization of extracellularlignin peroxidase from immobilized Phanerochaete chrysos-porium in a 10-L bioreactor. Enzyme Microb. Technol. 10:410-417.
25. Linko, S., L. C. Zhong, M. S. A. Leisola, Y.-Y. Linko, A.Fiechter, and P. Linko. 1987. Lignin peroxidase production byimmobilized Phanerochaete chrysosporium in repeated batchshake cultures, p. 209-213. In E. Odier (ed.), Lignin enzymicand microbial degradation. INRA Publications, Versailles,France.
26. Matusiewicz, H., and R. M. Barnes. 1985. Tree ring woodanalysis after hydrogen peroxide pressure decomposition withinductively coupled plasma atomic emission spectrometry andelectrothermal vaporization. Anal. Chem. 57:406-411.
27. Niku-Paavola, M.-L., E. Karhunen, P. Salola, and V. Raunio.1988. Ligninolytic enzymes of the white-rot fungus Phlebiaradiata. Biochem. J. 254:877-884.
28. Otjen, L., R. Blanchette, M. Effland, and G. Leatham. 1987.Assessment of 30 white-rot basidiomycetes for selective lignindegradation. Holzforschung 41:343-349.
29. Paszczynski, A., V.-B. Huynh, and R. Crawford. 1985. Enzy-matic activities of an extracellular manganese-dependent perox-idase from Phanerochaete chrysosporium. FEMS Microbiol.Lett. 29:37-41.
30. Paszczynski, A., V.-B. Huynh, and R. Crawford. 1986. Compar-ison of ligninase-1 and peroxidase-M2 from the white-rot fungusPhanerochaete chrysosporium. Arch. Biochem. Biophys. 244:750-765.
31. Reid, I. D. 1983. Effects of nitrogen supplements on degradation
APPL. ENVIRON. MICROBIOL.
on January 1, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

LIGNIN-DEGRADING WHITE ROT FUNGI
of aspen wood lignin and carbohydrate components by Phaner-ochaete chrysosporium. Appl. Environ. Microbiol. 45:830-837.
32. Taneda, K., M. Ota, and M. Nagashima. 1986. The radialdistribution and concentration of several chemical elements inwoods of five Japanese species. Mokuzai Gakkaishi 32:833-841.
33. Tien, M., and T. K. Kirk. 1983. Lignin-degrading enzyme fromhymenomycete Phanerochaete chrysosporium Burds. Science221:661-663.
34. Tien, M., and T. K. Kirk. 1983. Lignin-degrading enzyme fromPhanerochaete chrysosporium: purification, characterizationand catalytic properties of a unique H202-requiring oxygenase.Proc. Natl. Acad. Sci. USA 81:2280-2284.
35. Tien, M., T. K. Kirk, C. Bull, and J. A. Fee. 1986. Steady-state
and transient-state kinetic studies on the oxidation of 3,4-dimethoxybenzyl alcohol catalyzed by the ligninase of Phaner-ochaete chrysosporium Burds. J. Biol. Chem. 261:1687-1693.
36. Waldner, R., M. S. A. Leisola, and A. Fiechter. 1988. Compar-ison of ligninolytic activities of selected white-rot fungi. Appl.Microbiol. Biotechnol. 29:400-407.
37. Weinstein, D. A., K. Krisnangkura, M. B. Mayfield, and M. D.Gold. 1980. Metabolism of radiolabeled P-guaiacyl ether-linkedlignin dimeric compounds by Phanerochaete chrysosporium.Appl. Environ. Microbiol. 39:535-540.
38. Young, H. E., and V. P. Guinn. 1966. Chemical elements incomplete mature trees of seven species in Maine. Tappi 49:190-197.
VOL. 56, 1990 217
on January 1, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from