mitochondrial fission, fusion, and stressphm.utoronto.ca/~jeffh/mitofissionfusion.pdfmitochondrial...
TRANSCRIPT

Mitochondrial Fission, Fusion,and StressRichard J. Youle1* and Alexander M. van der Bliek2*
Mitochondrial fission and fusion play critical roles in maintaining functional mitochondria when cellsexperience metabolic or environmental stresses. Fusion helps mitigate stress by mixing the contentsof partially damaged mitochondria as a form of complementation. Fission is needed to create newmitochondria, but it also contributes to quality control by enabling the removal of damaged mitochondriaand can facilitate apoptosis during high levels of cellular stress. Disruptions in these processes affectnormal development, and they have been implicated in neurodegenerative diseases, such as Parkinson’s.
Mitochondria are double-membrane–bound subcellular organelles that pro-vide a host of metabolic functions,
including energy production through oxidativephosphorylation. Mitochondrial morphologiesvary widely among different cell types. Fibro-blast mitochondria, for example, are usually longfilaments (1 to 10 mm in length with a fairlyconstant diameter of ~700 nm), whereas hepato-cyte mitochondria are more uniformly spheres orovoids. When mitochondria are viewed in livecells, it becomes immediately apparent that theirmorphologies are far from static. Their shapeschange continually through the combined actionsof fission, fusion, and motility. Rapid fission andfusion of mitochondria in cultured fibroblastsallows for the complete redistribution of mito-chondrial green fluorescent protein (GFP) fromone mitochondrion to all the other mitochon-dria of a cell within an hour. The wide range ofmitochondrial lengths observed in different celltypes and under different conditions results fromchanges in the balance between the rates of mito-chondrial fission and fusion. Here, we discusshow fission and fusion contribute to mitochon-drial quality control and the responses of mam-malian cells to stress.
Mitochondrial Fusion and Fission ProteinsMitochondrial fission and fusion processes areboth mediated by large guanosine triphosphatases(GTPases) in the dynamin family that are wellconserved between yeast, flies, and mammals(1). Their combined actions divide and fuse thetwo lipid bilayers that surround mitochondria.The mitochondrial inner membrane, which en-closes the matrix, is folded into cristae that con-tain membrane-bound oxidative phosphorylationenzyme complexes and the bulk of the solubleelectron transport proteins such as cytochrome c,whereas the smooth mitochondrial outer mem-
brane encapsulates the inner membrane and anintermembrane space.
Fission is mediated by a cytosolic dynaminfamily member (Drp1 in worms, flies, and mam-mals and Dnm1 in yeast). Drp1 is recruited fromthe cytosol to form spirals around mitochondriathat constrict to sever both inner and outer mem-branes. Yeast share with mammals this core func-tion of Drp1 but have distinct accessory proteins.Mdv1 recruits Dnm1 to mitochondrial fissionsites in yeast, whereas Mid49, Mid51, and Mffrecruit Drp1 to mitochondria in mammals (2),often at sites where mitochondria make contactwith the endoplasmic reticulum (3). Fusion be-tweenmitochondrial outermembranes ismediatedbymembrane-anchored dynamin family membersnamed Mfn1 and Mfn2 in mammals, whereasfusion between mitochondrial inner membranesis mediated by a single dynamin family membercalled Opa1 in mammals. Mitochondrial fissionand fusion machineries are regulated by proteol-ysis and posttranslational modifications (1).
Mitochondrial fission is essential for growingand dividing cells to populate themwith adequatenumbers of mitochondria. It has been less clearwhy mitochondrial fission and fusion are alsoneeded for nonproliferating cells, but the impor-tance of these processes is evident from non-proliferating neurons, which cannot survive
without mitochondrial fission, and from two hu-man diseases, dominant optic atrophy andCharcotMarie Tooth disease type 2A, which are causedby fusion defects. The importance of mitochon-drial fusion for embryogenesis was shown withMfn1 and Mfn2 knock-out mice, which die inutero at midgestation because of a placental defi-ciency, whereas the Mfn1 Mfn2 double knock-out mice die even earlier in development (4).Mouse embryo fibroblasts (MEFs) derived fromthe double knock-out mice do survive in culture,despite a complete absence of fusion, but some oftheir mitochondria display a reduced mitochon-drial DNA (mtDNA) copy number and lose mem-brane potential, causing problems with adenosinetriphosphate (ATP) synthesis (5). Mitochondrialfusion is therefore not absolutely essential for cellsurvival in vitro, but it is required for embryonicdevelopment and for cell survival at later stagesin development (4). These differential require-ments for fusion may stem from higher demandson oxidative metabolism in different cell types oron other functions that are indirectly affected byfusion, such asmitochondrial motility in neurons.
Fusion Promotes ComplementationBetween Damaged MitochondriaMitochondria have their own small circular ge-nomes, encoding select subunits of ATP synthesisand electron transport proteins that form oxida-tive phosphorylation complexes with other sub-units encoded by the nuclear genome, as well astransfer and ribosomal RNAs (tRNAs and rRNAs)needed for their translation. A single somatic cellcan have thousands of copies of these genomes,which are grouped in protein-rich complexes callednucleoids, with between one and eight genomecopies per nucleoid (6). Mutations and deletionsthat occasionally arise in mitochondrial DNAyield a heteroplasmic mixture of wild-type andmutant mitochondrial genomes within one cell.Maternal inheritance of these mutations can causemitochondrial diseases, such as mitochondrialencephalomyopathy with lactic acidosis andstrokelike episodes (MELAS) and myoclonus
REVIEW
1Biochemistry Section, Surgical Neurology Branch, National In-stitute of Neurological Disorders and Stroke, National Institutesof Health, Bethesda, MD 20892, USA. 2Department of BiologicalChemistry, David Geffen School of Medicine at University ofCalifornia–Los Angeles, Los Angeles, CA 90095, USA.*To whom correspondence should be addressed. E-mail: [email protected] (R.J.Y.); [email protected] (A.M.v.d.B.)
Damaged
Complementation of mitochondrial function by fusion
Fusion is stimulatedby energy demand
and stress
Fission generatesnew organellesand facilitatesquality control
Healthy
Fig. 1. Fusion rescues stress by allowing functional mitochondria (green) to complement dysfunctionalmitochondria (yellow) by diffusion and sharing of components between organelles. Stress-induced hyper-fusion yields maximal potential (light green), whereas under relaxed conditions cells are able to segregatethe damaged (yellow) ones.
31 AUGUST 2012 VOL 337 SCIENCE www.sciencemag.org1062
on October 30, 2017
http://science.sciencem
ag.org/D
ownloaded from

epilepsy with ragged-red fibers (MERRF). For-tunately, mitochondria with mutant DNA can stillfuse with other mitochondria in the same cell, al-lowingmitochondria withwild-type DNA to com-pensate for defects in mitochondria with mutantDNA by sharing components as long as the muta-tion load remains below 80 to 90% per cell (7, 8).Because nucleoids do not appear to exchangeDNA (6),mitochondria in heteroplasmic cells com-plement one another by sharing RNA or proteincomponents. Fusion between mitochondria canalso rescue two mitochondria with mutations indifferent genes by cross-complementation to oneanother, and it can mitigate the effects of environ-mental damage through the exchange of proteinsand lipids with other mitochondria. Mitochondrialfusion can therefore maximize oxidative capacityin response to toxic stress, as long as the stress isbelow a critical threshold (Fig. 1).
Mitochondrial Morphology Is Controlledby MetabolismRates of mitochondrial fissionand fusion respond to changesin metabolism. Mitochondriabecome more fused when theyare forced to rely on oxidativephosphorylation by withdraw-ing glucose as a carbon source(9). Increased fusionmay be nec-essary to maximize the fidelityfor oxidative phosphorylationby stimulating complementa-tion among mitochondria (Fig.1). Fusion is also enhanced bytreatments that directly or in-directly inhibit protein synthe-sis and by starvation and mTOR(mammalian target of rapamycin)inhibition–induced autophagy(10–12). Starvation-induced au-tophagy may enhance fusion byincreasing the reliance on oxida-tive phosphorylation through themetabolism of lipids and pro-teins (9). Alternatively, starvationmay evoke a specific stress response called stress-induced mitochondrial hyperfusion (10), or it mayinhibit fission to protect mitochondria from au-tophagic catabolism when they are most needed(11, 12). Each of these effects is consistent with amodel in which mitochondrial dynamics helpmaximize the capacity for oxidative phosphoryl-ation under stressful conditions (Fig. 1).
Repairing Small Amounts ofMitochondrial DamageMitochondria continually produce highly reactivesuperoxide anions as a byproduct of electron trans-port during oxidative phosphorylation. These reactiveoxygen species (ROS) damage proteins, lipids, andDNA (Box 1). Damage to proteins in the electrontransport chain may worsen the situation by produc-ing evenmoreROS (13).Mitochondria use quality-control proteases to eliminate damaged proteins (14)
and respond to unfolded protein stress in thematrixthrough transcriptional induction of chaperone ex-pression (15). Damaged mitochondrial outer mem-brane proteins alsomay be removed by the ubiquitinproteasome quality-control pathway (16). Mito-chondria respond to genotoxic damage by some,but not all, of theDNA repair pathways found in thenucleus. These proteotoxic and genotoxic damage-response pathways target individual molecules forquality control, thereby rescuingmitochondriawithminor damage without the need for altered fissionor fusion rates (14). Another level of quality controlentails the wholesale elimination of mitochondriaby autophagy, a process that is linked to mitochon-drial fission and fusion.
Scrapping Mitochondria ThatAre Beyond RepairAutophagy is a well-established mechanism tocompensate for nutrient depletion by degradingcellular components and to protect cells from del-
eterious protein aggregates by encapsulating anddegrading them. Autophagy is also required formaintaining a healthy mitochondrial network, pre-sumably by eliminating old and damaged mito-chondria (17, 18). The importance of this processis shown by the accumulation of swollen and de-fective mitochondria in hepatocytes and MEFsfrom mice lacking the key autophagy gene Ulk1(17) and the appearance of deformed mitochon-dria in hepatic cells in Atg7-deficient mice (18).
The autophagic elimination of mitochondria,mitophagy, appears to be intimately linked to mito-chondrial fission and fusion processes. A study offibroblast mitochondrial dynamics showed that onein five daughter mitochondria is depolarized andeliminatedbymitophagy (19). Inmost fission events,one daughter mitochondrion is transiently hyper-polarized while the sister mitochondrion is hypo-polarized, suggesting that fission embodies a “stress
test” that could push a daughter mitochondrion tocompletely depolarize if it functions suboptimally.Mitophagy could be prevented with a dominant-negative mutant of Drp1, suggesting that fissionis required for mitophagy (19). Photodamagedmitochondria undergo selective mitophagy (20),which is also consistent with the model that fis-sion provides a form of quality control by segregat-ing damaged parts of mitochondria and targetingthem for elimination by autophagy (Fig. 2).
Recent work on two gene products mutated infamilial Parkinson’s disease, PINK1 and Parkin,yields insight into amolecular mechanism of qual-ity control via the elimination of damaged mito-chondria (Fig. 3). The abundance of the kinasePINK1 is constitutively repressed in healthymitochondria by import into the inner mitochon-drial membrane and degradation by the rhomboidprotease PARL. When a mitochondrion becomesuncoupled, protein import to the inner mitochon-drial membrane is prevented so PINK1 is diverted
from PARL and accumulates onthe outermitochondrialmembrane.This yields a sensor of mitochon-drial damage that can flag an in-dividual impaired mitochondrionin amilieu of healthy ones. PINK1on a damaged mitochondrion,through its kinase activity, recruitsthe E3 ligase Parkin from the cy-tosol specifically to that impairedmitochondrion (Fig. 3).Once there,Parkin ubiquitinates outer mito-chondrial membrane proteins andinduces autophagic elimination ofthe flagged mitochondrion (21).
This molecular pathway fitsnicely with the fission model (19)(Fig. 2) to yield the mitochondrialquality-control model (Fig. 3).However, mitochondria have tobe severely depolarized to accu-mulate PINK1, and the degree towhich this happens physiologicallyis not clear. At least in cultured tu-mor cells that can maintain robust
ATP levels byglycolysis,mitochondrial F1F0ATPasecan cleave ATP derived from glycolysis and recon-stitute membrane potential despite the completeloss of membrane potential maintenance throughrespiration (22). Furthermore, mitochondrial fusionas discussed previously can lead to compensationfor missing components, thereby rescuing im-paired organelles. These forces would be expectedto counteract damage-induced depolarization ofmitochondria and mitigate PINK1-mediated mito-phagy. The stress test on membrane potential dur-ing fission (Fig. 2), however, might overcome thoseforces to trigger complete depolarization.
Mutations in PINK1 (23) and Parkin (24) leadto early-onset autosomal recessive Parkinson’sdisease, suggesting that defects in mitochondrialquality control could cause certain forms of par-kinsonism and supporting more general modelsthat mitochondrial dysfunction is an etiology of
Box 1. Mitochondrial Stress
Various insults can cause damage:• Environmental (radiation, toxic chemicals)• Genetic (mutations in genes for metabolic processes or repair pathways)• Spontaneous (ROS generated as byproduct of electron transport)Types of damage:• DNA• Proteins• LipidsProblems caused by damage:• Loss of metabolic functions (ATP synthesis, etc.)• More ROS made by defective mitochondria• F1F0-ATPase may, instead of making ATP, consume ATP to generate
membrane potentialCellular responses to damage:• DNA repair• Proteases• Lipases• Mitochondrial unfolded protein response• Mitophagy• Apoptosis
www.sciencemag.org SCIENCE VOL 337 31 AUGUST 2012 1063
REVIEWon O
ctober 30, 2017
http://science.sciencemag.org/
Dow
nloaded from
substantia nigral neuron degeneration. PINK1-and Parkin-deficient Drosophila display muscleand neuron degeneration that is associated withswollen and defective mitochondria (25–27).Consistent with the model that mitochondrialfission and fusion promotes mitochondrial qual-ity control, inhibition of mitochondrial fusion orpromotion of mitochondrial fission compensatesfor deficiencies of PINK1 and Parkin in flies.Furthermore, Parkin overexpression in flies res-cues unfolded protein stress of mitochondriathrough autophagy (28), and stimulation ofautophagy rescues depolarized mitochondriaaccumulation in dopaminergic neurons fromParkin-deficient Drosophila (29).
Banish Mitochondria That TrulyAre UncoupledDefective mitochondria can be toxic by generatingexcessive amounts of ROS, by consuming ATPthrough reversal of ATP synthase, and by interferingwith a host of other metabolic processes (Box 1).Low levels of damage might be corrected by com-plementation through mitochondrial fusion, butbadly damagedmitochondriawill contaminate othermitochondria if they are allowed to rejoin themitochondrial network before their eliminationby autophagy. Several mechanisms are at work tostop this from happening. A first line of defenseis provided by a built-in requirement of the mito-chondrial inner membrane fusion machinery formembrane potential (30). Vertebrates have elabo-rated on thismechanism by providing a second lineof defense through proteolytic inactivation of theinner membrane fusion dynamin OPA1. Proteoly-sis is mediated by the mitochondrial inner mem-brane protease OMA1, which is rapidly activatedby low membrane potential and low levels of ATP(31, 32). The outer membranes of these mitochon-dria can still fuse, even without functional OPA1or membrane potential, but the inner membrane–bound matrix compartments do not fuse, resulting
in several matrix compartments surrounded by acommon outer membrane, like peas in a pod.
The last line of defense is provided by the Pink1and Parkin pathway through the ubiquitination ofthe mitochondrial outer membrane fusion proteinsMfn1 and Mfn2. Ubiquitination of these proteinsleads to their extraction from themembrane by p97and their degradation by proteasomes (16). In ad-dition, Pink1 and Parkin disrupt mitochondrial mo-tility by degrading the small GTPase Miro, whichserves as an adaptor for kinesin-dependent transport
and is also needed for mitochondrial fusion (33).Ultimately, uncoupled mitochondria lose both theirinner and outermembrane fusionmachineries, there-by preventing them from fusing with and poison-ing the healthy mitochondrial network. Purposefulsegregation and disposal of damaged mitochondriathrough changes in fission and fusion pathwaysare therefore integral parts of mitochondrial quality-control mechanisms.
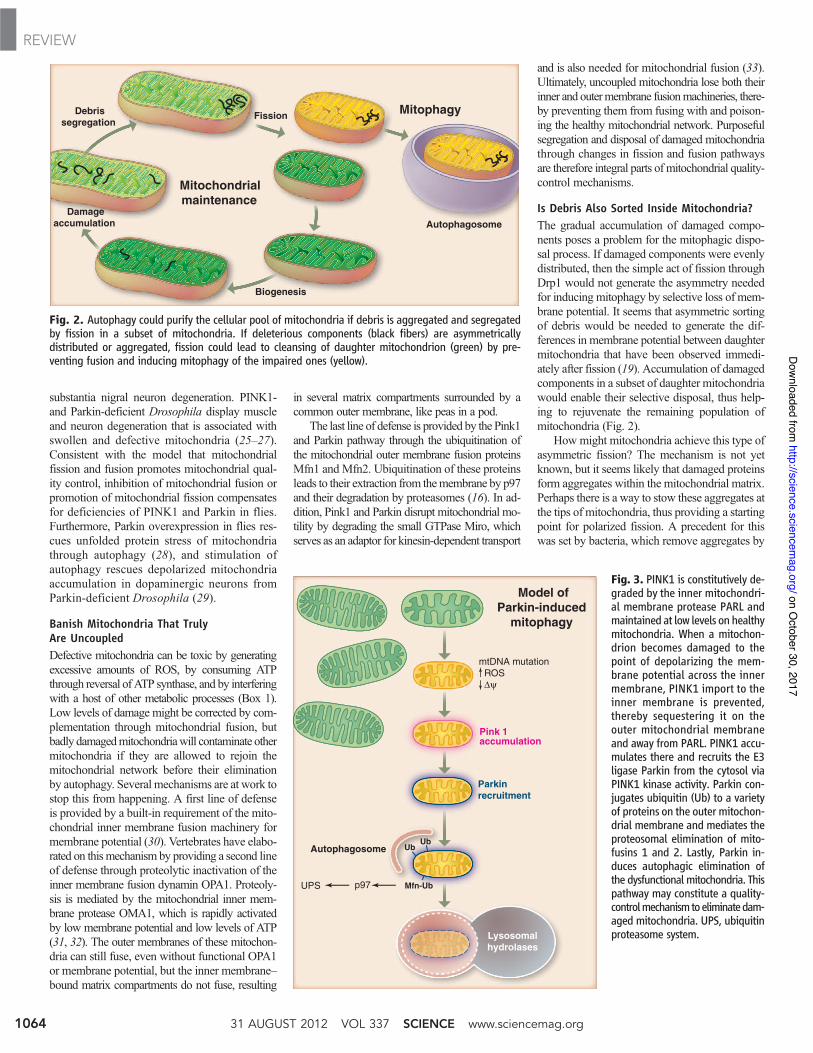
Is Debris Also Sorted Inside Mitochondria?The gradual accumulation of damaged compo-nents poses a problem for the mitophagic dispo-sal process. If damaged components were evenlydistributed, then the simple act of fission throughDrp1 would not generate the asymmetry neededfor inducing mitophagy by selective loss of mem-brane potential. It seems that asymmetric sortingof debris would be needed to generate the dif-ferences in membrane potential between daughtermitochondria that have been observed immedi-ately after fission (19). Accumulation of damagedcomponents in a subset of daughter mitochondriawould enable their selective disposal, thus help-ing to rejuvenate the remaining population ofmitochondria (Fig. 2).
How might mitochondria achieve this type ofasymmetric fission? The mechanism is not yetknown, but it seems likely that damaged proteinsform aggregates within the mitochondrial matrix.Perhaps there is a way to stow these aggregates atthe tips of mitochondria, thus providing a startingpoint for polarized fission. A precedent for thiswas set by bacteria, which remove aggregates by
Fig. 3. PINK1 is constitutively de-graded by the inner mitochondri-al membrane protease PARL andmaintained at low levels on healthymitochondria. When a mitochon-drion becomes damaged to thepoint of depolarizing the mem-brane potential across the innermembrane, PINK1 import to theinner membrane is prevented,thereby sequestering it on theouter mitochondrial membraneand away from PARL. PINK1 accu-mulates there and recruits the E3ligase Parkin from the cytosol viaPINK1 kinase activity. Parkin con-jugates ubiquitin (Ub) to a varietyof proteins on the outer mitochon-drial membrane and mediates theproteosomal elimination of mito-fusins 1 and 2. Lastly, Parkin in-duces autophagic elimination ofthe dysfunctional mitochondria. Thispathway may constitute a quality-controlmechanismtoeliminatedam-aged mitochondria. UPS, ubiquitinproteasome system.
Model ofParkin-induced
mitophagy
mtDNA mutation ROS ∆ψ
Pink 1accumulation
Parkin recruitment
Autophagosome
Lysosomalhydrolases
p97UPS
UbUb
Mfn-Ub
Debrissegregation
Damageaccumulation
Fission
Biogenesis
Mitochondrialmaintenance
Mitophagy
Autophagosome
Fig. 2. Autophagy could purify the cellular pool of mitochondria if debris is aggregated and segregatedby fission in a subset of mitochondria. If deleterious components (black fibers) are asymmetricallydistributed or aggregated, fission could lead to cleansing of daughter mitochondrion (green) by pre-venting fusion and inducing mitophagy of the impaired ones (yellow).
31 AUGUST 2012 VOL 337 SCIENCE www.sciencemag.org1064
REVIEWon O
ctober 30, 2017
http://science.sciencemag.org/
Dow
nloaded from

asymmetric fission, thus enhancing the growthrates of those daughter cells that do not receiveaggresomes (34). A similar asymmetry was ob-served during mammalian cell division, whereaggregates accumulate at the centrosome and areselectively inherited by one of the two daughtercells (35). If mitochondria also have such adeliberate mechanism, then they might have amechanism for inducing fission when too manyaggregates are formed inside mitochondria.
Such an inducing mechanism is suggested bygenetic studies showing that Pink1 and Parkin actupstream of the fission machinery inDrosophila.However, studies with mammalian cells haveonly shown effects of Pink1 and Parkin afterfission is completed. Mammalian cells may havedeveloped an additional, as yet undiscovered,mechanism to induce fission whenmitochondrialaggregates accumulate, analogous to the rapidproteolytic inactivation of the fusion machinerythrough Oma1-mediated proteolysis when mito-chondria lose membrane potential or ATP.
Aggregation of misfolded proteins in the cyto-sol is facilitated by p62 and NBR1, which can leadto their disposal by autophagy (36). Interestingly,p62 also accumulates on mitochondria after Pink1and Parkin activation. Once there, p62 triggersmitochondrial aggregation through its oligomer-ization domain (36). Mitochondrial aggregationmay be an indirect result of aggregating ubiquitin-ated proteins on the mitochondrial outer mem-brane to segregate debris before fission. Whenprotein damage accumulates, small vesicles budfrom the outermitochondrial surface. The traffickingof these vesicles to lysosomes suggests anotherand surprisingly direct pathway of mitochondrialdebris removal that is independent of Drp1,therefore independent of classic mitochondrialfission, and also independent of autophagy (37).
Selective Removal of Mutant mtDNACanmitophagy cleanse genotoxic stress in additionto proteotoxic stress? Mutations in mtDNA ac-cumulate as mammals age and could accumulategeneration after generation were it not for germlinepurification of mtDNA. Although the mechanismsare not yet known, mitochondrial genomes withstrong deleterious mutations can be removed dur-ing oogenesis (38, 39). Models for this cleansingmechanism include selective expansion of less im-paired mitochondria to populate oocytes, apopto-sis of oocytes with excessive mutant mtDNAs,and removal of poorly functioning mitochondriaby mitophagy. Whether mutated mtDNA is selec-tively removed from somatic cells is not known.
A requisite for elimination of deleterious mito-chondrial DNA mutations by mitophagy, be it inthe germ line or soma, is physical linkage betweenthe mutated mtDNA and the mutated gene product(40). Might there be a mechanism to identify mal-functioningnucleoids through their defective proteinproducts, for example, through physical associationwith protein aggregates? Integral inner membraneproteins diffuse much more slowly than solublematrix or intermembrane space proteins and there-
fore are more likely to be retained with their paren-tal nucleoid than soluble tRNAs aftermitochondrialfusion and fission events. This physical proximitymight link nucleoids with mutant gene productsthat affect protein coding sequences and facilitatetheir autophagic purification. Such differential dif-fusibility between integral membrane proteins andtRNAs might explain why mutations in tRNAs aremuch more common in human diseases than mu-tations in the integral protein components of theoxidative phosphorylationmachinery. Also, proteinaggregates may start to form immediately duringprotein synthesis, which is physically linked withnucleoids (41).Amechanism for purifyingmtDNA,by retainingmutant proteinswith their genome,mayprove to be an unexpected bonus of coupled tran-scription and translation to mitochondrial nucleoids.
Mitochondrial Fission and ApoptosisWhen all else fails, stressed cells undergo apopto-sis. In the past decade,many connections have beendiscovered between apoptosis and mitochondrialdynamics, as discussed more fully in this issue byHoppins and Nunnari. High levels of cell stressthat lead to apoptosis also lead to excessive fissionof mitochondria. This occurs almost simultane-ously with two steps of apoptosis that involvemitochondria: translocation from the cytosol tomitochondria of the pro-apoptotic Bcl-2 familymember Bax and cytochrome c release.WhenBaxtranslocates to mitochondria, it accumulates inconcentrated foci that colocalize with Drp1 andmitofusins. Inhibition of mitochondrial fission byDrp1 knock-down delays cytochrome c release,indicating that mitochondrial fission participates inBax-mediated permeabilization of the outer mito-chondrial membrane (42). The link may be thatBax is activated to oligomerize and release cyto-chrome c by membrane hemifusion intermediatesthat are formed during mitochondrial fission (43).Intriguingly, Bcl-2 familymembers also participateinmitochondrial fission and fusion in nonapoptoticcells (44). Thus, mitochondrial dynamics are in-volved not only in regulating individual mitochon-drial fidelity within cells but also at the whole-celllevel by participating in apoptotic cell death.
OutlookFusion allowsmitochondria to compensate for oneanother’s defects by sharing components and there-by helps maintain energy output in the face ofstress. However, when a certain threshold of dam-age is reached, mitochondria are eliminated whole-sale by autophagy. Fission segregates the mostseriously damaged mitochondria to preserve thehealth of the mitochondrial network in addition toregulating morphology and facilitating mitochon-drial trafficking. The highly dynamicmitochondrialfusion and fission cycle is proposed to balancetwo competing processes: compensation of dam-age by fusion and elimination of damage by fis-sion. Failure of these stress responses may lead toneuron death and neurodegenerative disorders. In-depth understanding of mitophagic processes couldaid the development of new treatments for mito-
chondrial and neurodegenerative diseases: It wasrecently shown that reactivation of autophagy canmitigate certain other diseases, such as musculardystrophies associated with mitophagy (45).
References and Notes1. S. Hoppins, L. Lackner, J. Nunnari, Annu. Rev. Biochem.
76, 751 (2007).2. K. Elgass, J. Pakay, M. T. Ryan, C. S. Palmer, Biochim.
Biophys. Acta, published online 10 May 2012.3. J. R. Friedman et al., Science 334, 358 (2011); 10.1126/
science.1207385.4. H. Chen, D. C. Chan, Ann. N. Y. Acad. Sci. 1201, 21 (2010).5. H. Chen, A. Chomyn, D. C. Chan, J. Biol. Chem. 280,
26185 (2005).6. E. A. Schon, R. W. Gilkerson, Biochim. Biophys. Acta
1800, 245 (2010).7. M. Yoneda, T. Miyatake, G. Attardi, Mol. Cell. Biol. 14,
2699 (1994).8. K. Nakada et al., Nat. Med. 7, 934 (2001).9. R. Rossignol et al., Cancer Res. 64, 985 (2004).
10. D. Tondera et al., EMBO J. 28, 1589 (2009).11. A. S. Rambold, B. Kostelecky, N. Elia, J. Lippincott-Schwartz,
Proc. Natl. Acad. Sci. U.S.A. 108, 10190 (2011).12. L. C. Gomes, G. Di Benedetto, L. Scorrano, Nat. Cell Biol.
13, 589 (2011).13. R. S. Balaban, S. Nemoto, T. Finkel, Cell 120, 483 (2005).14. M. J. Baker, T. Tatsuta, T. Langer, Cold Spring Harbor
Perspect. Biol. 3, a007559 (2011).15. A. M. Nargund, M. W. Pellegrino, C. J. Fiorese,
B. M. Baker, C. M. Haynes, Science 337, 587 (2012);10.1126/science.1223560.
16. A. Tanaka et al., J. Cell Biol. 191, 1367 (2010).17. D. F. Egan et al., Science 331, 456 (2011); 10.1126/
science.1196371.18. M. Komatsu et al., J. Cell Biol. 169, 425 (2005).19. G. Twig et al., EMBO J. 27, 433 (2008).20. I. Kim, J. J. Lemasters, Antioxid. Redox Signal. 14, 1919
(2011).21. D. Narendra, A. Tanaka, D. F. Suen, R. J. Youle,
J. Cell Biol. 183, 795 (2008).22. K. Buchet, C. Godinot, J. Biol. Chem. 273, 22983 (1998).23. E. M. Valente et al., Science 304, 1158 (2004); 10.1126/
science.1096284.24. T. Kitada et al., Nature 392, 605 (1998).25. J. C. Greene et al., Proc. Natl. Acad. Sci. U.S.A. 100,
4078 (2003).26. J. Park et al., Nature 441, 1157 (2006).27. I. E. Clark et al., Nature 441, 1162 (2006).28. I. Pimenta de Castro et al., Cell Death Differ. 19, 1308 (2012).29. J. L. Burman, S. Yu, A. C. Poole, R. B. Decal, L. Pallanck,
Proc. Natl. Acad. Sci. U.S.A. 109, 10438 (2012).30. S. Meeusen et al., Cell 127, 383 (2006).31. S. Ehses et al., J. Cell Biol. 187, 1023 (2009).32. B. Head, L. Griparic, M. Amiri, S. Gandre-Babbe,
A. M. van der Bliek, J. Cell Biol. 187, 959 (2009).33. X. Wang et al., Cell 147, 893 (2011).34. A. B. Lindner, R. Madden, A. Demarez, E. J. Stewart,
F. Taddei, Proc. Natl. Acad. Sci. U.S.A. 105, 3076 (2008).35. L. C. Fuentealba, E. Eivers, D. Geissert, V. Taelman,
E. M. De Robertis, Proc. Natl. Acad. Sci. U.S.A. 105, 7732(2008).
36. T. Johansen, T. Lamark, Autophagy 7, 279 (2011).37. V. Soubannier et al., Curr. Biol. 22, 135 (2012).38. J. B. Stewart et al., PLoS Biol. 6, e10 (2008).39. W. Fan et al., Science 319, 958 (2008).40. A. Kowald, T. B. Kirkwood, Commun. Integr. Biol. 4, 627
(2011).41. J. He et al., Nucleic Acids Res. 40, 6109 (2012).42. D. F. Suen, K. L. Norris, R. J. Youle, Genes Dev. 22,
1577 (2008).43. S. Montessuit et al., Cell 142, 889 (2010).44. A. Autret, S. J. Martin, Mol. Cell 36, 355 (2009).45. P. Grumati et al., Nat. Med. 16, 1313 (2010).
Acknowledgments: We thank members of the Youle lab forthoughtful comments. This work was supported by IntramuralProgram of the National Institute of Neurological Disordersand Stroke and grants from the NIH (GM051866) and theNSF (0552271) to A.M.v.d.B.
10.1126/science.1219855
www.sciencemag.org SCIENCE VOL 337 31 AUGUST 2012 1065
REVIEWon O
ctober 30, 2017
http://science.sciencemag.org/
Dow
nloaded from

Mitochondrial Fission, Fusion, and StressRichard J. Youle and Alexander M. van der Bliek
DOI: 10.1126/science.1219855 (6098), 1062-1065.337Science
participant in mitochondrial division and discuss how mitochondrial dynamics and cell death are linked. explain that the endoplasmic reticulum is an activeHoppins and NunnariParkinson's disease. In their Perspective,
responses to stress, interference with which are likely to play an important role in a variety of diseases including (p. 1062) review the importance of mitochondrial fusion and fission in cellularYoule and van der Bliekreactions.
mitochondria are remarkably dynamic, continually moving around the cytoplasm and undergoing fusion and fission are autonomous organelles with their own genomes. Within cells,−−the powerhouses of the cell−−Mitochondria
Mitochondrial Dynamics
ARTICLE TOOLS http://science.sciencemag.org/content/337/6098/1062
CONTENTRELATED http://science.sciencemag.org/content/sci/337/6098/1052.full
REFERENCEShttp://science.sciencemag.org/content/337/6098/1062#BIBLThis article cites 45 articles, 21 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.Sciencelicensee American Association for the Advancement of Science. No claim to original U.S. Government Works. The title Science, 1200 New York Avenue NW, Washington, DC 20005. 2017 © The Authors, some rights reserved; exclusive
(print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
on October 30, 2017
http://science.sciencem
ag.org/D
ownloaded from