mitochondria: more than just atp cows - sfrbm:...
TRANSCRIPT

Mitochondria:More Than Just ATP Cows
Paul S. Brookes, PhD.Anesthesiology, University of Rochester Medical Center
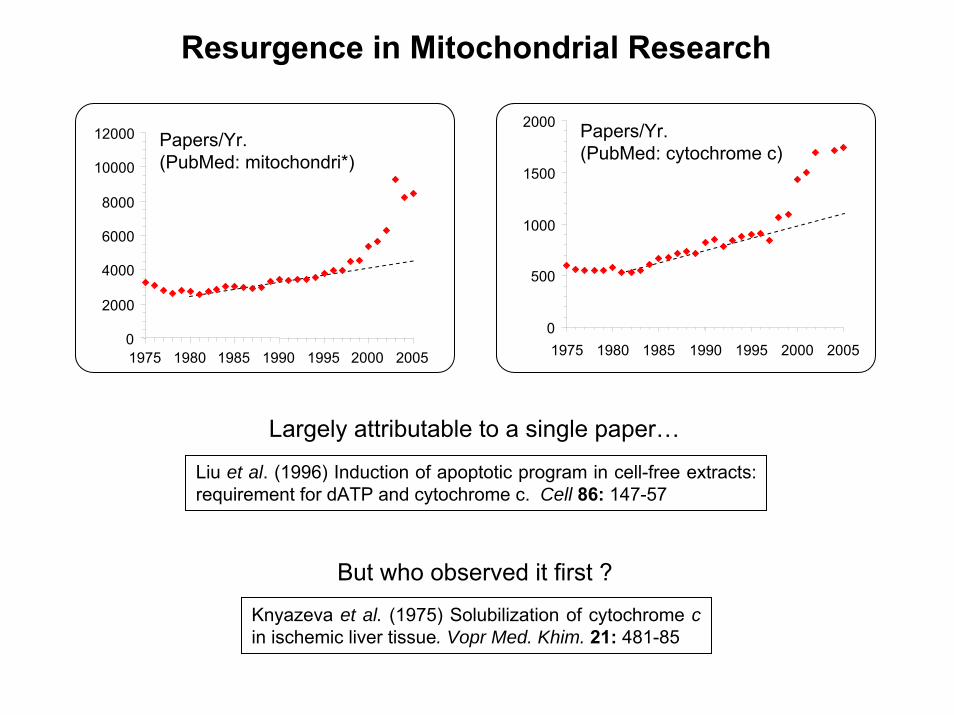
Resurgence in Mitochondrial Research
Papers/Yr.(PubMed: mitochondri*)
0
2000
4000
6000
8000
10000
12000
1975 1980 1985 1990 1995 2000 2005
0
500
1000
1500
2000
1975 1980 1985 1990 1995 2000 2005
Papers/Yr.(PubMed: cytochrome c)
Largely attributable to a single paper…
Liu et al. (1996) Induction of apoptotic program in cell-free extracts: requirement for dATP and cytochrome c. Cell 86: 147-57
Knyazeva et al. (1975) Solubilization of cytochrome cin ischemic liver tissue. Vopr Med. Khim. 21: 481-85
But who observed it first ?

“Text Book” Views of Mitochondria
1850: Mitochondria coined by microscopists, from Greek mitos: thread, chondros: grain
Frey & Mannella TIBS (2000) 25: 319-24
Updated Views - 3D electron tomography

Mitochondrial DNA (mtDNA)
Complex I II III IV VMw (~kDa) 900 140 250 200 600Subunits (mtDNA) 46(7) 4(0) 11(1) 13(3) 10(2)
Parseghian & Luhrs (2006) Biochem. Cell Biol. 84: 589-604Mitochondria contain histones…
CxICxI
CxIIICxIII
cc
CxIVCxIVCxIICxII
O2e- CxVCxV
H+
H+ H+
H+
ADPATP
YY CC NNAAWW
II
MMQQ
DDKK
GG
RR
LL
HHSS
EETT PP
Ori
12SrRNA 16SrRNA
FFVV
LL
COX1
ATP6/8
ND1
Cyt-b1D
Loop
ND2
ND3N
D4
ND5
ND6
COX2
COX3
HumanmtDNA
(16.6 kB)
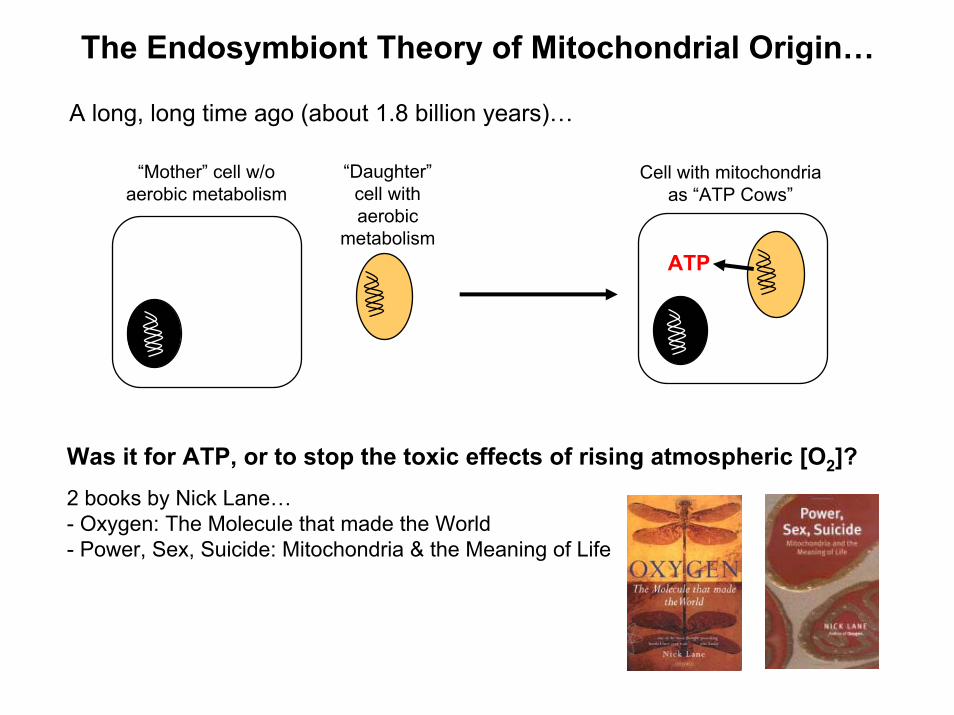
The Endosymbiont Theory of Mitochondrial Origin…
“Mother” cell w/oaerobic metabolism
“Daughter”cell withaerobic
metabolism
Cell with mitochondriaas “ATP Cows”
A long, long time ago (about 1.8 billion years)…
ATP
Was it for ATP, or to stop the toxic effects of rising atmospheric [O2]?2 books by Nick Lane…- Oxygen: The Molecule that made the World- Power, Sex, Suicide: Mitochondria & the Meaning of Life

H+ H+ H+
Q
O2 H2Oe-
e-
H+
ADP ATPFADH2
NADH
NAD+
FAD
TCACycle
H+
LEAK
Ca2+
ROSNO•
NO•
Outline

Model System: Cardiac Ischemia-Reperfusion Injury (IR)
Langendorff perfused rat heart:
0
50
100
150
10 min.
LVP
(mm
Hg)
PT Pore Opening↑ROS
↑[Ca2+]m
Murry et al. (1986) Circulation 74: 1124-36
0
50
100
150
& Ischemic Preconditioning (IPC)

Critical & Convergent Roles of NO• & Mitochondria in IPC
Diazoxide XenonVAs
plasma membrane
K+ATP
GSK-3βP
eNOS Akt
PKG(inactive)
PI3K
MAPK
PLC/PLD
GPCR
Adenosine, Angiotensin -II,Bradykinin,Acetylcholine,
Endothelin, Opioids
DAG
PDK-1
GFRs
Insulin,GF, IL-6
ETC
PKCε
Nitrite,NO• donors
Ca2+ PT
Pore
ROS
UCP
Diazoxide XenonVAs
plasma membrane
K+ATP
GSK-3βPP
eNOS AktAkt
PKG(inactive)
PI3K
MAPK
PLC/PLD
GPCR
Adenosine, Angiotensin -II,Bradykinin,Acetylcholine,
Endothelin, Opioids
DAG
PDK-1
GFRs
Insulin,GF, IL-6
ETC
PKCεPKCε
Nitrite,NO• donors
Ca2+
Ca2+ PT
Pore
ROS
UCP
mitochondrion
NO•
Jones & Bolli (2006)J. Mol. Cell. Cardiol. 40: 16-23
PDE5 Inhibitors eNOSDietary factorsACE Inhibitors
AT1 receptor blockadeNOS gene therapy
Statins, Exercise
Zaugg & Schaub (2003)J. Muscle Res. Cell Motil. 24: 219-49

Na+
H+H+
O2
ADPATP
Δψm+
–e-
Ca2+
MCUCa2+
Ca2+
Na+/Ca2+
Exch’
RaMNa+
H+
PTP
mRyRNa+/H+
Exch’
respiratorychain
Mitochondrial Ca2+ Uptake & Efflux Pathways
MCU: Mitochondrial Ca2+ Uniporter (high [Ca2+], quite slow)
RaM: Rapid Mode Ca2+ uptake (low [Ca2+], very fast)
mRyR: Mitochondrial Ryanodine Receptor
No protein identified
Found in excitable tissues

Succinate
Citrate
Isocitrate
α-KetoglutarateFumarate
Malate
Oxaloacetate
O-O
OO-
O-O
OO-
O-O
OO-
OH
O-O
OO-
O OH
OO-
O
-OO
O-
OHO
O-
O
-OO
O-
OO-
OO-
O
O
OO-
SS-CoA
OO
O-
Pyruvate
OO-
S-CoAAcetyl-CoA
SDH
MDH
CS
FH
Acon
ICDH
αKGDH
SCS
NADH, CO2
NADHCO2
NADH
NADH
GTP
FADH2
PDH
TCA Cycle… recent insights
OO-
OO-
Malonate
CO2
H2O2
H2O2
CO2
Fedotcheva et al. (2006)Free Rad. Biol Med. 41: 56-64
α-keto acids react readily with H2O2
HIF prolyl-hydroxylases use α-KG as a substrate and are inhibited by succinate
–OH
Degradation Transcription
Tumor Survival
CO2 O2PHD
Hif-1α Hif-1α
Selak et al. (2005) Cancer Cell. 7: 77-85
Succinate α-Ketoglutarate
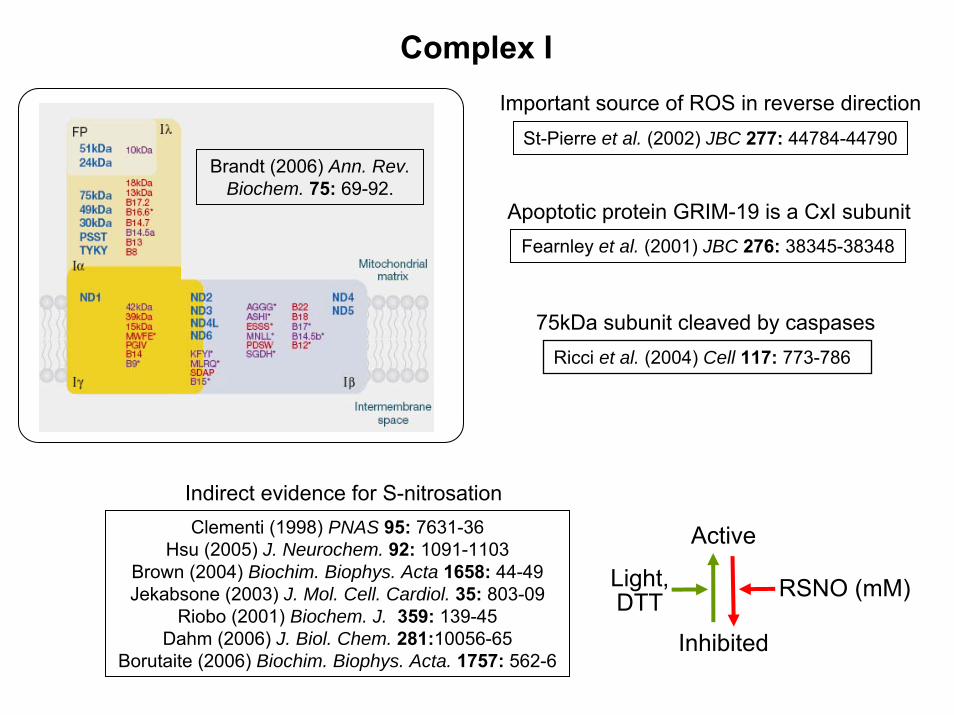
Complex IImportant source of ROS in reverse direction
St-Pierre et al. (2002) JBC 277: 44784-44790
Apoptotic protein GRIM-19 is a CxI subunitFearnley et al. (2001) JBC 276: 38345-38348
Brandt (2006) Ann. Rev.Biochem. 75: 69-92.
Clementi (1998) PNAS 95: 7631-36Hsu (2005) J. Neurochem. 92: 1091-1103
Brown (2004) Biochim. Biophys. Acta 1658: 44-49Jekabsone (2003) J. Mol. Cell. Cardiol. 35: 803-09
Riobo (2001) Biochem. J. 359: 139-45Dahm (2006) J. Biol. Chem. 281:10056-65
Borutaite (2006) Biochim. Biophys. Acta. 1757: 562-6
Indirect evidence for S-nitrosation
Inhibited
Light,DTT RSNO (mM)
Active
75kDa subunit cleaved by caspasesRicci et al. (2004) Cell 117: 773-786
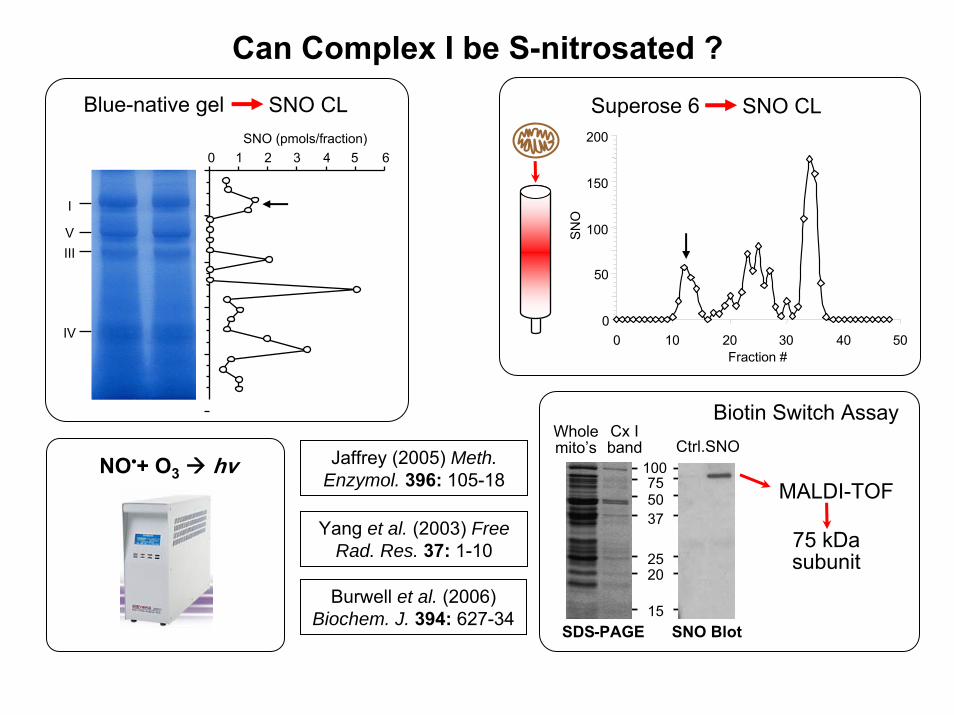
Can Complex I be S-nitrosated ?
I
VIII
IV
SNO (pmols/fraction)0 1 2 3 4 5 6
Blue-native gel SNO CL
NO•+ O3 hv
Superose 6 SNO CL
0
50
100
150
200
0 10 20 30 40 50Fraction #
SNO
SNO Blot
Ctrl.SNO
SDS-PAGE
Wholemito’s
100755037
2520
15
MALDI-TOF
Biotin Switch AssayCx Iband
75 kDasubunit
Burwell et al. (2006)Biochem. J. 394: 627-34
Yang et al. (2003) Free Rad. Res. 37: 1-10
Jaffrey (2005) Meth.Enzymol. 396: 105-18

Is Complex I S-nitrosation Physiologically Important?
0
5
10
15
20
25
IPC
pmol
s S
NO
/mg
prot
ein
Ctrl.
ND = notdetectable
ND
Ctrl. IPC
25
37
50
75100
Mitochondria from IPC hearts contain SNO
0
10
20
30
40
50
0 250 500[GSNO] (µM)
mol
SN
O/m
ol C
x I (
)
0
20
40
60
80
100
% C
ompl
ex I
Activ
ity (
)
S-nitrosation vs. Inhibition
Burwell et al. (2006) Biochem. J. 394: 627-34
Recovery from IR is enhanced in the dark
LVP
(mm
Hg)
10 min.
Light (8.7±1.7%)
Dark (24.6±2.3%)
100
0
100
0
100
0
100
00
20
40
60
80
100
Cel
l via
bilit
y (%
of c
ontro
l)
**
#*
HRIPC
GSNO(µM)
10 20 10 20
#* #
*
†#*
Nor
mox
ic C
ontro
l
mSNO(µM)
“mito-SNO” protects myocytes from IR injury
+ O
DQ
20
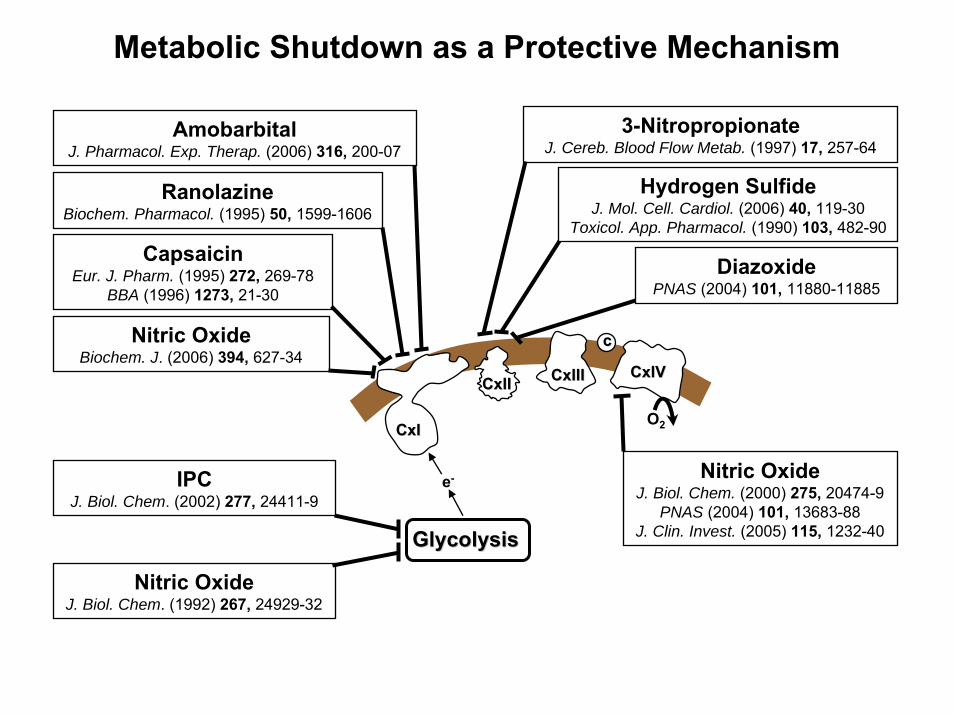
Metabolic Shutdown as a Protective Mechanism
Nitric OxideJ. Biol. Chem. (2000) 275, 20474-9
PNAS (2004) 101, 13683-88J. Clin. Invest. (2005) 115, 1232-40
Nitric OxideBiochem. J. (2006) 394, 627-34
AmobarbitalJ. Pharmacol. Exp. Therap. (2006) 316, 200-07
RanolazineBiochem. Pharmacol. (1995) 50, 1599-1606
CapsaicinEur. J. Pharm. (1995) 272, 269-78
BBA (1996) 1273, 21-30
CxICxI
CxIIICxIII
cc
CxIVCxIVCxIICxII
O2
GlycolysisGlycolysis
Nitric OxideJ. Biol. Chem. (1992) 267, 24929-32
IPCJ. Biol. Chem. (2002) 277, 24411-9
3-NitropropionateJ. Cereb. Blood Flow Metab. (1997) 17, 257-64
DiazoxidePNAS (2004) 101, 11880-11885
Hydrogen SulfideJ. Mol. Cell. Cardiol. (2006) 40, 119-30
Toxicol. App. Pharmacol. (1990) 103, 482-90
e-

Complex II
• mitoK+ATP channel opening ischemic preconditioning
• Drugs that open K+ATP (e.g. diazoxide) are Cx II inhibitors
• Cx II inhibitors (e.g. malonate) can mimic preconditioning• Splice variant of Cx II subunit 3 is within the SUR gene
PiTX
ATPK+
Ardehali et al. (2004) PNAS 101: 11880-5
Cx VCx II
ANT
mABC1InnerMembrane
Wohllk et al. (1998) Mol. Genet. Metab. 65: 187-90
Schafer et al. (1969) Biochem. Pharmacol. 18: 2678-81
Kir 6.1vs. SUR
ATPK+

H+ H+
O2
ADP
+
– CxII
FADH2
H+
Acetyl-CoA
Pyruvate
NADHNAD+
CxI
CxIII
CxIV CxV
PDH
ATP
e-
UCPH+
FAD
Q
QH2
QH•
H+
e-
cyt-c
Δψm
Q-cycle
α-KG
Citrate
Isocitrate
Succinate
OAA
CS
α-KGDH
ICDH
NADH
Enoyl-CoA
Acyl-CoA
Acyl-CoA DH
ETFQOR
Acyl-CoA -C2
NADH β-ox
Mito’ Sources of ROS – more than just CxI & CxIII
MonoamineOxidase B
DHODH
MDH
SDH
e-
Dihydro-Orotate Orotate
R–CH2–NH2 R–CH=O
H2O2
FAD FADH2
ROS Sources• Complex I• Complex III• Pyruvate Dehydrogenase• α-ketoglutarate Dehydrogenase• Acyl-CoA Dehydrogenase• Isocitrate Dehydrogenase• Dihydroorotate Dehydrogenase• β-oxidation e- Transfer Flavoprotein
(quinone oxidoreductase)• Monoamine Oxidase B

ROS Metalloproteases
Upstream signals(Ca2+, NO●, TNF etc.)
Phosphatases
PT poreapoptosis
NF-κB
CREB
Keap-1
Cell cycle,proliferation
Hif-1α
HO-1
MAPKs
Antioxidants(GSH, SOD etc)
Mitochondrial ROS & Cell Signaling

Mitochondrial ROS Generation & Hypoxia
Guzy et al. (2005) Cell Metab 1: 401-8
Bell et al. (2005) Mitochondrion 5: 322-32
Chandel et al. (2000) J. Biol. Chem. 275: 25130-8
Waypa & Schumacker (2002) Respir. Physiol. Neurobiol. 132: 81-91
Hypoxia
↑ROS Hif-1α
Current Dogma:
O2 (%)
Mito
RO
SG
ener
atio
n
Nor
mox
ia
Hyp
erox
ia
Hyp
oxia
1-5 20 50
H+
O2
H+
e-
H+
II
IIIIII
ccIVIV
Proposed Mechanism:
Q
H2O
H+
O2
H+
e-
H+
II
IIIIII
ccIVIV
QH•
NormoxiaElectrons flow all the way to O2 at complex IV, and fully
reduce it to H2O
O2O2
•-
HypoxiaLack of O2 at complex IV
causes a “back-up” of electrons in the chain, increasing lifetime of
semiquinone radical (QH•), which donates electrons
to O2, forming O2•-

Mitochondrial ROS Generation & Hypoxia
Prediction: Mitochondrial hypoxic signaling response is an inherent property of the respiratory chain. Mitochondria function autonomously in hypoxic signaling.
DCF Fluorescence
Chandel et al. (2000)J. Biol. Chem. 275: 25130-8
(results from Hep-3B cells)
Hif-1α Stabilization
But: ROS generation by isolated mitochondria at hypoxic [O2] (1% = 10μM) has never been measured.
And: kM of cytochrome c oxidase is ~1μM O2, but kM of the ROS generating system is unknown.
Probe specificity (esp. DCF)
Inhibitor specificity (rotenone, AA)
Probe location (mitoSOX Red)

Open Flow Respirometer
.
Air N2
Gas Inlet
Gas Exhaust
Stirrer motor& Stopper
StirringPropeller
O2 Electrode
HumidifierMass-flowControllers
Photomultiplier Tube
Fiber Optic Fluorimeter
O2
Cole et al. (1982) J. App. Physiol. 53: 1116-24
Brookes et al. (2003) J. Biol. Chem. 278: 31603-9
Hoffman et al. (2007) Am. J. Physiol. Heart. In-Press
Open-flow respirometry chamber: O2 consumed by mitochondria is replaced by O2 from headspace gas. This allows incubation at steady-state values of [O2] as low as 0.1uM. Simultaneous cytochrome spectral measurements inform on redox state of respiratory chain:
μM O2
% R
educ
tion
0
20
40
60
80
100
120
0 50 100 150 200 250
020406080
100120
0 2 4 6 8 100 2 4 6 8 10
Conventional respirometry chamber: [O2] is constantly changing, as mitochondria are consuming O2. Below 10% O2, the only way to maintain a relatively constant [O2] is to use very small amounts of mitochondria (imcompatible with ROS measurements).
[O2]
(μM
)
Time
All the interesting stuff happens in the last 10 sec. of the experiment!
0
200
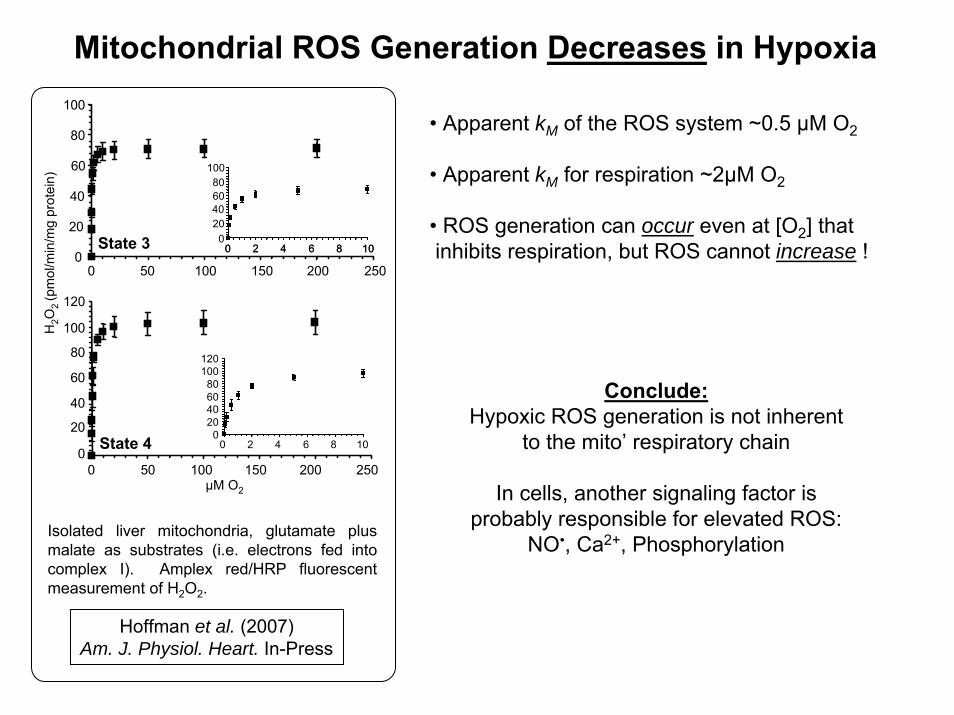
Mitochondrial ROS Generation Decreases in HypoxiaH
2O2
(pm
ol/m
in/m
g pr
otei
n)
State 3
State 40
20
40
60
80
100
120
0 50 100 150 200 250
020406080
100120
0 2 4 6 8 10
0
20
40
60
80
100
50 100 150 200 250
020406080
0 2 4 6 8 10
100
0 2 4 6 8 10
0
μM O2
• Apparent kM of the ROS system ~0.5 μM O2
• Apparent kM for respiration ~2μM O2
• ROS generation can occur even at [O2] thatinhibits respiration, but ROS cannot increase !
Isolated liver mitochondria, glutamate plus malate as substrates (i.e. electrons fed into complex I). Amplex red/HRP fluorescent measurement of H2O2.
Hoffman et al. (2007)Am. J. Physiol. Heart. In-Press
Conclude:Hypoxic ROS generation is not inherent
to the mito’ respiratory chain
In cells, another signaling factor is probably responsible for elevated ROS:
NO•, Ca2+, Phosphorylation

Ca2+
Ca2+
OxidantsAttractylosidePiRSSRATP + ADP DepletionNAD(P)+ > NAD(P)H
VDAC
ANT
Cyp-D
Hexokinase
Creatine
Kinase
Bclfamilyproteins
pH<7Mg2+
Cyclosporin ABongrekic AcidRSHATP + ADPNAD(P)H > NAD(P)+
++
– –
PBR
Permeability Transition Pore (classical view)
Brookes et al. (2004) Am. J. Physiol. 287: C817-C833
Opening of the PT pore is mechanistically linked to cytochrome crelease. Cyt-c release can occur without pore opening, andOMM swelling/rupture does not appear to play a major role

Mechanism by which cytochrome c exitsMitochondria during apoptosis is still elusive
Swelling ≠ Cytochrome c Release
Brookes et al. (2000) J. Biol. Chem. 275: 20474-9
5 min.
Ca2+
10%
ΔA
bs @
540n
m v
s. in
itial C
ytochrome C
release (% of m
aximum
)0
20
40
60
80
100
5 min.
Ca2+Ca2+
10%
ΔA
bs @
540n
m v
s. in
itial C
ytochrome C
release (% of m
aximum
)0
20
40
60
80
100
0
20
40
60
80
100
Swelling (% of maximum)
Cyt
-C R
elea
se (%
of m
axim
um)
0
20
40
60
80
100
0 20 40 60 80 100Swelling (% of maximum)
Cyt
-C R
elea
se (%
of m
axim
um)
0
20
40
60
80
100
0 20 40 60 80 1000
20
40
60
80
100
0 20 40 60 80 100

Components of the PT Pore: knockout mice data
ANT Kokoszka et al. (2004) Nature 427(6973): 461-5 Non-essential
Halestrap (2003) Curr. Med. Chem. 10: 1507-25
Maybe other MCF proteins can sustitute?Palmieri (2004) Pflugers Arch. 447: 689-709
Or maybe it is essential?
VDAC Krauskopf et al. (2006) Biochim. Biophys. Acta 1757: 590-5 Non-essential
Cyp-D Nakagawa et al. (2005) Nature 434(7033): 652-8
Baines et al. (2005) Nature 434(7033): 658-62
Essential for PT pore, but pore is only important in necrosis, not apoptosis
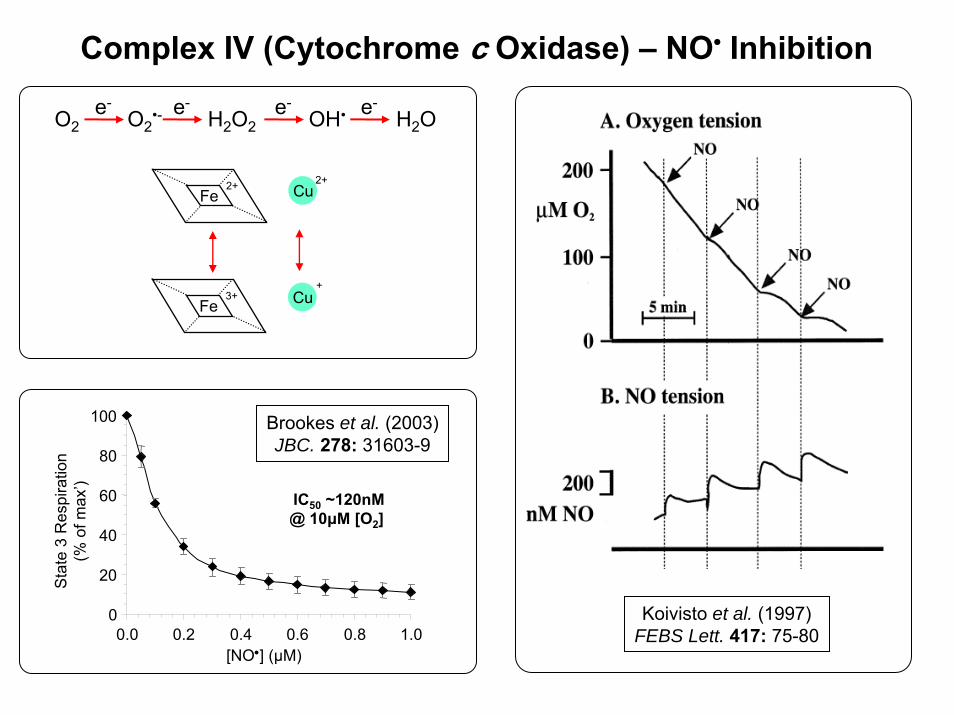
Complex IV (Cytochrome c Oxidase) – NO• Inhibition
O2 O2•- OH•H2O2 H2O
e- e- e- e-
Fe2+
Fe3+ Cu
Cu2+
+
Koivisto et al. (1997)FEBS Lett. 417: 75-80
0
20
40
60
80
100
0.0 0.2 0.4 0.6 0.8 1.0[NO•] (µM)
Sta
te 3
Res
pira
tion
(% o
f max
’)
IC50 ~120nM@ 10μM [O2]
Brookes et al. (2003)JBC. 278: 31603-9
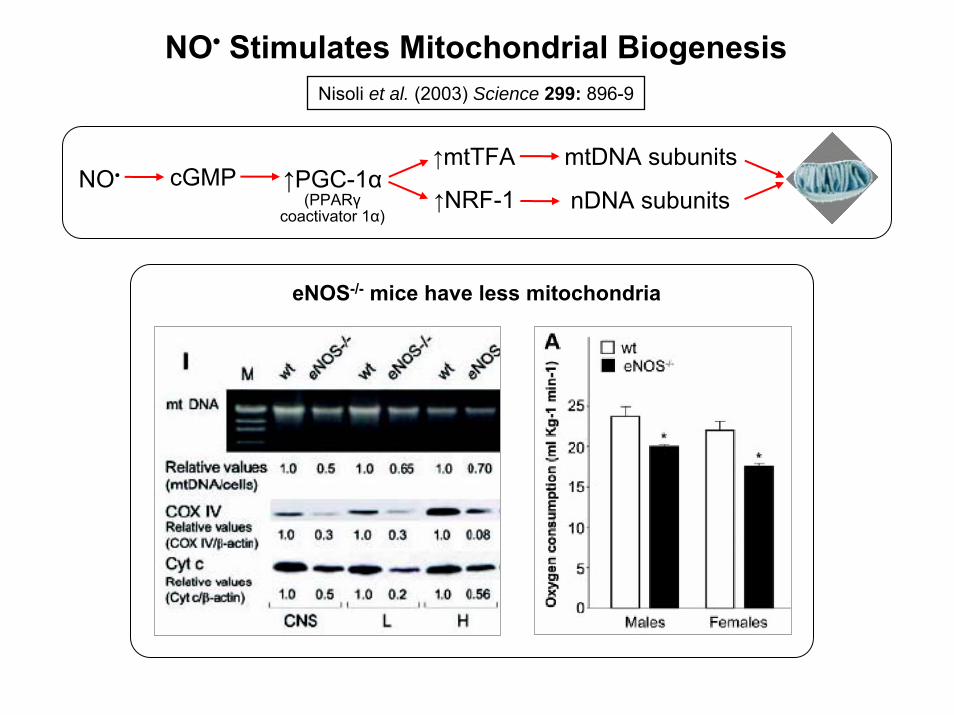
Nisoli et al. (2003) Science 299: 896-9
NO• Stimulates Mitochondrial Biogenesis
↑PGC-1α(PPARγ
coactivator 1α)↑NRF-1
↑mtTFA
nDNA subunits
mtDNA subunitsNO• cGMP
eNOS-/- mice have less mitochondria
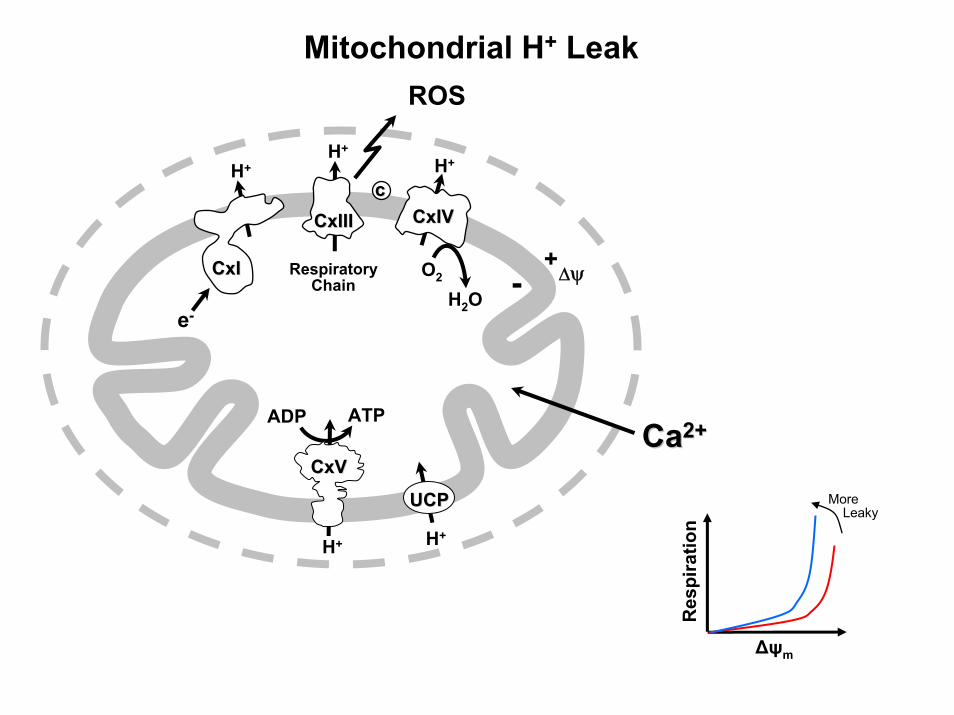
Mitochondrial H+ Leak
H+
ATPADP
RespiratoryChain
CaCa2+2+
H+
O2
H+
e-
H+
H2O
ROS
+- ΔψCxICxI
CxIIICxIIIcc
CxIVCxIV
CxVCxV
H+
UCPUCP
Δψm
Res
pira
tion
MoreLeaky
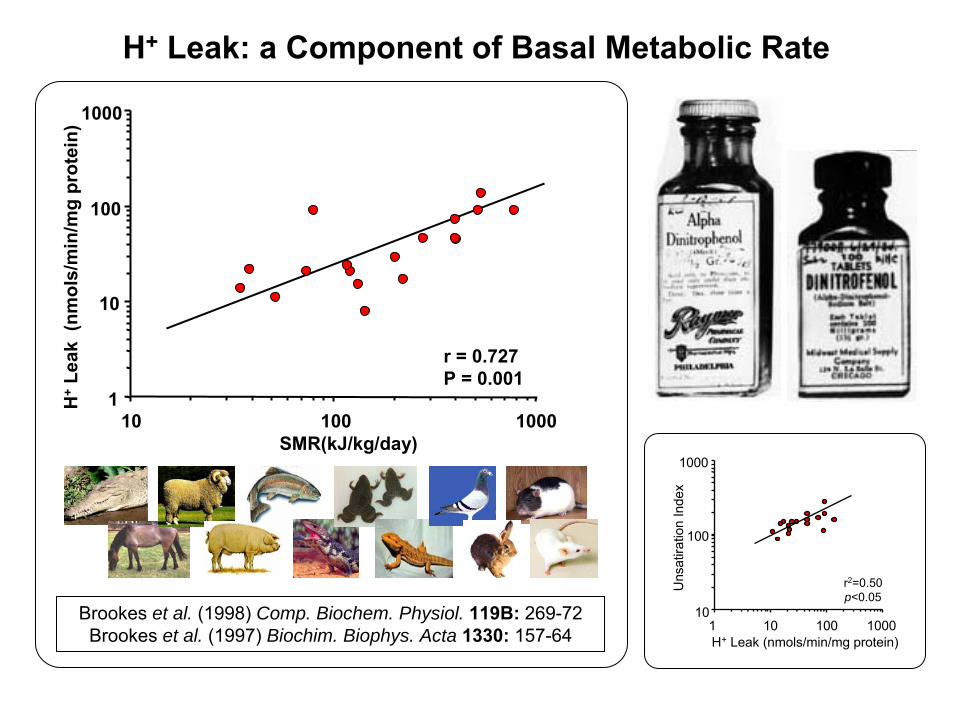
SMR(kJ/kg/day)
1
10
100
1000
10 100 1000
H+
Leak
(nm
ols/
min
/mg
prot
ein)
r = 0.727P = 0.001
Brookes et al. (1998) Comp. Biochem. Physiol. 119B: 269-72Brookes et al. (1997) Biochim. Biophys. Acta 1330: 157-64
H+ Leak: a Component of Basal Metabolic Rate
Uns
atira
tion
Inde
x
10
100
1000
1 10 100 1000H+ Leak (nmols/min/mg protein)
r2=0.50p<0.05

H+ Leak Mechanisms & Modulators
Uncouplingproteins
UCP
H+
GDP,Genipin
ROS,e- philes
Lipidbilayer
H+
Non-specificprotein boundary
H+
AMP ANTallosterism
ANT
H+AMP
CATR
TransientPT pore
ANT
VDAC
Cyp-D
CATR CsA,SfA
H+

Uncoupling Proteins: Location & Homology
UCP-2Fleury et al. (1997) Nature Genetics 15: 269-72
UCP-4
UCP-3Boss et al. (1997) FEBS Lett. 408: 39-42
UCP-1 Brown adipose “thermogenin”, 10% of protein in BAT mito’
Leukocytes, Brain, Kidney, Adipose, Sk-M, Heart
Sk-M, BAT, T-cells, Macrophages
Brain
PUMP Plants
Nicholls & Rial E (1999) J. Bioenerg. Biomembr. 31: 399-406
Bouillaud et al. (2001) Biochim. Biophys. Acta 1504: 107-19
Sanchis et al. (1998) J. Biol. Chem. 273: 34611-5 UCP-5 Brain
Jezek et al. (1996) J. Biol. Chem. 271: 32743-8
Stuart et al. (1999) Biochim. Biophys. Acta. 1413: 50-4 fishUCP
59% = UCP1
57% = UCP1, 71% = UCP2
32% = UCP1-3
38% = UCP1-3
60% = UCP1, 82% = UCP2

H+ Leak in IPC
Nadtochiy et al. (2006) Biochem. J. 395: 611-18
Membrane Potential (mV)
Res
pira
tion
(nm
ols
O2/m
in/m
g pr
otei
n)
CON
IPC+GDPIPC
25
37UCP2
50
0
10
20
30
40
75 100 125 150 175
UCP2 mRNA↑ in delayed IPCJ. Biol. Chem. (2005) 280: 33470-6
ROS, RNS, e-philes ↑H+ Leak/UCPs
Nature (2002) 415: 96-9
J. Biol. Chem. (2003) 278: 48534-45
J. Neurochem. (1998) 70: 2195-202
Car
diac
VO
2/ R
PP Con IPC
0.0
0.5
1.0
Pre 1st
IPC
*
2nd
IPC
*
3rd
IPC
*
Circ. Res. (2003) 93: 192–200
DNP/FCCP/Tg ↑UCP cardioprotection
Cardiovasc. Res. (2000) 47: 68-73
J. Mol. Cell. Cardiol. (2003) 35: 749-59
Circulation (2004) 110: 528-33
ROS / RNS are essential for IPCAntiox. Redox. Sig. (2004) 6: 393-404
Cardiovasc Res. (2006) 70: 231-9

Δψ (mV)
Res
pira
tion
(nm
ols
O2/m
in/m
g pr
otei
n) Ctrl
1µM LNO2
LNO2 + GDP
0
10
20
30
40
0 25 50 75 100 125 150 175
Added LNO2 protectsmyocytes from IR
20
40
60
80
100
Ctrl.
IR 500nMLN
O2
Cel
l via
bilit
y (%
)
0
*
Ctrl.
10.06
9 10 11 12 13t (min.) →
9.95 10.06 IPC
LNO2 is formed insidemitochondria during IPC
O
O- NO2
O
O- NO29,10 12,13
ROSRNS
LipidOxidation
UCPActivation
Mild H+ Leak(↓Δψm)
↓ROS↓[Ca2+]m
PT PoreIPC
*O
O- NO2
& Nitration
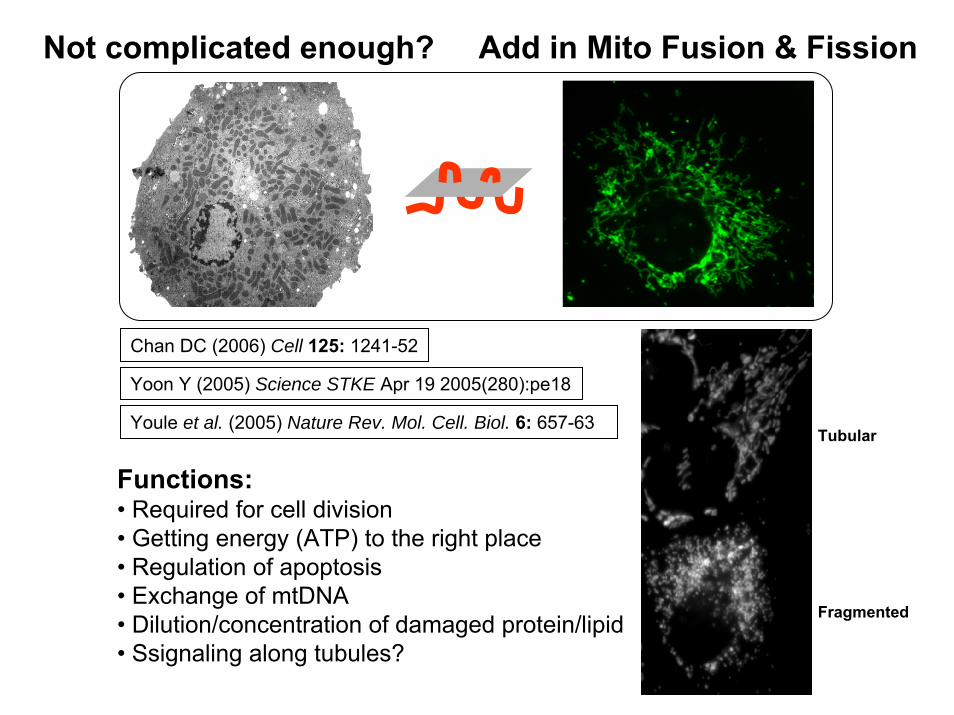
Not complicated enough? Add in Mito Fusion & Fission
Youle et al. (2005) Nature Rev. Mol. Cell. Biol. 6: 657-63
Functions:• Required for cell division• Getting energy (ATP) to the right place• Regulation of apoptosis• Exchange of mtDNA• Dilution/concentration of damaged protein/lipid• Ssignaling along tubules?
Chan DC (2006) Cell 125: 1241-52
Yoon Y (2005) Science STKE Apr 19 2005(280):pe18
Tubular
Fragmented

Mitochondrial Morphology: a Tightly Regulated Process
OPA-1 (mammalian homolog of Mgm-1p) Mito inner membrane GTPase, remodeling of IMM
Fragmentation essential for apoptosis Youle et al. (2001) Dev. Cell 1: 515-25
hFis-1 (human homolog of yeast Fis1p) 17 kDa mito’ outer membrane protein, recruits DLP1
Overexpress hFis1 fragmentation apoptosis Sugioka et al. (2004) JBC. 279: 52726-34
Mfn-1/2 (mitofusin, mammalian homolog of Fzo-1p) 86 kDa GTPase, tethers 2 mitochondria for fusion
Yu et al. (2005) J. Cell Sci. 118: 4141-51Overexpress Fzo-1p inhibits apoptosis
DLP-1 (dynamin-like protein, DRP-1, Dnm1-p) 80 kDa cytosolic GTPase, pinchase
Fragmentation essential for high glucose-induced ROS Yu et al. (2006) PNAS 103: 2653-8

H+
O2ADP
Δψm+
–
Cx II
NADH
NAD+
Cx I
Cx V
ATP
ANT
Cx III Cx IVCyt-c
Ca2+
UniporterCa2+/Na+
Exch’ UCP
FADH2FAD
TCACycle
H2O
ADP ATPH+Ca2+
VDAC
R+H+
H+ H+
Ca2+ Na+
H+
Rotenone,Amytal
Piericidin, MPP+
DNP
CCCP
FCCP
H+
H+
DIDS
Valinomycin K+
NigericinK+
H+
Attractyloside,Bongkrekic Acid
PAO, NEM
GDP,Genipin
Ru RedRu-360
CGP37157
TTFA, 3-NP,Malonate,Atpenins
Antimycin A,MyxothiazolStigmatellin
Cyanide,Azide,
CO, NO• Oligomycin
K+ATPChannel
K+
Diazoxide,Pinacidil,Nicorandil
5-HD,ATP
Glibenclamide
+
PT Pore
Cyp-DVDAC ANT
Cyclosporin A,N-methyl-val-CsA
Sanglifehrin A
Chloramphenicol
mRNA
MitochondrialProtein Synthesis
MitochondrialMitochondrialInhibitorsInhibitors
Abbreviations… 3-NP: 3-nitro propionic acid, 5-HD: 5-hydroxydecanoate, ANT: adenine nucleotide translocase, CCCP:carbonylcyanide m-chlorophenylhydrazone, Cx I: respiratory complex I etc., Cyp-D: cyclophilin D, Cyt-c: cytochrome c, DNP: dinitrophenol, DIDS: 4,4-diisothiocyanato-stilbene-2,2'disulphonate, FCCP: carbonyl cyanide p-[trifluoromethoxy]-phenyl-hydrazone, MPP+: 1-methyl-4-phenylpyridinium, NEM: N-ethylmaleimide, PAO: phenylarsine oxide, TTFA: thenoyl-trifluoroacetone, UCP: uncoupling protein, VDAC: volatege dependent anion channel.
Arsenite,Ketomethylvalerate
Mersalyl,NEM
PhosphateTransporter
Pi
Trans-aminase
Amino-oxyacetate Glutamate
Oxaloacetate
Ro-68-3400
α-CHC PyruvateTransporter
Pyr