micrococcus denitrificans - journal of bacteriology
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, June 1976, p. 1030-1036Copyright © 1976 American Society for Microbiology
Vol. 126, No. 3Printed in U.SA.
Pathway of Thiamine Pyrophosphate Synthesis inMicrococcus denitrificans
HIROSHI SANEMORI, YOSHIKO EGI, AND TAKASHI KAWASAKI*Department ofBiochemistry, Hiroshima University School of Medicine, Hiroshima 734, Japan
Received for publication 29 December 1975
The pathway of thiamine pyrophosphate (TPP) biosynthesis, which is formedeither from exogenously added thiamine or from the pyrimidine and thiazolemoieties of thiamine, in Micrococcus denitrificans was investigated. The follow-ing indirect evidence shows that thiamine pyrophosphokinase (EC 2.7.6.2) cata-lyzes the synthesis ofTPP from thiamine: (i) [35S]thiamine incubated with cellsof this microorganism was detected in the form of [VS]TPP or [35S]thiamine; (ii)thiamine gave a much faster rate of TPP synthesis than thiamine monophos-phate (TMP) when determined with the extracts; and (iii) a partially purifiedpreparation ofthe extracts can use thiamine, but not TMP, as the substrate. Theactivities of the four enzymes involved in TMP synthesis from pyrimidine andthiazole moieties of thiamine were detected in the extracts ofM. denitrificans.The extracts contained a high activity of the phosphatase, probably specific forTMP. After M. denitrificans cells were grown on a minimal medium containing3 mM adenosine, which causes derepression of de novo thiamine biosynthesis inEscherichia coli, the activities of the four enzymes involved with TMP synthe-sis, the TMP phosphatase, and the thiamine pyrophosphokinase were enhancedtwo- to threefold. These results indicate that TPP is synthesized directly fromthiamine without forming TMP as an intermediate and that de novo synthesis ofTPP from the pyrimidine and thiazole moieties involves the formation ofTMP,followed by hydrolysis to thiamine, which is then converted to TPP directly.Thus, the pathway of TPP synthesis from TMP synthesized de novo in M.denitrificans is different from that found in E. coli, in which TMP synthesizedde novo is converted directly to TPP without producing thiamine.
It has been well documented that thiaminepyrophosphate (TPP) acts as the coenzyme forpyruvate dehydrogenase (EC 1.2.4.1), oxoglu-tarate dehydrogenase (EC 1.2.4.2), pyruvatedecarboxylase (EC 4.1.1.1), transketolase (EC2.2.1.1), and several other enzymes.The pathway ofTPP biosynthesis in yeast (2-
4, 14, 15, 17, 26-28, 31) and Escherichia coli (10,12, 25), has been established and is shown inFig. 1. The four enzymes involved in the bio-synthetic process of thiamine monophosphate(TMP) from pyrimidine and thiazole moietiesof thiamine are the same in both organisms,whereas the reactions involved in TPP syn-thesis from TMP synthesized de novo are differ-ent in these two organisms. In yeast, TPP issynthesized from TMP via an intermediateproduction of free thiamine (2-4); that is, TMPis first dephosphorylated to thiamine before itis converted to TPP. Exogenously addedthiamine is, therefore, synthesized to TPP by aone-step reaction catalyzed by thiamine pyro-phosphokinase (EC 2.7.6.2) (13, 30, 31). In E.coli, TMP is directly phosphorylated to TPPwithout producing free thiamine and, there-
fore, thiamine added exogenously is trans-formed to TPP by two successive steps cata-lyzed by thiamine monophosphokinase (8, 20,21) and TMP kinase (20, 21, 23, 24), respec-tively.The enzymatic differences in the synthesis of
TPP between yeast and E. coli prompted us tostudy the reason why TPP synthesis fromthiamine proceeds by a one-step pyrophosphor-ylation reaction in yeast and, on the otherhand, by a two-step phosphorylation reactionsin E. coli. During the survey of different micro-organisms, it was also found that Micrococcusdenitrificans does contain the thiamine pyro-phosphokinase activity.The present paper describes evidence for the
existence of thiamine pyrophosphokinase in M.denitrificans as well as the pathway of de novosynthesis ofTPP from the pyrimidine and thia-zole moieties of thiamine.
MATERIALS AND METHODS
Organisms. M. denitrificans strain 12442 and E.coli K-12 strain 3301 were obtained from the Insti-
1030
on Decem
ber 14, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

PATHWAY OF TPP SYNTHESIS IN M. DENITRIFICANS 1031
I1) III)HMP--- HMP-P--A HMP-PP
-TMP---VII-- TPP
TbD11 ,/'- (VII) 2 V)(VI)
ThiamineFIG. 1. Pathway of TPP synthesis in yeast and
M. denitrificans (solid line) and E. coli (dottedline). (I) Hydroxymethylpyrimidine kinase; (II) hy-droxymethylpyrimidine phosphate kinase; (III) hy-droxyethylthiazole kinase; (IV) thiaminephosphatepyrophosphorylase; (V) a phosphatase (nonspecificin yeast; probably specific in M. denitrificans); (VI)thiamine pyrophosphokinase (yeast and M. denitri-ficans); (VII) thiamine monophosphokinase (E. coli);(VIII) TMP kinase (E. coli). HMP, Hydroxymethyl-pyrimidine or 2-methyl-4-amino-5-hydroxymethyl-pyrimidine (thiamine pyrimidine); HMP-P, HMPmonophosphate; HMP-PP, HMP pyrophosphate; Th,hydroxyethylthiazole or 4-methyl-5-hydroxyethyl-thiazole (thiamine thiazole); Th-P, Th monophos-phate.
tute for Fermentation, Osaka, Japan. Both are wildstrains.Chemicals. [35S]thiamine hydrochloride (100
mCi/mmol) was purchased from the RadiochemicalCentre, Amersham, England. Hydroxymethylpy-rimidine and hydroxyethylthiazole were generousgifts of S. Yurugi, Takeda Pharmaceutical Indus-tries, Ltd., Osaka. Hydroxymethylpyrimidine was
converted to its phosphate and pyrophosphate by themethod described previously (28). Hydroxyethyl-thiazole monophosphate was prepared from TMP bythe sulfite cleavage method (16). TMP and TPP hy-drochloride were purchased from Sigma ChemicalCo. The other chemicals used were of analyticalgrade.Growth of cells. M. denitrificans was grown aero-
bically at 37 C on a minimal medium (5) composed of(in grams per liter): sodium succinate 6H2O, 13.5;K2HPO4, 6.0; KH2PO4, 4.0; MgSO4-7H2O, 0.2; CaCl2,0.04; sodium molybdate, 0.15; MnSO4 4H2O, 0.001;NH4Cl, 3.2 (NH4NO3, 3.2, for anaerobic growth); andan iron-citrate solution, 5 ml, containing 1.1 g ofFeSO4 7H2O and 1.05 g of citric acid per liter.
E. coli K-12 was grown aerobically at 37 C on aminimal medium (6) containing 0.2% glucose as thesole of carbon source.When both bacteria were grown anaerobically,
each flask was filled to the neck with 400 ml of theminimal medium, followed by flushing with N2 gasprior to capping with a rubber stopper. The flaskswere incubated at 37 C after being inoculated with5-ml precultures of bacteria.
Cells of both bacteria were harvested at the latelogarithmic phase by centrifugation at 4 C.
Preparation of cell suspensions of M. denitrifi-cans. Harvested cells were washed once with themedium and then suspended in the same mediumused for growth. The cell suspension adjusted to anoptical density of 0.50 at 560 nm with a Spectronic 20spectrophotometer (Bausch and Lomb) was used foranalysis of [35S]thiamine uptake.
Preparation of crude extracts. Harvested cells ofM. denitrificans and E. coli were washed once in a
solution of 50 mM tris(hydroxymethyl)aminometh-ane (Tris)-hydrochloride (pH 7.5) and 1 mM dithio-threitol and then resuspended in the same buffersystem, followed by sonic treatment for 5 min at 4 C.After centrifugation for 20 min at 15,000 x g, thesupernatant fluids were used as crude extracts forthe assay of enzyme activities.
Protein concentration of the extracts was deter-mined by the method of Lowry et al. (18), withbovine serum albumin as a standard.
Assay of the enzyme activity to catalyze the syn-thesis of TPP from thiamine. The reaction mixturecontained 30 nmol of thiamine (or TMP), 2 Amol ofadenosine 5'-triphosphate (ATP), 10 ,umol ofMgSO4,100 ,umol of Tris-hydrochloride (pH 7.5), and analiquot of the enzyme preparation (3 mg of protein)in a total volume of 1.5 ml. To determine the activityin E. coli extracts, 150 ,umol of KCl was added. Thereaction was started by adding the enzyme prepara-tion, incubated for 15 min at 37 C, and then termi-nated by heating the mixture for 5 min at 90 C afterthe pH was adjusted to 5 with 1.5 ml of 0.01 N HCl.After removal of denatured proteins by centrifuga-tion, the TPP content was determined spectrophoto-metrically from the supernatant fluid by the methoddescribed previously (29).The kinase activity was expressed as nanomoles
of TPP formed per milligram of protein per hour.Assay of the activities of TMP-synthesizing en-
zymes. Activities of four enzymes involved in TMPsynthesis from hydroxymethylpyrimidine and hy-droxyethylthiazole (Fig. 1) were determined by pro-cedures described previously (9, 10).
Assay of the activity of TMP-phosphohydrolyz-ing enzyme. The reaction mixture was composed of0.1 ,±mol of TMP, 250 umol of Tris-hydrochloride(pH 7.5), and crude extract (2 mg of protein) in atotal volume of 5 ml. The reaction mixture wasincubated for 20 min at 37 C, and the amount ofthiamine liberated was deternined by the thio-chrome fluorescence method (7). The reaction ratedetermined under these conditions was linear up to5 mg ofprotein and also up to 60 min of incubation at37 C.
Analysis of intracellular forms of [35S]thiaminetransported. The M. denitrificans cell suspensionprepared as described above was incubated with 1,uM [35S]thiamine for 2 and 10 min at 37 C. The fateof [35Slthiamine taken up by M. denitrificans cellswas analyzed by the paper chromatographicmethod. The procedure employed was identical tothat used to study the fate of [35S]thiamine or[35S]hydroxyethylthiazole transported into E. colicells (11, 32).
RESULTSChromatographic analysis of [35S]thiamine
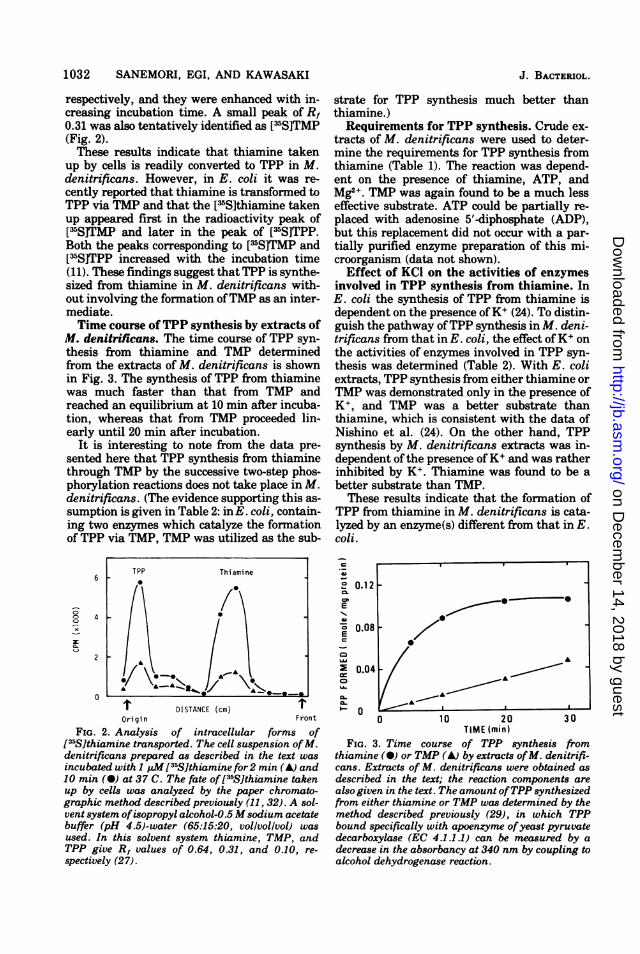
taken up by cells of M. denitrificans. Restingcells of M. denitrificans were incubated with[35S]thiamine in the presence of glucose, andthen the fate of [35S]thiamine after entry intothe cell was determined by paper chromato-graphic analysis. The two large radioactivitypeaks corresponding to Rf values 0.13 and 0.61were identified as [35S]TPP and ['5S]thiamine,
VOL. 126, 1976
on Decem
ber 14, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
1032 SANEMORI, EGI, AND KAWASAKI
respectively, and they were enhanced with in-creasing incubation time. A small peak of Rf0.31 was also tentatively identified as [35S]TMP(Fig. 2).These results indicate that thiamine taken
up by cells is readily converted to TPP in M.denitrificans. However, in E. coli it was re-cently reported that thiamine is transformed toTPP via TMP and that the [35S]thiamine takenup appeared first in the radioactivity peak of[35S]TMP and later in the peak of [35S]TPP.Both the peaks corresponding to [35SITMP and[15S]TPP increased with the incubation time(11). These findings suggest that TPP is synthe-sized from thiamine in M. denitrificans with-out involving the formation ofTMP as an inter-mediate.Time course ofTPP synthesis by extracts of
M. denitrificans. The time course of TPP syn-thesis from thiamine and TMP determinedfrom the extracts of M. denitrificans is shownin Fig. 3. The synthesis of TPP from thiaminewas much faster than that from TMP andreached an equilibrium at 10 min after incuba-tion, whereas that from TMP proceeded lin-early until 20 min after incubation.
It is interesting to note from the data pre-sented here that TPP synthesis from thiaminethrough TMP by the successive two-step phos-phorylation reactions does not take place in M.denitrificans. (The evidence supporting this as-sumption is given in Table 2: inE. coli, contain-ing two enzymes which catalyze the formationof TPP via TMP, TMP was utilized as the sub-
6
4
2
x
a-CL)
t DISTANCE (cm) tOrigin Front
FIG. 2. Analysis of intracellular forms of[35S]thiamine transported. The cell suspension ofM.denitrificans prepared as described in the text was
incubated with 1 pM[35Slthiamine for2 min (A) and10 min (-) at 37 C. The fate of[35S]thiamine takenup by cells was analyzed by the paper chromato-graphic method described previously (11, 32). A sol-vent system ofisopropyl alcohol-0.5 Msodium acetatebuffer (pH 4.5)-water (65:15:20, vollvollvol) was
used. In this solvent system thiamine, TMP, andTPP give Rf values of 0.64, 0.31, and 0.10, re-
spectively (27).
strate for TPP synthesis much better thanthiamine.)Requirements for TPP synthesis. Crude ex-
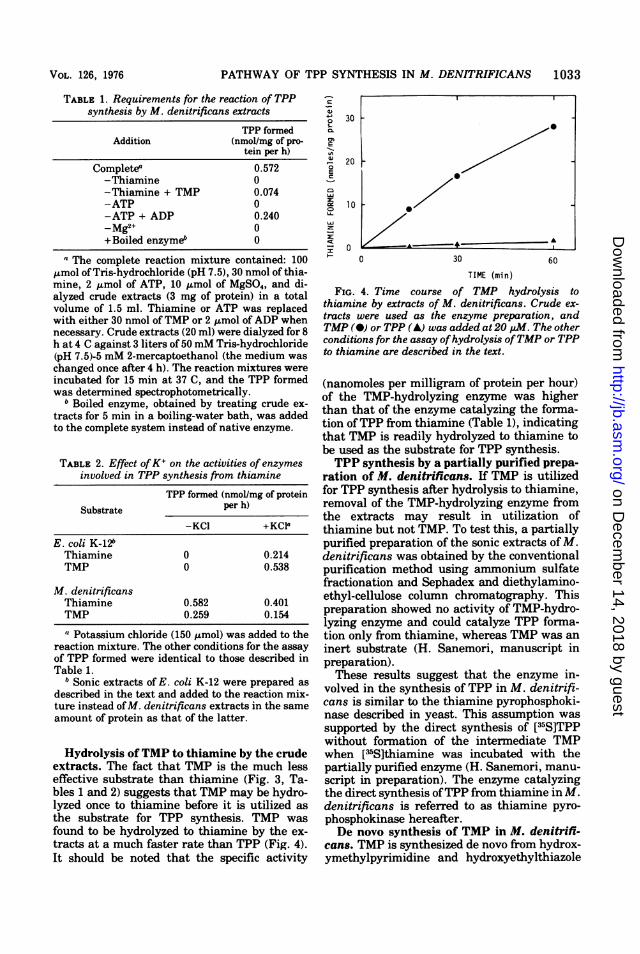
tracts of M. denitrificans were used to deter-mine the requirements for TPP synthesis fromthiamine (Table 1). The reaction was depend-ent on the presence of thiamine, ATP, andMg2+. TMP was again found to be a much lesseffective substrate. ATP could be partially re-placed with adenosine 5'-diphosphate (ADP),but this replacement did not occur with a par-tially purified enzyme preparation of this mi-croorganism (data not shown).
Effect of KCI on the activities of enzymesinvolved in TPP synthesis from thiamine. InE. coli the synthesis of TPP from thiamine isdependent on the presence ofK+ (24). To distin-guish the pathway ofTPP synthesis in M. deni-trificans from that in E. coli, the effect ofK+ onthe activities of enzymes involved in TPP syn-thesis was determined (Table 2). With E. coliextracts, TPP synthesis from either thiamine orTMP was demonstrated only in the presence ofK+, and TMP was a better substrate thanthiamine, which is consistent with the data ofNishino et al. (24). On the other hand, TPPsynthesis by M. denitrificans extracts was in-dependent of the presence ofK+ and was ratherinhibited by K+. Thiamine was found to be abetter substrate than TMP.These results indicate that the formation of
TPP from thiamine in M. denitrificans is cata-lyzed by an enzyme(s) different from that in E.coli.
° 0.12
Eas° 0.08 7E
E0.04cmo /
0 10 20 30TIME (min)
FIG. 3. Time course of TPP synthesis fromthiamine (a) or TMP (A) by extracts ofM. denitrifi-cans. Extracts of M. denitrificans were obtained asdescribed in the text; the reaction components arealso given in the text. The amount ofTPP synthesizedfrom either thiamine or TMP was determined by themethod described previously (29), in which TPPbound specifically with apoenzyme ofyeast pyruvatedecarboxylase (EC 4.1.1.1) can be measured by adecrease in the absorbancy at 340 nm by coupling toalcohol dehydrogenase reaction.
TPP Thiamine0
S
:/'\~~~~x
J. BACTERIOL.
on Decem
ber 14, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
PATHWAY OF TPP SYNTHESIS IN M. DENITRIFICANS 1033
TABLE 1. Requirements for the reaction of TPPsynthesis by M. denitrificans extracts
TPP formedAddition (nmol/mg of pro-
tein per h)
Completea 0.572-Thiamine 0-Thiamine + TMP 0.074-ATP 0-ATP + ADP 0.240-gM2+ 0+Boiled enzymeb 0
The complete reaction mixture contained: 100,umol ofTris-hydrochloride (pH 7.5), 30 nmol of thia-mine, 2 utmol of ATP, 10 ,tmol of MgSO4, and di-alyzed crude extracts (3 mg of protein) in a totalvolume of 1.5 ml. Thiamine or ATP was replacedwith either 30 nmol ofTMP or 2 ,Lmol of ADP whennecessary. Crude extracts (20 ml) were dialyzed for 8h at 4 C against 3 liters of 50mM Tris-hydrochloride(pH 7.5)-5 mM 2-mercaptoethanol (the medium waschanged once after 4 h). The reaction mixtures wereincubated for 15 min at 37 C, and the TPP formedwas determined spectrophotometrically.
b Boiled enzyme, obtained by treating crude ex-tracts for 5 min in a boiling-water bath, was addedto the complete system instead of native enzyme.
TABLE 2. Effect ofK+ on the activities of enzymesinvolved in TPP synthesis from thiamine
TPP formed (nmol/mg of proteinSubstrate per h)
-KC1 +KCIa
E. coli K-12bThiamine 0 0.214TMP 0 0.538
M. denitrificansThiamine 0.582 0.401TMP 0.259 0.154
{I Potassium chloride (150 ,umol) was added to thereaction mixture. The other conditions for the assayof TPP formed were identical to those described inTable 1.
b Sonic extracts of E. coli K-12 were prepared asdescribed in the text and added to the reaction mix-ture instead ofM. denitrificans extracts in the sameamount of protein as that of the latter.
Hydrolysis of TMP to thiamine by the crudeextracts. The fact that TMP is the much lesseffective substrate than thiamine (Fig. 3, Ta-bles 1 and 2) suggests that TMP may be hydro-lyzed once to thiamine before it is utilized asthe substrate for TPP synthesis. TMP wasfound to be hydrolyzed to thiamine by the ex-tracts at a much faster rate than TPP (Fig. 4).It should be noted that the specific activity
a
0
._
a
0
0)E
E
X:Ie
30
20~
10
0 k30 60
TIME (min)
FIG. 4. Time course of TMP hydrolysis tothiamine by extracts of M. denitrificans. Crude ex-
tracts were used as the enzyme preparation, andTMP (-) or TPP (A) was added at 20 pM. The otherconditions for the assay ofhydrolysis ofTMP or TPPto thiamine are described in the text.
(nanomoles per milligram of protein per hour)of the TMP-hydrolyzing enzyme was higherthan that of the enzyme catalyzing the formna-tion ofTPP from thiamine (Table 1), indicatingthat TMP is readily hydrolyzed to thiamine tobe used as the substrate for TPP synthesis.TPP synthesis by a partially purified prepa-
ration of M. denitrificans. If TMP is utilizedfor TPP synthesis after hydrolysis to thiamine,removal of the TMP-hydrolyzing enzyme fromthe extracts may result in utilization ofthiamine but not TMP. To test this, a partiallypurified preparation of the sonic extracts ofM.denitrificans was obtained by the conventionalpurification method using ammonium sulfatefractionation and Sephadex and diethylamino-ethyl-cellulose column chromatography. Thispreparation showed no activity of TMP-hydro-lyzing enzyme and could catalyze TPP forma-tion only from thiamine, whereas TMP was an
inert substrate (H. Sanemori, manuscript inpreparation).These results suggest that the enzyme in-
volved in the synthesis of TPP in M. denitrifi-cans is similar to the thiamine pyrophosphoki-nase described in yeast. This assumption was
supported by the direct synthesis of [3S]TPPwithout formation of the intermediate TMPwhen [35S]thiamine was incubated with thepartially purified enzyme (H. Sanemori, manu-
script in preparation). The enzyme catalyzingthe direct synthesis ofTPP from thiamine in M.denitrificans is referred to as thiamine pyro-phosphokinase hereafter.De novo synthesis of TMP in M. denitrifi-
cans. TMP is synthesized de novo from hydrox-ymethylpyrimidine and hydroxyethylthiazole
1-~~
Z-I + ,
VOL. 126, 1976
on Decem
ber 14, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

1034 SANEMORI, EGI, AND KAWASAKI
in yeast (2-4, 14, 26-28), as well as in E. coli(10, 25), as shown in Fig. 1. Activities of theoverall reaction and of the four enzymes in-volved in TMP synthesis were determined (Ta-ble 3). All of these activities were detected inextracts of M. denitrificans and were lowerthan those found in E. coli extracts. This sug-gests that a pathway of TMP synthesis fromhydroxymethylpyrimidine and hydroxyethyl-thiazole comparable to that in E. coli is operat-ing in M. denitrificans.
Effect of adenosine added to the growthmedium on TPP synthesis. It is important todetermine whether or not a pathway of de novo
TPP synthesis from TMP via thiamine is func-tioning physiologically in M. denitrificans.When adenosine or adenine is added to thegrowth medium, synthesis of the enzymes in-volved in TMP synthesis in E. coli can be dere-pressed, resulting in increases in the activitiesof these enzymes (9). With the extracts of M.denitrificans grown in the presence of 3 mMadenosine, activities of the overall reaction ofTPP synthesis, thiaminephosphate pyrophos-phorylase, TMP-phosphohydrolyzing enzyme,and thiamine pyrophosphokinase were mea-sured (Table 4). All of these activities werefound to be enhanced more than twofold in thepresence of adenosine.Adenine and adenosine were found to de-
crease the intracellular concentration of TPP(9, 22), which acts as a corepressor to bringabout the regulation of thiamine biosynthesis.Therefore, the results described above suggestthat the enzymes listed in Table 4 are involvedin the physiological pathway of TPP synthesisin M. denitrificans.TPP synthesis by cells of M. denitrificans
grown anaerobically. M. denitrificans cangrow well anaerobically on a medium contain-ing NH4NO3 as the terminal electron acceptorinstead of NH4Cl. Whether anaerobiosis of this
TABLE 3. Activities of enzymes involved in TMPsynthesisa
a The overall reaction ofthiamine synthesis from hydrox-ymethylpyrimidine and hydroxyethylthiazole (25) and theactivities of four enzymes involved in TMP synthesis weredetermined with crude extracts of E. coli K-12 and M.denitrificans as described previously (9, 10). They are givenas nanomoles per milligram of protein per hour.
Enzymes I through IV correspond to those illustrated inFig. 1. I, Hydroxymethylpyrimidine kinase; II, hydroxy-methylpyrimidine phosphate kinase; III, hydroxyethyl-thiazole kinase; IV, thiaminephosphate pyrophosphorylase.
TABLE 4. Effect of adenosine added to the growthmedium on TPP synthesis in M. denitrificans"
Thia-TMP py-TM mine
Addition Overall rophos- phos- minreaction phoryl- phatase phos-ase phoki-
nase
None 0.029 0.132 26.1 0.6733 mM adenosine 0.100 0.418 44.3 2.00
a M. denitrificans was grown in a minimal me-dium (5) supplemented with or without 3 mM adeno-sine, and crude extracts were prepared as describedin the text. The assay procedures for the enzymeactivities, as well as the overall reaction of TPPsynthesis, were as described in the text. Valuesgiven are nanomoles per milligram of protein perhour.
organism induces an alternative pathway ofTPP synthesis similar to that found in E. coliwas also tested. Extracts of M. denitrificanscells grown either aerobically on NH4Cl or an-aerobically on NH4NO3 showed almost identi-cal formation of TPP from thiamine (0.495 and0.552 nmol/mg of protein per h, respectively)and utilized TMP as a far less effective sub-strate than thiamine. Thus, the pathway ofTPP synthesis in M. denitrificans was not al-tered for growth under aerobic and anaerobicconditions.
DISCUSSIONThe pathway of enzymatic synthesis of TPP,
a key coenzyme in various enzyme reactions, isfairly well understood in E. coli (8, 12, 20-25)and yeast (for a review see reference 1). In E.coli, TPP is formed directly from TMP by denovo synthesis, whereas in yeast TPP forna-tion takes place through the intermediate pro-duction of the free form of thiamine. In thepresent study we present evidence that thereexist differences in TPP formation in E. coliand M. denitrificans, and the enzymatic se-quence of TPP formation in M. denitrificansfollows the same pathway as that elucidated foryeast.The data that support the direct formation of
TPP from thiamine are as follows. (i)[35S]thiamine incubated with M. denitrificanscells was detected in the cells of [35S]TPP or[35S]thiamine but not as [P5S]TMP (Fig. 2). (ii)The rate ofTPP synthesis was much faster fromthiamine than from TMP when determinedwith the extracts (Fig. 3, Tables 1, 2, and 4).(iii) A partially purified preparation ofM. deni-trificans extracts, which does not contain theactivity of TMP-phosphohydrolyzing enzyme,
J. BACTERIOL.
on Decem
ber 14, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

PATHWAY OF TPP SYNTHESIS IN M. DENITRIFICANS 1035
can convert only thiamine to TPP but not TMPto TPP and also can catalyze the formationof [35SITPP from [35S]thiamine without form-ing [35S]TMP (H. Sanemori, manuscript inpreparation). This evidence clarifies the pointthat the TPP synthesis from thiamine proceedsby a one-step pyrophosphorylation reactionwithout intermediate formation of TMP, likefound in yeast but not in E. coli. In addition tothis, TPP synthesis from thiamine by E. coliextracts showed a complete dependence on K+,whereas in the case ofM. denitrificans K+ par-tially inhibited TPP synthesis (Table 2). Thisindicates clearly that the enzyme system forTPP synthesis from thiamine in M. denitrifi-cans is different from that in E. coli.De novo TMP synthesis from hydroxymethyl-
pyrimidine and hydroxyethylthiazole in M.denitrificans occurs by the same pathway as inE. coli (10, 25) and yeast (for a review, seereference 1). The four enzymes involved in TMPsynthesis are hydroxymethylpyrimidine ki-nase, hydroxymethylpyrimidine phosphate ki-nase, hydroxyethylthiazole kinase, and thia-minephosphate pyrophosphorylase, as illus-trated in Fig. 1. TMP synthesized de novo in M.denitrificans should be hydrolyzed to thiamineby a phosphatase before conversion to thecoenzyme TPP; its formation is catalyzed bythiamine pyrophosphokinase as describedabove. If such a phosphatase specific for TMP,which is referred to as TMP phosphatase, ispresent and involved in de novo TPP synthesisfrom TMP via thiamine, the phosphataseshould be regulated by the same mechanismthat controls the other enzymes in the path-way. Adenosine, which causes derepression ofthiamine biosynthesis when added to thegrowth medium (22), resulted in an increase inthe activities ofTPP-synthesizing enzymes (Ta-ble 4). TMP phosphatase was found to be one ofthese enzymes, and it was also evident that thisTMP phosphatase enzyme was different fromother phosphatases. In an preliminary experi-ment, a protein peak exhibiting the activity ofTMP hydrolysis at pH 6.0 was separately ob-tained from the peak of acid phosphatase bySephadex G-200 column chromatography (datanot shown). In yeast (see reference 26 in review1) and parsley leaf (19), which contains thesame pathway for TPP synthesis as M. denitri-ficans, it was demonstrated that TMP synthe-sized de novo is hydrolyzed to thiamine by phos-phatases, including acid phosphatase withbroad substrate specificities.The results described above indicate the pres-
ence of a possible pathway of de novo TPPsynthesis in M. denitrificans, as illustrated inFig. 1. The reason why E. coli and M. denitrifi-
cans contain different pathways for TPP syn-thesis from thiamine has not yet been eluci-dated. A toxonomic survey of this pathway isnow under way.
ACKNOWLEDGMENTWe wish to thank Syed Raziuddin for his critical reading
of the manuscript.
LITERATURE CITED1. Brown, G. M. 1970. Biogenesis and metabolism of
thiamine, p. 369-381. In D. M. Greenberg (ed.), Meta-bolic pathways, 3rd ed., vol. 4. Academic Press Inc.,New York.
2. Camiener, G. W., and G. M. Brown. 1959. The biosyn-thesis of thiamine phosphates by extracts of bakers'yeast. J. Am. Chem. Soc. 81:3800.
3. Camiener, G. W., and G. M. Brown. 1960. The biosyn-thesis of thiamine. I. Enzymic formation of thiamineand phosphate esters of the pyrimidine moiety ofthiamine. J. Biol. Chem. 235:2404-2410.
4. Camiener, G. W., and G. M. Brown. 1960. The biosyn-thesis of thiamine. II. Fractionation of enzyme sys-tem and identification of thiazole monophosphate andthiamine monophosphate as intermediates. J. Biol.Chem. 235:2411-2417.
5. Chang, J. P., and J. G. Morris. 1962. Studies on theutilization of nitrate by Micrococcus denitrificans. J.Gen. Microbiol. 29:301-310.
6. Davis, B. D., and E. S. Mingioli. 1950. Mutants ofEscherichia coli requiring methionine or vitamin B,,.J. Bacteriol. 60:17-28.
7. Fujita, A. 1955. Thiaminase, p. 622-628. In S. P. Colo-wick and N. 0. Kaplan (ed.), Methods in enzymology,vol. 2. Academic Press Inc., New York.
8. Iwashima, A., H. Nishino, and Y. Nose. 1972. Conver-sion of thiamine to thiamine monophosphate by cell-free extracts of Escherichia coli. Biochim. Biophys.Acta 258:333-336.
9. Kawasaki, T., A. Iwashima, and Y. Nose. 1969. Regula-tion of thiamine biosynthesis in Escherichia coli. J.Biochem. (Tokyo) 65:407-416.
10. Kawasaki, T., T. Nakata, and Y. Nose. 1968. Geneticmapping with a thiamine-requiring auxotroph ofEscherichia coli K-12 defective in thiaminephosphatepyrophosphorylase. J. Bacteriol. 95:1483-1485.
11. Kawasaki, T., and K. Yamada. 1972. The uptake sys-tem of free thiamine in mutants of Escherichia coli.Biochem. Biophys. Res. Commun. 47:465-471.
12. Kayama, Y., and T. Kawasaki. 1973. Purification andproperties of thiaminephosphate pyrophosphorylaseofEscherichia coli. Arch. Biochem. Biophys. 158:242-248.
13. Kaziro, Y., R. Tanaka, Y. Mano, and N. Shimazono.1961. On the mechanism of transpyrophosphorylationin the biosynthesis of thiamine diphosphate. J. Bio-chem. (Tokyo) 49:472-476.
14. Leder, I. G. 1959. The enzymatic synthesis of thiaminemonophosphate. Biochem. Biophys. Res. Commun.1:63-66.
15. Leder, I. G. 1961. The enzymatic synthesis of thiaminemonophosphate. J. Biol. Chem. 236:3066-3071.
16. Leder, I. G. 1970. Preparation of -(4-methyl-5-thiazo-lyl)-ethyl phosphate, p. 166-167. In D. B. McCormickand L. D. Wright (ed.), Methods in enzymology, vol.18. Academic Press Inc., New York.
17. Lewin, L. M., and G. M. Brown. 1961. The biosynthesisof thiamine. III. Mechanism of enzymatic formationof the pyrophosphate ester of 2-methyl-4-amino-5-hy-droxymethylpyrimidine. J. Biol. Chem. 236:2768-2771.
VOL. 126, 1976
on Decem
ber 14, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

1036 SANEMORI, EGI, AND KAWASAKI
18. Lowry, 0. H., N. J. Rosebrough, A. L. Far, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:266-275.
19. Mitsuda, H., Y. Takii, K. Iwami, and K. Yasumoto.1975. Purification and properties of thiamine pyro-phosphokinase from parsley leaf. J. Nutr. Sci. Vitam-inol. 21:103-115.
20. Nakayama, H., and R. Hayashi. 1972. Biosynthesis ofthiamine pyrophosphate in Escherichia coli. J. Bacte-riol. 109:936-939.
21. Nakayama, H., and R. Hayashi. 1972. Biosyntheticpathway of thiamine pyrophosphate: a special refer-ence to the thiamine monophosphate-requiring mu-tant and the thiamine pyrophosphate-requiring mu-tant ofEscherichia coli. J. Bacteriol. 112:1118-1126.
22. Newell, P. C., and R. G. Tucker. 1966. The de-repres-sion ofthiamine biosynthesis by adenosine. A tool forinvestigating this biosynthetic pathway. Biochem. J.100:512-516.
23. Nishino, H. 1972. Biogenesis of cocarboxylase in Esche-richia coli. Partial purification and some properties ofthiamine monophosphate kinase. J. Biochem. (To-kyo) 72:1093-1100.
24. Nishino, H., A. Iwashima, and Y. Nose. 1971. Biogene-sis of cocarboxylase in Escherichia coli: a novel en-zyme catalyzing the formation of thiamine pyrophos-phate from thiamine monophosphate. Biochem. Bio-phys. Res. Commun. 45:363-368.
25. Now, Y., Y. Tokuda, M. Hirabayashi, and A. Iwash-ima. 1964. Thiamine biosynthesis from hydroxy-methylpyrimidine and thiazole by washed cells and
cell extracts of Escherichia coli and its mutants. J.Vitaminol. (Kyoto) 10:105-110.
26. Nose, Y., K. Ueda, and T. Kawasaki. 1959. Enzymicsynthesis of thiamine. Biochim. Biophys. Acta34:277-279.
27. Nose, Y., K. Ueda, and T. Kawasaki. 1961. Enzymaticsynthesis of thiamine. I. The enzymatic synthesis ofthiamine from pyrimidine and thiazole moieties by acell-free extract of yeast. J. Vitaminol. (Kyoto) 7:92-97.
28. Nose, Y., K. Ueda, T. Kawasaki, A. Iwashima, and T.Fujita. 1961. Enzymatic synthesis of thiamine. II.The thiamine synthesis from pyrimidine and thiazolephosphates and the enzymatic synthesis of pyrimi-dine mono- and diphosphate and thiazole monophos-phate. J. Vitaminol. (Kyoto) 7:98-114.
29. Sanemori, H., S. Yoshida, and T. Kawasaki. 1974.Studies on the binding of thiamine pyrophosphate toapoenzyme of yeast pyruvate decarboxylase. J. Bio-chem. (Tokyo) 75:123-129.
30. Shimazono, N., Y. Mano, R. Tanaka, and Y. Kaziro.1959. Mechanism of transpyrophosphorylation withthiaminokinase. J. Biochem. (Tokyo) 49:959-961.
31. Steyn-ParyE, E. S. 1952. Partial purification and prop-erties of thiaminokinase from yeast. Biochim. Bio.phys. Acta 8:310-324.
32. Yamasaki, H., H. Sanemori, K. Yamada, and T. Kawa-saki. 1973. Hydroxymethylthiazole uptake in Esche-richia coli: general properties and relationship be-tween uptake and thiamine biosynthesis. J. Bacte-riol. 116:1280-1286.
J. BACTERIOL.
on Decem
ber 14, 2018 by guesthttp://jb.asm
.org/D
ownloaded from