measuring growth, production, and yield of the giant kelp, macrocystis pyrifera
TRANSCRIPT

Measuring growth, production, and yield of the giant kelp, Macrocystis pyrifera
V. A . Gerard' & W . J . NorthCalifornia Institute of Technology, Kerckhoff Marine Laboratory, 101 Dahlia Street, Corona del Mar,CA 92625, U.S.A .I Present address: Maine Sciences Research Center, State University of New York, Stone Brook, NY 11794,U.S.A .
Keywords: seaweed, Macrocystis pyrifera, giant kelp, yield, productivity, growth, biomass
Introduction
Harvestable yield of the giant kelp, Macrocystispyrifera (L.) C . Agardh, is important to its use as acommercial source of phycocolloids and to devel-opment of marine biomass as an energy resource .Accurate prediction of yields from natural popula-tions and from crop plants on future marine farmsis necessary for optimum utilization of the naturalresource and for optimum design and managementof the cultivated resource . The nature of the yieldparameter - a single value integrating a number ofdynamic processes over a relatively long time peri-od - makes accurate prediction difficult . Temporaland spatial variability in growth, biomass produc-tion and loss, population structure and densitymight preclude reliable extrapolation of empiricalyield values to populations at other sites or times .To determine the magnitude of short-term tempor-al variability and better to define relationships be-tween kelp growth, biomass production and yield,we held a population of adult M. pyrifera underpartially controlled conditions . Nutrient supply,plant density, and holdfast depth were controlled .Irradiance and temperature were not controlled butwere monitored continuously . Growth and popula-tion parameters were measured over relativelyshort intervals . Harvestable yield was determinedempirically for the entire study period .
Materials and methods
We selected 63 adult M. pyrifera (25-43 frondsper plant) for transplanting during April, 1982 . Ten
Hydrobiologia 116/117, 321-324 (1984) .@ Dr W. Junk Publishers, Dordrecht . Printed in the Netherlands .
were from Laguna Beach, California . The remain-ing plants were from Isthmus Reef, Santa CatalinaIsland, California. All were collected from sub-strate depths of 9-15 m and were harvested prior totransplanting by cutting fronds 8 m above the hold-fast and removing the canopy portions . Fifty-one ofthe plants were transplanted to an experimentalenclosure located in Big Fisherman Cove, SantaCatalina Island. Twelve plants were transplanted toan unenclosed control site 40 m from the enclosure .None of the plants was exposed to air for more than1 min during transplanting. Holdfasts of all plantswere attached to buoys at 9 m depth and were even-ly spaced to give a plant density of 0 .43 m-2 .
The experimental enclosure consisted of a poly-vinyl chloride (PVC) bag, 15 m in diameter and12 m deep, suspended from a floating platform . Itwas deployed in January 1982, to allow leaching ofpotentially toxic substances prior to transplanting .The bag volume was 2 X 106 1. Seawater waspumped from 3 m depth outside the enclosure intothe bag at a rate of 100-150 1 s-l, giving a turnovertime of 4-6 h for the total volume . Most inflowingwater entered near the bottom of the bag, but part ofthe inflow was directed outward from the center ofthe enclosure at 0.5 m depth. Water flowed out of theenclosure via six mesh-covered ports in the bag,located near the waterline .
Nutrient concentrations within the enclosurewere increased during two periods by continuousaddition of chemical fertilizers (NaNO3 and NaH2-P04) to the inflowing water . Water samples werecollected 3 times weekly inside and outside theenclosure, and NO - and p043- concentrations weredetermined using a Technicon Auto-Analyzer . Wa-
3 22
ter temperatures inside and outside the enclosurewere recorded at 30-min intervals . Irradiance (400-700 nm) above the enclosure was measured with apyranometer and recorded at 30-min intervals .
Population parameters were determined forplants within the enclosure and for control plants .Monthly frond counts at I-m depth intervals be-tween the holdfast and canopy provided plant andfrond size distribution data . The frond wet-weight-to-length relationship was determined by measuringfronds collected from the enclosure . These data wereused to estimate biomass density at various timesduring the study . Frond growth was measured as aweekly increase in total stipe length of tagged fronds(initial length = 0 .5-9 m) . Standard growth rate (G)was calculated for each frond according to North(1971). Biomass production rates were estimatedusing frond elongation rates, size distributions, andthe frond weight-to-length relationship . Subcanopy
150-
0
x
April
May
June
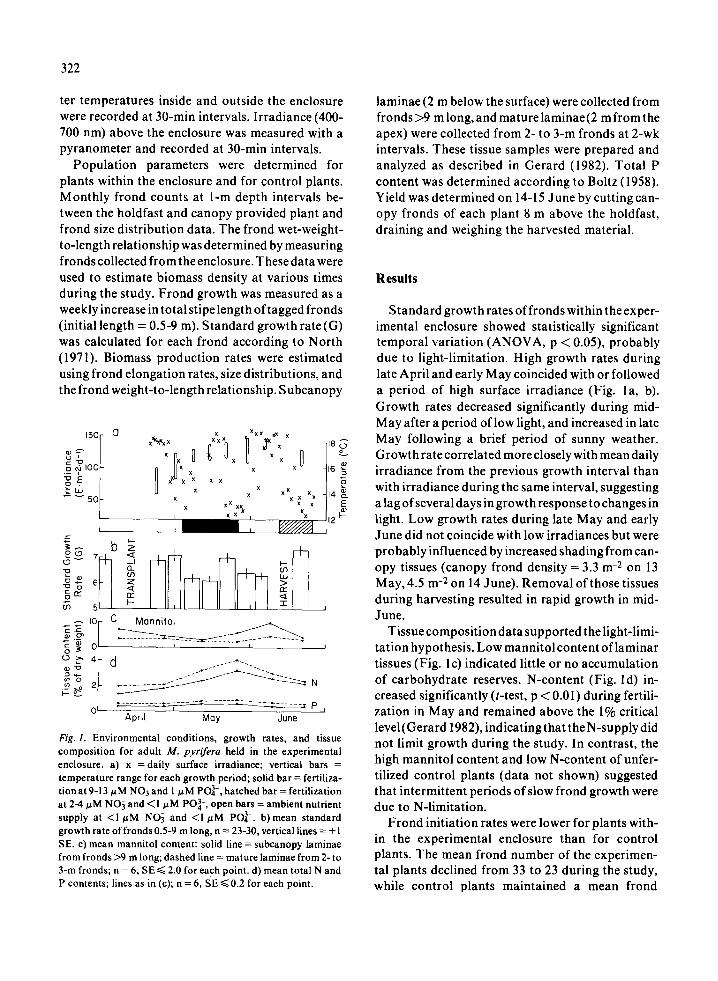
Fig. l . Environmental conditions, growth rates, and tissuecomposition for adult M. pyrifera held in the experimentalenclosure. a) x =daily surface irradiance; vertical bars =temperature range for each growth period ; solid bar = fertiliza-tion at 9-13 µM NO3 and 1 µM POL - , hatched bar = fertilizationat 2-4µM NO3 and <I µM p034-, open bars = ambient nutrientsupply at <1 µM N03 and <I µM PO4- . b) mean standardgrowth rate of fronds 0 .5-9 m long, n = 23-30, vertical lines = ± ISE. c) mean mannitol content: solid line = subcanopy laminaefrom fronds >9 m long ; dashed line = mature laminae from 2-to3-m fronds ; n = 6, SE < 2 .0 for each point . d) mean total N andP contents ; lines as in (c) ; n = 6, SE < 0 .2 for each point .
laminae (2 m below the surface) were collected fromfronds >9 m long, and mature laminae(2 mfrom theapex) were collected from 2- to 3-m fronds at 2-wkintervals . These tissue samples were prepared andanalyzed as described in Gerard (1982) . Total Pcontent was determined according to Boltz (1958) .Yield was determined on 14-15 June by cutting can-opy fronds of each plant 8 m above the holdfast,draining and weighing the harvested material .
Results
Standard growth rates of fronds within the exper-imental enclosure showed statistically significanttemporal variation (ANOVA, p < 0 .05), probablydue to light-limitation . High growth rates duringlate April and early May coincided with or followeda period of high surface irradiance (Fig . I a, b) .Growth rates decreased significantly during mid-May after a period of low light, and increased in lateMay following a brief period of sunny weather .Growth rate correlated more closely with mean dailyirradiance from the previous growth interval thanwith irradiance during the same interval, suggestinga lag of several days in growth response to changes inlight . Low growth rates during late May and earlyJune did not coincide with low irradiances but wereprobably influenced by increased shading from can-opy tissues (canopy frond density = 3.3 m -2 on 13May, 4.5 m -2 on 14 June) . Removal of those tissuesduring harvesting resulted in rapid growth in mid-June .
Tissue composition data supported the light-limi-tation hypothesis . Low mannitol content of laminartissues (Fig . lc) indicated little or no accumulationof carbohydrate reserves . N-content (Fig . Id) in-creased significantly (t-test, p < 0 .01) during fertili-zation in May and remained above the 1% criticallevel (Gerard 1982), indicating that the N-supply didnot limit growth during the study . In contrast, thehigh mannitol content and low N-content of unfer-tilized control plants (data not shown) suggestedthat intermittent periods of slow frond growth weredue to N-limitation .
Frond initiation rates were lower for plants with-in the experimental enclosure than for controlplants. The mean frond number of the experimen-tal plants declined from 33 to 23 during the study,while control plants maintained a mean frond
'(///AL Fa
Z0 0
a0o a~0 o
a-o_zQ
-}--U)
LLJ
ocn
5Qx
1 0 C
Mannitolc rn
0----------- -- ---- ----------
1
10 30
4- da3 0
a, oN a 2 NF O
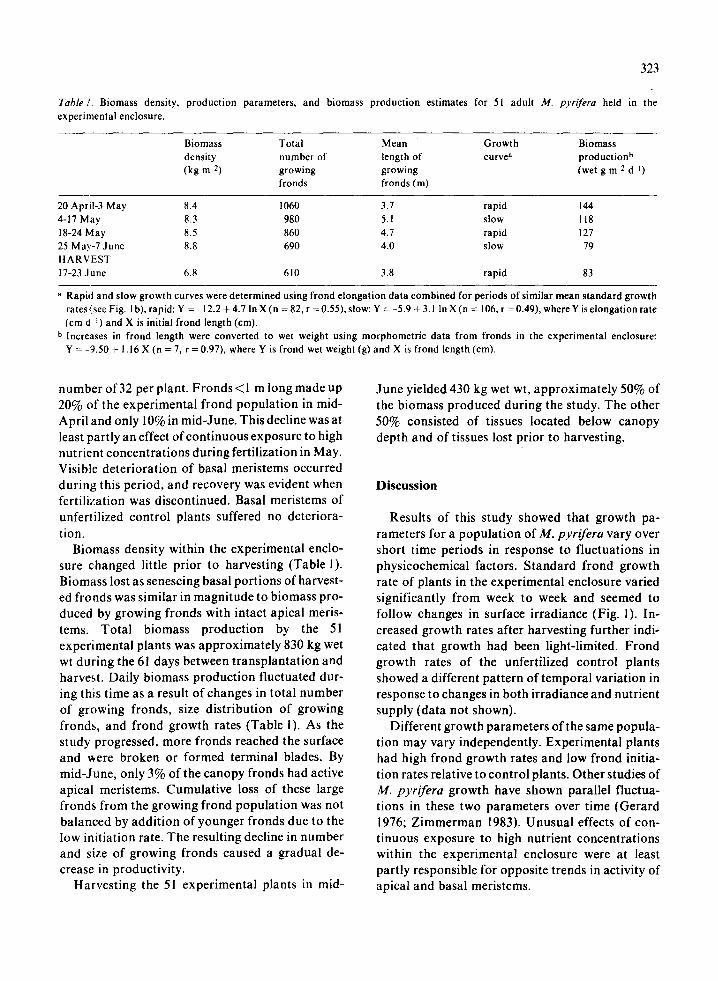
Table 1 . Biomass density, production parameters, and biomass production estimates for 51 adult M. pyrifera held in theexperimental enclosure .
number of 32 per plant . Fronds <1 m long made up20% of the experimental frond population in mid-April and only 10% in mid-June . This decline was atleast partly an effect of continuous exposure to highnutrient concentrations during fertilization in May .Visible deterioration of basal meristems occurredduring this period, and recovery was evident whenfertilization was discontinued . Basal meristems ofunfertilized control plants suffered no deteriora-tion .
Biomass density within the experimental enclo-sure changed little prior to harvesting (Table 1) .Biomass lost as senescing basal portions of harvest-ed fronds was similar in magnitude to biomass pro-duced by growing fronds with intact apical meris-tems . Total biomass production by the 51experimental plants was approximately 830 kg wetwt during the 61 days between transplantation andharvest. Daily biomass production fluctuated dur-ing this time as a result of changes in total numberof growing fronds, size distribution of growingfronds, and frond growth rates (Table 1) . As thestudy progressed, more fronds reached the surfaceand were broken or formed terminal blades . Bymid-June, only 3% of the canopy fronds had activeapical meristems . Cumulative loss of these largefronds from the growing frond population was notbalanced by addition of younger fronds due to thelow initiation rate . The resulting decline in numberand size of growing fronds caused a gradual de-crease in productivity .
Harvesting the 51 experimental plants in mid-
a Rapid and slow growth curves were determined using frond elongation data combined for periods of similar mean standard growthrates (see Fig . I b), rapid : Y =-12 .2 + 4 .7 In X (n = 82, r = 0 .55), slow : Y =-5 .9 + 3 .1 In X (n = 106, r =0 .49), where Y is elongation rate(cm d - 1 ) and X is initial frond length (cm) .
b Increases in frond length were converted to wet weight using morphometric data from fronds in the experimental enclosure :Y = -9 .50 + 1 .16 X (n = 7, r = 0 .97), where Y is frond wet weight (g) and X is frond length (cm) .
June yielded 430 kg wet wt, approximately 50% ofthe biomass produced during the study . The other50% consisted of tissues located below canopydepth and of tissues lost prior to harvesting .
Discussion
Results of this study showed that growth pa-rameters for a population of M. pyrifera vary overshort time periods in response to fluctuations inphysicochemical factors . Standard frond growthrate of plants in the experimental enclosure variedsignificantly from week to week and seemed tofollow changes in surface irradiance (Fig . 1) . In-creased growth rates after harvesting further indi-cated that growth had been light-limited . Frondgrowth rates of the unfertilized control plantsshowed a different pattern of temporal variation inresponse to changes in both irradiance and nutrientsupply (data not shown) .
Different growth parameters of the same popula-tion may vary independently . Experimental plantshad high frond growth rates and low frond initia-tion rates relative to control plants . Other studies ofM. pyrifera growth have shown parallel fluctua-tions in these two parameters over time (Gerard1976; Zimmerman 1983). Unusual effects of con-tinuous exposure to high nutrient concentrationswithin the experimental enclosure were at leastpartly responsible for opposite trends in activity ofapical and basal meristems .
323
Biomassdensity(kg m - z)
Totalnumber ofgrowingfronds
Meanlength ofgrowingfronds (m)
Growthcurvea
Biomassproductionb(wet g m -2 d-1 )
20 April-3 May 8 .4 1060 3 .7 rapid 1444-17 May 8 .3 980 5 .1 slow 11818-24 May 8 .5 860 4 .7 rapid 12725 May-7 June 8 .8 690 4 .0 slow 79HARVEST17-23 June 6.8 610 3 .8 rapid 83

324
Biomass production by plants within the experi-mental enclosure varied over short time periods,partly as a result of growth rate fluctuations (Ta-ble 1). Changes in population structure also con-tributed to changes in biomass production. Loss ofmeristematic activity in large fronds, due to break-age or terminal blade formation, and low frondinitiation rate resulted in a gradual decline in totalnumber of growing fronds, mean length of growingfronds, and biomass production . Biomass produc-tion is influenced by the size distribution of growingfronds, because large fronds with intact apical mer-istems usually increase in length and weight morerapidly than small fronds (North 1971 ; Gerard1976; Lobban 1978) .
Biomass was lost from plants within the experi-mental enclosure at a relatively constant rate, prim-arily by senescence of fronds cut during the pre-transplant harvesting . Some additional tissue waslost due to an undetermined disease which causedperforations like shot-holes in the laminae . Naturalpopulations typically show much greater temporalfluctuations in biomass loss, owing to effects ofstorms (Gerard 1976) and less frequently to nutrientstarvation (Jackson 1977) . Such major losses wouldbe much more important to biomass density andharvestable yield than was demonstrated by theunusually protected population used in this study .
Harvestable yield of plants within the experimen-tal enclosure was considerably lower than biomassproduction . Portions of harvested fronds remain-ing below canopy depth account for much of thisdifference. The empirically determined yield valuecould be extrapolated to an annual yield estimatefor the population if relatively constant year-roundrates of biomass production and loss were assumed .The short-term fluctuations in frond growth rateand biomass production demonstrated by theexperimental kelp population indicate the inaccu-racy of such an assumption. Further extrapolationof the yield measurement for this population underthis specific set of environmental conditions to yieldpredictions for other populations under differentconditions would be even less reliable .
An alternative method of predicting yields for
commercially harvested kelp populations is to de-fine relationships between growth parameters andpotential limiting factors, then use site-specific en-vironmental information to predict growth, bio-mass production and loss, and yield. Data on empir-ical growth, production, and yield, like thosepresented herein, could be used to test the accuracyof the predictions . We have recently begun work onthis task and look forward to the development of ourpredictive capabilities over the coming years .
Acknowledgements
We thank the Marine Biomass Project crew atKerckhoff Marine Laboratory for their efforts andendurance. M . Carr showed particular dedication tothe experimental enclosure . Dr . H. Kirkman assist-ed in field operations . D. Early computerized vo-luminous environmental data . Various GeneralElectric Company employees were responsible forthe operational reliability of the experimental en-closure. Funding was provided by the Gas ResearchInstitute .
References
Boltz, D . F., 1958 . Colorimetric determination of nonmetals .lnterscience Publ ., 321 pp .
Gerard, V . A ., 1976 . Some aspects of material dynamics andenergy flow in a kelp forest in Monterey Bay, California .Ph.D . Thesis, Univ. Calif., Santa Cruz, 173 pp .
Gerard, V . A ., 1982 . Growth and utilization of internal nitrogenreserves by the giant kelp Macro cystis pyrifera in a low nitro-gen environment . Mar. Biol . 66 : 27-35 .
Jackson, G . A ., 1977 . Nutrients and production of giant kelp,Macrocystis pyrifera, off southern California. Limnol .Oceanogr. 22: 979-995 .
Lobban, C . S ., 1978 . The growth and death of the Macrocystissporophyte (Phaeophyceae, Laminariales). Phycologia 17 :196-212 .
North, W . J ., 1971 . Growth of the mature giant kelp . NovaHedwigia 32 : 123-168 .
Zimmerman, R . C ., 1983 . Seasonal patterns in the productivityof a giant kelp (Macrocystis pyrifera) forest : the effect ofnutrient availability . Ph .D . Thesis, Univ. S . Calif ., 182 pp .