mathematical models of neospora caninum infection in dairy cattle: transmission and options for...
TRANSCRIPT

Mathematical models of Neospora caninum infection indairy cattle: transmission and options for control
N.P. Frencha, *, D. Clancyb, H.C. Davison c, A.J. Trees c
aDepartment of Clinical Veterinary Science, University of Liverpool, Leahurst, Neston, South Wirral, UKbStatistics and Operational Research Division, Department of Mathematical Sciences, University of Liverpool, Liverpool, UK
cVeterinary Parasitology, Liverpool School of Tropical Medicine/Faculty of Veterinary Science, University of Liverpool, Pembroke
Place, Liverpool, UK
Received 15 April 1999; received in revised form 21 July 1999; accepted 21 July 1999
Abstract
The transmission and control of Neospora caninum infection in dairy cattle was examined using deterministic andstochastic models. Parameter estimates were derived from recent studies conducted in the UK and from thepublished literature. Three routes of transmission were considered: maternal vertical transmission with a highprobability (0.95), horizontal transmission from infected cattle within the herd, and horizontal transmission from an
independent external source. Putative infection via pooled colostrum was used as an example of within-herdhorizontal transmission, and the recent ®nding that the dog is a de®nitive host of N. caninum supported theinclusion of an external independent source of infection. The predicted amount of horizontal transmission required
to maintain infection at levels commonly observed in ®eld studies in the UK and elsewhere, was consistent with thatobserved in studies of post-natal seroconversion (0.85±9.0 per 100 cow-years). A stochastic version of the model wasused to simulate the spread of infection in herds of 100 cattle, with a mean infection prevalence similar to that
observed in UK studies (around 20%). The distributions of infected and uninfected cattle corresponded closely toNormal distributions, with S.D.s of 6.3 and 7.0, respectively. Control measures were considered by altering birth,death and horizontal transmission parameters. A policy of annual culling of infected cattle very rapidly reduced the
prevalence of infection, and was shown to be the most e�ective method of control in the short term. Not breedingreplacements from infected cattle was also e�ective in the short term, particularly in herds with a higher turnover ofcattle. However, the long-term e�ectiveness of these measures depended on the amount and source of horizontalinfection. If the level of within-herd transmission was above a critical threshold, then a combination of reducing
within-herd, and blocking external sources of transmission was required to permanently eliminate infection. # 1999Australian Society for Parasitology Inc. Published by Elsevier Science Ltd. All rights reserved.
Keywords: Cattle; Horizontal and vertical transmission; Mathematical models; Neospora caninum
1. Introduction
Neospora caninum infection in dairy cattle is
now recognised as a major cause of abortion and
International Journal for Parasitology 29 (1999) 1691±1704
0020-7519/99/$20.00 # 1999 Australian Society for Parasitology Inc. Published by Elsevier Science Ltd. All rights reserved.
PII: S0020-7519(99 )00131-9
* Corresponding author. Tel.: +44-151-794-6031; fax: +44-
151-794-6028.
E-mail address: [email protected] (N.P. French)

economic loss to farmers worldwide [1±4]. Thishas stimulated research in a number of importantareas, including: quantifying the impact of N.caninum infection on production [5±8], develop-ing tools for identifying infected cattle [9] andimproving our understanding of the life-cycle ofthe parasite [10]. In this study we use a theoreti-cal modelling approach, based on experimentaland ®eld data, to examine the transmission of N.caninum in herds of dairy cattle and explore theimpact of a range of control options.
A previous modelling study of N. caninuminfection in closed dairy herds has explored therole of horizontal and vertical transmission in themaintenance of infection and the e�ects of pre-ferential selection of family lines on the trans-mission process (NP French, HC Davison, DClancy, M Begon, AJ Trees. Modelling ofNeospora species infection in dairy cattle: the im-portance of horizontal and vertical transmissionand di�erential culling. Proceedings of theSociety for Veterinary Epidemiology andPreventive Medicine, Ennis, 1998;113±22). Theconclusions of this work were that: (1) verticaltransmission alone would not sustain the infec-tion in a herd of cattle; and (2) in the absence ofany other route of transmission, preferentialselection of infected family lines would result in apotentially large, but transient, increase in theprevalence of infection.
Since this initial study was conducted, therehave been a number of developments that havegiven new insight into the epidemiology of the
disease. There is evidence of two potentialsources of horizontal transmission; via a caninede®nitive host [10] and cow-to-calf transmissionvia pooled colostrum or milk [11]. Furthermore,studies in the UK have given more precise esti-mates of transmission parameters (see [12], thisissue, for further details). This paper attempts touse our current understanding of this disease tomodify and improve existing models. Thisapproach is intended to complement and informfuture studies and be part of the long term goalof reducing the impact of N. caninum infectionon cattle production.
2. The basic deterministic model
2.1. Model structure
The model uses a system of non-linear di�eren-tial equations to describe the transmission of N.caninum infection between dairy cattle. Its beha-viour is explored using analytical and numericalmethods. Fig. 1 shows the structure of themodel. Our current understanding of the diseasesuggests that cattle are either persistently infected(Y) or uninfected (X), and there is no evidence ofa recovered or immune class. The birth rate par-ameter, r, is the rate at which female cattle areadded to the milking herd, and is r1 for infectedand r2 for uninfected cattle. Importantly, this isnot simply the birth rate of cattle in the herd, asthis would also include male calves and beef
Fig. 1. Schematic representation of a deterministic model of N. caninum.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±17041692

crosses. For simplicity, no time delay wasincluded to allow for the period between birthand ®rst calving, which is usually between 2 and3 years, although numerical simulations includingsuch a delay had little e�ect on the behaviour ofthe model. The death rate parameter m is the rateat which cattle are culled or removed from theherd, and is m1 for infected and m2 for uninfectedcattle. Both r and m are given values that areconsistent with national herd demographics andherd age structures in the UK. In order to modelthe impact of parasite virulence on both birth(e.g. abortion) and death (mortality and culling)processes, the birth and death parameters forinfected cattle are ®xed and the culling rate isalways greater than the birth rate m1>r1. Herdsize is then maintained by allowing the netgrowth of the uninfected population to vary.These demographic processes are consistent withthose observed in longitudinal ®eld studies [7].The e�ect of preferentially selecting infectedfamily lines has been described elsewhere, and isnot included in this study (NP French, HCDavison, D Clancy, M Begon, AJ Trees.Modelling of Neospora species infection in dairycattle: the importance of horizontal and verticaltransmission and di�erential culling. Proceedingsof the Society for Veterinary Epidemiology andPreventive Medicine, Ennis, 1998;113±22).
2.2. Routes of transmission
We consider three routes of transmission ofwhich only one, vertical transmission, has beenproven to occur in naturally infected cattle.
2.2.1. Vertical transmissionVertical transmission is de®ned here as the
transfer of infection from dam to calf via thetransplacental route. The probability of verticaltransmission (f) used in all models is 0.95, basedon a recent study conducted in the UK [12].Other studies have estimated this parameter to be0.81 [8] and 1.00 [6].
2.2.2. Horizontal transmission dependent on herdinfection prevalence: infection from within the herd
Here the force of infection is a function of cur-rent infection prevalence and is modelled using a`true' mass action process with a parameter b.The putative spread of infection via pooled colos-trum or milk [11] would be an example of such atransmission process whereby the rate of newinfections would depend on the current pro-portion of infected animals in the herd.
2.2.3. Horizontal transmission independent of herdinfection prevalence: infection from outside theherd
This type of transmission process is modelledusing a constant per-capita force of infectionwith a parameter t. An independent cycle ofinfection in another host [10, 13] would be anexample of this type of transmission.
At present, we have no information on therelative contribution of di�erent sources of hori-zontal infection, so neither b nor t can be givenreliable values based on either ®eld or experimen-tal data. However, we do have estimates of therate of post-natal seroconversion [12, 6]. Thesecan be used to explore the criteria necessary formaintaining stable equilibrium prevalences andto examine the role and behaviour of potentialsources of horizontal transmission.
2.3. Model equations
The model (Fig. 1) can be described by the sys-tem of di�erential equations:
dY=dt � r1fYÿ m1Y� bXY=N� tX, �1�
dX=dt � �r2 ÿ m2�X� r1�1ÿ f�Yÿ bXY=Nÿ tX,
where the net growth of the population ofuninfected cattle (r2ÿm2) is allowed to vary:
r2 ÿ m2 � ��m1 ÿ r1�Y �=X: �2�
Equivalently, we can model the prevalence ofinfection y:
dy=dt � r1fyÿ m1y� by�1ÿ y� � t�1ÿ y�, �3�where x=1ÿy.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±1704 1693
The basic reproductive ratio, Ro, for Eq. (1) is:
Ro � �r1f� b� tN �=m1, �4�which is the sum of the vertical and horizontaltransmission components divided by the averagelifespan of an infected individual.
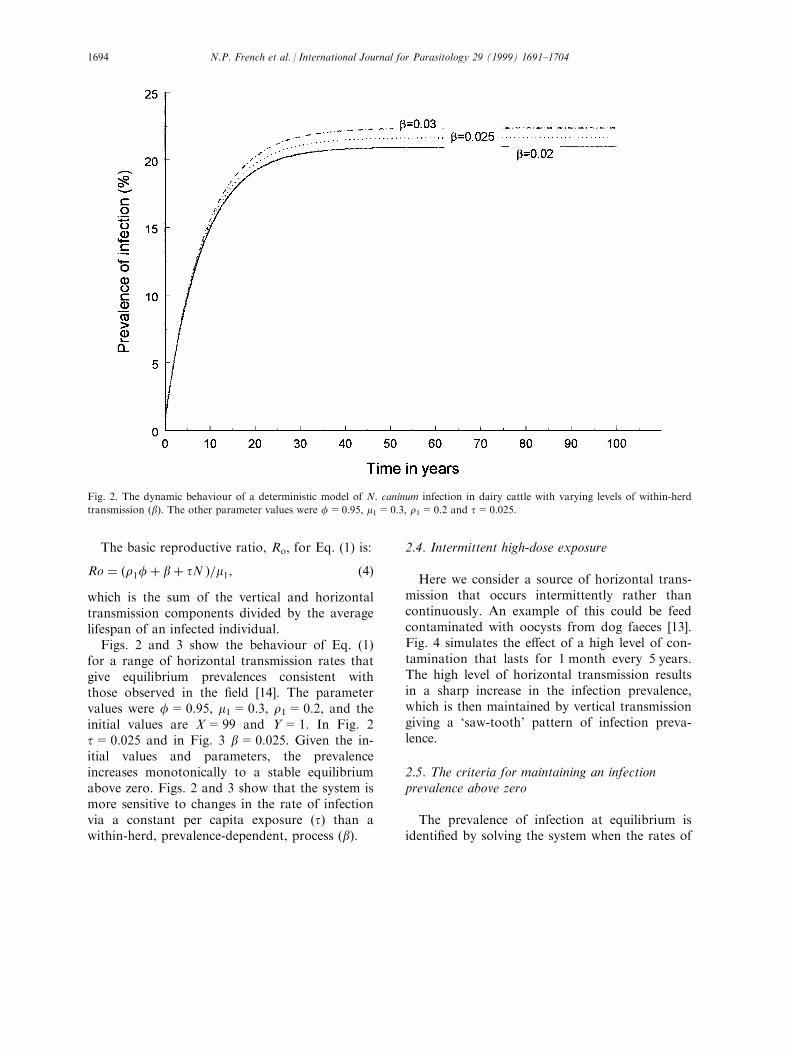
Figs. 2 and 3 show the behaviour of Eq. (1)for a range of horizontal transmission rates thatgive equilibrium prevalences consistent withthose observed in the ®eld [14]. The parametervalues were f=0.95, m1=0.3, r1=0.2, and theinitial values are X=99 and Y=1. In Fig. 2t=0.025 and in Fig. 3 b=0.025. Given the in-itial values and parameters, the prevalenceincreases monotonically to a stable equilibriumabove zero. Figs. 2 and 3 show that the system ismore sensitive to changes in the rate of infectionvia a constant per capita exposure (t) than awithin-herd, prevalence-dependent, process (b).
2.4. Intermittent high-dose exposure
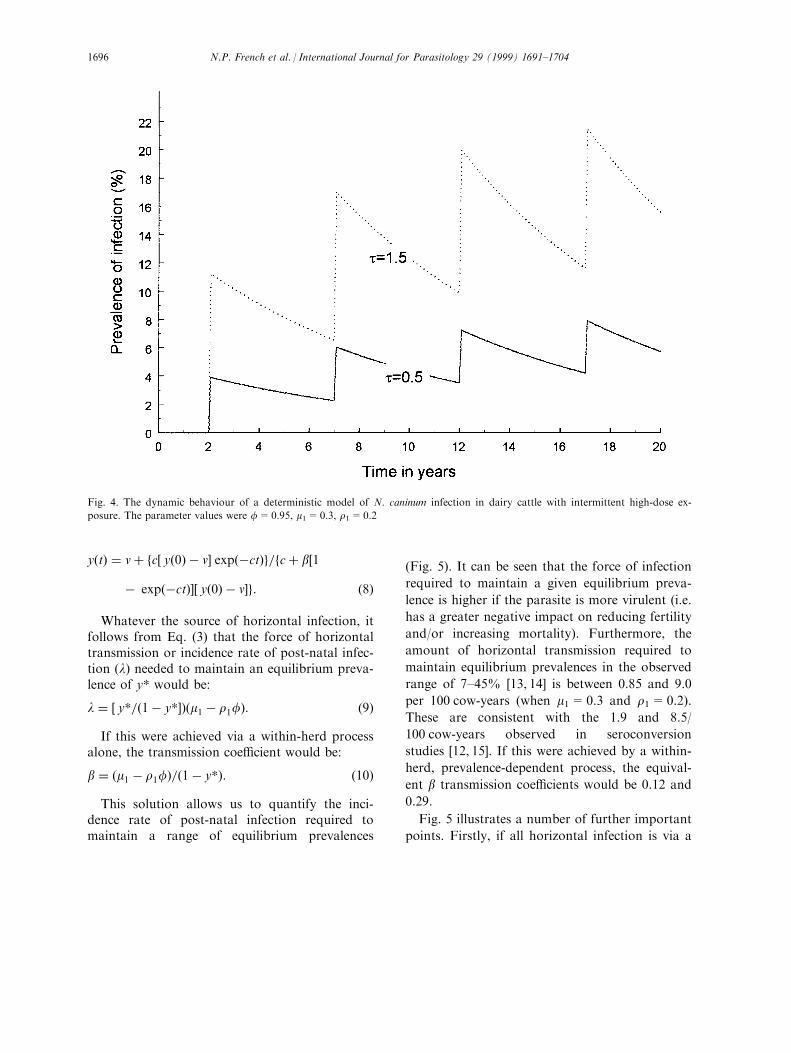
Here we consider a source of horizontal trans-mission that occurs intermittently rather thancontinuously. An example of this could be feedcontaminated with oocysts from dog faeces [13].Fig. 4 simulates the e�ect of a high level of con-tamination that lasts for 1 month every 5 years.The high level of horizontal transmission resultsin a sharp increase in the infection prevalence,which is then maintained by vertical transmissiongiving a `saw-tooth' pattern of infection preva-lence.
2.5. The criteria for maintaining an infectionprevalence above zero
The prevalence of infection at equilibrium isidenti®ed by solving the system when the rates of
Fig. 2. The dynamic behaviour of a deterministic model of N. caninum infection in dairy cattle with varying levels of within-herd
transmission (b). The other parameter values were f=0.95, m1=0.3, r1=0.2 and t=0.025.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±17041694
change are set to zero (dY/dt=dX/dt=0). If theonly route of horizontal transmission was via awithin-herd transmission process, then the preva-lence of infection at equilibrium y* would beeither endemically stable at:
y* � �r1fÿ m1 � b�=b �5�(provided that 0< m1ÿr1f< b)
or y* � 0:
If m1ÿr1f>b, then the equilibrium at y*=0 isstable.
If there is a combination of within-herd andconstant per capita horizontal transmission then,provided that r1f<=m1, the prevalence ofinfection at equilibrium would be the positive
root of the quadratic equation:
ÿb� y�2 � �b� r1fÿ m1 ÿ t�y� t � 0: �6�De®ning
k � r1fÿ m1 � bÿ t
and
c � �p k2 � 4tb�,then
y* � �k� c�=2b or �kÿ c�=2b: �7�
If y(0) is the value of y at time=0, andv=(k+ c)/2b is the positive equilibrium point,then the solution of Eq. (3) is:
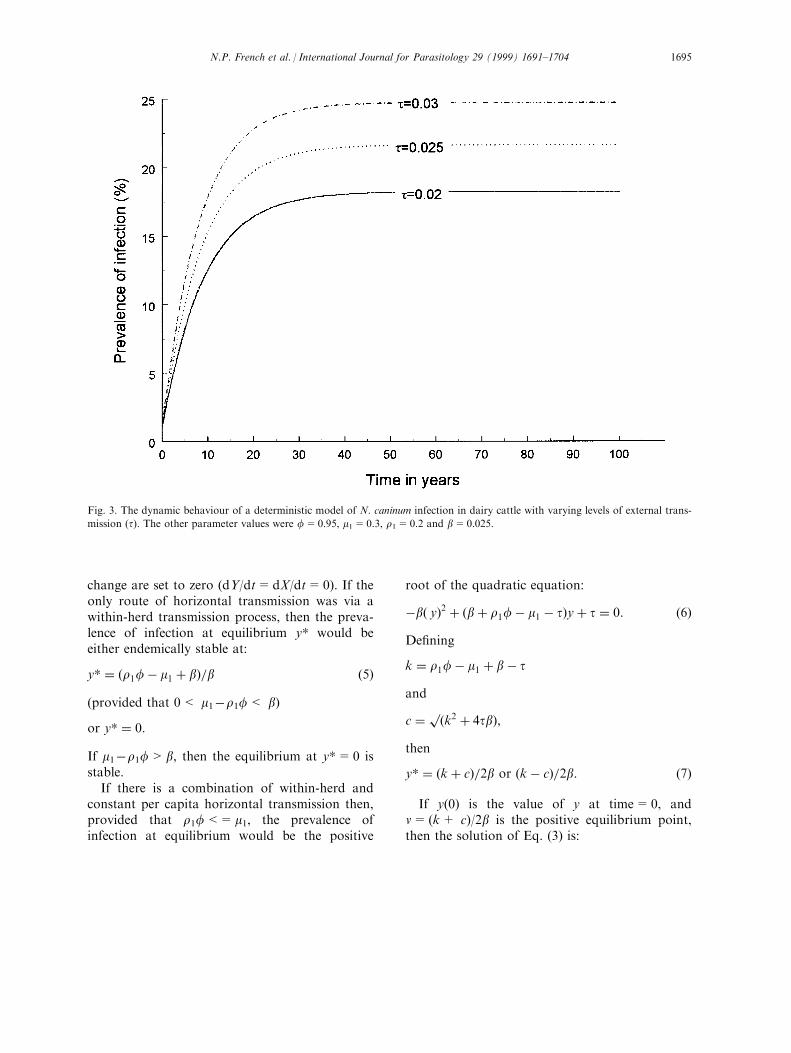
Fig. 3. The dynamic behaviour of a deterministic model of N. caninum infection in dairy cattle with varying levels of external trans-
mission (t). The other parameter values were f=0.95, m1=0.3, r1=0.2 and b=0.025.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±1704 1695
y�t� � v� fc� y�0� ÿ v� exp�ÿct�g=fc� b�1
ÿ exp�ÿct��� y�0� ÿ v�g: �8�
Whatever the source of horizontal infection, itfollows from Eq. (3) that the force of horizontaltransmission or incidence rate of post-natal infec-tion (l) needed to maintain an equilibrium preva-lence of y* would be:
l � � y*=�1ÿ y*���m1 ÿ r1f�: �9�If this were achieved via a within-herd process
alone, the transmission coe�cient would be:
b � �m1 ÿ r1f�=�1ÿ y*�: �10�This solution allows us to quantify the inci-
dence rate of post-natal infection required tomaintain a range of equilibrium prevalences
(Fig. 5). It can be seen that the force of infection
required to maintain a given equilibrium preva-
lence is higher if the parasite is more virulent (i.e.
has a greater negative impact on reducing fertility
and/or increasing mortality). Furthermore, the
amount of horizontal transmission required to
maintain equilibrium prevalences in the observed
range of 7±45% [13, 14] is between 0.85 and 9.0
per 100 cow-years (when m1=0.3 and r1=0.2).
These are consistent with the 1.9 and 8.5/
100 cow-years observed in seroconversion
studies [12, 15]. If this were achieved by a within-
herd, prevalence-dependent process, the equival-
ent b transmission coe�cients would be 0.12 and
0.29.
Fig. 5 illustrates a number of further important
points. Firstly, if all horizontal infection is via a
Fig. 4. The dynamic behaviour of a deterministic model of N. caninum infection in dairy cattle with intermittent high-dose ex-
posure. The parameter values were f=0.95, m1=0.3, r1=0.2
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±17041696
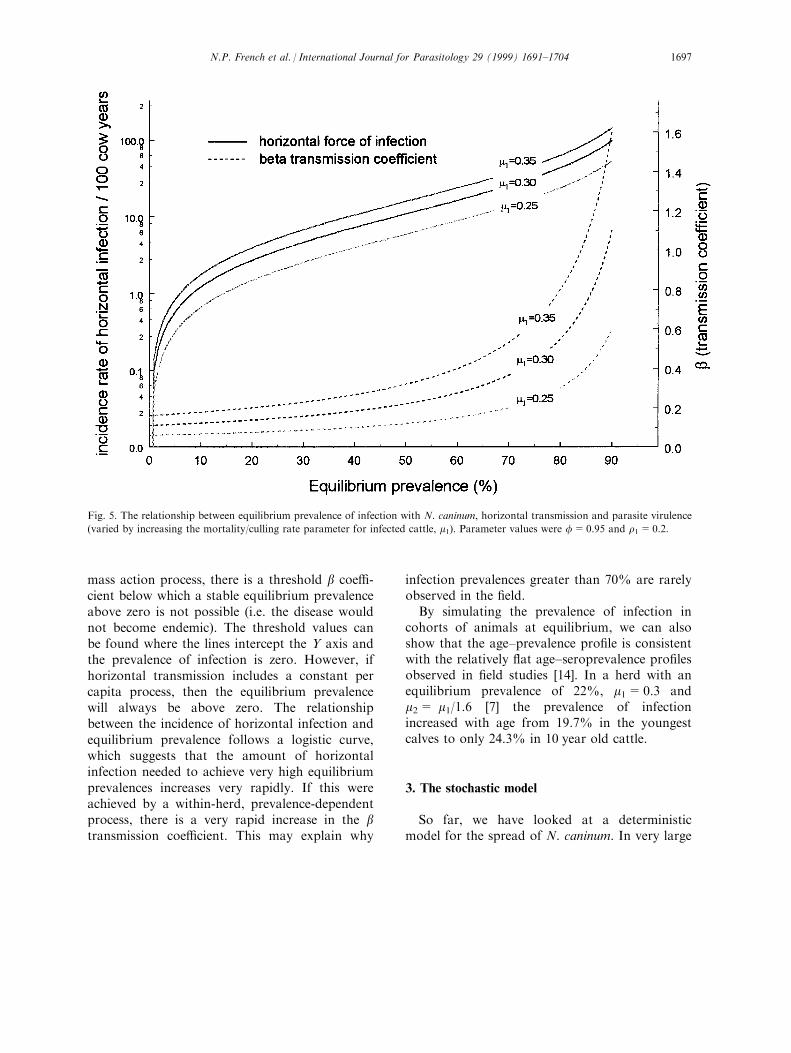
mass action process, there is a threshold b coe�-cient below which a stable equilibrium prevalenceabove zero is not possible (i.e. the disease wouldnot become endemic). The threshold values canbe found where the lines intercept the Y axis andthe prevalence of infection is zero. However, ifhorizontal transmission includes a constant percapita process, then the equilibrium prevalencewill always be above zero. The relationshipbetween the incidence of horizontal infection andequilibrium prevalence follows a logistic curve,which suggests that the amount of horizontalinfection needed to achieve very high equilibriumprevalences increases very rapidly. If this wereachieved by a within-herd, prevalence-dependentprocess, there is a very rapid increase in the btransmission coe�cient. This may explain why
infection prevalences greater than 70% are rarelyobserved in the ®eld.
By simulating the prevalence of infection incohorts of animals at equilibrium, we can alsoshow that the age±prevalence pro®le is consistentwith the relatively ¯at age±seroprevalence pro®lesobserved in ®eld studies [14]. In a herd with anequilibrium prevalence of 22%, m1=0.3 andm2= m1/1.6 [7] the prevalence of infectionincreased with age from 19.7% in the youngestcalves to only 24.3% in 10 year old cattle.
3. The stochastic model
So far, we have looked at a deterministicmodel for the spread of N. caninum. In very large
Fig. 5. The relationship between equilibrium prevalence of infection with N. caninum, horizontal transmission and parasite virulence
(varied by increasing the mortality/culling rate parameter for infected cattle, m1). Parameter values were f=0.95 and r1=0.2.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±1704 1697

populations, deterministic models can provide a
good description of the spread of disease.
However, for a herd of a few hundred animals,
the population is su�ciently small that stochastic
e�ects are important. Therefore, we now consider
a stochastic version of the model described in
Section 2. The rates shown in Fig. 1 are now
interpreted as average transition rates in a ran-
dom process. If the population at time t consists
of X uninfected and Y infected cattle, then in the
small time interval from t to t+Dt, the probabil-
ities of the various possible transitions are as
shown in Table 1.
For the deterministic model, we were able to
ensure that herd size remained constant by allow-
ing r2ÿm2 to vary according to Eq. (2). In our
stochastic model we will assume that all of the
parameters r1, m1, r2, m2, b, t, f, N remain con-
stant over time. The population size will be
allowed to vary between Nmin and Nmax.
Provided that Nmin<=X+ Y<=Nmax, the
population evolves according to the transition
rates given in Table 1. Whenever X+ Y= Nmin
and a death occurs, a new animal is instan-
taneously bought into the herd. This will be an
infected animal with probability P, uninfected
with probability 1ÿP, where P is the probability
that a purchased cow is infected prior to its arri-
val in the herd. Whenever X+ Y= Nmax and a
birth occurs, an animal is instantaneously culled.
This animal is chosen from the herd in such a
way that, with probability X/(X+ rY), a suscep-
tible animal is culled, while with probability rY/
(X+ rY), an infected animal is chosen (r being a
®xed parameter of the model).
Since herd size varies, the within-herd infectionrate bXY/N could perhaps be replaced by bXY/(X+ Y) but, since X+ Y is kept approximatelyconstant, this would make little di�erence, so forsimplicity we use bXY/N with N constant.
Using the same parameter values as in Section2, we take f=0.95, m1=0.3, r1=0.2, t=0.025,b=0.025. From Thurmond and Hietala [7], aplausible value for r is r=1.6. That is to say, theculling rate for infected cattle is 1.6 times thatfor uninfected animals. It therefore also seemssensible to take m2= m1/r=0.1875. We takeP=0.06 from estimates of the prevalence ofinfection in the national herd [16]. In our choiceof r2, we will be guided by Eq. (2). Using thesame equilibrium prevalence as in Section 2(v=0.22), we suppose that at equilibrium,r2ÿm2=(m1ÿr1)v/(1ÿv). We therefore taker2=0.1875+(0.1�0.22/0.78)10.2157, which isconsistent with the observation that uninfectedcattle have a lower risk of abortion comparedwith infected cattle [6]. Finally, we takeNmin=95 and Nmax=105 to ensure a herd sizeof around 100, and N=100.
To investigate the behaviour of the stochasticmodel, Monte Carlo simulation was used. Forthe above parameter values, 5000 simulationswere carried out, each with initial conditionsX=100, Y=0. The mean and S.D. of each of Xand Y, together with the correlation between Xand Y, were recorded at 1 year intervals over aperiod of 100 years. Fig. 6 shows the develop-ment over time of the mean number of infectedcattle in the herd, together with the mean totalherd size. An idea of the amount of randomvariability is given by the curves plotted two
Table 1
The probabilities of transition from one state to another in a stochastic model of N. caninum infection in dairy cattle
Event Probability
A susceptible gives birth to a susceptible (X, Y) 4 (X+1, Y) r2X DtAn infected gives birth to a susceptible (X, Y) 4 (X+1, Y) r1(1ÿf)Y DtAn infected gives birth to an infected (X, Y) 4 (X, Y+1) r1fY DtA susceptible dies or is culled (X, Y) 4 (Xÿ1, Y) m2X DtAn infected dies or is culled (X, Y) 4 (X, Yÿ1) m1Y DtAn infection occurs from outside the herd (X, Y) 4 (Xÿ1, Y+1) tX DtAn infection occurs from within the herd (X, Y) 4 (Xÿ1, Y+1) b(XY/N) Dt
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±17041698
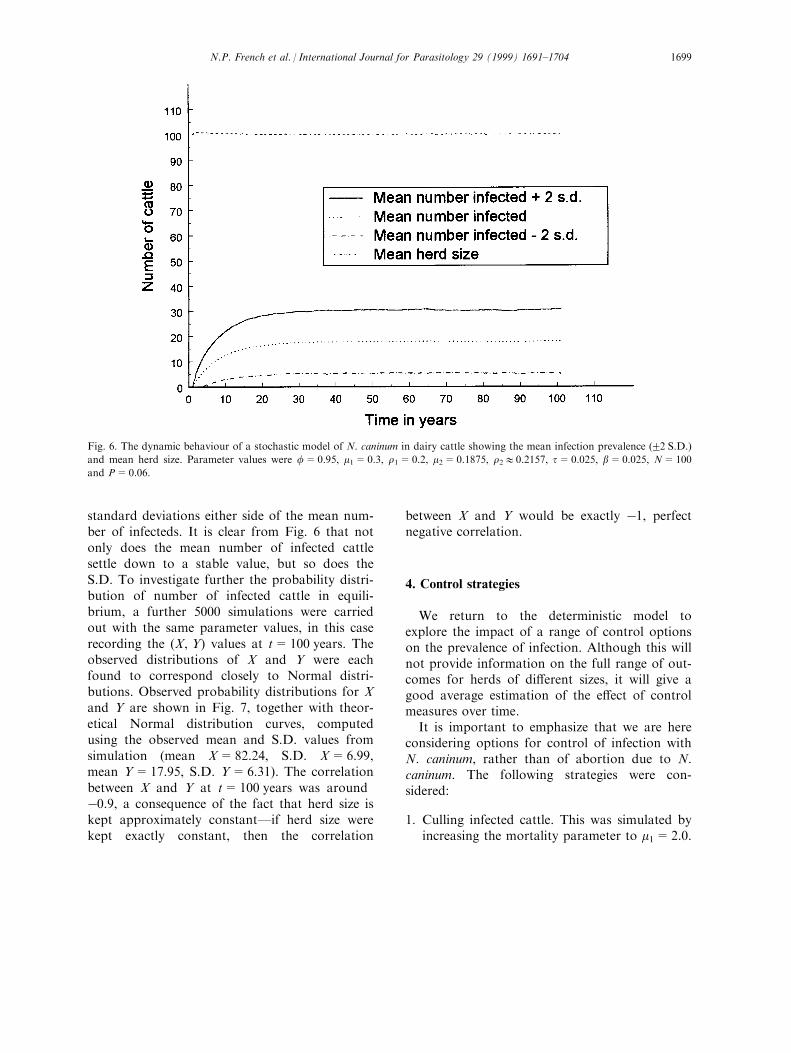
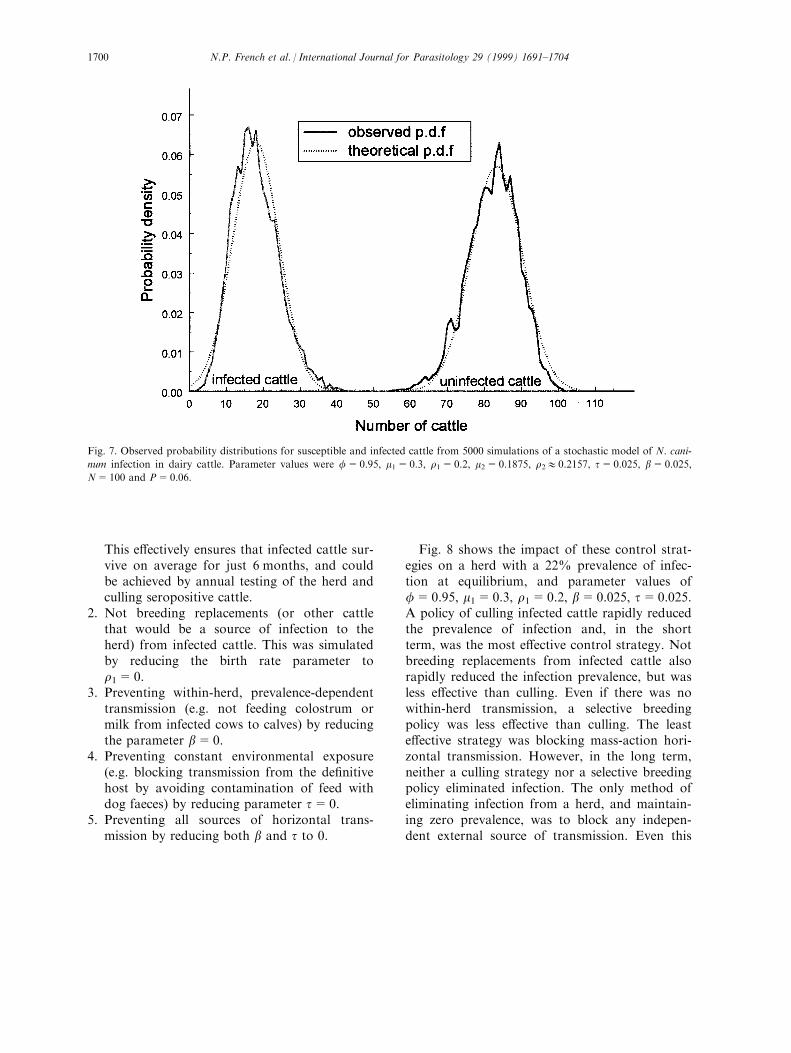
standard deviations either side of the mean num-ber of infecteds. It is clear from Fig. 6 that notonly does the mean number of infected cattlesettle down to a stable value, but so does theS.D. To investigate further the probability distri-bution of number of infected cattle in equili-brium, a further 5000 simulations were carriedout with the same parameter values, in this caserecording the (X, Y) values at t=100 years. Theobserved distributions of X and Y were eachfound to correspond closely to Normal distri-butions. Observed probability distributions for Xand Y are shown in Fig. 7, together with theor-etical Normal distribution curves, computedusing the observed mean and S.D. values fromsimulation (mean X=82.24, S.D. X=6.99,mean Y=17.95, S.D. Y=6.31). The correlationbetween X and Y at t=100 years was aroundÿ0.9, a consequence of the fact that herd size iskept approximately constantÐif herd size werekept exactly constant, then the correlation
between X and Y would be exactly ÿ1, perfectnegative correlation.
4. Control strategies
We return to the deterministic model toexplore the impact of a range of control optionson the prevalence of infection. Although this willnot provide information on the full range of out-comes for herds of di�erent sizes, it will give agood average estimation of the e�ect of controlmeasures over time.
It is important to emphasize that we are hereconsidering options for control of infection withN. caninum, rather than of abortion due to N.caninum. The following strategies were con-sidered:
1. Culling infected cattle. This was simulated byincreasing the mortality parameter to m1=2.0.
Fig. 6. The dynamic behaviour of a stochastic model of N. caninum in dairy cattle showing the mean infection prevalence (22 S.D.)
and mean herd size. Parameter values were f=0.95, m1=0.3, r1=0.2, m2=0.1875, r210.2157, t=0.025, b=0.025, N=100
and P=0.06.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±1704 1699
This e�ectively ensures that infected cattle sur-vive on average for just 6 months, and couldbe achieved by annual testing of the herd andculling seropositive cattle.
2. Not breeding replacements (or other cattlethat would be a source of infection to theherd) from infected cattle. This was simulatedby reducing the birth rate parameter tor1=0.
3. Preventing within-herd, prevalence-dependenttransmission (e.g. not feeding colostrum ormilk from infected cows to calves) by reducingthe parameter b=0.
4. Preventing constant environmental exposure(e.g. blocking transmission from the de®nitivehost by avoiding contamination of feed withdog faeces) by reducing parameter t=0.
5. Preventing all sources of horizontal trans-mission by reducing both b and t to 0.
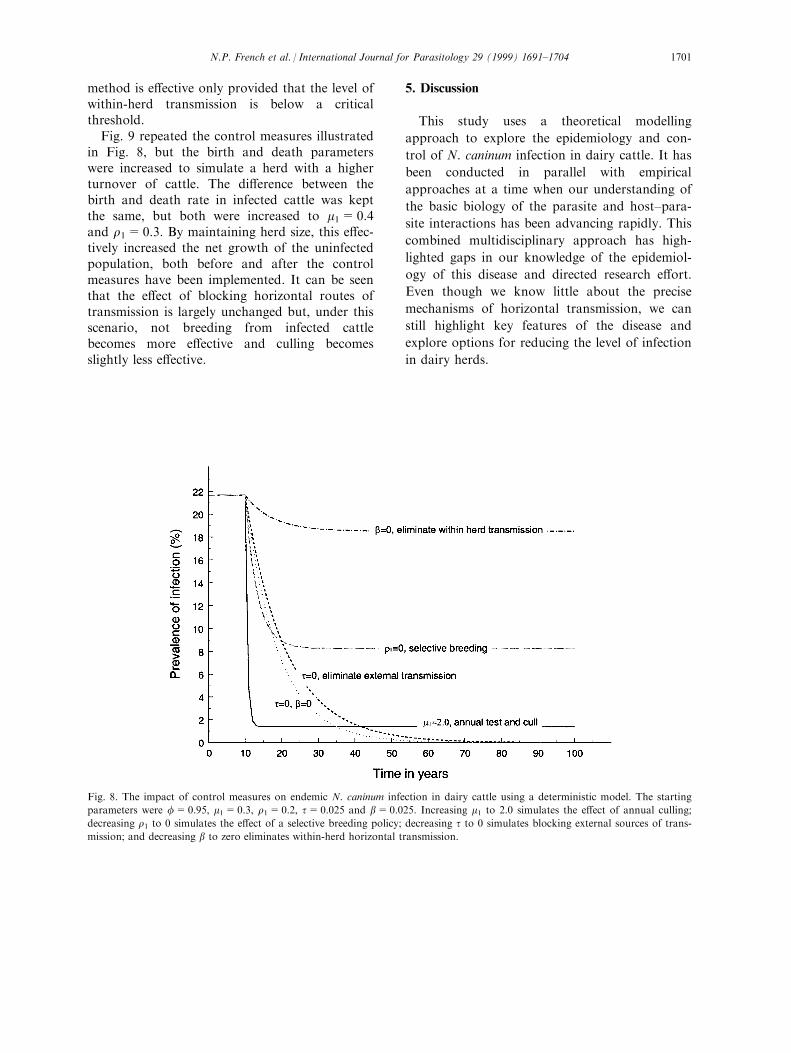
Fig. 8 shows the impact of these control strat-egies on a herd with a 22% prevalence of infec-tion at equilibrium, and parameter values off=0.95, m1=0.3, r1=0.2, b=0.025, t=0.025.A policy of culling infected cattle rapidly reducedthe prevalence of infection and, in the shortterm, was the most e�ective control strategy. Notbreeding replacements from infected cattle alsorapidly reduced the infection prevalence, but wasless e�ective than culling. Even if there was nowithin-herd transmission, a selective breedingpolicy was less e�ective than culling. The leaste�ective strategy was blocking mass-action hori-zontal transmission. However, in the long term,neither a culling strategy nor a selective breedingpolicy eliminated infection. The only method ofeliminating infection from a herd, and maintain-ing zero prevalence, was to block any indepen-dent external source of transmission. Even this
Fig. 7. Observed probability distributions for susceptible and infected cattle from 5000 simulations of a stochastic model of N. cani-
num infection in dairy cattle. Parameter values were f=0.95, m1=0.3, r1=0.2, m2=0.1875, r210.2157, t=0.025, b=0.025,
N=100 and P=0.06.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±17041700
method is e�ective only provided that the level ofwithin-herd transmission is below a criticalthreshold.
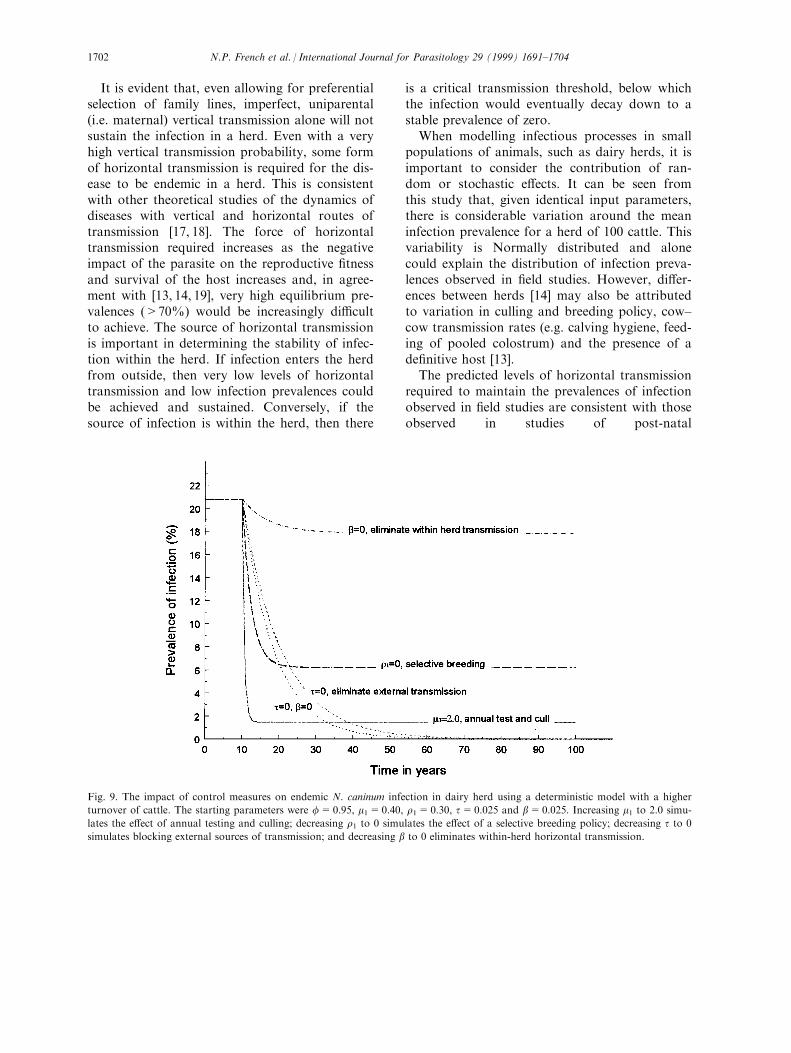
Fig. 9 repeated the control measures illustratedin Fig. 8, but the birth and death parameterswere increased to simulate a herd with a higherturnover of cattle. The di�erence between thebirth and death rate in infected cattle was keptthe same, but both were increased to m1=0.4and r1=0.3. By maintaining herd size, this e�ec-tively increased the net growth of the uninfectedpopulation, both before and after the controlmeasures have been implemented. It can be seenthat the e�ect of blocking horizontal routes oftransmission is largely unchanged but, under thisscenario, not breeding from infected cattlebecomes more e�ective and culling becomesslightly less e�ective.
5. Discussion
This study uses a theoretical modelling
approach to explore the epidemiology and con-
trol of N. caninum infection in dairy cattle. It has
been conducted in parallel with empirical
approaches at a time when our understanding of
the basic biology of the parasite and host±para-
site interactions has been advancing rapidly. This
combined multidisciplinary approach has high-
lighted gaps in our knowledge of the epidemiol-
ogy of this disease and directed research e�ort.
Even though we know little about the precise
mechanisms of horizontal transmission, we can
still highlight key features of the disease and
explore options for reducing the level of infection
in dairy herds.
Fig. 8. The impact of control measures on endemic N. caninum infection in dairy cattle using a deterministic model. The starting
parameters were f=0.95, m1=0.3, r1=0.2, t=0.025 and b=0.025. Increasing m1 to 2.0 simulates the e�ect of annual culling;
decreasing r1 to 0 simulates the e�ect of a selective breeding policy; decreasing t to 0 simulates blocking external sources of trans-
mission; and decreasing b to zero eliminates within-herd horizontal transmission.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±1704 1701
It is evident that, even allowing for preferentialselection of family lines, imperfect, uniparental(i.e. maternal) vertical transmission alone will notsustain the infection in a herd. Even with a veryhigh vertical transmission probability, some formof horizontal transmission is required for the dis-ease to be endemic in a herd. This is consistentwith other theoretical studies of the dynamics ofdiseases with vertical and horizontal routes oftransmission [17, 18]. The force of horizontaltransmission required increases as the negativeimpact of the parasite on the reproductive ®tnessand survival of the host increases and, in agree-ment with [13, 14, 19], very high equilibrium pre-valences (>70%) would be increasingly di�cultto achieve. The source of horizontal transmissionis important in determining the stability of infec-tion within the herd. If infection enters the herdfrom outside, then very low levels of horizontaltransmission and low infection prevalences couldbe achieved and sustained. Conversely, if thesource of infection is within the herd, then there
is a critical transmission threshold, below whichthe infection would eventually decay down to astable prevalence of zero.
When modelling infectious processes in smallpopulations of animals, such as dairy herds, it isimportant to consider the contribution of ran-dom or stochastic e�ects. It can be seen fromthis study that, given identical input parameters,there is considerable variation around the meaninfection prevalence for a herd of 100 cattle. Thisvariability is Normally distributed and alonecould explain the distribution of infection preva-lences observed in ®eld studies. However, di�er-ences between herds [14] may also be attributedto variation in culling and breeding policy, cow±cow transmission rates (e.g. calving hygiene, feed-ing of pooled colostrum) and the presence of ade®nitive host [13].
The predicted levels of horizontal transmissionrequired to maintain the prevalences of infectionobserved in ®eld studies are consistent with thoseobserved in studies of post-natal
Fig. 9. The impact of control measures on endemic N. caninum infection in dairy herd using a deterministic model with a higher
turnover of cattle. The starting parameters were f=0.95, m1=0.40, r1=0.30, t=0.025 and b=0.025. Increasing m1 to 2.0 simu-
lates the e�ect of annual testing and culling; decreasing r1 to 0 simulates the e�ect of a selective breeding policy; decreasing t to 0
simulates blocking external sources of transmission; and decreasing b to 0 eliminates within-herd horizontal transmission.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±17041702

seroconversion [6, 12, 15]. This implies that thedisease can be maintained at endemic equilibriumand, although N. caninum was only recentlydescribed in cattle, it is unlikely to be a long-term transient epidemic that would die out with-out intervention. However, we still need to testthe assumption that cattle that seroconvert haveacquired post-natal infections, have an increasedrisk of abortion and will vertically infect theiro�spring. We also need to determine the sourcesof horizontal infection. There is increasing evi-dence that the dog is a de®nitive host that couldbe an external source of infection to theherd [10, 13]. We have considered this by includ-ing constant and intermittent per-capita sourcesof infection in the model, with the assumptionthat the cycle of infection in the dog is main-tained independently of the herd of cattle.However, the cycle of infection in the dog couldalso be maintained by infection in the dairy herdthrough the consumption of infected placentae orother bovine material. If this was the case, thenthis route of transmission would be a delayedmass-action process dependent on herd infectionprevalence. Clearly, a greater understanding ofthe epidemiology of the disease in farm dogs andother potential wildlife hosts is required.
The ®rst paper describing colostrum as a po-tential source of infection considered it as a formof pseudo-vertical transmission [11]. We havealso considered colostrum and milk from infectedcattle as a source of within-herd horizontal trans-mission due to the common practice of feedingpooled colostrum and milk to neonatal calves,although, for simplicity, we have not modelledthis as an age-dependent process. Other forms ofwithin-herd horizontal transmission could alsooccur; for example, cow to cow transmission viainfected placentae or amniotic ¯uid [20], butfurther studies are needed to test such hypoth-eses.
The most e�ective control policy would needto take into account both the costs and bene®tsof each strategy. However, when considering theimpact of each measure on disease prevalencealone, the culling of infected cattle would appearto be the most e�ective in the short term(although it becomes less e�ective in herds with a
higher turnover of stock). The reason for this is
that culling removes both within-herd horizontal
and vertical sources of infection. The policy of
keeping infected cattle in the herd but not breed-
ing replacements from them would also be e�ec-
tive in the short term (and conversely becomes
more e�ective in herds with a higher turnover of
stock). However, this policy would not be as
e�ective as culling, because it would only directly
block vertical transmission and infected cattle
would remain in the herd for longer periods of
time. In the presence of horizontal transmission
(with the exception of within-herd transmission
below the critical threshold), neither culling nor
selective breeding would eliminate infection from
the herd. Culling would leave a residual level of
infection from external sources such as an inde-
pendent de®nitive host, and a policy of not
breeding from infected cattle would allow an ad-
ditional within-herd source of horizontal infec-
tion. In the presence of horizontal infection,
elimination of infection from a herd would there-
fore require a combination of control options.
The models developed here have provided a
mathematical framework for our understanding
of neosporosis in dairy cattle. They have incor-
porated some of the key features of the disease in
dairy cattle: (1) the very high probability of verti-
cal transmission, (2) the impact on mortality and
reproduction, and (3) the compensatory e�ect on
survival and reproduction of uninfected cattle.
We have also explored the role of horizontal
transmission and highlighted the need to identify
and quantify these sources of infection. The
models have been deliberately kept simple, with
few parameters and an analytical solution. Even
though many simplifying assumptions have been
made, the models behave in a plausible way and
the outputs are consistent with observational
®eld studies. Once we have a greater quantitative
understanding of the epidemiology, it may be
appropriate to build more complex models that
capture the age structure of the herd and describe
the cycles of infection in the putative de®nitive
host.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±1704 1703

Acknowledgements
Many thanks are due to Professor M. Begonfor his help in model development and his com-ments on the manuscript. Dr H. Davison wasfunded by the Ministry of Agriculture, Fisheriesand Food, UK.
References
[1] Anderson ML, Palmer CW, Thurmond MC et al.
Evaluation of abortions in cattle attributable to neos-
porosis in selected dairy herds in California. J Am Vet
Med Assoc 1995;207(9):1206.
[2] Otter A, Je�rey M, Gri�ths IB, Dubey JP. A survey of
the incidence of Neospora caninum infection in aborted
and stillborn bovine fetuses in England and Wales. Vet
Rec 1995;136:602±6.
[3] Thornton RN, Thompson EJ, Dubey JP. Neospora abor-
tion in New-Zealand cattle. New Zealand Vet J
1991;39:129±33.
[4] Boulton JG, Gill PA, Cook RW, Fraser GC, Harper P,
Dubey JP. Bovine neospora abortion in north-eastern
New-South-Wales. Aust. Vet J 1995;72:119±20.
[5] Thurmond MC, Hietala SK. E�ect of Neospora caninum
infection on milk production in ®rst-lactation dairy cows.
J Am Vet Med Assoc 1997;210:672±4.
[6] Thurmond MC, Hietala SK. E�ect of congenitally
acquired Neospora caninum infection on risk of abortion
and subsequent abortions in dairy cattle. Am J Vet Res
1997;58:1381±5.
[7] Thurmond MC, Hietala SK. Culling associated with
Neospora caninum infection in dairy cows. Am J Vet Res
1996;57:1559±62.
[8] Pare J, Thurmond MC, Hietala SK. Congenital
Neospora caninum infection in dairy cattle and associated
calfhood mortality. Can J Vet Res 1996;60:133±9.
[9] Williams DJ, McGarry J, Guy F, Barber J, Trees AJ.
Novel ELISA for detection of Neospora-speci®c anti-
bodies in cattle. Vet Rec 1997;140:328±31.
[10] McAllister MM, Dubey JP, Lindsay DS, Jolley WR,
Wills RA, McGuire AM. Dogs are de®nitive hosts of
Neospora caninum. Int J Parasitol 1998;28:1473±8.
[11] Uggla A, Stenlund S, Holmdahl OJM et al. Oral
Neospora caninum inoculation of neonatal calves. Int J
Parasitol 1998;28:1467±72.
[12] Davison HC, Otter A, Trees AJ. Estimation of vertical
and horizontal transmission parameters of Neospora
caninum in dairy cattle. Int J Parasitol 1999 (this issue).
[13] Pare J, Fecteau G, Fortin M, Marsolais G.
Seroepidemiologic study of Neospora caninum in dairy
herds. J Am Vet Med Assoc 1998;213:1595±8.
[14] Davison HC, French NP, Trees AJ. Herd-speci®c and
age-speci®c seroprevalence of Neospora caninum in 14
British dairy herds. Vet Rec 1999;44:547±50.
[15] Pare J, Thurmond MC, Hietala SK. Neospora caninum
antibodies in cows during pregnancy as a predictor of
congenital infection and abortion. J Parasitol
1997;83:82±7.
[16] Davison HC, Otter A, Trees AJ. Signi®cance of
Neospora caninum in British dairy cattle determined by
estimation of seroprevalence in normally-calving cattle
and aborting cattle. Int J Parasitol, in press.
[17] Lipsitch M, Nowak MA, Ebert D, May RM. The popu-
lation-dynamics of vertically and horizontally transmitted
parasites. Proc R Soc Lond Ser B 1995;260:321±7.
[18] Stringer SM, Hunter N, Woolhouse MEJ. A mathemat-
ical model of the dynamics of scrapie in a sheep ¯ock.
Math Biosci 1998;153:79±98.
[19] Thurmond MC, Hietala SK, Blanchard PC. Herd-based
diagnosis of Neospora caninum-induced endemic and epi-
demic abortion in cows and evidence for congenital and
postnatal transmission. J Vet Diagn Invest 1997;9:44±9.
[20] Ho MS, Barr BC, Tarantal AF et al. Detection of
Neospora from tissues of experimentally infected rhesus
macaques by PCR and speci®c DNA probe hybridiz-
ation. J Clin Microbiol 1997;35:1740±5.
N.P. French et al. / International Journal for Parasitology 29 (1999) 1691±17041704