marine cretaceous-paleogene biofacies and ichnofacies in southern tibet, china, and their...
TRANSCRIPT

ELSEVIER Marine Micropaleontology 32 (1997) 3-29
Marine Cretaceous-Paleogene biofacies and ichnofacies in southern Tibet, China, and their sedimentary significance
Zhou Zhicheng a,*, Helmut Willems b, Zhang Binggao a
a Nanjing Institute of Geology and Palaeontology, Academia Sinica, Nanjing 210008, China ’ Fachbereich 5-Geowissenschaften, Universitiit Bremen, 28334 Bremen, Germany
Received 1 June 1996; accepted 1 December 1996
Abstract
The study of biofacies is of great significance not only for biostratigraphy, but also for the interpretation of sedimentary environments and the reconstruction of paleogeography. The areas of Tuna, Gamba and Tingri of southern Tibet, China, were located in the north margin of the Indian Plate in the Cretaceous and Paleogene. The marine Cretaceous and Paleogene biofacies there can be divided into four groups using the following criteria: (a) the benthic biota and biogenic structures in the sediments; (b) the paleoecology of the main fossils; (c) the genesis of fossils. The four groups are euxinic biofacies, nektonic and planktic biofacies, benthic biofacies and redeposited biofacies, which include 24 main biofacies types. Like biofacies. ichnofacies are important indicators of paleoenvironments. The Cretaceous-Paleogene trace fossils in the Tuna and Gamba areas are attributable to Nereites, Zoophycos, Cruziana and Skolithos ichnofacies. Nereites ichnofacies indicates an open marine basin environment. Zoophycos and Cruziana ichnofacies indicate shelf margin and open sea shelf environments. The Skolithos ichnofacies reflects the environments of sand barrier deposited in the intertidal zone and the upper part of subtidal zone. The Zoophycos ichnofacies can further be subdivided into Thalassinoides-Planolites and Zoophycos-Chondrites subichnofacies which reflect aerobic and disaerobic conditions, respectively. The development of biofacies and ichnofacies in vertical profiles shows the sedimentary history from the Early Cretaceous to the Paleogene in Tuna, Gamba and Tingri areas. A regression occurred in Tuna and Gamba from the Neocomian to the Early Paleocene. The sedimentary environments went from euxinic basin and open marine basin, shelf margin, open sea margin, platform slope, platform margin, open marine platform to the sand barriers deposited in the intertidal zone or the upper part of subtidal zone. A transgression took place at the beginning of the middle Paleocene. The carbonate deposits formed in the open marine platform and open sea shelf environments overlay the sandstones of the Jidula Formation. The termination of marine deposits in these areas began in the Thanetian. The sedimentary history in the Tingri area differed from that in the Tuna and Gamba areas especially during middle Maastrichtian to Early Paleocene times. The sedimentary process in Tingri during this period took place in the lower part, the middle part of a fan and seaward prograding delta plains. The marine deposition in the Tingri area ended in Lutetian later than in the Tuna and Gamba areas.
Keywords: Cretaceous; Paleogene; biofacies; ichnofacies; Southern Tibet, China
* Corresponding author. E-mail: [email protected]
0377-8398/97/$17.00 I@ 1997 Elsevier Science B.V. All rights reserved. PIZ SO377-8398(97)0001 1-X

4 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29
1. Introduction
The Tuna, Gamba and Tingri areas of southern Ti- bet, China were located at the northern margin of the Indian Plate and belonged to the southern subzone of the Tethys-Himalaya during the Cretaceous. Many geologists have been attracted there and a great deal of researches on palaeontology, stratigraphy, sedi- mentology and tectonics has taken place over the last three decades because of the important tectonic location and the developed marine Cretaceous and Paleogene strata (He Yan et al., 1976; Wang Yujing, 1976; Wen Shixuan, 1974; Wan Xiaoqiao, 1987; Xu Yulin et al., 1989; Yin Jixiang, 1988; Liu and Ein- sele, 1994). The authors have made many scientific expeditions to southern Tibet since 1966 and they have obtained a large quantity of field samples; the results on biostratigraphy, carbonate microfacies and ichnology have been published (Zhou Zhicheng and Zhang Binggao, 1992; Willems and Zhang Binggao, 1993a,b; Willems, 1993; Zhang Binggao et al., 1994; Zhou Zhicheng, 1997). This paper is based on the former research work.
The Cretaceous to Paleogene is an important geo- historic time period. The Indian Plate drifted rapidly northwards and collided with the Eurasian Plate. During this event, great changes in paleogeogra-
phy and sedimentary environments took place and gave rise to the development of biofacies and ich- nofacies. This paper describes the main types of Cretaceous and Paleogene marine biofacies and ich- nofacies in detail and suggests their environmental significance. Also on the basis of environmental analysis of the biofacies and ichnofacies, the ma- rine sedimentary history in these areas during the Cretaceous-Paleogene is reconstructed.
2. Materials and methods
The samples for the studies of biofacies come from sections At and Gulu of the Tuna area, sections A, K, D, F, Zp and Zm of the Gamba area and sections L, M, Entong and Longjiang of the Tingri area. The samples and field photos for the studies of ichnofacies mainly come from section At of Tuna and sections A, K, D of Gamba (Fig. 1).
The following criteria for the classification of bio- facies have been taken into consideration: (a) the existence of benthic biota and biogenic structures in sediments (the biofacies is termed euxinic biofacies if there are no benthic fossils and biogenic structures in the sediments); (b) the paleoecologic features of the main fossils (according to this criterion, two groups of biofacies, nektonic and planktic biofacies
1 1 T-T /B E ‘I’ b I
-6amba4 3 - pT,HIMALAYA MTS.
0 30 60 km 1 Y adong
‘i. dj-
Fig. 1. Map showing the localities of sections. I = Section At; 2 = Section Gulu; 3 = Section A; 4 = Section K; 5 = Section Section F, 7 = Section Zp; 8 = Section Zm; 9 = Section L; 10 = Section M; II = Entong Section; 12 = Longjiang Section.
D: 6 =

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 5
and benthic biofacies are classified); and (c) the gen- esis of fossils (the fossils which were derived from resedimentation are referred to redeposited biofa- ties).
The classification of ichnofacies described in this paper is based on the classification by Seilacher (1967) Frey and Seilacher (1980) and Ekdale et al. (1984). The Cretaceous and Paleogene trace fossils can be divided into Nereites, Zoophycos, Cruziana and Skolithos ichnofacies.
When the biofacies and ichnofacies are used to interpret the sedimentary environments, the other factors such as diagenetic features and the textures and structures of rocks are also considered.
3. Stratigraphy
According to Willems and Zhang Binggao (1993a), the marine Cretaceous-Paleogene se- quences in Gamba and Tuna are formed of the Gamba Group (Neocomian-Santonian), the Zongshan For- mation (Campaniart-Maastrichtian), the Jidula For- mation (Maastrichtiar-Paleocene), the Zongpu For- mation (Paleocene-Thanetian) and the Zongpubei Formation (Thanetian).
The Gamba Group is composed of three forma- tions (in ascending order): Formation 1, the Gan- badongshan Formation, at least 760 m thick, con- sisting of siltstones and claystones intercalated by turbiditic sandstones; Formation 2, the Chaquiela Formation, consisting of about 560 m siltstones and claystones interstratified by many sandstone tur- bidites and sandstone dykes, and with increasing intercalations of lilmestones and marlstones in the upper part; Formation 3, the Ganbacunkou Forma- tion consisting of an interstratification of pelagic limestones, marlstones and fewer claystones. The Zongshan Formation with a thickness of at least 220 m is formed by three (in Gamba) or four (in Tuna) limestone packages rich in fossils, separated by cal- careous marlstones and marlstones. The 150-170 m thick Jidula Formation is composed of a sequence of ferruginous sandstones, in the middle part locally interbedded by black limestones and claystones. The Zongpu Formation., which is at least 250 m thick, comprises a variety of well-bedded limestones, but mainly nodular limestones and marlstones. The non- marine Zongpubei Formation with a thickness of at
least 160 m (in Gamba) consists of greenish-grey marlstones and red clay and siltstones.
Comparing the areas of Gamba and Tingri, some differences in the lithostratigraphic sequence can be recognized (Willems and Zhang Binggao, 1993b). In Tingri, the marine Cretaceous and Paleogene include the Gamba Group (Vraconian-Santonian), the Zongshan Formation (Santonian-Maastrichtian), the Zhepure Shanpo Formation (Maastrichtian- Paleocene), the Jidula Formation (Paleocene) and the Zhepure Shan Formation (Paleocene-Eocene). The Gamba Group with an investigated thickness of at least 625 m consists of marlstones which are interca- lated by continuously increasing layers of calcareous marlstones and limestones at the top. In compari- son to the type locality at Gamba, the Zongshan Formation is cut off by an erosional unconformity. The following 225 m thick Zhepure Shanpo For- mation in its lower part is composed of marlstones interbedded by turbiditic sandstones (often with ‘fin- ing upward’ cycles), and in the upper part it consists of mixed carbonatic siliciclastic resediments. The Jidula Formation consists of about 100 m of cal- careous sandstones intercalated by nodular marly sandstones and glauconitic sandstones. The 440 m thick Zhepure Shan Formation forming the summit of the Zhepure Mountain Range can be subdivided into five members (in ascending order): Dolomitic limestones and dolor&es (Member I); thick-bedded limestones (members II-III); nodular limestones and calcareous marlstones (Member IV); and very mas- sive limestones (Member V).
4. Biofacies
The 24 biofacies described in this paper do not cover all biofacies types, but are representative of the main biofacies types in the Cretaceous-Paleogene strata in the Tuna, Gamba and Tingri areas of southern Tibet. The environmental interpretation of the biofa- ties can provide the most important information about the sedimentary features and the paleogeography.
4.1. Euxinic biofacies
4.1.1. BF-I euxinic biofacies The euxinic biofacies coincides with the Letal-
pantostrat biofacies of Schafer (1963). There are no

6 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29
benthic organisms or biogenic sedimentary struc- tures in the continuous sedimentary sequence. This biofacies reflects a kind of stagnant, anaerobic and reducing condition which was not a suitable habi- tat for the benthic organisms. The black shale and mudstone in the Formation 1 of the Gamba Group contains this biofacies.
the limestones indicates depositional conditions with good water circulation and oxygenation.
4.2.2. BF-3 calcispheres (Plate I, 2)
4.2. Nektonic and planktic biofacies
4.2.1. BF-2 planktic foraminifers (Plate I, 1)
The BF-3 is characterized by predominant cal- cispheres and coexisting calpionellids. The former consist mainly of Pithonella sphaerica (Kaufmann) and P. ovalis (Kaufmann) (Plate I, 2). The latter Calpionellidae Bonet, includes Calpionella Lorenz, Stenosemellopsis Colom, Tintinnopsella Colom, En- dothicollaria Zhang et al. (Plate I, 3).
The fauna is mainly composed of planktic foramin- ifers (Heterohelix sp., Globotruncana, Dicarinella, Rotalipora, Hedbergella, Whiteinella, etc). The other components include a small quantity of smaller ben- thic foraminifers (Textularia), calcispheres, calpi- onellids and some fossil fragments such as echino- derms, molluscs, inoceramids and ostracodes derived from shallow water environments. The fossils occur in strongly bioturbated wackestones.
The other fossils consist of a small number of planktic and smaller benthic foraminifers and some fossil fragments which came from shallow water environments.
This biofacies is indicative of the environments with a good water circulation including the shelf margin, open sea shelf and platform slope.
4.2.3. BF-4$laments (Plate I, 4) This biofacies occurs in the environments of shelf Filaments originate from juvenile planktic or
margin, open sea shelf (Tuna and Gamba) and open pseudoplanktic larval stages of pelecypods (Fhigel, marine basin (Tingri) which are under the normal 1982). Wilson (1975) interpreted the filamentous wave base (Figs. 2-4). The strong bioturbation in micrite as ‘offshore basinal environment’. The bio-
FZ FACIES ZONES AND SUB -ZONES
: CUT - OFF LAGOONS, C
7 B RESTRICTED LAGOONS
A QUARTZ SAND BARRIERS
NOUSLY INFLUE PLATFORM PLATFORM
;.,
$px$~~ 4
* . . . . . . . . _r;. .:.. . PLATFORM SLOPE / PROXIMAL CONTINENTAL SLOPE
3 OPEN SEA SHELF (::~t~::::::::::..::[ffl 2 . . . . . . . . . . . . . . . 1
SHELF MARGIN / DISTAL CONTINENTAL SLOPE
Fig. 2. Summary of the eight facies zones (FZ) and further sub-zones differentiated in the facies models of Figs. 3 and 4 (after Willems, 1993).
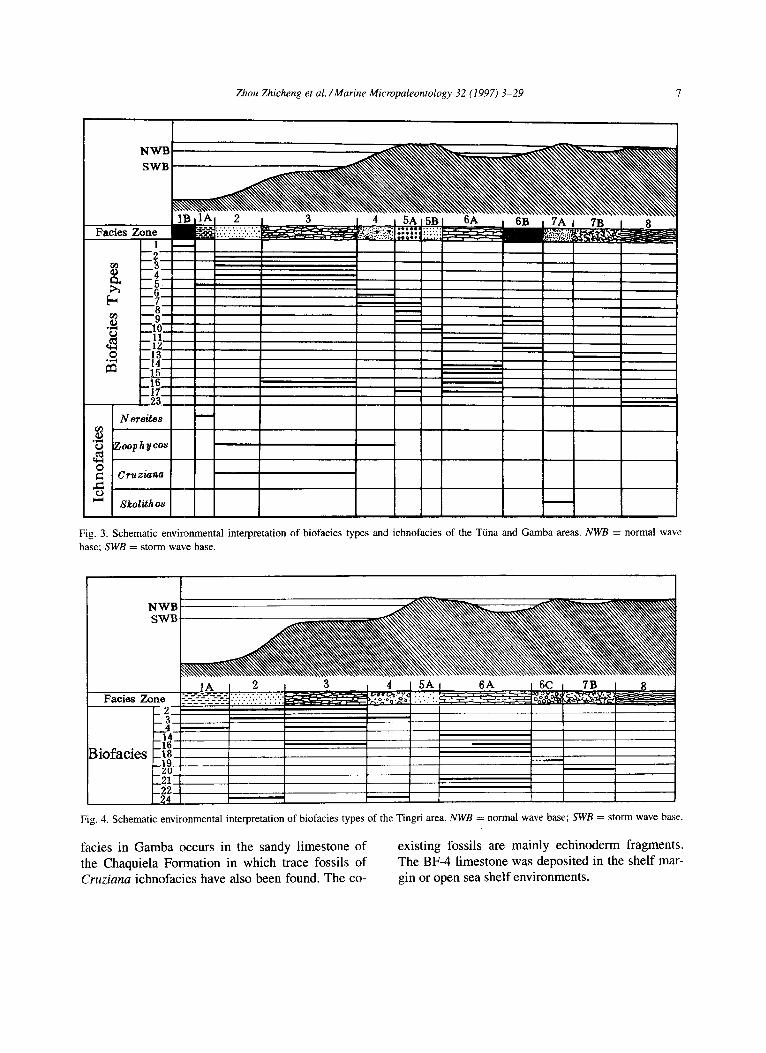
Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 7
NWBI SWB
M .4
2 zcwphyc~
I I I I H
Fig. 3. Schematic environmental interpretation of biofacies types and ichnofacies of the Tiina and Gamba areas. NWB = normal wave base; SWB = storm wave base.
NWB sWB
Fig. 4. Schematic environmental interpretation of biofacies types of the Tingri area. NWB = I normal wave base: SWB = storm wave base.
facies in Gamba occurs in the sandy limestone of the Chaquiela Formation in which trace fossils of Cruziana ichnofacies have also been found. The co-
existing fossils are mainly echinoderm fragments. The BF-4 limestone was deposited in the shelf mar- gin or open sea shelf environments.

8 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 9
The filamental biofacies in Tingri has been found in the marls at the upper part of the Gamba Group. The biota is briefly comprised of filaments, calci- spheres and planktic foraminifers, and the BF-4 is intercalated in BF-2 in vertical section. The biofa- ties indicates an open sea basin or open sea shelf environments.
4.2.4. BF-5 ammonites Ammonites are important fossils in Cretaceous
biostratigraphy. They are mostly found in the detrital rocks and marls of the Gamba Group. The mi- nor biota include bivalves, gastropods, echinoderms, belemnites and planktic foraminifers.
Trace fossils of Nereites and Cruziana ichnofacies have also been found in the deposits which often contain the ammoritic biofacies. The biofacies and the coexisting ichnofacies reflect the depositional environments of open marine basin, shelf margin and open sea shelf.
4.3. Benthic biofacies
4.3.1. BF-6 Goupillaudina-Sulcoperculina (Plate ZZ, 1)
The biota are characterized by the simultaneous occurrence of benthic organisms and planktic or- ganisms. The former include benthic foraminifers Goupillaudina, Sulcoperculina (Plate II, 1) and Ne- ojlabellina and other fossils such as molluscs, ostra- codes, brachiopods and echinoderms. The latter are composed of planktic foraminifers (Globotruncana), calcispheres, calpionellids and filaments. The fos- sils are found in strongly bioturbated wackestone, packstone and floatstone.
The composition of the biota shows that the biofa- ties is the typical transitional type of the benthic and planktic biofacies. The depositional environment was situated on the platform slope between the normal wave base and the storm wave base.
4.3.2. BF-7 Orbitoides-Sulcoperculina (Plate ZZ, 2) The benthic foraminifers include Orbitoides, SuZ-
coperculina, Goupillaudina, Miliolidae, Textularia and Rotaliidae. The other biota include echinoderms, molluscs, brachiopods, ostracodes and bryozoans. Some planktic foraminifers and calcispheres are also present. The bioclasts with micrite envelopes occur in the bioturbated wackestone, grainstone and pack- stone. The limestone also contains some fine-grained well rounded quartz.
BF-7 indicates shallower environments on the platform slope and high-energy shoals located at the platform margin.
4.3.3. BF-8 Orbitoides-MiZioZidae The biofacies is characterized by high diversity
of benthic biota including foraminifers (Orbitoides, Miliolidae, Sulcoperculina, Goupillaudina, Textu- Zaria), echinoderms, brachiopods and Rhodophyceae (Archaeolithothamnium). A few planktic foraminifers and calcispheres are also found in the limestone.
The features of biota and diagenic alternation indicate that sedimentation took place in the high- energy shoals above the normal wave base which was located at the platform margin.
4.3.4. BF-9 Orbitoides-Omphalocyclus (Plate ZZ, 3) The predominant components of the biota are
the larger benthic foraminifers Orbitoides media (d’Archiac) and Omphalocyclus macroporus (La- marck). The minor foraminifers include Sulcoper- culina, Rotaliidae and Miliolidae. Other biota in- clude echinoderms, brachiopods, molluscs and cal- careous algae (MarineZZa Zugeoni, Corallinaceae).
The bioclasts in grainstone, packstone, floatstone and rudstone were influenced by strong micritization. Sometimes, there was a high input of terrestrial materials during sedimentation. The sedimentation occurred in the high-energy shoals on the platform margin or a terrestrially influenced platform.
Plate I 1. Planktic foraminifers (Globotruncana) with some calcispheres. BF-2. Section D. Lower Zongshan Fm., Campanian. x 20. 2. Calcispheres (Pithonellu sphaerica, II ovalis), BF-3. Section At. Sample At38, Campanian. x6.5. 3. Calpionellids, BF-3. Section At, Sample At36, Campanian. x200. 4. Filaments with a few echinoderm fragments. BF-4. Section A, Fm. 2 of Gamba group. x5. 5. Keramsphaeru and Miliolidae. BF-21. Gulu Section, Paleocene. x 10. 6. Miscellanea, Sphaerogypsina and Verneuilina, BF-16. Section M, Sample Mr66, Paleocene. x20.

IO Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 11
4.3.5. BF-10 rudists (Plate III, 1) The biota is composed of unrounded and un-
sorted rudist fragments (Plate III, 1). The accessory fossils are benthic foraminifers (Orbitoides, Om- phalocyclus, Sulcoperculina and Miliolidae, etc.), echinoderms, brachiopods, Rhodophyceae (Archaeo- lithothamnium) and corals.
The boring structures showing geopetal fabrics can be found in the rudist fragments. Most of the bioclasts have the micrite envelopes. The diage- netic features are characterized by extensive mouldic porosity caused by selective solution.
The typical biofacies was formed in the high- energy shoals at the platform margin where rudist biostromes occasionally developed.
4.3.6. BF-11 Omphalocyclus-calcareous algae The benthic foraminifers consist mainly of Om-
phalocyclus and minor Orbitoides, Miliolidae and Rotaliidae. The calcareous algae include Coralli- naceae, Dasycladaceae and Udoteaceae (Halimeda and Ovulites). Other fossils include echinoderms, molluscs, rudists and sponges.
According to Elf-Aquitaine (1977, p. 24), Udotea- ceans tend to settle in shallow subtidal zones of the inner platform. The high diversity of biota and the presence of Udoteaceans show that this biofacies was produced in the open marine platform environment with good water circulation.
4.3.7. BF-12 Corallinaceans (Plate II, 4) Corallinaceans (Archaeolithothamnium, Litho-
thamnium and Lithophyllum) encrusted other bio- clasts such as rudists, corals and sponges, etc. to form rhodoids which show different growth forms: ramose, columnar, globular and laminated.
The accessory biota include other algae (Hali- meda, Ovulites, Solenoporaceae and Dasycladaceae), foraminifers (Omphalocyclus, Orbitoides, Milioli- dae and Rotalidae), echinoderms, ostracodes, in- oceramids, rudists, corals, sponges, stromatopoids,
serpulids, bryozoans and gastropods. The b&lasts with micrite envelopes occur in floatstone, rudstone, packstone and wackestone.
According to Bosence (1983, p. 226), the recent rhodoids of Florida occur in the sea grass bank near the coast and adjacent to ‘mud mounds’. The environment of rhodolite in Gamba is similar to that of recent rhodoids. The sedimentation of BF-12 took place on a nearshore marine platform which was influenced by tenigenous elastic input.
4.3.8. BF-13 Ovulites-ostracodes The biofacies is characterized by low biota di-
versity which consist of Ovulites, dasycladaceans, rudists, ostracodes and small-sized foraminifers (Verneuilina, Miliolidae, etc.). The limestones are intercalated in the sandstones of the Jidula Forma- tion and contain quite a few fine-grained to coarse- grained quartz and some pyrite. The contact be- tween these limestones and the underlying Lower Sandstone is marked by an erosional unconformity (Willems and Zhang Binggao, 1993a, fig. 6b).
According to Wray (1977, pp. 136, 155), Ovulites is an inhabitant of shallow subtidal (nearshore) la- goonal environments with temporary terrigenous in- flux, especially on the downwind side of protecting bars. This situation appears to hold true in the en- vironmental interpretation of BF- 13. The occurrence of limestones, the low biota diversity, the biotic as- pect and the presence of pyrite indicate a typical nearshore lagoonal environment influenced by ter- rigenous quartz input.
4.3.9. BF-14 Lockhartia-Dasycladaceae (Plate III, 2)
This biofacies is characterized by the abundance and high diversity of biota especially calcareous al- gae and benthic foraminifers. The former include Dasycladaceae (Neomeris, Trinocladus, Cymopo- ha, Fucoporella, Indopolia, etc.), Acetabulariaceae (Acicularia, Clypeina), Corallineae (Jania, Amphi-
Plate II 1. Goupillaudina and Sulcoperculina with some calcispheres and planktic foraminifers, BF-6. Section At, Sample At47, Campanian. x22. 2. Orbitoides, Sulcopenulina and Goupillaudina, BF-7, Section At, Sample At68, Campanian. x20. 3. Orbitoides and Omphalocyclus, BF-9. Section At, Sample At105, Campanian. x20. 4. The encrusting Archoeolithothamnium shows the globular growth type. BF-12. Section D, Sample Dr49, Maastrichtian. x20.
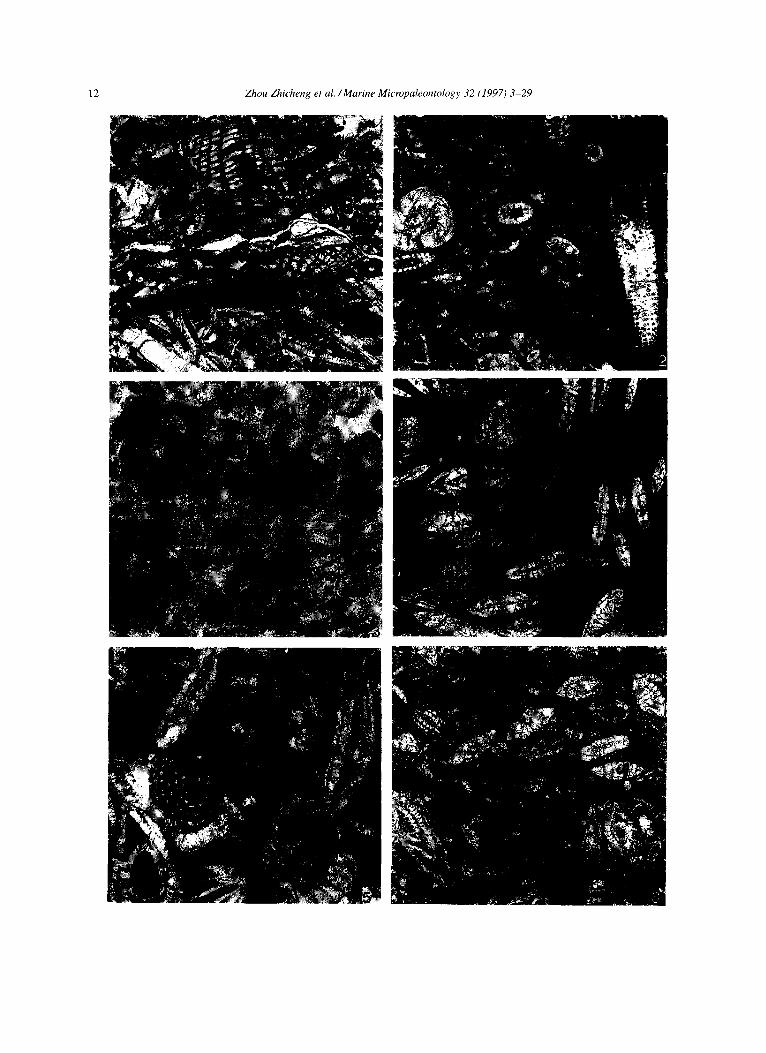
12 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 13
roa), Udoteaceae (Ovulites, Hulimedu) and the lat- ter include Lockhartia, Miliolidae, Vemeuilinidae, Discocyclina and Sphaerogypsina. Other fossils in- clude echinoderms, bivalves, gastropods, ostracodes, sponges and corals.
The high biota diversity shows that the primary environment had a good water circulation and the sedimentation took place in an open marine platform environment.
4.3.10. BF-15 Lockhartia-Verneuilinidue (Plate III,
3) The biota consist mainly of benthic foraminifers
including Lockhartia, Vemeuilinidae Miliolidae and Textularia, etc. The other biota are echinoderms, ostracodes and calcareous algae (Lithophyllum, Jania and Corallina subtilis). The limestones have been influenced by dolomitization.
The BF-15 is intercalated in BF-14 (Fig. 6). The biota diversity of BF-15 is much lower than that of BF-14. The marine platform was influenced by a small episodic transgression and regression in mid- dle Paleocene. The BF-14 was formed in the trans- gressive stage and the BF-1.5 was formed in the regressive stage. The depositional environment of BF-15 was shallower than that of BF-14.
4.3.11. BF-16 Miscellanea-Ranikothalia (Plate I, 6; Plate III, 5, 6)
This biofacies is characterized by abundant larger foraminifers including Miscellanea, Ranikothalia, Discocyclina, Operculina, Sphaerogypsina and Alve- olina. The small-si.sed foraminifers include Lockhar- tia, Miliolidae and Vemeuilinidae. The other biota are echinoderms, bivalves, gastropods, ostracodes and calcareous algae (Dasycladaceae indet., Furco- porella, Dissocladella, Distichoplax biserials, Litho- porella and Trinocladus).
The described assemblage of larger foraminifera is typical for open marine shelf areas (Hottinger,
1973, p. 445). After Reiss and Hottinger (1984, p. 282), during the middle to Late Paleocene, espe- cially, Ranikothalia associated with Operculina was a characteristic fauna for low-energy and soft bottom environments for water depths from 60 to 90 m. The existence of green algae indicates a good aeration and light penetration. As mentioned above, BF-16 is indicative of open marine shelf or carbonate platform environments.
4.3.12. BF-17 Alveolina-Orbitolites (Plate ZV 2) The biofacies consists mainly of larger fora-
minifers, especially Alveolina and Orbitolites. The other foraminifers include Opertorbitolites, Rotali- idae, Miliolidae and minor Vemeuilinidae and plank- tic foraminifers. The accessory biota are calcare- ous algae (Corallineae, Melobesieae indet., Dasy- cladaceae indet., Ovulites and Halimeda, and the fossils are found mainly in wackestone, floatstone and packstone.
According to Reiss and Hottinger (1984, p. 283) and Hottinger (1973, p. 446), the occurrence of alve- olinids and Orbitolites indicates a shallow and mod- erately agitated environment. Sedimentation took place in open marine platform or high-energy shoals.
4.3.13. BF-18 Halimeda-echinoderms (Plate IV 1) The biofacies mainly consists of Halimeda and
echinoderms. The foraminifers comprise the ben- thic small-sized foraminifers (Miliolidae, Vemeuilin- idae, Textularidae and Rotaliidae) and a few planktic foraminifers. Calcareous algae include Dasycladaceae (Pianella succincta), Melobesieae (Palaeolithothamnium) and Udoteceae (Hulimeda, Ovulites). Other fossils include bivalves, gastropods, ostracodes, bryozoans and brachiopods.
The bioclasts with micrite envelopes are found in wackestone, packstone, floatstone and rudstone. The limestones bearing BF-18 interfinger with ooid grainstone. The diagenetic features and the high
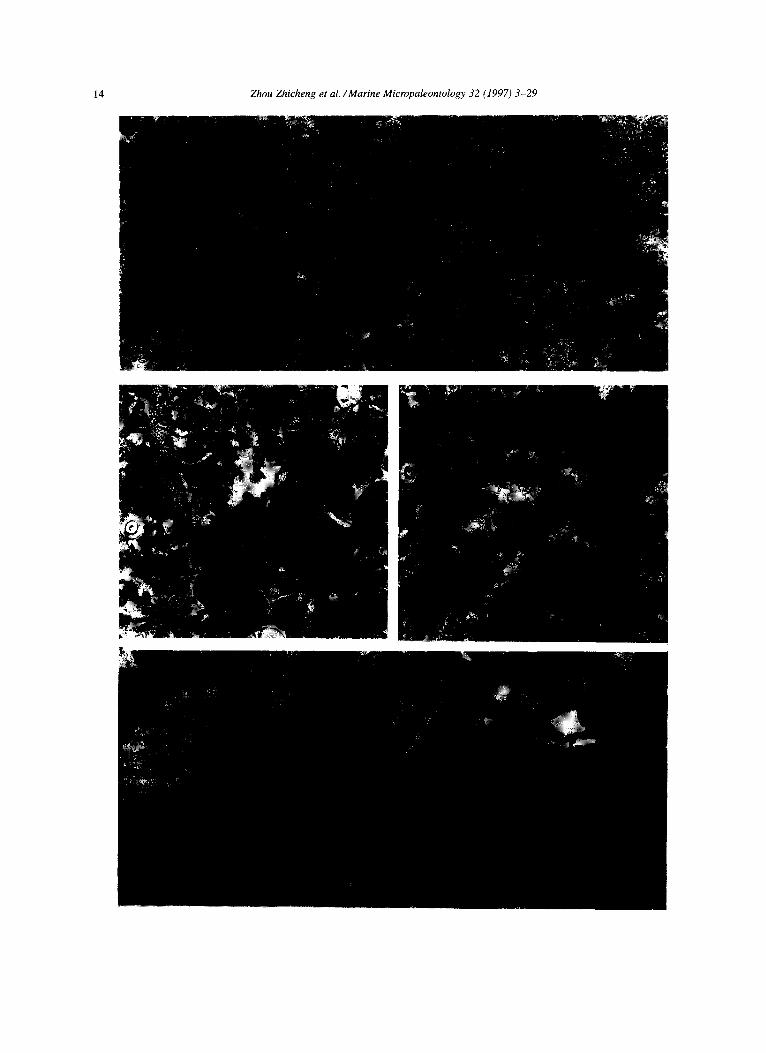
Plate III 1. Rudist fragments mainly consisting of radiolitids and hippuritids, BF-10. Section D, Sample Dr31, Maastrichtian. x.5 2. Lockhartia and Dasycladaceae, BF-14. Section Zm, Sample Zm14, Paleocene. x20. 3. Biota mainly consisting of Lockhartia, Verneuilina and miliolids, BF-15. Section Zp, Sample Zpl 1, Paleocene. x20. 4. Assilina. BF-22. Longjiang Section, Eocene. x 10. 5. Miscdanea and Alveolina. BF-16. Section F, Sample Ff33, Paleocene. x20. 6. Miscellanea and Discocyclina, BF-16, Section F, Sample Fr9, Paleocene. x 10.
14 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 15
biota diversity indicate that the sedimentation took place in the shallow and well circulated environment of the open marine platform adjacent to the ooid bars.
4.3.14. BF-19 Pseudolithothamnium-corals The biofacies is characterized by laminar rhodoids
which were formed by the encrusting Pseudoli- thothamnium album and minor Lithophyllum, Litho- thamnium and Archaeolithothamnium. Most rhodoid nuclei are composed of autochthonous or slightly re- worked corals (Pocilloporidae, Poritidae: Goniopora, Alveopora), sponges, Solenomeris and lithoclasts.
The rhodoids are embedded in bioclastic wacke/packstone. Other biota include Corallineae (Amphiroa, Jania), udoteacean (Halimeda), dasy- cladacean algae (Neomeris, Cymopolia, Clypeina), rotaliids, miliolids, verneuilinids, ostracodes, bi- valves, gastropods and echinoderms.
Solution in the freshwater vadose zone led to the intensive vuggy and mouldic porosity. The voids were filled with the granular calcite and vadose silts in a geopetal fabric. The diagenetic features indicate that the sedimentation took place in shallow and restricted environments. Compared with the model of tropic rhodolite environments (Bosence, 1983, pp. 226, 232), the most appropriate sedimentary environ- ment would be shallow subtidal rhodolite bars (patch reefs: FZ6B, Fig. 4) forming a barrier between the open marine platform (FZ6A) and restricted lagoon (FZ7B).
4.3.15. BF-20 Miliolidae-Dasycladaceae (Plate IV 3, 4)
The small-sized benthic foraminifers are com- posed of miliolids, vemeuilinids and Textularia. Dasycladacean algae occupy the main compo- nents of calcareous algae. They are Cymopo- ha, Dissocladella, Furcoporella, Neomeris, Trin- ocladus and Zndopolia (Plate IV, 4). Other algae include Udoteaceae (Halimeda, Ovulites), Melobe-
sieae (Archaeolithothamnium) and Squamariacaea (Pseudolithothamnium album). Other minor biota consist of echinoderms, bivalves, gastropods, ser- pulids and corals. The b&lasts are embedded in wackestone, packstone and floatstone.
It is suggested that the sedimentary environment was a restricted lagoonal area (FZ7B, Fig. 4) pro- tected from the open marine platform by rhodolite bars.
4.3.16. BF-21 Lockhartia-Keramosphaera (Plate I, 5)
The association is dominated by foraminifers consisting of Lockhartia, Keramosphaera, Sphaer- ogypsina and Miliolidae, etc. Calcareous algae in- clude Dasycladaceae (Trinocladus, Cymopolia, Dis- socladella), Udoteaceae (Halimeda, Ovulites) and Acetabulariaceae (Clypeina). Other biota are echino- derms, bivalves, gastropods and ostracodes.
According to (Reiss and Hottinger, 1984: 283), Daviesina and Lockhartia are typical of shallow sub- tidal environments with higher water agitation during the middle Paleocene and Early Eocene. The occur- rence of Lockhartia and the comparatively high biota diversity suggest that the sedimentary environment of BF-21 is an open marine platform (FZ6A, Fig. 4) with a good water circulation.
4.3.17. BF-22 Nummulites-Alveolina (Plate III, 4; Plate V 1, 3)
The biofacies is dominated by the abundant larger foraminifers (Nummulites, Alveolina, Orbitolites, As- silina and Discocyclina). The small-sized foramini- fers occur in the space among the larger foraminifers. They are Miliolidae, Vemeuilinidae, Textularridae, Lockhartia and a few planktic foraminifers. Other minor biota include echinoderms, bivalves, ostra- codes, bryozoans and calcareous algae (Lithoporella, Distichoplax biserials).
The dominant nummulitids and alveolinids lived during the middle Eocene as habitants of soft bot-
Plate IV 1. Halimeda, BE1 8, Section M, Sample Mrl 1, ?Danian. x 20. 2. Alveolina and Orbitolites, BF-17. Section F. Sample F16, Thanetian. x 10. 3. Miliolidae and Dasycladaceae. BF-20. Section M, Sample Mr42, ?Danian. x20. 4. Indopolia. BF-20. Section M. Sample Mr32, ?Danian. x20.

16 Zhou Zhicheng et ~1. /Marine Micropuleontology 32 (1997) 3-29

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 17
toms and/or areas with low water-energy in depth of 50-90 m (Reiss and Hottinger, 1984, p. 283). The depositional environment of BF-22 can be inter- preted as an open marine platform (FZ6A, Fig. 4) or a deeper part of a homocline ramp.
4.3.18. BF-23 bivalves-gastropods The biofacies is characterized by autochthonous
bivalves (oysters, Cardium, Mytilus, Nucula) and gastropods. There are some redeposited foramini- fers (Alveolina, Nummulites, Orbitolites, miliolids), echinoderms and bryozoans, etc. The fossils coex- ist with the radiaLfibrous ooids which are typical indicators of hypersaline coastal pond environments (Fliigel, 1982, p. 147). Another outstanding diage- netic feature is the high number of authigenic bipyra- midal quartz crystals often occurring in the centre of the ooids.
As above, the biofacies documents the last ma- rine ingressions into a restricted, slightly hypersaline cutoff lagoon or coastal pond (FZ8, Fig. 4).
4.4. Redeposited biofacies
4.4.1. BF-24 redeposited biofacies (Plate v 2) The redeposition, caused mostly by turbidite or
debris flow, led to the mixture of different biota de- rived from variant environments and variant strata. Sometimes, two different kinds of pebbles accumu- lated together. One came from the Zongshan Forma- tion of Maastrichtian age and formed in the shallow water environments. The others derived from pelagic deposits of Santonian-Campanian age. This typical biofacies in genesis is termed redeposited biofacies.
The biofacies is characterized by having poor sorting, roundness and chaotic texture and includes hemipelagic or pelagic biota and benthic biota.
According to the genetic features of this bio- facies the sedimentation took place mostly in the distal continental slope (FZ2, Fig. 4) or the proximal continental slope (FZ4, Fig. 4).
5. Ichnofacies
Trace fossils are regarded as a useful tool for interpreting the environmental factors such as bathymetry, sedimentation rate, oxygenation, sub- strates, energy level and salinity. The evolution of sedimentary environments and paleogeographic as- pects in southern Tibet during the period from Creta- ceous to Paleogene led not only to changes of biofa- ties, but also to changes of ichnofacies. Variant trace fossil assemblages have been found in the various kinds of deposits from different strata. Every trace- fossil assemblage is related to a certain sedimentary environment. According to Seilacher (1967), Frey and Seilacher (1980) and Ekdale et al. (1984), the trace fossil assemblages can be attributed to Nereites, Zoophycos, Cruziana and Skolithos ichnofacies.
5.1. Nereites ichnofacies
The trace fossils Phycosiphon sp. (Plate VI, 4) and Palaeophycos tubularis? have been found in the silt- stones in the lower part of Gamba Group of Neoco- mian age. A similar trace fossil assemblage occurs in the muddy siltstones and marls of Formation 2 of the Gamba Group aged middle-late Albian. The trace fossil assemblage includes Megagrapton? sp., Phy- cosiphon sp., Taenidium serpentinum (Plate VI, l), Planolites montanus, P. beverleyensis, Chondrites sp. and Palaeophycos tubularis? Most of the trace fossils of the assemblage are fodinichnia. Mega- grapton has been found in the flysch deposits of the Polish Carpathians (Ksiazkiewicz, 1970). Taeni- dium, Phycosiphon, Chondrites are main members of Nereites ichnofacies in the Ouachita Geosyncline (Chamberlain, 1978). The above mentioned trace fossil assemblages found in the Gamba Group can be assigned to the Nereites ichnofacies.
Nereites ichnofacies is usually considered to be the trace fossil assemblage produced on the soft sub- strate in the deep sea environment. The deposits in
Plate V 1. Nummulites and Alveolina. BF-22. Section M, Sample Mr84, Eocene. x 10. 2. Redeposited calcispheres, Textuluria and Omphalocyclus fragments, BF-24, Section L, Sample Lf 122, Maastrichtian-Paleocene. x 20. 3. Nummulites, Assilina and Discocyclina, BF-22. Entong Section, Sample 79DN16, Eocene. x 10. 4. Zoophycos sp., Zoophycos ichnofacies, Zoophycos-Chondrites suhichnofacies. Section At, Sample AT22, Campanian. 5. Chondrites sp. in thin section. Section A, Sample Ar7, Albian. x.5.

18 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 19
which the Nereites ichnofacies occurs contain am- monites, belemnites and planktic foraminifers. Com- pared with the facies model in this paper (Figs. 2 and 3), the appropriate sedimentary environment that the Nereites ichnofacies reflects is an open marine basin below lower wave base.
5.2. Zoophycos ichnofacies
The trace fossils Zoophycos (Plate V, 4), Chon- drites, Thalassinoides sp. and Planolites occur in Limestone 1 and Calcareous Marl 1 of the Zong- shan Formation in Gamba and Tiina. The trace fossil assemblage can be referred to the Zoophycos ichno- facies. An interesting phenomenon has been found in Section At in Tiina. The trace fossil assemblage in the limestones with lower terrigenous influx is strikingly different from that in the marlstones with higher terrigenous input. The trace fossil assemblage in limestones consists mainly of trace fossils of the upper tier which include Thalassinoides, Planolites and Phycodes? (Pla.te VI, 6). This trace fossil assem- blage is characterized by its high diversity and high abundance. By contrast to this, the trace fossil assem- blage in the marlstones contains mainly trace fossils of the deeper tier. They are Zoophycos and Chon- drites. The trace fossil assemblage is characterized by lower diversity and lower abundance. The two trace fossil assemblages alternate in the vertical sec- tion (Fig. 4). Brom.ley and Ekdale (1984) described the oxygen-controlled tiering relation of trace fos- sils in selected examples of Mesozoic marine strata that represent oxic to anoxic depositional environ- ments. When the oxygen content of bottom water decreases, the trace fossils Planolites and Thalassi- noides disappear from sections first and Chondrites and Zoophycos disalppear last. Therefore, Chondrites and Zoophycos are good indicators of dysaerobic or anaerobic conditions within the sedimentary environ-
ments. The Thalassinoides-Planolites assemblage is produced in aerobic environment. The alternation of the two trace fossil assemblages in section At was caused by the changes of oxygen content in bottom water. The Zoophycos ichnofacies in Tiina can be subdivided into Thalassinoides-Planolites subich- nofacies and Zoophycos-Chondrites subichnofacies which were representative of the aerobic and dysaer- obic conditions of the original depositional environ- ments, respectively.
According to Frey and Pemberton (1985, p. 99), the Zoophycos ichnofacies is typically portrayed as an intermediary between the Cruziana and Nere- ites ichnofacies, at a position corresponding more or less to the continental slope. The deposits bearing Zoophycos ichnofacies are rich in hemipelagic and pelagic organisms like planktic foraminifers, calci- spheres and calpionellids. The graded bedding pro- duced by distal turbidites and the bioturbated stmc- tures are present in the deposits. The sedimentary environments represented by Zoophycos ichnofacies in Tiina and Gamba correspond to shelf margin and open sea shelf environments reflected by BF-2 and BF-3.
5.3. Cruziana ichnofacies
Abundant trace fossils of this group have been found mainly in the siltstones, fine-grained sand- stones of Formation 2 of the Gamba Group. They include Gyrochorte comosa, Syringomorpha hori- zontalis, Fucusopsis sp., Teichichnus sp. (Plate VI, 5), Chondrites sp. (Plate V, 5; Plate VI, 3) and be- long to fugichnia. The trace fossil assemblage is characterized by high diversity and is referred to the Cruziana ichnofacies (Fig. 5).
Formation 2 of the Gamba Group contains abun- dant inorganic structures including cross-bedding, storm layers, slumping structures, channel deposits,
Plate VI 1. Taenidium serpentinum, Nereires ichnofacies. Section A, Sample Ar63, late Albian, polished section. x 1.5. 2. Arenicolites sp., Skolithos ichnofacies. Section D, Jidula Formation, Maastrichtian-Early Paleocene, x0.43. 3. Chondn’res sp., Tiina, late Albian? x0.67. 4. Phycosiphon sp., Nereites ichnofacies. Section K, Sample Krl, Neocomian. x 1.3. 5. Teichichnus sp., Cruziana ichnofacies. Section A, Sample Ar13, middle-late Albian, polished section. x 1. 6. Thalassinoides, Planolites and Phycodes? assemblage, Zoophycos facies, Thalassinoides-Planolites subichnofacies. Section At, Sample At1 6, Campanian. x 1.
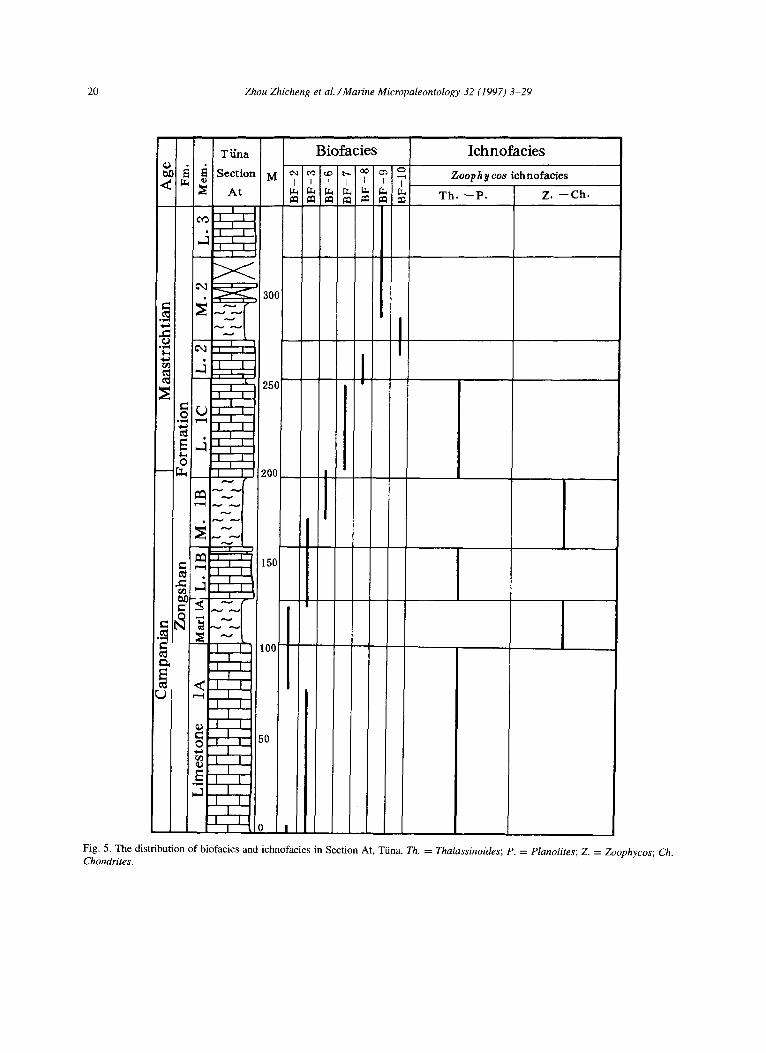
20 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29
Fig. 5. The distribution of biofacies and ichnofacies in Section At, Tiina. Th. = Z’halassinoides; P. = Planolites; Z. = Zoophycos; Ch. Chondrites.

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 21
sandy neptunic dikes, graded bedding and Bouma sequences. The sedimentary structures show that the deposits have been influenced by storms and tur- bidites. Commonly, Cruziana ichnofacies is found on open continental shelves. The Cruziana ichnofa- ties of the Gamba Group is produced on an open sea shelf, shelf margin where the activities of storm waves and turbidites reached.
5.4. Skolithos ichnc$acies
When geologists examine to the sandstones of the Jidula Formation in Gamba and Tuna, they will be impressed by the abundant, densely packed trace fossils in the sandstones. The vertical trace fossils include Arenicolites sp. (Plate VI, 2), Skolithos Zin- earis and Skolithos !jp., and are referred to the typical Skolithos ichnofacies.
As Frey and Pemberton (1984) and Read- ing (1986) indicated, the environments which the Skolithos ichnofacies represents are characterized by shifting substrates, moderate- to high-energy condi- tions and episodic (erosion or deposition. The envi- ronments include the foreshore and shoreface zones of beaches, bars and spits (Howard, 1972, 1975). The fine-medium g,rained sandstones of the Jidula Formation have medium to large cross-beddings at the lower part (Mernber 1) and medium to small di- mensional cross-beddings at the upper part (Member 3). The Skolithos ichnofacies and the physical sedi- mentary structures indicate that the sandstones were deposited in the high-energy environments of sand- barriers in subtidal shoreface zones and intertidal foreshore zones.
6. Sedimentary history
The preceding chapters describe the important Cretaceous-Paleogene biofacies and ichnofacies in Tuna, Gamba and Tingri and briefly indicate their environmental significance. The evolution of biofa- ties and ichnofacies in the vertical sections is impor- tant in understanding and reconstructing the marine sedimentary history from Cretaceous to Paleogene in southern Tibet (Figs. 5-9). The main facies zones are summarized in Fig. 2 (according to Willems, 1993). In this chapter, the outline of the sedimentary history is generalized.
6.1. Tiina, Gamba areas
6.1. I. Neocomian-Albian (basin environments) During the Neocomian-Albian (formations 1,2 of
the Gamba Group), basin environments were domi- nant. Based on the features of biofacies, ichnofacies and petrology, two different basin types can be dis- tinguished. One is the euxinic basin and the other is an open marine basin. The sedimentation may be related to the global oceanic anoxic event from Barremian to Albian (Schlager and Jenkyns, 1976; Arthur and Schlager, 1979).
The depositional conditions in the open marine basin differed from those of the euxinic basin by having a good water circulation and higher oxy- gen content. The deposits comprise siltstone, fine- grained sandstone and marlstones in which the nektonic and planktic fossils (ammonites, planktic foraminifers), benthic fossils (gastropods, bivalves) and biogenic structures (Nereites ichnofacies) can be found. The sedimentation was also influenced by distal turbidites. A short regression took place in the Albian. The Cruziana ichnofacies of Formation 2 of the Gamba Group indicates a shelf margin and open sea shelf environment.
6.1.2. Cenomanian-Campanian (shelfmargin, open sea shelf and platform slope environments)
The regression began in the Cenomanian and con- tinued to the Campanian leading to the transition of the environments from the basin to the prevailing shelf margin and open sea shelf. Carbonate deposits play an important role in the sedimentation of these environments. The deposits comprise mostly marl- stones and limestones (Formation 3 of the Gamba Group and the lower part of the Zongshan Forma- tion). The biofacies are mainly composed of BF-2 and BF-3. The Zoophycos ichnofacies at the lower part of the Zongshan Formation can be divided into the Thalassinoides-Planolites subichnofacies and the Zoophycos-Chondrites subichnofacies. The al- ternation of the two subichnofacies in the vertical section (Fig. 5) was caused by the episodic fluc- tuating of aerobic and disaerobic conditions at the sea floor. During the very late Campanian, the sed- imentation took place mainly in the platform slope located in the transitional facies zone between the open sea shelf and carbonate platform because of

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29
VJ T
-Y
I . . . . I 0 Fig. 6. The distribution of biofacies and ichnofacies in sections A, K, Gamba.
Ichnofacies
CTUZiaSU N eTe&?es
ichnofacies ichnofacies
I
the further regression. The biota in this environment represented by BF-6 are characterized by the co- existence of benthic organisms and the pelagic or hemipelagic organisms.
6.1.3. Maastrichtian (the development of a carbonate platfomt)
During the Maastrichtian, a carbonate platform was established and developed in Tiina and Gamba. The biota on the carbonate platform are character- ized by the predominant benthic organisms with high

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 23
Fig. 7. The distribution of biofacies and ichnofacies in sections D, Zp, Zm, F, Gamba.
diversity. In the early and middle Maastrichtian, the smaller foraminifers Miliolidae and the larger for- aminifers Orbitoides, Omphalocyclus are important rock-forming members, and in the late Maastrichtian the calcareous a1ga.e play a more important role in the composition of biota. The bioclasts usually with micrite envelopes show the depositional features in the shallow water environments. In general, the car-
bonate platform went ahead landwards from the early Maastrichtian to the late Maastrichtian. Two stages, platform margin and open marine platform, can be distinguished. The platform margin is represented by high-energy shoals (BF-7, BF-8, BF-9) and rudist biostromes (BF-10). In the late Maastrichtian, sed- imentary environments were deposited in the open marine platform including the carbonate platform

24 Zhou Zhicheng et al. /Marine Micropaleontology 32 (I 997) 3-29
~1 Tingri .x d Section I
3
-
M
-
LOO
8OC
500
400
100
0 -
3iofacies
Fig. 8. The distribution of biofacies in Section L, Tingri
and tenigenously influenced platform conditions. The biofacies (BF-11) formed in the carbonate plat- form consists mainly of Omphalocyclus and differ- ent kinds of calcareous algae (Corallinaceae, Dasy- cladaceae and Udoteaceae). Afterwards, the Coral- linaceae (BF-12) forming the rhodoids prevailed in the terrigenously influenced platform.
61.4. MaastrichtianlPaleocene boundary (sandstone accretions and lagoonal environment)
From the end of the Maastrichtian to the begin- ning of the Paleocene, the carbonate dominant sedi- mentation terminated because of the large quantities of terrigenous quartz sand input. The sand barriers
were deposited in the intertidal foreshore zone and the subtidal shoreface zone. The Skolithos ichnofa- ties formed under the shifting substrates; moderate- to high-energy and episodic erosion or deposition conditions flourished in the sandstones of the Jidula Formation. The sandstones are separated by a black pyrite-bearing limestone which contains a lower di- versity biota including Ovulites, ostracodes, dasy- cladaceans, rudists and smaller foraminifers. The limestones indicate a nearshore lagoonal environ- ment.
6.1 S. Middle Paleocene (open marine platform) The next transgression commenced at the begin-
ning of the middle Paleocene and resulted in the accumulation of carbonate deposits formed in the open marine platform. The marlstones and lime- stones of Member 1 of the Zongpu Formation (Fig. 6) are superimposed on the sandstones of the Jidula Formation. In the well circulated en- vironment, calcareous algae flourished again; they comprise Dasycladaceae, Acetabulariaceae, Coralli- naceae and Udoteaceae. The foraminifers Lockhartia and Verneuilina occupy an important position in the composition of the benthic foraminifers. The BF-14 and BF-1.5 are the representative biofacies during this period.
6.1.6. Thanetian (open sea shelf and open marine platform environments)
The transgression may reach a climax in the Thanetian when the sedimentary processes shifted from the open marine platform to the open sea shelf. The sedimentary processes are indicated by BF- 16 in which the larger foraminifers Miscellanea, Ranikothalia and Operculina occur. The transgres- sion only lasted a short time. From the transition during the Thanetian, the sedimentary processes returned from the open sea shelf to open marine platform again. The sedimentary environment is re- flected by the BF- 17 in which the larger foraminifers Alveolina and Orbitolites are dominant.
6.1.7. Latest Thanetian (jinal stages of marine sedimentation: cutoff lagoons and coastal ponds)
During the latest Thanetian, the continued re- gression of the sea resulted in the end of nor- mal marine sedimentation. Sedimentary processes

Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29 25
M
-
40 5.
Of
j0
1
Biofacies
LEGEND
E3 limestone
Ia nodular limestone
- - hJ_rl - marly limestone --
El dolomite
I, I N
1 marl
I I - L calcareous marl A_
El claystone
. . . . r-l siltstone . . . . . . .
. . . l-l .__ . . . fine - grained sandstone
. . I I -. ._ medium - gr&ed s&stone
coarse-grained sandstone
..___ _
cl .-..__ sandstone .._.. .
i-l Q septarium
PI thrusting
zig. 9. The distribution of biofacies in Section M, Tingri

26 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29
shifted from marginal marine depositional environ- upper half, whereas the sandstones of the Jidula For- ments, tidal flats and coastal ponds into cutoff hy- mation (Early Paleocene) developed from seaward persaline lagoons. As a result, the limestones of the prograding delta plains (Willems, 1993, p. 127). The Zongpu Formation were replaced by the marlstones sediments in the various evolutionary stages of the and clay/siltstones of the Zongpubei Formation and paleo-continental slope are different. They originated the biofacies rich in stenohaline organisms, espe- mainly from turbidite and debris flow sedimentation cially the larger foraminifers were replaced by the as well as normal sedimentation. The typical biofa- BF-23 containing mainly autochthonous euryhaline ties that resulted from the resedimentation can be bivalves and gastropods. attributed to the redeposited biofacies (BF-24).
As described above, two main cycles of transgres- sion and regression of the sea took place in Tuna and Gamba from the Cretaceous to Paleogene. The first cycle lasted from the Neocomian to the Maas- trichtian/Paleocene boundary. The second one lasted from the middle Paleocene to the end of the Thane- tian. The scope of the second transgression was much smaller than that of the first one. The marine sedimentary history ended at the Paleocene/Eocene boundary.
6.2.3. ?Danian (carbonate platform)
6.2. Tingri area
6.2.1. Late Albian-early Maastrichtian (open marine basin, basin margin and open sea shelf environments)
The sedimentary processes from the late Albian- Campanian occurred mainly in open marine basin, shelf margin and open sea shelf environments. The biota are characterized by pelagic and hemipelagic organisms (planktic foraminifers, filaments, calci- spheres, radiolarians and spicules). The biofacies are represented by BF-2, BF-3 and BF-4 (Fig. 8). The euxinic deposits found in Gamba from Neocomian to Albian have not been found in Tingri area during the same period. The basin environments in Tingri area had the better water circulation than that in the Gamba area.
The termination of the strong terrigenous material input and the beginning of the transgression led to the establishment of a stable carbonate platform in Tingri during ?Danian times. The deposits of the carbonate platform began with the ooid grainstone deposited in the high-energy shoal at the platform margin and the wackestone, packstone, floatstone and rudstone containing Halimeda, echinoderms and smaller foraminifers, Dasycladaceae, Melobesieae, formed in the open marine platform. Afterwards, rhodolite bars and patch reefs were produced on the open marine platform. The lagoons cutoff from the open sea by the rhodolite bars and patch reefs were formed simultaneously. The BF-20 is indicative of this kind of low-energy environment. With the increase in the transgression, the sedimentary pro- cesses shifted from the rodolite bars, patch reefs and lagoonal environments to the open marine platform. The changes of sedimentary environments resulted in the growth of the calcareous algae and foramin- ifers especially of Dasycladaceae and Lockhartia (BF-14).
6.2.4. Thanetian-Lutetian (diversification of larger foraminifers)
6.2.2. Middle Maastrichtian-Early Paleocene (continental slope)
While a carbonate platform was establishing in the Gamba area during the Maastrichtian, a continen- tal slope was developing in Tingri during the time from the middle Maastrichtian to the Early Pale- ocene. According to this model, the sediments of the Zhepure Shanpo Formation (from the middle Maas- trichtian to the Early Paleocene) represent deposits of a lower fan in the lower half, more of a mid-fan in the
A stable, carbonate platform only slightly differ- entiated morphologically (Read, 1982) is established during the period from the Thanetian to Lutetian. The sedimentary processes took place on an open marine platform for most of the time. A short trans- gression occurred in the middle and late Thanetian which resulted in the shifting of the sedimentary environments from the open marine platform to the open sea shelf. Consequently, the biota consisting mainly of the larger foraminifers Lockhartia and Keramosphaera (BF-21) were replaced by biota con- sisting mainly of larger foraminifers Miscellanea

Zhou Zhicheng et al./Marine Micropaleontology 32 (1997) 3-29 27
and Ranikothalia (BF-16). The sedimentary environ- ments returned to the open marine platform from Thanetian to Luteti.an. The biota are characterized by the very large Nummulites, Alveolina, Orbitolites, Assilina and Discocyclina (BF-22).
Because the non-marine Zongpubei Formation has not been found in Tingri, the marine sedimentary history there ended in the Lutetian according to the youngest deposits of Lutetian limestones.
7. Conclusions
The understanding of the sedimentary environ- ments and history of the Cretaceous and Paleogene in the Tuna, Gamba, and Tingri areas of southern Tibet is based on the detailed studies of biofacies and ichnofacies.
(1) Two major transgression/regression cycles can be distinguished from the Early Cretaceous to the Paleogene in the Gamba area. The first lasted from the Neocomian to the Cretaceous/Paleogene bound- ary and the second from the middle Paleocene to the Thanetian.
(2) The sedimentation processes in the Gamba area from Neocomian to late Albian took place briefly in basin environments from which two types, an euxinic basin and an open marine basin, can be recognized by the biofacies and ichnofacies. The former is reflected by the euxinic biofacies and litho- facies, while the latter contains ammonites biofacies and Nereites ichnolgcies. The Cruziana ichnofacies can be found from both the shelf margin and the open sea shelf.
(3) Calcispheres (calcareous dinoflagellates) flourished in the outer shelf environments from the Santonian to the Campanian. The Zoophycos ich- nofacies occurred in the zones from the open sea shelf to the platform margin from the Campanian to the lower Maastrichtian; this can be further divided into two subichnofacies: Thalassinoides-Planolites subichnofacies and Zoophycos-Chondrites subich- nofacies. Their alternation in the vertical section was caused by the fluctuation of the oxygen con- tent on the sea floor. The Th.-P. subichnofacies was produced under aerobic conditions, and the Z.-Ch. subichnofacies under disaerobic conditions. The transitional zone between the open sea shelf and the carbonate platform is also indicated by
the Goupillaudina-Sulcoperculina biofacies which is characterized by the coexistence of the benthic biota and the pelagic biota.
(4) Not until the Maastrichtian was a stable carbonate platform established in the Gamba area. The carbonate platform went through several stages: higher-energetic shoals and rudist biostromes, open marine platform and the terrigenously influenced platforms. The benthic foraminifers (Orbitoides, Omphalocyclus and Miliolidae), calcareous algae (Corallinaceae, Dasycladaceae and Udoteaceae), and rudists (hippuritids and radiolitids) are the most im- portant biota in these environments. The Coralli- naceans (Archaeolithothamnium, Lithothamnium and Lithophyllum) encrusted other bioclasts to form the rhodoids, which occurred in the nearshore marine platform.
(5) The occurrence of the thick-bedded Jidula sandstones from the late Maastrichtian to the Early Paleocene marked the end of the first regressive cycle in the Gamba area. The typical Skolithos ichnofacies, widely found in the sandstones, indi- cates nearshore environments in which sand barriers were built. Black limestones bearing an Ovulites- ostracodes biofacies are intercalated in the Jidula sandstones. They were produced in lagoonal envi- ronments with stagnating water circulation.
(6) In Tingri, the sedimentation processes during the time from the Vraconian to the Campanian were similar to those of Gamba. After the diastema of early Maastrichtian, heavy redeposition took place on a paleo-continental slope in the Tingri area.
(7) At the beginning of the middle Paleocene, a major transgression took place in the areas of Gamba and Tingri. In the Tingri area, a stable carbonate platform was formed. First, Halimedu-echinoderms biofacies was produced in the open marine platform and this was followed by the biofacies of Pseu- dolithothamnium album and corals produced in the rhodolite bars and patch reefs in the nearshore en- vironments. The larger foraminifers, Miscellanea, Ranikothalia, Opeculina, Discocyclina, and rotali- ids expanded in both Gamba and Tingri during the Thanetian.
(8) The termination of the marine sedimentary history ended in the Thanetian in the Gamba area, which was earlier than in the Tingri area. There, larger foraminifers Nummulites, Assilina and Aster-

28 Zhou Zhicheng et al. /Marine Micropaleontology 32 (1997) 3-29
ocyclina spread widely into the Lutetian which was the last stage of the marine sedimentation in Tingri.
Acknowledgements
We thank the German Science Foundation (Wi 725/5), the Max-Planck-Gesellschaft, and the Academia Sinica for their financial support of the Ti- bet Expeditions. We thank Prof. D.G. Jenkins, Prof. W. Berggren and Prof. Wang Pinxian for their com- ments and suggestions leading to the improvement of this paper.
References
Arthur, M.A., Schlager, SO., 1979. Cretaceous ‘Oceanic Anoxic Events’ as causal factors in development of reef-reservoired giant oil fields. AAPG Bull. 63 (6), 870-885.
Bosence, D.W.J., 1983. The occurrence and ecology of Re- cent rhodoliths: a review. In: Peryt, T. (Ed.), Coated Granis. Springer, Berlin, pp. 225-242.
Bromley, R.G., Ekdale, A.A., 1984. Chondrites: a trace fossil indicator of anoxia in sediments. Science 224, 872-874.
Chamberlain, C.K. (Ed.), 1978. A Guidebook to the Trace Fossils and Paleoecology of the Ouachita Geosyncline. SEPM, Tulsa, OK, 68 pp.
Ekdale, A.A., Bromley, R.G., Pemberton, S.G., 1984. Ichnology. SEPM, Tulsa, OK, 317 pp.
Elf-Aquitaine, 1977. Essai de caracterisation sedimentologique des dep6ts carbonates (An attempt at sedimentological charac- terisation of carbonate deposits). 2: Elements d’interpretation, 231 pp.
Flugel, E., 1982. Microfacies Analysis of Limestones. Springer, Berlin, 633 pp.
Frey, R.W., Pemberton, S.G., 1984. Trace fossil facies models, In: Walker, R.G. (Ed.), Facies Models (2nd ed.). Geosci. Can. Reprint Ser. 1, 189-207.
Frey, R.W., Pemberton, S.G., 1985. Biogenic structures in out- crops and cores. 1. Approaches to ichnology. Bull. Can. Pet. Geol. 33 (l), 72-l 15.
Frey, R.W., Seilacher, A., 1980. Uniformity in marine inverte- brate ichnology. Lethaia 13, 183-207.
He Yan, Zhang Binggao, Hu Lanying and Sheng Jinzhang, 1976. Mesozoic and Cenozoic Foraminifera from the Mount Jolmo Lungma Region. In: A Report of Scientific Expedition in the Mount Jolmo Lungma Region (1966- 1968), Palaeontology. Fast. II. Science Press, Beijing, pp. 1-124 (in Chinese).
Hottinger, L., 1973. Selected Paleogene larger foraminifera. In: Hallam, A. (Ed.), Atlas of Palaeobiogeography. Elsevier, Am- sterdam, pp. 443-452.
Howard, J.D., 1972. Trace fossils as criteria for recognizing an- cient shorelines. In: Rigby, J.K., Hamblin, W.K. (Eds.), Recog- nition of Ancient Sedimentary Environment. SEPM Spec. Publ. 16, 215-225.
Howard, J.D., 1975. The sedimentological significance of trace fossils. In: Frey, R.W. (Ed.), The Study of Trace Fossils. Springer, New York, NY, pp. 131-146.
Ksiazkiewicz, M., 1970. Observations on the ichnofauna of the Polish Carpathians. In: Crimes, T.P., Harper, J.C. (Eds.), Trace Fossils. Geol. J. Spec. Issue 3, 283-322.
Liu, G., Einsele, G., 1994. Sedimentary history of the Tethyan basin in the Tibetan Himalayas. Geol. Rundsch. 83 (1). 32-61.
Read, J.F., 1982. Carbonate platforms of passive (extension- al) continental margins: types, characteristics and evolution. Tectonophysics 81, 195-212.
Reading, H.G. (Ed.), 1986. Sedimentary Environments and Fa- ties. Blackwell, Oxford, 615 pp.
Reiss, Z., Hottinger, L., 1984. The Gulf of Aqaba. Ecological Micropaleontology. Springer, Berlin, 283 pp.
Schafer, W., 1963. Biozonose und Biofacies in marinen Bere- ich, Aufsatze u. Reden Senckenbergische Naturforsch. Ges., Kramer, Frankfurt a. Main, 37 pp.
Schlager, S.O., Jenkyns, H.C., 1976. Cretaceous oceanic anoxic events - causes and consequences. Geol. Mijnbouw 55, 179- 184.
Seilacher, A., 1967. Bathymetry of trace fossils. Mar. Geol. 5, 413-428.
Wen Shixuan, 1974. Stratigraphy of the Mount Jolmo Lungma Region. In: A Report of the Scientific Expedition in the Mount Jolmo Lungma Region (1966- 1968). Geology. Science Press, Beijing, pp. 1-231 (in Chinese).
Wan, Xiaoqiao, 1987. Foraminifera biostratigraphy and paleo- geography of the Tertiary in Tibet. Geoscience (China) 1 (I), 1-47 (in Chinese, with English abstract)..
Wang Yujing, 1976. Late Cretaceous and Paleogene Calcareous Algae from the Mount Jolmo Lungma Region. In: A Report of Scientific Expedition in the Mount Jolmo Lungma Region (1966-1968) Palaeontology. Fast. II. Science Press, Beijing, pp. l-24 (in Chinese).
Willems, H., 1993. Sedimentary History of the Tethys Himalaya Continental Margin in South Tibet (Gamba, Tingri) during Up- per Cretaceous and Paleogene (Xizang Autonomous Region, PR China). In: Willems, H. (Ed.), Geoscientific Investigations in the Tethyan Himalayas. Ber. Fachbereich Geowiss. Univ. Bremen 38,49-181.
Willems, H., Zhang Binggao, 1993a. Cretaceous and Paleo- gene Sediments of the Tibetan Tethys Himalaya in the Area of Gamba (South Tibet, PR China). In: Willems, H. (Ed.), Geoscientific Investigations in the Tethyas Himalayas. Ber. Fachbereich Geowiss. Univ. Bremen 38, 3-28.
Willems, H., Zhang Binggao, 1993b. Cretaceous and Paleogene Sediments of the Tibetan, Tethys Himalaya in the Area of Tin- gri (South Tibet, PR China). In: Willems, H. (Ed.), Geoscien- tific investigations in the Tethyan Himalayas. Ber. Fachbereich Geowiss. Univ. Bremen 38, 29-48.
Wilson, J.L., 1975. Carbonate Facies in Geologic History. Springer, Berlin, 471 pp.
Wray, J.L., 1977. Calcareous algae. Dev. Paleontol. Strat. 4, 185
PP. Xu Yulin, Wan Xiaoqiao, Gou Zonghai and Zhang Qihua, 1989.
The Jurassic, Cretaceous and Tertiary biostratum in Tibet.

Zhou Zhicheng et al/Marine Micropaleontology 32 (1997) 3-29 29
China Univ. Geosci. Press, 147 pp. (in Chinese, with English abstract).
Yin, Jixiang, 1988. The evolutionary outline on stratigraphy and geology of Southern Tethyan Domain in the Qinghai-Xizang (Tibet) Plateau. Bull. Inst. Geol. Acad. Sin. Sci. Press 3, l-25 (in Chinese, with English abstract)..
Zhang, Binggao, Zhou, IZhicheng, Willems, H., 1994. Discovery of Late Cretaceous calpionellids and related microfossils from Tuna and Gamba regi~ons, Southern Tibet. Acta Micropalaeon-
tol. Sin. 11 (4), 413-428 (in Chinese, with English abstract). Zhou Zhicheng, 1997. Cretaceous trace fossils from Tibet. N. Jb.
Geol. Pallontol. Abh. 203(2), 145-172. Zhou Zhicheng and Zhang Binggao, 1992. The depositional fea-
tures and the environmental significance of Cretaceous and Paleogene in Southern Xizang. In: China Society of the Qinghai-Xizang Plateau Research (Ed.), Proc. First Symp. on the Qinghai-Xizang Plateau, pp. 280-286 (in Chinese, with English abstract).